 Gina Leisching
Gina Leisching Ian Wiid
Ian Wiid- SAMRC Centre for TB Research, DST/NRF Centre of Excellence for Biomedical Tuberculosis Research, Division of Molecular Biology and Human Genetics, Faculty of Medicine and Health Sciences, Stellenbosch University, Cape Town, South Africa
The type I IFN response quickly became associated with its role in the innate immune response to viral infection. The past few years have seen the significance of IFNs expand in breadth to include non-viral pathogens. Previous work has identified that following viral infection, type I IFN signaling induces the production of the 2′-5′-oligoadenylate synthetase (OAS) family, which include OAS1, OAS2, OAS3, and OAS-like (OASL) protein. OASL was identified to be strongly induced following viral infection through engaging the RNA sensor RIG-I and increasing signaling through this pathway to enhance the anti-viral type I IFN response. Surprisingly, infection with viral dsDNA revealed an IFN inhibitory role and therefore pro-viral function of OASL through the inhibition of the cGAS cytosolic DNA sensing mechanism. Intracellular bacteria are able to activate the cytosolic DNA sensing pathway, however the role of OASL during bacterial infection is largely unknown. Vacuolar pathogenic microbes such as mycobacteria induce OASL early post infection, where it functions in a prosurvival fashion by inhibiting autophagic mechanisms and antimicrobial peptide expression. This suggests an underestimated role of OASL in the innate immune response to infection with a variety of pathogens and points to OASL-associated modulation of the type I IFN response. OASL may therefore play a critical role in defining the outcome of infection. We provide a brief update on the recent developments of the OAS family of proteins in response to DNA and RNA virus infections, as well as discuss evidence of Oasl expression in response to a number of cytosolic and vacuolar replicating bacterial pathogens.
Introduction
The 2′-5′-oligoadenylate synthetases (OAS) were the first interferon (IFN)-induced antiviral proteins described that are regulated by both type I and II IFNs (Zhou et al., 1997; Eskildsen et al., 2003). Originally, type I IFNs were discovered as mediators of the host response to viral infection, however it is now well-know that type I IFN induction is central to the genetic response of both viral and bacterial pathogens (Uematsu and Akira, 2006). The OAS family of proteins consists of OAS1, OAS2, OAS3, and OAS-like (OASL) protein. The OAS proteins exhibit anti-viral functions through indirectly impeding the translation of viral nucleic acids through the activation of RNase L. OASL on the other hand, is related to the OAS family by a N-terminal OAS-like domain but lacks synthetase activity. The lack of enzymatic activity is by no means an indication of a lack of function, as recently an anti-viral role was ascribed that is underpinned by the ability of OASL to enhance of IFN I signaling (Melchjorsen et al., 2009; Zhu et al., 2014, 2015; Choi et al., 2015). On the heels of this discovery, it was observed that this antiviral function is specific to infection with RNA viruses, however when challenged with a DNA virus, OASL exerts a prosurvival function through the inhibition of IFN I signaling (Ghosh et al., 2016). Not only does this reveal a contrasting bifunctional role of OASL, it also suggests that it is able to modulate the type I IFN response when challenged with RNA and DNA pathogens. Further, this suggests the possibility that intracellular pathogens such as bacteria, once phagocytosed, induce a similar, if not an identical response when bacterial DNA enters the host cytosol. The biological role of the OAS family during microbial infection is not well documented, however recent work has revealed a dichotomous function of OASL in promoting intracellular mycobacterial survival that is closely related with stimulator of interferon genes (STING; also known as TMEM173), IFN secretion, the RNase L endocytic pathway, and autophagy. Here we provide a brief update on the recent developments of the OAS family of proteins in response to DNA and RNA virus infections, as well as discuss evidence of Oasl expression in response to a number of intracellular bacterial pathogens.
