Species Diversity and Paleoecology of Late Pleistocene Horses From Southern Mexico
 Eduardo Jiménez-Hidalgo1*
Eduardo Jiménez-Hidalgo1*  Gerardo Carbot-Chanona2*
Gerardo Carbot-Chanona2*  Rosalía Guerrero-Arenas1
Rosalía Guerrero-Arenas1  Victor Manuel Bravo-Cuevas3 Genevieve Safi Holdridge4
Victor Manuel Bravo-Cuevas3 Genevieve Safi Holdridge4  Isabel Israde-Alcántara5
Isabel Israde-Alcántara5- 1Laboratororio de Paleobiología, Instituto de Recursos, Universidad del Mar, Puerto Escondido, Oaxaca, Mexico
- 2Museo de Paleontología “Eliseo Palacios Aguilera”, Dirección de Paleontología, Secretaría de Medio Ambiente e Historia Natural, Tuxtla Gutiérrez, Mexico
- 3Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo, Pachuca, Mexico
- 4Department of Geoscience and Center for Urban Network Evolution, Aarhus University, Aarhus, Denmark
- 5Instituto de Investigaciones en Ciencias de la Tierra, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Mexico
Equids are among the most common mammals found in faunal assemblages of Late Pleistocene age in Mexico. Much of what is known about the Equus species is the result of studies conducted in central and northern Mexico; much less is known about species in lower latitudes of Mexico. Here we describe three species that inhabited Oaxaca and Chiapas states. The fossil localities are in northwestern and central Oaxaca, as well as the central part of Chiapas. In Oaxaca, the largest species, Equus mexicanus, and the medium-sized Equus conversidens are represented by mandibles, skulls, diverse isolated teeth and some postcranial bones, while the smallest species, Haringtonhippus francisci is represented by a skull fragment and few isolated teeth. In Chiapas, E. mexicanus is represented by a mandible and several isolated teeth, E. conversidens by several mandibles and diverse isolated teeth, and H. francisci by isolated teeth and two mandibles. AMS radiocarbon and uranium dating of some of the equid localities in Oaxaca and Chiapas indicate that they were at least present since ~44,000 Cal BP years, they were common around 30,000 Cal BP years, and were still present at the end of the Pleistocene, around 12,000 years ago. The record of H. francisci from Chiapas is the youngest in North America. A cluster analysis of extended mesowear data and a discriminant analysis showed that Equus conversidens from Chiapas was obligate grazer, whilst the rest of the equids were variable grazers. Geographic distribution of localities in southern Mexico indicates that during the Pleistocene the equid species moved across the Transvolcanic Belt-Sierra Madre del Sur temperate biogeographic corridor and the Tamaulipas-Central America Gulf Lowlands tropical corridor.
Introduction
Equids are among the most common mammals in the Pleistocene Mexican faunal assemblages. Based on morphological characters, three or four Pleistocene horse species have been recently identified in Mexico. Alberdi et al. (2014) recognized Equus conversidens, Equus mexicanus, and Equus cedralensis. Priego-Vargas et al. (2017) recognized the previously mentioned species plus Equus francisci (see their table 10). Barrón-Ortiz et al. (2017), based on linear and geometric morphometric analyses of Equus specimens from the western interior of North America recognized three morphological groups of Equus in northeastern Mexico (San Josecito Cave, Nuevo León, and Cedral localities of San Luis Potosí) and they taxonomically identify such groups as E. ferus, E. conversidens and E. cedralensis; additionally, based on mitochondrial DNA analyses they recognized two equid clades [Caballine and New World Stilt-legged horses (NWSL)] and they referred the NWSL clade to E. conversidens. By means of paleogenomic and morphometric analyses, a new genus of NWSL horse for the Pleistocene of North America was erected: Haringtonhippus francisci. This species was reported (as Equus conversidens) from San Josecito Cave, Nuevo Léon, based upon a short mitochondrial DNA sequence (Barrón-Ortiz et al., 2017); this record was subsumed into H. francisci by Heintzman et al. (2017). Finally, Barrón-Ortiz et al. (2019) based on a morphology-based tree, determined that Equus comprises eight species, including H. francisci, and suggested that Haringtonhippus should be considered a synonym of Equus, but the authors do not formally synonymize both taxa.
At present there are 23 main Pleistocene equid localities in Mexico (Sánchez Salinas et al., 2016; Priego-Vargas et al., 2017); of them, just five (21.73%) were previously reported from southern Mexico (Doutt and Black, 1962; Pérez Crespo et al., 2008; Priego-Vargas et al., 2017; Díaz-Sibaja et al., 2018a). In most of these reports, the equids are only mentioned or briefly described.
In Oaxaca, Doutt and Black (1962) reported a molar of E. mexicanus near Yolomécatl (Figure 1), but they did not describe it. Many years later, Pérez Crespo et al. (2008) enlisted the Pleistocene mammalian localities from Oaxaca, including the horses (Equus mexicanus and E. excelsus); Jiménez-Hidalgo et al. (2012), briefly described the horses from the Viko vijin local fauna of northwestern Oaxaca (Equus mexicanus and E. conversidens) and Viñas-Vallverdú et al. (2017) enlisted Equus sp. as part of the Chazumba faunal assemblage. Finally, Díaz-Sibaja et al. (2018a) described some postcranial elements that were identified as Equus cf. E. conversidens from central Oaxaca (Figure 1).
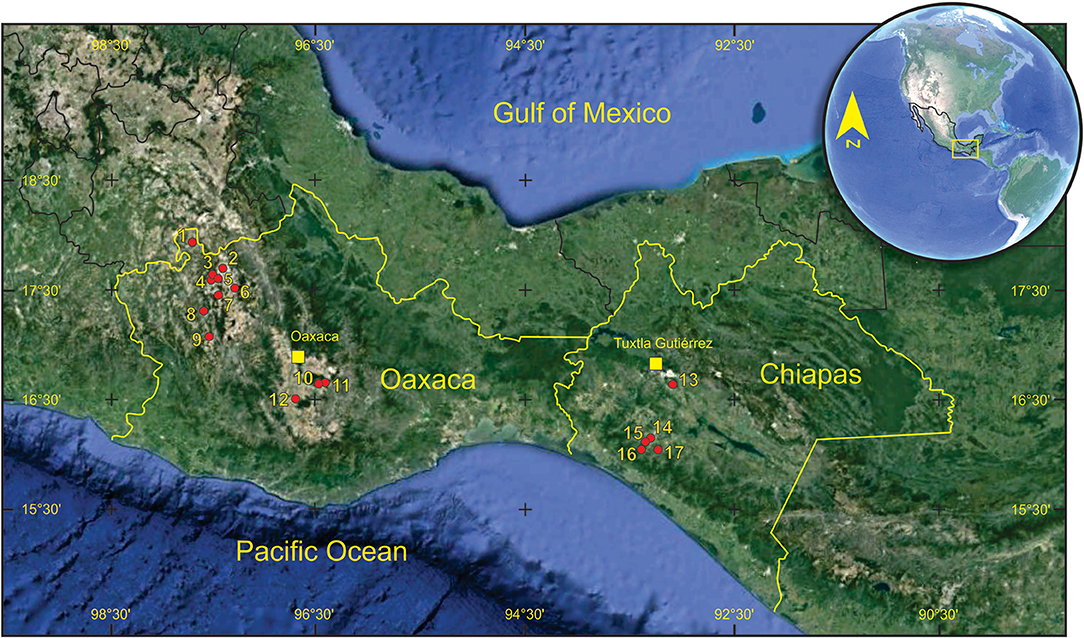
Figure 1. Main Late Pleistocene equid localities from Oaxaca and Chiapas. 1. Chazumba, 2. Concepción Buenavista, 3. Río Sabinal, 4. Río Salado, 5. Cañada del Misterio, 6. Sandage, 7. Río Tejupam; 8. Yolomécatl, 9. Magdalena Peñasco, 10. Güilla, 11. San Dionisio Ocotepec, 12. San Martín de los Cansecos, 13, Nandachuquí, 14. Los Mangos, 15. Gliptodonte, 16. La Simpatía, 17. La Tejería.
In Chiapas, Gómez-Pérez and Carbot-Chanona (2012) briefly described a specimen of Equus conversidens from the Villaflores municipality; the next year, Carbot-Chanona and Ovalles-Damián (2013) enlisted the vertebrate fossil record from Chiapas, including E. conversidens. Two years later, Pérez-Crespo et al. (2015) determined the dietary habits of some mammalian taxa from the Pleistocene of central Chiapas (Figure 1), including four specimens of E. conversidens. Finally, Bravo-Cuevas and Jiménez-Hidalgo (2018) enlisted the records of Pleistocene mammals from several areas of central and southern Mexico, including the equids.
This meager knowledge of southern Pleistocene horses compared to what is known from central and northern Mexico impedes our understanding of the inter-specific disparity of the recognized Mexican species, and the magnitude of the intraspecific variation along the Mexican territory. Furthermore, at present it is not possible to fully comprehend the dietary plasticity of Mexican species nor their biogeographic patterns during the Pleistocene.
Thus, the aims of this paper are: (1) to describe the equid species that inhabited northwestern and central Oaxaca, and central Chiapas during the Pleistocene; (2) to disclose the probable dietary preferences and body mass of the equids from Chiapas and Oaxaca; and (3) to comment about the geographic distribution of Mexican equids during the Pleistocene and the habitat where they roamed in southern Mexico.
Materials and Methods
Studied Specimens
The specimens from Oaxaca are housed at Laboratorio de Paleobiología, Universidad del Mar, Campus Puerto Escondido, Oaxaca, Mexico, under the acronym UMPE. Specimens from Chiapas are housed at Colección Paleontológica of the Museo de Paleontología “Eliseo Palacios Aguilera,” Tuxtla Gutiérrez, Chiapas, Mexico, under the acronym IHNFG. They were collected at some of the localities shown in Figure 1.
Since we evaluated morphological characters of teeth and bones of fossil horses from southern Mexico and to avoid the unnecessary proliferation of names for the Mexican equid species, we follow the taxonomic scheme of Priego-Vargas et al. (2017, table 10) that recognized four Pleistocene Equus species for Mexico: Equus conversidens, E. mexicanus, E. francisci, and E. cedralensis. Since Heintzman et al. (2017) transferred E. francisci to Haringtonhippus, instead of E. francisci, we used the name H. francisci.
We preferred not to synonymize Haringtonhippus to Equus, as suggested by Barrón-Ortiz et al. (2019), given the profound discrepancies between the genomic and morphologic analyses about the phylogenetic position of H. francisci, which reflect that the definition of Equus is a work still in progress (Barrón-Ortiz et al., 2019). Future analyses combining morphology and genomics will shed light about the phylogenetic relationships of H. francisci.
In order to disclose the taxonomic identity of the equid specimens studied, they were compared to those housed at the Colección de Macromamíferos, Museo de Paleontología, Universidad Autónoma del Estado de Hidalgo (UAHMP), the Vertebrate Paleontology Collection of the Natural History Museum of the Los Angeles County (LACM/CIT), and a cast of the holotype of Equus mexicanus (IGM 4009) housed at the Colección Nacional de Paleontología, Instituto de Geología, UNAM. Likewise, anatomical descriptions and illustrations of the Mexican Pleistocene species were used (Hibbard, 1955; Reynoso-Rosales and Montellano-Ballesteros, 1994; Alberdi et al., 2014; Priego-Vargas et al., 2017).
Measurements and Dental Nomenclature
Mandibular and skull measurements are in mm and follow the system of measurements for Equus bones and teeth (Eisenmann, 2009).
Teeth measurements are in mm and were taken following Eisenmann and Mashkour (2000).
Several measurements were taken from a basicranium specimen (Figure 2).
