Effects of extrinsic factors on some rumination patterns: A review
 Fabio Abeni
Fabio Abeni- Centro di ricerca Zootecnia e Acquacoltura, Consiglio per la ricerca in agricoltura e l’analisi dell’economia agraria (CREA), Lodi, Italy
The rumen and its activity, rumination, are the characterizing traits of the suborder Ruminantia, and it is accompanied by related feeding habits and consequent animal behavior. Several extrinsic (not related to the animal itself) factors affect rumination behavior; most are reflected in rumination timing (considering overall daily duration as well as circadian differences in rumination patterns): age, environmental factors, and diet. For these reasons, great efforts have been sustained at the research level for monitoring rumination patterns. Currently, some research outcomes are applied at the farm level; others are still at the experimental level. All of these efforts are finalized mainly for the use of rumination pattern recording as an effective prediction tool for the early detection of health and welfare problems, both in a single head and at the herd level. Moreover, knowledge of the effects of extrinsic factors on rumination physiology represents a great challenge for improving the efficiency of ruminant livestock management, from the diet to the housing system, from parasites to heat stress. The present review deals mainly with the worldwide raised ruminant species.
Introduction on rumination and its monitoring: Methodological aspects and new opportunities from precision livestock farming
Interest in rumination has grown in recent years with the development and application of new technologies that allow the opportunity to better study, and sometimes then apply at the farm level, the relationships among the factors (intrinsic and extrinsic to the animal) affecting this important and specific feeding behavior. This interest is confirmed by a simple examination of the scientific literature between 2008 and 2021 by the bibliometrix R package (Aria and Cuccurullo, 2017). Starting from a simple association between “rumination” and “cattle”, the trend analysis of the extracted papers in Scopus reports a growth rate of 15.14% on a per-year basis, from 2008 to 2021, for the number of scientific papers on this topic. To begin our analysis of the extrinsic factors affecting rumination time, we searched and examined the main keywords in recent literature with relevant issues. Behind the generic term “cattle”, the dairy cow is the main area of interest for rumination, followed by “behavior”, “performance”, “system”, and “milk production,” which highlights how this kind of research is strongly related to the productive system. The same equilibrium among words was confirmed by the analysis of their growth pattern from 2008 to 2021 on Scopus.
Rumination is a specific feeding function that characterizes ruminants. Rumen and its activity, rumination, are the characterizing traits of the suborder Ruminantia, and it is accompanied byrelated feeding habits and consequent animal behavior. Rumination consists of a sequence of events involving ingesta, which is firstly regurgitated from the rumen, then re-masticated, and then re-swallowed (Beauchemin, 2018). During rumination, mastication is slower and more consistent when compared with eating (Beauchemin, 2018). In the mouth, the regurgitated bolus is re-masticated for about 30 to 70 s before being re-swallowed; the successive bolus will be regurgitated after 2 to 4 s. The rumination takes place in bouts lasting from 30 s to 2 h, reaching up to 20 rumination bouts/day (Beauchemin, 2018). Rumen and rumination are pivotal for the activity of microbial digestion, starting from the first months of life (Moraïs and Mizrahi, 2019).
The current literature indicates that maximum daily physiological rumination time (RT) ranges from 10 to 12 h/d (Beauchemin, 2018). However, total daily rumination time (TDRT) is not enough for a deep rumination pattern evaluation. As an overall daily activity index, TDRT may be a poorly useful and effective tool in predicting which factors may affect rumination activity. For this reason, this index, although on the basis of the other ones, has been modified, combining it according to some daily farm routines and to feeding management and ration nutritional profile (see the next section).
Contrary to the great amount of research into dairy and beef cattle, research on buffalo rumination is scarce. Launching “buffalo” and “rumination” in Scopus after very few recent references, we found the papers of Ali et al. (1990) and Odyuo et al. (1991), which represent the first steps in our knowledge about RT in buffalo according to parity and day–night pattern. Two papers by Galloso-Hernández et al. (2020; 2021) and one by Athaíde et al. (2020) reported the rumination response of buffalo under heat stress conditions. Only recently, Quddus et al. (2022) verified the performance of neck and ear tags in buffalo to estimate rumination time, finding a Pearson’s correlation of 0.85 and a concordance correlation coefficient (CCC) of 0.91 with visual observation. In small ruminants, the factors affecting rumination time and its derivative indexes are similar to those already examined in cattle; the specific interest for sheep is evidenced by the presence of 29 papers from 2008 to 2021, which represent 2% of the share in the considered period.
Indexes from rumination monitoring and combined indexes from rumination and other monitoring activities
One of the first approaches adopted to study rumination behavior was based on scanning techniques, where the subjects were time-lapse monitored, for example, every 5 min. The animals were considered to continuously ruminate if seen ruminating in the successive time scan, as described by Nielsen et al. (2000). In this method, rumination bouts were defined (and their duration calculated in a visual observation session) starting from the first time the animals were seen ruminating, and then considered as ruminating until the last time they were seen ruminating. Clearly, the precision of this kind of data is poor if compared to other currently available data from automated monitoring systems. Moreover, with the time-lapse video recording technique, there is a need for a visual interpretation of videos for behavioral pattern interpretation and for data collection.
Another possible approach to rumination monitoring may be that at the herd or group level by recording the percentage of animals ruminating at a specific time point (Fiol et al., 2019). This method could be useful if the aim is herd/cluster monitoring because we do not have single-head records.
Today, in both research and herd management, the possibility to automatically monitor the rumination patterns of each individual is a reality, which greatly increases the opportunity for farmers, technicians, and researchers to improve their work.
As rumen physiology items play a pivotal role in the effectiveness of early detection of health and welfare problems in ruminant livestock, the continuous recording of rumination and of other related indexes has a high impact in the field. The availability of precision livestock farming (PLF) tools for RT monitoring to a continuous extent gives us the opportunity to analyze daily RT throughout the day (Soriani et al., 2013; Abeni and Galli, 2017; Marino et al., 2021), and to derive the measure of its variability, improving the effectiveness and data quality on daily rumination timing. In fact, it is important to analyze the RT within the day because, namely during specific situations like heat stress, cattle can spread its RT according to the different environmental comfort conditions in the housing system (Abeni and Galli, 2017). Marchesini et al. (2018) created two different indexes for RT (as well as for activity recording): the dishomogeneity index within a day for rumination (DDR), and the dishomogeneity index within a period (i.e., days of observation) for rumination (DR). The DDR gives us a measure of the RT distribution within a day, whereas the DR gives us how RT changes day-by-day because of a possible problem of the ruminant to cope with extrinsic stressors (dietary, environmental, management, etc.), or as a possible outcome of intrinsic changes (for example, the approach of parturition).
Continuous rumination activity recording, carried out by some commercial tools, can be examined in its different components, which could be related to different problems. The TDRT is affected by the chemical and physical features of the diet as well as by several stressors; however, for a better understanding of the possible causes of a deviation from the normality, it is necessary to deconstruct TDRT relating it with other factors (Tomczak et al., 2019), as well as to calculate some useful derivative indexes considering other available data from the herd management system (see below and Figure 1).
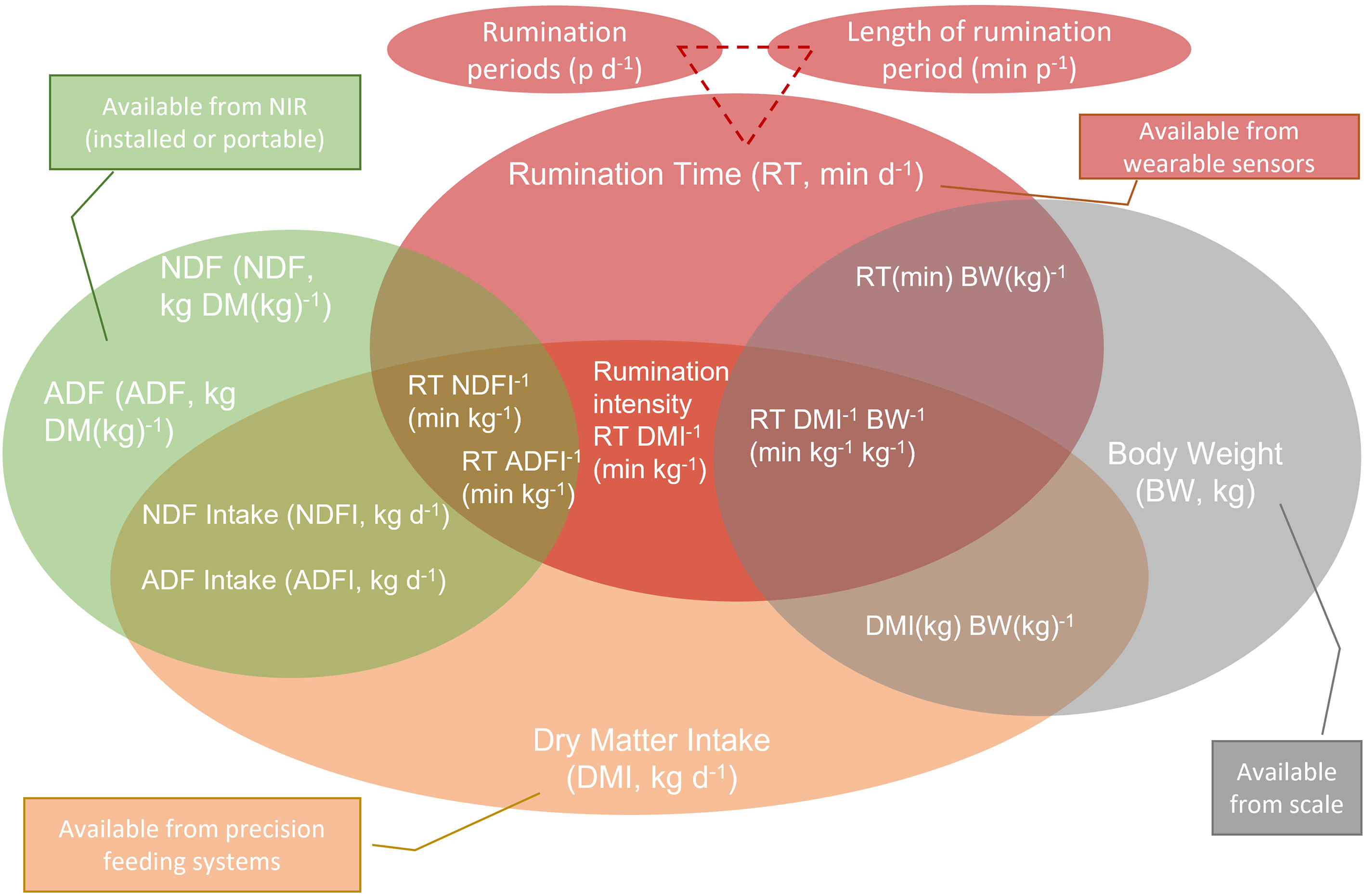
Figure 1 A picture of rumination-derived data available from digital on-farm technologies.