IFN/RNase L/OASL Antiviral Mechanism
The control of viral infection is essentially modulated in two parts, which upon entry of viral dsRNA, become activated and positively stimulate one another (Figure 1A). The first is the canonical OAS3/RNase L pathway which relies on the binding of OAS3 directly to dsRNA (Li et al., 2016). Previously it was thought that OAS1, 2, and 3 were involved in activating RNase L during viral infection, until recently Li and colleagues observed that OAS3 is mainly responsible for producing the 2–5A RNase L activators (Li et al., 2016). The control of infection is then achieved through 2–5A binding to RNase L, causing its dimerization and enabling its endoribonuclease activity (Tanaka et al., 2004). The second antiviral mechanism is stimulated by these cleavage products which then activate the cytoplasmic recognition receptors known as the retinoic acid-inducible gene I (RIG-1) receptor (RLR) (Yoneyama et al., 2004) and the melanoma differentiation-associated gene 5 (MDA5). The downstream signaling adaptor MAVS induces the translocation of interferon regulatory factor (IRF) 3 to the nucleus and induces the transcription of type I IFNs. The production of type I IFNs then enhances the activation of the RNase L degradative pathway (Silverman, 2007) and controls viral replication. It was observed that type I IFN signaling induces OASL through IRF3. OASL, which when present in the cytosol, binds directly to RIG-1 and mimics polyubiquitin, thereby sensitizing RIG-1 activation by viral RNA and enhancing antiviral signaling (Zhu et al., 2014). Thus during infection, OASL functions to enhance type I IFN signaling and suppress replication of RNA viruses. It has since been put forward that the anti-viral capability of OASL be harnessed with the potential for developing broad acting antiviral therapy (Zhu et al., 2015).
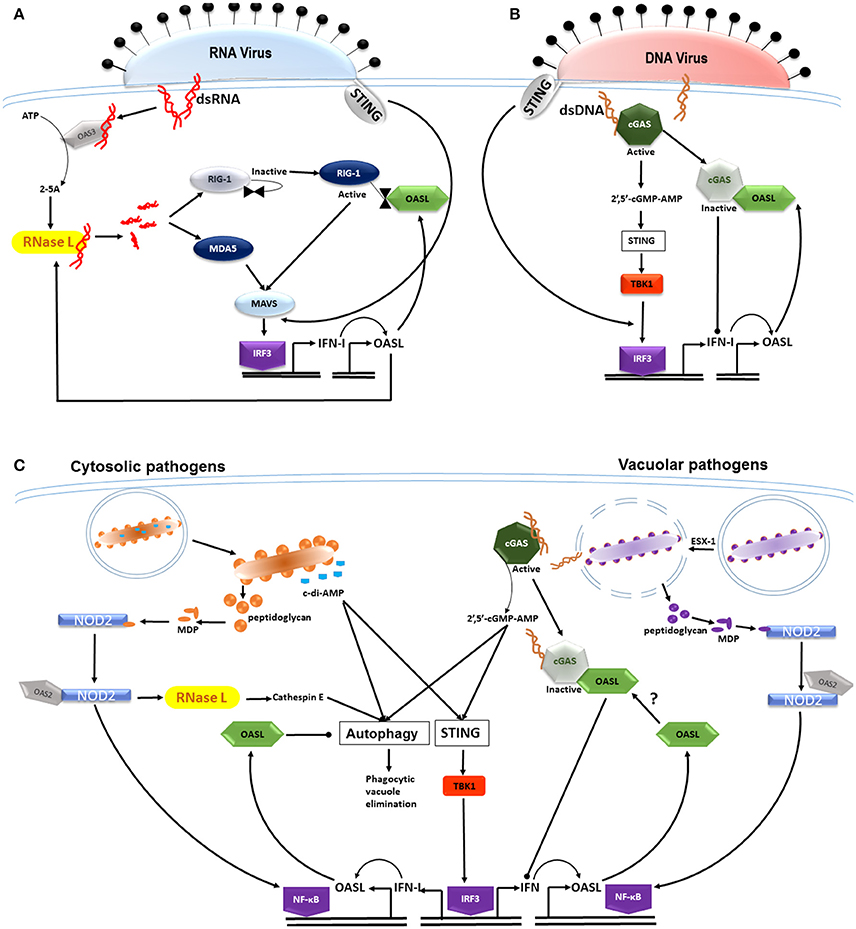
Figure 1. OASL differentially modulates type I IFN expression in response to RNA and DNA in the host cytosol. (A) Viral dsRNA binds to and activates OAS3 producing 2–5A activators of RNase L. RNase L functions to cleave viral RNA which then activate the cytoplasmic recognition receptors RIG-1 and MDA5. Subsequent signaling through the MAVS adaptor induces the translocation of interferon regulatory factor (IRF) 3 to the nucleus, resulting in the transcription of type I IFNs (STING-independent). IFN signaling induces OASL production, which when present in the cytosol, binds to and activates RIG-1 signaling, thereby enhancing IFN secretion and controlling viral replication. (B) The entry of viral dsDNA is sensed by the cytoplasmic DNA sensor cGAS, which through cGAMP induces the translocation of IRF-3 to the nucleus in a STING-independent manner. IFN induction and signaling induces OASL, which then binds to and inactivates cGAS, thereby inhibiting IFN production, allowing for the infection to persist. Although, IFN-I induction induced by both RNA and DNA viruses is STING-independent, STING is able to sense membrane fusion events at the cell membrane where it is able to induce