 Alma L. Saucedo1†
Alma L. Saucedo1† Eric E. Hernández-Domínguez1†
Eric E. Hernández-Domínguez1† Luis A. de Luna-Valdez2
Luis A. de Luna-Valdez2 Angel A. Guevara-García2
Angel A. Guevara-García2 Abraham Escobedo-Moratilla1
Abraham Escobedo-Moratilla1 Esaú Bojorquéz-Velázquez1
Esaú Bojorquéz-Velázquez1 Federico del Río-Portilla3
Federico del Río-Portilla3 Daniel A. Fernández-Velasco4
Daniel A. Fernández-Velasco4 Ana P. Barba de la Rosa1*
Ana P. Barba de la Rosa1*- 1Department of Molecular Biology, Instituto Potosino de Investigación Científica y Tecnológica, A.C., San Luis Potosí, México
- 2Instituto de Biotecnología, Universidad Nacional Autónoma de México, Cuernavaca, México
- 3Instituto de Química, Universidad Nacional Autónoma de México, Ciudad de México, México
- 4Laboratorio de Fisicoquímica e Ingeniería de Proteínas, Departamento de Bioquímica, Facultad de Medicina, Universidad Nacional Autónoma de México, Ciudad de México, México
Late embryogenesis abundant (LEA) proteins are part of a large protein family that protect other proteins from aggregation due to desiccation or osmotic stresses. Recently, the Amaranthus cruentus seed proteome was characterized by 2D-PAGE and one highly accumulated protein spot was identified as a LEA protein and was named AcLEA. In this work, AcLEA cDNA was cloned into an expression vector and the recombinant protein was purified and characterized. AcLEA encodes a 172 amino acid polypeptide with a predicted molecular mass of 18.34 kDa and estimated pI of 8.58. Phylogenetic analysis revealed that AcLEA is evolutionarily close to the LEA3 group. Structural characteristics were revealed by nuclear magnetic resonance and circular dichroism methods. We have shown that recombinant AcLEA is an intrinsically disordered protein in solution even at high salinity and osmotic pressures, but it has a strong tendency to take a secondary structure, mainly folded as α-helix, when an inductive additive is present. Recombinant AcLEA function was evaluated using Escherichia coli as in vivo model showing the important protection role against desiccation, oxidant conditions, and osmotic stress. AcLEA recombinant protein was localized in cytoplasm of Nicotiana benthamiana protoplasts and orthologs were detected in seeds of wild and domesticated amaranth species. Interestingly AcLEA was detected in leaves, stems, and roots but only in plants subjected to salt stress. This fact could indicate the important role of AcLEA protection during plant stress in all amaranth species studied.
Introduction
Seeds can withstand the loss of cellular water during the maturation phase of their development by the accumulation of high levels of ubiquitous proteins named late embryogenesis abundant (LEA) proteins (Ali-Benali et al., 2005; Dalal et al., 2009; Liu et al., 2013; Avelange-Macherel et al., 2015). LEA proteins were originally discovered in cotton (Gossypium hirsutum) seeds (Dure, 1989), but their accumulation is not only related to the development of desiccation tolerance in orthodox seeds (desiccation-tolerant seeds). LEA proteins are also induced upon water-related stress in plant vegetative tissues and in other anhydrobiotic organisms such as eubacteria, rotifers, nematodes, tardigrades, arthropods (Ingram and Bartels, 1996; Browne et al., 2002; Hundertmark and Hincha, 2008; Campos et al., 2013; Hatanaka et al., 2014; van Leeuwen et al., 2016). In some microorganisms, LEA proteins are reported in response to water limitation, which suggests that they have an important role in desiccation tolerance (Tunnacliffe and Wise, 2007; Tunnacliffe et al., 2010; Hand et al., 2011). In spite of their widely recognized importance for desiccation tolerance, the molecular function of LEA proteins is only starting to emerge, with a variety of functions in agreement with their diversity (Battaglia and Covarrubias, 2013).
The distinctive features of LEA proteins are their high hydrophilicity due to a high percentage of charged amino acids such as alanine, serine/threonine and the absence or very low content of non-polar amino acids (tryptophan and cysteine). The presence of repeated motifs, which tend to form secondary structures, has detected in LEA proteins (Dure, 1989; Garay-Arroyo et al., 2000; Tunnacliffe and Wise, 2007). Although LEA proteins are intrinsically disordered proteins (IDP) in aqueous solutions (Wolkers et al., 2001; Goyal et al., 2003; Boucher et al., 2010; Tompa and Kovacs, 2010; Popova et al., 2011), they may acquire some structure folding into α-helical conformations during partial or complete dehydration (Shih et al., 2004; Tolleter et al., 2007; Hincha and Thalhammer, 2012).
Several hundreds of LEA protein sequences have been gathered in a dedicated database1 and bioinformatics analyses have shown that each LEA class can be clearly characterized by a unique set of physico-chemical properties. This has led to the classification of LEA proteins into 12 non-overlapping classes with distinct properties (Battaglia et al., 2008; Hunault and Jaspard, 2010; Jaspard et al., 2012).
Although quite a few LEAs have been characterized, the functions of most members of the LEA family remain unknown (Cao and Li, 2015). Transgenic Arabidopsis thaliana plants overexpressing the Nicotiana tabacum NtLEA7-3 gene are much more resistant to cold, drought, and salt stresses (Gai et al., 2011). Tomato LEA25 increases the salt and chilling stress tolerance when overexpressed in yeast (Imai et al., 1996). Wheat and rice over-expressing HVA1 gene (encoding an LEA protein from barley) are more tolerant to drought and salt stress (Xu et al., 1996; Sivamani et al., 2000). Olvera-Carrillo et al. (2010) reported that in A. thaliana, the accumulation of AtLEA4 protein leads to a drought tolerant phenotype. The overexpression of BnLEA4-1 from Brassica napus in Escherichia coli can enhance bacterial cellular tolerance to temperature and salt stresses (Dalal et al., 2009).
On the other hand, LEA proteins have a broad subcellular distribution; they are present in cytosol, mitochondria, chloroplasts, endoplasmic reticulum, and nucleus (Candat et al., 2014) and the specific modes of their action could be related to their intracellular location. The biological activity of these proteins seems to be associated with the stabilization of membranes during cell drying (Tolleter et al., 2010), and assistance of the transport of proteins during stress conditions (Chakrabortee et al., 2010).
Amaranth, a member of Amaranthaceae family, is a plant that has been cultivated and used since ancient times by Mexican and Central American civilizations. In the last decades, the nutritional role of amaranth seeds from different species has been revalued, particularly for A. hypochondriacus and A. cruentus, not only because of their high protein content and their contribution of essential amino acids, like lysine and methionine (compared to other grains), but also for their antioxidant compounds (Becker et al., 1981; Rastogi and Shukla, 2013), and bioactive peptides (Silva-Sánchez et al., 2008). Current interest in amaranth plants is also related to their extraordinary adaptability to grow in adverse weather conditions (Brenner et al., 2000). Amaranth is resistant against several types of stresses such as pest (Valdes-Rodríguez et al., 2007), heat (Maughan et al., 2009), drought (Huerta-Ocampo et al., 2011), and salinity (Aguilar-Hernández et al., 2011; Huerta-Ocampo et al., 2014). The recent report of Amaranthus cruentus seed proteome by 2D-PAGE revealed the over-accumulation of one spot identified as a LEA protein (Maldonado-Cervantes et al., 2014). In the present study, we have cloned the corresponding LEA cDNA from A. cruentus (AcLEA, GenBank accession no. KX852451), and the recombinant protein was expressed in E. coli. Nuclear Magnetic Resonance (NMR) and Circular Dichroism (CD) were used as tools to study the structural characteristics of this particular AcLEA protein. Its functional activity was evaluated in vivo using E. coli as model.
According to its amino acid sequence, AcLEA protein belongs to the Group 3, its hydrophilic nature and spectroscopic characteristics being ad hoc with IDP molecules, but exhibiting a high content of α-helix in the presence of trifluoroethanol (TFE). Overexpression of AcLEA in E. coli conferred resistance to desiccation, osmotic and oxidative stress to the bacterial cells. When accumulated in a heterologous system (Nicotiana benthamiana protoplasts) the amaranth protein was found to be distributed in the cytoplasm of protoplasts. Western blot analyses disclosed that AcLEA protein accumulated in seeds of wild and domesticated amaranth species. Accumulation of AcLEA in leaves, stems, and roots was observed only in plants subjected to salinity stress.
Materials and Methods
RNA Extraction and Cloning of the cDNA Encoding AcLEA
Immature seeds (15 days after anthesis) of Amaranthus cruentus were used to extract total RNA with TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) and cDNA was synthesized as previously reported (Maldonado-Cervantes et al., 2014). AcLEA cDNA was amplified using specific primers containing NdeI (5′-CATATGGCATCACATGGTCAGAGT-3′) and XhoI (5′-CTCGAGCTAGGGCCTAGTAGTCTTAATTGGATC-3′) restriction sites. cDNA amplification was performed using Platinum Taq DNA polymerase (Invitrogen), under standard reaction conditions. The amplified PCR product was cloned into the plasmid pGEM-T-Easy (Promega Corp., Madison, WI, USA). AcLEA cDNA was excised from pGEM using NdeI and XhoI (New England Biolabs, Ipswich, MA, USA) restriction enzymes. Digested fragments were purified and subcloned into pET28 expression vector restricted with NdeI-XhoI (Novagen-Merck, Darmstadt, Germany) containing the His-Tag at N-terminal. Vector was modified to have a recognition cleavage site within the amino acid sequence LeuGluValLeuPheGln/GlyPro specific for human rhinovirus 3C protease as PreScission Protease (PSP), and ending with the pET28mod vector. The resulting plasmid pET28mod-AcLEA was sequenced in both directions to confirm the AcLEA cDNA identity.
Alternatively, the AcLEA cDNA was PCR flanked with attB1 and attB2 recombination sites for generation of an entry clone using the gateway system entry vector pDONR-Zeo (Karimi et al., 2007), which was later used to generate the expression vector pEarlyGate 103-AcLEA.
Physicochemical Properties and Phylogenetic Analyses
Protein hydrophilicity analysis was performed to obtain the hydropathy plots with the Kyte and Doolittle (1982) values from the Expasy ProtScale Tool2 (Gasteiger et al., 2005). Grand average of hydropathicity (GRAVY) and instability index were calculated using the ProtParam software3. Sequence similarities were determined using the BLAST program and the GenBank database on the NCBI web server. MUSCLE 3.8.31 (Edgar, 2004) was used to perform multiple sequence alignments of full-length protein sequences. The phylogenetic analyses of the LEA proteins based on amino acid sequences were carried out using the neighbor-joining method (Saitou and Nei, 1987). AcLEA protein classification was done comparing its sequence to those available in the LEA Proteins Data Base4 (Hunault and Jaspard, 2010) and the Pfam server5 (Finn et al., 2015).
AcLEA related protein sequences were retrieved from the recently reported genome of Amaranthus hypochondriacus (Clouse et al., 2016) deposited at Phytozome v12.06.
Expression and Purification of the Recombinant AcLEA Protein
Recombinant AcLEA protein (rAcLEA) was up-accumulated in BL21 (DE3) E. coli cells (Novagen) transformed with the expression vector pET28mod-AcLEA. LB media supplemented with kanamycin was used to grow cells at 37°C. Overnight cultures were diluted 100-fold using fresh LB medium, and incubation was continued until optical density (OD600) reached 0.5–0.6. At this point, 0.1 mM isopropyl thio-β-D-galacto-pyranoside (IPTG, Sigma–Aldrich, St. Louis, MO, USA) was added to induce the protein expression. After further 4 h of incubation at 28°C, cells were harvested by centrifugation at 3,000 × g for 15 min at 4°C.
