 Ouahiba Khaib Dit Naib1
Ouahiba Khaib Dit Naib1 Mourad Aribi1,2*
Mourad Aribi1,2* Aicha Idder3Amel Chiali4Hakim Sairi4Isabelle Touitou5,6,7
Aicha Idder3Amel Chiali4Hakim Sairi4Isabelle Touitou5,6,7 Gérard Lefranc8
Gérard Lefranc8 Mouna Barat-Houari5
Mouna Barat-Houari5
- 1Laboratory of Applied Molecular Biology and Immunology. Department of Biology, Abou-Bekr Belkaïd University, Tlemcen, Algeria
- 2Departments of Pharmacy and Oral Medicine, Abou-Bekr Belkaïd University, Tlemcen, Algeria
- 3Etablissement Hospitalier Spécialisé en Ophtalmologie, Clinique Hamou Boutlelis, Oran, Algeria
- 4Service de Dermatologie, Centre Hospitalo-Universitaire d’Oran, Oran, Algeria
- 5Unité Médicale des Maladies Auto-inflammatoires, Département de Génétique, CHRU, Montpellier, France
- 6Université Montpellier 1, Montpellier, France
- 7Génétique des Maladies Auto-inflammatoires et des Ostéo-arthropathies chroniques, INSERM U844, Montpellier, France
- 8Laboratoire d’Immunogénétique Moléculaire, Institut de Génétique Humaine, CNRS UPR 1142, et Université Montpellier 2, Montpellier, France
Objective: We have conducted the first study of the association of interleukin (IL)-10, tumor necrosis factor alpha (TNF-α), and IL23R-IL12RB2 region single nucleotide polymorphisms (SNPs) with Behçet’s disease (BD) in Western Algeria.
Methods: A total of 51 BD patients and 96 unrelated controls from West region of Algeria were genotyped by direct sequencing for 11 SNPs including 2 SNPs from the IL10 promoter [c.-819T > C (rs1800871), c.-592A > C (rs1800872)], 6 SNPs from the TNF-α promoter [c.-1211T > C (rs1799964), c.-1043C > A (rs1800630), c.-1037C > T (rs1799724), c.-556G > A (rs1800750), c.-488G > A (rs1800629), and c.-418G > A (rs361525)], and 3 SNPs from the IL23R-IL12RB2 region [g.67747415A > C (rs12119179), g.67740092G > A (rs11209032), and g.67760140T > C (rs924080)].
Results: The minor alleles c.-819T and c.-592A were significantly associated with BD [odds ratio (OR) = 2.18; 95% confidence interval (CI) 1.28–3.73, p = 0.003]; whereas, there was weaker association between TNF-α promoter SNPs or IL23R-IL12RB2 region and disease risk.
Conclusion: Unlike the TNF-α and the IL23R-IL12RB2 region SNPs, the two IL10 SNPs were strongly associated with BD. The -819T, and -592A alleles and the -819TT, -819CT, and -592AA and -592CA genotypes seem to be highly involved in the risk of developing of BD in the population of Western Algeria.
Introduction
Behçet’s disease (BD) is a systemic inflammatory multifactorial disease (1). It is characterized by recurrent episodes of oral and genital ulceration, skin, and ocular lesions (2). The disease is now recognized as a systemic vasculitis, given that it can affect other tissues and organs including blood vessels, the digestive tract, and nervous system (3, 4).
The etiology of BD is not fully elucidated. There is a hypothesis that a pathogenic autoimmune process of BD is triggered by an infectious or environmental agent, in individuals genetically predisposed (5). The most strongly genetic factor associated with the disease is HLA B51 (5, 6). This association was initially described in 1973 (7) and subsequently confirmed in different ethnic groups (6, 8, 9).
However, the association between BD and HLA B51 represents only 20% in the siblings of patients with the disease (10), and 50% of cases with BD are negative for this allele (8, 9). These observations suggest the existence of other risk factors outside the HLA region. In fact, several recent genome-wide association studies have identified additional and new genomic regions that predispose to the disease (11–14).
The BD causes inflammation and chronic immune activation within small blood vessels (15–17). The site of inflammation is usually characterized by infiltration of immune cells as well as by highly elevated levels of different cytokines (18). IL-10 is one of the most important cytokines that has been observed at increased level in the serum and active lesions of BD patients (19–21). TNF-α and soluble TNF-α receptors are also elevated in the sera of patients with BD (22–24). Both IL-10 and TNF-α cytokines have been shown to play an important role in the immunopathology of autoimmune diseases (25–28), and an opposite roles in the inflammatory responses (29, 30). An autoregulatory loop appears to exist in whereby TNF-α induces IL-10 production, which ultimately reduces TNF-α synthesis (31, 32). It has been suggested that the increase of IL-10 may down-regulate the expression of NO, prompting the protective role of elevation of IL-10 (33). Additionally, treatment with anti-TNF-α monoclonal antibodies has resulted in improvement of various manifestations of BD (34, 35).
IL-10 and TNF-α production may be regulated at the transcriptional level. Thus, several single nucleotide polymorphisms (SNPs) at the promoter of IL10 and TNF-α gene have been shown to be associated with changes in the expression levels of IL-10 and TNF-α production (36, 37). On the other hand, numerous recent studies have demonstrated an association between BD and several IL10 (13, 14, 38, 39) and TNF-α (39–43) SNPs in different ethnic groups. However, to date, there are no analogous or identical investigations in Algeria.
Two others cytokines, IL-23 and IL-12, may play an important role in BD pathogenesis; their levels are elevated in BD patients (44–46). IL-23 drives and promotes the development of a unique T-helper cell population that produces IL-17, Th17 cells. These IL-23-driven Th17 cells are highly pathogenic and elicit IL-17-dependent inflammation in autoimmune diseases (47). IL-12, a heterodimeric cytokine, is of crucial relevance to cell-mediated immunity and Th1 differentiation (48). This cytokine exerts its biological effects via binding to a heterodimeric receptor consisting of IL12RB2 and IL-12RB1 subunits.
The effect of IL-23 and IL-12 is mediated through the IL-23 and the IL-12 receptor (IL-23R, IL-12RB1). The genes that encode these receptors are adjacent on chromosome 1p31; a GWAS Studies revealed that IL23R-IL12RB2 region is associated with BD (13, 14). Nevertheless, its association in the pathogenesis of BD remains to be confirmed in different ethnic groups. In this context, we examined genetic association for 11 SNPs in IL10, TNF-α, and IL23R-IL12RB2 candidate genes with BD in Western Algeria.
Materials and Methods
Patients and Subjects
Fifty-one (51) unrelated BD patients and age- and sex-matched 96 healthy controls originate from the Western Algeria were recruited for a case-control study at the Oran Ophthalmic Hamou Boutlelis Hospital, the Department of Dermatology of Oran Medical Centre University, and the Oran Blood Transfusion Centre (Algeria). Among the 51 patients, 11 DNA belonging to Algerian origin, were selected from the biobank DNA for Genetics Laboratory of Autoinflammatory Diseases, Arnaud de Villeneuve Hospital, Montpellier (France).
Consent was signed by each participant or participant’s parent or legal guardian if entrant is a minor, under the Rules of Ethics and Professional Conduct. Patient characteristics were recorded using a questionnaire. The diagnosis of patients was based especially on the criteria proposed in 1990 (49). The control group was composed of healthy subjects without a family history of autoinflammatory diseases, and selected from the same population. This work was approved by the Institutional Ethics Board of Tlemcen Abou-Bekr Belkaïd University.
Genotyping
Each DNA was genotyped for 11 SNPs, including two IL10 promoter SNPs [c.-819T > C (rs1800871), c.-592A > C (rs1800872)], six SNPs from the TNF-α promoter [c.-1211T > C (rs1799964), c.-1043C > A (rs1800630), c.-1037C > T (rs1799724), c.-556G > A (rs1800750), c.-488G > A (rs1800629), and c.-418G > A (rs361525)], and three SNPs from the IL23R-IL12RB2 region [g.67747415A > C (rs12119179), g.67740092G > A (rs11209032), and g.67760140T > C (rs924080)].