IRF3. (C) Cyclic dinucleotides in the form of c-di-AMP are produced by diverse bacterial species and activate autophagic mechanisms as well as the STING/TBK1/IRF3 axis in a CpG-independent manner. The innate response to both cytosolic and vacuolar bacterial pathogens is enhanced through muramyl dipeptide (MDP), a breakdown product of bacterial peptidoglycan. MDP activates NOD2 which is then bound by OAS2, subsequently activating the inflammasome through NF-κB. NOD2 enhances RNase L activity which increases expression of cathepsin E- a mediator in the phagocytosis of bacteria. Vacuolar pathogens such as M. tuberculosis are able to perforate the phagosome, a process mediated by the ESX-1 secretion system. Bacterial dsDNA then gains entry into the cytosol binding to and activating cGAS, thus inducing signaling through the STING/TBK1/IRF3 axis. OASL production follows IFN signaling which inhibits autophagic mechanisms and antimicrobial peptide expression, thus promoting intracellular bacterial survival. It is unknown whether there is an association between cGAS and OASL and whether this influences type 1 IFN secretion.
cGAS/IFN/OASL Proviral Mechanism
Unlike infection with RNA viruses, much less is known about the host cells response to infection with DNA viruses. Cyclic GMP-AMP (cGAMP) synthase (cGAS) is a sensor of cytoplasmic DNA, usually of microbial origin and has recently been thoroughly reviewed (Chen et al., 2016; Patrick et al., 2016). Of interest however is the structurally homologous nature of cGAS to the OASs and the recent observation that it is able to bind directly to viral DNA(Sun et al., 2013). Once bound, 2′,5′-cGMP-AMP (cGAMP) is produced and signals through TBK1 and STING, inducing type I IFN production through the activation of IRF3 (Figure 1B). As with RNA virus infections, the presence of IFN signaling then rapidly induces OASL which binds to, and deactivates cGAS (Ghosh et al., 2016). The deactivation of cGAS consequently inhibits IFN production which then allows for the persistence of the DNA virus infection. It is accepted that signaling through cGAS during the initial stages of infection is accompanied by the downstream activation of STING where it plays an essential role in type I interferon-dependant innate immunity in response to intracellular DNA (Ishikawa et al., 2009). However, it is suspected that cGAS-dependant IFN induction by DNA viruses such as Vaccinia and HSV is independent of STING stimulation but this is yet to be confirmed (Ghosh et al., 2016). It is however important to note that STING is able to sense membrane-fusion events that are associated with viral entry into the host cell that is independent of nucleic acid sensing, but is still able to induce type I IFNs (Holm et al., 2012). Moreover, STING is unlikely to be required for IFN induction in response to RNA or RNA viruses presumably because viruses can activate IRF3 through RIG-1 and its downstream signaling adaptor MAVS (Chiu et al., 2009). This points to the possibility of a virus-specific response whereby STING-dependent type 1 IFN production is bypassed.
