 Enrique Balderas
Enrique Balderas Jin Zhang
Jin Zhang Enrico Stefani
Enrico Stefani Ligia Toro
Ligia Toro- 1Department of Anesthesiology, University of California, Los Angeles, Los Angeles, CA, USA
- 2Deparment of Molecular and Medical Pharmacology, University of California, Los Angeles, Los Angeles, CA, USA
- 3Department of Physiology, University of California, Los Angeles, Los Angeles, CA, USA
- 4Brain Research Institute, University of California, Los Angeles, Los Angeles, CA, USA
- 5Cardiovascular Research Laboratory, University of California, Los Angeles, Los Angeles, CA, USA
Since its discovery in a glioma cell line 15 years ago, mitochondrial BKCa channel (mitoBKCa) has been studied in brain cells and cardiomyocytes sharing general biophysical properties such as high K+ conductance (~300 pS), voltage-dependency and Ca2+-sensitivity. Main advances in deciphering the molecular composition of mitoBKCa have included establishing that it is encoded by the Kcnma1 gene, that a C-terminal splice insert confers mitoBKCa ability to be targeted to cardiac mitochondria, and evidence for its potential coassembly with β subunits. Notoriously, β1 subunit directly interacts with cytochrome c oxidase and mitoBKCa can be modulated by substrates of the respiratory chain. mitoBKCa channel has a central role in protecting the heart from ischemia, where pharmacological activation of the channel impacts the generation of reactive oxygen species and mitochondrial Ca2+ preventing cell death likely by impeding uncontrolled opening of the mitochondrial transition pore. Supporting this view, inhibition of mitoBKCa with Iberiotoxin, enhances cytochrome c release from glioma mitochondria. Many tantalizing questions remain open. Some of them are: how is mitoBKCa coupled to the respiratory chain? Does mitoBKCa play non-conduction roles in mitochondria physiology? Which are the functional partners of mitoBKCa? What are the roles of mitoBKCa in other cell types? Answers to these questions are essential to define the impact of mitoBKCa channel in mitochondria biology and disease.
Introduction
Mitochondria are key organelles defining cell fate and thus, much interest has developed in understanding the functional role of proteins present at its membranes. Mitochondria are shaped by an outer membrane, and an internal membrane that separates the intermembrane space and the matrix. The internal membrane is particularly important because it houses the respiratory chain protein complexes where reactive oxygen species and ATP are produced and also serves to delimit a Ca2+ storage space, making mitochondria not only producers of ATP but also regulators of Ca2+ and redox homeostasis. The list of proteins uncovered at the inner membrane keeps increasing, among them, K+ selective channels. A recent review by Szabo and Zoratti (2014) summarizes the evidence for the presence/role of: ATP-sensitive (KATP), small-conductance calcium-activated (SKCa), intermediate-conductance calcium-activated (IKCa), large-conductance, voltage and calcium-activated (BKCa), voltage-gated 1.3 (Kv1.3), two-pore domain acid-sensitive type 3 (TASK-3), and pH-sensitive K+channels. The subject of this review is the mammalian mitochondrial BKCa channel (mitoBKCa) that has sparked much interest primarily because of its role in protecting the heart from ischemic insult, as first demonstrated by Xu et al. (2002).
General Design of BKCa Channels
BKCa channels at the plasma membrane are characterized by having a large conductance, and by sensing changes in membrane potential and intracellular calcium (for a recent review see Contreras et al., 2013). Structure-function studies have ascribed these properties to distinct domains of the 7 transmembrane (S0–S7) α subunit -encoded by the Kcnma1 gene- that has an extracellular N-terminus and an intracellular C-terminus (Figure 1A). Four α subunits form a functional channel. The voltage sensing domain encompasses S0–S4 segments, the pore/gate domain includes S5–S6 and corresponding linker which lines the pore selectivity filter of the tetrameric channel, and the Ca2+sensing domain is located at the C-terminus. Pore residues located extracellularly comprise the receptor for pore blockers, Charybdotoxin (ChTx) and Iberiotoxin (IbTx) (Gao and Garcia, 2003; Banerjee et al., 2013). The intracellular C-terminus, which occupies two thirds of the whole protein, contains two regions that can sense Ca2+ known as the regulators of K+ conductance (RCK) 1 and 2. Mutagenesis studies have shown that RCK1 contains two critical aspartates (D362/D367) while RCK2 contains 5 consecutive aspartates in the “Ca2+ bowl” that together are sufficient for BKCa activation at physiological Ca2+ concentrations (Schreiber and Salkoff, 1997; Xia et al., 2002). However, recent crystal structures have only detected a single site of Ca2+ binding located in the “Ca2+ bowl” and utilizing two main-chain carbonyl oxygens of Q889 and D892 and side-chain carboxylate oxygens of D895 and D897 (underlined in 889QFLDQDDDDDPDT901) (Yuan et al., 2010, 2012). In addition to Ca2+, BKCa can also be activated by Mg2+ in the millimolar range. Interestingly, residues of distinct α subunits form part of the Mg2+ sensor, namely D99 and N172 from the voltage sensing domain of one subunit with E374 and E399 from the RCK1 domain of a different subunit (Shi et al., 2002; Yang et al., 2008).
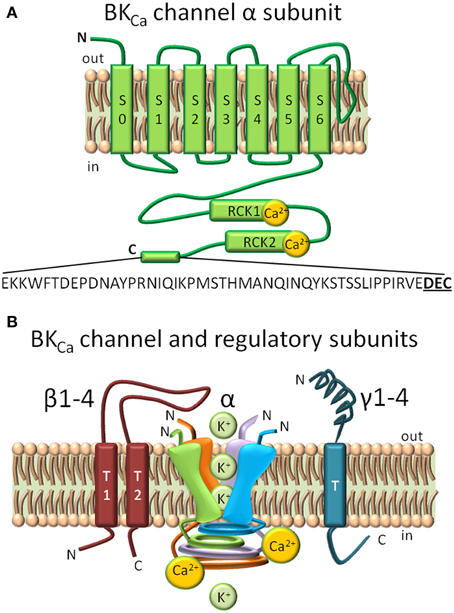
Figure 1. Structural domains in BKCa channels and regulatory subunits. (A) BKCa is composed by 7 transmembrane domains (S0–S7) and a long intracellular C-terminus. S0–S4 form the voltage sensing domain, and S5–S6 conform the pore-gating domain. Ca2+ biding sites are highlighted in the Regulator of Potassium Conductance (RCK) 1 and RCK2 domains. A C-terminal 50 amino acid splice insert, DEC, is highlighted. (B) Regulatory BKCa subunits. Homotetramer model of the pore-forming α subunit, the two spanning domain regulatory β subunits (1–4), and single spanning domain γ (1–4) subunits. The loop of β4 subunit confers to BKCa α subunit its resistance to toxin inhibition (Meera et al., 2000).
The Kcnma1 gene when transcribed can undergo extensive alternative splicing that give rise to multiple BKCa channel isoforms with varied functional characteristics including voltage/Ca2+ sensitivities, response to phosphorylation and arachidonic acid modulation, and subcellular localizations, including targeting to mitochondria as discussed later in this review (Saito et al., 1997; Tian et al., 2001; Zarei et al., 2004; Ma et al., 2007; Li et al., 2010; Singh et al., 2013).
BKCa channel functional heterogeneity is further increased by its association with modulatory β (1–4) or γ (Yan and Aldrich, 2010, 2012) subunits (Figure 1B) that are mostly tissue-specific and greatly modify functional and pharmacological characteristics like kinetics, Ca2+/V sensitivities, and toxin blockade (Knaus et al., 1994; Wallner et al., 1999; Brenner et al., 2000; Meera et al., 2000; Uebele et al., 2000). Beta subunits (β1,β2,β4) can also act as modulators of channel density at the plasma membrane via endocytic processes (Toro et al., 2006; Zarei et al., 2007; Shruti et al., 2012; Cox et al., 2014). The relatively high tissue specificity of β subunits make them key in defining the function of BKCa channels in different organs, for example the abundance of β1 subunit in smooth muscle make them essential to maintain a healthy vascular tone.