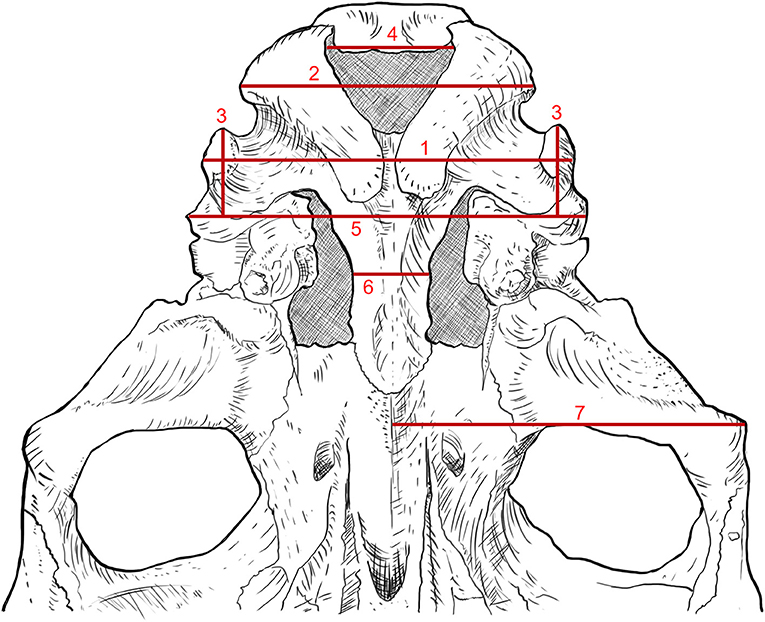
Figure 2. Measurements of the basicranium of equid species. 1. Width across jugal processes. 2. Width of occipital condyles. 3. Width of jugal apophyses. 4 width of foramen magnum. 5. Width across mastoid processes. 6. Minimum width of the basilar process. 7. Width across middle line of basisphenoid to the temporal articular processes.
All measurements were taken with digital calipers and recorded within 0.01 mm of accuracy.
Dental nomenclature follows Evander (2004), and Reynoso-Rosales and Montellano-Ballesteros (1994). The used curvature index of teeth follows MacFadden and Carranza-Castañeda (2002).
Paleoecological Aspects
Dietary Guilds
Mesowear analysis
An extended mesowear analysis was performed to include P4/p4 to M3/m3 (Kaiser and Fortelius, 2003; Kaiser and Solounias, 2003). We excluded teeth no yet in occlusion or showing initial wear or with damage on the occlusal surface, and those with a persisting crown height of <15 mm (Fortelius and Solounias, 2000; Kaiser and Fortelius, 2003; Kaiser and Solounias, 2003). A total of 112 cheek teeth were available for this study: 31 upper and 29 lower cheek teeth from Oaxaca (E. mexicanus, 13 upper, 16 lower; E. conversidens, 18 upper, 10 lower; H. francisci, three lower), and 39 upper and 13 lower cheek teeth from Chiapas (E. mexicanus, three upper; E. conversidens, 25 upper, eight lower; H. francisci, 11 upper, five lower). A hierarchical cluster analysis with Euclidean distance and complete linkage was performed on the mesowear variables perHigh, perSharp, perRound, perBlunt, of 35 extant species of herbivores with known dietary preferences (Fortelius and Solounias, 2000; Schulz and Kaiser, 2012) and five extinct Equus species (Table S1). The mesowear values of Equus grevyi and E. burchelli of Fortelius and Solounias (2000) were replaced by those of E. grevyi and E. quagga (= E. burchelli) of Schulz and Kaiser (2012).
We only coded the less-worn buccal cusp of each tooth, either the paracone or metacone in the P4-M3 and the protoconid or hypoconid in the p4-m3. The scored mesowear variables included the cusp shape (sharp, rounded, or blunt) and the occlusal relief (high or low). A sharp cusp is one that ends in a point and has no rounded area between the mesial and distal facet, a rounded cusp has a distinctly rounded apex but retains facets on the lower slopes and a blunt cusp is nearly flat and there are no facets (see Barrón-Ortiz et al., 2014). The occlusal relief depends on how high the cusp rises above the valley between paracone and metacone or protoconid and hypoconid (see Kaiser and Fortelius, 2003). All the scored data were converted to percentages, thus giving the variables perHigh, perSharp, perRound, perBlunt (Table S1). The dataset was analyzed with R software version 3.6.0 (R Core Team, 2014), using the library “fpc” (flexible procedures for clustering) version 2.2-3. Data were normalized, Euclidean distance and complete linkage were used to generate the clusters. A bootstrap analysis was performed (100 replicates) to identify how reliable were the groups (Zumel and Mount, 2014; the used script is in Supplementary Material 1).
Discriminant function analysis
To determine the accuracy of the classification among dietary guilds of the studied species, a discriminant function analysis was performed with the database used for the mesowear analysis, only using the dataset of extant species with known dietary habits as training set. Then, we ran a second analysis to predict the feeding group of the studied specimens. We used the linear discriminant method and common covariance as the data were of the same order of magnitude and were in covariance (Díaz-Sibaja et al., 2018b). Dietary category was the grouping variable (obligate grazer, variable grazer, mixed feeder, and browser). Posterior probabilities were calculated based on Bayes rules. We used the Wilk's lambda test (α = 0.05) to detect statistical differences between the multivariate centroids of the dietary categories. The analyses were carried out with JMP 8.0 (SAS Institute, 2019).
The cross-validation of the discriminant model was performed with the R library “flipMultivariates” version 0.1, using leave-one-out cross-validation.
Body Mass Estimation
To estimate the probable body mass of the fossil horses from southern Mexico, we used the predictive equations of Janis (1990) for perissodactyls and hyracoids only:
1. Log mass = (2.887 × log second upper molar width) + 1.345 (r2 = 0.993).
2. Log mass = (3.010 × log second lower molar length) + 1.216 (r2 = 0.986).
3. Log mass = (3.090 × log fourth lower premolar length) + 1.290 (r2 = 0.986).
In order to adhere to the equation specifications, the measurements of the teeth were converted to cm, and the estimated body mass to kg.
To detect if the estimated mean body mass of the identified species was statistically different, we used the Mann-Whitney U-test.
The datasets generated for this study can be found in the accompanying tables and in the Supplementary Materials and can be freely downloaded.
Stratigraphy of Fossil Localities
Stratigraphic information is available for some of the studied fossil localities. In the Sandage locality, municipality of Coixtlahuaca, northwestern Oaxaca (Figure 1), an Equus mexicanus maxillary fragment with teeth was discovered within an A horizon of a paleosol. The A horizon is thick (> 1 m), dark, organic rich, having a silty loam texture, a prismatic structure and evidence for slickensides. The enamel of one molar was dated by AMS, offering an age of 29,426–30,123 Cal BP. The paleosol is covered by several meters of alluvial sediments.
Also, in northwestern Oaxaca, in the Mixteca region, there are two fossil localities within the municipality of San Antonio Acutla. In the Cañada del Misterio locality (Figure 1), diverse E. conversidens and E. mexicanus specimens were collected from a medium-grained pinkish sand measuring 40 cm thick, which gradually changes from fine-grained sand to a microconglomerate with clasts measuring up to 3.0 cm in diameter. The fossiliferous bed is unconformably underlain by a succession of light brown silty and clayey beds. Organic sediment and a snail shell of the fossiliferous bed were dated by AMS, returning ages of 43,030–44,510 Cal BP and 43,120–44,830 Cal BP, respectively. At another site in Cañada del Misterio, an Equus mandible was collected on the top of an 11.0 cm thick microconglomerate containing clasts of up to 3.0 cm in diameter, which was covered by 25 cm of dark gray fine silt with abundant charcoal fragments. One of these fragments was dated by AMS, giving an age of 29,320–29,765 Cal BP.
In the Llano de Hueso locality, also in the municipality of San Antonio Acutla, an Equus conversidens skull and some E. mexicanus isolated teeth were recovered in the top of a poorly sorted fine- to medium- grained sand, 30 cm of thick, with medium-grained sand of paleochannels. The sand bed gradually becomes siltier toward the top. The E. conversidens skull was dated by means of uranium series, offering an age of 24,650–27,305 Cal BP (Ordoñez-Regil et al., 2016).
In the Río Salado locality (Figure 1), within Santiago Teotongo and San Antonio Acutla municipalities, several isolated teeth and some postcranial bones of E. conversidens, H. francisci, and E. mexicanus has been collected from fine-grained silty sand. A tooth of E. conversidens collected from the top of the stratigraphic sequence was dated by uranium series, giving an age of 11,380–12,620 Cal BP (Ordoñez-Regil et al., 2016).
In the Río Tejupam locality (Figure 1), within the Tejupam municipality, an H. francisci basicranium, as well as several E. conversidens and E. mexicanus specimens were collected from fine-grained silty sand and silty clay beds 25 cm thick (Jiménez-Hidalgo et al., 2011). Organic matter from the silty clayey sediments was dated by AMS returning an age of 19,584–20,180 Cal BP. The deposit of silty-clay is unconformably overlain by a 30 cm thick silty clay bed with dissecation marks. A massive fine-grained sand that interbeds with silty sediments with abundant charcoal overlays the previous bed. One charcoal piece was dated by AMS and showed an age of 6,880–7,005 Cal BP.
In the Güilla locality of central Oaxaca (Figure 1), an E. mexicanus upper teeth series were collected from the bottom of a well-rounded, imbricated alluvial and fluvial conglomerate of andesite and limestone clasts ranging 6.0–8.0 cm in diameter and of 4.0 m in thickness.
In Los Mangos locality of central Chiapas (Figure 1), several teeth and some mandibles of H. francisci, E. conversidens, and E. mexicanus were collected from a 75 cm thick deposit of silty sediments which was overlaid with coarse sand lenses in a silty matrix. Organic sediment from the top of the fossiliferous sediment was dated through AMS, showing an age of 12,720–12,820 Cal BP.
Associated Fauna
In the Acutla localities the associated fauna consists of gastropods of the families Physidae and Succineidae, Coelocentrum, Polygyra couloni, Mesomphix (Omphalina) lucubratus and Gyraulus parvus, the bivalve Pisidium, the lizard Sceloporus, the ground sloth Nothrotheriops, the lagomorph Sylvilagus floridanus, the rodents Microtus mexicanus, Peromyscus difficilis, and Reithrodontomys cf. R. megalotis, the tapir Tapirus, the cervid Odocoileus virginianus, the bovid Bison antiquus and the proboscideans Cuvieronius hyodon and Mammuthus columbi (Jiménez-Hidalgo et al., 2011, 2013).
In the Río Salado locality, the Equus specimens are associated with Glyptotherium cylindricum and the camels Hemiauchenia macrocephala and Camelops hesternus. The mammals Bison antiquus and M. columbi are also associated with the equids in the Acutla, Río Salado and Río Tejupam localities. Additional taxa from Río Tejupam include the ostracod Candona, the gastropods G. parvus, Planorbella trivolvis, Lymnaeidae and Poligyridae gastropods, the bivalve Pisidium casertanum, the salamander Ambystoma, and the rodents M. mexicanus, Neotoma mexicana, Neotomodon alstoni, Peromyscus difficilis, and the rabbit S. floridanus (Jiménez-Hidalgo et al., 2011, 2013; Guerrero-Arenas and Jiménez-Hidalgo, 2015).
The associated fauna collected from Los Mangos locality include the glyptodont Glyptotherium cylindricum, the deer Odocoileus virginianus, the capybara Neochoerus aesopi and the bovid Bison sp. (Gómez-Pérez and Carbot-Chanona, 2012; Carbot-Chanona et al., in press).
Results
Systematic Paleontology
Order Perissodactyla Owen, 1848
Family Equidae Gray, 1821
Tribe Equini Gray, 1821
Equus Linnaeus, 1758
Equus conversidens Owen, 1869
Referred Material
Chiapas state: locality Los Mangos. IHNFG-5351, rM2. locality Gliptodonte. IHNFG-2683, lP3-M3 teeth series, lM1-M3 teeth series, lm3, lm1, or lm2; IHNFG-2856, left mandible with p2, p4-m3; IHNFG-2685, lP2-M3 and rP2-M3 teeth series, rp2-m1 teeth series; IHNFG-4148, lp2-m3 and rp2-m3 teeth series, IHNFG-4162, lp4; IHNFG-5762, lP4; IHNFG-5803, lm1-m3 teeth series. locality La Tejería 2. IHNFG-5758, lP3-M3 and rP2-M3 teeth series, lp3-m3 teeth series and rm3. locality La Simpatía. IHNFG-5789, rP4; IHNFG-5804, rp3. Oaxaca state: Locality Oax-4 Río Salado. UMPE 014, lM3; UMPE 448, lm3; UMPE 450, lP4-M1, p3; UMPE 452, lM1; UMPE 470, rP4; UMPE 472 rm2; UMPE 484, rm3; locality Oax-5 Llano de Hueso. UMPE 077, skull; UMPE 476, lm3; UMPE 477 rm1; UMPE 479 lm3; UMPE 481, lM2; UMPE 535, rM1. Locality Oax-6 Cañada del Misterio. UMPE 453, rm1; UMPE 454, rm3; UMPE 478, lM3; UMPE 482, lm3; UMPE 508, rP4; UMPE 542, rp3 UMPE 625, P2-M3 and rostral fragment. Locality Oax-7 Río Tejupam. UMPE 008 left maxillary fragment with DP4-M3; UMPE 539 lM1; UMPE 569, rP3; UMPE 620, left mandible with p2-m3 and incisors; UMPE 898, lm3. Locality Oax-8 Magdalena Peñasco. UMPE 455 rM2; UMPE 457, rm2; UMPE 458, rm2. Locality Oax-17 Río Sabinal. UMPE 531, rM2. Locality San Martin de los Cansecos. UMPE 911, cast of lm3.
