Drought Facilitates Species Invasions in an Urban Stream: Results From a Long-Term Study of Tropical Island Fish Assemblage Structure
 Alonso Ramírez1*†
Alonso Ramírez1*†  Pablo E. Gutiérrez-Fonseca1
Pablo E. Gutiérrez-Fonseca1  Sean P. Kelly1†
Sean P. Kelly1†  Augustin C. Engman1,2
Augustin C. Engman1,2  Karleen Wagner1 Keysa G. Rosas1,3 Natalia Rodríguez1
Karleen Wagner1 Keysa G. Rosas1,3 Natalia Rodríguez1- 1Department of Environmental Sciences, University of Puerto Rico, San Juan, Puerto Rico
- 2North Carolina Cooperative Fish and Wildlife Research Unit, Department of Applied Ecology, North Carolina State University, Raleigh, NC, United States
- 3Odum School of Ecology, University of Georgia, Athens, GA, United States
The natural flow regime is a key regulator of the dynamics of stream communities and ecosystem processes. Native fish assemblages are evolutionarily adapted to their local flow regime. In the Caribbean, streams are vulnerable to changes in climate that will alter their flow regimes. We assessed long-term patterns in fish assemblages (2008–2016) in an urban stream in San Juan, Puerto Rico. We hypothesized that drought periods would result in negative effects on native fish species and positive effects on introduced species. Fish assemblages were sampled annually from 2008 to 2016 by backpack electrofishing, with additional sampling during 2015, which was a drought year. Cumulative dry season discharge was consistently over 300 m3/s from 2008 to 2013, except from 2014 to 2016 when it dropped below 170 m3/s. Thirteen species were found, including most native species reported for Puerto Rico and introduced species (e.g., Cichlids, Poeciliids, and Loracariids). Native species were dominant in abundance during most years, except during drought and post-drought sampling events when introduced species became dominant. Introduced species increased in richness with time from less than two species between 2008 and 2013 to seven in 2015. The increase in introduced species was mostly attributed to the appearance of several species of cichlids toward the end of the study. Cluster analysis divided the data set in two groups: drought and non-drought sampling events. Introduced cichlids and the native A. monticola accounted for more than 60% of assemblage dissimilarity. Fish assemblage abundance and richness were negatively related to several components of stream hydrology (e.g., cumulative daily stream discharge, the number of major floods). A significant change in size over time was found for the native A. monticola, which decreased in size during the drought period. Overall, our study documents drought-facilitated invasions by several aggressive and highly ecologically competitive species, highlights the importance of the natural flow regime as a source of environmental resilience to invasions in tropical island streams, and provides an example of how climate change has and will likely continue to alter Caribbean stream fish assemblages.
Introduction
The natural flow regime plays an important role in the dynamics of stream communities and ecosystem processes via its effects on the physical environment (Bunn and Arthington, 2002; Poff and Zimmerman, 2010). Flow is a major driver of the availability and distribution of the habitats (e.g., floodplains, pools, and riffles) that are important to the success of many aquatic populations and the natural flow regime provides the background for which species adapt to particular environments (Lytle and Poff, 2004). Changes in flow patterns result in diverse impacts on aquatic communities depending on the frequency, duration, and magnitude of divergence from a natural regime and certain organisms may be able to adapt to novel conditions through plasticity in morphology, behavior, or life history (Poff and Zimmerman, 2010; Mims and Olden, 2012).
Native fish assemblages are composed of populations that are evolutionarily adapted to the local natural flow regime, which in some regions of the world includes large and rapid fluctuations in hydrology associated with variability in rainfall (Lytle and Poff, 2004). The natural flow regimes of tropical oceanic island streams, and many mountainous continental coastal areas, are typically flashy due to intense storms, steep catchments, and relatively small watersheds. Urban streams are also characterized by a flashy flow regime due to high proportions of impervious surfaces within their watersheds (Walsh et al., 2005; Ramírez et al., 2009). However, on oceanic islands that receive high precipitation urban and non-urban watersheds have been shown to have flashy hydrographs (Brasher, 2003; Ramírez et al., 2009). Given that native fish fauna in these regions are often diadromous (i.e., migrate to the ocean to complete their life cycle), a natural flow regime and longitudinal connectivity with the ocean appear to be more important than land use in maintaining native fish assemblages (Brasher, 2003; Engman and Ramírez, 2012).
Caribbean islands are vulnerable to climate change and extreme climatic events (Nurse et al., 2014; Khalyani et al., 2016). Future rainfall projections include changes in monthly, seasonal, and annual rainfall by the end of the century under global climate change scenarios (Van Beusekom et al., 2016). Projections show that by 2071–2100, mean annual rainfall will decrease from 25 to 50%, primarily because precipitation is expected to decrease by nearly 35% from August to October (Campbell et al., 2011). This decrease in precipitation will result in severe reductions of annual stream flow. For example, hydrologic models project total flow reductions of 49–88% under high greenhouse gas emission scenarios and 39–79% under low emission scenarios (Van Beusekom et al., 2016). The Greater Antilles have already begun to experience major reductions in precipitation during recent years (Comarazamy and González, 2011; Stephenson et al., 2014), with 2015 categorized the year with the strongest drought in the last 50 years (Herrera and Ault, 2017). These climate change-induced alterations to the natural flow regime could create niche opportunities for non-native fish species (Olden et al., 2006), which could lead to the emergence of novel stream fish assemblages.
In this study, we examined long-term variation in fish assemblage structure using a 9-year (2008–2016) data set of fish samples collected in an urban tropical stream in San Juan, Puerto Rico. During the study, the island was affected by severe drought conditions that started in 2013, and were particularly severe during 2015 (Herrera and Ault, 2017). We saw this historical drought event as an opportunity to assess the way in which climate change is altering the flow regime of an urban stream and how the stream fish assemblage responded to these changes. We expected drought periods to have less variable flow regimes, altered stream physicochemistry, and reduced habitat availability for fishes. We hypothesized that drought would have negative effects on native fish species, resulting in decreases in richness and abundance and also altering fish size structure. In contrast, we expect increases in richness and abundance of introduced species in response to the altered flow regime.