Posttranslational modifications like lipidation and phosphorylation add another regulatory layer to BKCa function. For example, in the α subunit, internal myristoylation at N-terminal intracellular loops slows down the activation kinetics of BKCa channel and reduces its cell surface expression promoting endocytosis via a clathrin mediated mechanism (Alioua et al., 2011). Palmitoylation at S0–S1 linker also reduces cell surface expression, and at the spliced exon STREX (inserted at the C-terminus of BKCa) produces channels resistant to protein kinase C induced inhibition (Jeffries et al., 2010; Zhou et al., 2012). With respect to β subunits, juxtamembrane palmitoylation of the β4 subunit at its C-terminus promotes surface expression of BKCa α subunits but only when the latter contain a 50 amino acid C-terminal splice insert, named DEC (Chen et al., 2013). Most of the above findings have been obtained in heterologous expression systems; the next challenge is to define the physiological or pathophysiological impact that these processes have in different organs of the body.
Discovery of BKCa Channel Activity at the Inner Mitochondrial Membrane and Biophysical Properties
The first evidence showing that BKCa channel with a conductance of about 300 pS (in 150 mM KCl) was present at the inner mitochondrial membrane was given by Siemen and coworkers in the late 90's; the channel was characterized using mitochondria devoid of external membranes (mitoplasts) of the glioma cell line, LN-229, and the patch clamp technique (Siemen et al., 1999). Since then, mitochondrial BKCa channels (mitoBKCa) with similar conductances ranging from 200 to 307 pS have been detected in other systems using channel reconstitution in lipid bilayers or by patch clamping mitoplasts. A single report shows a 564 pS channel in mitochondria of brain (Table 1).
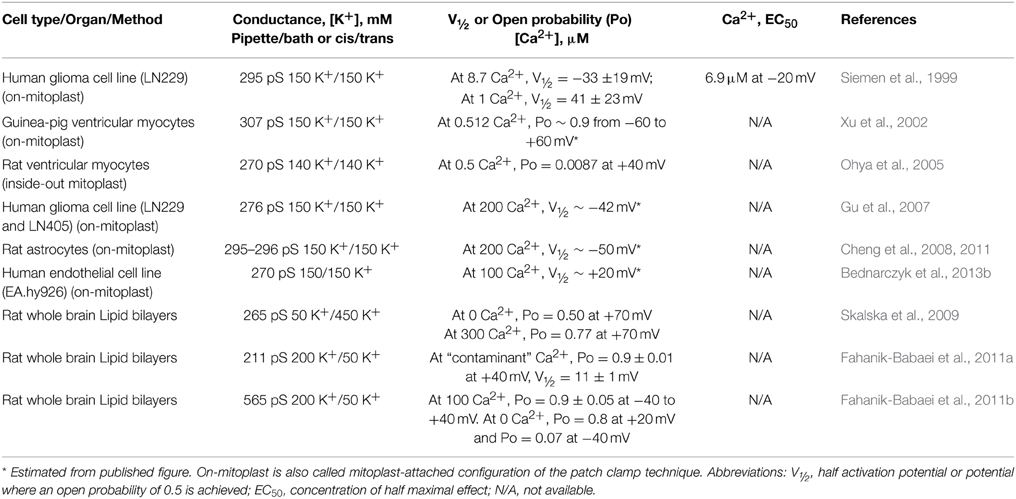
Table 1. mitoBKCa biophysical properties in mammals.
We now know that mitoBKCa and plasma membrane BKCa channel pore-forming α subunits are encoded by the same gene (Kcnma1) (Singh et al., 2013) explaining why they share common basic biophysical properties including a large conductance, and being responsive to voltage and Ca2+, although specific values may vary. A comparison of plasma membrane BKCa and mitoBKCa properties in a human glioma cell line (LN 229) shows that the conductance of the former was 199 ± 8 pS and of the latter was 278 ± 10 pS. Although both channels were voltage/Ca2+ dependent, their sensitivities were different. In the inside-out configuration, plasma membrane BKCa displayed a low sensitivity to voltage as it displayed a low open probability (Po) even at high potentials (Po < 0.1 at +80 mV and ~0.4 at 100 mV) and 400 μM Ca2+ facing the cytosolic side of the channel. mitoBKCa, on the other hand, recorded on the on-mitoplast configuration and the same Ca2+ in the bath solution only needed a depolarization to −40 mV to reach a Po of ~0.6 (Gu et al., 2014). Although in this configuration the precise Ca2+ concentration at the matrix side of the channel is difficult to establish, the authors showed that by decreasing Ca2+ in the bath to “zero” the channel Po decreased to ≤0.05 (see below for discussion of mitoBKCa orientation). Thus, assuming that both classes of channels could sense the same Ca2+ concentration, one possible explanation to these differences is that multiple BKCa isoforms exist, both at the plasma membrane and in mitochondria, which could result from a combination of factors including splice variation, association with auxiliary subunits or posttranslational modifications affecting how they respond to voltage and Ca2+.
Along the above point of view, Table 1 shows that different cell types appear to express mitoBKCa channels with varied voltage and Ca2+ sensitivities. For example, the cardiac mitoBKCa channel from guinea pig had a particularly high Po of ~0.9 within a large voltage range (−60 to +60 mV) at 0.5 μM [Ca2+] suggesting that its molecular composition (e.g., association with auxiliary subunits) may be substantially distinct from that expressed in glioma mitochondria which at 1 μM Ca2+ displays a Po of 0.5 at +41 mV (half activation potential, V1/2 = 41 mV) (note that in both cases, experiments were performed in the on-mitoplast mode and Ca2+ was changed in the bath solution; thus, the exact Ca2+ concentration in the matrix side is unknown). This variability is not exclusive of mitoBKCa as different isoforms are also detected at the plasma membrane even within the same cell type. For example, BKCa channels from coronary smooth muscle can display a predominant V½ of −98 mV but also −66 mV, −38 mV, −16 mV and 21 mV (at 18 μM Ca2+) (Tanaka et al., 1997) that could be explained by α + β1 channels with different β1 subunit stoichiometry; while skeletal muscle BKCa single channels can have four-fold differences in K0.5 for Ca2+ with an average of 14 ± 7 μM at +30 mV (McManus and Magleby, 1991).
Obviously, a detailed biophysical and molecular characterization of mitoBKCais needed for each cell type to be able to understand the basis of their function.
Orientation of mitoBKCa; is the Ca2+ Sensor Facing the Mitochondrial Matrix?
Siemen et al. (1999) showed in on-mitoplast patches that increasing Ca2+ in the bath increased channel Po; the EC50 for Ca2+ measured at +60 mV was ~0.9 μM. The usage of a Ca2+ ionophore was dispensable, and thus, it was assumed that the normal Ca2+ import mitochondrial mechanisms were sufficient to increase matrix Ca2+ in the vicinity of mitoBKCa Ca2+sensor. In addition, mitoBKCa was blocked by ChTx (the toxin binds plasma membrane BKCa at the extracellular pore vestibule) applied to the patch pipette. ChTx displayed an EC50 of ~1.5 nM and a Hill coefficient of 1.2 consistent with the sensitivity reported for plasma membrane BKCa channels formed by α or α + β1 subunits (Meera et al., 2000). These results supported the idea that mitoBKCa ChTx receptor (in the pore vestibule) is facing the mitochondrial intermembrane space and thus, the Ca2+ sensor (located in the opposite side of BKCa protein) is facing the mitochondrial matrix.