Genotyping was performed at the Laboratory of Genetics of Autoinflammatory Diseases, Arnaud de Villeneuve Hospital, Montpellier (France). Genomic DNA was isolated from peripheral blood, drawed on EDTA anti-coagulant, using QIAamp DNA Blood Kits (Qiagen, Valencia, CA, USA). The DNA samples were then dosed by spectrophotometry ND-1000 (Nano Drop Technologies, Wilmington, DE, USA) at 260 and 280 nm. The DNA concentration and ratio OD260/OD280 were estimated for each sample (50).
The DNA samples were subsequently amplified in a Applied Biosystems Thermocycler (Applied Biosystems, Foster City, CA, USA) in a 15 μL reaction volume containing 50 ng DNA, 2X Promega PCR Master Mix, and 25 μM of each primer (Table 1). The PCR programs were as follows: after a denaturation phase of 15 min at 95°C, the samples were subjected to 35 amplification cycles followed by a final elongation step of 7 min at 72°C. Each cycle comprises 30 s denaturation at 95°C, 30 s of primer annealing at 60°C, and 1 min extension at 72°C.
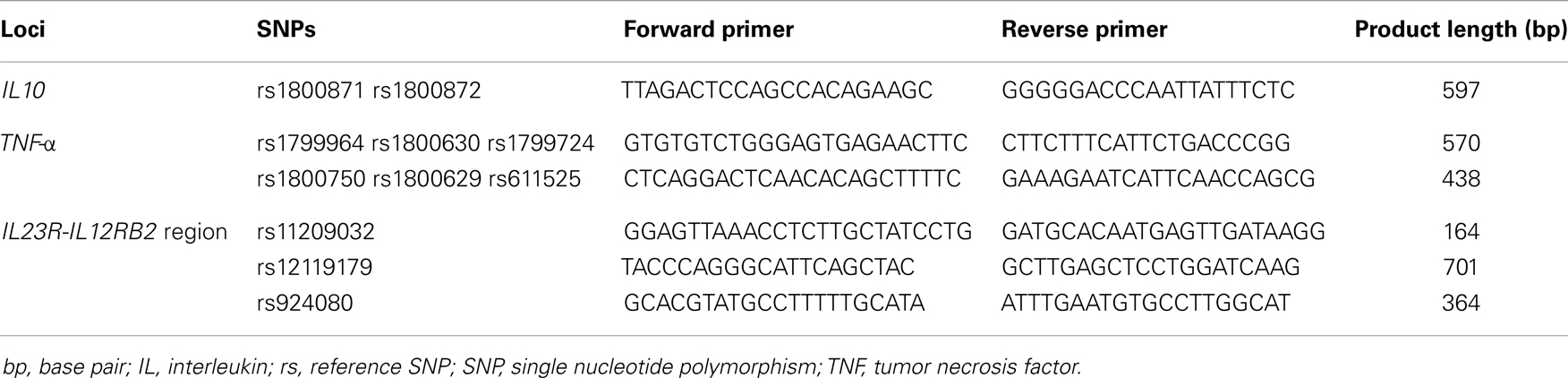
Table 1. Primers sequence and length product.
After checking the quality and size of the PCR products by agarose gel (1.5%) electrophoresis, SNPs genotyping was performed by direct sequencing using the BigDye Terminator version 3.1 (BDT v3.1) Cycle Sequencing Kit, followed by capillary electrophoresis on an ABI 3100XL Genetic Analyzer, according to the manufacturer’s recommendations (Applied Biosystems, Foster City, CA, USA) (Figure 1).
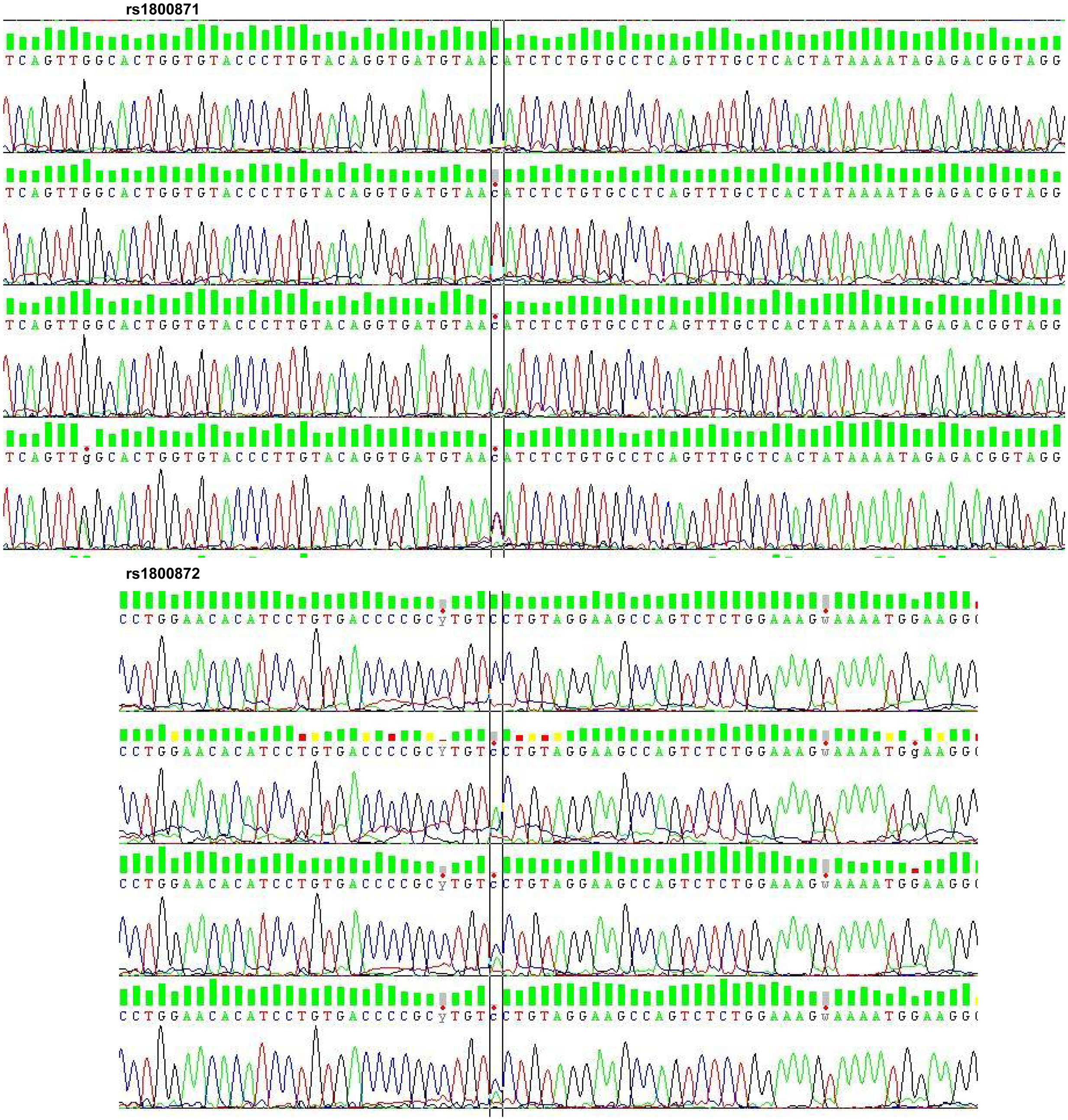
Figure 1. Electropherogram of rs1800871 and rs1800872. rs, reference SNP; SNP, single nucleotide polymorphism.
Statistical Analysis
Comparisons of allele and genotype frequencies between groups (patients versus control subjects, and between the patient’s groups according to different clinical features) were performed using the Chi-square or Fisher’s exact tests. The association analysis was carried out by Odds ratio (OR) and corresponding 95% confidence interval (95% CI). Statistical analyses were performed using GraphPad Prism Version 5.04 (GraphPad Software, Inc., La Jolla, CA, USA) and Epi Info 2000 Version 1.0 for Windows (Epi Info, Atlanta, GA, USA) software.
Results
Table 2 shows the description of the clinical characteristics of the patients with BD of the current study. The mean age (±SD) of the patients at disease onset was 26 ± 11 years. Predominant lesions were oral ulcers (100%), cutaneous lesions (86.27%), genital ulcers (82.35%), eye lesions (62.74%), and arthritis (58.82%).
