Global versus local processing: seeing the left side of the forest and the right side of the trees
- 1Department of Psychology, Dalhousie University, Nova Scotia, Canada
- 2Department of Communication Sciences and Disorders, University of South Carolina, Columbia, SC, USA
- 3WJB Dorn VA Medical Center, Columbia, SC, USA
- 4Department of Psychology, University of South Carolina, Columbia, SC, USA
Previous studies using hierarchical figures (where a large global shape is composed of a series of smaller local shapes) suggest that performance is better for local features presented in the right relative to left visual field, whereas the opposite pattern is observed for global features. However, these previous studies have focused on effects between hemifields. Recent data from patients with neurological damage suggest that local deficits can be allocentric (e.g., following left hemisphere injury, individuals are relatively slow to detect features on the right side of an object, regardless of visual field). Therefore, we decided to extend previous global versus local research by also observing local performance within hemifields. Specifically, on each trial we presented two hierarchical figures (one in each hemifield), but crucially the left and right side of each item were composed of different local features. In this task, the participant simply reports if a circle is present, regardless of location or whether this is a local or global feature. We observed that both neurologically healthy individuals, as well as an individual with brain injury, were relatively better detecting local information on the right side of objects, regardless of spatial location, while both showed better performance for global stimuli in the left visual field. This work is consistent with recent work in patients with neurological damage, and provides a new paradigm for exploring hemispheric specialization.
Introduction
While the human brain appears roughly symmetrical, there is clear evidence that there is some hemispheric specialization, for example consider the well-established fact that the left hemisphere plays a dominant role for language in most individuals. There is also evidence that perceptual processing may exhibit hemispheric specialization. A popular concept is that patients with left hemisphere lesions have difficulty with local details while patients with right hemisphere damage have difficulty with global form. Support for this notion comes from studies where patients are shown hierarchical figures (large shapes composed of smaller shapes, as seen in the insets of Figure 2) and asked to report either the small or large shapes. Individuals with left hemisphere damage are slower to report local elements while those with right hemisphere damage are relatively slower to report the large item (Delis et al., 1986; Robertson et al., 1988). Similar, though small, effects have been reported in healthy adults (Yovel et al., 2001), where individuals perform better for local information in the right visual field (presumably reflecting left hemisphere specialization), while performance is for global information is better in the left visual field (presumably due to the right hemisphere's specialization). However, previous studies contrasting local and global processing in healthy adults have focused on performance between hemifields, though careful review of literature for individuals with brain injury suggest local processing may be object-centered (with left hemisphere tuned for the right side of stimuli, regardless of hemifield). Our aim was to develop a paradigm that is intuitive for the participant yet allows us to observe global as well as both egocentric (between hemifields) local and allocentric (object-centered) local performance in a single session.
Numerous studies of healthy individuals as well as patients with neurological damage suggest that the right hemisphere is biased toward global information whereas the left hemisphere appears to be more specialized for local information, as observed following stroke or hemispherectomy (see Yovel et al., 2001 for a review). Classically, these studies have used compound hierarchical figures (Navon, 1977), where small letters are grouped to form a larger letter, with individuals with right hemisphere injury being particularly impaired with recognizing the larger letter whereas right injury impairs performance with local items. These findings have been taken to suggest that left hemisphere is biased toward processing high spatial frequencies, while the right is biased toward low spatial frequencies (Sergent, 1982).
While Sergent (1982) suggests a static model for hemispheric specialization, some evidence suggests that the calibration of “local” and “global” scale is dynamic and is determined by the task at hand (Robertson and Ivry, 2000). For example, Shulman and Wilson (1987) presented individuals with hierarchical figures while also detecting low contrast sinusoidal gratings. They found that when individuals were asked to attend to the global letters, they were more sensitive to detect the low frequency gratings. On the other hand, attending to the local letters improved detection of the high frequency gratings. Flevaris et al. (2010) recently demonstrated that this can be reversed and lateralized: attending the high frequency gratings improves sensitivity to the local elements especially for items in the right hemifield while attending low spatial frequency gratings reduces errors for global targets but more so for items presented in the left hemifield.
Robertson and Ivry (2000) provide a model to interpret these findings. They hypothesize that that the visual system employs an initial spatial frequency filter based on the task at hand. If accurate, this suggests that blocked studies may be manipulating low level perception (with filter optimized in anticipation of upcoming target scale), as well as higher-level processing. Robertson and Ivry (2000) describe a thought-experiment where participants are shown a hierarchical figure, but do not know whether the target will be defined at the global or local level. They propose that in this case the initial filter would be set broadly, allowing each hemisphere to exhibit its preference (e.g., left hemisphere showing a local benefit, while the right hemisphere exhibits a global preference).