Materials and Methods
This study was conducted within the Río Piedras watershed (Figure 1), which drains the San Juan Metropolitan Area of Puerto Rico, a location with a population density of 3,733 people/km2 (Lugo et al., 2011). With steep slopes in its headwaters (at ~150 m elevation) and a gradient that decreases as it enters the coastal plain, the general topography of the Río Piedras is similar to other watersheds in Puerto Rico (De Jesús-Crespo and Ramírez, 2011). The watershed has been heavily modified by urbanization, many of its tributaries have been channelized and concrete lined, some ephemeral and intermittent streams have been paved over, and there have been considerable land movements that reshaped the local topography (Lugo et al., 2011). Impervious surfaces cover 49% of the entire watershed (De Jesús-Crespo and Ramírez, 2011), which results in high runoff rates (e.g., 72% of rainfall becomes stream flow; Osterkamp, 2001). Municipal infrastructure provides drinking water to the entire watershed, but sewage collection is not provided for headwater areas, which rely on septic systems. Sewage and storm water systems are combined in some areas, a fact that becomes evident after heavy rains (Ramírez et al., 2014). The watershed is no longer used as a source of water and no water treatment facilities discharge into the stream (Lugo et al., 2011). Therefore, changes in stream hydrology are the result of precipitation events and not management activities.
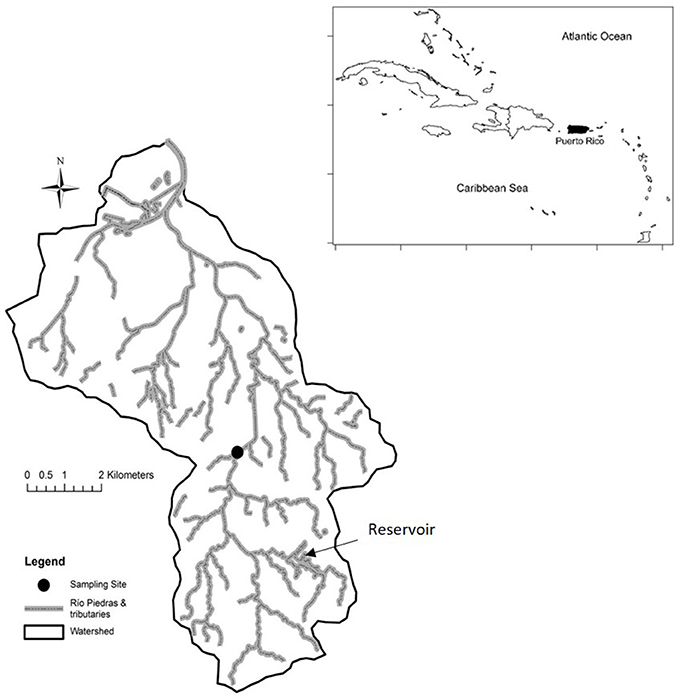
Figure 1. Map of the Río Piedras Watershed and Puerto Rico. The point in the middle of the watershed represents the location of our study site.
The native freshwater fish fauna of Puerto Rico is representative of oceanic islands in general because it is relatively species poor and all species are diadromous, migrating between marine and freshwater environments during their life cycle. In addition, this group of species is widely present in streams throughout the Caribbean region (Nordlie, 2012; Cooney and Kwak, 2013). At least nine species are known to occur in Puerto Rico: Bigmouth Sleeper (Gobiomorus dormitor), Smallscaled Spinycheek Sleeper (Eleotris perniger), Fat Sleeper (Dormitator maculatus), Mountain Mullet (Agonostomus monticola), American Eel (Anguilla rostrata), River Goby (Awaous banana), and Sirajo Goby (Sicydium spp.) (Cooney and Kwak, 2013). Sirajo Goby is a complex of morphologically similar species, which includes up to four species: Sicydium buscki, Sicydium gilberti, Sicydium plumieri, and Sicydium punctatum (Watson, 2000). Reliable identification is possible only at the adult stage by microscopic examination of upper jaw dentition (Watson, 2000). Introduced species richness is greater than native species richness in Puerto Rico, up to 70 species have been found in streams and reservoirs around the island (Neal et al., 2009). All native fishes and up to six introduced species have been previously reported for the Río Piedras watershed (Ramírez et al., 2009).
The study site is in the central part of the watershed, along the main stem of the Río Piedras (18°22'00.99“N−66°03'47.58”W, Figure 1). The site is upstream of the oldest and most densely developed part of the watershed, in the El Señorial neighborhood. The drainage area upstream from the sampling site has 35% impervious surface cover (Ramírez et al., 2014), a drainage area of 19.4 km2, and a total population of 30,786 (Lugo et al., 2011). The site is located at 60 m elevation, has an average yearly discharge of 0.61 m3/s (18-year record), and the stream channel (4–6 m wide) is composed of a series of riffles and pools with natural substrate dominated by boulders and cobble in riffles and sand in pools. Water chemistry reflects the urbanized watershed, with solute concentrations (e.g., NO3: >3 ppm, Ca: >25 ppm; Ramírez et al., 2014). Riparian vegetation is present, covering a strip of 10 to 15 m at each side, and composed of trees, grasses, and shrubs that contribute large amounts of organic matter. Flow data were obtained from the United States Geological Survey (USGS) gauging station at Hato Rey, located 7.5 km downstream from the study site (USGS 50049100). Although the El Señorial gauge (USGS 50048770) is closer to our study site, it has large data gaps.
Sampling
Ethics Statement
Research was conducted following standard protocols for fish surveys without specimen collections.
Physical and chemical characteristics of water have been monitored weekly in the Rio Piedras since 2011 (Ramírez et al., 2014). From this long-term monitoring, here we report monthly averages based on weekly measurements. Samples were collected from the center of the channel and measurements for water pH, specific conductivity, and temperature were taken in the field using a YSI Model 63 multi-parameter instrument (YSI Incorporates, Yellow Springs, Ohio). Total suspended solids were measured by filtering one liter of water through pre-combusted 0.70-μm glass fiber filters and obtaining dry weight.
Fish assemblages were sampled by backpack electrofishing using a Smith-Root model 12-B and following the standardized three-pass protocol suggested by Kwak et al. (2007). Block nets were placed at each end of a 100-m study reach to prevent fish from entering or exiting the area. Electrofishing started at the downstream end with one crewmember using the electrofisher moving the anode in a constant zigzag as the operator moved upstream. Additional crewmembers carried dip nets and buckets to collect fish. Each sampling pass along the reach was timed and completed in 30 min. All specimens captured were stored in buckets, and at the end of each pass fish were identified to species and measured (total length) to the nearest 0.1 mm. Fish from the first and second passes were held in mesh baskets in the stream and outside the study reach until the end of pass three, when all fish were released. Sampling was conducted annually from 2008 to 2016, except during the 2015 drought, when we added three additional samples to better characterize drought conditions.