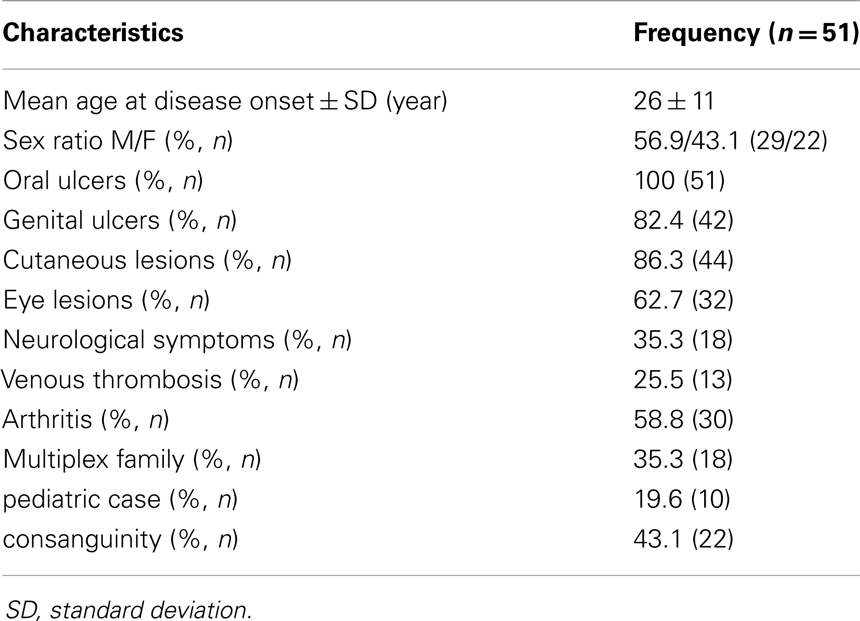
Table 2. Clinical and demographic features of the Behçet patients of the current study.
The distribution of alleles and genotypes frequencies of IL10 promoter SNPs c.-819C > T (rs1800871) and c.-592C > A (rs1800872) showed that the two SNPs were in total linkage disequilibrium in our sample. For this, reason the results of one SNP c.-819C > T will be considered (Table 3).
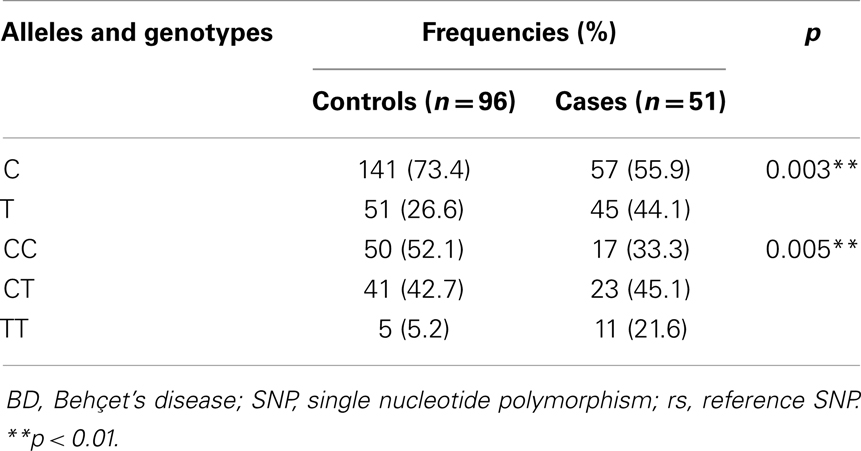
Table 3. Allelic and genotypic frequencies of rs1800871 variant in BD patients and controls.
The allele frequencies were significantly different in patients compared to controls. As indicated in Table 3, the frequencies of c.-819T (rs1800871) allele, and of the -819TT, -819CT (rs1800871) genotypes were significantly increased in patients than in controls (p = 0.003 and p = 0.005, respectively). Additionally, these SNPs was significantly associated with the disease (c.-819T; OR = 2.18, 95% CI 1.28–3.73, p < 0.01; -819TT and -819CT, OR = 2.17, 95% CI 1.01–4.69, p < 0.05) (Figure 2).
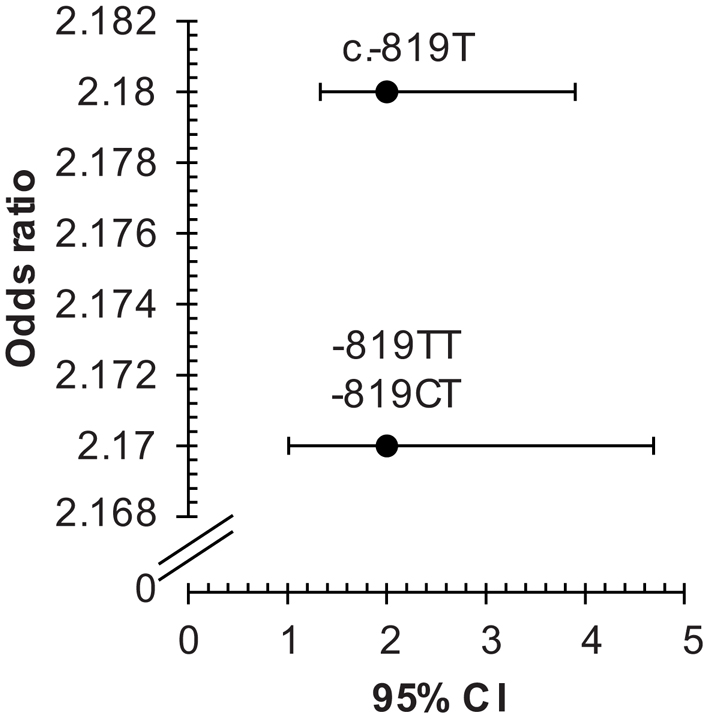
Figure 2. Odds ratios for associations between IL10 c.-819C > T, and -819TT, -819CT (rs1800871) with Behçet’s disease. The two SNPs c.-819C > T and c.-592C > A are in total linkage disequilibrium in our sample; therefore, the results of only one SNP c.-819C > T is considered. CI, confidence interval; rs, reference SNP; SNP, single nucleotide polymorphism.
A subset analysis was performed to examine the difference in allele frequencies in clinical subsets of BD (Table 4). We observed a significant association between c.-819T and all classes; nevertheless, the association was slightly lower for the ocular lesion (OR = 1.55, 95% CI 0.81–2.96, p > 0.05). Additionally, the association was more significant for the Genital ulcers (OR = 2.21; 95% CI 1.29–4.04, p = 0.002).
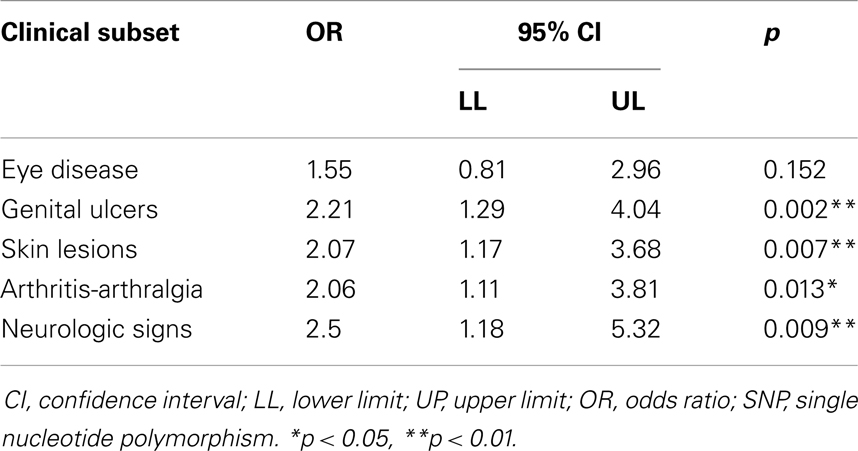
Table 4. Association analysis of clinical subclasses with IL10 c.-819T SNP in patients with Behçet’s disease.
We reported in Tables 5 and 6 that all IL23R-IL12RB2 SNPs alleles and genotypes, respectively, were not significantly associated with the disease (OR > 1, p > 0.05). The minor allele frequencies were different in the two groups, but this difference did not reach statistical significance (p > 0.05).