Few years later Xu et al. (2002) investigated the presence of mitoBKCa in ventricular mitoplasts from guinea pig also using on-mitoplast patches and isotonic 150 mM KCl. Ventricular myocytes contained mitochondria rich in mitoBKCa whose conductance (~300 pS) was similar to that observed in the glioma cells (295 pS) (Table 1). Ventricular mitoBKCa activity was completely abolished by 200 nM ChTx applied to the patch pipette and the ensemble average patch currents augmented by increasing bath Ca2+ from 0.5 to 40 μM. In this case, the usage of a Ca2+ ionophore was also dispensable. Assuming that matrix Ca2+ was in equilibrium with the bath solution (aided by native Ca2+ import mechanisms), the results further supported the picture of mitoBKCa with its ChTx receptor facing the intermitochondrial space and thus, its Ca2+ sensor facing the matrix side.
Other studies have supported the orientation of mitoBKCa as being the pore vestibule facing the mitochondrial intermembrane space and the Ca2+ sensor in the matrix. Direct evidence using mitoplasts: (1) In the human glioma cell line LN-229, on-mitoplast patches showed an increased channel Po with matrix depolarization and exposing the external side of the mitoplast membrane to ChTx (using the whole-mitoplast or outside-out configurations and applying ChTx to the bath) reduced mitoBKCa activity (Gu et al., 2007, 2014); (2) In rat astrocyte mitoplasts, IbTx added to the pipette in the on-mitoplast configuration reduced mitoBKCa activity (Cheng et al., 2008), while depolarization of the inner side of the mitoplast (matrix side) promoted increased channel activity (Cheng et al., 2011). Indirect evidence using intact rat brain mitochondria: Ca2+-induced depolarization was prevented by IbTx (50 nM) and ChTx (200 nM) (Skalska et al., 2009). Because the outer mitochondrial membrane is permeable to proteins up to 5 kDa and IbTx as well as ChTx are ~4.2 kDa, both toxins are expected to traverse the outer membrane and reach the channel pore facing the intermembrane space; this arrangement would place the C-terminus facing the matrix.
The evidence supporting an opposite orientation, i.e., a C-terminus facing the intermembrane space is less clear. Recent recordings in inside-out mitoplast patches from a human astrocytoma cell line (U-87 MG) show a mitoBKCa channel that was activated by hyperpolarization of the matrix side (positive potentials applied to the pipette interior) and blocked by IbTx also applied to the matrix side (bath solution) (Bednarczyk et al., 2013a). Assuming that this channel is encoded by the Kcnma1 gene (encoding plasma membrane BKCa) (see below Singh et al., 2013), the above results would imply a pore vestibule facing the matrix and a C-terminus facing the intermembrane space. However, decreasing matrix Ca2+ decreased channel activity making the authors suggest that both IbTx and Ca2+ binding sites were probably facing the mitochondrial matrix. This hypothesis would support the existence of a mitoBKCa channel unrelated to the plasma membrane BKCa, where the pore vestibule and the Ca2+ sensor are facing opposite sides of the plasma membrane (see Figure 1). It is evident that further studies on the orientation of mitoBKCa channel at the inner mitochondrial membrane are needed.
mitoBKCa Molecular Origin
mitoBKCa molecular origin has been recently defined as the Kcnma1 gene, which also encodes plasma membrane BKCa (Singh et al., 2013). Utilizing an exon-scanning RT-PCR strategy of the mouse Kcnma1 gene and ventriculocyte mRNAs (these cardiac cells were an excellent system for this quest since they are characterized by their lack of BKCa channel activity at the plasma membrane) Singh et al., found transcript expression of 3 alternatively spliced exons, STREX, SV27 (27 amino acid insert) and the C-terminal 50 amino acid “DEC” exon. Notoriously, the amount of DEC exon transcripts equaled that of the total BKCa predicting a mitoBKCa tetrameric structure constituted by two subunits of BKCa variant containing DEC insert. Indeed, expression of BKCa constructs containing the DEC exon was sufficient for BKCa targeting to mitochondria in adult ventriculocytes. Mass spectrometry analysis and functional data using BKCa knockout mice further confirmed mitoBKCa as being encoded by the Kcnma1 gene.
Interestingly, the ability of DEC exon for targeting mitoBKCa to mitochondria may vary depending on the cell type and/or its association with other subunits. In fact, expression in CHO cells of a BKCa-DEC variant (containing additional alternatively spliced exons including SV27) cloned from hair cells showed prominent expression in Mitotracker labeled mitochondria but also at the cell periphery (Kathiresan et al., 2009). In contrast, expression of a DEC variant in COS-cells yielded signals confined to the endoplasmic reticulum (Ma et al., 2007). Thus, the DEC sequence may not be the only factor defining the targeting of BKCa-DEC to mitochondria in all cell types but cell-specific mechanisms may exist that facilitate its mitochondrial delivery.
mitoBKCa and Regulatory Subunits
BKCa regulatory β subunits are expressed in mitochondria from heart, skeletal muscle, endothelial cells and brain as discussed below. Whether BKCa γ subunits exist in mitochondria is still unknown.
In the heart, β1 subunit was found in mitoplasts isolated from rat ventricular myocytes and a two hybrid system showed its direct interaction with cytochrome c oxidase subunit I (Ohya et al., 2005). These results demonstrated the localization of β1 in the mitochondrial inner membrane and associated with a component of the respiratory chain. Furthermore, functional experiments showed that in rat ventricular mitoplasts mitoBKCa activity was enhanced by stimulation with 30 μM estradiol in the presence of 500 nM Ca2+ (Ohya et al., 2005). Under these conditions, estradiol is known to activate plasma membrane BKCa channels in complex with β 1 subunits (Valverde et al., 1999). Interestingly, in mitoplasts of astrocytes the stimulatory effect of 30 μM β-estradiol on mitoBKCa was transient resulting in a final inhibition of channel activity that was resilient to a second β-estradiol stimulus and was only partially restored by increasing Ca2+ in the matrix side of inside-out patches (Thiede et al., 2012).
In cultured pulmonary artery smooth muscle, β 1 subunit was found to play a role in 11,12-epoxyeicosatrienoic acid induced depolarization of mitochondrial membrane potential as this effect was abrogated in the β1−/− animal (Loot et al., 2012).
In a human endothelial cell line, β2 was detected in lysates of mitochondria and mitoplasts. However, this subunit seems not to be forming complex with mitoBKCa in this cell line because the reported channel activity (Bednarczyk et al., 2013b) does not show classical time-dependent inactivation conferred by β2 (Wallner et al., 1999; Benzinger et al., 2006).
In rat soleus muscle, β4 is visualized both at the plasma membrane and in mitochondria using immunocytochemistry; and by immunoblotting as a protein of ~26 kDa using purified mitochondria (Skalska et al., 2008).
In the brain, β4 subunit is clearly localized to mitochondria of the gigantocellular reticular nucleus and in mitochondria of the pons. Because β 4 could not be proteolyzed in intact mitochondria but only after detergent solubilization, it was speculated that it must be internal to the outer mitochondrial membrane. Western blot analysis of brain homogenates and mitochondrial fractions further confirmed the presence of β4 in mitochondria. Interestingly, β2 was also found in brain mitochondrial fractions but not β1 nor β3 subunits (Piwonska et al., 2008). β4 has also been observed in mitochondria of rat hippocampal neurons by immunochemistry and as a protein of ~26 kDa in mitochondrial lysates of whole brain (Skalska et al., 2009). Thus, it is possible that β4 forms part of the mitoBKCa complex at the inner mitochondrial membrane of various types of neurons.