For structural studies cell pellets were resuspended in native buffer (150 mM NaCl, 50 mM Tris-HCl, pH 8) and for antibodies production cells pellets were resuspended in denaturing lysis buffer (500 mM NaCl, 6 M guanidine hydrochloride, 20 mM sodium phosphate, pH 7.8). Resuspended pellets were sonicated for 45 s (Misonix Sonicator 3000, Cole-Parmer, Vernon Hills, IL, USA) in ice bath. Antibodies were obtained as described in Supplementary Information. The soluble fraction was separated by centrifugation at 20,000 × g for 30 min at 4°C. Recombinant six-His-tagged AcLEA (rHis-AcLEA, 20.7 kDa) was purified by metal-chelate affinity chromatography (IMAC) using the Ni-NTA agarose purification system (Novex, Thermo Fischer Scientific Inc., Waltham, MA, USA), and eluted with five volumes of native (150 mM NaCl, 50 mM Tris-HCl, pH 8.0) or denaturing elution buffer (500 mM NaCl, 8 M urea, 20 mM sodium phosphate, pH 4.0). In both native and denaturing purifications, buffer exchange to 150 mM NaCl, 50 mM Tris-HCl, pH 8.0, was performed by dialysis using a 5 kDa cut-off membrane (Merck Millipore, Billerica, MA, USA), and cleavage of His-Tag was carried out overnight at 4°C. After cleavage, a second step of IMAC purification was carried out under native conditions (150 mM NaCl, 50 mM Tris-HCl, pH 8.0) in order to obtain native rAcLEA. Since rAcLEA was found to be weakly bounded to the resin, a native buffer containing 20 mM imidazole was used for protein elution. Finally, PD10 desalting columns (GE Healthcare, Piscataway, NJ, USA) were used to remove buffer components. For NMR spectroscopic and CD analyses, an additional purification step of rAcLEA was done using FPLC chromatography with a Sephacryl S-100 column (GE Healthcare) with a mobile phase of 10 mM sodium phosphate pH 7.0 (Sigma–Aldrich). All rAcLEA purification steps were followed by 12% SDS-PAGE gels stained with Coomassie Blue.
Recombinant proteins, excised from gel and/or in solution after chromatography purification were reduced with 10 mM DTT followed by protein alkylation with 55 mM iodoacetamide, and finally digested with trypsin (Promega, Madison, WI, USA) in an overnight reaction at 37°C. MS was carried out with a SYNAPT-HDMS (Waters Corp.) coupled to a nano-ACQUITY-UPLC system as described in Supplementary Information.
Structural and Functional Characteristics of AcLEA
NMR and CD Analyses
Lyophilized rAcLEA purified under native conditions was dissolved in H2O/D2O (95:5) to prepare a solution at final concentration of 1 mM, and transferred to a 3 mm tube. For 1H-NMR water suppression signal was performed using the double-pulsed field gradient spin echo sequence (DPFGSE). Fourier transformation was applied to FID file and data were analyzed with the NUTS Data Processing Software (Acorn NMR Inc., Livermore, CA, USA). Proton nuclear magnetic resonance spectra (1H-NMR) were acquired on a 500 MHz Varian Innova spectrometer (Varian, Palo Alto, CA, USA) at 298 K.
Circular dichroism spectra were recorded on a Chirascan Circular Dichroism Spectrometer (Applied Photophysics, Leatherhead, UK), equipped with a Peltier cell holder for control of temperature. A stock solution of rAcLEA protein (0.4 mg/ml) was prepared in 10 mM phosphate buffer pH 8.0. Far UV CD spectra were obtained using a quartz cell with a light path of 1 mm in the 200-260 nm range with a bandwidth of 1.0 nm and a digital resolution of 0.5 s per point. Temperature-induced conformational changes were simultaneously recorded at 210, 222 and 230 nm using a heating rate of 1°C/min in the 20 to 70°C range. After the heating ramp, the sample was cooled to 20°C then far UV CD spectra was taken to determine the reversibility of the conformational changes. CD spectra in the near UV region covering the 250-350 nm range were recorded using a quartz cell with a 10 mm path length, bandwidth of 2.0 nm and 1.0 s time per point. Molar ellipticity values, [θ], were calculated from measured θ using the equation:
where θ is the measured ellipticity in degrees, M is the protein molecular weight, c is the protein concentration in mg/ml, and l is the path length. Estimation of secondary structure was performed using the CDNN algorithm (Bohm et al., 1992) using a spectral window data from 200 to 260 nm. Five spectra were recorded for each experimental condition.
Assay of Protective Role of AcLEA in E. coli
Transformed E. coli BL21 cells carrying the plasmid pET28mod-AcLEA and the empty plasmid pET28mod (control) were grown in LB liquid medium supplemented with 37 μg/ml kanamycin overnight at 37°C. For both bacterial cultures, an aliquot was diluted 100-fold using fresh liquid LB with antibiotic and allowed to grow for 2–3 h at 37°C. When OD600 reached 0.5–0.6, IPTG was added to a final concentration of 0.1 mM and cultures were kept at 28°C for 2 h, for rAcLEA protein induction. At this point stress treatments were analyzed. To test the function of AcLEA protein to prevent desiccation stress, E. coli cells were dried at 40°C for 2 h in a flat plates under the laminar flow hood. After drying, cells were rehydrated in 200 μl of liquid LB media. Re-suspended cells were spread over Petri dishes containing LB, antibiotic, and IPTG, then were incubated overnight at 37°C. The number of colony former units (CFU) was used to compare viability (Dalal et al., 2009; He et al., 2012). Salinity stress was assessed with different concentrations of NaCl (0.4, 0.6, and 0.8 M). Sorbitol (0.6, 0.8, and 1.0 M) and PEG 4000 (5, 10, and 20% w/v) were used to decrease osmotic potential and mimic dehydration, and H2O2 (0.1, 0.5, and 1.0 M) was tested to promote oxidant conditions. In all experiments, absorbance at 600 nm (OD600) was used to measure the bacterial growth in liquid media (Liu and Zheng, 2005; Wu et al., 2014; Hu et al., 2016). All experiments were carried out in three biological replicates each replicate was done at least three times.
Localization In vivo Using Nicotiana benthamiana Protoplasts
The expression vector pEarlyGate103-AcLEA was transferred to Agrobacterium tumefaciens C58C1 by electroporation in 0.1 cm gap cuvettes. A single colony was used to inoculate LB broth supplemented with ampicillin (100 μg/ml), rifampicin (100 μg/ml), and kanamycin (50 μg/ml). The inoculated broth was cultivated at 30°C overnight. To ensure high expression levels of the recombinant protein, A. tumefaciens cells containing the expression clone were used along the helper strain p19 (Voinnet et al., 2003). A. tumefaciens cells were harvested by centrifugation at 1,400 × g at room temperature and resuspended in an aqueous solution of 10 mM MgCl2. Dilutions were made to adjust a final OD600 of 1.0 in the infiltration solution of both the p19 helper strain and the experimental strain (carry on the pEarlyGate103-AcLEA expression vector). Then acetosyringone (50 μg/ml) was added to the infiltration solution and incubated at room temperature for 3 h. This bacterial solution was used to infiltrate N. benthamiana leaves and the treated plants were incubated for 96 h in regular growth conditions (26°C and 16/8 h light/dark cycle) prior the protein extraction or protoplasts preparation.
Total protein was extracted from infiltrated leaves by 10 min incubation in extraction buffer (70 mM Tris-HCl, pH 8.0, 1 mM MgCl2, 25 mM KCl, 5 mM NaEDTA⋅2H2O, 0.25 mM sucrose, 7.5 mM DTT, 0.1% v/v Triton X-100) followed by centrifugation at 16,000 × g for 10 min at 4°C. The protein extracts were analyzed by Western blot using anti-GFP (Invitrogen) and anti-AcLEA specific antibodies. Protoplasts were released from the leaf tissue by incubation in enzyme solution composed of 0.5 M mannitol, 1% w/v cellulase R10 (KARLAN Research Products Corp., Cottonwood, AZ, USA) and 0.05% w/v macerozyme R10 (KARLAN Research) and leaf tissue was incubated in this solution for 3 h at constant agitation (1,400 × g). Confocal microscopy images were obtained with an Olympus FV1000 microscope (Olympus, Center Valley, PA, USA) using excitation lasers of 633 nm for chlorophyll and 514 nm for GFP.
Detection of AcLEA in Seeds, Leaves, Stems, and Roots from Different Amaranth Species
Proteins from seeds, leaves, stems, and roots were extracted from wild (A. hybridus and A. powellii) and domesticated (A. cruentus and A. hypochondriacus) amaranth species.
Seeds were milled under liquid nitrogen in order to obtain a fine powder and proteins were extracted according to their solubility properties. Aqueous soluble proteins were extracted with buffer containing 10% glycerol, 0.1 M Tris-HCl, pH 8.0 in a relation 1:20 (flour/buffer). Suspension was mixed with vortex for 15 min at 4°C and centrifuged at 17,000 × g at 4°C, supernatant was recovered and named as hydrophilic fraction. Resulting pellet was resuspended in 7 M urea, 2 M thiourea, 2% CHAPS (w/v), 2% Triton X-100, 0.05 M DTT and mixed as indicated above. The solubilized proteins (hydrophobic fraction) were recovered by centrifugation for 15 min at 17,000 × g at 4°C.
Proteins from leaves, stems, and roots were extracted from plants growing under normal and salt stress conditions. Seeds were germinated and seedlings were transferred to plastic pots with soil (Peat Moss Tourbe, Premier Horticulture, Rivière-du-Loup, QC, Canada). Amaranth seedlings were divided in two groups; the control and the salt-stressed groups, which were watered with water and water containing 150 mM NaCl (EC 16.9–17.2 ds/m), respectively. Samples from control and salt-stressed plants were collected next day after salt-stress imposition. Tissues were collected from three biological replicates containing three plants for each replicate. Samples were collected and immediately frozen in liquid nitrogen and milled to a fine powder as reported before (Huerta-Ocampo et al., 2014). The powder was suspended in extraction buffer (1:10 w/v) containing 7 M urea, 2 M thiourea, 2% Triton X-100, and 0.1 M 2-mercaptoethanol. The mixture was sonicated (GE-505, Ultrasonic Processor, Sonics & Materials, Inc., Newtown, CT, USA) for 15 min at 4°C and centrifuged as above.
Proteins (10 μg) were separated in a 12% SDS-PAGE and resolved at 75/150 V for 90 min and then transferred to a PVDF membrane using a Trans-Blot SD semi-dry transfer cell (Bio-Rad, Hercules, CA, USA) for 45 min at 15 V in transfer buffer (25 mM Tris, 192 mM glycine). Membranes were blocked for 2 h with 5% defatted milk in TBS containing 0.1% Tween-20 (TBST), washed three times for 10 min with TBST and incubated with anti-AcLEA IgG rabbit polyclonal antibody for 2 h at 1:1,000 dilution in TBST. Membranes were washed three times for 10 min each with TBST, incubated with anti-rabbit IgG-alkaline phosphatase antibody (Sigma–Aldrich) for 90 min at 1:10,000 dilution in TBST. After membranes were washed three times for 10 min with TBST. Western blots were revealed with alkaline phosphatase buffer (0.1 M Tris, pH 9.5, 0.1 M NaCl, 5 mM MgCl2) and 0.5 mM BCIP, 0.4 mM NBT for 10–20 min at 37°C.
Results and Discussion
AcLEA Cloning and Recombinant Protein Expression in E. coli System
Bioinformatics analyses, using LC-MS/MS information (Maldonado-Cervantes et al., 2014) and the A. hypochondriacus transcriptome (Delano-Frier et al., 2011), allowed us to design specific primers for cloning the full-length AcLEA cDNA. Amplified AcLEA fragment was ligated into pET28mod vector (Supplementary Figures S1A,B).
AcLEA cDNA contains an ORF of 516 bp that codifies for a 172 amino acids protein with a molecular mass calculated of 18.34 kDa and a theoretical pI of 8.58, values that corresponded to experimental data previously reported (Maldonado-Cervantes et al., 2014). The sequence (Supplementary Figure S2A) was deposited in the GenBank with access code KX852451. In order to identify AcLEA similar proteins and consensus sequences, a search was performed using protein BLAST algorithm and multiple alignment was carried out with the sequences of the highest similarity matches (Figure 1A). A search of related sequences in the LEAPdb database (Hunault and Jaspard, 2010) confirmed that all these sequences are grouped in the LEA_4 Pfam (PF02987). According to the classification proposed by Battaglia et al. (2008), in this family are included LEA proteins from Group 3, such as the cotton protein D-7 (Dure, 1993). Group 3 LEA proteins are characterized by a repetitive motif of 11 amino acids TAQAAKDKTSE (motif 3) in the middle of the sequence that is preceded or followed by ATEAAKQKASE (motif 5); in the N-terminal region is usually conserved the SYKAGETKGRKT (motif 4); meanwhile GGVLQQTGEQV (motif 1) and AADAVKHTLGM (motif 2) are frequently observed in the C-terminal. In many proteins motifs 3 and 5 are present more than once. Motifs 1 to 5 were detected in AcLEA wherein the motif arrangement is M4-M5-M3-M1-M2 with only one complete motif of each type (Figure 1A). On the other hand, the motifs arrangement for LEA group 6 is in the order M3-M1-M2-M4 (Rivera-Najera et al., 2014).