Description
Skull
The specimen UMPE 077 is somewhat dorsoventrally crushed (Figure 3). The skull is broken behind the retro-articular processes of the temporal bones. The nasal notch is retracted to a position that lies dorsal to the mesostyle of P2. The rostrum is deep and the facial crest begins at the level of the mesostyle of P4. Compared to extant Equus, the muzzle is short. The infraorbital foramen is located dorsal to the metastyle of P3. Measurements of the skull are in Table 1.
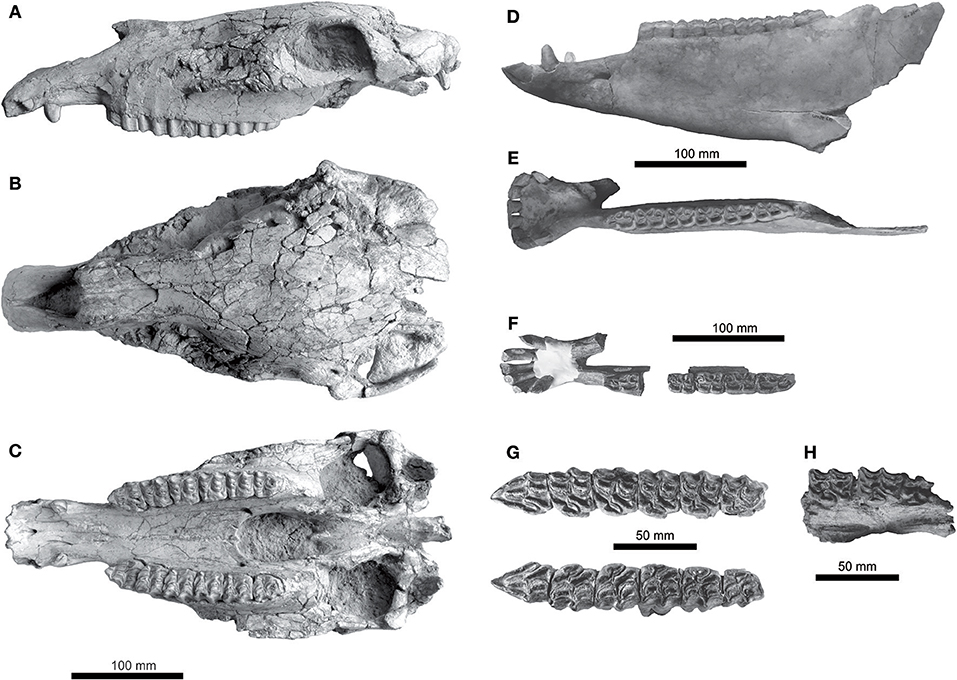
Figure 3. Equus conversidens from southern Mexico. UMPE 077, skull in (A) lateral, (B) dorsal, and (C) palatal views. UMPE 620, left mandible in (D) lateral and (E) dorsal views. IHNFG-2856, left mandible in (F) dorsal view. IHNFG-2685, upper teeth series in (G) occlusal view. UMPE 008, left maxillary with DP4- M2 in (H) occlusal view.
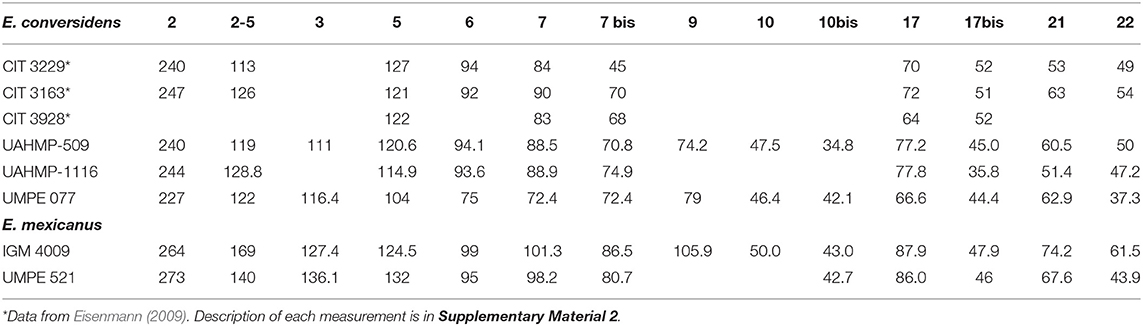
Table 1. Skull measurements of the equids from the Pleistocene of southern Mexico and other selected specimens from North America.
The palatine is lateromedially concave and narrow; the major palatine foramina and the rostral margin of the choanae are located at the level of the M2's protocone. The teeth series converge rostrally, especially the right one (Figure 3). The mandibular fossa is shallow.
Mandible
The mandibles are somewhat stout (Figure 3), increasing in height from the premolars to the molars (Table 2). The mental foramen is ovoid in outline and well-developed, and it is below the diastema, which is short. The incisor's arcade is quadrangular in outline. The symphysis is relatively slender (Table S2).
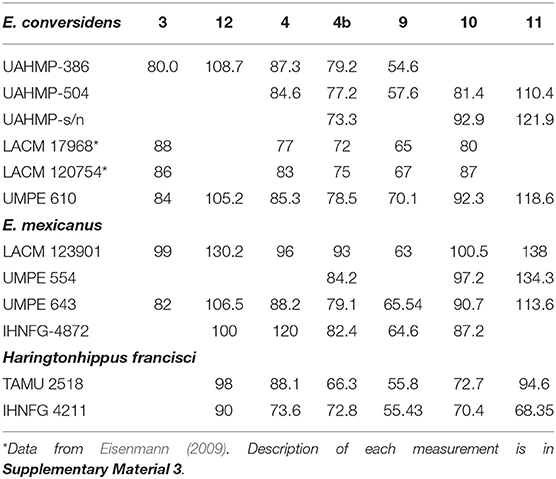
Table 2. Measurements of equid mandibles from the Pleistocene of southern Mexico and other selected specimens from North America.
Upper Teeth
The P2 is triangular in outline, rostro-caudally short (L/B mean ratio of 1.398), the anterior accessory rib is slightly developed, the parastyle is rounded to slightly acute on its rostral portion; the mesostyle is rectangular in outline and the metastyle is slightly developed. The pli protoloph and the pli protoconule are deep; the pli postfossette and pli hypostyle are also deep. The protocone varies from long to rounded. The plicaballine is moderately developed (Figure 3).
The P3 and P4 are quadrangular (Table S2), with quadrangular or rounded parastyle and mesostyle, the metastyle is slightly developed; there are 2 or 3 folds in the caudal part of the prefossette, the pli protoloph is deep and the pli protoconule could be shallow or deep. The pli postfossette is deep and the pli hypostyle is shallow or deep. The protocone is elongated and slightly curved on its lingual side, and its length represents around 50% the total tooth length (Table S2).
The M1 and M2 are quadrangular (Table S2), the parastyle is rounded, the mesostyle is quadrangular to rounded and the metastyle is slightly developed; there are three or no folds in the caudal part of the prefossette, the pli protoloph is shallow to absent; the pli protoconule is deep; the pli postfossette generally is deep, but it is absent in one specimen; the pli hypostyle is shallow or absent. The protocone is elongated and usually flat, but in few specimens, it is slightly curved in its lingual part (Figure 3), its length represents slightly less than the 50% of the total tooth length. There is no pli caballine in these molars.
The M3 is triangular in outline and small (Table S2), its parastyle and mesostyle are rounded, and the metastyle is not present. There are 1–3 folds in the caudal part of the prefossette, the pli protoloph is absent, the pli protoconule can be shallow or deep. The pli postfossette is shallow to absent; the pli hypostyle is deep or absent. The protocone is long, flat, slightly curved or sinuous in its labial side (Figure 3). The M3's have a curvature index of 40.
Lower Teeth
The incisors lack infundibula; the i3 is separated by the canine by a short diastema (Figure 3). In males the canine is well-developed and bears a crest that runs all along the crown height.
The p2 is triangular in outline (Table S3), the metaconid is rounded and the metastylid has an acute tip, the linguaflexid is “U”-shaped, there is a rounded entoconid; the postflexid is very long. The ectoflexid is shallow and does not penetrate the isthmus. There is a faint pli caballinid.
The p3 and p4 are rectangular, the metaconid is rounded and the metastylid is triangular (Figure 3), the linguaflexid is “V”-shaped; the postflexid is long and with a somewhat sinuous labial margin; usually, the ectoflexid is moderately deep and does not penetrate the isthmus; the pli caballinid is slightly developed.
The m1 and m2 are also rectangular (Table S3) and the m3 is triangular in outline. They have a rounded metaconid and a triangular metastylid; the linguaflexid is “V” or “U”-shaped (Figure 3); the postflexid is long and less sinuous than in the premolars or almost flat; the entoflexid is moderately deep or deep and can or cannot penetrate the isthmus. The pli caballinid can be present or absent.
Discussion
The studied specimens have a close morphology and dimensions when compared to fossil material of Equus conversidens from central and northern Mexico. In this species the muzzle is short and tapers rostrally, the infraorbital foramen is set high, the facial crest extends anteriorly to a position dorsal to P4, and the teeth series converge rostrally, as in UMPE 077 (Figure 3; Table 1). The studied upper teeth from Oaxaca and Chiapas are of medium size compared to the other Mexican Pleistocene species (Table S2), the P2 is rostro-caudally short and the M3 is small. The mandible is stout and has a similar size to those identified as E. conversidens (Table 2); the lower teeth have an elongated protocone that is slightly curved on its lingual side, the premolars and molars have few folds in the inner side of the fossettes, they have V-shaped linguaflexids and moderately deep ectoflexids; the size of the teeth is similar to those of E. conversidens (Table S3) and intermediate in size between E. mexicanus, E. cedralensis, and H. francisci (Azzaroli, 1998; Bravo-Cuevas et al., 2011; Alberdi et al., 2014; Priego-Vargas et al., 2017).
The above described combination of characters identifies the studied specimens as Equus conversidens.
In the studied sample, the M3 of E. conversidens has the lowest curvature index (40 vs. 70 and 50 in the other species). If this curvature value is also present in specimens from other localities, it could be useful to identify isolated M3 of this species.
On the other hand, the presence/absence of incisors' infundibula is very variable within horse species, and it also changes with tooth wear (Barrón-Ortiz et al., 2017). Thus, we do not consider it as a useful taxonomic character, but we believe it is important to describe the character state of the sample from southern Mexico.
Equus mexicanus (Hibbard, 1955)
Referred Material
Chiapas state: locality Los Mangos. IHNFG-4872, mandible with both series of DP2-M2, the left incisors and both canine roots; IHNFG-4886, lP3 or lP4; IHNFG-5660 rP4. Locality Nandachuqui. IHNFG-694, rP2; IHNFG-691, rM1 or rM2. Locality La Simpatía. IHNFG-5352, rM3. Locality La Tejería. IHNFG-4700, Mx. Oaxaca state: Locality Sandage. UMPE 908, lP4-M3 and rM1-M3 teeth series. Locality Oax-4 Río Salado. UMPE 011, lM1; UMPE 900 rm3; UMPE 447, calcaneum; UMPE 489, left metatarsal III. Locality Oax-5 Llano de Hueso. UMPE lP4; UMPE 474, P3; UMPE 480, Mx; UMPE 490, right mandible with p2-m2; UMPE 903, rP4. Locality Oax-6 Cañada del Misterio. UMPE 546, rp3; UMPE 643, dentary with both p2-m3 teeth series, the canines and incisors; UMPE 897, lM2; UMPE 901, lP2; UMPE 902, rm2. Locality Oax-7 Río Tejupam. UMPE 505, rM2. Locality Oax-17 Río Sabinal. UMPE 521, skull; UMPE 554, left mandible with p3-m3; UMPE 567, lP2; UMPE 611, lm2. Locality Güilla. UMPE 907, lP2-M3 and rP2-Px-M2.