β4 subunit expression in the glia appears to be disease dependent as it was not found expressed in glia of adult normal brain (Piwonska et al., 2008) but it has been detected in mitochondria of a human gliobastoma cell line (U-87 MG) where it comigrates with cytochrome c oxidase subunit I (Bednarczyk et al., 2013a). Yet, the functional impact that β4 may have on mitoBKCa activity in glioma cells and in neurons needs to be established.
From the above studies it appears that mitochondrial β subunits show tissue specificity, a characteristic of their plasma membrane counterparts. However, more studies are needed to assess this hypothesis or to establish their functional role in mitochondria. Gene silencing models could be ideal for this task.
mitoBKCa Channel and Protection from Ischemic Insult
mitoBKCachannel was first related to cardiac protection from global ischemia and reperfusion injury by Xu et al. (2002) using a BKCa opener, NS1619 (10–30 μM). The drug used to precondition the heart prior ischemia and reperfusion, improved left ventricular developed pressure and decreased infarct size. Both effects were abolished with 1 μM Paxilline, an inhibitor of BKCa. Several factors support the notion that NS1619 was opening BKCa located in mitochondria: (a) the fact that NS1619 could not be targeting plasma membrane BKCa, as adult cardiomyocytes are known for their lack of sarcolemmal BKCa expression/activity (Singh et al., 2013; Schmitt et al., 2014); (b) mitochondrial K+ uptake was accelerated by NS1619 and decelerated by blocking BKCa with 100 nM IbTx; and (c) the protective effect of preconditioning the heart with NS1619 on reperfusion was not related to relaxation of the vasculature, where BKCa channel is abundant. Since then, other groups have confirmed and expanded these results (Table 2).
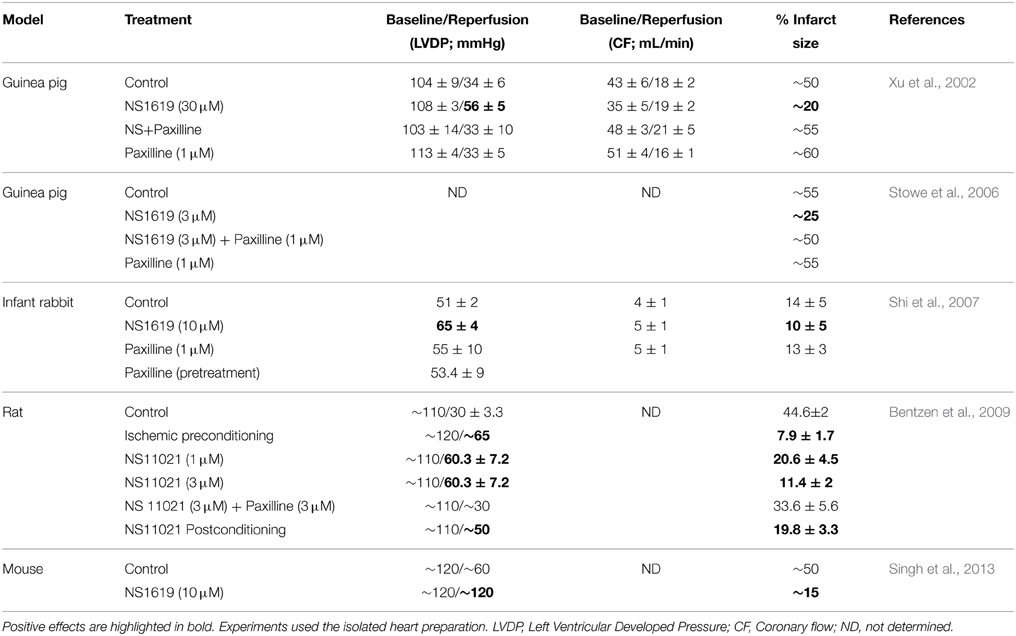
Table 2. BKCa channel agonists and cardioprotection.
Stowe et al. (2006) confirmed an improved left ventricular developed pressure by 3 μM NS1619 preconditioning and measured mitochondrial Ca2+ and ROS production. In situ recording of these parameters during the ischemia/reperfusion protocol (in the left ventricle of the isolated, perfused heart) demonstrated that both parameters decreased by NS1619 preconditioning both during the ischemia period and ~10 min after reperfusion started.
The cardioprotective effect of BKCa activation, in improving left ventricular developed pressure and reducing infarct size, has also been observed with BKCa opener NS11021 (1–3 μM) (Bentzen et al., 2009) and naringenin (4 μM) (Testai et al., 2013). Importantly, NS11021 is also effective when hearts or isolated cells are treated postischemia or postmetabolic inhibition and during reperfusion or re-energization, respectively (Bentzen et al., 2009; Borchert et al., 2013). These findings make BKCa an excellent target to improve cardiac function after an ischemic event as it occurs during heart infarct.
Interestingly, the protective effect of NS1619 may not be mediated by mitoBKCa in all systems like in primary rat cortical neurons. In this type of neurons, preconditioning with NS1619 (150 μM) caused mitochondrial depolarization (consistent with K+ influx) but this depolarization was not prevented by 5 min preincubation with 20 μM Paxilline (Gaspar et al., 2009). An alternative explanation to this negative result could be that Paxilline needs longer time to diffuse through the plasma membrane and reach mitoBKCa at the mitochondrial inner membrane.
Although BKCa channel activity is indeed inhibited by Paxilline (Zhou and Lingle, 2014), its exclusive use as pharmacological indicator of BKCa functional role in mitochondria or elsewhere needs to be taken with caution. This assertion is underscored by recent studies, where isoflurane preconditioning protected the heart from ischemic insult, showing that Paxilline (1 μM) abolished the anesthetic cardioprotective effect equally well in wild type and Kcnma1−/− mice (Wojtovich et al., 2011, 2013).
It is also important to highlight that the specific action of NS1619 on BKCa channel activation has been questioned (Szewczyk et al., 2010). Concentrations of NS1619 above 10 μM inhibited SERCA with a consequent Ca2+ overload in sarcoplasmic reticulum and posterior cytosolic contamination (Wrzosek, 2014). Other authors also reported non-specific effects of NS1619 at concentrations ≥10 μM such as inhibition of mitochondrial respiratory chain (~15–30% inhibition at 10 μM) (Kicinska and Szewczyk, 2004; Cancherini et al., 2007), and H+/K+ leak with 50–100 μM of the drug (Aldakkak et al., 2010). Therefore, the use of low concentrations of NS1919 (i.e., ≤10 μM) in conjunction with the use of genetically modified models is desirable.
Conclusive evidence for the role of BKCa in cardioprotection has come from studies using BKCa knockout mouse models (Kcnma1−/−) whose hearts are not protected from ischemic injury by NS1619 (5–10 μM) or NS11021 (500 nM) (Singh et al., 2013; Wojtovich et al., 2013), or by ischemic preconditioning (Soltysinska et al., 2014) as revealed by measurements of heart function and infarct size in isolated perfused hearts. Mitochondrial BKCa of cardiomyocytes (Singh et al., 2013; Soltysinska et al., 2014) as well as BKCa expressed in cardiac neurons (Wojtovich et al., 2013) contribute to the cardioprotective effects. A role of BKCa expressed in cardiac neurons is supported by the fact that an inhibitor of neural transmission prevents protection by NS1619 (Wojtovich et al., 2013). Supporting a role for mitoBKCa are measurements in isolated mitochondria at 10 min of reperfusion showing improved Ca2+ retention capacity with NS1619 preconditioning a property that was absent in the knockout animal (Singh et al., 2013). In addition, a model of anoxia/reoxygenation of isolated mitochondria (mimicking the isolated heart ischemia/reperfusion model) showed that the cardioprotective effect of preconditioning could be related to a mitoBKCa-mediated decrease of ROS production, as the production of ROS postanoxia was higher in the KO animal (Soltysinska et al., 2014).