Yet, it should be noted that previous studies exploring hemispheric specialization have suggested that these effects are subtle and difficult to detect. For example, in a meta-analysis of 16 previous studies Yovel et al. (2001) note that the majority failed to find unequivocal support for this double dissociation. Based on this review and additional new experiments, Yovel et al. (2001) suggest that prior studies often failed to find effects because of the paradigm used. Specifically, they argue that effects are smaller when the scale of the target is unknown (when a target can occur at either local or global level, which they refer to as divided attention) than when the scale is blocked so that the participant only needs to search for targets on one scale during each trial (focused attention). They note that during divided attention “one hemisphere would be disadvantaged by the necessity to switch attention from its preferred level to the target level and also by poorer processing of the target once attention is switched.” While this claim makes several assumptions (e.g., that a hemisphere can only process at one level, versus processing in parallel but with different efficiencies; and switching costs to shift levels), it does provide an explanation for their findings. However, Robertson and Ivry's suggestions that divided tasks may lead to different bottom-up tuning for local and global stimuli does provide an alternative hypothesis: focused tasks engage a narrowly tuned initial spatial filter, whereas the divided task necessitates a broader initial filter. If accurate, this makes interpretation of focused tasks more difficult as it is unclear whether the effects found in divided tasks reflect asymmetries in bottom-up perceptual filtering or top-down target selection.
In any case, Yovel et al. (2001) provide indisputable evidence that current paradigms used in healthy adults to detect global versus local asymmetries only find subtle effects. A careful review of the literature from patients with neurological injury suggests an alternative explanation for these small effects. Specifically, whereas most right hemisphere patients who exhibit deficits of spatial perception exhibit egocentric neglect (missing information on their contralesional side), some patients exhibit allocentric spatial neglect, where they fail to detect features on the contralesional side of an object, regardless of its location in space (for reviews, see Driver and Pouget, 2000; Marsh and Hillis, 2008; Karnath and Rorden, 2011). While egocentric spatial neglect is typically considered more frequent after right hemisphere injury, the allocentric form appears to be somewhat more common after left hemisphere (Kleinman et al., 2007) versus right hemisphere (Hillis et al., 2005) injury. The performance of these patients suggests that each hemisphere may have allocentric local processing, which is not bounded by hemifield. In other words, parts of the left hemisphere process the right side of objects, whereas the right hemisphere preferentially processes the left side of objects (though perhaps with less efficiency than the left hemisphere). This could explain the poor track record of previous studies that sought lateral biases in local perceptual processing. In those studies local features are found on both sides of the target objects, presumably projecting to each hemisphere's allocentric processing. Here we directly test whether this dissociation is found in neurologically healthy adults.
Our aim was to explore the laterality of global and local information using a hierarchical figure task where the participant is not explicitly required to focus on local elements on some trials and global features on other trials. Specifically, we created a task where, on every trial, the participant was shown two compound objects (one in each hemifield), with the task being to report the presence or absence of a circle (regardless of whether it was a local or global feature). Unlike previous studies, in our design the participant is unable to anticipate whether the relevant information in an upcoming trial will be high or low frequency. Furthermore, the laterality of the local stimulus is not only at the level of hemifield of presentation but also of the side of the global object. Unlike any prior study in healthy adults, we will be able to examine the prior finding of hemifield effects as well as those within objects.
As noted, evidence for hemispheric specialization of local and global information comes from both healthy individuals as well as patients with neurological damage. Therefore, our first aim was to implement this task in neurologically healthy individuals. However, we also examined the behavior of a patient to demonstrate that this new paradigm overcomes a major obstacle in applying the focused attention hierarchical figure task in a neuropsychological setting. In particular, in conventional tasks patients must be told in some blocks to ignore large stimuli while reporting small stimuli; and then forced to reverse this pattern in other blocks. This task switching can be challenging for patients who perseverate. Furthermore, it makes interpreting data difficult, as it is unclear whether slow performance on one task reflects poor performance at the attended level, or difficulty in inhibiting the unattended level. Previous studies have tried to tease apart these inhibition effects by contrasting congruent (where local and global features identify the same letter) to incongruent trials (where different letters are presented at the two scales). However, such manipulations can also encourage patients to perseverate (as the unattended dimension is predictive of the correct answer on the congruent trials). Our novel solution to this problem is to always have a consistent task—the participant is asked to always report whether a circle is present, regardless of whether it is at the local or global level. This allows us to interleave trials with global targets with other trials that have local targets.
A novel aspect of our paradigm is the fact that the local features are different on the left and right side of each global object. Yovel et al. (2001) note that many studies of global and local perception have failed to find effects in neurologically healthy adults, though they suggest effects are most robust in divided attention designs. An alternative explanation is that the left hemisphere local specialization may apply to both visual hemifields. Evidence for this comes from Kleinman et al. (2007), who found that individuals with left hemisphere injury often neglect local information on the right side of items. In other words, patients with right hemisphere injury appear to have difficulty with relatively leftward global items, but those with left hemisphere damage appear to have difficulty with local information presented on the right side of objects.