Data Analysis
Using daily discharge data from the Hato Rey gauge station, we calculated cumulative daily discharge for the 6 months prior to each sampling event as a descriptor of stream hydrology. Most sampling events were in midsummer, so our cumulative daily discharge metric generally covered the dry season. Hydrological variability was characterized in two ways. First, we counted the number of floods during the same 6 months. We classified floods as discharges equal to or larger than 6 m3/s, which is the bank full flow of this site. Second, we applied the Richards-Baker Flashiness Index (Baker et al., 2004) by summing the absolute values of daily flow differences and dividing by the sum of the daily flows for the same period.
Fish abundance was calculated as total catch per sampling event, as sampling effort and the sampling reach was maintained constant over the 9 years. Thus, abundances reported here are minimum estimates of fish abundance in the study reach. Fish richness was calculated as total species richness obtained during the three passes. Individuals from the genus Sicydium are reported as a single taxon here (Sicydium spp.) because field identification is not possible (Watson, 2000).
Cluster analysis was used to identify groups among sampling events (e.g., years, except for 2015) based on abundance similarities. Ward's method was selected as a linkage method and Euclidean matrix as a dissimilarity distance measurement. Analysis of similarities (ANOSIM; Clarke and Warwick, 2001) was used to compare differences in assemblage structure among groups. ANOSIM scores fall between 0 and 1, with higher values corresponding to greater dissimilarity among groups. Similarity percentage-species contributions (SIMPER) procedure (Clarke and Warwick, 2001) was used to identify fish taxa that were most responsible for dissimilarities observed between groups. Linear regression analyses were used to relate fish abundance, richness, and individual size with hydrological variables.
Analyses were conducted with R statistical software (R Development Core Team, 2011, Version 2.13.1). For regression analyses, all data were tested for normality using the Shapiro-Wilk Test. Fish richness and abundance data were transformed by applying log(x) and log(x + 1.5) functions, respectively. P-values less than or equal to 0.05 were considered significant.
Results
Stream discharge in the Río Piedras reflected two contrasting climatic periods, “normal” wet years from 2008 to 2013 and then drought years from 2014 to 2016 (Figure 2). Peak drought was reached during 2015, when all months were below the long-term average discharge (Figure 2A). Cumulative discharge during the 6 months before sampling (mostly dry season) was consistently over 300 m3/s from 2008 to 2013, except for 2009 when it dropped to 200 m3/s (Figure 2B). In contrast, during the period from 2014 to 2016 it was below 170 m3/s (Figure 2A). The flashiness index did not reflect hydrological changes in the stream, showing similar values for all years and sampling events (Figure 2C). However, the number of floods recorded for the same 6-month period was considerably higher from 2008 to 2013 (except for 2009) relative to 2014–2016 (Figure 2D). Although 2015 was the only year recognized as a severe drought, both 2014 and 2016 had similar discharge characteristics to 2015 (Figure 2).
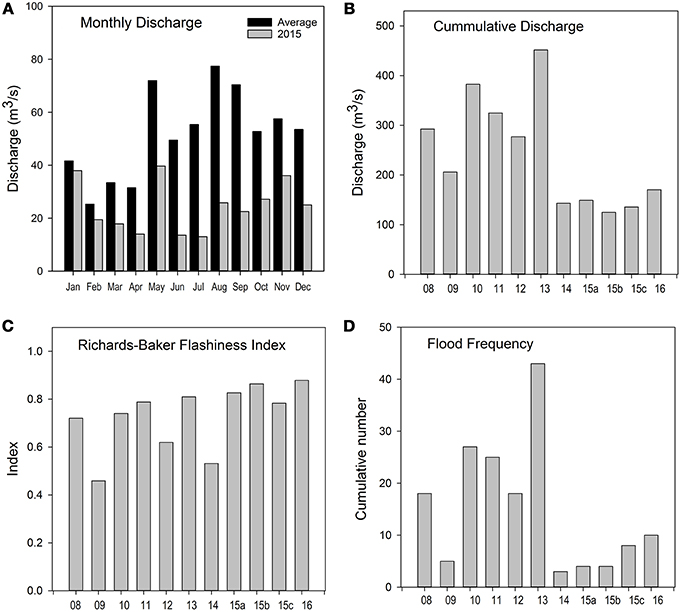
Figure 2. Discharge characteristics during the study: (A) Long-term average monthly discharge (2008–2014) relative to the drought (2015). Conditions for the 6 months prior to fish sampling: (B) cumulative daily discharge (m3/s), (C) Richards-Baker Flashiness index, and (D) flood frequency, number of floods defined as bank full events (larger than 6 m3/s). Data from the United States Geological Survey gaging station at Hato Rey, located 7.5 km downstream from the study site (USGS 50049100).
Water physical and chemical characteristics were not strongly affected by drought. Water temperature presented annual seasonality, with maximum values during summer and slightly higher temperatures during 2015 (Figure 3A). Conductivity and total suspended solids (Figures 3B,D) were not noticeably different during drought years, but water pH decreased slightly during 2015–2016 (Figure 3C).
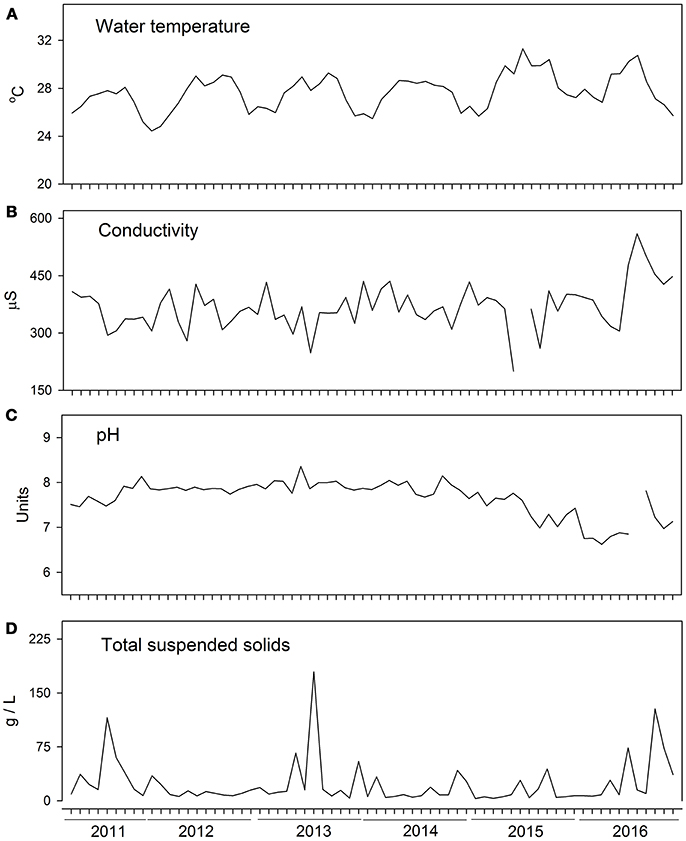
Figure 3. Physical and chemical characteristics of the water, weekly measurements from 2011 to 2016. (A) Water temperature, (B) Specific conductivity, (C) Water pH, (D) Total suspended solids.