mitoBKCa and ROS Production
The knockout animal studies described above point to mitoBKCa expression and ROS reduction as mitochondrial mechanisms playing a role in heart protection by ischemic preconditioning. The role of mitoBKCa activation in the regulation of ROS production has also been examined in heart mitochondria under basal conditions and has been found to depend on the ROS producing conditions. In succinate energized mitochondria, where ROS production at site I is enhanced by reversed electron flow, putative stimulation of mitoBKCa channel with 30 μM NS1619 produced a profound reduction of H2O2 production rate. Further, this decrease was partially overcome by pretreatment with 5 μM Paxilline (Heinen et al., 2007a). In contrast, when reverse electron flow was blocked by rotenone (i.e., succinate + rotenone), NS1619 increased H2O2 production rate and this effect was abolished by Paxilline (Heinen et al., 2007b). A decrease in ROS production by stimulating ROS production at complex I of the respiratory chain was also observed in brain mitochondria where activating mitoBKCa channel either with 10 μM CGS7184 or 3 μM NS1619 reduced H2O2 production mediated by malate-glutamate or succinate. The CGS7184-mediated decrease in ROS production was abolished by 50 nM IbTx supporting the involvement of mitoBKCa (Kulawiak et al., 2008). Experiments using the knockout models should provide definite proof of mitoBKCa role in the maintenance of normal levels of ROS to impede cell damage.
mitoBKCa Channel and Mitochondrial Transition Pore (mPTP)
Under stress conditions, opening of mPTP allows the passage of small molecules with osmotic consequences for the cell like swelling and rupture of the mitochondrial outer membrane that permits liberation of apoptotic factors including cytochrome c causing cell death. The key elements responsible to keep mPTP closed are the mitochondrial Ca2+ content and ROS levels. Interestingly, mitoBKCa activity has been related to both ROS production as described above and to the regulation of mitochondrial Ca2+ content. Direct measurement of Ca2+ retention capacity (CRC) of cardiac mitochondria revealed that the protective effect of NS1619 preconditioning (increased CRC) was absent in the BKCa KO animal (Singh et al., 2013). These results imply that opening of BKCa to certain extent can protect mitochondria from uncontrolled mPTP opening. In line with this view, the proapoptotic protein Bax (Bcl-2 associated protein X) directly inhibits mitoBKCa activity in astrocyte mitoplasts but does not by itself produce mPTP electrical activity; and thus, Bax has been proposed to produce mPTP opening via inhibition of mitoBKCa (Cheng et al., 2011). In fact, inhibition of mitoBKCa with IbTx reduces the amount of Ca2+ necessary to depolarize brain mitochondria (a measure of increased mPTP activity) (Cheng et al., 2008) and increases cytochrome c release (from GL261 glioma mitochondria), a landmark of mPTP opening and apoptosis (Cheng et al., 2011).
From the above studies, it is evident that there is a physiological coupling between mitoBKCa and mPTP. Recent evidence indicates that mPTP is formed by dimers of the ATP-synthase (Giorgio et al., 2013). It would be interesting to test the hypothesis of a physical interaction between mitoBKCa and the ATP-synthase. The functional coupling of mitoBKCa with other components of the respiratory chain has been highlighted by the fact that mitoBKCa activity is affected by substrates of the respiratory chain in human astrocytoma U-87 MG cells (Bednarczyk et al., 2013a). The precise mechanisms of this coupling remain to be elucidated.
Proposed Mechanisms of mitoBKCa Channel Regulation of Common Pathological Conditions
In mitochondria isolated from brain of diabetic rats, a K+ channel of 46 pS conductance that is inhibited by IbTx, a specific blocker of BKCa, has been identified. This channel is voltage dependent with an effective valence of ~4.7 in contrast to ~2.6 of mitoBKCa from healthy brain (Noursadeghi et al., 2014). At present, it is difficult to ascribe a specific mechanism to this molecular switch. One possibility is that a subconductance of mitoBKCa becomes much more stable under diabetic conditions inasmuch a ~50 pS subconductance state of mitoBKCa has been reported in cardiac mitoplasts (Xu et al., 2002).
Cancerous tumor cells are highly resistant to hypoxia suggesting the presence of a mechanism(s) that prevents their death. mitoBKCa channels are expressed in cancerous cell lines (Siemen et al., 1999) and could participate in such mechanism as they are activated by hypoxia in mitoplasts derived from human glioma LN-229 cells and astrocytes (Gu et al., 2007, 2014; Cheng et al., 2008). Notably, hypoxia also reduces mPTP electrical activity in mitoplasts of liver mitochondria and delays mPTP opening in intact liver mitochondria as measured by Ca2+-induced membrane depolarization (Cheng et al., 2008). How would mitoBKCa sense oxygen levels during hypoxia? It is possible that chronic hypoxia-induced heme oxygenase-1 binds to the mitochondrial channel much like heme oxygenase-2 does with plasma membrane BKCa (Williams et al., 2004) promoting CO production and activation of mitoBKCa.
Supporting the view that mitoBKCa may regulate mitochondrial function as a redox sensor is the fact that its electrical activity can be inhibited by 300 nM hemin, a byproduct of hemoglobin (Augustynek et al., 2014) with oxidative properties that increases drastically (~10 mM) during hemolysis like the one that occurs during hemorraghic stroke.
Concluding Remarks and Perspectives
As a general conclusion, we can state that BKCa channel is present in the inner mitochondrial membrane of various cell types and in different species. The discovery that mitoBKCa is encoded by the same gene as the plasma membrane BKCa (Kcnma1) is just the beginning in our understanding of the molecular composition of mitoBKCa isoforms in different tissues that seem to display a spectrum of biophysical characteristics just like the plasma membrane channel. A detailed biophysical characterization in native mitochondria and in genetically engineered organelles should help in correlating distinct properties with molecular composition.
An immediate specific question to resolve is, for example, the orientation of mitoBKCa in the inner mitochondrial membrane. Experiments in inside-out patches where matrix and intermembrane spaces can have defined Ca2+ concentrations should solve this problem. Moreover, they will allow defining with certainty V1/2 and Ca2+ EC50 values for mitoBKCa in each cell type. At present, most of the experiments have been done in the on-mitoplast configuration and this information is lacking. Other points to address are: whether the BKCa γ subunit is present in mitochondria and if it forms part of mitoBKCa complex, or if β subunits have alternative functions in mitochondria.
Many other questions remain to be solved such as the physiological/pathophysiological role of mitoBKCa channel in different tissues, its subunit composition in different cell types, mechanisms of mitochondrial targeting, and interaction with mitochondrial protein complexes.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by NIH R01 HL107418 (ES and LT), UC MEXUS-CONACYT Postdoctoral Fellowship FE 13-248 (EB) and AHA-WSA Postdoctoral Fellowship 15POST22490015 (EB).