Performing rumination bout counts recorded by visual scanning systems is a scarcely precise technique and is not easily feasible at the farm level. It is possible to approximate the RT calculation, and at the same time, gives the number of bouts within a standard time (Nielsen et al., 2000). Nowadays, the introduction of continuous rumination pattern–recording tools allows us to detect hourly (sometimes bi-hourly) rumination time (HRT, min/h). This gives us the opportunity to assess some possible interactions between daily farm management routines (milking sessions, ration administration, etc.) and environmental conditions (Abeni and Galli, 2017). For example, the partition of RT between daytime and nighttime may be particularly useful in assessing how the cows in their environment recover from heat stress during the warmest summer period when this activity tends to shift from the day to the night hours (Soriani et al., 2013; Abeni and Galli, 2017). The following is a list of RT indexes, indicating the use and effectiveness of each.
Rumination time per kilogram of dry matter intake (RTDMI, min/kg DMI) is the total daily RT divided by the total daily DMI of the ration. This is an intuitive correction to normalize the recorded RT according to the DMI intake of the ruminant, generally affected by its body size and its individual (genetic and non-genetic) productive potential. It is also defined as rumination intensity (Watt et al., 2015).
Rumination time per kilogram NDF intake (RTNDFI, min/kg NDF intake) represents the normalization according to the total daily NDF intake, which depends on the DMI and the ration composition (mainly from the forage quality). A variant of this index is the rumination time per kilogram of NDF assayed with a heat-stable amylase and expressed inclusive of residual ash (aNDF).
Rumination time per kilogram of indigestible NDF intake (iNDFI) (RTiNDFI, min/kg iNDF intake) represents the adjustment for the indigestible NDF, generally associated with the forage degree of lignification, due to the phenological stage as well as to the hay/haylage/silage production technique.
Rumination time per kilogram of forage NDF intake (RTfNDFI, min/kg fNDF intake), as reported by Jiang et al. (2017), is more related to the quality of forage NDF.
Another way to relate the RT response to diet quality is to combine a chemical attribute (NDF) to the physical dimension of particle size (peNDF) fractions as determined by the Penn State Particle Separator (Zebeli et al., 2012), where peNDF1.18 is the dietary physically effective NDF inclusive of particles retained above the mesh with >1.18-mm holes, and peNDF8.0 is the dietary physically effective NDF inclusive of particles retained above the mesh with >8-mm holes. This way, the rumination time per gram of peNDF8.0 and rumination time per gram of peNDF1.18 can be calculated (Cao et al., 2013). Yang and Beauchemin (2007) reported a Pearson’s correlation coefficient of 0.59 between peNDF8.0 and rumination intensity expressed as min/kg DMI in dairy cows. With the improvement of knowledge on rumen dynamics, the most recent research considers the role of NDF above a mesh with 4.0-mm holes (peNDF4.0), and peNDF8.0. Therefore, we can consider the following indexes as reported by Jiang et al. (2017):
-rumination time per kilogram of physically effective NDF4.0 intake (RTpeNDF4.0I, min/kg peNDF4.0 intake);
-rumination time per kilogram of physically effective NDF8.0 intake (RTpeNDF8.0I, min/kg peNDF8.0 intake);
-rumination time per kilogram of digestible organic matter intake (DOMI) (RTDOMI, min/kg DOM intake), which is another way to link RT to ration quality and digestibility;
-rumination time per kilogram NDFI per 100 kg body weight (BW; RTNDFIBW, min/kg NDF intake per 100 kg BW), which is an essential adjustment that links the response to forage quality (NDF) to the size of the animal; this index is highly useful in growing ruminants (heifers, steers), but it is also important to evaluate RT in dairy cows at different parities as well as at different lactation and/or pregnancy stages; and
-rumination time per kilogram ADF (RTADFI, min/kg ADF intake), as reported by Jiang et al. (2017).
An example of some of these items is reported in Table 1 (adapted from Jardstedt et al., 2018).
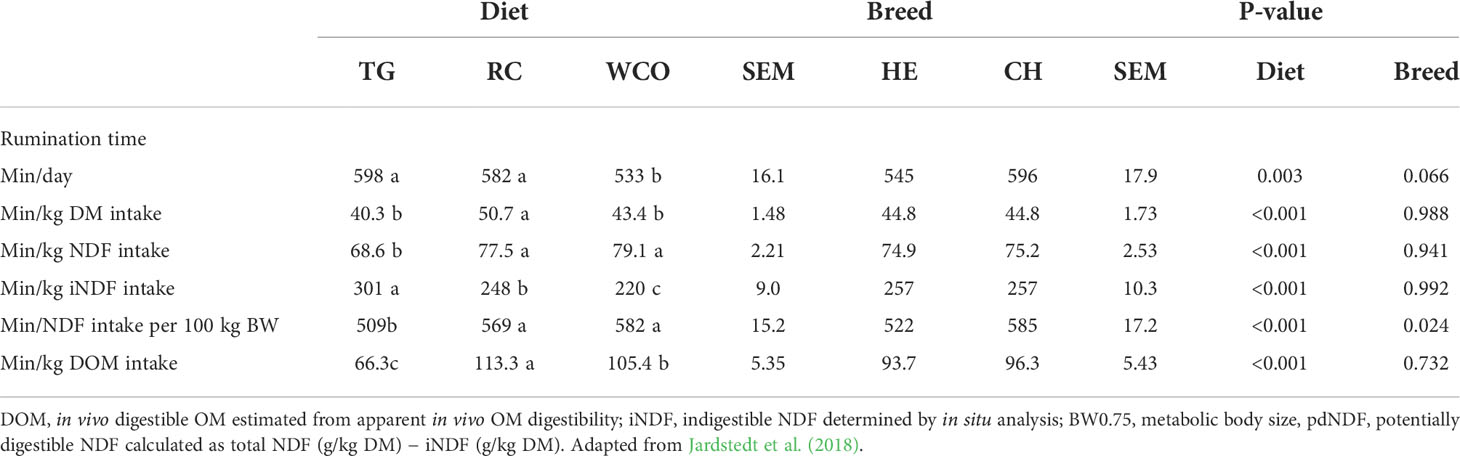
Table 1 Different ways to report the rumination time in beef cows of Hereford (HE) and Charolais (CH) breeds fed timothy silage (TG), reed canarygrass silage (RC), and whole-crop oat silage plus urea (WCO).
Other important items that can be really useful to understand the ruminant feeding behavior (as evidenced by Jiang et al., 2017) are the following:
- the number of ruminating periods (rumination periods per day), with ruminating period defined as at least 5 min of rumination occurring after at least 5 min without ruminating activity (Krause and Combs, 2003) and
- chewing time per ruminating period (min/ruminating period).
There is one system that also gives the number of boluses and the total chews (in addition to the rumination time); when these items were included in a model to predict health problems, the model reached the highest accuracy in early detection of sick cows (Fadul et al., 2022).
Nowadays, near-infrared reflectance (NIR) tools have improved the possibility of analyzing nutritional features of diets, allowing a more precise calculation of the previously listed indexes, detecting changes in real time, and are sometimes installed within systems that allow instantaneous correction in ingredient loading according to actually assessed features (namely, DM). NIR tools can be portable or mounted on a total mixed ration (TMR) mixer wagon. Moreover, NIR outcomes could be connected by an Internet of Things (IoT), as suggested by Rego et al. (2020) to give DM and NDF of the diet and therefore, supporting the calculation in real time of the derivative indexes as suggested in Figure 1.
Technologies and tools for automated recording
In the past, the studies on rumination patterns relied essentially on visual observation recording. Recently, Stygar et al. (2021) reported in their review a commentary on PLF tools for animal welfare assessment, classifying them as high- or low-performance tools, in comparison to the visual observation set as the gold standard. The performance was statistically defined by Pearson correlation coefficient (r), Spearman’s rank correlation coefficient (rs), concordance correlation coefficient (CCC, with a goal of > 0.90 according to Akoglu, 2018), sensitivity (Se), specificity (Sp), or the area under the receiver operating characteristic curve (AUC, with a goal of > 0.90); coefficient of determination (R2), and Cohen’s kappa coefficient (κ, with a goal of > 0.81); mean bias from the Bland–Altman plots (B–A plots) included zero with the 95% interval of agreement; and the significance tests for the intercept and slope of linear regression (I/S) that did not differ significantly from 0 or 1. Several tools for automated recording of this feeding behavior are available.
Comparison among the performance recorded for some available tools
Table 2 reports the prediction effectiveness of statistical indexes of some commercially available tools for rumination recording. The use of a collar-mounted microphone was tested in commercial farm environments by Ambriz-Vilchis et al. (2015). They reported a different reliability of the automated recorded RT (compared with visual observation or video recording) when cows were fed a partial mixed ration (PMR) integrated at stalls or when the cows grazed and were fed indoors with an integrative ration.