We sampled thirteen fish species in the Río Piedras including most of the native freshwater fish species reported for Puerto Rico (Table 1). Introduced species were dominated by Cichlids, Poeciliids, and Loracariid catfishes; families known to be aquarium-trade species (Kwak et al., 2016). Cichlids were the most diverse group among introduced species, with a total of four species (Table 1).
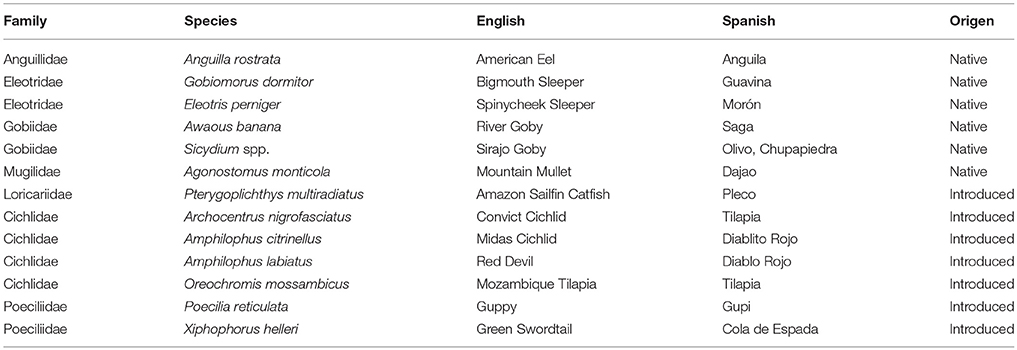
Table 1. Fish species found at the study reach in the Rio Piedras during the study.
Total fish abundance ranged from only 25 individuals during 2013 to 260 in 2009. Native species were numerically dominant during most years, except in 2015 and 2016 when introduced species became dominant (Figure 4A). Among native species, A. monticola was the most abundant in all years. Most other natives had a peak abundance during 2009, but they had similarly low abundances during the subsequent years (Figure 5). 2013 was a year with low abundances of most natives, except for E. perniger (Figure 5C).
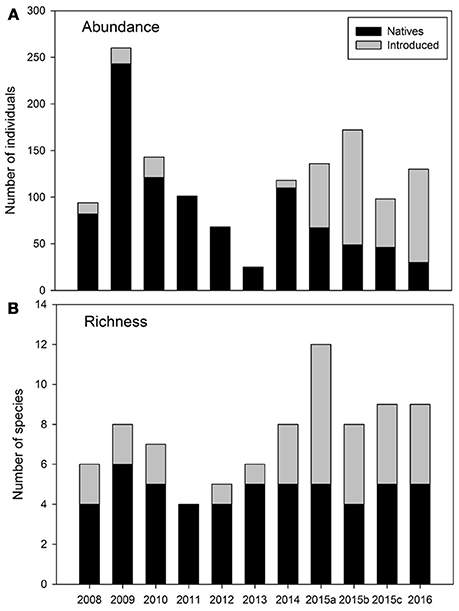
Figure 4. Annual changes in total fish abundance (A) and total species richness (B) divided by natives (dark column) and introduced (gray column) species for the Rio Piedras Watershed. Note that three sampling events were conducted during 2015.
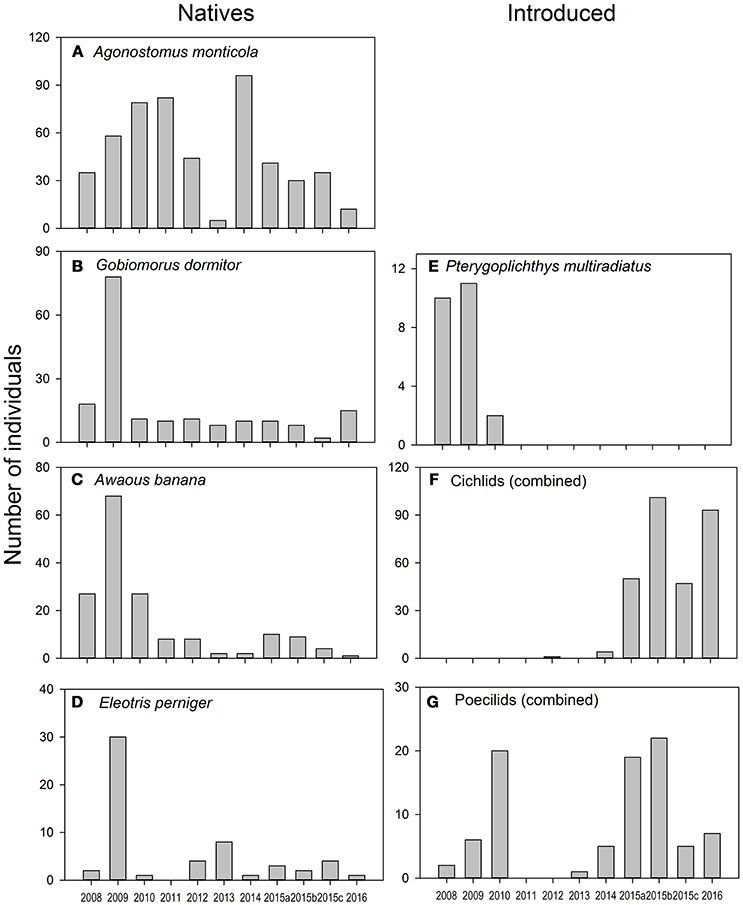
Figure 5. Annual changes in the abundance of dominant species in the Rio Piedras, divided by natives and introduced species. Columns are annual total abundance. Agonostomus monticula (A), Gobiomorus dormitor (B), Awaous banana (C), Eleotris perniger (D), Pterygoplichthys multiradiatus (E), combined abundance of Cichlids (F) and Poecilids (G).