cGAS/STING/IFN/OASL Probacterial Mechanism
Phagocytic cells are not only able to discriminate between viral and bacterial infection, but also extracellular microbes from intracellular replicating pathogens. Pattern recognition receptors (PRRs) are central to distinguishing these signals and sensing conserved motifs presented by microbes (Gordon, 2002; Meylan et al., 2006; Kawai and Akira, 2010). The membrane-bound Toll-like receptors (TLRs) monitor extracellular activity and phagolysosomal compartments by recognizing pathogen associated molecular patterns (PAMPs) that include flagella, bacterial lipoprotein, lipopolysaccharide (LPS), and CpG DNA (Akira et al., 2006) and are responsible for inducing proinflammatory genes. The soluble, cytosolic NOD-like receptors (NLRs) recognize cell wall fragments from both Gram-negative and Gram-positive bacteria (Hasegawa et al., 2006; Shaw et al., 2008), as well as acid-fast mycobacteria (Jo, 2008) that activate the inflammasome (Figure 1C). Muramyl dipeptide (MDP), a peptidoglycan break down product, is one such cell wall fragment which binds to and activates NOD2 (Girardin et al., 2003). It was observed that OAS2 binds NOD2 and enhances RNase L activity (Dugan et al., 2009) which increases expression of cathepsin E, an important mediator of autophagy following the phagocytosis of bacteria (Tsukuba et al., 2013). Additionally, the activation of NOD2 leads to the activation of the NF-κB transcription factor which is associated with the release of cytokines and control of the infection. Once phagocytosed, most pathogenic intracellular bacteria are able to mediate the phagosome breach, which in the process leads to content leakage in the form of dsDNA into the host cytosol(Vance et al., 2009; Manzanillo et al., 2012). Vacuolar pathogens such as Mycobacterium tuberculosis perforate the phagosome, a process mediated by the ESX-1 secretion system(Manzanillo et al., 2012). In this way, extracellular mycobacterial DNA is able to enter the host cytosol, inducing IFN-β signaling through the Sting/Tbk1/Irf3 axis. Bacterial pathogens such as Listeria monocytogenes that replicate in the cytoplasm induce IFN-β transcription as part of the cytosolic surveillance pathway (CSP) (Henry and Monack, 2007) which induces signaling through the very same axis. Thus the inflammatory response induced by infection with cytosolic or vacuolar pathogens converge through this signaling axis. The signaling events downstream of this have however remained unclear until recently.
As with viral dsDNA, it was observed that the entry of foreign bacterial dsDNA is met with the DNA sensor cGAS, which then signals through cGAMP, activating the Sting/Tbk1/Irf3 axis which will culminate in type I IFN secretion. It is now known that IFN-α IFN-β secretion in response to bacterial infection is in fact a pathogenic mechanism that enhances bacterial replication (Stanley et al., 2007). Moreover, OASL is also produced as a result of signaling through this axis and aids in promoting a favorable intracellular milieu for bacterial survival. In line with this, the biological role of OASL during mycobacterial infection, specifically with the vacuolar pathogen M. leprae was recently elucidated (de Toledo-Pinto et al., 2016). The authors suggest that the presence of OASL is required for intracellular M. leprae survival which appears to be dependent on inhibiting autophagic mechanisms, thus preventing clearance of the microbe. Since this is the first study observing the OASL-(vacuolar) microbe link, the next question that arises is whether OASL is relevant during infection with other vacuolar pathogens and those which replicate in the cytoplasm. A search of the literature revealed the upregulation of OASL or the murine equivalent Oasl2 through transcriptome analysis (Table 1) in a number of infection models. Its upregulation has been observed in response to a number of Gram-positive and Gram-negative bacteria from as early as 4 hpi. These pathogens include Chlamydia trachomatis (Lad et al., 2005), Chlamydia pneumonia (Eickhoff et al., 2007), L. monocytogenes (Kutsch et al., 2008), Brucella abortus (Roux et al., 2007), and Lactobacillus acidophilus (Weiss et al., 2010). Although, this search was not exhaustive, it suggests that OASL more than likely plays a role during infection with a wide range of bacterial species. What needs to be determined is whether OASL binds to and inactivates cGAS, thus inhibiting STING-dependant type I IFN production. This is what is observed during infection with HSV and Vaccinia DNA viruses (Ghosh et al., 2016), however an identical response cannot be assumed since it was determined that OASL knock-down did not influence IFN-β release during M. leprae infection (de Toledo-Pinto et al., 2016).