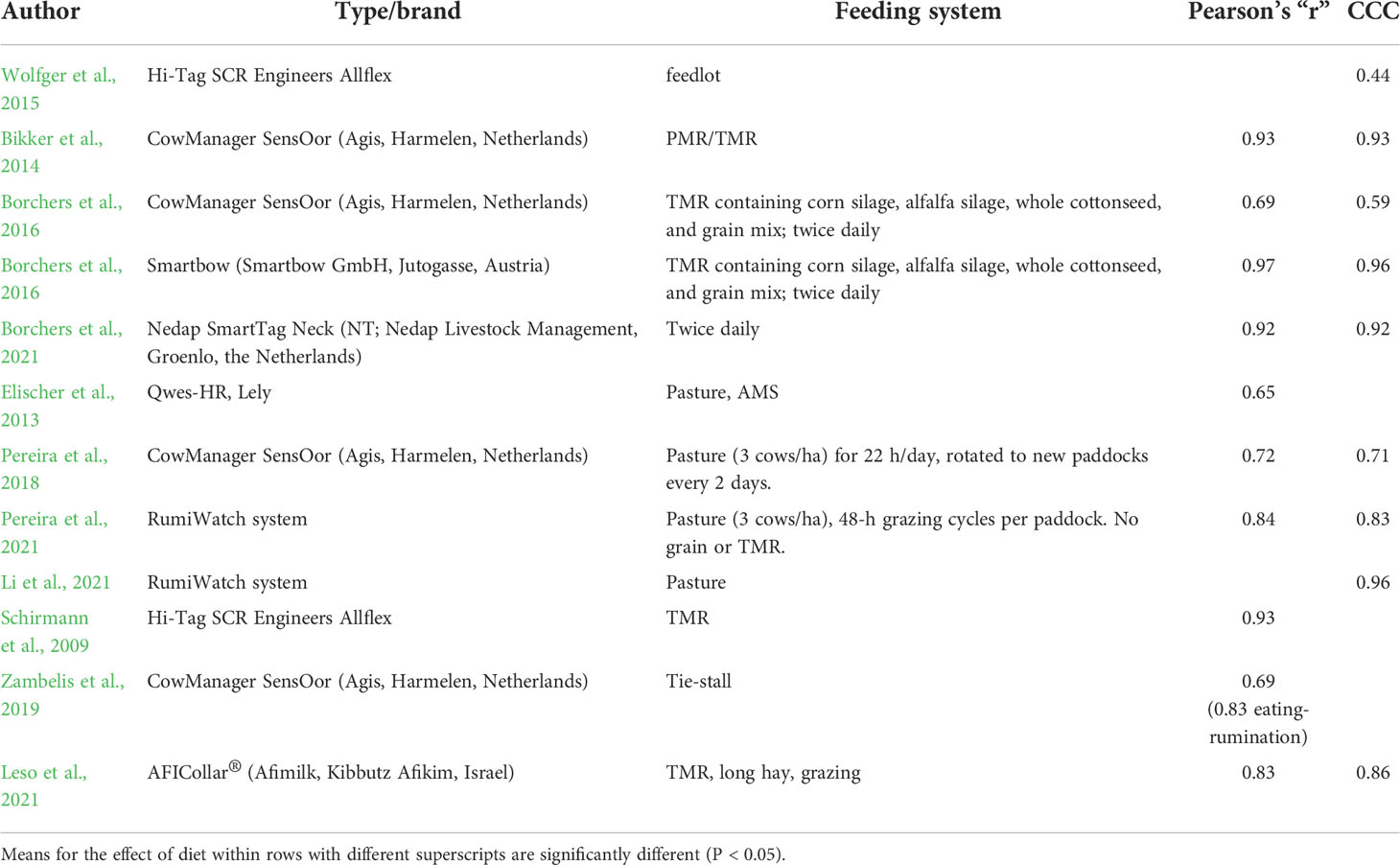
Table 2 Predictive statistics performance of some commercially available tools for rumination recording.
Zehner et al. (2017) defined the use of a noseband pressure sensor; Pereira et al. (2021) reported how the same tool was able to give rumination time (min/day) and the number of rumination cycles/5 min, which represents additional information to clarify the rumination response to external factors. Li et al. (2021) recently tested this device for monitoring the feeding behavior of grazing dairy cows. This device was also tested for water buffalo by Guccione et al. (2019).
For tie stall–housed dairy cows, the assessment of rumination time recording by an ear-tag accelerometer was validated by Zambelis et al. (2019), but they did not report the CCC.
Borchers et al. (2016) validated the rumination time recording with two triaxial accelerometers, concluding that visually recorded rumination behaviors were strongly correlated with the Smartbow ® (r = 0.97, CCC = 0.96) and weakly correlated with the CowManager® sensor (r = 0.69, CCC = 0.59). More recently, they validated a neck tag for rumination time recording, with good performance as confirmed by high Pearson’s (r = 0.92) and CCC (0.92) parameters (Borchers et al., 2021). Wolfger et al. (2015) validated the use of a 3D axial accelerometer also in feedlot cattle, but they obtained a low CCC.
An important part of the system is the software that manages raw data to extract information. Analyzing the performance of a new sensor, Leso et al. (2021) compared two different versions of the software for the same device and highlighted the higher reliability in detecting rumination patterns in the newest version.
An important issue is the possibility of these new tools detecting a significant difference between two (or more) groups of observations when the entity is small; practically, this involves how they can support a discrimination in both experimental and field conditions. The smallest difference in TDRT detected as significant (P < 0.05) among the consulted literature was found in Lockard et al. (2020), where a 21 min/day of difference between the two experimental thesis (dietary treatment in heat-stressed feedlot steers) was evaluated by the SCR system.
A picture (not exhaustive) of the actual availability of data from different tools and their possible integration to generate new data and indexes is reported in Figure 1.
Chemical and physical dietary features affecting rumination
According to Jiang et al. (2017) and White et al. (2017a), the main dietary factors that may affect rumination items in dairy cows are the following:
a. Forage species and their phenological stage at harvest.
b. Feed/diet composition (dietary roughage level as %; cottonseed content, expressed as % of DM in TMR; total NDF, NDF from forage, and CP in TMR expressed as % of DM; NDF and starch digestibility; and the % of wet forage, as silage or green fodder, expressed as % of DM in the TMR).
c. Particle size (diet mean particle size, MPS; % of particles with a length > 19 and 8 mm; NDF content in particle with length > 8 mm).
d. Feed/diet management (preparation, distribution, and productive level).
Forage species and their phenological stage at harvest
Rumination pattern modifications due to different forage sources are related to fecal particle size in pregnant beef cattle. For example, feeding reed canarygrass silage (RC) was associated with longer rumination time per kilogram DMI than feeding timothy silage and whole crop oat silage, because of the higher concentration of NDF in RC (Jardstedt et al., 2018). At the same time, differences in RTNDFI were justified by the differences in iNDF concentration per kilogram NDF of the diet (Jardstedt et al., 2018). The different forage sources were reflected in fecal particle sizes, with lower values for RC (Jardstedt et al., 2018).
Different grass silage maturities affected rumination values in dairy cows, but to a different extent for each item (Rinne et al., 2002), from early to late harvest class: + 17.6% for TDRT; + 27.3% for min/kg DMI; + 3.5% for min/kg NDF intake; and – 30.0% for min/kg indigestible NDF intake. As evidenced in Figure 2, reporting the values published by Rinne et al. (2002), rumination intensity may be a useful index to assess, also at farm level, the relationship between cow feeding behavior and diet (namely, forage) quality.
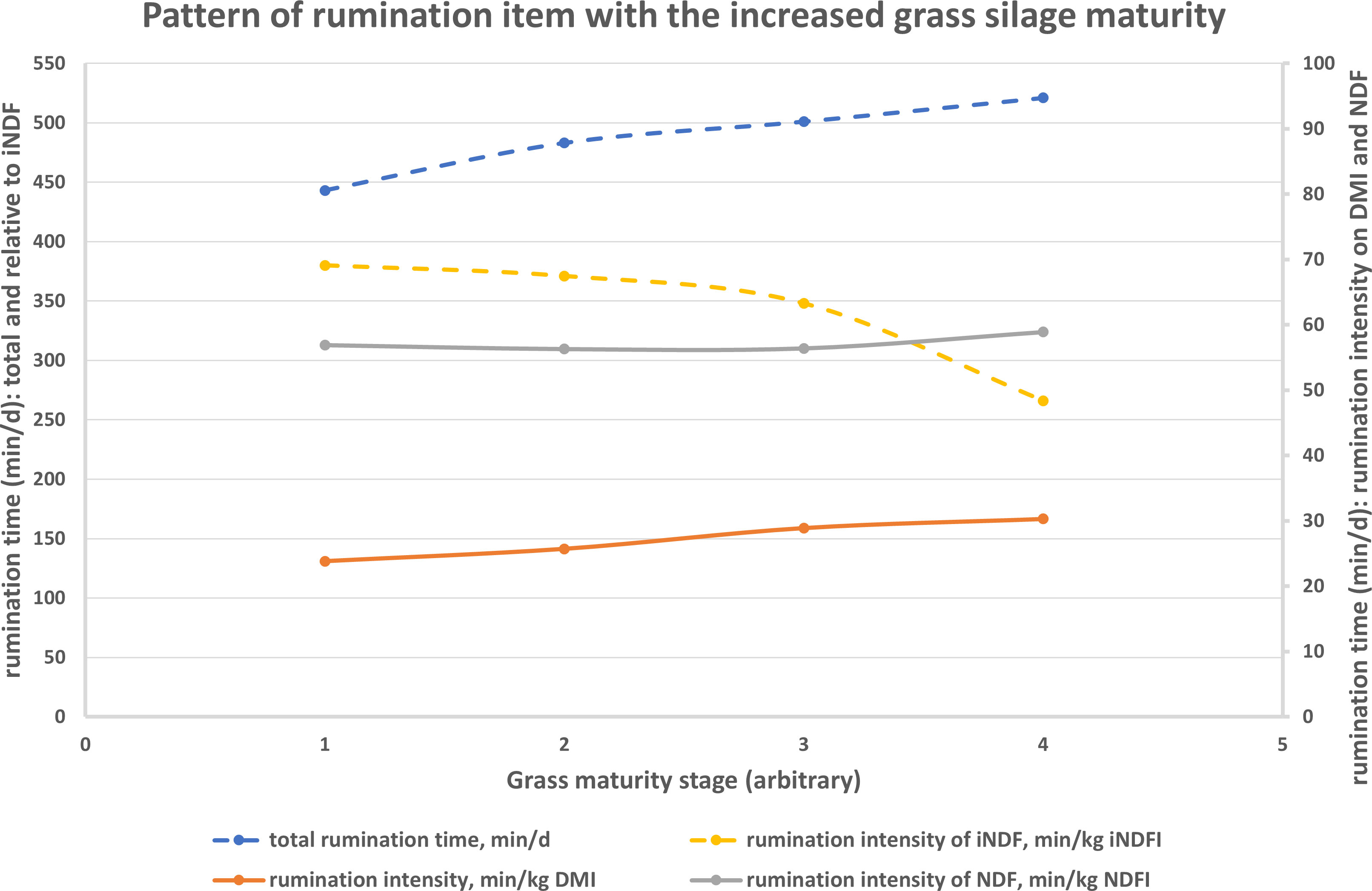
Figure 2 Relationship between advancing grass silage maturity and rumination items in lactating dairy cows according to the values published by Rinne et al. (2002).