References
Aldakkak, M., Stowe, D. F., Cheng, Q., Kwok, W. M., and Camara, A. K. (2010). Mitochondrial matrix K+ flux independent of large-conductance Ca2+-activated K+ channel opening. Am. J. Physiol. Cell. Physiol. 298, C530–C541. doi: 10.1152/ajpcell.00468.2009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Alioua, A., Li, M., Wu, Y., Stefani, E., and Toro, L. (2011). Unconventional myristoylation of large-conductance Ca(2)-activated K channel (Slo1) via serine/threonine residues regulates channel surface expression. Proc. Natl. Acad. Sci. U.S.A. 108, 10744–10749. doi: 10.1073/pnas.1008863108
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Augustynek, B., Kudin, A. P., Bednarczyk, P., Szewczyk, A., and Kunz, W. S. (2014). Hemin inhibits the large conductance potassium channel in brain mitochondria: a putative novel mechanism of neurodegeneration. Exp. Neurol. 257, 70–75. doi: 10.1016/j.expneurol.2014.04.022
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Banerjee, A., Lee, A., Campbell, E., and MacKinnon, R. (2013). Structure of a pore-blocking toxin in complex with a eukaryotic voltage-dependent K(+) channel. Elife 2:e00594. doi: 10.7554/eLife.00594
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bednarczyk, P., Koziel, A., Jarmuszkiewicz, W., and Szewczyk, A. (2013b). Large-conductance Ca(2)(+)-activated potassium channel in mitochondria of endothelial EA.hy926 cells. Am. J. Physiol. Heart Circ. Physiol. 304, H1415–H1427. doi: 10.1152/ajpheart.00976.2012
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bednarczyk, P., Wieckowski, M. R., Broszkiewicz, M., Skowronek, K., Siemen, D., and Szewczyk, A. (2013a). Putative structural and functional coupling of the mitochondrial BK channel to the respiratory chain. PLoS ONE 8:e68125. doi: 10.1371/journal.pone.0068125
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bentzen, B. H., Osadchii, O., Jespersen, T., Hansen, R. S., Olesen, S. P., and Grunnet, M. (2009). Activation of big conductance Ca(2+)-activated K(+) channels (BK) protects the heart against ischemia-reperfusion injury. Pflug. Arch. 457, 979–988. doi: 10.1007/s00424-008-0583-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Benzinger, G. R., Xia, X. M., and Lingle, C. J. (2006). Direct observation of a preinactivated, open state in BK channels with beta2 subunits. J. Gen. Physiol. 127, 119–131. doi: 10.1085/jgp.200509425
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Borchert, G. H., Hlavackova, M., and Kolar, F. (2013). Pharmacological activation of mitochondrial BK(Ca) channels protects isolated cardiomyocytes against simulated reperfusion-induced injury. Exp. Biol. Med. (Maywood.) 238, 233–241. doi: 10.1177/1535370212474596
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brenner, R., Jegla, T. J., Wickenden, A., Liu, Y., and Aldrich, R. W. (2000). Cloning and functional characterization of novel large conductance calcium-activated potassium channel beta subunits, hKCNMB3 and hKCNMB4. J. Biol. Chem. 275, 6453–6461. doi: 10.1074/jbc.275.9.6453
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cancherini, D. V., Queliconi, B. B., and Kowaltowski, A. J. (2007). Pharmacological and physiological stimuli do not promote Ca(2+)-sensitive K+ channel activity in isolated heart mitochondria. Cardiovasc. Res. 73, 720–728. doi: 10.1016/j.cardiores.2006.11.035
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, L., Bi, D., Tian, L., McClafferty, H., Steeb, F., Ruth, P., et al. (2013). Palmitoylation of the beta4-subunit regulates surface expression of large conductance calcium-activated potassium channel splice variants. J. Biol. Chem. 288, 13136–13144. doi: 10.1074/jbc.M113.461830
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cheng, Y., Gu, X. Q., Bednarczyk, P., Wiedemann, F. R., Haddad, G. G., and Siemen, D. (2008). Hypoxia increases activity of the BK-channel in the inner mitochondrial membrane and reduces activity of the permeability transition pore. Cell. Physiol. Biochem. 22, 127–136. doi: 10.1159/000149790
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cheng, Y., Gulbins, E., and Siemen, D. (2011). Activation of the permeability transition pore by Bax via inhibition of the mitochondrial BK channel. Cell. Physiol. Biochem. 27, 191–200. doi: 10.1159/000327944
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Contreras, G. F., Castillo, K., Enrique, N., Carrasquel-Ursulaez, W., Castillo, J. P., Milesi, V., et al. (2013). A BK (Slo1) channel journey from molecule to physiology. Channels (Austin.) 7, 442–458. doi: 10.4161/chan.26242
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cox, N., Toro, B., Pacheco-Otalora, L. F., Garrido-Sanabria, E. R., and Zarei, M. M. (2014). An endoplasmic reticulum trafficking signal regulates surface expression of beta4 subunit of a voltage- and Ca(2)(+)-activated K(+) channel. Brain Res. 1553, 12–23. doi: 10.1016/j.brainres.2014.01.028
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fahanik-Babaei, J., Eliassi, A., Jafari, A., Sauve, R., Salari, S., and Saghiri, R. (2011a). Electro-pharmacological profile of a mitochondrial inner membrane big-potassium channel from rat brain. Biochim. Biophys. Acta 1808, 454–460. doi: 10.1016/j.bbamem.2010.10.005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fahanik-Babaei, J., Eliassi, A., and Saghiri, R. (2011b). How many types of large conductance Ca(+)(2)-activated potassium channels exist in brain mitochondrial inner membrane: evidence for a new mitochondrial large conductance Ca(2)(+)-activated potassium channel in brain mitochondria. Neuroscience 199, 125–132. doi: 10.1016/j.neuroscience.2011.09.055
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gao, Y. D., and Garcia, M. L. (2003). Interaction of agitoxin2, charybdotoxin, and iberiotoxin with potassium channels: selectivity between voltage-gated and Maxi-K channels. Proteins 52, 146–154. doi: 10.1002/prot.10341
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gaspar, T., Domoki, F., Lenti, L., Katakam, P. V., Snipes, J. A., Bari, F., et al. (2009). Immediate neuronal preconditioning by NS1619. Brain Res. 1285, 196–207. doi: 10.1016/j.brainres.2009.06.008
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Giorgio, V., von Stockum, S., Antoniel, M., Fabbro, A., Fogolari, F., Forte, M., et al. (2013). Dimers of mitochondrial ATP synthase form the permeability transition pore. Proc. Natl. Acad. Sci. U.S.A. 110, 5887–5892. doi: 10.1073/pnas.1217823110
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gu, X. Q., Pamenter, M. E., Siemen, D., Sun, X., and Haddad, G. G. (2014). Mitochondrial but not plasmalemmal BK channels are hypoxia-sensitive in human glioma. Glia 62, 504–513. doi: 10.1002/glia.22620
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gu, X. Q., Siemen, D., Parvez, S., Cheng, Y., Xue, J., Zhou, D., et al. (2007). Hypoxia increases BK channel activity in the inner mitochondrial membrane. Biochem. Biophys. Res. Commun. 358, 311–316. doi: 10.1016/j.bbrc.2007.04.110
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heinen, A., Aldakkak, M., Stowe, D. F., Rhodes, S. S., Riess, M. L., Varadarajan, S. G., et al. (2007a). Reverse electron flow-induced ROS production is attenuated by activation of mitochondrial Ca2+-sensitive K+ channels. Am. J. Physiol. Heart Circ. Physiol. 293, H1400–H1407. doi: 10.1152/ajpheart.00198.2007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heinen, A., Camara, A. K., Aldakkak, M., Rhodes, S. S., Riess, M. L., and Stowe, D. F. (2007b). Mitochondrial Ca2+-induced K+ influx increases respiration and enhances ROS production while maintaining membrane potential. Am. J. Physiol. Cell. Physiol. 292, C148–C156. doi: 10.1152/ajpcell.00215.2006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jeffries, O., Geiger, N., Rowe, I. C., Tian, L., McClafferty, H., Chen, L., et al. (2010). Palmitoylation of the S0-S1 linker regulates cell surface expression of voltage- and calcium-activated potassium (BK) channels. J. Biol. Chem. 285, 33307–33314. doi: 10.1074/jbc.M110.153940
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kathiresan, T., Harvey, M., Orchard, S., Sakai, Y., and Sokolowski, B. (2009). A protein interaction network for the large conductance Ca(2+)-activated K(+) channel in the mouse cochlea. Mol. Cell. Proteomics. 8, 1972–1987. doi: 10.1074/mcp.M800495-MCP200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kicinska, A., and Szewczyk, A. (2004). Large-conductance potassium cation channel opener NS1619 inhibits cardiac mitochondria respiratory chain. Toxicol. Mech. Methods 14, 59–61. doi: 10.1080/15376520490257482
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Knaus, H. G., Folander, K., Garcia-Calvo, M., Garcia, M. L., Kaczorowski, G. J., Smith, M., et al. (1994). Primary sequence and immunological characterization of β-subunit of high conductance Ca2+-activated K+ channel from smooth muscle. J. Biol. Chem. 269, 17274–17278.