Methods
Participants
All protocols of recruitment, reimbursement, and testing were approved by our institutional review board. We tested 12 neurologically healthy university graduate students (one male, mean age of 22.25 with a SD of 0.625). Each student received 10 dollars for volunteering in this study. In addition, the participants were told that the individual with the best performance (as determined by dividing mean response time [RT] by mean error rate) would receive an additional forty-dollar award. This incentive was designed to encourage high performance on this task. In addition, we tested a male patient with neurological damage who was 59 years old at the time of the MRI imaging and first testing session (and 60 at the time of the second testing session). His chronic stroke damage can be seen in Figure 1. The patient exhibited full visual fields, as assessed by confrontation testing. No personal neglect was exhibited (correctly identifying six regions on contralesional side of body, Cocchini et al., 2001). No clear spatial bias was seen on a conventional object cancellation task (detecting 41 of 60 occurrences of the letter “A” in a cluttered field of distractors, missing nine on the left half of the page and 10 on the right half, 14 of the missed targets were along the bottom edge of the page). The patient did report weakness of the left arm and some pain in the left leg, and attributed these to a spinal injury. The patient received 20 dollars compensation for each session.
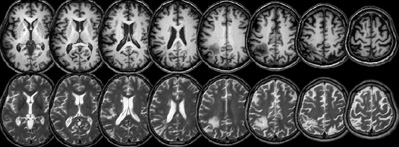
Figure 1. Axial MRI slices from the patient with neurological damage are shown in neurological convention (with the left side of each slice corresponding to the left side of the brain). Both the T1 (top row) and T2 (bottom row) images have been normalized to MNI stereotaxic space, with slices corresponding to Z = 8, 16, 24, 32, 40, 50, 60 mm.
Medical Imaging
MRI scans were acquired for the patient using a Siemens Trio 3T system with a 12-channel head coil. We acquired a T1-weighted 3D MP-RAGE with a TI of 900 ms, a TR of 2250 ms between TFE shots and a 9°, with a very short TE (4.52 ms). A total of 160 1 mm sagittal slices were acquired, each with a 256 × 256 matrix and 256 × 256 mm FOV. In addition, a T2-weighted SPACE (Sampling Perfection with Application optimized Contrasts using different flip-angle Evolution) 3D TSE volume was acquired, using a TR of 3200 ms, a TE of 354 ms, 256 × 256 matrix scan with 160 slices yielding a 1 × 1 × 1 mm resolution. Both images were acquired using the same slice positioning parameters. Images were processed using SPM5 (Wellcome Department of Imaging Neuroscience). The images were converted from DICOM to NIfTI format using dcm2nii (Rorden et al., 2007), and the origin was manually set to designate the Anterior Commissure (with identical transforms applied to both images, ensuring a good starting estimate for the subsequent steps). First, the T2 image was co-registered to match the alignment of the T1 image. Next, the T1 image was normalized to stereotaxic space using SPM5's in-built unified segmentation and normalization routines. The resulting spatial transforms were applied to both images. Note that this unified segmentation routine is very robust even with extensive brain injury (Crinion et al., 2007). The resulting images are illustrated in Figure 1.
Apparatus
Stimuli were presented from the built-in 15.4″ LCD display of a Compaq R3000 using a Python program written by the first author. Examples of these displays are shown along the bottom of Figure 2. Each of the small (local) stimuli measured 1.2° of visual angle and the larger (global) stimuli had a diameter that subtended 8.6°, with the centers of the global objects positioned 5.9° left and right of fixation. There were eight possible shapes presented, the target circle, and the distractors: square, diamond, triangle, and four different orientations of “D” (0, 90, 180, and 270°).
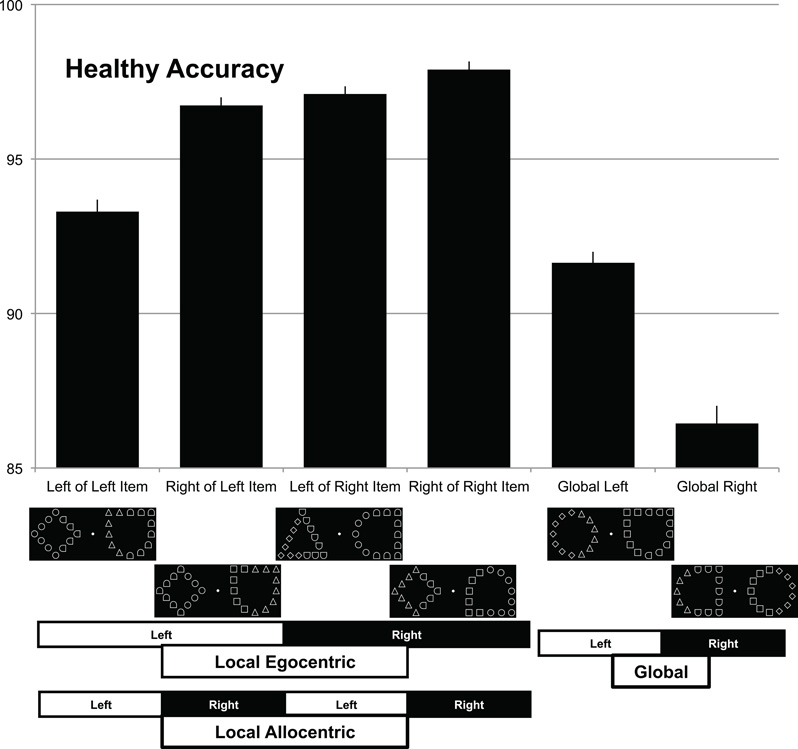
Figure 2. Performance for healthy individuals performing this task. Percent accuracy for targets appearing at different locations (left vs. right) and different scales (local vs. global). The task was to press a green button whenever a circle was present, regardless of whether this was a local feature (four leftmost columns) or a global feature (two rightmost columns), and to press a red button if there was no circle present (not shown). Bars show performance for different target locations, with example displays shown beneath each column (with error bars showing normalized standard error, as suggested by Loftus and Masson, 1994). Note that local features are detected more accurately when they appear on the right relative to more leftward locations. This appears to occur both within objects (allocentric) and between visual fields (egocentric). Global form shows the opposite pattern with global targets detected more accurately on the left than the right visual field.