Dietary fiber source affects rumination timing also in finishing steers. The replacement of grass hay with wood chips did not affect growth performance (probably because of the low level of inclusion), but it reduced DMI and increased resting time, with a drop in ruminating activity (Beretta et al., 2021).
Feed/diet composition
In the study by Jiang et al. (2017) on dairy cows, dietary roughage level did not affect RT (min/day), nor the number of ruminating periods (rumination periods per day), chewing time per ruminating period (min/ruminating period), RTNDFI, and total chewing time per kilogram of ADF intake, even if these results seem surprising. On the other hand, they reported how finely dietary roughage level linearly and negatively affected RTfNDFI, RTpeNDF4.0I, and RTpeNDF8.0I.
The percentage of peNDF (from 6.4% to 15.4%) in the diet of beef heifers linearly increases TDRT and RT per kilogram DM, but decreases RT per kilogram NDF (Llonch et al., 2020). The linear effect of barley silage inclusion from 0 to 12% of DM in feedlot cattle diet was evidenced by Chibisa et al. (2020), with a change from 246 to 316 min/day. The effect of forage inclusion level in dairy cow diet on RT was confirmed by Krogstad et al. (2021), evidencing how it did not interact with the form of dried distiller grains and solubles included in the TMR.
Nielsen et al. (2000) did not find differences in dairy cows fed two diets markedly different in concentrate content for a number of bouts and their duration recorded in the 6 h after the morning TMR administration. Those attempts would probably be more successful with current tools for automated recording of those items. Zebeli et al. (2012) reported the regression to estimate rumination time from peNDF>8 (see Figure 3).
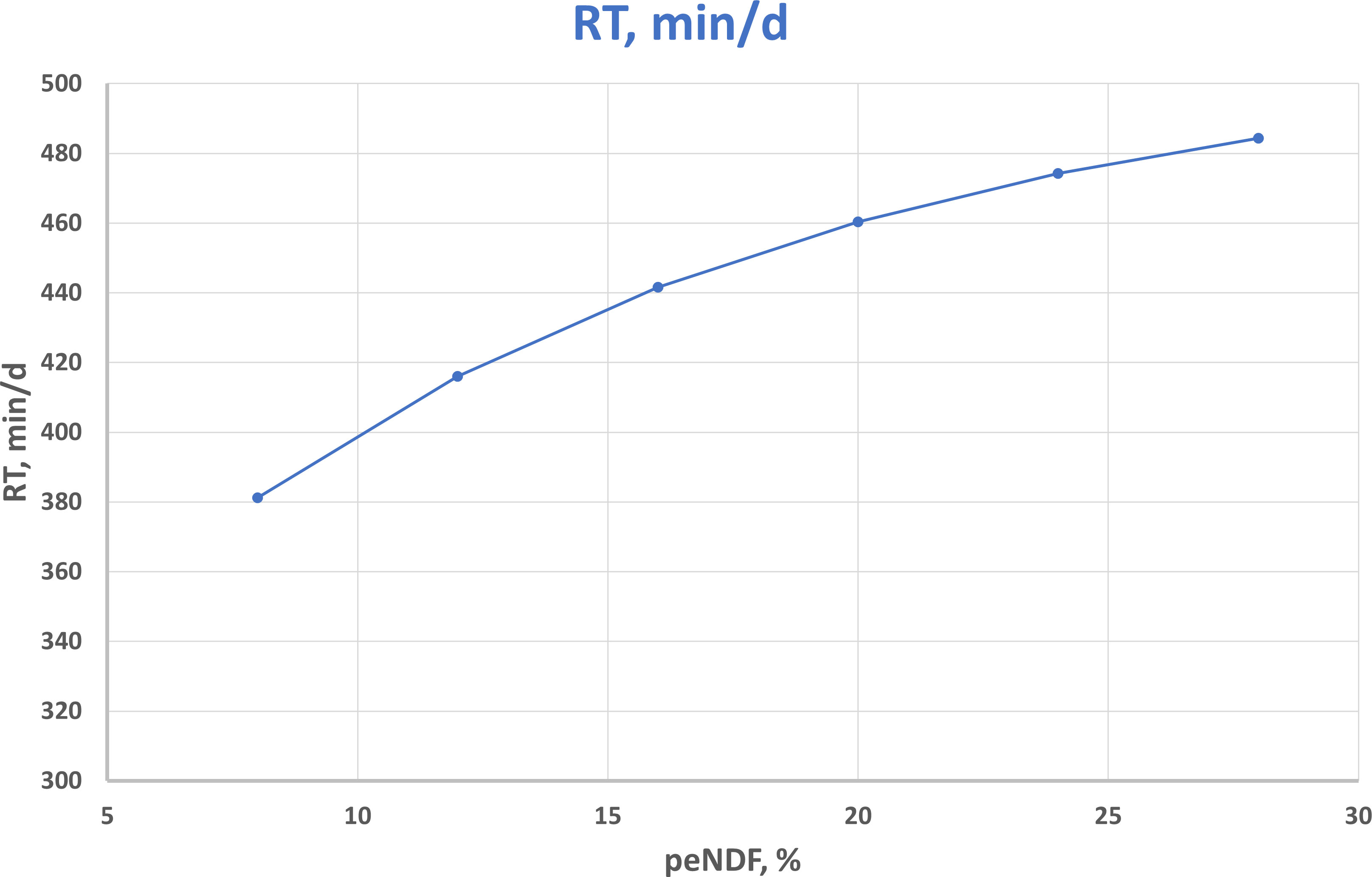
Figure 3 Relationship between physically effective NDF (peNDF) and rumination time in dairy cow according to the equation by Zebeli et al. (2012) where rumination time (RT, min/day) = 512.7 – 243.0 * exp -0.0768 * peNDF(%).
Bae et al. (1979) focused their attention on the relationships between hay intake and the different items of rumination in sheep. At that time, the reference for the dietary fiber supplied by hay was defined as the cell wall component (CWC). Rumination time, the number of chews during rumination, and the number of rumination bouts were quadratically increased with the increase in total hay intake when those items divided by the grams of CWC were inversely related to forage intake (Bae et al., 1979). At the same time, the number of chews per minute and per bolus increased, and the number of boluses per minute of rumination time decreased as hay intake increased (Bae et al., 1979).
From the feeding trials in buffalo husbandry, we only report how whole raw soybean grain (WRS) reduced the rumination activity in a study involving four castrated Murrah buffalo males in a 4 × 4 Latin square design with the other three diets differing in supplement content: control (not supplement), soybean oil, WRS, and calcium salts of fatty acids (de Aragão Bulcão et al., 2021). This very low number of subjects makes it difficult to conclude how rumination should be considered a good signal of poor welfare in buffalo; further research should be carried out for a deeper knowledge of the application in this species.
Particle size
Diet forage NDF and mean particle size are the main factors that affect TDRT (Beauchemin, 2018). The role of corn silage particle size in determining peNDF leads to a growing interest in the effects of its processing at harvest. This action is not only related to the theoretical length of cut (TLOC), but also to the presence of a kernel processor. The effects of different combinations of these factors in dairy cows were reported by Salvati et al. (2021): The kernel processor decreased at the same as the TLOC and TDRT, but within a same TLOC, the kernel processor decrease TLOC was associated with an increased TDRT.
White et al. (2017b) also proposed a useful model to predict, and then monitor, daily rumination time according to those extrinsic dietary factors reported in the formula:
Rumination time, min/day = −357 − 16.7 × MPS + 4.34 × 19-mm + 2.49 × 8-mm + 71.5 × DMI − 1.54 × (DMI × DMI) + 4.78 × NDF − 1.68 × dNDF − 2.35 × dStarch
where MPS is the mean particle size (mm) estimated for materials retained in each sieve of the Penn State Particle Separator, 19-mm and 8-mm are the percentage of TMR retained above mesh with 19- and 8-mm holes, dNDF is the rumen-degraded NDF, and dStarch is rumen-degraded starch.
Gentry et al. (2016) reported how particle size may affect RT in finishing steers. They showed how the grinding processing affected the daily RT, but also other rumination patterns related to the day of treatment, DM, NDF, and peNDF (Gentry et al., 2016). Marchesini et al. (2020) did not find any difference in RT, DDR, and DR in fattening bulls fed the same diet processed for a short (20 min) or long (30 min) time by the feeding mixer wagon.
Recent studies on factors affecting goat RT reported a significant effect of forage particle size (5 vs. 20 mm of the theoretical length of cut) and no significant effect of acid detergent lignin (ADL) on the aNDF ratio in forage (Cao et al., 2013). The effect of forage particle size was confirmed also by Arowolo et al. (2020) but with different absolute values of RT/day.
Feed/diet management
Figure 4 clearly shows the relationship between RT, both as absolute daily value and as rumination intensity per kilogram of DMI, and milk yield according to the equations by Byskov et al. (2015). It is clearly represented how RT is positively related to milk yield; on the other hand, the negative relationship between rumination intensity and milk yield is mainly due to the increase in DMI and concentrates that leads to higher milk yield and implies a partial decline in RT per DMI unit.