FIGURE 1. (A) MUSCLE multiple sequence alignment of AcLEA isolated from immature seeds of Amaranthus cruentus. The red boxes show the category and position of the conserved motifs. (B) Phylogenetic tree constructed using the neighbor-joining method based on the multiple sequences alignment. Accessions numbers of published sequences in the GenBank are as follows: Amaranthus cruentus (AcLEA, KX852451), Beta vulgaris LEA_Dc3 (XP_010691209.1), Vitis amurensis LEA5 (ADY17817.1), Brassica napus LEA76, Vitis vinifera (XP_002285360.1), Arabidopsis thaliana LEA7 (AT1G00010), Arabidopsis thaliana LEA 1X08_A (AT1G01470), Arabidopsis lyrata subsp. lyrata (ARALDYDRAFT_47395), Cicer arietinum LEA3 (XP_004506901.1), Medicago truncatula LEA3 (XP_013454682.1), Lupinus angustifolius LEA3 (XP_019454903.1), Arachis hypogaea LEA3 (ADQ91835.1), Oryza sativa Indica LEA3 (CAA92106.1), Oryza sativa Japonica LEA3 (ABS44867.1), Triticum aestivum (AHZ35571.1), PVLEA4-25 and PvLEA18 from Phaseolus vulgaris (AAC49862.1 and AAC49859.1, respectively), Zea mays (NP_001150813.1), Catharanthus roseus (AAY84145.1), Solanum tuberosum LEA 2-like (XP_006364193.1), Solanum lycopersicum (NP_001238798.1), Capsicum annuum DC3 (XP_016562822.1), Nicotiana sylvestris Dc3-like (XP_009770536.1), Nicotiana tabacum Dc3-like (XP_016459037.1).
With these sequences was constructed the phylogenetic tree in which was also included commercial crops such as: Zea mays (NP_001150813.1), Phaseolus vulgaris (PvLEA_18 and PVLEA4-25), Triticum aestivum (AHZ35571.1), Oryza sativa (LEA_3 ABS44867.1 and CAA92106.1), and other crops such a Vitis amurensis (ADY17817.1), Vitis vinifera (XP_002285360.1), Nicotiana sylvestris (XP_009770536.1), Nicotiana tabacum (XP_016459037.1), Capsicum annuum (XP_016562822.1), Catharanthus roseus (AAY84145.1), Camelina sativa (010487398.1), Solanum tuberosum (XP_006364193.1), Arabidopsis lyrata subsp. lyrata (ARALYDRAFT_474395), among others. The phylogenetic tree shows that LEA denominated Dc3 from B. vulgaris (sugar beet, XP_010691209.1) shared the highest homology with AcLEA (Figure 1B), while the LEA-14 from A. thaliana (1X08, At1g01470) which structure has been reported (Singh et al., 2005) showed low similarity with AcLEA as well as for LEA proteins from the commercial cereals and legumes.
AcLEA shares a similar amino acid composition as other LEA proteins, being rich in alanine (19.2%), lysine (14.0%), glutamic acid (9.9%), glutamine (9.3%), threonine (9.3%), and glycine (8.1%) (Battaglia et al., 2008; Denekamp et al., 2010). The total number of negatively charged residues (Asp and Glu) is 27, meanwhile positive charged residues (Arg and Lys) is 29. Another characteristic of AcLEA is the lack of Trp and Cys residues. AcLEA has an aliphatic index of 29.36 with a grand average of hydropathicity (GRAVY) computed of -1.23, indicating a higher abundance of hydrophilic amino acids. Based on the AcLEA amino acid sequence, the hydropathic profile was calculated using the Kyte and Doolittle (1982) values, results showing that the hydrophilic character of this protein is clearly exhibited (Supplementary Figure S2B), as well AcLEA was predicted as disordered structure (Supplementary Figure S2C). The term hydrophilins was coined to the group of proteins with an average hydrophilicity index >1 and at least 6% Gly. Since hydrophilicity index is 1.23 and the Gly content is 8.1%, AcLEA protein fits in the definition of hydrophilins (Garay-Arroyo et al., 2000).
Protein Expression and Purification
Two distinctive bands putatively corresponding to the recombinant AcLEA were detected in SDS-PAGE, one of them was located at 21.4 kDa, correlating with the molecular weight expected for recombinant protein linked to His-tag, the second band was located at 16.0 kDa (Figure 2). The identities of these two bands were successfully identified by LC-MS/MS and bioinformatics analysis using A. hypochondriacus database (Supplementary Figure S3). Sequences of the matched peptides as well MASCOT scores are shown in Table 1. Data confirm that both the 21.4 and 16.0 kDa bands corresponded to AcLEA. Nevertheless peptides in the N-terminal region were not detected in the 16.0 kDa product, indicating that this short protein is a truncated fragment lacking the N-terminal region.
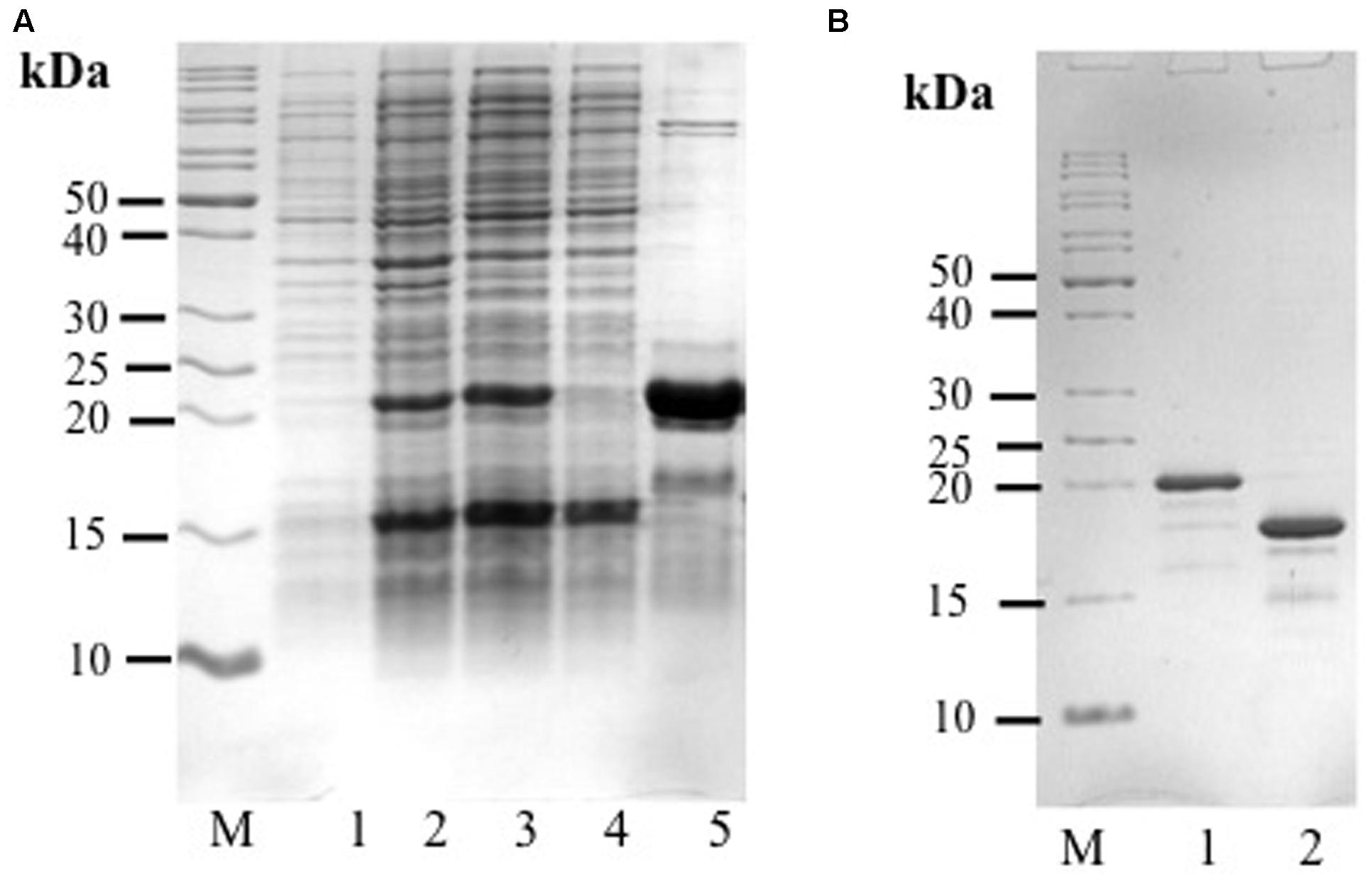
FIGURE 2. (A) Analysis of His-rAcLEA expression and purification. Lane M = molecular weight marker, Lane 1 = total proteins from non-induced BL21 cells, Lane 2 = total proteins from BL21 cells after induction with IPTG, Lane 3 = soluble proteins from Lane 2, Lane 4 = fraction not retained from Ni2+-column, Lane 5 = fraction retained in Ni2+-column and eluted with 300 mM imidazole. (B) His-tag cleavage with PreScission Protease (PSP), Lane M = molecular weight marker, Lane 1 = His-Tag rAcLEA, Lane 2 = rAcLEA.
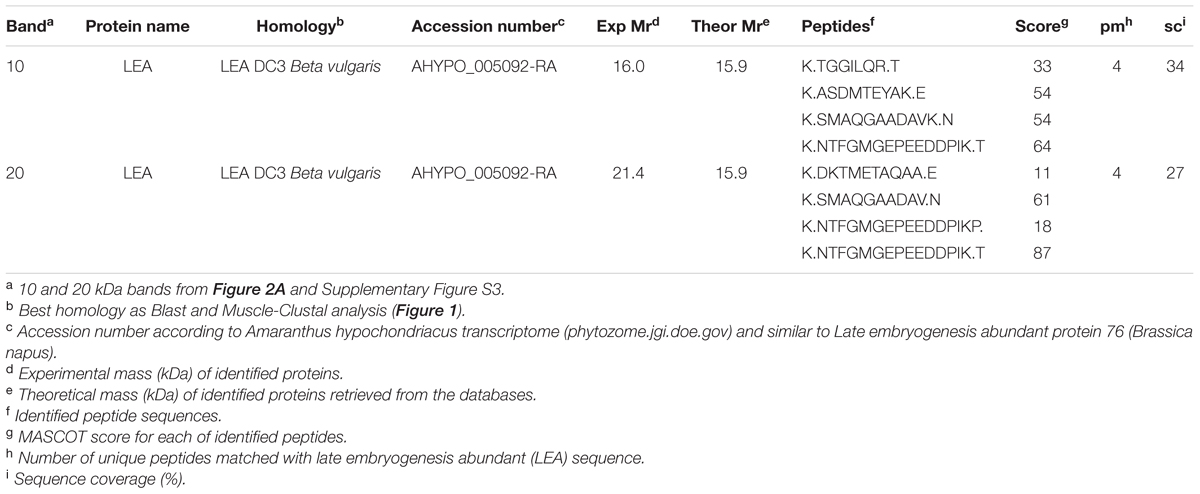
TABLE 1. Recombinant AcLEA protein identities by LC-MS/MS.
The 21.4 kDa His-rAcLEA was retained on the Ni2+ column and was eluted continuously with successive low concentrations of imidazole (50 mM) washes, but high imidazole concentration (300 mM) was required to completely recover the rAcLEA (Figure 2A and Supplementary Figure S4). The 15.4 kDa rAcLEA was not retained by Ni2+ column, confirming that this protein is a truncated fragment lacking of N-terminal His-Tag, which was confirmed by MS/MS analysis (Table 1 and Supplementary Figure S3). After exchange buffer by dialysis, the His-Tag was removed by PSP protease cleavage and rAcLEA purification was carried out again using Ni2+-NTA resin (Figure 2B). Retention of cleaved rAcLEA in this stationary phase can be explained because after the proteolysis cleavage residues added to the N-terminal include Gly-Pro-His, and since AcLEA possesses a His in position 4 (Met-Ala-Ser-His), this combination of two histidine residues in relative positions 1–4 seems to be responsible for the rAcLEA binding to Ni2+-NTA resin.
For spectroscopic analysis it is desirable to have a protein purity greater than 98% (Acton et al., 2005). To ensure this experimental condition it was necessary to use a final chromatographic purification step based on molecular exclusion. Both rAcLEA proteins purified by native and denaturing conditions were eluted in a Sephacryl S-100 column with 10 mM sodium phosphates buffer at pH 7.0 as mobile phase; no changes were detected in retention time between them. Typical chromatographic profile shows only one well-defined peak and rAcLEA showed higher purity (Supplementary Figures S5A,B).