Procedure
The participant was seated in front of the computer (60 cm from screen) with the buttons on the right side. The red button was nearest the participant and the green button was behind it. A white fixation dot was present on the screen at all times. Each trial was initiated by pressing a yellow button (controlled by the experimenter for the patient, otherwise the trials were initiated by the participant with their left hand). The two bilateral hierarchical figures were then displayed, and remained visible until a response was made. The participant reported the presence or absence of a circle by pressing either a red or green 6.35 cm button (www.ablenetinc.com). For each subject, the mapping of red and green to present and absent was consistent throughout testing, but this mapping was counterbalanced across participants. A target was present on half of the trials. Each possible position for a target was equally represented (two global positions and four local positions). The neurologically healthy controls completed 10 blocks, each with 48 trials, while the patient completed five blocks of 48 trials in a single session. The participants were free to take a break at any time and the block separation was invisible to the participant. Upon inspection of the data, the patient was run through another session of 240 trials approximately 5 months later in order to get a better estimate of the effects (for a total of 480 trials for all participants).
Results
We examined the RTs and accuracies for trials where a target was present. Initial cleaning of the data involved removing the first two trials for each run of trials because these had extraordinarily long RTs and were used to familiarize the subject with the task. One healthy participant had substantially lower overall accuracy than the next lowest (68.8% vs. 92.7%) and was, therefore, removed as an outlier. Subsequently, a statistical outlier rejection was achieved by removing all RTs greater than 2.326 SD from the mean for each condition (Van Selst and Jolicoeur, 1994). This eliminated 3.38% of the trials from the control participant data (4.2% with patient) and they were all trials easily identified as outliers from visual inspection. The mean accuracies for control subjects are shown in Figure 2 while the patient accuracies are in Figure 3.
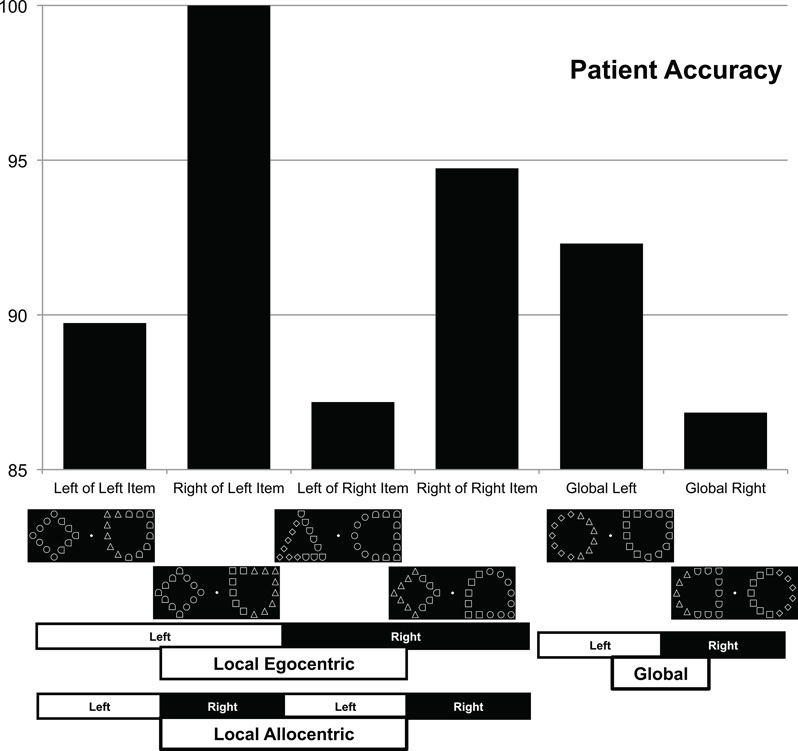
Figure 3. Performance for a single patient with neurological damage, using the same layout as Figure 2. Note that this individual exhibits the same pattern of allocentric and global performance as healthy adults, yet a reversed pattern for egocentric local items (finding local items more accurately if they appear in the left [ipsilesional] versus right [contralesional] visual field).