Fish assemblage composition ranged from four species in 2011 to 12 in the first sampling of 2015 (2015a, Figure 4B). Native fish richness ranged from four to six species. Inter-annual variation in native richness was mostly due to a few elusive taxa, Anguilla and Sicydium, which were rarely captured and always had low abundances. Native species abundance was characterized as being variable between years (Figures 5A–D). Introduced species increased in richness with time from less than two species between 2008 and 2013 to seven in 2015a (Figure 4B). The increase in introduced species was mostly attributed to the appearance of several species of cichlids toward the end of the study. Despite the overall increase in introduced fish richness, we did observe the loss of some species. Pterygoplichthys multiradiatus, an introduced catfish, was very conspicuous during the first 3 years of the study, but has not been found in the study reach since 2010 (Figure 5E). Nevertheless, we have collected this species in other reaches within the same watershed. Cichlids followed the opposite pattern, they were absent at the beginning of the study and became dominant in the last few sampling events (Figure 5F). Poeciliids were variable over time, peaking in abundance during 2010 and 2015 (Figure 5G).
Cluster analysis divided the data set in two groups, the various samplings during 2015 and 2016 formed one group, while samples from the remaining years formed another group (Figure 6). Differences between the groups were confirmed with ANOSIM, with a high dissimilarity value (R = 0.57; p < 0.05). Cichlids and A. monticola accounted for more than 60% of assemblage dissimilarity between the two groups, according to SIMPER analysis. Cichlids dominated the 2015–2016 group (drought group) and native species the 2008–2014 group (non-drought group, Figure 6).
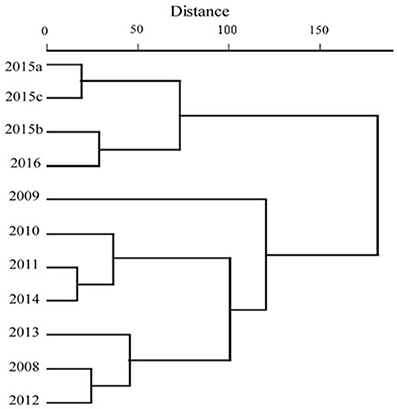
Figure 6. Cluster analysis, using Ward's method, showing two major groups based on fish abundance and composition: non-drought years (2008–2014) and drought (2015).
Fish assemblage abundance and richness were both negatively related with several components of stream hydrology during the 6 months prior to sampling. Cumulative daily stream discharge was negatively related to introduced fish abundance (R2 = −0.48; p = 0.02), total fish richness (R2 = −0.48; p = 0.02), and introduced fish richness (R2 = −0.51; p = 0.01). The number of floods was negatively related to total fish abundance (R2 = −0.40; p = 0.04), introduced fish abundance (R2 = −0.43; p = 0.03), total fish richness (R2 = −0.41; p = 0.03), and introduced fish richness (R2 = −0.44; p = 0.03). Flashiness did not vary greatly during the study, including during drought years (average index 0.73, during drought = 0.83), and was only found to be negatively related to native fish abundance (R2 = −0.57; p = 0.01).
Size distribution was homogeneous for the majority of the species over the 9 years of sampling. Five taxa were collected consistently enough to provide sufficient data to test for changes in total length over the eleven sampling dates. Native species included A. monticola, G. dormitor, and A. banana, which had minimum and maximum total lengths ranging from 19 to 330 mm, 54 to 419 mm, and 55 to 240 mm, respectively. Introduced taxa included Poeciliidae and Cichlidae, which were found to be generally smaller, with minimum and maximum total lengths ranging from 13 to 140 mm and 10 to 230 mm, respectively. Out of these five taxa, a significant change in size over time was only observed for A. monticola. There was only a modest trend of decreasing median total length in A. monticola when analyzing the entire 9-year sampling period (R2 = −0.34; p = 0.06; Figure 7). However, there was a significant decrease in the last seven sampling events from 2012 to 2016 (R2 = −0.85; p < 0.01; Figure 7), which is consistent with the last drought period reported for the island.
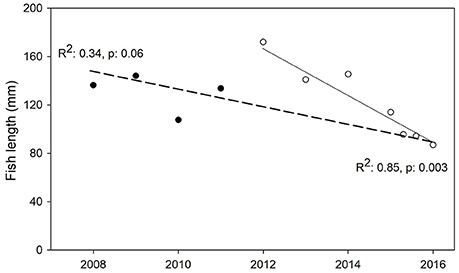
Figure 7. Median total length of Agonostomus monticola (Mountain Mullet) for each sampling date. Linear regression for the entire period (2008–2016) was not significant for changes in fish total length (dashed line, R2 = −0.34, p = 0.06). However, there was a significant decrease in median body length for the period 2012–2016 (solid line, R2 = −0.85, p < 0.01).
Discussion
Our long-term fish monitoring study revealed high inter-annual variability of fish occurrence and abundance in the Río Piedras watershed and that fish assemblage structure in this urban ecosystem is significantly affected by the flow regime. We argue that observed responses were the result of changes in hydrology due to precipitation patters, as water physical and chemical variables changed little during the study period and the watershed lacks water intakes or diversions. The Río Piedras fish assemblage is composed of a mix of native, diadromous species (typical of oceanic tropical islands around the world) and introduced species, which likely entered the system as aquarium or angler releases. Over the course of this study, an extreme drought reduced the overall discharge and frequency of floods in this stream which, caused a shift in the fish assemblage from being dominated by native species in terms of richness and abundance to introduced species. Our initial hypothesis that native fishes would be negatively affected by drought, while introduced species would benefit from it, was supported by the observed changes in fish populations. Drought effects at the population and assemblage levels seemed to be cumulative across years, as they were much more evident in 2015 than earlier during this drought period (e.g., 2013–2014).