Nuclear Magnetic Resonance Spectroscopy
rAcLEA obtained under native conditions was used to evaluate the structural conformation of the recombinant protein by proton nuclear magnetic resonance. Uni-dimensional 1H-NMR spectrum provides general overviews of protein structure because chemical shifts values are strongly related with the presence of different elements of secondary structure (Wishart et al., 1991; Mielke and Krishnan, 2009). Particularly, HN amide protons are widespread from 6 to 11 ppm in proteins with a well-defined tri-dimensional folding with a high content of α-helix and β-strand. In contrast, HN resonances of unfolded proteins with a random coil conformation collapse in a narrow region around 7–8 ppm (Singh et al., 2005). Figure 3 shows the 1H-NMR spectrum of native rAcLEA, as can be observed amide and aromatic protons are distributed between 6.8 and 8.6 ppm, suggesting a random conformation. Moreover, Hα resonances around 4.1 ppm have also a compact distribution, which is consistent with random coil as well the absence of splitting due to coupling in aliphatic signals in the 0.8–2.0 ppm range. This spectroscopic patterns indicate that methyl and methylene groups present in aliphatic amino acid lateral chains have free rotation without limitations due to steric impediment, suggesting that rAcLEA in the experimental conditions tested lacks secondary and tertiary structure. In fact, rAcLEA possesses the typical NMR profile for IDP previously observed in a LEA protein of T. aestivum (Sasaki et al., 2014) and a dehydrin of A. thaliana (Agoston et al., 2011).
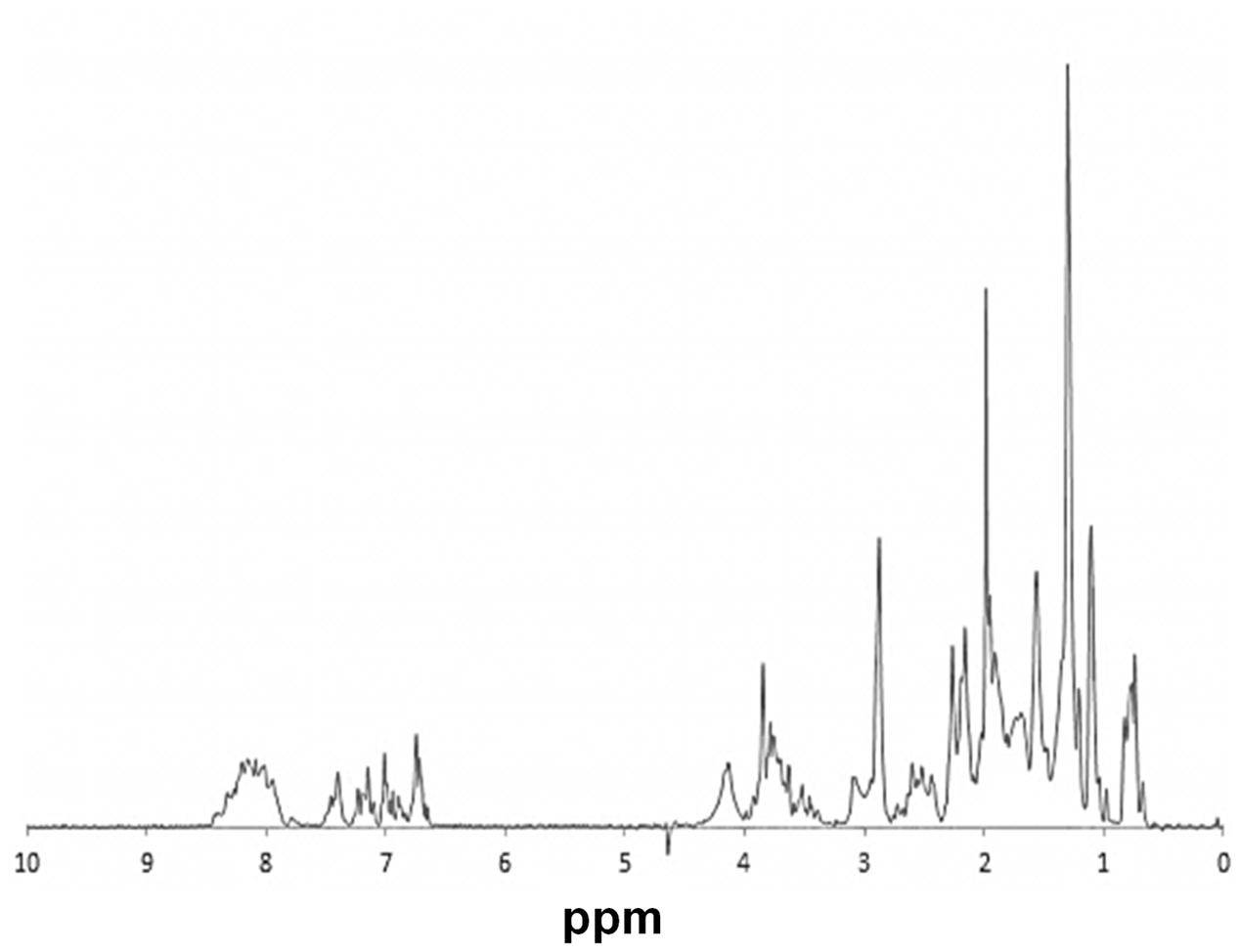
FIGURE 3. NMR. 1H-NMR spectrum of HisAcLEA purified from native conditions. Narrow signal distribution in amide region, between 6.5 and 8.5 ppm, strongly suggest the lack of a well-defined tridimentional structure distintive of intrinsically disordered proteins.
Circular Dichroism Spectroscopy
The amino acid composition of AcLEA is rich in α-helix promoters such as Ala (19.0%), Met (5.2%), Glu (9.8%), Gln (9.2%), Thr (9.2%), and Lys (13.8%), nevertheless the Gly content is high (8.1%) this amino acid does not have a high propensity for secondary structure formation (Serrano et al., 1992; Creighton, 1993). As observed in other LEA proteins, secondary structure prediction indicates the formation of vast segments of helical structures reaching up to 80% α-helix content. Interestingly, NMR data (Figure 3) showed that in the experimental tested conditions rAcLEA has the spectral profile of an IDP. Therefore, in order to further explore the conformational properties of rAcLEA, CD spectra were recorded in the far UV region. rAcLEA was dissolved in 10 mM phosphate buffer pH 8.0 at different NaCl or sorbitol concentrations (Furuki et al., 2011; Wu et al., 2014; Warner et al., 2016). As shown in Figure 4A, the AcLEA spectra were not modified by NaCl nor sorbitol presence. All these CD spectra show a negative signal near 200 nm and weak bands in the 210–220 region, suggesting a low secondary structure content. In agreement, the deconvolution of the spectra using the CDNN program (Bohm et al., 1992) indicates a limited content of secondary structure (Supplementary Table S1). Because it is well known that the temperature-induced conformational changes (Soulages et al., 2002), then the curves as a function of temperature at different wavelengths (210, 222, and 230 nm) were followed. For all samples at all the wavelengths tested, the ellipticity signal barely changed with temperature (Figure 4B). In agreement, the spectra obtained at 20°C before and after the heating cycle as well as that obtained at 75°C were very similar (Supplementary Figure S6). The lack of a temperature-induced transition strongly suggests that if secondary structure segments are formed, those segments are fluctuating and do not participate in the compact core structure.
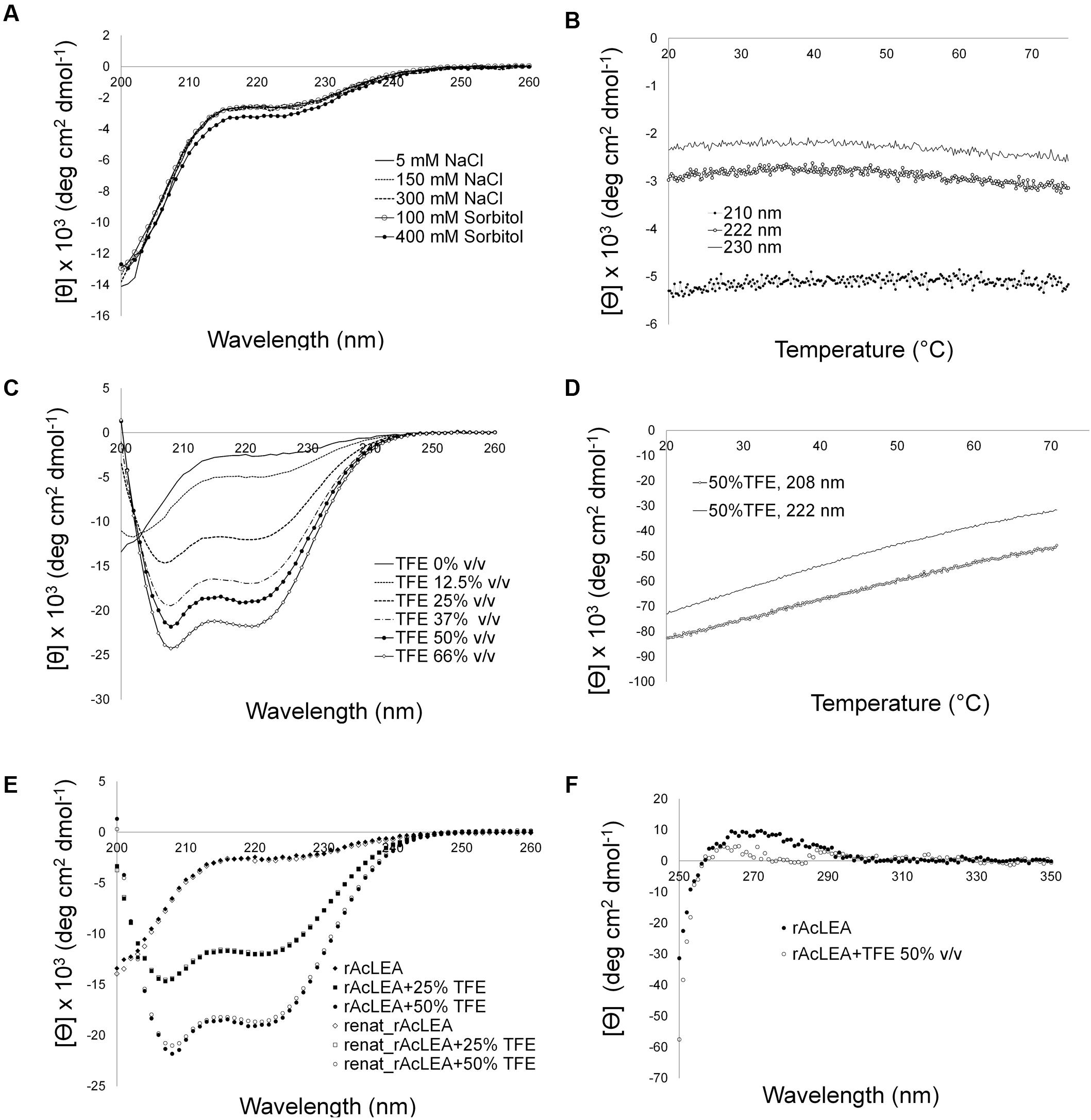
FIGURE 4. Circular dichroism spectroscopy of rAcLEA under different environment conditions. (A) Far circular dichroism (CD) spectra of rAcLEA in presence of NaCl and sorbitol. (B) Recombinant AcLEA (5 mM NaCl, 10 mM phosphates, pH 8, melting monitoring at 210, 222, and 230 nm. (C) CD spectra of rAcLEA at different concentrations of trifluoroethanol (TFE), an additive that induce α-helix folding. (D) Melting denaturing of α-helix rAcLEA folded with 50% v/v TFE. (E) Folding recovery after melting. (F) Near CD spectra of rAcLEA in buffer solution with 50% v/v TFE.
It is well established that TFE can induce α-helix folding in peptides (Buck, 1998; Boswell et al., 2014), as well as in unstructured proteins with a predisposition to form secondary structure such as LEA proteins (Shih et al., 2004; Rivera-Najera et al., 2014). Therefore the effect of TFE was evaluated on the rAcLEA conformation. Far UV CD spectra clearly show the tendency of rAcLEA to adopt helical structure as TFE concentration increases (Figure 4C). At TFE concentrations higher than 25%, the CD spectra of rAcLEA show the distinctive minima at 208 and 222 nm characteristic of α-helix structures (Muller et al., 2008). As TFE concentration increased up to 66% a gain of helical structure up 70.7% and a decreased in all the other types of secondary structure were observed (Figure 5 and Supplementary Table S1), this result being quantitatively confirmed using the CDNN program (Supplementary Table S1). In order to determine if this increase in helical content was accompanied with the formation of a structured core, the effect of temperature on rAcLEA dissolved in 50% TFE was assayed. It was found that the ellipticity signal at 208 and 222 nm was lost in a non-cooperative way (Figure 4D) and changes in CD signal were fully reversible at 25 and 50% TFE (Figure 4E). This strongly suggests that the helical segments induced by the addition of TFE are not arranged in a well-folded tertiary structure. To further explore the formation of tertiary structure, the CD spectra of rAcLEA in the aromatic region were also determined. In the absence of TFE, rAcLEA showed a weak signal in the region corresponding to Tyr and Phe residues, the intensity at 270 nm band being further decreased in the presence of TFE (Figure 4F), thus confirming the absence of TFE-induced tertiary structure formation.