Description
Skull
The specimen UMPE 521 is dorsally crushed, so, it is not possible to observe any undistorted feature. It is large, caudally, the skull preserves the left temporal articular tubercle and the retro-articular process (Figure 4). The premaxillaries are stout. The nasal notch is retracted to a position that lies dorsal to the mesostyle of P2. Rostrally, the facial crest ends between the P4 and the M1. The muzzle is short (Table 1), as in the skull of E. conversidens.
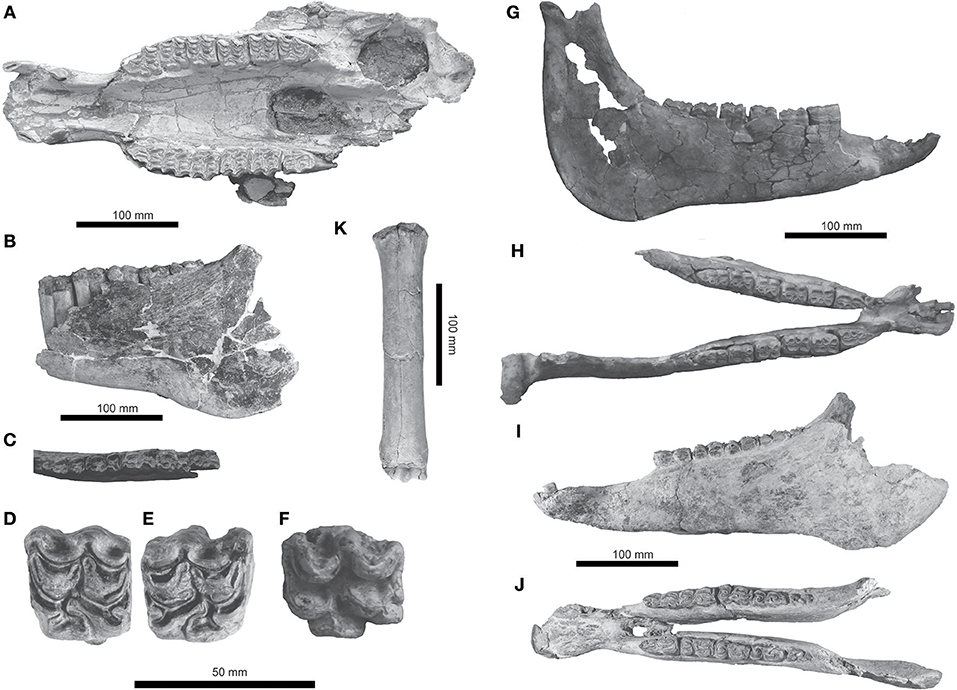
Figure 4. Equus mexicanus from southern Mexico. UMPE 521, skull in (A) palatal view. UMPE 554, left mandible in (B) lateral and (C) occlusal views. (D) IHNFG-4886, M1; (E) IHNFG-5660, P4, and (F) UMPE 908, P4 in occlusal views. UMPE 643, mandible in (G) lateral and (H) dorsal views. IHNFG-4872, mandible in (I) lateral and (J) dorsal views. UMPE 489, left metatarsal in (K) cranial view.
The palatine is wide (Table 1); the anterior margin of choanae is immediately behind M2's protocone (Figure 4). The upper teeth series seems straight. The mandibular fossa is shallow.
Mandible
The mandible is somewhat stout, its height increases caudally along the teeth series (Table 2). The diastema is short. The incisor's arcade has a “U” outline. The symphysis is somewhat slender. The articular process is very similar to extant Equus.
Upper Teeth
The P2 is long and triangular in outline (Table S2), the anterior accessory rib is slightly developed, the parastyle is acute to slightly rounded, the mesostyle is rectangular in outline (Figure 4), there is a deep pli protoloph and pli protoconule; the pli postfossette and pli hypostyle are also deep. The protocone is subrounded with a wide isthmus. The plicaballine is moderately to well-developed.
The P3 and P4 are quadrangular, with quadrangular parastyle and mesostyle, there are 4–5 folds in the caudal part of the prefossette, and the pli protoloph and the pli protoconule are deep. The pli postfossette and pli hypostyle are also deep. The protocone is elongated and slightly curved on its lingual side, its length represents around 2.41 of the total length of these premolars. The plicaballine is single and moderately developed.
The M1 and M2 are quadrangular, the parastyle is rounded to quadrangular, the mesostyle is rectangular in outline, there is 1–3 folds in the caudal part of the prefossette, the pli protoloph is very variable, it can be absent, be moderately deep or deep; the pli protoconule can be shallow to deep. The pli postfossette varies from moderately deep to very deep; the pli hypostyle varies from shallow to deep. The protocone is elongated and can be slightly curved to flat on its lingual side; its length represents around 2.14 of the total teeth length. The pli caballine is faint to absent, only in the M2 UMPE 505 it is well-developed.
The M3 is triangular in outline (Table S2); it has a rounded parastyle, a rounded to slightly quadrangular mesostyle and there is no metastyle (Figure 4). There are one to four shallow folds in the caudal part of the prefossette, the pli protoloph is shallow, the pli protoconule can be shallow or deep. The pli postfossete is faint to absent, the pli hypostyle is deep. The protocone is long and flat to slightly curved lingually. The M3's have a curvature index of 50.
Lower Teeth
The incisors lack infundibula; the i3 is separated by the canine by a very short diastema. In females the canine is a very small conical tooth.
The p2 is triangular in outline (Table S3), the metaconid is rounded and the metastylid has an acute tip, the linguaflexid is “V”-shaped, there is a rounded entoconid; the postflexid is very long. The ectoflexid is shallow and does not penetrate the isthmus.
The p3 and p4 are rectangular (Table S3), the metaconid is rounded and the metastylid is triangular to ovoid with an acute tip in outline, the linguaflexid is variable, can have a “V” shape, or is “U”-shaped; the postflexid is very long and with a sinuous labial margin; the ectoflexid is moderately deep and does not penetrate the isthmus; the pli caballinid can be present or absent.
The deciduous premolars of IHNFG-4872 are large and their morphology is very similar to those of the permanent premolars (Figure 4), the ectoflexid is deep and penetrates the isthmus. Their measurements are in Table S4.
The m1 and m2 are almost quadrangular and the m3 is triangular in outline. They have a rounded metaconid and a triangular to slightly rounded metastylid; the linguaflexid is “V” or “U”-shaped, the postflexid is long and less sinuous than in the premolars; the ectoflexid is deep and penetrates the isthmus. The pli caballinid is present to slightly developed.
Postcranial Bones
The partial left calcaneus UMPE 447 is large, stout and with a quadrangular outline; the sustentaculum is broken. The left metatarsal UMPE 489 is stout and dorsoventrally short (Figure 4); its length is 263.1 mm, the proximal width is 55.3 mm, diaphysis width at middle is 34.49 mm; distal width is 52 mm.
Discussion
The skull UMPE 521 shares with the holotype of Equus mexicanus its large size, a nasal notch that is retracted to a position that lies dorsal to the P2 and a facial crest that ends between the P4 and the M1 (Hibbard, 1955); the mandibles from Chiapas and Oaxaca share their large size (Table 2) and robustness with this species (Azzaroli, 1998).
The upper teeth from southern Mexico have a large size with moderately to complexly plicated enamel on fossettes, long, flat to somewhat curved protocones, a deep pli protoloph and pli hypostyle on premolars and a long P2; these features are present in E. mexicanus, as are those of the lower molars, such as a large size, a predominantly triangular metastylid, and deep to moderately deep ectoflexids.
The limbs bones are large and stout, as those of E. mexicanus from Cedral, San Luis Potosí (Alberdi et al., 2014).
The P2s of E. mexicanus and H. francisci are longer than those of E. conversidens; by the same token, the occlusal area of the M3 in these species is comparatively larger than that of E. conversidens (Table S2) and its curvature index is higher.
The above described combination of characters allows this sample to be identified as Equus mexicanus.
Haringtonhippus (Heintzman et al., 2017)
Haringtonhippus francisci (Hay, 1915)
Referred Material
Chiapas state: locality Los Mangos. IHNFG-4211, dentary with both p2-m3 teeth series, the canines and incisors; IHNFG-4698, lP2-M3 and rP2-M2 teeth series; IHNFG-4700, rP2; IHNFG-4211, mandible with lp3-m1, rp2-m3, i1-2 and the canines; IHNFG-4699, mandible with lp2-m3, incisive fragments, and canines. Locality Gliptodonte. IHNFG-4155, lP2-M1 teeth series and rP4, rM2; IHNFG-2708, Mx. Oaxaca state: locality Oax-5 Llano de Hueso. UMPE 906, rP4. Locality Oax-6 Cañada del Misterio. UMPE 905, lP3. Locality Oax-7 Río Tejupam. UMPE 561, basicranium. Oax-17 Río Sabinal. UMPE 904, lP4.
Description
Skull
The available skull is broken, only the basicranium is preserved (Figure 5); the left bones are slightly distorted; the bones seem somewhat slender. The zygomatic arches are missing. The caudal part of the frontal is present, it is flat as in extant Equus; the parietals are somewhat bulbous, and the median sagittal crest is slightly developed. The foramina that are present at the base of the temporal crest are large. Measurements of the basicranium are in Table 3.
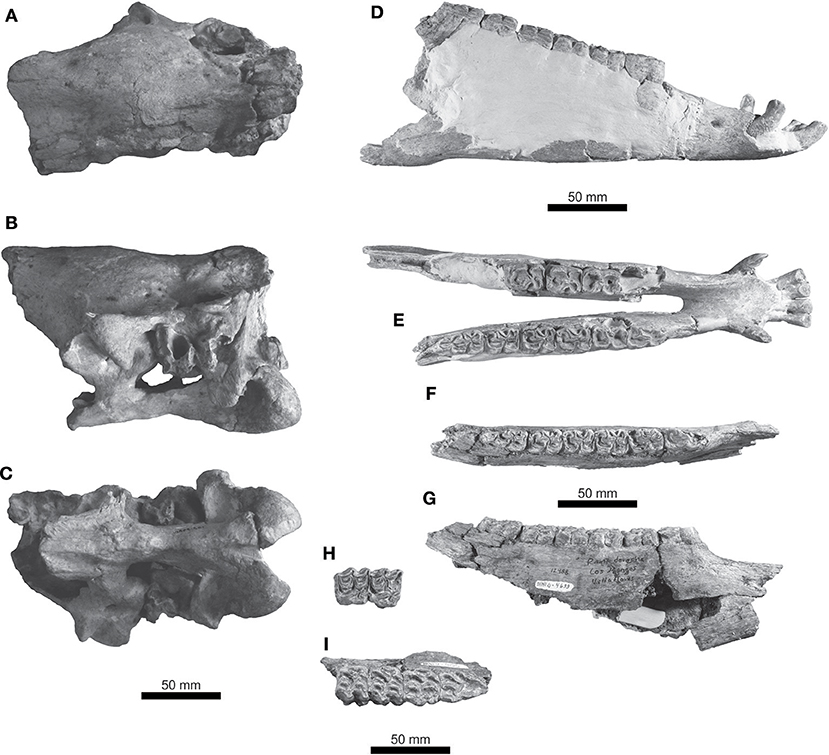
Figure 5. Haringtonhippus francisci from southern Mexico. UMPE 561, basicranium in (A) dorsal, (B) lateral, and (C) ventral views. IHNFG-4211, mandible in (D) lateral and (E) dorsal views. IHNFG-4699, mandible in (F) lateral and (G) dorsal views. IHNFG-4155, right P4-M1 in (H) occlusal view and left P2-M1 in (I) occlusal view.
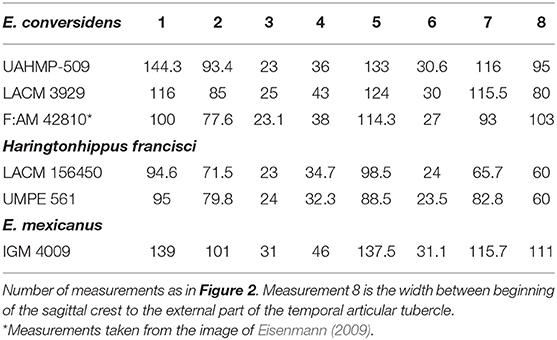
Table 3. Basicranium measurements of Pleistocene equid species present in Mexico.