The shift from a native-dominated fish assemblage to one dominated by introduced species was largely driven by invasions of Cichlids and declines in the native A. monticola population. Changes in the population of A. monticola over time at our study site could be the result of interactions with introduced cichlids, which became dominant during drier years. Introduced taxa commonly compete with native species for habitat and resources and this can have negative effects on native fish populations and significantly alter food web dynamics in aquatic ecosystems (Martin et al., 2010). Moreover, all four of the cichlids that occurred in our reach have been rated as having a high, very high, or extreme potential risk for invasion in other tropical contexts (Corfield et al., 2008). Competition among introduced and native taxa may be reduced during natural flow conditions through interspecies habitat segregation (e.g., pools and rapids), but augmented during droughts when space, habitat variability, and food resources become limited (Propst and Gido, 2004; Engman and Ramírez, 2012). A. monticola are known to prefer fast-moving water in riffles where they can feed on drifting macroinvertebrates in well-oxygenated water (Phillip, 1993). In Trinidad, A. monticola were found to have significantly different diets during contrasting flow conditions. In the low flow conditions of the dry season they fed mostly on algae, while during high flow conditions in the wet season their diet was dominated by insects and prawns (Phillip, 1993). Introduced cichlids, however, have been found to flourish in low flow and altered conditions because they prefer more lentic habitats where they consume a diversity of resources and are tolerant of poorly oxygenated waters (Canonico et al., 2005; Corfield et al., 2008). Engman and Ramírez (2012) found fish biomass to be negatively associated with increasing stream channel homogeneity within the Río Piedras watershed, and this loss of habitat diversity is similar to what was observed in our study during low flow conditions.
We hypothesize that low flow conditions in the Río Piedras could lead to more direct interactions (e.g., competition, niche overlap) between A. monticola and cichlids, resulting in the observed shift in size structure toward smaller individuals of A. monticola. Three potential causes for the observed shift in A. monticola size structure are: (1) direct mortality of large individuals, (2) decreased growth rates, and (3) displacement or emigration of larger individuals from the reach. None of these three causes are mutually exclusive and all three could be related to niche overlap with introduced Cichlidae under low flow conditions. For example, interspecific competition for space within the reach could prevent A. monticola from accessing microhabitats with high dissolved oxygen concentrations—which would be in short supply during drought conditions—and cause mortality in larger individuals. Additionally, diet overlap between A. monticola individuals and the introduced species could depress the growth rates and prevent small individuals from reaching larger size classes. Finally, larger A. monticola individuals could have simply emigrated from the reach because of competition for food or space with the introduced species.
Results of this study provide evidence that the natural flow regime of oceanic tropical island streams is a source of environmental resistance to invasion. Tropical wet island streams are naturally subject to rapid fluctuations in stream discharge, as the result of frequent storms (Ramírez et al., 2009). In short and steep watersheds, floods are a source of disturbance and can displace individuals downstream and increase mortality in populations. This is the case for Puerto Rico, where heavy rainfall results in flash floods and major disturbances that can decrease fish densities up to 46% in natural rivers within protected areas (Smith and Kwak, 2015). However, recruitment from the marine environment appears to be a source of resilience for these native diadromous species even in the face of extreme disturbances (Smith and Kwak, 2015). Moreover, native fish taxa on oceanic islands are morphologically and behaviorally adapted to the natural flow regime. For example, they are often benthically oriented and dorso-ventrally flattened to hold their positions during floods (Engman and Ramírez, 2012). Contrary to native species, introduced species in this system complete their entire life cycle in freshwater ecosystems, most lack successful adaptations to survive large floods, and in the case of our study, became invasive only in a hydrologically altered environment (i.e., during drought). Cichlids that invaded our study reach likely originated from an established population present in the Las Curias reservoir, an unmanaged reservoir located upstream in the watershed (Figure 1), well before the drought. There are established records of Oreochromis mossambicus in the Las Curias reservoir since 2006 (USGS) and Amphilophus spp. since 2011 (personal observations by the authors). So, although there were potential source populations of invasive species in an upstream lentic environment for many years they were likely unable to survive or become established in our riverine study reach until a drought provided favorable hydrological conditions.
Natural flow regimes are generally considered favorable to native fish assemblages due to evolutionary adaptations in morphology and behavior that allow them to outcompete introduced taxa adapted to different hydrologic conditions (Lytle and Poff, 2004; Propst and Gido, 2004; Engman and Ramírez, 2012). Results from a 9-year-long study in the San Juan River, New Mexico, showed that native fish species density was ten times greater during high flows when compared to low flows (Propst and Gido, 2004). Propst and Gido (2004) also found that non-native fish densities were negatively associated with high flows, but positively associated with extended periods of low flow during summer months. Under low flow conditions streams will lose habitat heterogeneity that can be important for the life histories of native fish species (Magoulick and Kobza, 2003; Fisk et al., 2015). Flow reduction is also known to function as an ecological filter for species assemblages, increasing similarity in composition among habitats, or pools in streams (Chase, 2007). In our urban watershed in Puerto Rico, introduced species were found to be more abundant in stream reaches that had increased habitat homogeneity associated with channelization (Engman and Ramírez, 2012). Despite the loss of habitat, all native fish taxa were found to be present within this heavily urbanized watershed, perhaps due to the potential for the recruitment of individuals from downstream estuaries (Engman and Ramírez, 2012). In addition to their dependence on marine environments, many diadromous species use floods as cues for locating rivers and for upstream migration. For example, A. monticola in Puerto Rico have limited movement, except after high flows that trigger their upstream movement (Smith and Kwak, 2015). Abundance and diversity of native and non-native fish assemblages within the Rio Piedras watershed therefore depend greatly on cues from large floods and habitat heterogeneity, both associated with hydrology.
Droughts causing low flow conditions are irregular events and their effects on freshwater ecosystems could be transitory and infrequent. However, current climate change predictions indicate that droughts will become more common in the Caribbean and many regions around the tropics (Neelin et al., 2006; Campbell et al., 2011). With more frequent droughts, changes in hydrological conditions are expected to be more frequent in Puerto Rico, which will have negative impacts on native aquatic organisms. At the same time, other human activities, such as water abstraction (i.e., high rates of constant water withdrawals) can exacerbate actual droughts (e.g., increasing the intensity or duration of low flow conditions) and cause drought-like conditions in streams even when precipitation is not excessively low. High water abstraction rates can impact stream longitudinal connectivity and thus alter movement between larval and adult habitats. Crook et al. (2007) showed that 70% of the mean flow generated within El Yunque National Forest in Puerto Rico is collectively diverted by water intakes before it reaches the estuary. The authors also showed that shrimps are exceptionally vulnerable to the effects of water withdrawal during times of low stream flow (Crook et al., 2007, 2009). Furthermore, a recent study by Engman et al. (2017) indicated that migrations of post-larval amphidromous fish in Puerto Rico were positively related to river discharge, which implies that drought and water abstraction could negatively affect the early life stages of fish populations. Recent studies in the Amazon basin provide additional support for the importance of drought in influencing fish assemblage composition in tropical streams. Röpke et al. (2017) studied river-floodplain ecosystems in the Amazon, finding that climate-driven hydrological changes are related to changes in fish assemblage composition, with extreme drought conditions generating the largest changes in composition.