The data from the neurologically healthy controls was subjected to multi-level modeling (linear mixed effects analyses) with subject as a random factor. Additive models were constructed first to get individual main effects and interactions added afterwards. For RT data natural log RT were analyzed and the key statistic is a t value for the effect in the model. Note that there is no degrees of freedom value given for the t because an exact amount is not known for mixed effects models (Pinheiro and Bates, 2000). In our case (and most designs like this) the df is a relatively large value because each observation contributes. We will calculate probabilities assuming a conservative df of 61 (significant t's are two or greater, after Baayen, 2008). Accuracy data were analyzed with multi-level logistic regression.
An analysis of target scale (local vs. global) and corresponding target side (left vs. right) was conducted first. Note that for local targets the side was based on an object-centered representation (e.g., left local target values included both trials where the target was on the left side of the right object or the left side of the left object), and that collapsing the data in this fashion increases statistical power by increasing the number of samples and also controls for visual eccentricity. The analysis of RTs revealed no main effects (Scale t = 1.75, p = 0.085, se = 0.011; Side t = 0.30, p = 0.976, se = 0.010), nor was an interaction observed (t = 0.38, p = 0.71, se = 0.022). Using the model predictions and transforming back to RT it was found that these values were 1019 ms for global left and 1028 ms for global right, 1004 ms for local left and local right. A similar analysis of the accuracy of response revealed a main effect for the factor scale (z = 6.30, p < 0.0001), with more accurate performance for the local items. There was no main effect for the factor side (z = 0.088, p = 0.93), with roughly equal performance for targets appearing on the left or right side. However, we did observe the predicted interaction between scale and side (z = 3.42 p = 0.0006), with global items detected at a higher percentage (logit) on the left, 93.0% (2.59), relative to right visual field, 88.3% (2.03), and local items being more accurately detected on the right 97.8% (3.78) side of an object than on the left side of an object 95.7% (3.10).
A subsequent analysis of target scale (local vs. global) and global stimulus side (left vs. right) was conducted to see if the established finding that performance is better on targets presented to the right visual field was replicated. Unlike the prior analysis, left and right of the global object is collapsed across and only left and right visual field is preserved. The analysis of RTs again revealed no main effect of scale (t = 1.72, p = 0.09, se = 0.011) but a main effect of side (t = 2.33, p = 0.023, se = 0.010) with right faster than left. That main effect was qualified by an interaction (t = 2.17, p = 0.034, se = 0.022) because at the global level RTs were shorter to the left (1018 ms) than right (1027 ms) but at the local level this was reversed (1023 ms left vs. 985 ms right). A similar analysis computed for the accuracy of response found a main effect for scale (z = 6.30, p < 0.0001). There was no main effect for the factor side (z = 0.04, p = 0.97). The predicted interaction between scale and side was found (z = 3.56, p = 0.0004). This confirmed the finding in RT with global items detected more accurately on the left (logit) at 93.0% (2.59) relative to right visual field 88.3% (2.03) and local items being more accurately detected on the right side 97.9% (3.82) side than on the left 95.6% (3.08).
The complete descriptive information for all six conditions is shown in Table 1. With visual inspection suggesting an expected benefit for visual eccentricity (performance is improved when local targets are closer to the fovea), though note this effect is orthogonal to our primary effects of interest.
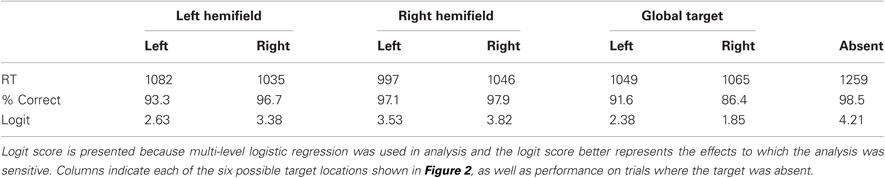
Table 1. The pattern of performance observed for 12 neurologically healthy individuals, reporting the response time (RT, in ms derived from mean log RT), percent accuracy (% Correct) and logit accuracy.
Patient data analysis was similar to the control participants using generalized linear modeling without the multiple levels. Analysis of log target RTs with the factors of side (local level was object side) and scale revealed a main effect of target scale (global vs. local), t(209) = 5.49, p < 0.0001, se = 0.058. Specifically, the patient was faster to detect local targets (2363 ms) than global targets (3261 ms). This pattern of performance is typical for sparse Navon figures (Lamb and Robertson, 1988; Dalrymple et al., 2007). There was no main effect of side, t(203) = 1.34, p = 0.18, se = 0.055 but there was an interaction between scale and side. t(209) = 2.38, p = 0.018, se = 0.12. RT and accuracy scores are presented in Table 2. It can be seen that with the global stimuli performance is poorer on the right side, but with the local stimuli performance is poorer on the left side. A similar analysis was performed on accuracy with no main effects (Scale z = 0.855, p = 0.392, se = 0.49; Side z = 1.15, p = 0.252, se = 0.50) but there was an interaction that confirmed the one found in RTs, z = 1.97, p = 0.049, se = 1.11. There was better performance for left global targets [92.3% (2.48)] than right global targets [86.8% (1.89)] and better performance for right local targets [97.4% (3.62)] than left local targets [88.4% (2.04)].