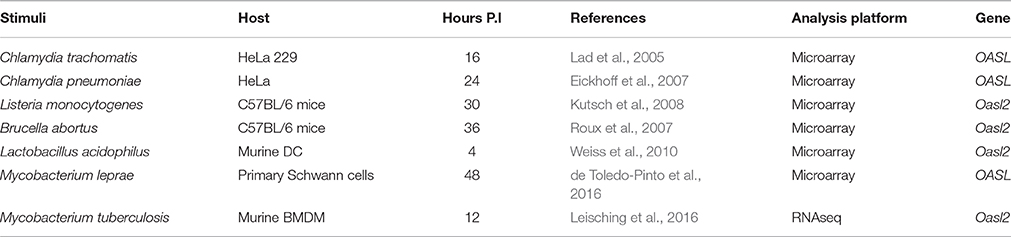
Table 1. Vacuolar and cytosolic bacterial pathogens that induce OASL/Oasl2.
The balance between type I and type II IFNs is particularly important for controlling intracellular bacterial infections, notably M. tuberculosis, M. leprae, and L. monocytogenes, and since OASL plays a significant role in modulating type I IFN production is can be said that the expression of this protein may be central to controlling viral and bacterial infections. Collectively, the above evidence clearly points to an underestimated role of OASL in the innate immune response to infection with a variety of pathogens. Not only is OASL expression closely related to the secretion of type I IFNs, its role is clearly diverse with it interacting closely with the RIG-I pathway, autophagy, and DNA sensing through cGAS.
Author Contributions
GL wrote the manuscript. IW and BB provided substantial contributions to the framework of the manuscript, as well as revision of the work, and approval of the final draft.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by the Department of Science and Technology/National Research 480 Foundation (DST/NRF) and the South African Medical Research Council (SAMRC).
References
Akira, S., Uematsu, S., and Takeuchi, O. (2006). Pathogen recognition and innate immunity. Cell 124, 783–801. doi: 10.1016/j.cell.2006.02.015
Chen, Q., Sun, L., and Chen, Z. J. (2016). Regulation and function of the cGAS-STING pathway of cytosolic DNA sensing. Nat. Immunol. 17, 1142–1149. doi: 10.1038/ni.3558
Chiu, Y.-H., MacMillan, J. B., and Chen, Z. J. (2009). RNA polymerase III detects cytosolic DNA and induces type I interferons through the RIG-I pathway. Cell 138, 576–591. doi: 10.1016/j.cell.2009.06.015
Choi, U. Y., Kang, J.-S., Hwang, Y. S., and Kim, Y.-J. (2015). Oligoadenylate synthase-like (OASL) proteins: dual functions and associations with diseases. Exp. Mol. Med. 47, e144. doi: 10.1038/emm.2014.110
de Toledo-Pinto, T. G., Ferreira, A. B., Ribeiro-Alves, M., Rodrigues, L. S., Batista-Silva, L. R., and Ozorio Moraes, M. (2016). STING-dependent 2′-5′ oligoadenylate synthetase-like production is required for intracellular mycobacterium leprae survival. J. Infect. Dis. 214, 311–320. doi: 10.1093/infdis/jiw144
Dugan, J. W., Albor, A., David, L., Fowlkes, J., Blackledge, M. T., and Davey, M. P. (2009). Nucleotide oligomerization domain-2 interacts with 2′-5′-oligoadenylate synthetase type 2 and enhances RNase-L function in THP-1 cells. Mol. Immunol. 47, 560–566. doi: 10.1016/j.molimm.2009.09.025
Eickhoff, M., Thalmann, J., Hess, S., Martin, M., Laue, T., Kruppa, J., et al. (2007). Host cell responses to Chlamydia pneumoniae in gamma interferon-induced persistence overlap those of productive infection and are linked to genes involved in apoptosis, cell cycle, and metabolism. Infect. Immun. 75, 2853–2863. doi: 10.1128/IAI.01045-06
Eskildsen, S., Justesen, J., Schierup, M. H., and Hartmann, R. (2003). Characterization of the 2′–5′-oligoadenylate synthetase ubiquitin-like family. Nucleic Acids Res. 31, 3166–3173. doi: 10.1093/nar/gkg427
Ghosh, A., Sampath, P., Zhu, J., Hornung, V., Thorne, S., and Sarkar, S. N. (2016). Modulation of cellular immune response by 2′-5′ Oligoadenylate Synthetase - like (OASL) proteins during DNA virus infection. J. Immunol. 196(Suppl. 1), 61.6.