The external auditory meatus is faintly ovoid in outline; the mastoid process is triangular in outline as in extant Equus.
The nuchal crest is missing, the occipital condyles are very similar to the ones of extant Equus, the jugal processes are dorsoventrally short; the basioccipital bone is slender, as is the preserved portion of the basisphenoid. The retroarticular processes are also dorsoventrally short. The preserved part of the mandibular fossa is deeper than in E. conversidens and E. mexicanus specimens from Oaxaca.
Mandible
There are two mandible fragments in the sample (Figure 5). They are small and somewhat stout; they increase in height from the premolars to molars (Table 2). The mental foramen is ovoid in outline and well-developed.
Upper Teeth
The P2 is triangular in outline and long (Table S2), the anterior accessory rib is slightly developed, the parastyle is rounded, the mesostyle is rectangular to slightly rounded and the metastyle is poorly developed (Figure 5). The pli protoloph and the pli protoconule are moderately deep; the pli posftossette and pli hypostyle are also moderately deep. The protocone is short. The plicaballine is well-developed in IHNFG-4698, but is absent in IHNFG-4155, which has more worn teeth.
The P3 and P4 are quadrangular, with quadrangular to subrounded parastyle and mesostyle; the metastyle is slightly developed (Figure 5). There are 2 or 3 folds in the caudal part of the prefossette, the pli protoloph is deep to shallow, the pli protoconule is deep. The pli posfossette is deep to moderately deep and the pli hypostyle is shallow. The protocone is elongated and slightly curved on its lingual side, and its length represents around 43.3% of the total length of these premolars (Table S2); the pli caballine is absent.
The M1 and M2 are quadrangular (Table S2), the parastyle is rounded, the mesostyle is quadrangular and the metastyle is slightly developed. There are two or no folds on the caudal portion of the prefossete, the pli protoloph is deep to shallow, the pli protoconule, the pli postfossete and the pli hypostyle are shallow. The protocone is elongated and flat to slightly concave in its lingual part; it represents around 45% of the total tooth length. The pli caballine is absent.
The M3 is triangular in outline, the parastyle and mesostyle are quadrangular in outline, the metastyle is absent. There is 1-fold in the caudal part of the prefossette, the pli protoloph is somewhat deep, the pli protoconule is very deep; the pli postfossette is absent and the pli hypostyle is deep. The protocone is very long and flat. The M3 has a curvature index of 70.
Lower Teeth
The incisors lack infundibula. In males the canine is well-developed and bears a crest that runs all along the crown.
The p2 is triangular in outline (Table S3), the metaconid is rounded and the metastylid has a sub-acute tip; the linguaflexid has an open “V” shape, the entoconid is rounded and the postflexid is long. The ectoflexid is moderately deep and does not penetrate the isthmus. There is not pli caballinid.
The p3 and p4 are rectangular (Table S3), the metaconid is rounded and the metastylid is triangular, the linguaflexid is predominantly “V”-shaped, but few are “U”-shaped; the postflexid is relatively long and almost flat; the ectoflexid is moderately deep and does not penetrate the isthmus. There is no pli caballinid.
The m1 and m2 also are rectangular (Table S3) and the m3 is very elongated and triangular in outline. These molars have a rounded or sub-acute metaconid and a triangular to subrounded metastylid; the linguaflexid is “V” or slightly “U”-shaped; the postflexid is somewhat long and flat or slightly concave; the ectoflexid is deep and penetrate the isthmus. The pli caballinid is absent.
Discussion
The basicranium from Oaxaca is most similar in size to a skull identified as H. francisci (LACM(CIT) 109/156450) from the Pleistocene of Nevada (Heintzman et al., 2017); UMPE 561 is smaller than those of E. conversidens and E. mexicanus (Table 3). The mandible measurements of IHNFG-4211 are like those of the mandible TAMU 2518, which is part of the holotype of H. francisci (Table 2).
The upper molars from Chiapas and Oaxaca share with E. cedralensis and H. francisci their small size (Table S2), their flat or somewhat curved and elongated protocone, the few plications on the fossettes, the absence of plicaballine and the very elongated protocone of M3. The lower molars are also of small size (Table S3) and share with the small equid species a rounded metaconid and a predominantly acute metastylid, and a lack of pli caballinid (Hay, 1915; Lundelius and Stevens, 1970; Alberdi et al., 2014; Priego-Vargas et al., 2017).
Teeth of H. francisci and E. cedralensis have a very similar occlusal pattern and size (Barrón-Ortiz et al., 2017; Priego-Vargas et al., 2017), but according to Alberdi et al. (2014), E. cedralensis does not possess slender metapodials as H. francisci, but shorter and stouter ones. The studied sample of small teeth was not associated with any metapodial; nonetheless, the basicranium and the mandible are morphologically similar and about the same size as those of H. francisci, and giving that this small species was the first erected (Hay, 1915), we identify the small specimens from southern Mexico as H. francisci.
Paleoecological Aspects of Equid Species From Southern Mexico
Dietary Guilds
The cluster of the mesowear analysis shows five main groups (Figure 6); one containing the hypergrazer Bison bison and Equus conversidens from Hidalgo (LD = 0.86), one with obligate grazers and variable grazer species (LD = 3.3), one with exclusively variable grazers (LD = 3.3), another with variable grazers and mixed feeder species (LD = 2.4), and a cluster with browser species (LD = 2.0). The bootstrap nodal support of these five main clusters range from 0.70 to 0.95, meanwhile the support of the equid species from southern Mexico ranges from 0.62 to 0.97 and those of the Pleistocene equids from other regions of Mexico varies from 0.68 to 0.93 (Figure 6).
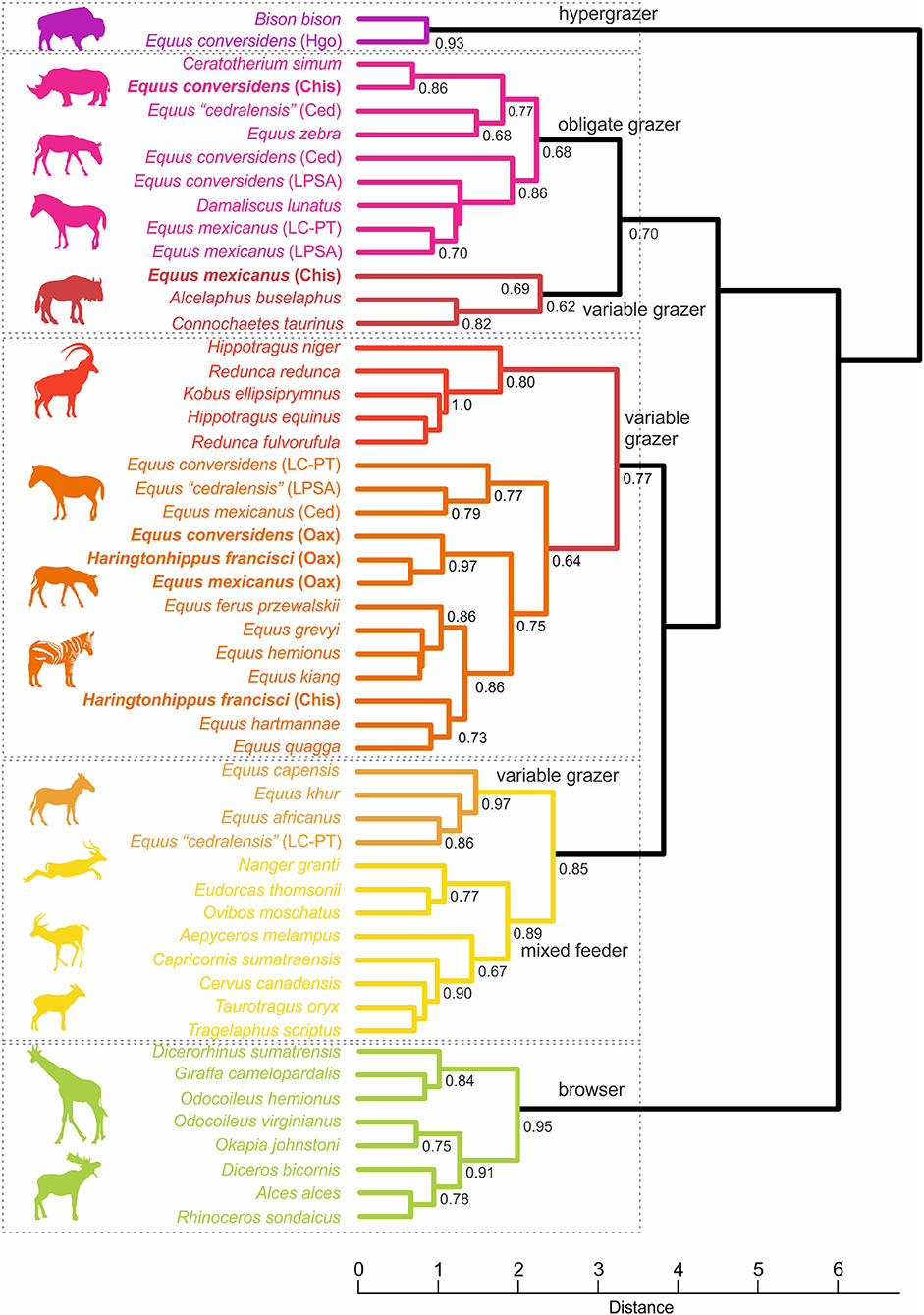
Figure 6. Hierarchical cluster of the mesowear analysis of diverse extant and extinct equid species and other extant herbivorous mammals. The equid species of this study are in bold. Hgo, Hidalgo; Chis, Chiapas; Ced, Cedral locality in San Luis Potosí; LPSA, La Piedad de Santana locality in Michoacán; LC-PT, La Cinta-Portalitos locality in Michoacán; Oax, Oaxaca.
Most of the studied samples are distributed in the clusters of variable grazers, except for the sample of Equus conversidens from Chiapas that is in the cluster of obligate grazers. None of the samples are in the cluster of the extant mixed-feeders or typical browsers.
The Oaxacan samples of Equus mexicanus, E. conversidens and H. francisci, showed a comparable occlusal relief and commonly have rounded cusps. These samples are joined in a group (LD = 1.0), which in turn integrates a subcluster with the Przewalski's wild horse E. ferus przewalskii, the giant wild ass E. kiang, the onager E. hemionus, and the zebras E. grevyi, E. hartmannae, and E. quagga (LD = 28); in this subcluster is H. francisci from Chiapas (LD = 1.2) (Figure 6).
The sample of Equus conversidens from Chiapas is in the cluster of the obligate grazers. It is grouped with the white rhinoceros Ceratotherium simum (LD = 0.73), integrating a subcluster that includes E. zebra and E. cedralensis from Cedral in north-central Mexico. The sample of Equus mexicanus from Chiapas is grouped with the variable grazers Alcelaphus buselaphus and Connochaetes taurinus (Figure 6).
It is important to note that given the small teeth samples (n = 3) of E. mexicanus from Chiapas and H. francisci from Oaxaca, these results are preliminary.
In the discriminant function analysis, the sample of E. conversidens from Chiapas was classified as obligate grazer with a posterior probability of 100%, meanwhile that from Oaxaca was classified as variable grazer with a probability of 99% (Figure 7). Haringtonhippus francisci from Chiapas was classified as variable grazer with a probability of 90% and that from Oaxaca was classified in the same guild with 99%. The samples of E. mexicanus from Chiapas and Oaxaca were classified as variable grazers with a posterior probability of 60 and 99%, respectively (Figure 7).