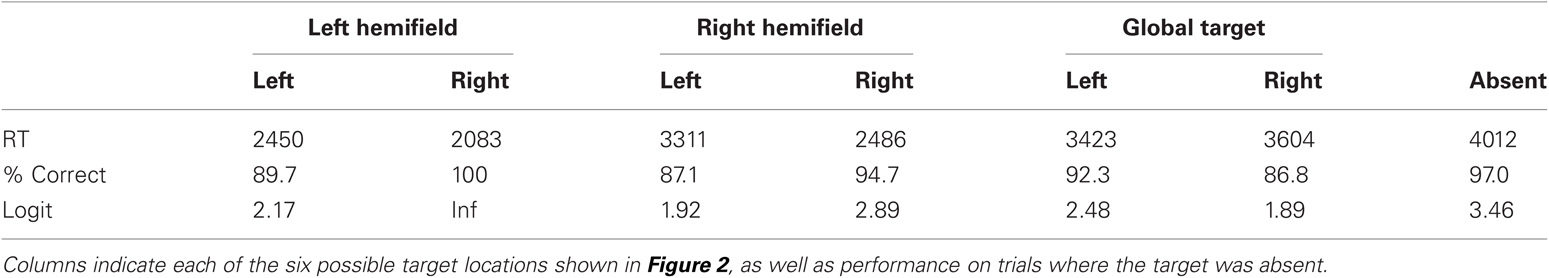
Table 2. Performance for a single patient with neurological damage, revealing the Response Time (RT, in ms derived from mean log RT), percent accuracy and logit accuracy.
As before, there was also an analysis of target scale and global stimulus side. Global and local effects were confirmed because essentially that analysis is just replicated here but the interaction between scale and side was not significant in RT [t(209) = 0.99, p = 0.32, se = 0.11] or accuracy (z = 0.017, p = 0.99, se = 1.01). Instead, there was a main effect of side in RT [t(209) = 3.54, p = 0.0005, se = 0.053] with left (2590 ms) faster than right (3039 ms). There was no effect of side in accuracy (z = 1.226, p = 0.22, se = 0.496) but general trends in accuracy confirmed the RT finding so it was not a speed-accuracy tradeoff.
The patient data suggests that the source of the local performance benefit in the right hemifield and that on the right side of an object found in the control participants may have different neural underpinnings.
Discussion
Our goal was to test the laterality effects predicted by Robertson and Ivry (2000) of local stimuli being processed by the left hemisphere and global stimuli processed by the right hemisphere with two new design features. The laterality effects in the visual field were replicated in healthy participants along with a new finding of object-centered effects where local and global stimuli were differentially processed within an item (e.g., best performance for local information on the right of an object, regardless of whether the object is in the left or right hemifield). Furthermore the method was shown to be amenable to data collection from a patient with neurological damage. However, because the single patient's performance was better with right local items that were part of an object that could appear in either hemifield, but not when the laterality of the local item was defined by visual hemifield, a common cause can be questioned. This study replicates effects previously described in both patients and healthy individuals while controlling for the expected scale of the targets (in previous studies the participants only needed to attend to local elements in some trials and global elements in other trials). Although these findings are novel in terms of the hierarchical figure literature, below we describe how such a finding is actually supported by the deficits observed in patients with neurological damage.
Individuals with damage to posterior brain regions often have difficulty perceiving visually presented stimuli. There are three well-known principles for the deficits observed. First, patients tend to ignore stimuli presented on their contralesional side (a symptom referred to as contralesional neglect). Second, chronic contralesional neglect is more commonly seen following right hemisphere injury. Third, patients with right hemisphere injury tend to have difficulty perceiving global form, while individuals with left hemisphere injury have difficulty perceiving local detail. While the literature notes exceptions to each of these rules-of-thumb, we describe a single patient with a pattern of deficits that does not cleanly fit any of these heuristics. Specifically, using a novel experimental pattern we have identified an individual with chronic left hemisphere injury who exhibits contralesional neglect for global information, yet ipsilesional neglect for local information. We believe these symptoms can best be explained by the normal hemispheric specialization for local and global object recognition.
Patients with neglect tend to ignore stimuli presented on one side. Classically, neglect is associated with right hemisphere injury, and typically these individuals ignore information on the contralesional side of space (Hillis et al., 2005). However, there are clearly exceptions to this rule. For example, Kleinman et al. (2007) observed a high incidence of neglect in patients with acute left hemisphere injury, with 9 of 47 individuals exhibiting neglect—while the same group using a similar protocol (Hillis et al., 2005) reported neglect in 16 of 50 individuals with right hemisphere injury. Crucially, the form of neglect between these two groups was different: with the majority of left hemisphere neglect being allocentric (missing information on one side of individual items) whereas those with acute right hemisphere damage tended to exhibit egocentric neglect (ignoring items on one side of space). Therefore, at least at the acute stage, there is evidence that left hemisphere neglect is prevalent, albeit in a form that is less overtly obvious and not often revealed by classic clinical testing (with most bedside tests sensitive to egocentric deficits).