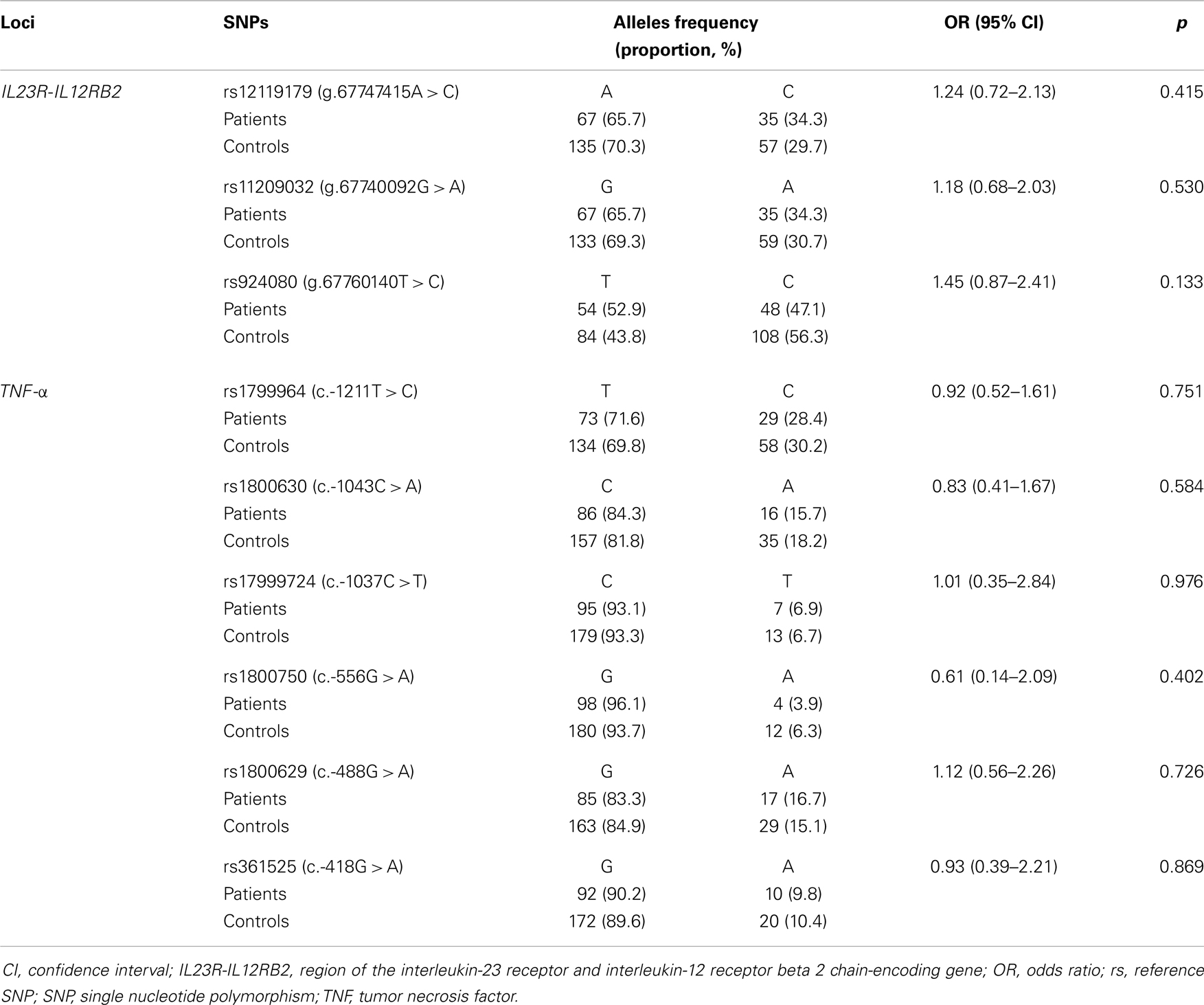
Table 5. IL23R-IL12RB2 and TNF-α allelic frequencies in patients with Behçet’s disease.
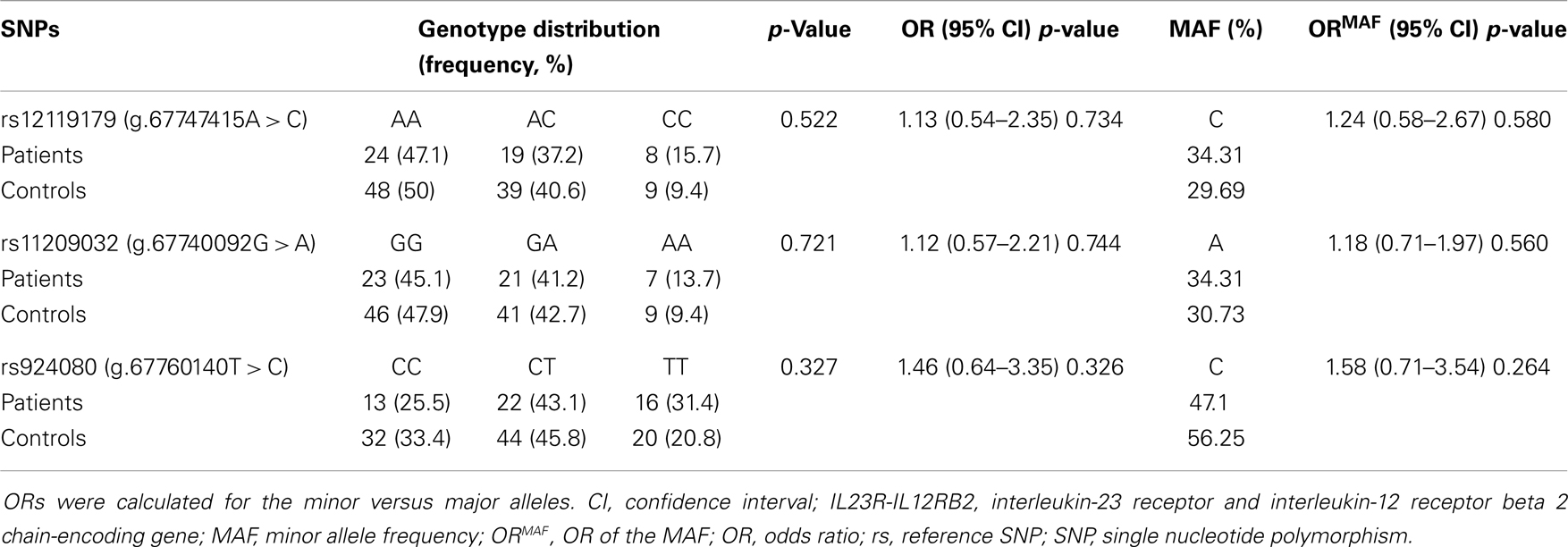
Table 6. The distribution of IL23R-IL12RB2 genotypes in patients with Behçet disease.
As indicated in Tables 5 and 7, alleles and genotypes of the TNF-α polymorphisms display similar distributions in patients and controls (p > 0.05). Except for c.-1037T and c.-488A all others TNF-α alleles were not associated with BD (OR < 1).