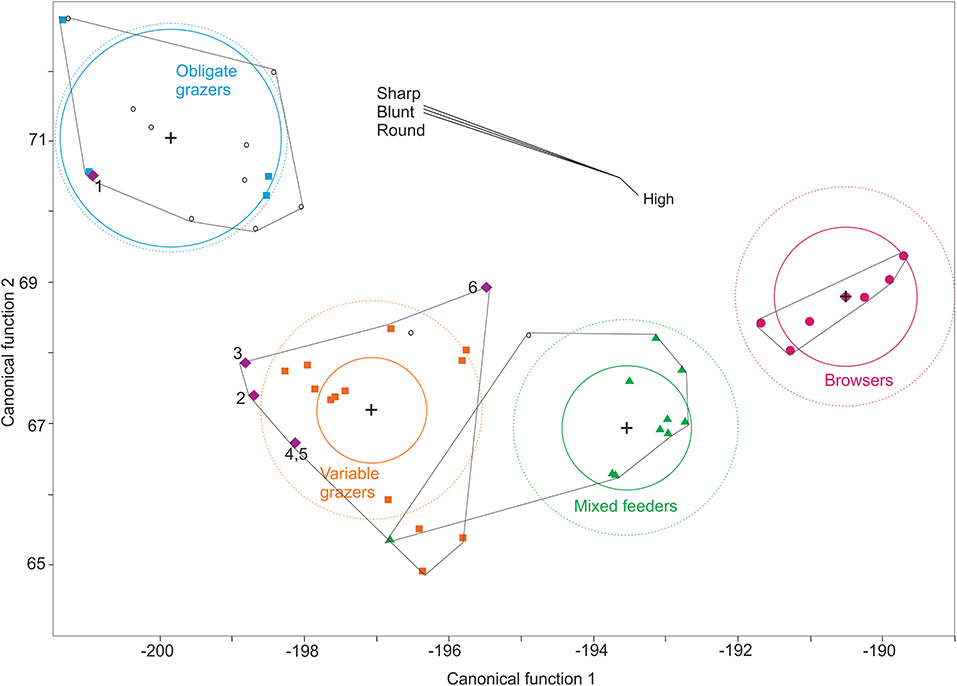
Figure 7. Canonical plot of the discriminant analysis of the mesowear data used in this paper. Chiapas: 1. Equus conversidens, 3. Haringtonippus francisci, 6. E. mexicanus. Oaxaca: 2. E. conversidens, 4. H. francisci, 5. E. mexicanus. Obligate grazers, blue squares; variable grazers, orange squares; mixed feeders, green triangles; browsers, fuchsia dots. Open circles are the Equus species from central and northern Mexico. Crosses show the centroid of each dietary group. In each dietary category, the inner circle represents the 95% confident region that contain the true mean of the group; the exterior dotted circle represents the region estimated to contain the 50% of the population for that group. The biplot lines (High, Sharp, Blunt, Round) indicate the coordinate directions in canonical space. Convex hulls for each diet category are outlined. Canonical function 1 explains 84.88% of the variation and canonical function 2 explains 14.99%.
The centroids of each dietary category showed statistically significant differences between groups (prob > F ≤ 0.0001).
Estimated Body Mass
The estimated body masses of the equid species from Chiapas and Oaxaca are found in Table S5. The highest estimations are those based on the length of p4, whereas similar body mass estimations resulted with the measurements of the second upper and lower molar.
The lowest body mass estimations are those of H. francisci (167–251 kg) and the highest are those of E. mexicanus (305–458.21 kg); E. conversidens has intermediate mass estimations (242.71–326.48 kg). There is a difference of 75.8–193 kg between the body mass estimations of E. mexicanus and E. conversidens, and of 51.6–75.5 kg between the estimations of E. conversidens and H. francisci (Table S5).
Discussion
Species Richness and Geochronology
Three horse species inhabited the territory that now constitutes the states of Chiapas and Oaxaca during the Late Pleistocene: the largest is Equus mexicanus, the medium-sized is E. conversidens, and the smallest is H. francisci.
The dating of several of the fossiliferous localities indicate that in Oaxaca, E. conversidens and E. mexicanus were present since at least 44,510–43,030 Cal BP, they were common between 30,123 and 19,584 Cal BP, and that E. conversidens persisted until 12,620–11,380 years BP. Haringtonhippus francisci was present in the Mixteca region of Oaxaca between 19,584 and 20,180 Cal BP. By the same token, the three identified equid species were present in Chiapas at least until 12,720 Cal BP.
The equids E. conversidens and E. mexicanus from Oaxaca are the oldest dated records in Mexico. Previously, both species were recovered from sediments that are around five meters above a 49,724 ± 2,074 Cal BP dated bed in Huexoyucán, Tlaxcala, in central Mexico (Tovar et al., 2013; Sánchez Salinas et al., 2016). Equus conversidens, E. mexicanus, and E. cedralensis have been recorded in sediments of around 24,000 Cal BP in La Cinta-Portalitos, west-central Mexico (Marín-Leyva et al., 2016; Díaz-Sibaja et al., 2018b), and they were also collected in beds bracketed by dates of 37,694 ± 1,963 years BP and 21,468 ± 458 Cal BP in Cedral, northern Mexico (Alberdi et al., 2014). Additionally, E. conversidens was recorded in strata having an age between 11,000 and 27,000 Cal BP in San Josecito Cave in northern Mexico (Arroyo-Cabrales et al., 1995; Arroyo Cabrales and Johnson, 2003), and it also has been collected in sediments that are three meters above a bed dated in 34,512 ± 220 Cal BP from Santa Cruz Nuevo, Puebla state, central Mexico (Tovar et al., 2014).
The dated record of H. francisci from Chiapas is the youngest in North America, since the Last Appearing Datum (LAD) from Eastern Beringia is 14.4 14C ka BP, the LAD in Gypsum Cave, Nevada is 13.1 14C ka BP (Heintzman et al., 2017).
Paleoecological Aspects
Dietary Guilds
The bootstrap values of the five main recovered clusters (hypergrazer, obligate grazer/variable grazer, variable grazer, variable grazer/mixed feeder, and browser) range from 0.70 to 0.95, whilst those for the Pleistocene equids from southern Mexico ranges from 0.62 to 0.97 (Figure 6); these values indicate a good nodal support for the recovered clusters, with the exception of 0.62, which can be considered as moderate (Zumel and Mount, 2014). Probably, the lower nodal support of 0.62 of E. mexicanus from Chiapas and the variable grazers is related to its small teeth sample (n = 3).
The Oaxacan samples of Equus mexicanus, E. conversidens, and H. francisci are clustered together and they integrate a subcluster with E. ferus przewalskii, E. kiang, E. hemionus, and the zebras E. grevyi, E. hartmannae, and E. quagga; in this subcluster also is H. francisci from Chiapas. The extant horses clustered with the Oaxacan samples are almost exclusively grazers, although the wild horse, the giant ass, and the onager can browse, whereas the zebras consume a wide variety of grasses (Kingdon, 2015; King et al., 2015). In the mesowear analysis of Schulz and Kaiser (2012), these extant equids cluster with the variable grazers Alcelaphus buselaphus and Connochaetes taurinus, but in our analysis both bovids formed a cluster with E. mexicanus from Chiapas; both bovids shows seasonal and geographical variability in their diet (Gagnon and Chew, 2000).
The sample of E. conversidens from Chiapas clusters in the obligate grazers (Figure 6), with the white rhinoceros Ceratotherium simum, integrating a subcluster that also includes E. cedralensis from Cedral. In the contiguous cluster is E. conversidens from Cedral, E. conversidens from La Piedad-Santa Ana, E. mexicanus from La Cinta-Portalitos and E. mexicanus from La Piedad-Santa Ana. The white rhinoceros is a typical pure grazer that consumes short and tall grasses depending on the season (Macdonald, 2006).
In our cluster analysis, the extant abrasion-dominated grazer species are at one end of the dendogram and the attrition-dominated browsers are at the other end; two extant variable grazer species (Equus africanus and E. khur) clusters with the mixed-feeders and two other species (Alcelaphus buselaphus and Connochaetes taurinus) cluster with the obligate grazers; in between there is a cluster composed of exclusively variable grazers (Figure 6). E africanus and E. khur have a higher proportion of high valleys and sharp edges compared to A. buselaphus and C. taurinus (Table S1); this indicates that these Equus species have a less abrasive diet than the bovids.
The topology of our dendrogram seems to reflect how abrasive are the dietary resources taken by the considered ungulate species. Thus, E. conversidens from Chiapas should had a more abrasive diet that E. mexicanus from Chiapas, and both should have a more abrasive diet than the Pleistocene equids from Oaxaca; H. francisci from Chiapas should had the least abrasive diet (Figure 6).
As can be observed, the four Pleistocene equid species from Mexico show a dietary plasticity, they can be an obligate grazer in some regions of the country, or variable grazer in another regions (Figure 6).
Regarding the discriminant analysis, the results showed a high percentage of correct classification within dietary guilds (97.1%). After performing the cross-validation analysis this high percentage of correct classification persist (94%) (Table S5). Except for E. conversidens from Chiapas, the rest of the samples were classified as variable grazers (Figure 7), and as was previously mentioned, there are statistically significant differences between the centroids of each guild. Equus mexicanus from Chiapas was classified as a variable grazer with a probability of 60% and as mixed feeder with a probability of 40%. The mesowear signal of E. conversidens from Chiapas as an obligate grazer is consistent with the results obtained by stable isotopes (Pérez-Crespo et al., 2015).
The extinct species Equus capensis was classified as a mixed- feeder with a probability of 80%. Regarding the species from central Mexico, E. cedralensis from La Cinta-Portalitos was classified as a variable grazer (99% of probability), E. conversidens from La Cinta-Portalitos was classified as a grazer (77% of probability) or variable grazer (23% of probability). The rest of the species were classified as obligate grazers (probability of 92–100%). This is in accordance to what has been previously published of these species in other regions of Mexico (see Table S1 for references).
Estimated Body Masses
The estimated body mass of Equus mexicanus from southern Mexico ranged from 305 to 458.21 kg, that for E. conversidens is 242.71–326.48 kg and the body mass of H. francisci ranged from 167 to 251 kg (Table S6). The estimated mean body size differences of these species are statistically significant for the M2 and p4 estimations (Table S7); the Mann-Whitney U-test could not be calculated with equation (2) given the small sample of m2's for H. francisci and E. mexicanus.
The mean estimated body mass based on M2 of E. conversidens is 23.52% less compared to that of E. mexicanus, and the estimated body mass of H. francisci is around 21% less than that of E. conversidens; for the mean body masses based on m2, the body mass of E. conversidens is 28.70% less than that of E. mexicanus and the one of H. francisci is 23% less than the one of E. conversidens.
Body mass is highly linked with the ecological adaptations such as diet of the herbivorous mammals (Maiorama, 1990); hence, the estimated body mass differences between the identified equid species from the Late Pleistocene of Chiapas and Oaxaca, probable diminished the resources competition between them, allowing to inhabit common areas, at least during some periods of time.
Here, estimated body masses values are like those previously reported for E. conversidens of 229.5 kg from La Piedad-Santana and 307.9 kg from La Cinta-Portalitos, and the body mass of E. mexicanus (476.7 kg) from La Piedad-Santana, in west-central Mexico, all based on measurements of postcranial bones (Marín-Leyva et al., 2016). These authors also reported a body mass of 126 and 145.5 kg for E. cedralensis, the other small-sized equid.
The body masses of the equids from Oaxaca and Chiapas are also like the previous estimations derived from Cedral specimens of northern Mexico: 354.33–532.28 kg for E. mexicanus, 215.63–282.52 kg for E. conversidens, and 91.83–169.59 kg for E. cedralensis (Alberdi et al., 2014).
Paleoenvironments and Habitats of Pleistocene Equids From Southern Mexico
Evidence for slickensides in the A horizon of Sandage locality of Coixtlahuaca, suggests that dry-wet seasonality was important around 30,000 CAL BP. A thick A horizon may reflect fast turnover typical of grasslands or result from a cumuli environment. Thick, dark A horizons with slickensides are characteristic of some Mollisols, which typically develop under grasslands, although in some cases they have been observed forming in marshes or under forests (Soil Survey Staff, 1999).
The textural features of the fossiliferous sediments from San Antonio Acutla, Santiago Teotongo and the sandy sediments of Tejupam, indicate that they were deposited as part of a perennial, very low energy fluvial system during some time; additionally, the presence of caliche or nodular calcretes in some beds suggests low humidity or precipitation during other times. By the same token, the silty clayey beds of Tejupam indicate the presence of some perennial water bodies (marshes) during some time intervals.
The associated fauna of San Antonio Acutla, Río Salado, and Tejupam localities of Oaxaca, include extant ostracod, charophyte and gastropod species that now live in quiet and shallow water, litter and deciduous forests (Pilsbry, 1946; Turthill et al., 1964; Schultz and Cheatum, 1970), salamanders that live in shallow ponds, with abundant aquatic vegetation (Duellman and Trueb, 1994); rodents that inhabit dry, semiarid hills to montane forests or deserts, salt marshes, and pine-oak forests (Davis and Follansbee, 1945; Webster and Knox, 1982; Fernández et al., 2010), open woodland or shrub vegetation types (Cornely and Baker, 1986).