Overall, our study documents drought-facilitated invasions by several aggressive and very ecologically competitive species, highlights the importance of the natural flow regime as a source of environmental resistance to invasions in tropical island streams, and provides an example of how climate change has and will likely continue to alter Caribbean stream fish assemblages. We predict that, in the absence of major droughts in the near future, the fish assemblage of the Río Piedras will be resilient to this invasion via rapid recovery of the native fish populations through recruitment. However, continued unimpeded freshwater-marine connectivity is necessary for this recovery to occur.
Author Contributions
All authors (AR, PG, SK, AE, KW, KR, NR) participated of the following: Substantial contributions to the conception or design of the work; or the acquisition, analysis, or interpretation of data for the work; and drafting the work or revising it critically for important intellectual content; and final approval of the version to be published; and agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This project was partially funded by the Luquillo Long-Term Ecological Research (LTER) program and the San Juan Urban Long-Term Research Area (ULTRA) program, both funded by the US National Science Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was made possible thanks to help provided by multiple members of our laboratory. We appreciate help and technical support by T. Kwak and his lab members at North Carolina State University.
References
Baker, D. B., Richards, R. P., Loftus, T. T., and Kramer, J. W. (2004). A new flashiness index: characteristics and applications to Midwestern rivers and streams. J. Am. Water Resour. Assoc. 40, 503–522. doi: 10.1111/j.1752-1688.2004.tb01046.x
Brasher, A. M. D. (2003). Impacts of human disturbances on biotic communities in Hawaiian streams. Bioscience 53, 1052–1060. doi: 10.1641/0006-3568(2003)053[1052:IOHDOB]2.0.CO;2
Bunn, S. E., and Arthington, A. H. (2002). Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manage. 30, 492–507. doi: 10.1007/s00267-002-2737-0
Campbell, J. D., Taylor, M. A., Stephenson, T. S., Watson, R. A., and Whyte, F. S. (2011). Future climate of the Caribbean from a regional climate model. Int. J. Climatol. 31, 1866–1878. doi: 10.1002/joc.2200
Canonico, G. C., Arthington, A. H., McCrary, J. K., and Thieme, M. L. (2005). The effects of introduced tilapias on native biodiversity. Aquatic conservation: Mar. Freshw. Ecosystems 15, 463–483. doi: 10.1002/aqc.699
Chase, J. M. (2007). Drought mediates the importance of stochastic community assembly. Proc. Nat. Acad. Sci. U.S.A. 104, 17430–17434. doi: 10.1073/pnas.0704350104
Clarke, K. R., and Warwick, R. M. (2001). Change in Marine Communities: an Approach to Statistical Analysis and Interpretation. London: Plymouth Marine Laboratory.
Cooney, P. B., and Kwak, T. J. (2013). Spatial extent and dynamics of dam impacts on tropical island freshwater fish assemblages. BioScience 63, 176–190. doi: 10.1525/bio.2013.63.3.6
Comarazamy, D. E., and González, J. E. (2011). Regional long-term climate change (1950–2000) in the midtropical Atlantic and its impacts on the hydrological cycle of Puerto Rico. J. Geophys. Res. 116:D00Q05. doi: 10.1029/2010JD015414
Corfield, J., Diggles, B., Jubb, C., McDowall, R. M., Moore, A., Richards, A., et al. (2008). Review of the Impacts of Introduced Ornamental Fish Species that have Established Wild Populations in Australia. Prepared for the Australian Government Department of the Environment, Water, Heritage, and the Arts.
Crook, K. E., Pringle, C. M., and Freeman, M. C. (2009). A method to assess longitudinal riverine connectivity in tropical streams dominated by migratory biota. Aquatic Conservation: Mar. Freshw. Ecosystems 19, 714–723. doi: 10.1002/aqc.1025
Crook, K. E., Scatena, F. N., and Pringle, C. M. (2007). Water Withdrawn From the Luquillo Experimental Forest, 2004. General Technical Report Rio IITF-GTR-34. Rio Piedras, PR: US Department of Agriculture, Forest Service, International Institute of Tropical Forestry.
De Jesús-Crespo, R., and Ramírez, A. (2011). Effects of urbanization on stream physicochemistry and macroinvertebrate assemblages in a tropical urban watershed in Puerto Rico. J. North Am. Benthol. Soc. 30, 739–750. doi: 10.1899/10-081.1
Engman, A. C., Kwak, T. J., and Fischer, J. R. (2017). Recruitment phenology and pelagic larval duration in Caribbean amphidromous fishes. Freshw. Sci. 36, 851–865. doi: 10.1086/694176
Engman, A. C., and Ramírez, A. (2012). Fish assemblage structure in urban streams of Puerto Rico: the importance of reach- and catchment-scale abiotic factors. Hydrobiologia 693, 141–155. doi: 10.1007/s10750-012-1100-6
Fisk, J. M., Kwak, T. J., and Heise, R. J. (2015). Effects of regulated river flows on habitat suitability for the robust redhorse. Trans. Am. Fish. Soc. 144, 792–806. doi: 10.1080/00028487.2015.1042557
Herrera, D., and Ault, T. (2017). Insights from a new high-resolution drought atlas for the Caribbean spanning 1950 to 2016. J. Clim. 30, 7801–7825. doi: 10.1175/JCLI-D-16-0838.1
Khalyani, A. H., Gould, W. A., Harmsen, E., Terando, A., Quinones, M., and Collazo, J. A. (2016). Climate change implications for tropical islands: interpolating and interpreting statistically downscaled GCM projections for management and planning. J. Appl. Meteorol. Climatol. 55, 265–282. doi: 10.1175/JAMC-D-15-0182.1
Kwak, T. J., Cooney, P. B., and Brown, C. H. (2007). Fishery Population and Habitat Assessment in Puerto Rico Streams: Phase 1 Final Report. Federal Aid in Sport Fish Restoration Project F-50 Final Report. Marine Resources Division, Puerto Rico Department of Natural and Environmental Resources, San Juan.