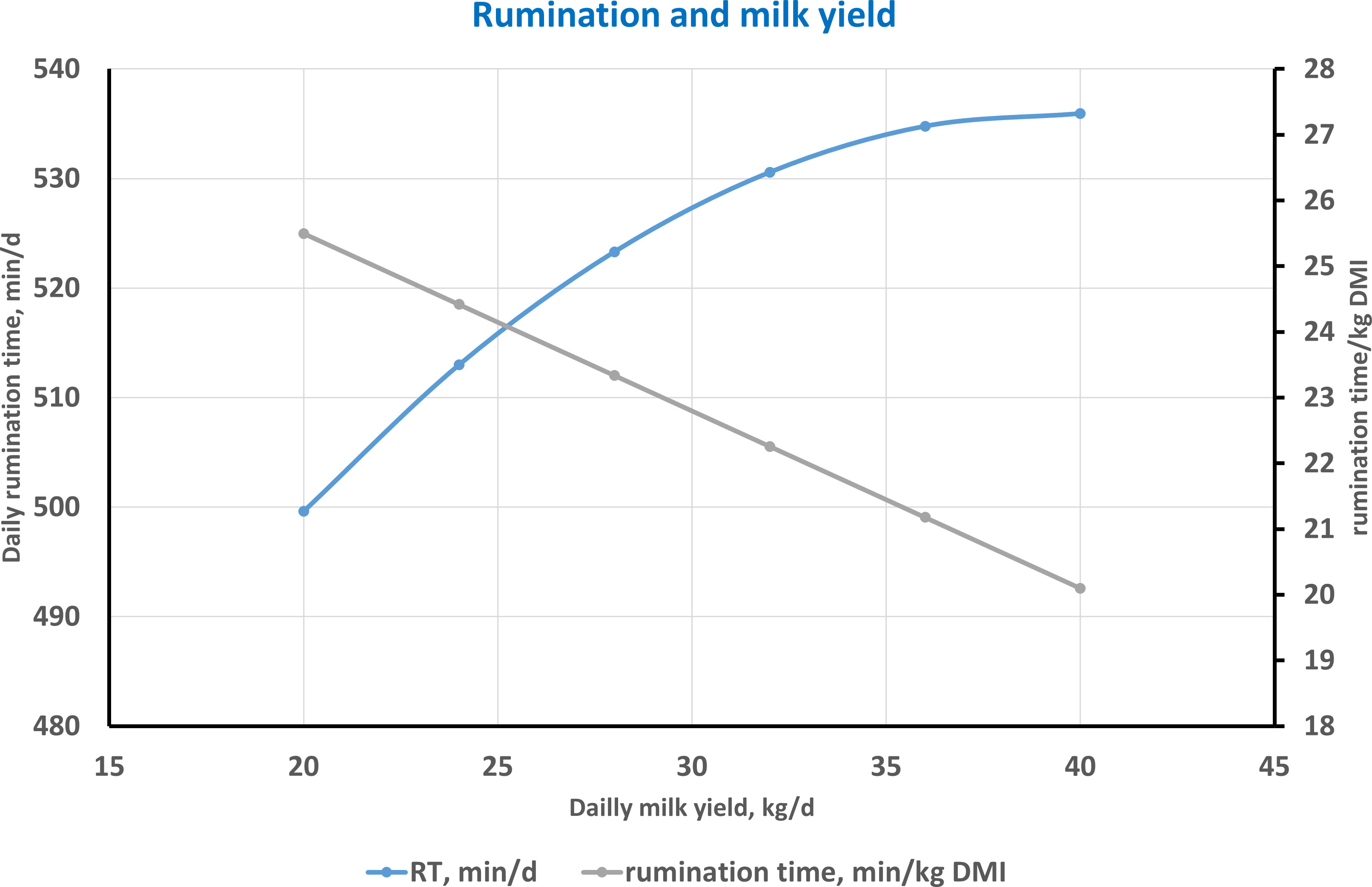
Figure 4 Relationship between daily milk yield and rumination time (RT) expressed as minutes per day and as minutes per kilogram of dry matter intake (DMI) according to the equations by Byskov et al. (2015).
In addition, rumination time affects the acid–base balance of rumen fluid. In cattle, this is essentially due to the high rate of saliva secretion during rumination. However, some differences in the reported salivation rate between beef and dairy cattle must be considered. In feedlot cattle, González et al. (2012) reported a value of 25 ml/min of ruminating steer, which was similar to that from an eating steer (20 ml/min) and about two times greater than the one during resting (10 ml/min). In dairy cows, Jiang et al. (2017) reported quite different (absolute) values as well as a different ranking in salivation rate for different cow states and according to the percentage of roughage in the diet: 192.4 to 201.2 ml/min during eating, and 128.4 to 139.6 ml/min during resting. Saliva has a high buffering capacity due to its high, although variable (Castillo-Lopez et al., 2021), concentration of bicarbonate and phosphate, as reviewed by González et al. (2012). Therefore, RT monitoring may be very important in fattening beef cattle because of the acidogenic diets generally supplied to this category to check if the acid production may be compensated by the saliva buffering activity. This is also an important issue for lactating dairy cows, where the adoption of a low forage-to-concentrate ratio often leads to acidosis or sub-acute ruminal acidosis (SARA), a condition where ruminal pH drops for several hours below the value of 5.5 (Oetzel, 2017). A quantitative scoring of rumen acidity was demonstrated to be associated with a different rumination profile throughout the day, with a proportional RT reduction for each quartile increase in rumen acidity (Saha et al., 2019).
Higher daily meal administration frequency improves the synchronization between bouts of acid production (from ration fermentation) and saliva production, also improving the organic acid absorption from the rumen (González et al., 2012). On the other hand, the effect of a direct addition of a ruminal buffer, such as bicarbonate, can negatively affect feeding behavior by reducing rumination and increasing meal size (as per kilogram of feed for each meal; González et al., 2012).
To prevent the excessive decrease in rumen pH caused by acidogenic diets, some additives could be used. Magrin et al. (2018) used a live yeast strain of Saccharomyces cerevisiae in a high-concentrate diet fed to finishing Charolais bulls. They showed how this dietary supplementation did not affect rumen pH, but it prevented the drop in RT, expressed as rumination intensity, at the end of the fourth month of the fattening period (12.8 vs. 15.9 min kg/DMI in control and yeast-treated finishing bulls, respectively).
In our evaluation, it is important to consider how rumination may be affected by other environmental factors related to the animal lying ability, when this feeding behavior can be facilitated. Additionally, as suggested by Beauchemin (2018), the different comminution of feed during eating (generally related to specific physical features of the forage) may confound the results reported on rumination time.
If rumination time, in the evaluation of dietary treatments, is strongly affected by the interaction with the first chewing during eating in response to dietary features, maybe RT could be less biased in evaluating the response to other environmental factors less affected by this eating–ruminating interaction.
Non-dietary environmental factors affecting rumination
Rumen activity and rumination time as affected by cattle housing and herd management
Rumination time may be affected by housing as well as by several other management factors.
In young cattle, rumination may be affected by competitive housing conditions (Fiol et al., 2019). In particular, the rumination time during the first 5 h after feed delivery was significantly higher in dominating heifers compared with subordinate heifers. This is probably the consequence of an increased dry matter intake of the first cows at feed delivery time; therefore, the practical value of a big difference in rumination time in the first hours after feed distribution could be considered as an effective detection tool of overcrowded groups, at least in the feeding area. Overcrowding of lactating dairy cows from a stocking density above 130% (compared to 100%) did not affect TDRT but caused a decrease in the proportion of RT within a stall as a percentage of the TDRT, suggesting an adaptation of ruminating behavior of the cows according to the more competitive conditions (Krawczel et al., 2012).
Sjostrom et al. (2019) reported how cows housed outdoors (on straw packs) had lower rumination from 0800 to 1000 h, and from 1800 to 2000 h, when compared to those housed indoors (in a compost-bedded pack barn), although the daily RT was higher for outdoors-kept cows; the same ranking was confirmed for RT per kilogram DMI (27.7 vs. 26.0 min/kg DMI for outdoors and indoors, respectively). These results suggest caution in the interpretation of RT data in relation to the housing system as well as according to feeding delivery. Moreover, this states how important the joint RT analysis is to hourly variability for the real detection of rumen physiology disorders rather than artifacts.
The same researchers studied how rumination time may be affected at pasture by horn fly disturbance: Daily RT was negatively correlated with the presence of horn flies (Sjostrom et al., 2016). Woolley et al. (2018) reported a decreased RT in grazing cows when treated with repellent against pest flies (therefore less disturbed); their results, differing from the previous study, were justified by increased grazing activity when cows were less disturbed by flies. The interaction between rumination and flies appears specific for each management condition. Regarding environmental enrichment, Park et al. (2020) reported a positive effect from the availability of a brush on feedlot steer behavior with an increased RT. This result may be related to fewer stereotypes and less aggressive behavior of the steers with brush availability compared to those without brush, suggesting the importance of pen comfort also for fattening cattle.
Because rumination has a peak at approximately 4 h after feeding and rumination is often related to specific body positions (standing; lying and its laterality), there is a high relationship between feeding timing and frequency, as well as with the time spent lying, but Schirmann et al. (2012) reported a weak positive correlation between rumination and lying time. These relationships might be useful to improve feeding management and coordinate it with the other tasks in and around the barn with the aim of guaranteeing the required amount of daily time for lying and ruminating.
These findings confirm how daily RT indexes are suitable markers in animal welfare evaluation, as well as in early detection of stress-coping aptitude by individuals or groups in farm conditions. On the other hand, RT may not be considered a reliable biomarker of a generic chronic stress, being particularly affected by the type of stress source, as recently demonstrated by Grelet et al. (2022).
Rumination time and grazing management
Among the factors affecting rumination timing patterns, grazing management is often poorly investigated. However, two specific papers from New Zealand (Gregorini et al., 2012; Gregorini et al., 2013) reported how grazing schedule according to milking timing may affect total daily RT, with higher values when grazing time was not restricted compared to both two time intervals of 4 h or a single 8-h time interval of access to pasture between milking (Gregorini et al., 2012). In those conditions, age, breed, and genetic merit did not affect total RT, but are reflected in different rumination patterns, with evident differentiation in younger cows that exhibit lower values in the latter part of the day (Gregorini et al., 2013). From a practical point of view, the ability to monitor this pattern may be helpful in labor planning (first for milking) in pasture-based systems and in the interpretation of data from a grazing management system.
The study by Yang et al. (2021) focused on the effect of a different shrub availability on the time partition among grazing, walking, and ruminating/resting in yak; in their case, a decrease in rumination time with the higher shrub availability was the consequence of an increased walking activity as an adaptive strategy to increase food intake.
Rumen activity, rumination time, and parasites
The presence of parasites affects the gastrointestinal physiology. Gibb et al. (2005) reported a trend for an improved RT from the treatment with anthelmintic products in dairy cows, but only during July, probably as a consequence of a treatment by time effect in the summer season. Högberg et al. (2021) reported a transient effect of a similar treatment, but in young steers with a challenge of gastrointestinal-nematode larvae. The effect of a gastrointestinal nematode infection does not necessarily affect mean daily RT; the main effect was observed on an increased variability in RT especially during the first 40 days of infection (Högberg et al., 2021). This is essentially due to a subsequent adaptation pathway of the gastrointestinal tract (GIT) tissues to the infection (Högberg et al., 2021). These few available data support the hypothesis of a possible role of RT in gastrointestinal parasite detection in the first stage of the problem.