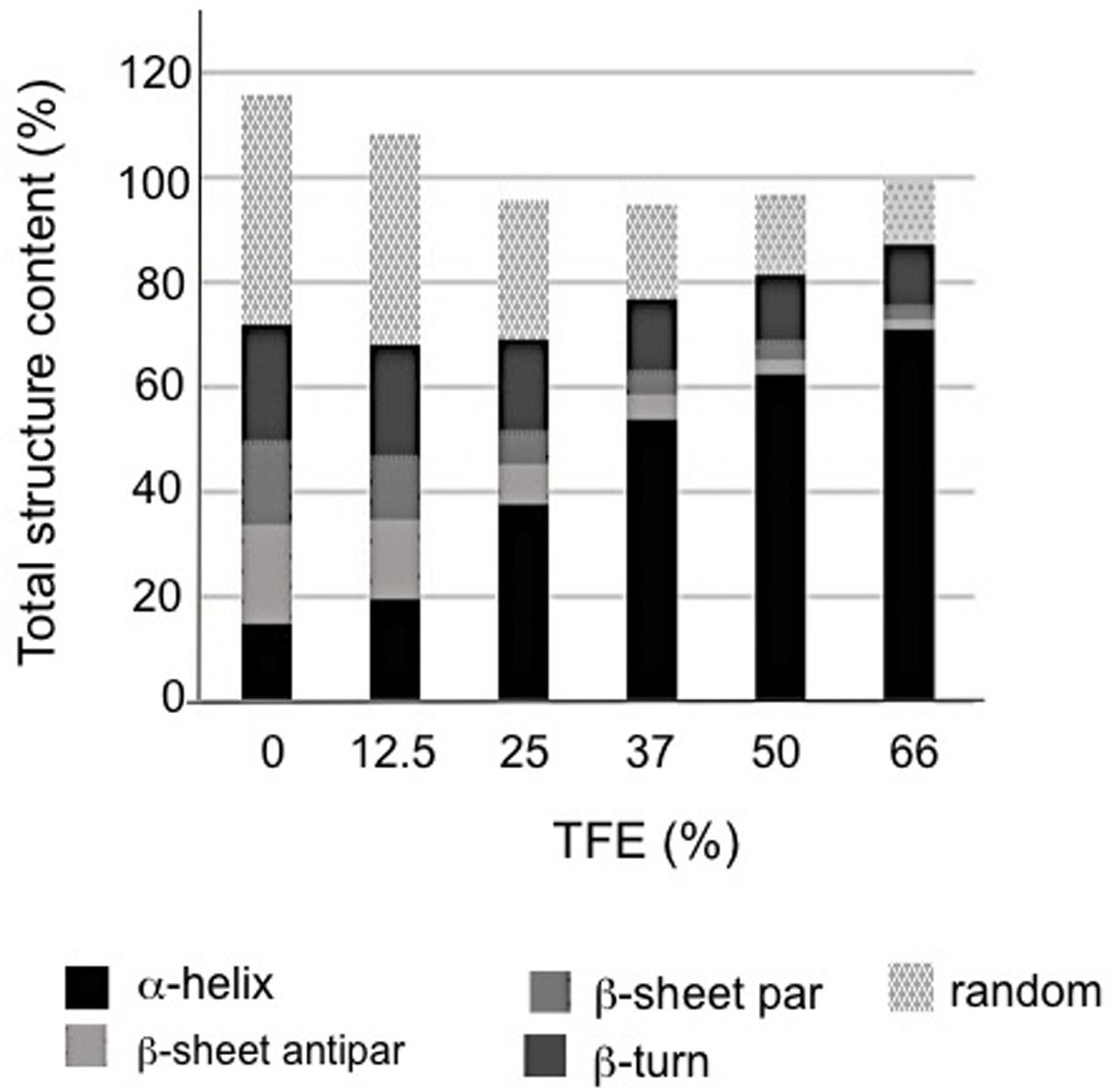
FIGURE 5. Structural composition vs. TFE concentration in rAcLEA solution as calculated from respective CD data using CDNN software.
Biological Properties of AcLEA In vivo Using E. coli as a Model
It has been demonstrated that the expression system of E. coli is a simple, convenient, and effective model to determine the function of recombinant proteins (Liu and Zheng, 2005). So we used the transformed E. coli DE3 cells to evaluate their tolerance to diverse types of abiotic stress conditions (desiccation, NaCl, H2O2, sorbitol, and PEG-4000).
Figure 6A shows the growth kinetics of control E. coli cells transformed with empty plasmid (control) and pET28mod-AcLEA plasmid. It has been reported that expression of LEA (group 1) genes from plants has no effect on the growth kinetics of transformed E. coli or yeast cells (Lan et al., 2005; Campos et al., 2006; Dang et al., 2014). These results are in agreement with our results; however, Warner et al. (2016) reported that induction of AfrLEA-1 (Artemia franciscana LEA group 1) was associated with inhibition of Top10F′ E. coli on account of basic pI of AfrLEA-1. Curiously AcLEA has also a basic pI but we did not observe such cell growth inhibition.
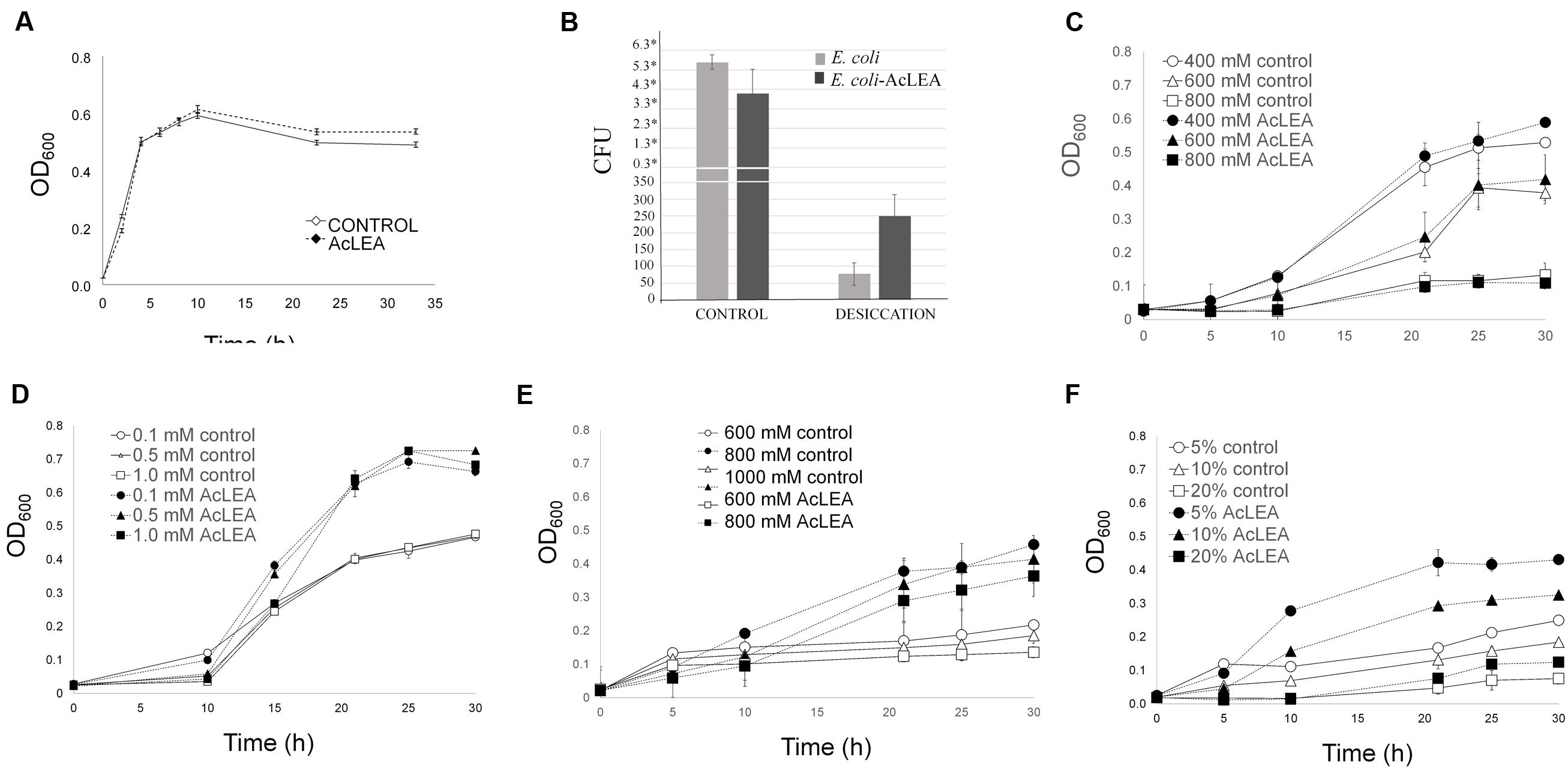
FIGURE 6. Escherichia coli growth in different stress conditons. Cells were grown overnight in LB medium (with antibiotic) and their concentration was determined. An equal number of cells were added to flask with LB (control) and LB containing: NaCl, H2O2, sorbitol or PEG as indicated. (A) Growth kinetics of E. coli transformed with pET28mod (control) and pET28mod-AcLEA. Cells were grown until optical density at 600 nm reached 0.6 then 0.1 mM IPTG was and incubated for 2 h at 28°C. (B) Cell viability related to colony former units (CFU) before (control) and after desiccation. ∗ ×106 cells. (C) Effect of salinity stress induced by NaCl. (D) Oxidant stress caused by additon of H2O2. (E,F) Effect of dehydration stress and osmotic potential stress simulated by the additon of sorbitol and PEG-4000, respectively. Each curve/column represents an average of three biological experiments with three replicates each. Bars in each figure represent the standard deviation.
It has been suggested that hydrophilic and heat-stable proteins may modify the structure of other proteins and bind water directly to attenuate the damage caused by desiccation (Houde et al., 1992). Figure 6B shows a clear difference in the number of E. coli viable cells before and after desiccation stress. Before drying process very similar CFU (expressed in ×106 units) were obtained for control and AcLEA expressing cells, but after desiccation, although only a very small fraction of cells survived, the number of CFU expressing AcLEA were three times higher than in control cells. This result suggests that AcLEA expression E. coli improved its survival capacity after desiccation. On the other hand, it is well known that the E. coli growth rate is strongly influenced by the salt content present in the growth medium (Gowrishankar, 1985). Lan et al. (2005) and Reddy et al. (2012) showed that overexpression of LEA group 1 from plants in E. coli provides an increased tolerance to the harmful effects of high salinity environments. Liu and Zheng (2005) indicated that the expression of PM2, a LEA group 3 from soybean, enhances salt tolerance of E. coli cells and that the 22-mer repeat region is an important functional region in this protein. As shown in Figure 6C, the E. coli growth was inhibited by addition of NaCl and contrarily to other reports, the expression of AcLEA did not change this behavior. Because AcLEA has been classified as LEA Group 3 it was expected that it would participated in the protection of cells against salt stress, however, the differences in the amino acid sequences detected in AcLEA (Figure 1A) could be responsible for this observed difference.
Low ROS concentrations can act as messengers to regulate biological process, while high ROS concentrations can have very harmful effects and dehydration will disrupt the metabolism of seeds leading to high ROS production (Bailly et al., 2008). In Figure 6D is shown that even at high H2O2 concentration, AcLEA conferred a significant tolerance to E. coli cells. On the other hand, Warner et al. (2016) reported that in general E. coli strains tolerate low sorbitol concentrations. Our results showed that AcLEA was able to overcome the negative sorbitol effect on E. coli growth even at 1 M concentration (Figure 6E). E. coli growth was also tested in the presence of PEG as a compound that decreases the osmotic potential of the cells. As shown in Figure 6F, the accumulation of AcLEA improved the growth cell supporting an osmoprotection function.