Girardin, S. E., Boneca, I. G., Viala, J., Chamaillard, M., Labigne, A., and Sansonetti, P. J. (2003). Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J. Biol. Chem. 278, 8869–8872. doi: 10.1074/jbc.C200651200
Gordon, S. (2002). Pattern recognition receptors: doubling up for the innate immune response. Cell 111, 927–930. doi: 10.1016/S0092-8674(02)01201-1
Hasegawa, M., Yang, K., Hashimoto, M., Park, J.-H., Kim, Y.-G., Fujimoto, Y., et al. (2006). Differential release and distribution of Nod1 and Nod2 immunostimulatory molecules among bacterial species and environments. J. Biol. Chem. 281, 29054–29063. doi: 10.1074/jbc.M602638200
Henry, T., and Monack, D. M. (2007). Activation of the inflammasome upon Francisella tularensis infection: interplay of innate immune pathways and virulence factors. Cell. Microbiol. 9, 2543–2551. doi: 10.1111/j.1462-5822.2007.01022.x
Holm, C. K., Jensen, S. B., Jakobsen, M. R., Cheshenko, N., Horan, K. A., and Yarovinsky, T. O. (2012). Virus-cell fusion as a trigger of innate immunity dependent on the adaptor STING. Nat. Immunol. 13, 737–743. doi: 10.1038/ni.2350
Ishikawa, H., Ma, Z., and Barber, G. N. (2009). STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 461, 788–792. doi: 10.1038/nature08476
Jo, E.-K. (2008). Mycobacterial interaction with innate receptors: TLRs, C-type lectins, and NLRs. Curr. Opin. Infect. Dis. 21, 279–286. doi: 10.1097/QCO.0b013e3282f88b5d
Kawai, T., and Akira, S. (2010). The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat. Immunol. 11, 373–384. doi: 10.1038/ni.1863
Kutsch, S., Degrandi, D., and Pfeffer, K. (2008). Immediate lymphotoxin β receptor-mediated transcriptional response in host defense against L. monocytogenes. Immunobiology 213, 353–366. doi: 10.1016/j.imbio.2007.10.011
Lad, S. P., Fukuda, E. Y., Li, J., Luis, M., and Li, E. (2005). Up-regulation of the JAK/STAT1 signal pathway during Chlamydia trachomatis infection. J. Immunol. 174, 7186–7193. doi: 10.4049/jimmunol.174.11.7186
Leisching, G., Pietersen, R.-D., van Heerden, C., van Helden, P., Wiid, I., and Baker, B. (2016). RNAseq reveals hypervirulence-specific host responses to M. tuberculosis infection. Virulence. doi: 10.1080/21505594.2016.1250994.[Epub ahead of print].
Li, Y., Banerjee, S., Wang, Y., Goldstein, S. A., Dong, B., and Weiss, S. R. (2016). Activation of RNase L is dependent on OAS3 expression during infection with diverse human viruses. Proc. Natl. Acad. Sci. U.S.A. 113, 2241–2246. doi: 10.1073/pnas.1519657113
Manzanillo, P. S., Shiloh, M. U., Portnoy, D. A., and Cox, J. S. (2012). Mycobacterium tuberculosis activates the DNA-dependent cytosolic surveillance pathway within macrophages. Cell Host Microbe 11, 469–480. doi: 10.1016/j.chom.2012.03.007
Melchjorsen, J., Kristiansen, H., Christiansen, R., Rintahaka, J., Matikainen, S., Paludan, S., et al. (2009). Differential regulation of the OASL and OAS1 genes in response to viral infections. J. Interferon Cytokine Res. 29, 199–208. doi: 10.1089/jir.2008.0050
Meylan, E., Tschopp, J., and Karin, M. (2006). Intracellular pattern recognition receptors in the host response. Nature 442, 39–44. doi: 10.1038/nature04946
Patrick, K. L., Bell, S. L., and Watson, R. O. (2016). For better or worse: cytosolic DNA sensing during intracellular bacterial infection induces potent innate immune responses. J. Mol. Biol. 428, 3372–3386 doi: 10.1016/j.jmb.2016.04.030