Rumen activity and rumination time as affected by climate
Rumen activity may be affected in its different items by heat stress. Rumen volatile fatty acid (VFA) production and absorption may be affected both directly and indirectly by heat stress. In an experimental trial, a high environmental temperature reduced ruminal butyrate absorption (Bedford et al., 2020). A reduced butyrate absorption in heat-stressed heifers was related to a decreased expression of transporters responsible for the transport of VFA metabolites into the blood (Bedford et al., 2020), and this did not take place in pair-fed heifers with the same reduced feed intake. This difference in heifers with the same DMI reduction, but evoked in two distinct ways, is probably the result of a backlog of butyrate metabolite export in heat-stressed animals. The authors suggested that during heat stress, an increased acetate absorption in turn led to increased conversion of both acetate and butyrate to β-hydroxybutyrate (BHB), and thus decreased the expression of transporters to move BHB out of the epithelial cell. These effects create a high concentration gradient that impairs butyrate absorption from the rumen (Bedford et al., 2020).
Soriani et al. (2013) reported a proportional decrease in total daily RT with increased Temperature Humidity Index (THI), jointly with a slight shift in RT from daytime to nighttime (reaching 63.2% of the daily RT value); this pattern was confirmed by Abeni and Galli (2017). This is the evidence of an adaptation behavioral pattern of cows, probably also related to a change in the daily timing in DMI.
Environmental climatic patterns may affect the possibility to use RT as a marker for some health problems. A good example comes from Held-Montaldo et al. (2021) that evidenced how climate (expressed as season) affected RT evaluation in detecting metritis. Monitoring a total of 103 multiparous cows, RT was affected in both prepartum and postpartum during autumn calving season, with an average of 5% to 6% decline (on a daily basis) in cows that experienced metritis, but this did not take place in spring calving season. In the same paper, this was clearly attributed to the THI level between 2 and 10 days after calving (Held-Montaldo et al., 2021). According to these results, it is important to consider RT pattern as a marker of different co-existing factors affecting it; therefore, great prudence must be used in the RT data interpretation.
The role of RT as a suitable marker in cattle welfare assessment was also confirmed by Magrin et al. (2017) in finishing young bulls that, when exposed to THI over the physiological thermal comfort threshold, maintained an acceptable RT (close to 10 min/h) with ceiling fans to mitigate heat stress, compared to control bulls (with RT below 6 min/h).
When we take into account a ruminant particularly adapted to extreme (cold) environmental conditions as is the yak, rumination must be viewed with care in its interpretation as a feeding behavior marker. For example, the paper by Liu et al. (2019) reported a positive correlation between environmental temperature and rumination time; however, their conditions were not comparable to those reported in the formerly quoted research with Bos taurus breeds. This comparison was between a very cold winter and a summer remarkably under thermoneutral conditions for Bos taurus breeds. Therefore, the interpretation of that study on yak suggests a relationship between rumination time and an improvement in grazing conditions that become more favorable for an increased forage intake, and therefore for rumination.
Dealing with climate challenges, Kennedy (1985) analyzed the effect of cold exposure on the main measurements of rumination behavior in sheep: chews/day, chews/g CWC intake, duration (min/day), chews/min, chews/cycle, and cycles. The results evidenced how cold exposure (1–4°C for four periods of 45 days compared to 22–25°C) did not affect (P > 0.05) any of the measured indexes of rumination.
Rumination in extensive agroforestry and silvopastoral systems
In extensive agroforestry and pastoral systems, here considered across all ruminant species, rumination can be considered as an animal welfare marker in an integrated approach, where not only dietary factors are important, and where climate is only one aspect of the interaction between the animal and the environment. In these cases, also from a scientific perspective, the individual RT is less suitable than the percentage of ruminating animals (both standing and lying) in the different daily timings (Tenffen de Sousa et al., 2021). In this context, RT evaluated through the different stages of the day and season may offer an assessment of animal welfare and adaptation (also from a foraging point of view) in cattle under different silvopastoral systems.
The main body of results from agroforestry systems with dairy cattle was from Brazil. At the agroforestry level, the percentage of heifers and cows ruminating in a specific time is considered a reliable marker (at the research level) of a successful strategy to mitigate heat stress (HS), generally by shading trees (Giro et al., 2019; Carnevalli et al., 2020; Deniz et al., 2020; Tenffen de Sousa et al., 2021). Extensive grazing systems for beef and dairy cattle are generally perceived (by the consumer) to be welfare-friendly for the animal. However, people often forget the stressful conditions due to climate, not only by air temperature, but also by direct solar radiation, on grazing ruminants. The response of cattle to shade is clearly positive, with higher percentages of ruminating animals and longer RT than those not in tree-shaded groups.
As previously stated for other ruminants, there is a growing interest in silvopastoral systems in buffalo husbandry. Water buffalo RT (visually assessed from 0600 to 1800 h) did not differ among heifers in different HS conditions under conventional or silvopastoral systems; however, this evaluation was evidenced in a very limited number of heifers (n = 9) and for a limited daytime (Galloso-Hernández et al., 2021). The same small sample size was reported in a previous study by the same researchers (Galloso-Hernández et al., 2020), but with a significant increase of RT (0600 to 1800 h) in moderate HS compared to intense HS, confirming the “adaptation strategy” already well described in cows (Calamari et al., 2014; Abeni and Galli, 2017). Athaíde et al. (2020) highlighted a positive effect of shade availability in a comparison between two groups of 10 buffaloes each.
Conclusion
The rumination activity patterns in several ruminant species, by monitoring their rumination time and its many derivative indexes, are nowadays easily available; this allows farmers and their consultants to detect welfare and health problems early and in a more improved manner, increasing their effectiveness in preventing losses in production efficiency. If dietary factors are generally the first sources of variability in these items, this review highlights how other biotic (parasites) and abiotic (environmental climate, housing facilities, and management) conditions may affect RT in several species. Along with this, the present review also underlines the lack of current knowledge and the need to carry out more research on this topic and on its applications at the farm level.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Funding
This work was supported by the project “Agricoltura Digitale–AGRIDIGIT”, subproject “Tecnologie digitali nella filiera del latte bovino e bufalino–PLF4Milk” (MiPAAF, D.M. 36507/7305/2018 del 20/12/2018).
Acknowledgments
I want to thank Prof. Pasquale De Palo (Università degli Studi di Bari “Aldo Moro”) for his helpful suggestions after the reading of a first draft of this paper.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abeni F., Galli A. (2017). Monitoring cow activity and rumination time for an early detection of heat stress in dairy cow. Int. J. Biometeorol. 61 (3), 417–425. doi: 10.1007/s00484-016-1222-z
Akoglu H. (2018). User’s guide to correlation coefficients. Turk. J. Emerg. Med. 18 (3), 91–93. doi: 10.1016/j.tjem.2018.08.001
Ali A., Raza S. H., Ghaffar A. (1990). Eating and rumination in relation to age of lactating buffalo. Appl. Anim. Behav. Sci. 28 (3), 273–279. doi: 10.1016/0168-1591(90)90106-N
Ambriz-Vilchis V., Jessop N. S., Fawcett R. H., Shaw D. J., Macrae A. I. (2015). Comparison of rumination activity measured using rumination collars against direct visual observations and analysis of video recordings of dairy cows in commercial farm environments. J. Dairy Sci. 98 (3), 1750–1758. doi: 10.3168/jds.2014-8565
Aria M., Cuccurullo C. (2017). Bibliometrix: An r-tool for comprehensive science mapping analysis. J. Informetrics 11 (4), 959–975. doi: 10.1016/j.joi.2017.08.007
Arowolo M. A., Yang S., Wang M., He J. H., Wang C., Wang R., et al. (2020). The effect of forage theoretical cut lengths on chewing activity, rumen fermentation, dissolved gases, and methane emissions in goats. Anim. Feed Sci. Technol. 263, 114454. doi: 10.1016/j.anifeedsci.2020.114454
Athaíde L. G., Joset W. C. L., de Almeida J. C. F., Pantoja M. H. D. A., Noronha R. D. P. P., Bezerra A. S., et al. (2020). Thermoregulatory and behavioral responses of buffaloes with and without direct sun exposure during abnormal environmental condition in marajó island, pará, Brazil. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.522551
Bae D. H., Welch J. G., Smith A. M. (1979). Forage intake and rumination by sheep. J. Anim. Sci. 49 (5), 1292–1299. doi: 10.2527/jas1979.4951292x
Beauchemin K. A. (2018). Invited review: Current perspectives on eating and rumination activity in dairy cows. J. Dairy Sci. 101 (6), 4762–4784. doi: 10.3168/jds.2017-13706
Bedford A., Beckett L., Harthan L., Wang C., Jiang N., Schramm H., et al. (2020). Ruminal volatile fatty acid absorption is affected by elevated ambient temperature. Sci. Rep. 10 (10), 13092. doi: 10.1038/s41598-020-69915-x
Beretta V., Simeone A., Franco J., Bentancur O., Casaretto A., Mondelli S., et al. (2021). Effect of totally replacing hay by wood chips and feed delivery system on performance, behavior and carcass traits of finishing steers. Livestock Sci. 243, 104384. doi: 10.1016/j.livsci.2020.104384
Bikker J. P., Van Laar H., Rump P., Doorenbos J., Van Meurs K., Griffioen G. M., et al. (2014). Evaluation of an ear-attached movement sensor to record cow feeding behavior and activity. J. Dairy Sci. 97 (5), 2974–2979. doi: 10.3168/jds.2013-7560
Borchers M. R., Chang Y. M., Tsai I. C., Wadsworth B. A., Bewley J. M. (2016). A validation of technologies monitoring dairy cow feeding, ruminating, and lying behaviors. J. Dairy Sci. 99 (9), 7458–7466. doi: 10.3168/jds.2015-10843
Borchers M. R., Gavigan S., Harbers A., Bewley J. (2021). An evaluation of a novel device for measuring eating, rumination, and inactive behaviors in lactating Holstein dairy cattle. Animal 15, 100008. doi: 10.1016/j.animal.2020.100008