Our new paradigm provides a method to disentangle global processing, local egocentric and local object-centered processing. While previous work has demonstrated that local performance is better in the right hemisphere than the left, our work extends this to show that right-sided benefits for local perception can be found within a hemifield. While these findings are clearly consistent with the notion of object-centered processing, there is one other model that could also explain the behavior we observed in healthy participants. Specifically, it is logically possible that the local processing benefit acts as a gradient—with better performance the more rightward in the visual field. This would account for our observed data (Figure 2), where accuracy increases from left to right. Such effects have been clearly established in neglect (Karnath and Niemeier, 2002) and can explain many of the allocentric effects attributed to neglect patients (Driver and Pouget, 2000). This explanation is very parsimonious, as it does not require positing independent egocentric and allocentric systems. On the other hand, data from the single neurological patient we observed does seem to provide some suggestion that one can see normal local allocentric biases without normal local egocentric biases. While this finding comes from a single patient, it does establish a method to dissociate these competing accounts. Likewise, future studies in healthy adults could present stimuli at different eccentricities across trials to directly adjudicate between these models. Indeed, inspection of the RT data in our healthy controls demonstrates that eccentricity appears to play a major factor in performance. While our analysis was designed to collapse across eccentricity (for both egocentric and allocentric performance) and combine RT and accuracy information, a dataset where eccentricity is manipulated could prove useful for testing between these two theories.
One nice feature of our paradigm is that the participant always does a single task: detecting the presence or absence of a circle. There is no need to explicitly tell the participant when to switch between local or global scales. This paradigm avoids the confound that switching tasks may invoke a different initial spatial frequency filter (as suggested by Robertson and Ivry, 2000). In addition, previous designs where task switching is required may incur larger between task-interference effects. We suggest that our participants' behavior replicates the pattern predicted by Robertson and Ivry, (2000) thought-experiment: performance for global information is better in the left visual field (presumably reflecting the right hemisphere's preference for global information) while perception of local elements is better if they occur on the right half of an object (which we infer as reflecting the left hemisphere's preference for local items).
As reviewed earlier, lateralized differences in local and global processing have often been interpreted as demonstrating specialization in processing of spatial frequency (Sergent, 1982; Robertson and Ivry, 2000). We note that there are alternative explanations for these effects. Indeed, one potential criticism regarding the “global” compound stimuli is that their grouping is implicit, whereas the shape is explicit. According to this model, the global stimuli differ not only in spatial scale, but also in the Gestalt binding required, which may be more efficient in the right hemisphere (Atchley and Atchley, 1998; Corballis et al., 1999). Alternatively, Kosslyn et al. (1998) interprets positron emission tomography data as demonstrating that the right hemisphere is more activated during a coordinate task (is a bar near or far from an ×) whereas the left hemisphere was more activated in a categorical task (is the bar above or below an ×). Likewise, Bradshaw and Nettleton (1983), describe the right hemisphere as better for configural, holistic processing while the left hemisphere is tuned for detailed analytical processing. New paradigms, such as the one described here will help elucidate the true functional differences between the two hemispheres.
Our primary aim was to extend previous work on local versus global specialization by coding allocentric local information. However, we note that our paradigm could be extended to examine whether global processing also exhibits allocentric properties. For example, consider an experiment where the task is to detect whether an object has any curved edges, such that a D and its mirror are targets whereas a K and its mirror do not have curved edges. With these stimuli, one could have both global and local elements that are defined as targets by their left and right side.
Although we provide additional evidence for hemispheric specialization, our interpretation does rely on one alteration to the common conception of hemispheric specialization: the left hemisphere benefit for local information is not bound by hemifield. Rather, the left hemisphere appears to provide a benefit for features on the right side of objects as well as a benefit for features in the right hemifield (potentially reflecting an object-centered frame of reference). While previous studies generated global hierarchical figures based on a single local shape, our study used different shapes for the left and right half of the object. This allows our paradigm to reveal an effect that is compatible with an object-centric frame of reference for the local bias. This finding is consistent with the work of Kleinman et al. (2007), where neglect following left hemisphere injury tended to be allocentric rather than egocentric (e.g., patients would neglect the right half of objects, rather than the right side of space).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Institutes of Health (R01 NS054266).
References
Atchley, R., and Atchley, P. (1998). Hemispheric specialization in the detection of subjective objects. Neuropsychologia 36, 1373–1386.
Baayen, R. H. (2008). Analyzing Linguistic Data: A Practical Introduction to Statistics. New York, NY: Cambridge University Press.
Bradshaw John, L., and Norman C. Nettleton. (1983). Human Cerebral Asymmetry. Englewood Cliffs, NJ: Prentice Hall Press. ISBN: 9780134446462
Cocchini, G., Beschin, N., and Jehkonen, M. (2001). The fluff test: a simple task to assess body representation neglect. Neuropsychol. Rehabil. 11, 17–31.