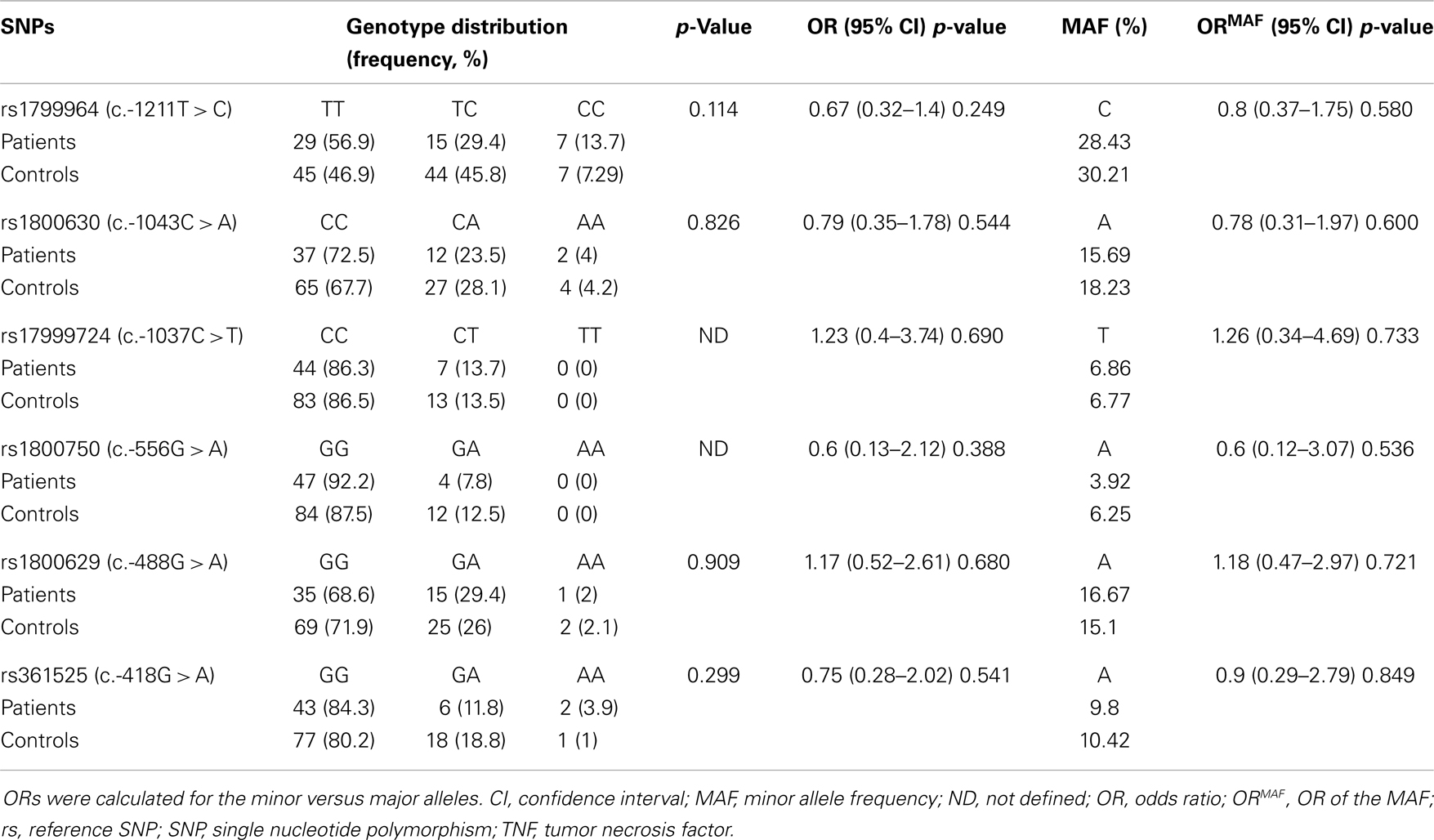
Table 7. The distribution of TNF-α genotypes in patients with Behçet disease.
Discussion
To date, the etiopathogenesis of BD is not fully elucidated. Researches in recent decades have shown the complex role of genetic factors in the development of the disease. We analyze the association between BD and 11 SNPs in IL10, TNF-α, and IL23R-IL12RB2 candidate genes in the Western Algeria population.
This is the first report demonstrating that the c.-819T and c.-592A alleles were associated with BD in Algeria.
Previous genetic studies have shown a strong association of many IL10 variants with BD in different ethnic groups. Recent genome-wide association study conducted by Mizuki et al. (14) in a Japanese cohort, including 612 individuals with BD and 740 unaffected individuals controls from different ethnic groups, has shown a significant difference between the two groups for five IL10 SNPs. The two SNPs analyzed in our study showed an association (OR = 2.18, 95% CI 1.28–3.73, p < 0.01). On the other hand, a strong association has also been highlighted for IL10 rs1518111 (OR = 1.45, 95% CI 1.34–1.58) in a genome-wide study performed in 311,459 SNPs in 1215 individuals with BD and 1278 healthy controls from Turkey (13). Our results are consistent with those obtained by Wallace et al. (38) who studied the segregation of two IL10 promoter SNPs, rs1800871 and rs1800896 in 178 cases and 295 controls from two populations, including Arab Middle East and United Kingdom. A strong association of the 819T allele has been observed in United Kingdom patients (OR 1.5, 95% CI 1.1–2).
Our results are, however, in disagreement with those of the study of Ates et al. (39) in which no significant association was revealed by exploring three IL10 SNPs (-1082G > A, rs1800896, c.-819C > T, rs1800871, and c.-592C > A, rs1800872) in 102 patients with BD and 102 controls from Turkey. These conflicting results may be explained by ethnic differences.
Several studies have shown the association of the two (c.-819 C > T, c.-592C > A) studied SNPs with various inflammatory diseases, but also with cancer (51–53), periodontitis (54), and docetaxel-induced liver injury (55). These associations suggested that the two SNPs can play an important role in the expression of IL10. In fact, it has been previously reported that the SNP c.-819C > T and/or c.-592C > A alleles affect the transcription of IL10 (56). Other studies performed on three SNPs at position -1082 A > G, -819C > T, and -592C > A in the promoter region of the IL10 gene have shown that the expression levels of IL-10 was significantly different according to the some haplotypes (57, 58). Finally, it has been reported that the disease-associated rs1518111 allele seems to be associated with low IL-10 mRNA expression and protein production (13). Indeed at the SNP rs1518111 locus, the rate of transcript of the G allele is higher than that of the A allele and patients homozygous for the A allele produce less IL-10 than those who are heterozygous or homozygous for the G allele.
The result of subset analysis suggests that the risk allele might predispose to genital ulcers, skin lesions, neurologic signs, and arthritis-arthralgia, but weakly to eye complications; the association was more significant for the genital ulcers (OR = 2.21, p = 0.002). Our results are in agreement with those of previous study (21) that showed a variable increase in mRNA expression within all BD lesions, including oral and genital ulcers, pseudofolliculitis lesions, and lesions at the site of pathergy testing.
Recent GWAS study from Turkey and Japan revealed IL12R-IL23RB2 SNPs in association with BD. Three SNPs were strongly associated with the disease, including rs924080 (OR = 1.28, p = 6.69 × 10−9) (13), rs12119179 (p = 2.7 × 10−8), and rs1495965 (OR = 1.35, p = 1.9 × 10−11) (14), but no significant association was found in a Korean cohort.
Our results showed no significant association between BD and rs12119179, g.67740092G > A (rs11209032), and g.67760140T > C (rs924080) SNPs in the IL23R-IL12RB2 region. In Iranian study (59), six SNPs in IL23R-IL12RB2 were found to be associated with BD; the most significant of which were rs17375018 (OR = 1.51, p = 1.93 × 10−6), rs7517847 (OR = 1.48, p = 1.23 × 10−6), and rs924080 (OR = 1.29, p = 1.78 × 10−5). Others studies have also identified a strong relationship between polymorphisms of IL23R and BD (60–62). These associations may suggest an important role of Th17 cells that express the IL-23R on their surface. Kim et al. (62) studied the interaction of specific IL17A, IL23R, and STAT4 (signal transducers and activators of transcription 4) SNPs in intestinal BD Korean patients; they suggest that the IL-23/IL-17 axis plays a significant role in disease pathogenesis.
IL-12 has been implicated in the pathogenesis of a multitude of diverse autoimmune diseases (63, 64). IL12RB2 constitute a risk factor for primary biliary cirrhosis, with the reported top associated SNPs mainly located in intronic sequences (65–67).
The genetic architecture and modularity of human autoimmune diseases is very complex. The functional implications of most of these associations are not yet clarified. Identify candidate causal SNPs and pathways (ICSN Pathway) analysis may act as a powerful guide to further research into the functional and immunological ramifications of these associations.