In situ Localization of AcLEA
To decipher the subcellular localization of AcLEA protein, the corresponding coding sequence was fused with green fluorescent protein (AcLEA-GFP) in vectors designed for transient transgene expression in N. benthamiana leaf protoplasts. Confocal microscopy images (Figure 7A) of protoplasts from leaves infiltrated with the expression vector pEarlyGate103-AcLEA clearly show that AcLEA protein exhibits a cytosolic localization in these conditions. The accumulation of AcLEA and GFP proteins in infiltrated leaves was confirmed by immunodetection analysis (Figure 7B). It is noted that cytosolic LEA proteins could be involved in stress protection not only within the cytosol itself but also at the level of membranes delimiting the organelles such as mitochondria, chloroplasts, endoplasmic reticulum, and nucleus (Candat et al., 2014).
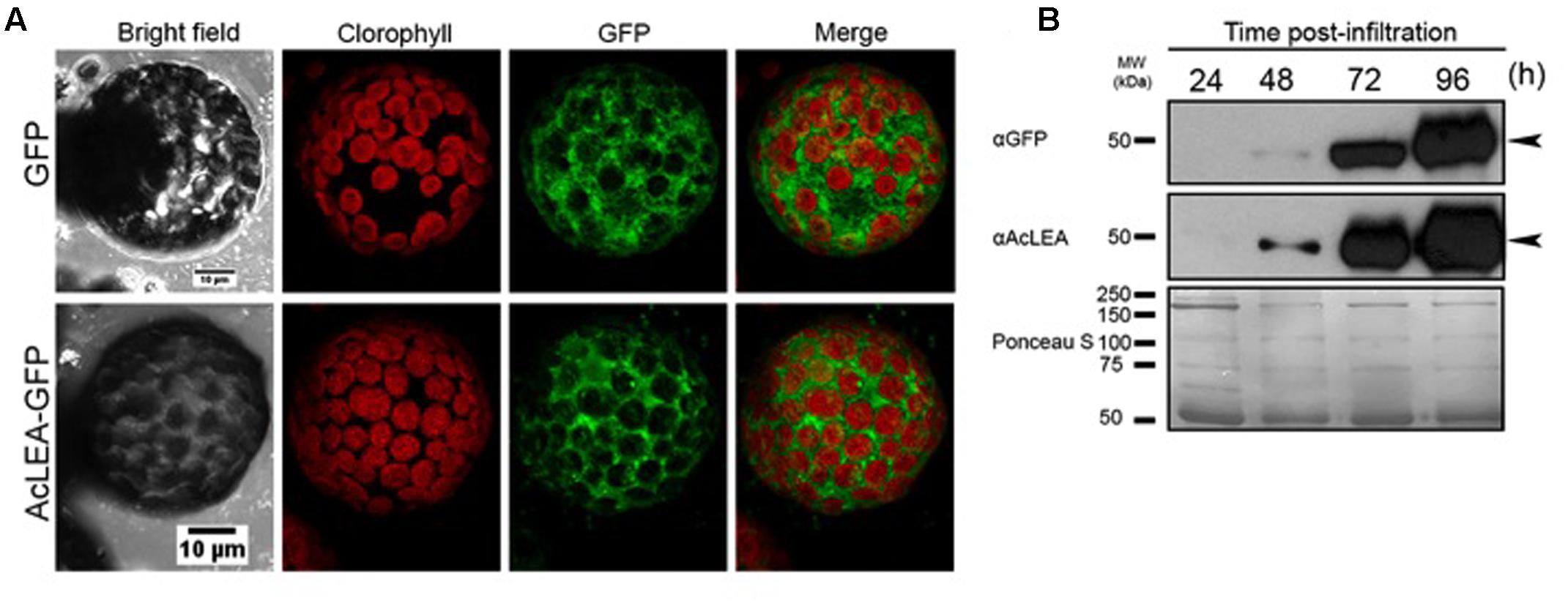
FIGURE 7. AcLEA subcellular localization using the Nicotiana benthamiana system. (A) Confocal microscopy images of protoplasts from N. benthamiana leaves infiltrated with the expression vector pEarlyGate103 (GFP) and pEarlyGate10-AcLEA-GFP. Images were captured 96 h after infiltration of indicated constructs in bright field and in fluorescence using an excitation laser of 488 and emission filters BA655-755 for chlorophyll and BA505-605 for GFP. A merged image is also shown. (B) Immunoblots of kinetics of expression and accumulation of proteins infiltrated at the indicated times. Below immunodetection autoradiographs, the Ponceau S stained membrane is shown as loading control.
AcLEA Localization in Amaranth Seeds and Plant Tissues
Anti-AcLEA antibodies were sensitive to detect the corresponding polypeptides in extracts from seed proteins from different amaranth wild and domesticated species. Among all species analyzed, no differences in abundance were observed in seeds (Figures 8A,B and Supplementary Figure S7). This could indicate that AcLEA plays an important function most likely during seed drying process. To identify all sequences related with LEA proteins, we carried out a search in phytozome database7. Sixty matches were retrieved but only one of those sequences (AHYPO_005092) was identical to AcLEA (Supplementary Figure S8), which correlates with the Western blot analysis where only one reactive band was observed (Figure 8B).
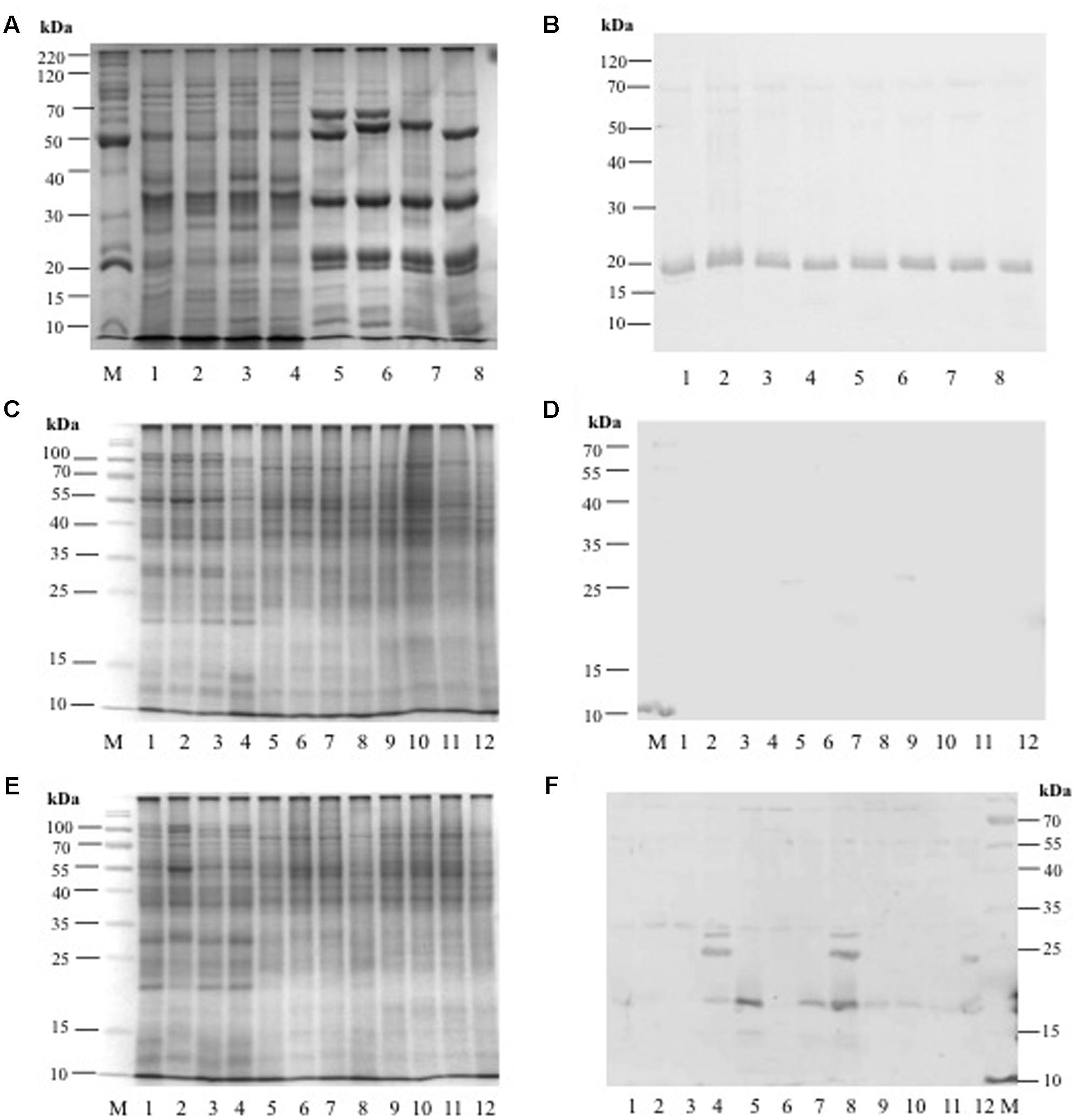
FIGURE 8. (A) SDS-PAGE profile from amaranth seed storage proteins: Lane M = molecular weigh marker, Lanes 1–4 = hydrophilic proteins from: A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively. Lanes 5–8 = hydrophobic proteins from: A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively. (B) Western Blot analysis against anti-AcLEA. (C) SDS-PAGE profile from amaranth leaves, stems, and roots from plants growing under normal conditions: Lane M = molecular weight marker; Lanes 1–4 = leaf proteins from: A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively; Lanes 5–8 = stem proteins from: A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively; Lanes 9–12 = root proteins from: A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively. (D) Western blot analysis against AcLEA. (E) SDS-PAGE profile from amaranth leaves, stems, and roots from plants subjected to salinity stress: Lane M = molecular weight marker; Lanes 1–4 = leaf proteins from of A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively; Lanes 5–8 = stem proteins from of A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively; Lanes 9–12 = root proteins from of A. hybridus, A. powellii, A. cruentus, and A. hypochondriacus, respectively. (F) Western blot analysis against AcLEA.
The abundance of AcLEA was tested also on leaves, stems, and roots of wild and domesticated amaranth species. Under normal conditions of plant growth of watering, AcLEA was not detected (Figures 8C,D). But very interestingly, when plants were subjected to salinity stress, we observed the accumulation of AcLEA (Figures 8E,F). As shown in Figure 8F, AcLEA accumulation was observed in A. hypochondriacus leaves in the expected size (19 kDa, Supplementary Figure S7) but also two more bands around 25 and 30 kDa were observed. In leaves of wild species, the 19 kDa band was barely observed, but in stems a strong band was observed in the wild species A. hybridus and the domesticated A. cruentus and A. hypochondriacus. Meanwhile in roots the 19 kDa band was detected in all species, but at much lower accumulation. These results have shown that AcLEA is conserved in seeds among amaranth species, but that AcLEA plays an important function in response to plant stress and its tissue-specific accumulation was observed.
Conclusion
We present the isolation, cloning, structural and functional characterization of the first LEA from Amaranthus species (AcLEA). The deduced amino acid sequence of this gene showed that AcLEA belongs to the LEA proteins group 3 and structural analysis in solution has shown that it belongs to IDPs lacking of a well-defined secondary or tertiary structure, but has a strong tendency to adopt a helical conformation. Using E. coli as in vivo model to evaluate the AcLEA function it was shown that this protein displayed a protective effect against desiccation, osmotic, and oxidative stresses. In N. benthamiana leaf protoplasts AcLEA was observed as being localized to the cytosol. Moreover, AcLEA was detected in different tissues from wild and domesticated amaranth species suggesting the important function of AcLEA protein as osmoprotectant during seed desiccation. But interestingly, AcLEA was accumulated in leaves and stems in response to salt stress. These results highlighted the importance of AcLEA as an important protein for stress protection in amaranth species.