Kulawiak, B., Kudin, A. P., Szewczyk, A., and Kunz, W. S. (2008). BK channel openers inhibit ROS production of isolated rat brain mitochondria. Exp. Neurol. 212, 543–547. doi: 10.1016/j.expneurol.2008.05.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, J., Al-Khalili, O., Ramosevac, S., Eaton, D. C., and Denson, D. D. (2010). Protein-protein interaction between cPLA2 and splice variants of alpha-subunit of BK channels. Am. J. Physiol. Cell. Physiol. 298, C251–C262. doi: 10.1152/ajpcell.00221.2009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Loot, A. E., Moneke, I., Keseru, B., Oelze, M., Syzonenko, T., Daiber, A., et al. (2012). 11,12-EET stimulates the association of BK channel alpha and beta(1) subunits in mitochondria to induce pulmonary vasoconstriction. PLoS ONE 7:e46065. doi: 10.1371/journal.pone.0046065
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ma, D., Nakata, T., Zhang, G., Hoshi, T., Li, M., and Shikano, S. (2007). Differential trafficking of carboxyl isoforms of Ca2+-gated (Slo1) potassium channels. FEBS Lett. 581, 1000–1008. doi: 10.1016/j.febslet.2007.01.077
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
McManus, O. B., and Magleby, K. L. (1991). Accounting for the Ca2+-dependent kinetics of single large-conductance Ca2+-activated K+ channels in rat skeletal muscle. J. Physiol. 443, 739–777. doi: 10.1113/jphysiol.1991.sp018861
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Meera, P., Wallner, M., and Toro, L. (2000). A neuronal beta subunit (KCNMB4) makes the large conductance, voltage- and Ca2+-activated K+ channel resistant to charybdotoxin and iberiotoxin. Proc. Natl. Acad. Sci. U.S.A. 97, 5562–5567. doi: 10.1073/pnas.100118597
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Noursadeghi, E., Jafari, A., Saghiri, R., Sauve, R., and Eliassi, A. (2014). Impairment of brain mitochondrial charybdotoxin- and ATP-insensitive BK channel activities in diabetes. Neuromol. Med. 16, 862–871. doi: 10.1007/s12017-014-8334-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ohya, S., Kuwata, Y., Sakamoto, K., Muraki, K., and Imaizumi, Y. (2005). Cardioprotective effects of estradiol include the activation of large-conductance Ca(2+)-activated K(+) channels in cardiac mitochondria. Am. J. Physiol. Heart Circ. Physiol. 289, H1635–H1642. doi: 10.1152/ajpheart.00016.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Piwonska, M., Wilczek, E., Szewczyk, A., and Wilczynski, G. M. (2008). Differential distribution of Ca2+-activated potassium channel beta4 subunit in rat brain: immunolocalization in neuronal mitochondria. Neuroscience 153, 446–460. doi: 10.1016/j.neuroscience.2008.01.050
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Saito, M., Nelson, C., Salkoff, L., and Lingle, C. J. (1997). A cysteine-rich domain defined by a novel exon in a slo variant in rat adrenal chromaffin cells and PC12 cells. J. Biol. Chem. 272, 11710–11717. doi: 10.1074/jbc.272.18.11710
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schmitt, N., Grunnet, M., and Olesen, S. P. (2014). Cardiac potassium channel subtypes: new roles in repolarization and arrhythmia. Physiol. Rev. 94, 609–653. doi: 10.1152/physrev.00022.2013
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schreiber, M., and Salkoff, L. (1997). A novel calcium-sensing domain in the BK channel. Biophys. J. 73, 1355–1363. doi: 10.1016/S0006-3495(97)78168-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shi, J., Krishnamoorthy, G., Yang, Y., Hu, L., Chaturvedi, N., Harilal, D., et al. (2002). Mechanism of magnesium activation of calcium-activated potassium channels. Nature 418, 876–880. doi: 10.1038/nature00941
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shi, Y., Jiang, M. T., Su, J., Hutchins, W., Konorev, E., and Baker, J. E. (2007). Mitochondrial big conductance KCa channel and cardioprotection in infant rabbit heart. J. Cardiovasc. Pharmacol. 50, 497–502. doi: 10.1097/FJC.0b013e318137991d
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shruti, S., Urban-Ciecko, J., Fitzpatrick, J. A., Brenner, R., Bruchez, M. P., and Barth, A. L. (2012). The brain-specific Beta4 subunit downregulates BK channel cell surface expression. PLoS ONE 7:e33429. doi: 10.1371/journal.pone.0033429
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Siemen, D., Loupatatzis, C., Borecky, J., Gulbins, E., and Lang, F. (1999). Ca2+-Activated K Channel of the BK-Type in the inner mitochondrial membrane of a human glioma cell line. Biochim. Biophys. Res. Comm. 257, 549–554. doi: 10.1006/bbrc.1999.0496
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Singh, H., Lu, R., Bopassa, J. C., Meredith, A. L., Stefani, E., and Toro, L. (2013). mitoBKCa is encoded by the Kcnma1 gene, and a splicing sequence defines its mitochondrial location. Proc. Natl. Acad. Sci. U.S.A. 110, 10836–10841. doi: 10.1073/pnas.1302028110
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Skalska, J., Bednarczyk, P., Piwonska, M., Kulawiak, B., Wilczynski, G., Dolowy, K., et al. (2009). Calcium ions regulate K(+) uptake into brain mitochondria: the evidence for a novel potassium channel. Int. J. Mol. Sci. 10, 1104–1120. doi: 10.3390/ijms10031104
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Skalska, J., Piwonska, M., Wyroba, E., Surmacz, L., Wieczorek, R., Koszela-Piotrowska, I., et al. (2008). A novel potassium channel in skeletal muscle mitochondria. Biochim. Biophys. Acta 1777, 651–659. doi: 10.1016/j.bbabio.2008.05.007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Soltysinska, E., Bentzen, B. H., Barthmes, M., Hattel, H., Thrush, A. B., Harper, M. E., et al. (2014). KCNMA1 encoded cardiac BK channels afford protection against ischemia-reperfusion injury. PLoS ONE 9:e103402. doi: 10.1371/journal.pone.0103402
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Stowe, D. F., Aldakkak, M., Camara, A. K., Riess, M. L., Heinen, A., Varadarajan, S. G., et al. (2006). Cardiac mitochondrial preconditioning by Big Ca2+-sensitive K+ channel opening requires superoxide radical generation. Am. J. Physiol. Heart Circ. Physiol. 290, H434–H440. doi: 10.1152/ajpheart.00763.2005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Szabo, I., and Zoratti, M. (2014). Mitochondrial channels: ion fluxes and more. Physiol. Rev. 94, 519–608. doi: 10.1152/physrev.00021.2013
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Szewczyk, A., Kajma, A., Malinska, D., Wrzosek, A., Bednarczyk, P., Zablocka, B., et al. (2010). Pharmacology of mitochondrial potassium channels: dark side of the field. FEBS Lett. 584, 2063–2069. doi: 10.1016/j.febslet.2010.02.048
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tanaka, Y., Meera, P., Song, M., Knaus, H.-G., and Toro, L. (1997). Molecular constituents of maxi KCa channels in human coronary smooth muscle. Predominant α + β subunit complexes. J. Physiol. 502, 545–557. doi: 10.1111/j.1469-7793.1997.545bj.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Testai, L., Martelli, A., Marino, A., D'Antongiovanni, V., Ciregia, F., Giusti, L., et al. (2013). The activation of mitochondrial BK potassium channels contributes to the protective effects of naringenin against myocardial ischemia/reperfusion injury. Biochem. Pharmacol. 85, 1634–1643. doi: 10.1016/j.bcp.2013.03.018