No significant associations were found between BD and studied TNF-α polymorphisms. These SNPs have been studied in various ethnic groups for possible association with BD. However, the allelic and genotypic associations of these studies have been contradictory. In Korean patients, TNF-α c.-1043A (rs1800630) allele was associated with an increased risk of BD (OR = 1.4, p = 0.030) (68). However, no significant association was found in meta-analysis studies for this SNP (42). Additionally, it has been reported a significant associations between c.-1037T allele (rs1799724) (OR = 0.76, 95% CI 0.58–0.98), c.-488G allele (rs1800629) (OR = 1.8, p = 0.010) (68), and c.-418A allele (rs361525) (OR = 1.51, 95% CI 1.12–2.04) (42), and BD. Moreover, no significant associations were identified with other TNF-α promoter polymorphisms, such as c.-1037T (rs1799724), c.-488A (rs1800629), and c.-556G > A (rs1800750) alleles with BD in Moroccan patients (43). The TNF-α c.-1211C allele (rs1799964) presented a significant association with BD in several populations, including Turkish (p = 0.023) (69), Korean (p = 0.030, OR = 1.4) (68), and UK white Caucasoid population (RR = 2.3, p = 0.00004) (40). The frequency of the TNF-α c.-1211C allele was significantly higher in Behcet’s patients than in healthy controls in Moroccan and Tunisian populations (OR = 1.65, p = 0.015; OR = 1.68, p = 0.02, respectively) (41, 43) and in meta-analysis (OR = 1.35, 95% CI 1.09–1.68) (42). This polymorphism has been associated with several extra-intestinal manifestations of Crohn’s disease, including uveitis, erythema nodosum, and large joint arthropathy (70), all of which are known to be associated with BD. Further investigation is necessary to determine the functional significance of TNF-α c.-1691042C and how it participates in the inflammatory dysregulation associated with BD.
Thus, polymorphisms at positions c.-1211T > C, c.-1043C > A, c.-1037C > T, and c.-488G > A have been associated with increased transcriptional activity and production of TNF-α in some studies (37, 71), in contrast to others (72–75). The over production of TNF-α during the course of BD may result in other TNF-α polymorphisms or post-transcriptional mechanisms. Furthermore, TNF-α production is not only under the control of the promoter region of TNF-α, and it may also result from complex cis and trans interactions among other cytokines.
TNF-α is encoded in the HLA complex on chromosome 6, a region that has long been known to be associated with BD. This gene-dense region, presentes a strong linkage disequilibrium (76). The association between BD and TNF-α could therefore be a result of linkage disequilibrium with alleles within this group. So it will be interesting to investigate other genes polymorphism among this region in our population.
In conclusion, we replicate the associations between BD and the SNPs from the IL23R-IL12RB2 region and c.-1037C > T and c.-488G > A TNF-α promoter SNPs. IL10 promoter SNPs (rs1800871 and rs1800872) is strongly associated with BD in the population of the Western Algeria. It would be interesting to study other SNPs to identify additional associations with BD in the studied population.
Authors Contribution
Mourad Aribi, Gérard Lefranc are Principal Investigators of the study, participated in its design and execution and helped draft the manuscript and critically reviewed it for intellectual content; Mouna Barat-Houari participated in the design of the study, carried out genetic analyses, and helped draft the manuscript; Ouahiba Khaib Dit Naib wrote the manuscript and carried out genetic analyses; Aicha Idder, Amel Chiali, and Hakim Sairi are responsible for the recruitment of eligible patients and their families; Isabelle Touitou conceived of the study, participated in its design, and coordination. All the authors read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to the patients, their families and the healthy controls for their participation. They would also like to address special thanks to Nathalie Ruiz-Pallares, Department of Genetics, CHRU, Montpellier, and all the staff of the Oran Blood Transfusion Centre for their help during this study.
References
1. McGonagle D, McDermott MF. A proposed classification of the immunological diseases. PLoS Med (2006) 3:e297. doi:10.1371/journal.pmed.0030297
2. Krause I, Weinberger A. Behçet’s disease. Curr Opin Rheumatol (2008) 20:82–7. doi:10.1097/BOR.0b013e3282f154d1
3. Suzuki Kurokawa M, Suzuki N. Behcet’s disease. Clin Exp Med (2004) 4:10–20. doi:10.1007/s10238-004-0033-4
4. Yazici H, Esen F. Mortality in Behçet’s syndrome. Clin Exp Rheumatol (2008) 26(Suppl 51):S138–40.
5. Fietta P. Behçet’s disease: familial clustering and immunogenetics. Clin Exp Rheumatol (2005) 23(Suppl 38):S96–105.
6. de Menthon M, Lavalley MP, Maldini C, Guillevin L, Mahr A. HLA-B51/B5 and the risk of Behçet’s disease: a systematic review and meta-analysis of case-control genetic association studies. Arthritis Rheum (2009) 61:1287–96. doi:10.1002/art.24642
7. Ohno S, Aoki K, Sugiura S, Nakayama E, Itakura K, Aizawa M. HLA-B5 and Behçet’s disease. Lancet (1973) 302:1383–4. doi:10.1016/S0140-6736(73)93343-6
8. Ohno S, Ohguchi M, Hirose S, Matsuda H, Wakisaka A, Aizawa M. Close association of HLA-Bw51 with Behçet’s disease. Arch Ophthalmol (1982) 100:1455–8. doi:10.1001/archopht.1982.01030040433013
9. Mizuki N, Inoko H, Mizuki N, Tanaka H, Kera J, Tsuiji K, et al. Human leukocyte antigen serologic and DNA typing of Behçet’s disease and its primary association with B51. Invest Ophthalmol Vis Sci (1992) 33:3332–40.
10. Yazici H, Fresko I, Yurdakul S. Behcet’s syndrome: disease manifestations, management, and advances in treatment. Nat Clin Pract Rheumatol (2007) 3:148–55. doi:10.1038/ncprheum0436
11. Karasneh J, Gül A, Ollier WE, Silman AJ, Worthington J. Whole-genome screening for susceptibility genes in multicase families with Behçet’s disease. Arthritis Rheum (2005) 52:1836–42. doi:10.1002/art.21060
12. Fei Y, Webb R, Cobb BL, Direskeneli H, Saruhan-Direskeneli G, Sawalha AH. Identification of novel genetic susceptibility loci for Behçet’s disease using a genome-wide association study. Arthritis Res Ther (2009) 11:R66. doi:10.1186/ar2695
13. Remmers EF, Cosan F, Kirino Y, Ombrello MJ, Abaci N, Satorius C, et al. Genome-wide association study identifies variants in the MHC class I, IL10, and IL23R-IL12RB2 regions associated with Behçet’s disease. Nat Genet (2010) 42:698–702. doi:10.1038/ng.625
14. Mizuki N, Meguro A, Ota M, Ohno S, Shiota T, Kawagoe T, et al. Genome-wide association studies identify IL23R-IL12RB2 and IL10 as Behçet’s disease susceptibility loci. Nat Genet (2010) 42:703–6. doi:10.1038/ng.624
15. Esin S, Gül A, Hodara V, Jeddi-Tehrani M, Dilsen N, Koniçe M, et al. Peripheral blood T cell expansions in patients with Behcet’s disease. Clin Exp Immunol (1997) 107:520–7.
16. Freysdottir J, Lau S, Fortune F. Gammadelta T cells in Behçet’s disease (BD) and recurrent aphthous stomatitis (RAS). Clin Exp Immunol (1999) 118:451–7.
17. Direskeneli H. Behcet’s disease: infectious aetiolozgy, new autoantigens, and HLA-B51. Ann Rheum Dis (2001) 60:996–1002. doi:10.1136/ard.60.11.996
18. Kulaber A, Tugal-Tutkun I, Yentür SP, Akman-Demir G, Kaneko F, Gül A, et al. Pro-inflammatory cellular immune response in Behçet’s disease. Rheumatol Int (2007) 27:1113–8. doi:10.1007/s00296-007-0367-9
19. Raziuddin S, al-Dalaan A, Bahabri S, Siraj AK, al-Sedairy S. Divergent cytokine production profile in Behçet’s disease. Altered Th1/Th2 cell cytokine pattern. J Rheumatol (1998) 25:329–33.