Byskov M. V., Nadeau E., Johansson B. E. O., Nørgaard P. (2015). Variations in automatically recorded rumination time as explained by variations in intake of dietary fractions and milk production, and between-cow variation. J. Dairy Sci. 98 (6), 3926–3937. doi: 10.3168/jds.2014-8012
Cao Y. C., Gao Y., Xu M., Liu N. N., Zhao X. H., Liu C. J., et al. (2013). Effect of ADL to aNDF ratio and ryegrass particle length on chewing, ruminal fermentation, and in situ degradability in goats. Anim. Feed Sci. Technol. 186, 112–119. doi: 10.1016/j.anifeedsci.2013.08.010
Calamari L., Soriani N., Panella G., Petrera F., Minuti A., Trevisi E. (2014). Rumination time around calving: An early signal to detect cows at greater risk of disease. J. Dairy Sci. 97 (6), 3635–3647. doi: 10.3168/jds.2013-7709
Carnevalli R. A., de Mello A. C. T., Coletti A. J., Garcia L. F., Xavier D. B. (2020). Shade controls the ruminating and idleness times of dairy heifers in tropical integrated systems. Agrofor Syst. 94 (3), 779–790. doi: 10.1007/s10457-019-00448-7
Castillo-Lopez E., Petri R. M., Ricci S., Rivera-Chacon R., Sener-Aydemir A., Sharma S., et al. (2021). Dynamic changes in salivation, salivary composition, and rumen fermentation associated with duration of high-grain feeding in cows. J. Dairy Sci. 104 (4), 4875–4892. doi: 10.3168/jds.2020-19142
Chibisa G. E., Beauchemin K. A., Koenig K. M., Penner G. B. (2020). Optimum roughage proportion in barley-based feedlot cattle diets: Total tract nutrient digestibility, rumination, ruminal acidosis, short-chain fatty absorption, and gastrointestinal tract barrier function. J. Anim. Sci. 98 (6), 1–14. doi: 10.1093/jas/skaa160
de Aragão Bulcão L. F., Alba H. D. R., de Carvalho G. G. P., de Araújo M.L.G.M.L., Gandra J. R., Ribeiro C. V. D. M., et al. (2021). Digestion, ruminal metabolism, and feeding behavior of buffaloes fed diets supplemented with soybean oil, whole and raw soybean, and calcium salts of fatty acids. Trop. Anim. Health Product. 53 (2), 216. doi: 10.1007/s11250-021-02654-x
Deniz M., Schmitt Filho A. L., Hötzel M. J., de Sousa K. T., Pinheiro Machado Filho L. C., Sinisgalli P. A. (2020). Microclimate and pasture area preferences by dairy cows under high biodiversity silvopastoral system in southern Brazil. Int. J. Biometeorol. 64 (11), 1877–1887. doi: 10.1007/s00484-020-01975-0
Elischer M. F., Arceo M. E., Karcher E. L., Siegford J. M. (2013). Validating the accuracy of activity and rumination monitor data from dairy cows housed in a pasture-based automatic milking system. J. Dairy Sci. 96 (10), 6412–6422. doi: 10.3168/jds.2013-6790
Fadul M., D’Andrea L., Alsaaod M., Borriello G., Di Lori A., Stucki D., et al. (2022). Assessment of feeding, ruminating and locomotion behaviors in dairy cows around calving – a retrospective clinical study to early detect spontaneous disease appearance. PloS One 17 (3), e0264834. doi: 10.1371/journal.pone.0264834
Fiol C., Aguerre M., Carriquiry M., Ungerfeld R. (2019). Social dominance affects intake rate and behavioral time budget in pre-pubertal dairy heifers allocated in continuous competitive situations. Animal 13 (6), 1297–1303. doi: 10.1017/S1751731118002835
Galloso-Hernández M. A., Rodríguez-Estévez V., Alvarez-Díaz C. A., Soca-Pérez M., Dublin D., Iglesias-Gómez J., et al. (2020). Effect of silvopastoral systems in the thermoregulatory and feeding behaviors of water buffaloes under different conditions of heat stress. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.00393
Galloso-Hernández M. A., Soca-Pérez M., Dublin D., Alvarez-Díaz C. A., Iglesias-Gómez J., Díaz-Gaona C., et al. (2021). Thermoregulatory and feeding behavior under different management and heat stress conditions in heifer water buffalo (Bubalus bubalis) in the tropics. Animals 11, 1162. doi: 10.3390/ani11041162
Gentry W. W., Weiss C. P., Meredith C. M., McCollum F. T., Cole N. A., Jennings J. S. (2016). Effects of roughage inclusion and particle size on performance and rumination behavior of finishing beef steers. J. Anim. Sci. 94 (11), 4759–4770. doi: 10.2527/jas2016-0734
Gibb M. J., Huckle C. A., Forbes A. B. (2005). Effects of sequential treatments with eprinomectin on performance and grazing behaviour in dairy cattle under daily-paddock stocking management. Vet. Parasitol. 133 (1), 79–90. doi: 10.1016/j.vetpar.2004.11.038
Giro A., Macedo Pezzopane J. R., Barioni Junior W., de Faria Pedroso A., Prudêncio Lemes A., Botta D., et al. (2019). Behavior and body surface temperature of beef cattle in integrated crop-livestock systems with or without tree shading. Sci. Total Environ. 684, 587–596. doi: 10.1016/j.scitotenv.2019.05.377
González L. A., Manteca X., Calsamiglia S., Schwartzkopf-Genswein K. S., Ferret A. (2012). Ruminal acidosis in feedlot cattle: Interplay between feed ingredients, rumen function and feeding behavior (a review). Anim. Feed Sci. Technol. 172, 66–79. doi: 10.1016/j.anifeedsci.2011.12.009
Gregorini P., DelaRue B., McLeod K., Clark C. E. F., Glassey C. B., Jago J. (2012). Rumination behavior of grazing dairy cows in response to restricted time at pasture. Livestock Sci. 146 (1), 95–98. doi: 10.1016/j.livsci.2012.02.020
Gregorini P., Dela Rue B., Pourau M., Glassey C., Jago J. (2013). A note on rumination behavior of dairy cows under intensive grazing systems. Livestock Sci. 158 (1-3), 151–156. doi: 10.1016/j.livsci.2013.10.012
Grelet C., Vanden Dries V., Leblois J., Wavreille J., Mirabito L., Soyeurt H., et al. (2022). Identification of chronic stress biomarkers in dairy cows. Animal 16 (5), 100502. doi: 10.1016/j.animal.2022.100502
Guccione J., D’Andrea L., Alsaaod M., Borriello G., Steiner A., Ciaramella P. (2019). Validation of a noseband pressure sensor algorithm as a tool for evaluation of feeding behaviour in dairy Mediterranean buffalo (Bubalus bubalis). J. Dairy Res. 86 (1), 40–42. doi: 10.1017/S0022029919000074
Held-Montaldo R., Cartes D., Sepúlveda-Varas P. (2021). Behavioral changes in dairy cows with metritis in seasonal calving pasture-based dairy system. J. Dairy Sci. 104 (11), 12066–12078. doi: 10.3168/jds.2021-20424
Högberg N., Hessle A., Lidfors L., Baltrušis P., Claerebout E., Höglund J. (2021). Subclinical nematode parasitism affects activity and rumination patterns in first-season grazing cattle. Animal 15, 100237. doi: 10.1016/j.animal.2021.100237
Jardstedt M., Hessle A., Nørgaard P., Frendberg L., Nadeau E. (2018). Intake and feed utilization in two breeds of pregnant beef cows fed forages with high-fiber concentrations. J. Anim. Sci. 96 (8), 3398–3411. doi: 10.1093/jas/sky199
Jiang F. G., Lin X. Y., Yan Z. G., Hu Z. Y., Liu G. M., Sun Y. D., et al. (2017). Effect of dietary roughage level on chewing activity, ruminal pH, and saliva secretion in lactating Holstein cows. J. Dairy Sci. 100 (4), 2660–2671. doi: 10.3168/jds.2016-11559
Kennedy P. M. (1985). Influences of cold exposure on digestion of organic matter, rates of digesta in the gastro-intestinal tract, and feeding and rumination behaviour in sheep given four forage diets in the chopped, or ground and pelleted form. Br. J. Nutr. 53 (1), 159–173. doi: 10.1079/BJN19850020
Krause K. M., Combs D. K. (2003). Effects of forage particle size, forage source, and grain fermentability on performance and ruminal pH in midlactation cows. J. Dairy Sci. 86 (4), 1382–1397. doi: 10.3168/jds.S0022-0302(03)73722-9
Krawczel P. D., Klaiber L. B., Butzler R. E., Klaiber L. M., Dann H. M., Mooney C. S., et al. (2012). Short-term increases in stocking density affect the lying and social behavior, but not the productivity, of lactating Holstein dairy cows. J. Dairy Sci. 95, 4298–4308. doi: 10.3168/jds.2011-4687
Krogstad K. C., Herrick K. J., Morris D. L., Hanford K. J., Kononoff P. J. (2021). The effects of pelleted dried distillers grains and solubles fed with different forage concentrations on rumen fermentation, feeding behavior, and milk production of lactating dairy cows. J. Dairy Sci. 104 (6), 6633–6645. doi: 10.3168/jds.2020-19592
Leso L., Becciolini V., Rossi G., Camiciottoli S., Barbari M. (2021). Validation of a commercial collar-based sensor for monitoring eating and ruminating behaviour of dairy cows. Animals 11, 2852. doi: 10.3390/ani11102852
Li Z., Cheng L., Cullen B. (2021). Validation and use of the RumiWatch noseband sensor for monitoring grazing behaviours of lactating dairy cows. Dairy 2, 104–111. doi: 10.3390/dairy2010010
Liu P., Ding L., Zhou Y., Jing X., Degen A. A. (2019). Behavioural characteristics of yaks grazing summer and winter pastures on the qinghai-Tibetan plateau. Appl. Anim. Behav. Sci. 218, 104826. doi: 10.1016/j.applanim.2019.06.007
Llonch L., Castillejos L., Ferret A. (2020). Increasing the content of physically effective fiber in high-concentrate diets fed to beef heifers affects intake, sorting behavior, time spent ruminating, and rumen pH. J. Anim. Sci. 98 (6), 1–9. doi: 10.1093/jas/skaa192
Lockard C. L., Lockard C. G., Paulus-Compart D. M., Jennings J. S. (2020). Effects of a yeast-based additive complex on performance, heat stress behaviors, and carcass characteristics of feedlot steers. Livestock Sci. 236, 104052. doi: 10.1016/j.livsci.2020.104052
Magrin L., Brscic M., Lora I., Rumor C., Tondello L., Cozzi G., et al. (2017). Effect of a ceiling fan ventilation system on finishing young bulls’ health, behaviour and growth performance. Animal 11 (6), 1084–1092. doi: 10.1017/S1751731116002482
Magrin L., Gottardo F., Fiore E., Gianesella M., Martin B., Chevaux E., et al. (2018). Use of a live yeast strain of Saccharomyces cerevisiae in a high-concentrate diet fed to finishing charolais bulls: Effects on growth, slaughter performance, behavior, and rumen environment. Anim. Feed Sci. Technol. 241, 84–93. doi: 10.1016/j.anifeedsci.2018.04.021
Marchesini G., Cortese M., Ughelini N., Ricci R., Chinello M., Contiero B., et al. (2020). Effect of total mixed ration processing time on ration consistency and beef cattle performance during the early fattening period. Anim. Feed Sci. Technol. 262, 114421. doi: 10.1016/j.anifeedsci.2020.114421
Marchesini G., Mottaran D., Contiero B., Schiavon E., Segato S., Garbin E., et al. (2018). Use of rumination and activity data as health status and performance indicators in beef cattle during the early fattening period. Vet. J. 231, 41–47. doi: 10.1016/j.tvjl.2017.11.013
Marino R., Petrera F., Speroni M., Rutigliano T., Galli A., Abeni F. (2021). Unraveling the relationship between milk yield and quality at the test day with rumination time recorded by a PLF technology. Animals 11 (5), 1583, 1–14. doi: 10.3390/ani11061583
Mendes A. M. D., Silva D. K. D., Ferreira M. D., Veras A. S., Ferreira G. D. G., Vigoderis R. B., et al. (2013). Ingestive behaviour of pastured crossbred dairy cows offered different supplement types. Trop. Anim. Health Prod. 45 (1), 231–237. doi: 10.1007/s11250-012-0195-0
Moraïs S., Mizrahi I. (2019). Islands in the stream: from individual to communal fiber degradation in the rumen ecosystem. FEMS Microbiol. Rev. 43, 362–379. doi: 10.1093/femsre/fuz007. fuz007.