Author Contributions
AS and APBR conceived and designed the work. AS and EEHD cloned the AcLEA gene in E. coli system. LdL-V and AG-G, carried out N. benthamiana transient transformation. AS and DF-V carried out CD analysis and AS and FdR-P carried out the NMR analysis. AS and AE-M prepared the anti-AcLEA antibodies in rabbits, EB-V conducted Western blot analysis. AS and APBR drafted the manuscript and all authors reviewed and approval the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
AS and EH give thanks to CONACYT-Mexico postdoctoral fellow 232286 and 290754, respectively. LdL-V thanks CONACYT-Mexico by the doctoral fellowship 240088. We also thank to CONACYT-Mexico Grants 56787 (Laboratory for Nanoscience and Nanotechnology Research-LINAN) and 204373 (Infrastructure project, INFRA-2013-01), and 251848 (CB-2015). We thank to Alberto Barrera-Pacheco for his technical assistance in MS/MS analysis. LdL-V and AG-G thank to LNMA for confocal microscopy images. We thank to Fabiola Veana Hernández for her technical assistance. We thank to Project “Problemas Nacionales-Amaranto en la soberanía alimentaria” No. 248415.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00497/full#supplementary-material
Footnotes
- ^http://forge.info.univ-angers.fr/~gh/Leadb
- ^http://web.expasy.org/protscale/
- ^http://web.expasy.org/protparam/
- ^http://forge.info.univ-angers.fr/~gh/Leadb/index.php
- ^http://pfam.xfam.org/
- ^http://www.phytozome.net
- ^https://phytozome.jgi.doe.gov/pz/portal.html
References
Acton, T. B., Gunsalus, K. C., Xiao, R., Ma, L. C., Aramini, J., Baran, M. C., et al. (2005). Robotic cloning and protein production platform of the northeast structural genomics consortium. Methods Enzymol. 394, 210–243. doi: 10.1016/S0076-6879(05)94008-1
Agoston, B. S., Kovacs, D., Tompa, P., and Perczel, A. (2011). Full backbone assignment and dynamics of the intrinsically disordered dehydrin ERD14. Biomol. NMR Assign. 5, 189–193. doi: 10.1007/s12104-011-9297-2
Aguilar-Hernández, H. S., Santos, L., Leon-Galvan, F., Barrera-Pacheco, A., Espitia-Rangel, E., De Leon-Rodriguez, A., et al. (2011). Identification of calcium stress induced genes in amaranth leaves through suppression subtractive hybridization. J. Plant Physiol. 168, 2102–2109. doi: 10.1016/j.jplph.2011.06.006
Ali-Benali, M. A., Alary, R., Joudrier, P., and Gautier, M. F. (2005). Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. Biophys. Acta 1730, 56–65. doi: 10.1016/j.bbaexp.2005.05.011
Avelange-Macherel, M. H., Payet, N., Lalanne, D., Neveu, M., Tolleter, D., Burstin, J., et al. (2015). Variability within a pea core collection of LEAM and HSP22, two mitochondrial seed proteins involved in stress tolerance. Plant Cell Environ. 38, 1299–1311. doi: 10.1111/pce.12480
Bailly, C., El-Maarouf-Bouteau, H., and Corbineau, F. (2008). From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C R Biol. 331, 806–814. doi: 10.1016/j.crvi.2008.07.022
Battaglia, M., and Covarrubias, A. A. (2013). Late embryogenesis abundant (LEA) proteins in legumes. Front. Plant Sci. 4:190. doi: 10.3389/fpls.2013.00190
Battaglia, M., Olvera-Carrillo, Y., Garciarrubio, A., Campos, F., and Covarrubias, A. A. (2008). The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 148, 6–24. doi: 10.1104/pp.108.120725
Becker, R., Wheeler, E. L., Lorenz, K. A., Staffor, E., Grosjean, O. K., Betschart, A. A., et al. (1981). A compositional study of amaranth grain. J. Food Sci. 46, 1175–1180. doi: 10.1111/j.1365-2621.1981.tb03018.x
Bohm, G., Muhr, R., and Jaenicke, R. (1992). Quantitative analysis of protien far UV ciruclar dichroism spectra by neural networks. Protein Eng. 5, 191–195. doi: 10.1093/protein/5.3.191
Boswell, L. C., Menze, M. A., and Hand, S. C. (2014). Group 3 late embryogenesis abundant proteins from embryos of Artemia franciscana: structural properties and protective abilities during desiccation. Physiol. Biochem. Zool. 87, 640–651. doi: 10.1086/676936
Boucher, V., Buitink, J., Lin, X., Boudet, J., Hoekstra, F. A., Hundertmark, M., et al. (2010). MtM25 is an atypical hydrophobic late embryogenesis-abundant protein that dissociates cold and desiccation-aggregated proteins. Plant Cell Environ. 33, 418–430. doi: 10.1111/j.1365-3040.2009.02093.x
Brenner, D. M., Baltensperger, D. D., Kulakow, P. A., Lehmann, J. W., Myers, R. L., Slabbert, M. M., et al. (2000). “Genetic resources and breeding of amaranthus,” in Plant Breeding Reviews, Vol. 19, ed. J. Janick (Oxford: John Wiley & Sons, Inc).
Browne, J., Tunnacliffe, A., and Burnell, A. (2002). Anhydrobiosis: plant desiccation gene found in a nematode. Nature 416:38. doi: 10.1038/416038a
Buck, M. (1998). Trifluoroethanol and colleagues: cosolvents come of age. Recent studies with peptides and proteins. Q. Rev. Biophys. 31, 297–355. doi: 10.1017/S003358359800345X
Campos, F., Cuevas-Velazquez, C., Fares, M. A., Reyes, J. L., and Covarrubias, A. A. (2013). Group 1 LEA proteins, an ancestral plant protein group, are also present in other eukaryotes, and in the archeae and bacteria domains. Mol. Genet. Genomics 288, 503–517. doi: 10.1007/s00438-013-0768-2
Campos, F., Zamudio, F., and Covarrubias, A. A. (2006). Two different late embryogenesis abundant proteins from Arabidopsis thaliana contain specific domains that inhibit Escherichia coli growth. Biochem. Biophys. Res. Commun. 342, 406–413. doi: 10.1016/j.bbrc.2006.01.151
Candat, A., Paszkiewicz, G., Neveu, M., Gautier, R., Logan, D. C., Avelange-Macherel, M.-H., et al. (2014). The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 26, 3148–3166. doi: 10.1105/tpc.114.127316
Cao, J., and Li, X. (2015). Identification and phylogenetic analysis of late embryogenesis abundant proteins family in tomato (Solanum lycopersicum). Planta 241, 757–772. doi: 10.1007/s00425-014-2215-y
Chakrabortee, S., Meersman, F., Schierle, G. S. K., Bertoncini, C. W., McGee, B., Kaminski, C. F., et al. (2010). Catalytic and chaperone-like functions in an intrinsically disordered protein associated with desiccation tolerance. Proc. Natl. Acad. Sci. U.S.A. 107, 16084–16089. doi: 10.1073/pnas.1006276107
Clouse, J. W., Adhikary, D., Page, J. T., Ramaraj, T., Deholos, M. K., Udall, J. A., et al. (2016). The amaranth genome: genome, transcriptome, and physical map assembly. Plant Genome 9, 1–14. doi: 10.3835/plantgenome2015.07.0062
Creighton, T. (1993). Proteins. Structure and Molecular Properties, Second Edn. New York, NY: W. H. Freeman.
Dalal, M., Tayal, D., Chinnusamy, V., and Bansal, K. C. (2009). Abiotic stress and ABA-inducible Group 4 LEA from Brassica napus plays a key role in salt and drought tolerance. J. Biotechnol. 139, 137–145. doi: 10.1016/j.jbiotec.2008.09.014
Dang, N. X., Popova, A. V., Hundertmark, M., and Hincha, D. K. (2014). Functional characterization of selected LEA proteins from Arabidopsis thaliana in yeast and in vitro. Planta 240, 325–336. doi: 10.1007/s00425-014-2089-z
Delano-Frier, J. P., Aviles-Arnaut, H., Casarrubias-Castillo, K., Casique-Arroyo, G., Castrillon-Arbelaez, P. A., Herrera-Estrella, L., et al. (2011). Transcriptomic analysis of grain amaranth (Amaranthus hypochondriacus) using 454 pyrosequencing: comparison with A. tuberculatus, expression profiling in stems and in response to biotic and abiotic stress. BMC Genomics 12:363. doi: 10.1186/1471-2164-12-363
Denekamp, N. Y., Reinhardt, R., Kube, M., and Lubzens, E. (2010). Late embryogenesis abundant (LEA) proteins in nondesiccated, encysted, and diapausing embryos of rotifers. Biol. Reprod. 82, 714–724. doi: 10.1095/biolreprod.109.081091
Dure, L. III (1989). Common amino acid sequence domains among the LEA proteins of higher plants. Plant Mol. Biol. 12, 475–486. doi: 10.1007/BF00036962
Dure, L. (1993). A repeating 11-mer amino acid motif and plant desiccation. Plant J. 3, 363–369. doi: 10.1046/j.1365-313X.1993.t01-19-00999.x
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Finn, R. D., Coggill, P., Eberhardt, R. Y., Eddy, S. R., Mistry, J., Mitchell, A. L., et al. (2015). The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 44, D279–D285. doi: 10.1093/nar/gkv1344
Furuki, T., Shimizu, T., Kikawada, T., Okuda, T., and Sakurai, M. (2011). Salt effects on the structural and thermodynamic properties of a group 3 LEA protein model peptide. Biochemistry 50, 7093–7103. doi: 10.1021/bi200719s
Gai, Y. P., Ji, X. L., Lu, W., Han, X. J., Yang, G. D., and Zheng, C. C. (2011). A novel late embryogenesis abundant like protein associated with chilling stress in Nicotiana tabacum cv. bright yellow-2 cell suspension culture. Mol. Cell. Proteom. 10, M111.010363. doi: 10.1074/mcp.M111.010363
Garay-Arroyo, A., Colmenero-Flores, J. M., Garciarrubio, A., and Covarrubias, A. A. (2000). Highly hydrophilic proteins in prokaryotes and eukaryotes are common during conditions of water deficit. J. Biol. Chem. 275, 5668–5674. doi: 10.1074/jbc.275.8.5668
Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M. R., Appel, R. D., et al. (2005). “Protein identification and analysis tools on the ExPASy server,” in The Proteomics Protocols Handbook, eds J. M. Walker and N. J. Totowa (New York City, NY: Humana Press Inc), 571–607. doi: 10.1385/1-59259-890-0:571
Gowrishankar, J. (1985). Identification of osmoresponsive genes in Escherichia coli: evidence for participation of potassium and proline transport systems in osmoregulation. J. Bacteriol. 164, 434–445.