This associated fauna suggests that in these Oaxacan fossil localities there were patchy habitats with grasses and weedy habitats, and temperate forests composed by conifer species with shrubs and grasses. Leaf litter allowed the conditions for terrestrial gastropods. Perennial or intermittent water bodies provided the habitat for the freshwater mollusks, which is also indicated by the stratigraphy. These heterogeneous habitats allowed the maintenance of diverse herbivore mammals, including the equids.
In Chiapas, the fine-grained fossil sediments of Los Mangos locality indicate that they were deposited in a water pond that later changed to a low flow fluvial regime. The associated Glyptotherium and Neochoerus suggest a perennial body water with subaqueous vegetation.
Geographic Distribution of Mexican Equids During the Late Pleistocene
The described equid species had a wide geographic distribution in Mexico during the Late Pleistocene, with most of the localities above 1,000 masl, in the Mexican highlands (Figure 8).
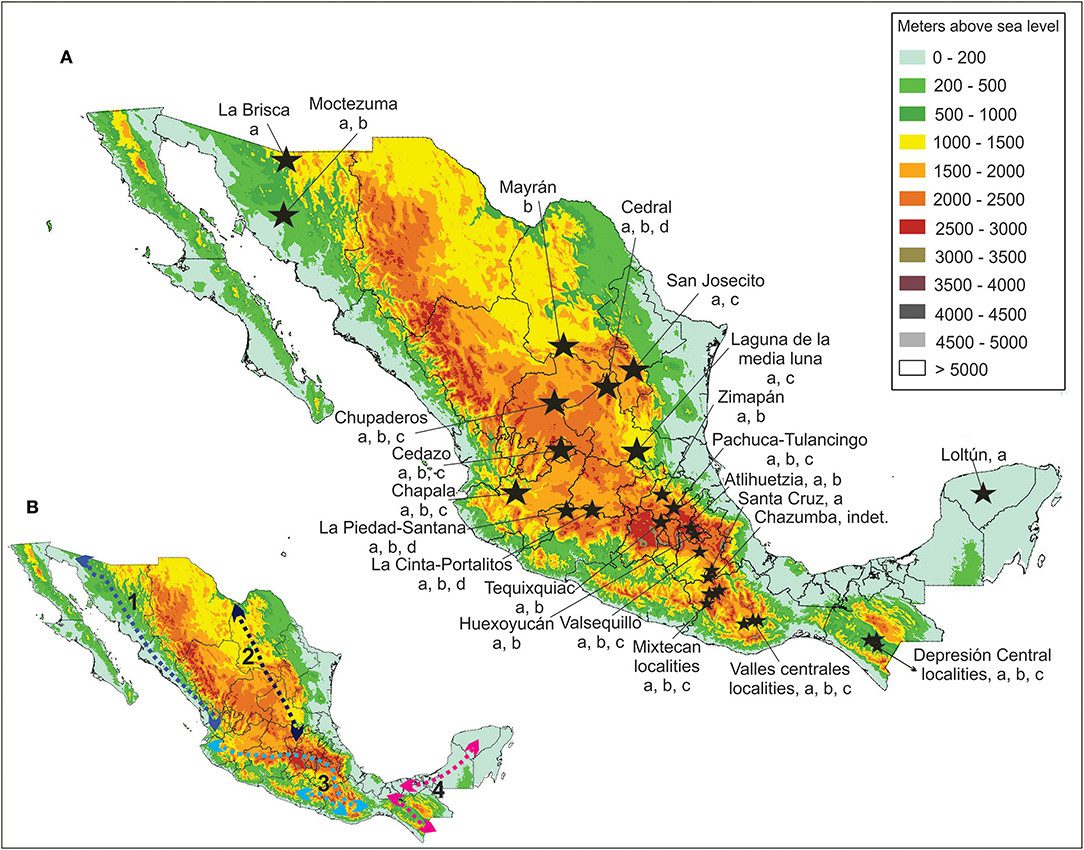
Figure 8. (A) Main Late Pleistocene Mexican equid localities. a, Equus conversidens; b, E. mexicanus; c, Haringtonhippus francisci; d, E. cedralensis. Data from Priego-Vargas et al. (2017). (B) Mexican biogeographic corridors used by Pleistocene equids. 1. Sonora, Central America Pacific lowlands; 2. Rocky Mountains, Sierra Madre Occidental; 3. Transvolcanic Belt, Sierra Madre del Sur; 4. Tamaulipas, Central America Gulf Lowlands.
The climatic oscillations of the Late Pleistocene, as well as the rugged terrain of the Mexican highlands, generated heterogeneous habitats all along the country (Mastretta-Yanes et al., 2015); which, as was described before, in Chiapas and Oaxaca consisted of some areas with grasses, woodlands and forests, with fluvial systems and permanent water bodies during the last 44,000 years. These patchy habitats allowed a rich array of herbivorous and carnivorous mammals to inhabit in Mexico during the Pleistocene.
The geographic distribution of the Late Pleistocene equid fossil localities indicate that Equus mexicanus, E. conversidens and H. francisci could move across Mexico by the Rocky Mountains—Sierra Madre Occidental and the Transvolcanic Belt—Sierra Madre del Sur temperate corridors, and the Sonora—Central America Pacific lowlands and the Tamaulipas—Central America Gulf Lowlands tropical corridors (see Ceballos et al., 2010).
Conclusions
Three Pleistocene equid species were identified in southern Mexico, the largest is Equus mexicanus, the medium-sized is E. conversidens and the smallest is H. francisci. In Oaxaca E. conversidens and E. mexicanus were present since at least 43,000 Cal BP years and E. conversidens persisted until 12,620–11,380 years BP. These three identified equid species were present in Chiapas at least until 12,720 Cal BP. The record of H. francisci from Chiapas is the youngest in North America.
The studied equid specimens from Chiapas and Oaxaca were mainly variable grazers, meanwhile those from central or northern Mexico were mainly obligate grazers. This indicates a dietary plasticity for Equus mexicanus, E. conversidens, and H. francisci; this plasticity has already been observed for the first two species in central and northern Mexico. In our analysis H. francisci from southern Mexico and E. cedralensis from Michoacan had similar dietary habits.
In southern Mexico, the identified equid species could move across the Transvolcanic Belt—Sierra Madre del Sur temperate corridor and the Tamaulipas—Central America Gulf Lowlands tropical corridor during the Late Pleistocene.
As in other regions of Mexico, these equids species were a common component of the Late Pleistocene landscapes of southern Mexico.
Data Availability Statement
All datasets generated for this study are included in the manuscript/Supplementary Files.
Author Contributions
EJ-H and GC-C conceived the study. EJ-H wrote the manuscript with the input of all the authors. RG-A, GC-C, EJ-H, produced the figures. EJ-H and GC-C measured the specimens. II-A and GH described the stratigraphy of the fossil localities and dated some localities. RG-A described the associated fauna and their habitat preferences. VB-C, GC-C, and EJ-H described and interpreted the mesowear cluster. EJ-H performed the discriminant analysis. All authors contributed to the interpretations of results and to editing of the text.
Funding
This paper was part of Conacyt-Ciencia Básica 2008 project N°101626, Conacyt-Ciencia Básica 2015 projects 255883 and 257647. Chiapas Goverment funded the collecting of specimens through project Prospección y Resguardo del Patrimonio Paleontológico de Chiapas.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor and reviewer, ES, declared their involvement as co-editors in the Research Topic, and confirm the absence of any other collaboration.
Acknowledgments
We acknowledge the kind invitation of G. Semprebon, R. Bernor, L. Avilla, F. Rivals, and E. Scott to participate in this Research Topic. Thanks to M. Díaz de León for his help with the R software and to the students that helped during the collecting of the Equus fossils. G. Goeckner provided proofreading assistance. L. Gómez-Pérez took the pictures of the Chiapas specimens. J. E. Hernández-Cortez drew Figure 2. We acknowledge the comments of two Reviewers that helped to improve this paper.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00394/full#supplementary-material
Abbreviations
Teeth abbreviations are: C/c, Canines; I/I, Incisors; P/p, Premolars; M/m, Molars upper/lower. The number indicates the position of the tooth in the tooth row. Other abbreviations are: AMS, accelerator mass spectrometry, C14 dating; Cal BP, calendar years before present; l, left; LD, linkage distance; masl, meters above sea level; perBlunt, percentage of blunt cusps; perHigh, percentage of high occlusal relief; perLow, percentage of low occlusal relief; perRound, percentage of rounded cusps; perSharp, percentage of sharp cusps; r, right.
References
Alberdi, M. T., Arroyo-Cabrales, J., Marín-Leyva, A. H., and Polaco, O. J. (2014). Study of cedral horses and their place in the Mexican Quaternary. Rev. Mex. Cienc. Geol. 31, 221–237.
Arroyo Cabrales, J., and Johnson, E. (2003). Catálogo de los ejemplares tipo procedentes de la Cueva de San Josecito, Nuevo León, México. Rev. Mex. Cienc. Geol. 20, 79–93.
Arroyo-Cabrales, J., Johnson, E., Haas, H., De Los Rios-Paredes, M., Ralph, R. W., and Hartwell, W. T. (1995). First radiocarbon dates for San Josecito Cave, Nuevo Leon, Mexico. Quat. Res. 43, 255–258. doi: 10.1006/qres.1995.1026
Azzaroli, A. (1998). The genus Equus in North America: the Pleistocene species. Palaeontogr. Ital. 85, 1–60.
Barrón-Ortiz, C. I., Avilla, L. S., Jass, C. N., Bravo-Cuevas, V. M., Machado, H., and Mothé, D. (2019). What is Equus? Reconciling taxonomy and phylogenetic analyses. Front. Ecol. Evol. 7:343. doi: 10.3389/fevo.2019.00343
Barrón-Ortiz, C. I., Rodrigues, A. T., Theodor, J. M., Kooyman, B. P., Yang, D. Y., and Speller, C. F. (2017). Cheek tooth morphology and ancient mitochondrial DNA of late Pleistocene horses from the western interior of North America: implications for the taxonomy of North American Late Pleistocene Equus. PLoS ONE 12:8. doi: 10.1371/journal.pone.0183045
Barrón-Ortiz, C. R., Theodor, J. M., and Arroyo-Cabrales, J. (2014). Dietary resource partitioning in the Late Pleistocene horses from Cedral, north-central Mexico: evidence from the study of dental wear. Rev. Mex. Cienc. Geol. 31, 260–269.
Bravo-Cuevas, V. M., and Jiménez-Hidalgo, E. (2018). “Advances on the paleobiology of late Pleistocene mammals from central and southern Mexico,” in The Pleistocene, Geography, Geology and Fauna, eds G. Huard and J. Gareau (New York, NY: Nova Science Publishers),277–313.
Bravo-Cuevas, V. M., Jiménez-Hidalgo, E., and Priego-Vargas, J. (2011). Taxonomía y hábito alimentario de Equus conversidens (Perissodactyla, Equidae) del Pleistoceno tardío (Rancholabreano) de Hidalgo, Centro de México. Rev. Mex. Cienc. Geol. 8, 65–82.
Carbot-Chanona, G., Eng-Ponce, J., and Gómez-Pérez, L. E. (in press). Description of the Neochoerus specimens from the Late Pleistocene (Rancholabrean) of Chiapas, comments on the taxonomic identity of the fossil capybaras from other Mexican localities. Bol. Soc. Geol. Mex. 72, 1–17.
Carbot-Chanona, G., and Ovalles-Damián, E. (2013). “Registro de vertebrados fósiles,” in La biodiversidad en Chiapas: Estudio de Estado, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO)/Gobierno del Estado de Chiapas (Tuxtla Gutiérrez), 261–274.