20. Hamzaoui K, Hamzaoui A, Guemira F, Bessioud M, Hamza M, Ayed K. Cytokine profile in Behçet’s disease patients. Relationship with disease activity. Scand J Rheumatol (2002) 31:205–10. doi:10.1080/030097402320318387
21. Ben Ahmed M, Houman H, Miled M, Dellagi K, Louzir H. Involvement of chemokines and Th1 cytokines in the pathogenesis of mucocutaneous lesions of Behçet’s disease. Arthritis Rheum (2004) 50:2291–5. doi:10.1002/art.20334
22. Lee EB, Kim JY, Lee YJ, Park MH, Song YW. TNF and TNF receptor polymorphisms in Korean Behcet’s disease patients. Hum Immunol (2003) 64:614–20. doi:10.1016/S0198-8859(03)00057-0
23. Akdeniz N, Esrefoglu M, Keles MS, Karakuzu A, Atasoy M. Serum interleukin-2, interleukin-6, tumour necrosis factor-alpha and nitric oxide levels in patients with Behcet’s disease. Ann Acad Med Singapore (2004) 33:596–9.
24. Oztas MO, Onder M, Gurer MA, Bukan N, Sancak B. Serum interleukin 18 and tumour necrosis factor-alpha levels are increased in Behcet’s disease. Clin Exp Dermatol (2005) 30:61–3. doi:10.1111/j.1365-2230.2004.01684.x
25. Vassalli P. The pathophysiology of tumor necrosis factors. Annu Rev Immunol (1992) 10:411–52. doi:10.1146/annurev.iy.10.040192.002211
26. Hofmann SR, Rösen-Wolff A, Tsokos GC, Hedrich CM. Biological properties and regulation of IL-10 related cytokines and their contribution to autoimmune disease and tissue injury. Clin Immunol (2012) 143:116–27. doi:10.1016/j.clim.2012.02.005
27. Iyer SS, Cheng G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit Rev Immunol (2012) 32:23–63. doi:10.1615/CritRevImmunol.v32.i1.30
28. Dai H, Ciric B, Zhang GX, Rostami A. Interleukin-10 plays a crucial role in suppression of experimental autoimmune encephalomyelitis by Bowman-Birk inhibitor. J Neuroimmunol (2012) 245:1–7. doi:10.1016/j.jneuroim.2012.01.005
29. Makhatadze NJ. Tumor necrosis factor locus: genetic organisation and biological implications. Hum Immunol (1998) 59:571–9. doi:10.1016/S0198-8859(98)00056-1
30. Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol (2001) 19:683–765. doi:10.1146/annurev.immunol.19.1.683
31. Lio D, Candore G, Colombo A, Colonna Romano G, Gervasi F, Marino V, et al. A genetically determined high setting of TNF alpha influences immunological parameters of HLA-B8, DR3 positive subjects: implications for autoimmunity. Hum Immunol (2001) 62:705–13. doi:10.1016/S0198-8859(01)00264-6
32. Candore G, Lio D, Colonna Romano G, Caruso C. Pathogenesis of autoimmune diseases associated with 8.1 ancestral haplotype: effect of multiple gene interactions. Autoimmun Rev (2002) 1:29–35. doi:10.1016/S1568-9972(01)00004-0
33. Belguendouz H, Messaoudène D, Lahmar K, Ahmedi L, Medjeber O, Hartani D, et al. Interferon-γ and nitric oxide production during Behçet uveitis: immunomodulatory effect of interleukin-10. J Interferon Cytokine Res (2011) 31:643–51. doi:10.1089/jir.2010.0148
34. Arayssi T, Hamra R, Homeidan F, Uthman I, Awwad ST, Mroue K, et al. The efficacy of a single dose of infliximab in the treatment of Behcet’s disease uveitis. Clin Exp Rheumatol (2005) 23:427.
35. Sfikakis PP, Markomichelakis N, Alpsoy E, Assaad-Khalil S, Bodaghi B, Gul A, et al. Anti-TNF therapy in the management of Behcet’s disease review and basis for recommendations. Rheumatology (Oxford) (2007) 46:736–41. doi:10.1093/rheumatology/kem034
36. Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet (1997) 24:1–8. doi:10.1111/j.1365-2370.1997.tb00001.x
37. Wilson AG, Symons JA, McDowell TL, McDevitt HO, Duff GW. Effects of a polymorphism in the human tumor necrosis factor a promoter on transcriptional activation. Proc Natl Acad Sci U S A (1997) 94:3195–9. doi:10.1073/pnas.94.7.3195
38. Wallace GR, Kondeatis E, Vaughan RW, Verity DH, Chen Y, Fortune F, et al. IL-10 genotype analysis in patients with Behçet’s disease. Hum Immunol (2007) 68:122–7. doi:10.1016/j.humimm.2006.11.010
39. Ates O, Dalyan L, Hatemi G, Hamuryudan V, Topal-Sarikaya A. Analyses of functional IL10 and TNF-α genotypes in Behçet’s syndrome. Mol Biol Rep (2010) 37:3637–41. doi:10.1007/s11033-010-0015-4
40. Ahmad T, Wallace GR, James T, Neville M, Bunce M, Mulcahy-Hawes K, et al. Mapping the HLA association in Behçet’s disease: a role for tumor necrosis factor polymorphisms? Arthritis Rheum (2003) 48:807–13. doi:10.1002/art.10815
41. Kamoun M, Chelbi H, Houman MH, Lacheb J, Hamzaoui K. Tumor necrosis factor gene polymorphisms in Tunisian patients with Behcet’s disease. Hum Immunol (2007) 683:201–5. doi:10.1016/j.humimm.2006.12.006
42. Touma Z, Farra C, Hamdan A, Shamseddeen W, Uthman I, Hourani H, et al. TNF Polymorphisms in patients with Behçet’s disease: a meta-analysis. Arch Med Res (2010) 41:142–6. doi:10.1016/j.arcmed.2010.02.002
43. Radouane A, Oudghiri M, Chakib A, Bennani S, Touitou I, Barat-Houari M. SNPs in the TNF-a gene promoter associated with Behçet’s disease in Moroccan patients. Rheumatology (Oxford) (2012) 51:1595–9. doi:10.1093/rheumatology/kes141
44. Guenane H, Hartani D, Chachoua L, Lahlou-Boukoffa OS, Mazari F, Touil-Boukoffa C. Production of Th1/Th2 cytokines and nitric oxide in Behçet’s uveitis and idiopathic uveitis. J Fr Ophtalmol (2006) 29:146–52. doi:JFO-02-2006-29-2-0181-5512-101019-200508206
45. Belguendouz H, Messaoudene D, Hartani D, Chachoua L, Ahmedi ML, Lahmar-Belguendouz K, et al. Effect of corticotherapy on interleukin-8 and -12 and nitric oxide production during Behçet and idiopathic uveitis. J Fr Ophtalmol (2008) 31:387–95. doi:10.1016/S0181-5512(08)71433-5
46. Chi W, Yang P, Zhu X, Wang Y, Chen L, Huang X, et al. Production of interleukin-17 in Behcet’s disease is inhibited by cyclosporin A. Mol Vis (2010) 19:880–6.
47. Langrish CL, Chen Y, Blumenschein WM, Mattson J, Basham B, Sedgwick JD, et al. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J Exp Med (2005) 201:233–40. doi:10.1084/jem.20041257
48. Collison LW, Vignali DA. Interleukin-35: odd one out or part of the family? Immunol Rev (2008) 226:248–62. doi:10.1111/j.1600-065X.2008.00704.x
49. International Study Group for Behçet’s Disease. Criteria for diagnosis of Behçet’s disease. International Study Group for Behçet’s Disease. Lancet (1990) 335:1078–80.
50. Glasel JA. Validity of nucleic acid purities monitored by 260nm/280nm absorbance ratios. Biotechniques (1995) 18:62–3.