Roux, C. M., Rolán, H. G., Santos, R. L., Beremand, P. D., Thomas, T. L., and Tsolis, R. M. (2007). Brucella requires a functional Type IV secretion system to elicit innate immune responses in mice. Cell. Microbiol. 9, 1851–1869. doi: 10.1111/j.1462-5822.2007.00922.x
Shaw, M. H., Reimer, T., Kim, Y.-G., and Nuñez, G. (2008). NOD-like receptors (NLRs): bona fide intracellular microbial sensors. Curr. Opin. Immunol. 20, 377–382. doi: 10.1016/j.coi.2008.06.001
Silverman, R. H. (2007). Viral encounters with 2′, 5′-oligoadenylate synthetase and RNase L during the interferon antiviral response. J. Virol. 81, 12720–12729. doi: 10.1128/JVI.01471-07
Stanley, S. A., Johndrow, J. E., Manzanillo, P., and Cox, J. S. (2007). The type I IFN response to infection with Mycobacterium tuberculosis requires ESX-1-mediated secretion and contributes to pathogenesis. J. Immunol. 178, 3143–3152. doi: 10.4049/jimmunol.178.5.3143
Sun, L., Wu, J., Du, F., Chen, X., and Chen, Z. J. (2013). Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339, 786–791. doi: 10.1126/science.1232458
Tanaka, N., Nakanishi, M., Kusakabe, Y., Goto, Y., Kitade, Y., and Nakamura, K. T. (2004). Structural basis for recognition of 2′,5′-linked oligoadenylates by human ribonuclease L. EMBO J. 23, 3929–3938. doi: 10.1038/sj.emboj.7600420
Tsukuba, T., Yanagawa, M., Kadowaki, T., Takii, R., Okamoto, Y., Sakai, E., et al. (2013). Cathepsin E deficiency impairs autophagic proteolysis in macrophages. PLoS ONE 8:e82415. doi: 10.1371/journal.pone.0082415
Uematsu, S., and Akira, S. (2006). Toll-like receptors and innate immunity. J. Mol. Med. 84, 712–725. doi: 10.1007/s00109-006-0084-y
Vance, R. E., Isberg, R. R., and Portnoy, D. A. (2009). Patterns of pathogenesis: discrimination of pathogenic and nonpathogenic microbes by the innate immune system. Cell Host Microbe 6, 10–21. doi: 10.1016/j.chom.2009.06.007
Weiss, G., Rasmussen, S., Zeuthen, L. H., Nielsen, B. N., Jarmer, H., and FrøkiÃČÂęr, H. (2010). Lactobacillus acidophilus induces virus immune defence genes in murine dendritic cells by a Toll-like receptor-2-dependent mechanism. Immunology 131, 268–281. doi: 10.1111/j.1365-2567.2010.03301.x
Yoneyama, M., Kikuchi, M., Natsukawa, T., Shinobu, N., Imaizumi, T., Miyagishi, M., et al. (2004). The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 5, 730–737. doi: 10.1038/ni1087
Zhou, A., Paranjape, J., Brown, T. L., Nie, H., Naik, S., and Colmenares, C. (1997). Interferon action and apoptosis are defective in mice devoid of 2′,5′-oligoadenylate-dependent RNase L. EMBO J. 16, 6355–6363. doi: 10.1093/emboj/16.21.6355
Zhu, J., Ghosh, A., and Sarkar, S. N. (2015). OASL—a new player in controlling antiviral innate immunity. Curr. Opin. Virol. 12, 15–19. doi: 10.1016/j.coviro.2015.01.010
Keywords: OASL, OAS, cGAS, STING, Type I IFN, mycobacteria, pathogenesis
Citation: Leisching G, Wiid I and Baker B (2017) The Association of OASL and Type I Interferons in the Pathogenesis and Survival of Intracellular Replicating Bacterial Species. Front. Cell. Infect. Microbiol. 7:196. doi: 10.3389/fcimb.2017.00196
Received: 09 December 2016; Accepted: 04 May 2017;
Published: 19 May 2017.
Edited by:
Yousef Abu Kwaik, University of Louisville, United StatesReviewed by:
Stephanie M. Seveau, Ohio State University, United StatesAlan G. Goodman, Washington State University, United States
Copyright © 2017 Leisching, Wiid and Baker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gina Leisching, Z2luYWxAc3VuLmFjLnph