Ceballos, G., Arroyo-Cabrales, J., and Ponce, E. (2010). Effects of Pleistocene environmental changes on the distribution and community structure of the mammalian fauna of Mexico. Quat. Res. 73, 464–473. doi: 10.1016/j.yqres.2010.02.006
Cornely, J. E., and Baker, R. J. (1986). Neotoma mexicana. Mamm. Species 262, 1–7. doi: 10.2307/3504054
Davis, W. B., and Follansbee, L. A. (1945). The mexican volcano mouse. Neotomodon. J. Mammal. 26, 401–411. doi: 10.2307/1375160
Díaz-Sibaja, R., Jiménez-Hidalgo, E., and García-Zepeda, M. L. (2018a). Una nueva localidad fosilífera en Oaxaca (México) y el registro más austral de Bison latifrons. Implicaciones paleobiogeográficas, paleoecológicas y paleoambientales. Bol. Soc. Geol. Mex. 70, 201–222. doi: 10.18268/BSGM2018v70n1a12
Díaz-Sibaja, R., Jiménez-Hidalgo, E., Ponce-Saavedra, J., and García-Zepeda, M. L. (2018b). A combined mesowear analysis of Mexican Bison antiquus shows a generalist diet with geographical variation. J. Paleontol. 92, 1130–1139. doi: 10.1017/jpa.2018.19
Doutt, J. K., and Black, C. C. (1962). A new Pleistocene locality in the State of Oaxaca, Mexico. J. Mammal. 43, 414–415. doi: 10.2307/1376950
Duellman, W. E., and Trueb, L. (1994). Biology of Amphibians. Baltimore: Johns Hopkins University Press.
Eisenmann, V. (2009). System of Measurements for Equus Bones and Teeth. Available online at: http://vera-eisenmann.com (accessed on December 14, 2018).
Eisenmann, V., and Mashkour, M. (2000). “Database for teeth and limb bones of modern hemiones,” in Fiches D'Osteologie Animale Pour L'Archéologie, Série B: Mammifères, eds J. Desse and N. Desse-Berset (Juan-les-Pins: Centre de Recherches Archéologiques du CNRS, APDCA), 1–46.
Evander, R. L. (2004). Chapter 16: a revised dental nomenclature for fossil horses. Bull. Amer. Mus. Nat. Hist. 285, 209–218. doi: 10.1206/0003-0090(2004)285<0209:C>2.0.CO;2
Fernández, J. A., García-Campusano, F., and Hafner, M. S. (2010). Peromyscus difficilis (Rodentia: Cricetidae). Mamm. Species 42, 220–229. doi: 10.1644/867.1
Fortelius, M., and Solounias, N. (2000). Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Am. Mus. Novit. 3301, 1–36. doi: 10.1206/0003-0082(2000)301<0001:FCOUMU>2.0.CO;2
Gagnon, M., and Chew, A. E. (2000). Dietary preferences in extant African Bovidae. J. Mammal. 81, 490–5112. doi: 10.1644/1545-1542(2000)081<0490:DPIEAB>2.0.CO;2
Gómez-Pérez, L. E., and Carbot-Chanona, G. (2012). Contribución al estudio de los megamamíferos del Pleistoceno Tardío del municipio de Villaflores, Chiapas, México. Lacandonia 6, 31–41.
Guerrero-Arenas, R., and Jiménez-Hidalgo, E. (2015). “Las comunidades del Pleistoceno tardío de la Mixteca alta oaxaqueña: cómo en pasado nos permite conservar el presente,” in El pasado lejano de la Mixteca, comp. ed R. O. Escamilla (Huajuapan de León: Universidad Tecnológica de la Mixteca, 25–38.
Hay, O. P. (1915). Contributions to the knowledge of the mammals of the Pleistocene of North America. Proc. U. S. National Mus. 48, 515–575. doi: 10.5479/si.00963801.48-2086.515
Heintzman, P. D., Zazula, G. D., Mac Phee, R. D. E., Scott, E., Cahill, J. A., McHorse, B. K., et al. (2017). A new genus of horse from Pleistocene North America. eLife 6, 1–43, doi: 10.7554/eLife.29944
Hibbard, C. W. (1955). Pleistocene vertebrates from the upper becerra formation, valley of tequixquiac, Mexico, with notes on other Pleistocene Forms. Univ. Mich. Contr. Mus. Paleontol. 21, 47–96.
Janis, C. (1990). “Correlation of cranial and dental variables with body size in ungulates and macropodoids,” in Body Size in Mammalian Paleobiology: Estimation and Biological Implications, ed J. Damuth and MacFadden B. J. (London, Cambridge University Press), 255–299.
Jiménez-Hidalgo, Cabrera-Pérez, L., Macfdden, B. J., and Guerrero-Arenas, R. (2013). First record of Bison antiquus from the late Pleistocene of southern Mexico. J. S. Am. Earth Sci. 42, 83–90. doi: 10.1016/j.jsames.2012.07.011
Jiménez-Hidalgo, E., Campos-Camacho, M. A., Guerrero-Arenas, R., and Cabrera-Pérez, L. (2012). The Late Pleistocene Equids From Northwestern Oaxaca, Southern Mexico. Geological Society of America, Abstracts with Programs 44, 10.
Jiménez-Hidalgo, Guerrero-Arenas, R., Macfdden, B. J., and Cabrera-Pérez, L. (2011). The late pleistocene (Rancholabrean) Viko Vijin local fauna from la Mixteca Alta, northwestern Oaxaca, southern Mexico. Rev. Bras. Paleontol. 14, 15–28. doi: 10.4072/rbp.2011.1.02
Kaiser, T. M., and Fortelius, M. (2003). Differential mesowear in occluding upper and lower molars: opening mesowear analysis for lower molars and premolars in hypsodonts horses. J. Morphol. 258, 67–83. doi: 10.1002/jmor.10125
Kaiser, T. M., and Franz-Odendaal, T. A. (2004). A mixed-feeding Equus species from the Middle Pleistocene of South Africa. Quat. Res. 62, 316–323. doi: 10.1016/j.yqres.2004.09.002
Kaiser, T. M., and Solounias, N. (2003). Extending the tooth mesowear method to extinct and extant equids. Geodiversitas 25, 321–345.
King, S. R. B., Boyd, L., Zimmermann, W., and Kendall, B. E. (2015). Equus ferus (Errata Version Published in 2016). The IUCN Red List of Threatened Species 2015: e.T41763A97204950. Available online at: http://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T41763A45172856.en
Lundelius, E. L., and Stevens, M. S. (1970). Equus francisci, a small stilt-legged horse, middle Pleistocene of Texas. J. Paleontol. 44, 148–153.
Macdonald, D. W. (2006). The Princeton Encyclopedia of Mammals. New Jersey, NJ: Princeton University Press.
MacFadden, B. J., and Carranza-Castañeda, O. (2002). Cranium of Dinohippus mexicanus (Mammalia: Equidae) from the early Pliocene (latest Hemphillian) of central Mexico, and the origin of Equus. Bull. Florida Mus. Nat. Hist. 43, 163–185.
Maiorama, V. C. (1990). “Evolutionary strategies and body size in a guild of mammals,” in Body Size in Mammalian Paleobiology, eds J. Damuth and B. J. MacFadden (New York, NJ: Cambridge University Press), 69–102.
Marín-Leyva, A. H., DeMiguel, D., García-Zepeda, M. L., Ponce-Saavedra, J., Arroyo-Cabrales, J., Schaaf, P., et al. (2016). Dietary adaptability of Late Pleistocene Equus from west central Mexico. Palaeogeogr. Palaeoclimatol. Palaeoecol. 441, 748–757. doi: 10.1016/j.palaeo.2015.10.019
Mastretta-Yanes, A., Moreno-Letelier, A., Pinero, D., Jorgensen, T. H., and Emerson, B. C. (2015). Biodiversity in the Mexican highlands and the interaction of geology, geography and climate within the Trans-Mexican Volcanic Belt. J. Biogeogr. 42, 1586–1600. doi: 10.1111/jbi.12546
Ordoñez-Regil, E., Almazán-Torres, M. G., Jiménez-Hidalgo, E., and Tenorio, D. (2016). Radiometric dating of Late Pleistocene mammal bones from La Mixteca Alta Oaxaqueña. J. Archaeol. Sci. Rep. 9, 160–167. doi: 10.1016/j.jasrep.2016.07.021
Pérez Crespo, A., Arroyo-Cabrales, J., and Santos Moreno, A. (2008). Generalidades de los mamíferos del Pleistoceno Tardío de Oaxaca. Nat. y Des. 6, 5–11.
Pérez-Crespo, V. A., Carbot-Chanona, G., Morales-Puente, P., Cienfuegos-Alvarado, E., and Otero, F. J. (2015). Paleoambiente de la Depresión Central de Chiapas, con base en isótopos estables de carbono y oxígeno. Rev. Mex. Cienc. Geol. 32, 273–282.
Pilsbry, H. A. (1946). Land Mollusca of North America (North of Mexico). Philadelphia, PA Academy of Natural Sciences.
Priego-Vargas, J., Bravo-Cuevas, V. M., and Jiménez-Hidalgo, E. (2017). Revisión taxonómica de los équidos del Pleistoceno de México con base en la morfología dental. Rev. Bras. Paleontol. 20, 239–268. doi: 10.4072/rbp.2017.2.07
R Core Team (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/
Reynoso-Rosales, V. H., and Montellano-Ballesteros, M. (1994). Revisión de los équidos de la Fauna local de Cedazo del Pleistoceno de Aguascalientes México. Rev. Mex. Cienc. Geol. 11, 87–105.
Sánchez Salinas, M., Jiménez Hidalgo, E., and Castañeda Posadas, C. (2016). Mamíferos fósiles del Pleistoceno tardío (Rancholabreano) de San Mateo Huexoyucán, Tlaxcala, México. Bol. Soc. Geol. Mex. 68, 497–514.
SAS Institute (2019). JMP 8.0. Available online at: https://www.jmp.com.
Schultz, G. E., and Cheatum, E. P. (1970). Bison occidentalis and associated invertebrates from the Late Wisconsin of Randall County, Texas. J. Paleontol. 44, 836–850.
Schulz, E., and Kaiser, T. M. (2012). Historical distribution, habitat requirements and feeding ecology of the genus Equus (Perissodactyla). Mammal. Rev. 43, 111–123. doi: 10.1111/j.1365-2907.2012.00210.x
Soil Survey Staff (1999). Soil Taxonomy a Basic System of Soil Classification for Making and Interpreting Soil Surveys. Washington, DC: United States Department of Agriculture, Agriculture Handbook, 436.
Tovar, R. E., Sedov, S., Montellano-Ballesteros, M., and Solleiro, E. (2014). The alluvial sequence of Santa Cruz Nuevo, Mexico: paleoecological interpretation of fossil vertebrates and pedosediments from marine isotope stage 3. Rev. Mex. Cienc. Geol. 31, 407–419.
Tovar, R. E., Sedov, S., Solís, B., and Solleiro, E. (2013). Dark humic alluvial paleosols in Central and Southern Mexico: micromorphological indicators of Late Pleistocene megafauna habitats. Span. J. Soil Sci. 3, 217–235. doi: 10.3232/SJSS.2013.V3.N3.06
Turthill, S. J., Clayton, L., and Laird, W. M. (1964). A comparison of a fossil Pleistocene molluscan fauna from North Dakota with a recent molluscan fauna from Minessota. Am. Midl. Nat. 71, 344–362. doi: 10.2307/2423292
Viñas-Vallverdú, R., Arroyo-Cabrales, J., Rivera-González, I. I., Xosé Pedro, R. Á, Rubio-Mora, A., Eudave-Eusebio, I. N., et al. (2017). Recent archaeo-palaeontological findings from Barranca del Muerto site, Santiago Chazumba, Oaxaca, México. Quat. Int. 431, 168–180. doi: 10.1016/j.quaint.2015.04.055
Webster, D., and Knox, D. (1982). Reithrodontomys megalotis. Mamm. Species 167, 1–5. doi: 10.2307/3504020
Keywords: Equus, Pleistocene, Mexico, Oaxaca, Chiapas, paleoecology, taxonomy
Citation: Jiménez-Hidalgo E, Carbot-Chanona G, Guerrero-Arenas R, Bravo-Cuevas VM, Holdridge GS and Israde-Alcántara I (2019) Species Diversity and Paleoecology of Late Pleistocene Horses From Southern Mexico. Front. Ecol. Evol. 7:394. doi: 10.3389/fevo.2019.00394
Received: 29 January 2019; Accepted: 04 October 2019;
Published: 18 October 2019.
Edited by:
Leonardo Santos Avilla, Universidade Federal Do Estado Do Rio de Janeiro, BrazilReviewed by:
Manuel F. G. Weinkauf, Université de Genève, SwitzerlandEric Scott, California State University, San Bernardino, United States
Copyright © 2019 Jiménez-Hidalgo, Carbot-Chanona, Guerrero-Arenas, Bravo-Cuevas, Holdridge and Israde-Alcántara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eduardo Jiménez-Hidalgo, eduardojh@zicatela.umar.mx; Gerardo Carbot-Chanona, gfcarbot@gmail.com