51. Ding Q, Fan B, Fan Z, Ding L, Li F, Tu W, et al. Interleukin-10-819C >T polymorphism contributed to cancer risk: evidence from 29 studies. Cytokine (2012) 61:139–45. doi:10.1016/j.cyto.2012.09.008
52. Xue H, Wang YC, Lin B, An J, Chen L, Chen J, et al. A meta-analysis of interleukin-10 -592 promoter polymorphism associated with gastric cancer risk. PLoS One (2012) 7:e39868. doi:10.1371/journal.pone.0039868
53. Li D, He Q, Li R, Xu X, Chen B, Xie A. Interleukin-10 promoter polymorphisms in Chinese patients with Parkinson’s disease. Neurosci Lett (2012) 513:183–6. doi:10.1016/j.neulet.2012.02.033
54. Zhong Q, Ding C, Wang M, Sun Y, Xu Y. Interleukin-10 gene polymorphisms and chronic/aggressive periodontitis susceptibility: a meta-analysis based on 14 case-control studies. Cytokine (2012) 60:47–54. doi:10.1016/j.cyto.2012.05.014
55. Liang X, Zhang J, Zhu Y, Lu Y, Zhou X, Wang Z, et al. Specific genetic polymorphisms of IL10-592AA and IL10-819 TT genotypes lead to the key role for inducing docetaxel-induced liver injury in breast cancer patients. Clin Transl Oncol (2013) 15:331–4. doi:10.1007/s12094-012-0936-6
56. Temple SE, Lim E, Cheong KY, Almeida CA, Price P, Ardlie KG, et al. Alleles carried at positions -819 and -592 of the IL10 promoter affect transcription following stimulation of peripheral blood cells with Streptococcus pneumoniae. Immunogenetics (2003) 55:629–32. doi:10.1007/s00251-003-0621-6
57. Lin WP, Lin JH, Chen XW, Wu CY, Zhang LQ, Huang ZD, et al. Interleukin-10 promoter polymorphisms associated with susceptibility to lumbar disc degeneration in a Chinese cohort. Genet Mol Res (2011) 10:1719–27. doi:10.4238/vol10-3gmr1283
58. Tesse R, Del Vecchio GC, De Mattia D, Sangerardi M, Valente F, Giordano P. Association of interleukin-(IL)10 haplotypes and serum IL-10 levels in the progression of childhood immune thrombocytopenic purpura. Gene (2012) 505:53–6. doi:10.1016/j.gene.2012.05.050
59. Xavier JM, Shahram F, Davatchi F, Rosa A, Crespo J, Abdollahi BS, et al. Association study of IL10 and IL23R-IL12RB2 in Iranian patients with Behçet’s disease. Arthritis Rheum (2012) 64:2761–72. doi:10.1002/art.34437
60. Jang WC, Nam YH, Ahn YC, Lee SH, Park SH, Choe JY, et al. Interleukin-17F gene polymorphisms in Korean patients with Behcet’s disease. Rheumato Int (2008) 29:173–8. doi:10.1007/s00296-008-0664-y
61. Jiang Z, Yang P, Hou S, Du L, Xie L, Zhou H, et al. IL-23R gene confers susceptibility to Behcet’s disease in a Chinese Han population. Ann Rheum Dis (2010) 69:1325–8. doi:10.1136/ard.2009.119420
62. Kim ES, Kim SW, Moon CM, Park JJ, Kim TI, Kim WH, et al. Interactions between IL17A, IL23R, and STAT4 polymorphisms confer susceptibility to intestinal Behcet’s disease in Korean population. Life Sci (2012) 90:740–6. doi:10.1016/j.lfs.2012.03.017
63. Kastelein RA, Hunter CA, Cua DJ. Discovery and biology of IL-23 and IL-27: related but functionally distinct regulators of inflammation. Annu Rev Immunol (2007) 25:221–42. doi:10.1146/annurev.immunol.22.012703.104758
64. Paunovic V, Carroll HP, Vandenbroeck K, Gadina M. Signalling, inflammation and arthritis: crossed signals: the role of interleukin (IL)-12, -17, -23 and -27 in autoimmunity. Rheumatology (2008) 47:771–6. doi:10.1093/rheumatology/kem352
65. Hirschfield GM, Liu X, Xu C, Lu Y, Xie G, Lu Y, et al. Primary biliary cirrosis associated with HLA, IL12A and IL12RB2 variants. N Engl J Med (2009) 360:2544–55. doi:10.1056/NEJMoa0810440
66. Liu X, Invernizzi P, Lu Y, Kosoy R, Lu Y, Bianchi I, et al. Genome-wide meta-analyses identify three loci associated with primary biliary cirrosis. Nat Genet (2010) 42:658–60. doi:10.1038/ng.627
67. Mells GF, Floyd JA, Morley KI, Cordell HJ, Franklin CS, Shin SY, et al. Genome-wide association study identifies 12 new susceptibility loci for primary biliary cirrhosis. Nat Genet (2011) 43:329–32. doi:10.1038/ng.789
68. Park K, Kim N, Nam J, Bang D, Lee ES. Association of TNFA promoter region haplotype in Behçet’s disease. J Korean Med Sci (2006) 21:596–601. doi:10.3346/jkms.2006.21.4.596
69. Akman A, Sallakci N, Kacaroglu H, Tosun O, Yavuzer U, Alpsoy E, et al. Relationship between periodontal findings and the TNF-alpha gene 1031T/C polymorphism in Turkish patients with Behcet’s disease. J Eur Acad Dermatol Venereol (2008) 22:950–7. doi:10.1111/j.1468-3083.2008.02678.x
70. Orchard TR, Chua CN, Ahmad T, Cheng H, Welsh KI, Jewell DP. Uveitis and erythema nodosum in inflammatory bowel isease: clinical features and the role of HLA genes. Gastroenterology (2002) 123:714–8. doi:10.1053/gast.2002.35396
71. Higuchi T, Seki N, Kamizono S, Yamada A, Kimura A, Kato H, et al. Polymorphism of the 5’ flanking region of the human tumor necrosis factor (TNF)-alpha gene in Japanese. Tissue Antigens (1998) 51:605–12. doi:10.1111/j.1399-0039.1998.tb03002.x
72. Brinkman BM, Zuijdeest D, Kaijzel EL, Breedveld FC, Verweij CL. Relevance of the tumor necrosis factor alpha (TNF alpha) -308 promoter polymorphism in TNF alpha gene regulation. J Inflamm (1995) 46:32–41.
73. Uglialoro AM, Turbay D, Pesavento PA, Delgado JC, McKenzie FE, Gribben JG, et al. Identification of three new single nucleotide polymorphisms in the human tumor necrosis factor-alpha gene promoter. Tissue Antigens (1998) 52:359–67. doi:10.1111/j.1399-0039.1998.tb03056.x
74. Skoog T, van’t Hooft FM, Kallin B, Jovinge S, Boquist S, Nilsson J, et al. A common functional polymorphism (C – >A substitution at position -863) in the promoter region of the tumour necrosis factor-alpha (TNF-alpha) gene associated with reduced circulating levels of TNF-alpha. Hum Mol Genet (1999) 8:1443–9. doi:10.1093/hmg/8.8.1443
75. Kaijzel EL, Bayley JP, van Krugten MV, Smith L, van de Linde P, Bakker AM, et al. Allele-specific quantification of tumor necrosis factor alpha (TNF) transcription and the role of promoter polymorphisms in rheumatoid arthritis patients and healthy individuals. Genes Immun (2001) 2:135–44. doi:10.1038/sj.gene.6363747
Keywords: Behçet’s disease, genetic association, IL10, TNF-α, IL23R-IL12RB2, single nucleotide polymorphism, Western Algeria
Citation: Khaib Dit Naib O, Aribi M, Idder A, Chiali A, Sairi H, Touitou I, Lefranc G and Barat-Houari M (2013) Association analysis of IL10, TNF-α, and IL23R-IL12RB2 SNPs with Behçet’s disease risk in Western Algeria. Front. Immunol. 4:342. doi: 10.3389/fimmu.2013.00342
Received: 27 July 2013; Accepted: 06 October 2013;
Published online: 21 October 2013.
Edited by:
Giuseppe Alvise Ramirez, Università Vita-Salute San Raffaele, ItalyReviewed by:
Fulvio D’Acquisto, Queen Mary University of London, UKAngelo A. Manfredi, Università Vita-Salute San Raffaele, Italy
Copyright: © 2013 Khaib Dit Naib, Aribi, Idder, Chiali, Sairi, Touitou, Lefranc and Barat-Houari. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mourad Aribi, Laboratory of Applied Molecular Biology and Immunology, Abou-Bekr Belkaïd University of Tlemcen, Rocade # 2, Tlemcen, TLM 13000, Algeria e-mail: m_aribi@mail.univ-tlemcen.dz; m_aribi@yahoo.fr