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thiede, A., Gellerich, F. N., Schonfeld, P., and Siemen, D. (2012). Complex effects of 17beta-estradiol on mitochondrial function. Biochim. Biophys. Acta 1817, 1747–1753. doi: 10.1016/j.bbabio.2012.02.034
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tian, L., Duncan, R. R., Hammond, M. S., Coghill, L. S., Wen, H., Rusinova, R., et al. (2001). Alternative splicing switches potassium channel sensitivity to protein phosphorylation. J. Biol. Chem. 276, 7717–7720. doi: 10.1074/jbc.C000741200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Toro, B., Cox, N., Wilson, R. J., Garrido-Sanabria, E., Stefani, E., Toro, L., et al. (2006). KCNMB1 regulates surface expression of a voltage and Ca2+-activated K+ channel via endocytic trafficking signals. Neuroscience 142, 661–669. doi: 10.1016/j.neuroscience.2006.06.061
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Uebele, V. N., Lagrutta, A., Wade, T., Figueroa, D. J., Liu, Y., McKenna, E., et al. (2000). Cloning and functional expression of 2 families of {beta}-subunits of the large conductance calcium-activated K+ channel. J. Biol. Chem. 275, 23211–23218. doi: 10.1074/jbc.M910187199
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Valverde, M. A., Rojas, P., Amigo, J., Cosmelli, D., Orio, P., Bahamonde, M. I., et al. (1999). Acute activation of maxi-K channels (hSlo) by estradiol binding to the beta subunit [In Process Citation]. Science 285, 1929–1931. doi: 10.1126/science.285.5435.1929
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wallner, M., Meera, P., and Toro, L. (1999). Molecular basis of fast inactivation in voltage and Ca2+-activated K+ channels: a transmembrane β –subunit homolog. Proc. Natl. Acad. Sci. U.S.A. 96, 4137–4142. doi: 10.1073/pnas.96.7.4137
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Williams, S. E., Wootton, P., Mason, H. S., Bould, J., Iles, D. E., Riccardi, D., et al. (2004). Hemoxygenase-2 is an oxygen sensor for a calcium-sensitive potassium channel. Science 306, 2093–2097. doi: 10.1126/science.1105010
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wojtovich, A. P., Nadtochiy, S. M., Urciuoli, W. R., Smith, C. O., Grunnet, M., Nehrke, K., et al. (2013). A non-cardiomyocyte autonomous mechanism of cardioprotection involving the SLO1 BK channel. PeerJ. 1:e48. doi: 10.7717/peerj.48
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wojtovich, A. P., Sherman, T. A., Nadtochiy, S. M., Urciuoli, W. R., Brookes, P. S., and Nehrke, K. (2011). SLO-2 is cytoprotective and contributes to mitochondrial potassium transport. PLoS ONE 6:e28287. doi: 10.1371/journal.pone.0028287
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wrzosek, A. (2014). The potassium channel opener NS1619 modulates calcium homeostasis in muscle cells by inhibiting SERCA. Cell Calcium 56, 14–24. doi: 10.1016/j.ceca.2014.03.005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Xia, X. M., Zeng, X., and Lingle, C. J. (2002). Multiple regulatory sites in large-conductance calcium-activated potassium channels. Nature 418, 880–884. doi: 10.1038/nature00956
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Xu, W., Liu, Y., Wang, S., McDonald, T., Van Eyk, J. E., Sidor, A., et al. (2002). Cytoprotective role of Ca2+- activated K+ channels in the cardiac inner mitochondrial membrane. Science 298, 1029–1033. doi: 10.1126/science.1074360
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yan, J., and Aldrich, R. W. (2010). LRRC26 auxiliary protein allows BK channel activation at resting voltage without calcium. Nature 466, 513–516. doi: 10.1038/nature09162
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yan, J., and Aldrich, R. W. (2012). BK potassium channel modulation by leucine-rich repeat-containing proteins. Proc. Natl. Acad. Sci. U.S.A. 109, 7917–7922. doi: 10.1073/pnas.1205435109
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yang, H., Shi, J., Zhang, G., Yang, J., Delaloye, K., and Cui, J. (2008). Activation of Slo1 BK channels by Mg2+ coordinated between the voltage sensor and RCK1 domains. Nat. Struct. Mol. Biol. 15, 1152–1159. doi: 10.1038/nsmb.1507
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yuan, P., Leonetti, M. D., Hsiung, Y., and MacKinnon, R. (2012). Open structure of the Ca2+ gating ring in the high-conductance Ca2+-activated K+ channel. Nature 481, 94–97. doi: 10.1038/nature10670
Yuan, P., Leonetti, M. D., Pico, A. R., Hsiung, Y., and MacKinnon, R. (2010). Structure of the human BK channel Ca2+-activation apparatus at 3.0 a resolution. Science 329, 182–186. doi: 10.1126/science.1190414
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zarei, M. M., Eghbali, M., Alioua, A., Song, M., Knaus, H. G., Stefani, E., et al. (2004). An endoplasmic reticulum trafficking signal prevents surface expression of a voltage- and Ca2+-activated K+ channel splice variant. Proc. Natl. Acad. Sci. U.S.A. 101, 10072–10077. doi: 10.1073/pnas.0302919101
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zarei, M. M., Song, M., Wilson, R. J., Cox, N., Colom, L., Knaus, H.-G., et al. (2007). Endocytic trafficking signals in KCNMB2 regulate surface expression of a voltage and Ca2+-Activated K+ channel. Neuroscience 147, 80–89. doi: 10.1016/j.neuroscience.2007.04.019
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhou, X., Wulfsen, I., Korth, M., McClafferty, H., Lukowski, R., Shipston, M. J., et al. (2012). Palmitoylation and membrane association of the stress axis regulated insert (STREX) controls BK channel regulation by protein kinase C. J. Biol. Chem. 287, 32161–32171. doi: 10.1074/jbc.M112.386359
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhou, Y., and Lingle, C. J. (2014). Paxilline inhibits BK channels by an almost exclusively closed-channel block mechanism. J. Gen. Physiol. 144, 415–440. doi: 10.1085/jgp.201411259
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: mitochondria, potassium channels, BK channels, MaxiK channels, subunit composition, permeability transition pore, ischemia reperfusion injury
Citation: Balderas E, Zhang J, Stefani E and Toro L (2015) Mitochondrial BKCa channel. Front. Physiol. 6:104. doi: 10.3389/fphys.2015.00104
Received: 29 January 2015; Accepted: 13 March 2015;
Published: 31 March 2015.
Edited by:
Alex M. Dopico, The University of Tennessee Health Science Center, USAReviewed by:
Adam Szewczyk, Nencki Institute of Experimental Biology, PolandAnna Bukiya, The University of Tennessee Health Science Center, USA
Copyright © 2015 Balderas, Zhang, Stefani and Toro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ligia Toro, Division of Molecular Medicine, Department of Anesthesiology, David Geffen School of Medicine at UCLA, BH-509A CHS, Box 957115, Los Angeles, CA 90095-7115, USA ltoro@ucla.edu