Corballis, P. M., Fendrich, R., Shapley, R. M., and Gazzaniga, M. S. (1999). Illusory contour perception and amodal boundary completion: evidence of a dissociation following callosotomy. J. Cogn. Neurosci. 11, 459–466.
Crinion, J., Ashburner, J., Leff, A., Brett, M., Price, C., and Friston, K. (2007). Spatial normalization of lesioned brains: performance evaluation and impact on fMRI analyses. Neuroimage 37, 866–875.
Dalrymple, K. A., Kingstone, A., and Barton, J. J. (2007). Seeing trees OR seeing forests in simultanagnosia: attentional capture can be local or global. Neuropsychologia 45, 871–875.
Delis, D. C., Robertson, L. C., and Efron, R. (1986). Hemispheric specialization of memory for visual hierarchical stimuli. Neuropsychologia 24, 205–214.
Driver, J., and Pouget, A. (2000). Object-centered visual neglect, or relative egocentric neglect? J. Cogn. Neurosci. 12, 542–545.
Flevaris, A. V., Bentin, S., and Robertson, L. C. (2010). Local or global? Attentional selection of spatial frequencies binds shapes to hierarchical levels. Psychol. Sci. 21, 424–431.
Hillis, A. E., Newhart, M., Heidler, J., Barker, P. B., Herskovits, E. H., and Degaonkar, M. (2005). Anatomy of spatial attention: insights from perfusion imaging and hemispatial neglect in acute stroke. J. Neurosci. 25, 3161–3167.
Karnath, H. O., and Niemeier, M. (2002). Task-dependent differences in the exploratory behaviour of patients with spatial neglect. Neuropsychologia 40, 1577–1585.
Karnath, H. O., and Rorden, C. (2011). The anatomy of spatial neglect. Neuropsychologia PMID: 21756924
Kleinman, J. T., Newhart, M., Davis, C., Heidler-Gary, J., Gottesman, R. F., and Hillis, A. E. (2007). Right hemispatial neglect: frequency and characterization following acute left hemisphere stroke. Brain Cogn. 64, 50–59.
Kosslyn, S. M., Thompson, W. L., Gitelman, D. R., and Alpert, N. M. (1998). Neural systems that encode categorical versus coordinate spatial relations: PET investigations. Psychobiology 26, 333–347.
Lamb, M. R., and Robertson, L. C. (1988). The processing of hierarchical stimuli: effects of retinal locus, locational uncertainty, and stimulus identity. Percept. Psychophys. 44, 172–181.
Loftus, G. R., and Masson, M. E. (1994). Using confidence intervals in within-subject designs. Psychon. Bull. Rev. 1, 476–490.
Marsh, E. B., and Hillis, A. E. (2008). Dissociation between egocentric and allocentric visuospatial and tactile neglect in acute stroke. Cortex 44, 1215–1220.
Navon, D. (1977). Forest before trees: the precedence of global features in visual perception. Cogn. Psychol. 9, 353–383.
Pinheiro, J. C., and Bates, D. M. (2000). Mixed Effects Models in S and S-Plus. New York, NY: Springer.
Robertson, L. C., and Ivry, R. (2000). Hemispheric asymmetry: attention to visual and auditory primitives. Curr. Dir. Psychol. Sci. 9, 59–63.
Robertson, L. C., Lamb, M. R., and Knight, R. T. (1988). Effects of lesions of temporal-parietal junction on perceptual and attentional processing in humans. J. Neurosci. 8, 3757–3769.
Rorden, C., Karnath, H. O., and Bonilha, L. (2007). Improving lesion-symptom mapping. J. Cogn. Neurosci. 19, 1081–1088.
Sergent, J. (1982). The cerebral balance of power: confrontation or cooperation? J. Exp. Psychol. Hum. Percept. Perform. 8, 253–272.
Shulman, G. L., and Wilson, J. (1987). Spatial frequency and selective attention to local and global information. Perception 16, 89–101.
Van Selst, M., and Jolicoeur, P. (1994). A solution to the effect of sample size on outlier elimination. Q. J. Exp. Psychol. 47A, 631–650.
Keywords: spatial selection, visual attention
Citation: Christie J, Ginsberg JP, Steedman J, Fridriksson J, Bonilha L and Rorden C (2012) Global versus local processing: seeing the left side of the forest and the right side of the trees. Front. Hum. Neurosci. 6:28. doi: 10.3389/fnhum.2012.00028
Received: 23 November 2011; Accepted: 08 February 2012;
Published online: 22 February 2012.
Edited by:
Kenneth Hugdahl, University of Bergen, NorwayReviewed by:
Jonathan Kleinman, Johns Hopkins, USAPaul M. Corballis, University of Auckland, New Zealand
Copyright: © 2012 Christie, Ginsberg, Steedman, Fridriksson, Bonilha and Rorden. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Christopher Rorden, Department of Psychology, McCausland Center for Brain Imaging, University of South Carolina, Columbia, SC 29208, USA. e-mail: rorden@sc.edu