Moyo M., Adekunle Adebayo R., Verla Nsahlai I. (2019). Effects of diet and roughage quality, and period of the day on diurnal feeding behaviour patterns of sheep and goats under subtropical conditions. Asian-Australas. J. Anim. Sci. 32 (5), 675–690. doi: 10.5713/ajas.17.0901
Nielsen B. L., Veerkamp R. F., Lawrence A. B. (2000). Effects of genotype, feed type and lactational stage on the time budget of dairy cows. Acta Agric. Scand. Sect. A Anim. Sci. 50 (4), 272–278. doi: 10.1080/090647000750069467
Odyuo L. T., Jana D. N., Das N. (1991). Food intake and rumination behaviour of lactating and non-lactating buffaloes reared under a loose housing system. Appl. Anim. Behav. Sci. 31 (1), 35–41. doi: 10.1016/0168-1591(91)90151-M
Oetzel G. R. (2017). Diagnosis and management of subacute ruminal acidosis in dairy herds. Vet. Clin. Food Anim. 33, 463–480. doi: 10.1016/j.cvfa.2017.06.004
Park R. M., Schubach K. M., Cooke R. F., Herring A. D., Jennings J. S., Daigle C. L. (2020). Impact of a cattle brush on feedlot steer behavior, productivity and stress physiology. Appl. Anim. Behav. Sci. 228, 104995. doi: 10.1016/j.applanim.2020.104995
Pereira G. M., Heins B. J., Endres M. I. (2018). Validation of an ear-tag accelerometer sensor to determine rumination, eating, and activity behaviors of grazing dairy cattle. J. Dairy Sci. 101 (3), 2492–2495. doi: 10.3168/jds.2016-12534
Pereira G. M., Sharpe K. T., Heins B. J. (2021). Evaluation of the RumiWatch system as a benchmark to monitor feeding and locomotion behaviors of grazing dairy cows. J. Dairy Sci. 104 (3), 3736–3750. doi: 10.3168/jds.2020-18952
Quddus R. A., Ahmad N., Khalique A., Bhatti J. A. (2022). Validation of NEDAP monitoring technology for measurements of feeding, rumination, lying, and standing behaviors, and comparison with visual observation and video recording in buffaloes. Animals 12, 578. doi: 10.3390/ani12050578
Rego G., Ferrero F., Valledor M., Campo J. C., Forcada S., Royo L. J., et al. (2020). A portable IoT NIR spectroscopic system to analyze the quality of dairy farm forage. Comput. Electron. Agric. 175, 105578. doi: 10.1016/j.compag.2020.105578
Rinne M., Huhtanen P., Jaakkola S. (2002). Digestive processes of dairy cows fed silages harvested at four stages of grass maturity. J. Anim. Sci. 80 (7), 1986–1998. doi: 10.2527/2002.8071986x
Saha S., Gallo L., Bittante G., Schiavon S., Bergamaschi M., Gianesella M., et al. (2019). A study on the effects of rumen acidity on rumination time and yield, composition, and technological properties of milk from early lactating Holstein cows. Animals 9, 66. doi: 10.3390/ani9020066
Salvati G. G. S., Santos W. P., Silveira J. M., Gritti V. C., Arthur B. A. V., Salvo P. A. R., et al. (2021). Effect of kernel processing and particle size of whole-plant corn silage with vitreous endosperm on dairy cow performance. J. Dairy Sci. 104 (2), 1794–1810. doi: 10.3168/jds.2020-19428
Schirmann K., Chapinal N., Weary D. M., Heuwieser W., von Keyserlingk M. A. G. (2012). Rumination and its relationship to feeding and lying behavior in Holstein dairy cows. J. Dairy Sci. 95 (6), 3212–3217. doi: 10.3168/jds.2011-4741
Schirmann K., von Keyserlingk M. A. G., Weary D. M., Veira D. M., Heuwieser W. (2009). Technical note: Validation of a system for monitoring rumination in dairy cows. J. Dairy Sci. 92 (12), 6052–6055. doi: 10.3168/jds.2009-2361
Sjostrom L. S., Heins B. J., Endres M. I., Moon R. D., Paulson J. C. (2016). Short communication: Relationship of activity and rumination to abundance of pest flies among organically certified cows fed 3 levels of concentrate. J. Dairy Sci. 99 (12), 9942–9948. doi: 10.3168/jds.2016-11038
Sjostrom L. S., Heins B. J., Endres M. I., Moon R. D., Sorge U. S. (2019). Effects of winter housing system on hygiene, udder health, frostbite, and rumination of dairy cows. J. Dairy Sci. 102 (11), 10606–10615. doi: 10.3168/jds.2018-15759
Soriani N., Panella G., Calamari L. (2013). Rumination time during the summer season and its relationships with metabolic conditions and milk production. J. Dairy Sci. 96 (8), 5082–5094. doi: 10.3168/jds.2013-6620
Stygar A. H., Gómez Y., Berteselli G. V., Dalla Costa E., Canali E., Niemi J. K., et al. (2021). A systematic review on commercially available and validated sensor technologies for welfare assessment of dairy cattle. Front. Vet. Sci. 8. doi: 10.3389/fvets.2021.634338
Tenffen de Sousa K., Deniz M., Martinez do Vale M., Dittrich J. R., Hötzel M. J. (2021). Influence of microclimate on dairy cows’ behavior in three pasture systems during the winter in south Brazil. J. Thermal Biol. 97, 102873. doi: 10.1016/j.jtherbio.2021.102873
Tomczak D. J., Lockard C. L., Jennings J. S., Richeson J. T. (2019). Performance, rumination, and rumen pH responses to different dietary energy density and feed management strategies in auction-derived feedlot cattle. J. Anim. Sci. 97 (11), 4682–4690. doi: 10.1093/jas/skz323
Watt L. J., Clark C. E. F., Krebs G. L., Petzel C. E., Nielsen S., Utsumi S. A. (2015). Differential rumination, intake, and enteric methane production of dairy cows in a pasture-based automatic milking system. J. Dairy Sci. 98 (10), 7248–7263. doi: 10.3168/jds.2015-9463
White R. R., Hall M. B., Firkins J. L., Kononoff P. J. (2017a). Physically adjusted neutral detergent fiber system for lactating dairy cow rations. I: Deriving equations that identify factors that influence effectiveness of fiber. J. Dairy Sci. 100 (12), 9551–9568. doi: 10.3168/jds.2017-12766
White R. R., Hall M. B., Firkins J., Kononoff P. J. (2017b). Physically adjusted neutral detergent fiber system for lactating dairy cow rations. II: Development of feeding recommendations. J. Dairy Sci. 100 (12), 9569–9584. doi: 10.3168/jds.2017-12766
Wolfger B., Timsit E., Pajor E., Cook N., Barkema H., Orsel K. (2015). Accuracy of an ear tag-attached accelerometer to monitor rumination and feeding behavior in feedlot cattle. J. Anim. Sci. 93 (6), 3164–3168. doi: 10.2527/jas2018-8802
Woolley C. E., Lachance S., DeVries T. J., Bergeron R. (2018). Behavioural and physiological responses to pest flies in pastured dairy cows treated with a natural repellent. Appl. Anim. Behav. Sci. 207, 1–7. doi: 10.1016/j.applanim.2018.07.009
Yang W. Z., Beauchemin K. A. (2007). Altering physically effective fiber intake through forage proportion and particle length: Chewing and ruminal pH. J. Dairy Sci. 90 (6), 2826–2838. doi: 10.3168/jds.2007-0032
Yang C., Tsedan G., Fan Q., Wang S., Wang Z., Chang S., et al. (2021). Behavioral patterns of yaks (Bos grunniens) grazing on alpine shrub meadows of the qinghai-Tibetan plateau. Appl. Anim. Behav. Sci. 234, 105182. doi: 10.1016/j.applanim.2020.105182
Zambelis A., Wolfe T., Vasseur E. (2019). Technical note: Validation of an ear-tag accelerometer to identify feeding and activity behaviors of tiestall-housed dairy cattle. J. Dairy Sci. 102 (5), 4536–4540. doi: 10.3168/jds.2018-15766
Zebeli Q., Aschenbach J. R., Tafaj M., Boguhn J., Ametaj B. N., Drochner W. (2012). Invited review: Role of physically effective fiber and estimation of dietary fiber adequacy in high-producing dairy cattle. J. Dairy Sci. 95 (3), 1041–1056. doi: 10.3168/jds.2011-4421
Keywords: cattle, environmental factors, feeding behavior, precision livestock farming, rumination
Citation: Abeni F (2022) Effects of extrinsic factors on some rumination patterns: A review. Front. Anim. Sci. 3:1047829. doi: 10.3389/fanim.2022.1047829
Received: 18 September 2022; Accepted: 13 October 2022;
Published: 07 November 2022.
Edited by:
Adem Kara, Erzurum Technical University, TurkeyReviewed by:
Paolo Bani, Catholic University of the Sacred Heart, ItalyEmre Sahin, Bingöl University, Turkey
Recep Gümüş, Cumhuriyet University, Turkey
Copyright © 2022 Abeni. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fabio Abeni, fabiopalmiro.abeni@crea.gov.it