Kwak, T. J., Engman, A. C., Fischer, J. R., and Lilyestrom, C. G. (2016). “Drivers of Caribbean freshwater ecosystems and fisheries,” in Freshwater, Fish and the Future: Proceedings of the Global Cross-Sectoral Conference, eds W.W. Taylor, D.M. Bartley, C.I. Goddard, N.J. Leonard, and R. Welcomme (Bethesda, MD: Food and Agriculture Organization of the United Nations, Rome; Michigan State University, East Lansing; and American Fisheries Society).
Lugo, A. E., Ramos-González, O. M., and Rodríguez-Pedraza, C. (2011). The Rio Piedras Watershed and its Surrounding Environment. FS-980, U.S. Department of Agriculture, Forest Service, Río Piedras, Puerto Rico.
Lytle, D. A., and Poff, N. L. (2004). Adaptation to natural flow regimes. Trends Ecol. Evol. 19, 94–100. doi: 10.1016/j.tree.2003.10.002
Magoulick, D. D., and Kobza, R. M. (2003). The role of refugia for fishes during drought: a review and synthesis. Freshw. Biol. 48, 1186–1198. doi: 10.1046/j.1365-2427.2003.01089.x
Martin, C. W., Valentine, M. M., and Valentine, J. F. (2010). Competitive interactions between invasive Nile tilapia and native fish: the potential for altered trophic exchange and modification of food webs. PLoS ONE 5:e14395. doi: 10.1371/journal.pone.0014395
Mims, M. C., and Olden, J. D. (2012). Life history theory predicts fish assemblage response to hydrologic regimes. Ecology 93, 35–45. doi: 10.1890/11-0370.1
Neal, J. W., Lilyestrom, C. G., and Kwak, T. J. (2009). Factors influencing tropical island freshwater fishes: species, status, and management implications in Puerto Rico. Fisheries 34, 546–554. doi: 10.1577/1548-8446-34.11.546
Neelin, J. D., Münnich, M., Su, H., Meyerson, J. E., and Holloway, C. E. (2006). Tropical drying trends in global warming models and observations. Proc. Nat. Acad. Sci. U.S.A. 103, 6110–6115. doi: 10.1073/pnas.0601798103
Nordlie, F. G. (2012). Life-history characteristics of eleotrid fishes of the western hemisphere, and perils of life in a vanishing environment. Rev. Fish Biol. Fisher. 22, 189–224. doi: 10.1007/s11160-011-9229-3
Nurse, L. A., McLean, R. F., Agard, J., Briguglio, L. P., Duvat-Magnan, V., Pelesikoti, N., et al. (2014). “Small islands,” in Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, et al. (Cambridge, UK; NY: Cambridge University Press), 1613–1654.
Olden, J. D., Poff, N. L., and Bestgen, K. R. (2006). Life-history strategies predict fish invasions and extirpations in the Colorado River Basin. Ecol. Monogr. 76, 25–40. doi: 10.1890/05-0330
Osterkamp, W. R. (2001). Earth Surface Processes, Materials Use, And Urban Development: A Case Study of the San Juan Metropolitan Area, northeastern Puerto Rico. Tucson, AZ: Water Resources Division, US Geological Survey.
Phillip, D. A. (1993). Reproduction and feeding of the Mountain Mullet, Agonostomus monticola, in Trinidad, West Indies. Environ. Biol. Fishes 37, 47–55. doi: 10.1007/BF00000711
Poff, N. L., and Zimmerman, J. K. H. (2010). Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshw. Biol. 55, 194–205. doi: 10.1111/j.1365-2427.2009.02272.x
Propst, D. L., and Gido, K. B. (2004). Responses of native and nonnative fishes to natural flow regime mimicry in the San Juan River. Trans. Am. Fish. Soc. 133, 922–931. doi: 10.1577/T03-057.1
Ramírez, A., de Jesús-Crespo, R., Martinó-Cardona, D. M., Martínez-Rivera, N., and Burgos-Caraballo, S. (2009). Urban streams in Puerto Rico: what can we learn from the tropics? J. North Am. Benthol. Soc. 28, 1070–1079. doi: 10.1899/08-165.1
Ramírez, A., Rosas, K. G., Lugo, A. E., and Ramos González, O. M. (2014). Spatio-temporal variation in stream water chemistry in a tropical urban watershed. Ecol. Soc. 19:45. doi: 10.5751/ES-06481-190245
Röpke, C. P., Amadio, S., Zuanon, J., Ferreira, E. J., Deus, C. P., Pires, T. H., et al. (2017). Simultaneous abrupt shifts in hydrology and fish assemblage structure in a floodplain lake in the central Amazon. Sci. Rep. 7:40170. doi: 10.1038/srep40170
Smith, W. E., and Kwak, T. J. (2015). Tropical insular fish assemblages are resilient to flood disturbance. Ecosphere 6, 1–16. doi: 10.1890/ES15-00224.1
Stephenson, T. S., Vincent, L. A., Allen, T., Van Meerbeeck, C. J., McLean, N., Peterson, T. C., et al. (2014). Changes in extreme temperature and precipitation in the Caribbean region, 1961–2010. Int. J. Climatol. 34, 2957–2971. doi: 10.1002/joc.3889
Van Beusekom, A. E., Gould, W. A., Terando, A. J., and Collazo, J. A. (2016). Climate change and water resources in a tropical island system: propagation of uncertainty from statistically downscaled climate models to hydrologic models. Int. J. Climatol. 36, 3370–3383. doi: 10.1002/joc.4560
Keywords: climate change, tropical streams, disturbance, resistance, natural flow regime
Citation: Ramírez A, Gutiérrez-Fonseca PE, Kelly SP, Engman AC, Wagner K, Rosas KG and Rodríguez N (2018) Drought Facilitates Species Invasions in an Urban Stream: Results From a Long-Term Study of Tropical Island Fish Assemblage Structure. Front. Ecol. Evol. 6:115. doi: 10.3389/fevo.2018.00115
Received: 25 October 2017; Accepted: 19 July 2018;
Published: 08 August 2018.
Edited by:
Diane E. Pataki, University of Utah, United StatesReviewed by:
Olyssa Starry, Portland State University, United StatesLilian Casatti, Universidade de São Paulo, Brazil
Copyright © 2018 Ramírez, Gutiérrez-Fonseca, Kelly, Engman, Wagner, Rosas and Rodríguez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alonso Ramírez, aramirez@ramirezlab.net
†Present Address: Alonso Ramirez, Department of Applied Ecology, North Carolina State University, Raleigh, NC, United States
Sean P. Kelly, Department of Life Science, Tunghai University, Taichung, Taiwan