Goyal, K., Tisi, L., Basran, A., Browne, J., Burnell, A., Zurdo, J., et al. (2003). Transition from natively unfolded to folded state induced by desiccation in an anhydrobiotic nematode protein. J. Biol. Chem. 278, 12977–12984. doi: 10.1074/jbc.M212007200
Hand, S. C., Menze, M. A., Toner, M., Boswell, L., and Moore, D. (2011). LEA proteins during water stress: not just for plants anymore. Annu. Rev. Physiol. 73, 115–134. doi: 10.1146/annurev-physiol-012110-142203
Hatanaka, R., Furuki, T., Zhimizu, T., Takezawa, D., Kikawada, T., Sakurai, M., et al. (2014). Biochemical and structural characterization of an endoplasmic reticulum-localized late embryogenesis abundant (LEA) protein from the liverwort Marchantia polymorpha. Biochem. Biophys. Res. Commun. 454, 588–593. doi: 10.1016/j.bbrc.2014.10.130
He, S., Tan, L., Hu, Z., Chen, G., Wang, G., and Hu, T. (2012). Molecular characterization and functional analysis by heterologous expression in E. coli under diverse abiotic stresses for OsLEA5, the atypical hydrophobic LEA protein from Oryza sativa L. Mol. Genet. Genomics 287, 39–54. doi: 10.1007/s00438-011-0660-x
Hincha, D. K., and Thalhammer, A. (2012). LEA proteins: IDPs with versatile functions in cellular dehydration tolerance. Biochem. Soc. Trans. 40, 1000–1003. doi: 10.1042/BST20120109
Houde, M., Danyluk, J., Laliberte, J. F., Rassart, E., Dhindsa, R. S., and Sarhan, F. (1992). Cloning, characterization, and expression of a cDNA encoding a 50-kilodalton protein specifically induced by cold acclimation in wheat. Plant Physiol. 99, 1381–1387. doi: 10.1104/pp.99.4.1381
Hu, T., Zhou, N., Fu, M., Qin, J., and Huang, X. (2016). Characterization of OsLEA1a and its inhibitory effect on the resistance of E. coli to diverse abiotic stresses. Int. J. Biol. Macromol. 91, 1010–1017. doi: 10.1016/j.ijbiomac.2016.06.056
Huerta-Ocampo, J. A., Barrera-Pacheco, A., Mendoza-Hernández, C. S., Espitia-Rangel, E., Mock, H. P., and Barba de la Rosa, A. P. (2014). Salt stress-induced alterations in the root proteome of Amaranthus cruentus L. J. Proteome Res. 13, 3607–3627. doi: 10.1021/pr500153m
Huerta-Ocampo, J. A., Leon-Galvan, M. F., Ortega-Cruz, L. B., Barrera-Pacheco, A., De Leon-Rodriguez, A., Mendoza-Hernandez, G., et al. (2011). Water stress induces up-regulation of DOF1 and MIF1 transcription factors and down-regulation of proteins involved in secondary metabolism in amaranth roots (Amaranthus hypochondriacus L.). Plant Biol. 13, 472–482. doi: 10.1111/j.1438-8677.2010.00391.x
Hunault, G., and Jaspard, E. (2010). LEAPdb: a database for the late embryogenesis abundant proteins. BMC Genomics 11:221. doi: 10.1186/1471-2164-11-221
Hundertmark, M., and Hincha, D. K. (2008). LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 9:118. doi: 10.1186/1471-2164-9-118
Imai, R., Chang, L., Ohta, A., Bray, E. A., and Takagi, M. (1996). A lea-class gene of tomato confers salt and freezing tolerance when expressed in Saccharomyces cerevisiae. Gene 170, 243–248. doi: 10.1016/0378-1119(95)00868-3
Ingram, J., and Bartels, D. (1996). The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 377–403. doi: 10.1146/annurev.arplant.47.1.377
Jaspard, E., Macherel, D., and Hunault, G. (2012). Computational and statistical analyses of amino acid usage and physic-chemical properties of the twelve late embryogenesis abundant protein classes. PLoS ONE 7:e36868. doi: 10.1371/journal.pone.0036968
Karimi, M., Depicker, A., and Hilson, P. (2007). Recombinational cloning with plant gateway vectors. Plant Physiol. 145, 1144–1154. doi: 10.1104/pp.107.106989
Kyte, J., and Doolittle, R. F. (1982). A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 157, 105–132. doi: 10.1016/0022-2836(82)90515-0
Lan, Y., Cai, D., and Zheng, Y.-Z. (2005). Expression in Escherichia coli of three different soybean late embryogenesis abundant (LEA) genes to investigate enhanced stress tolerance. J. Integr. Plant Biol. 47, 613–621. doi: 10.1111/j.1744-7909.2005.00025.x
Liu, Y., Wang, L., Xing, X., Sun, L., Pan, J., Kong, X., et al. (2013). ZmLEA3, a multifunctional Group 3 LEA protein from maize (Zea mays L.), is involved in biotic and abiotic stresses. Plant Cell Physiol. 54, 944–959. doi: 10.1093/pcp/pct047
Liu, Y.-L., and Zheng, Y.-Z. (2005). PM2, a group 3 LEA protein from soybean, and its 22-mer repeating region confer salt tolerance in Escherichia coli. Biochem. Biophys. Res. Commun. 331, 325–332. doi: 10.1016/j.bbrc.2005.03.165
Maldonado-Cervantes, E., Huerta-Ocampo, J. A., Montero-Moran, G. M., Barrera-Pacheco, A., Espitia-Rangel, E., and Barba de la Rosa, A. P. (2014). Characterization of Amaranthus cruentus L. seed proteins by 2-DE and LC/MS-MS: identification and cloning of a novel late embryogenesis-abundant protein. J. Cereal Sci. 60, 172–178. doi: 10.1016/j.jcs.2014.02.008
Maughan, P. J., Yourstone, S. M., Jellen, E. N., and Udall, J. A. (2009). SNP discovery via genomic reduction, barcoding, and 454-Pyrosequencing in amaranth. Plant Genome 2, 260–270. doi: 10.3835/plantgenome2009.08.0022
Mielke, S. P., and Krishnan, V. V. (2009). Characterization of protein secondary structure from NMR chemical shifts. Prog. Nucl. Magn. Reson. Spectrosc. 54, 141–165. doi: 10.1016/j.pnmrs.2008.06.002
Muller, I., Sarramégna, V., Renault, M., Lafaquière, V., Sebai, S., Milon, A., et al. (2008). The full-length Mu-opioid receptor: a conformational study by circular dichorism in trifluoroethanol and membrane-mimetic environments. J. Membrane Biol. 223, 49–57. doi: 10.1007/s00232-008-9112-x
Olvera-Carrillo, Y., Campos, F., Reyes, J. L., Garciarrubio, A., and Covarrubias, A. A. (2010). Functional analysis of the Group 4 late embryogenesis abundant proteins reveals their relevance in the adaptive response during water deficit in Arabidopsis. Plant Physiol. 154, 373–390. doi: 10.1104/pp.110.158964
Popova, A. V., Hundertmark, M., Seckler, R., and Hincha, D. K. (2011). Structural transitions in the intrinsically disordered plant dehydration stress protein LEA7 upon drying are modulated by the presence of membranes. Biochim. Biophys. Acta 1808, 1879–1887. doi: 10.1016/j.bbamem.2011.03.009
Rastogi, A., and Shukla, S. (2013). Amaranth: a new millennium crop of nutraceutical values. Crit. Rev. Food Sci. Nutr. 53, 109–125. doi: 10.1080/10408398.2010.517876
Reddy, P. S., Reddy, G. M., Pandey, P., Chandrasekhar, K., and Reddy, M. K. (2012). Cloning and molecular characterization of a gene encoding late embryogenesis abundant protein from Pennisetum glaucum: protection against abiotic stresses. Mol. Biol. Rep. 39, 7163–7174. doi: 10.1007/s11033-012-1548-5
Rivera-Najera, L. Y., Saab-Rincon, G., Battaglia, M., Amero, C., Pulido, N. O., Garcia-Hernandez, E., et al. (2014). A group 6 late embryogenesis abundant protein from common bean Is a disordered protein with extended helical structure and oligomer-forming properties. J. Biol. Chem. 289, 31995–32009. doi: 10.1074/jbc.M114.583369
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstruction phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Sasaki, K., Christov, N. K., Tsuda, S., and Imai, R. (2014). Identification of a novel LEA protein involved in freezing tolerance in wheat. Plant Cell Physiol. 55, 136–147. doi: 10.1093/pcp/pct164
Serrano, L., Sancho, J., Hirshberg, M., and Fersht, A. R. (1992). Alpha-helix stability in proteins. I. Empirical correlations concerning substitution of side-chains at the N and C-caps and the replacement of alanine by glycine or serine at solvent-exposed surfaces. J. Mol. Biol. 227, 544–559. doi: 10.1016/0022-2836(92)90906-Z
Shih, M. D., Lin, S. D., Hsieh, J. S., Tsou, C. H., Chow, T. Y., Lin, T. P., et al. (2004). Gene cloning and characterization of a soybean (Glycine max L.) LEA protein, GmPM16. Plant Mol. Biol. 56, 689–703. doi: 10.1007/s11103-004-4680-3
Silva-Sánchez, C., Barba de la Rosa, A. P., León-Galván, M. F., de Lumen, B. O., De León-Rodríguez, A., and González de Mejía, E. (2008). Bioactive peptides in amaranth (Amaranthus hypochondriacus) seed. J. Agric. Food Chem. 56, 1233–1240. doi: 10.1021/jf072911z
Singh, S., Cornilescu, C. C., Tyler, R. C., Cornilescu, G., Tonelli, M., Lee, M. S., et al. (2005). Solution structure of a late embryogenesis abundant protein (LEA14) from Arabidopsis thaliana, a cellular stress-related protein. Protein Sci. 2005, 2601–2609. doi: 10.1110/ps.051579205
Sivamani, E., Bahieldin, A., Wraith, J. M., Al-Niemi, T., Dyer, W. E., Ho, T., et al. (2000). Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 155, 1–9. doi: 10.1016/S0168-9452(99)00247-2
Soulages, J. L., Kim, K., Walters, C., and Cushman, J. C. (2002). Temperature-induced extended helix/random coil transitions in a group 1 late embryogenesis-abundant protein from soybean. Plant. Physiol. 128, 822–832. doi: 10.1104/pp.010521
Tolleter, D., Hincha, D. K., and Macherel, D. (2010). A mitochondrial late embryogenesis abundant protein stabilizes model membranes in the dry state. Biochim. Biophys. Acta 1798, 1926–1933. doi: 10.1016/j.bbamem.2010.06.029
Tolleter, D., Jaquinod, M., Mangavel, C., Passirani, C., Saulnier, P., Manon, S., et al. (2007). Structure and function of a mitochondrial late embryogenesis abundant protein are revealed by desiccation. Plant Cell 19, 1580–1589. doi: 10.1105/tpc.107.050104
Tompa, P., and Kovacs, D. (2010). Intrinsically disordered chaperones in plants and animals. Biochem. Cell Biol. 88, 167–174. doi: 10.1139/o09-163
Tunnacliffe, A., Hincha, D. K., Leprince, O., and Macherel, D. (2010). “LEA proteins: versa-tility of form and function,” in Sleeping Beauties: Dormancy and Resistance in Harsh Environments, eds E. Lubzens, J. Cerda, and M. Clark (Berlin: Springer), 91–108. doi: 10.1007/978-3-642-12422-8_6
Tunnacliffe, A., and Wise, M. J. (2007). The continuing conundrum of the LEA proteins. Naturwissenschaften 94, 791–812. doi: 10.1007/s00114-007-0254-y
Valdes-Rodríguez, S., Guerrero-Rangel, A., Melgoza-Villagomez, C., Chagolla-Lopez, A., Delgado-Vargas, F., Martinez-Gallardo, N., et al. (2007). Cloning of a cDNA encoding a cystatin from grain amaranth (Amaranthus hypochondriacus) showing a tissue-specific expression that is modified by germination and abiotic stress. Plant Physiol. Biochem. 45, 790–798. doi: 10.1016/j.plaphy.2007.07.007
van Leeuwen, M. R., Wyatt, T. T., van Doorn, T. M., Lugones, L. G., Wösten, H. A. B., and Dijksterhuis, J. (2016). Hydrophilins in the filamentous fungus Neosartorya fischeri (Aspergillus fischeri) have protective activity against several types of microbial water stress. Environ. Microbiol. Rep. 8, 45–52. doi: 10.1111/1758-2229.12349
Voinnet, O., Rivas, S., Mestre, P., and Baulcombe, D. (2003). An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 33, 949–956. doi: 10.1046/j.1365-313X.2003.01676.x
Warner, A. H., Guo, Z.-H., Moshi, S., Hudson, J. W., and Kozarova, A. (2016). Study of model systems to test the potential function of Artemia group 1 late embryogenesis abundant (LEA) proteins. Cell Stress Chaperones 21, 139–154. doi: 10.1007/s12192-015-0647-3
Wishart, D. S., Sykes, B. D., and Richards, F. M. (1991). Simple techniques for the quantification of protein secondary structure by H-1 NMR spectroscopy. FEBS Lett. 293, 1–2. doi: 10.1016/0014-5793(91)81155-2
Wolkers, W. F., McCready, S., Brandt, W. F., Lindsey, G. G., and Hoekstra, F. A. (2001). Isolation and characterization of a D-7 LEA protein from pollen that stabilizes glasses in vitro. Biochim. Biophys. Acta 1544, 196–206. doi: 10.1016/S0167-4838(00)00220-X
Wu, Y., Liu, C., Kuang, J., Ge, Q., Zhang, Y., and Wang, Z. (2014). Overexpression of SmLEA enhances salt and drought tolerance in Escherichia coli and Salvia miltiorrhiza. Protoplasma 251, 1191–1199. doi: 10.1007/s00709-014-0626-z
Keywords: amaranth seeds, circular dichroism, intrinsically disordered proteins (IDP), late embryogenesis abundant (LEA) proteins, nuclear magnetic resonance, Western blot
Citation: Saucedo AL, Hernández-Domínguez EE, de Luna-Valdez LA, Guevara-García AA, Escobedo-Moratilla A, Bojorquéz-Velázquez E, del Río-Portilla F, Fernández-Velasco DA and Barba de la Rosa AP (2017) Insights on Structure and Function of a Late Embryogenesis Abundant Protein from Amaranthus cruentus: An Intrinsically Disordered Protein Involved in Protection against Desiccation, Oxidant Conditions, and Osmotic Stress. Front. Plant Sci. 8:497. doi: 10.3389/fpls.2017.00497
Received: 22 December 2016; Accepted: 22 March 2017;
Published: 07 April 2017.
Edited by:
Dominique Job, Centre National de la Recherche Scientifique, FranceReviewed by:
Julia Buitink, National Institute for Agricultural Research, FranceTiago Santana Balbuena, São Paulo State University, Brazil
Copyright © 2017 Saucedo, Hernández-Domínguez, de Luna-Valdez, Guevara-García, Escobedo-Moratilla, Bojorquéz-Velázquez, del Río-Portilla, Fernández-Velasco and Barba de la Rosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana P. Barba de la Rosa, apbarba@ipicyt.edu.mx
†Present address: Alma L. Saucedo, Consejo Nacional de Ciencia y Tecnología, Laboratorio de RMN, Departamento de Química Analítica Facultad de Medicina, Universidad Autónoma de Nuevo León, Av. Madero y Av. Gonzalitos s/n, Colonia Mitras Centro, Monterrey, México; Eric E. Hernández-Domínguez, Consejo Nacional de Ciencia y Tecnología-Instituto de Ecología, A.C. Carretera antigua a Coatepec 351, El Haya, Xalapa de Enríquez, México