Gut Microbiota Dysbiosis in BK Polyomavirus-Infected Renal Transplant Recipients: A Case-Control Study
 Jian Zhang
Jian Zhang Hao Qin1
Hao Qin1  Mingyu Chang
Mingyu Chang- 1Department of Urology, Beijing Friendship Hospital, Capital Medical University, Beijing, China
- 2Beijing Key Laboratory of Tolerance Induction and Organ Protection in Transplantation, Beijing, China
Background: BK polyomavirus infection results in renal allograft dysfunction, and it is important to find methods of prediction and treatment. As a regulator of host immunity, changes in the gut microbiota are associated with a variety of infections. However, the correlation between microbiota dysbiosis and posttransplant BK polyomavirus infection was rarely studied. Thus, this study aimed to characterize the gut microbiota in BK polyomavirus-infected renal transplant recipients in order to explore the biomarkers that might be potential therapeutic targets and establish a prediction model for posttransplant BK polyomavirus infection based on the gut microbiota.
Methods: We compared the gut microbial communities of 25 BK polyomavirus-infected renal transplant recipients with 23 characteristic-matched controls, applying the 16S ribosomal RNA gene amplicon sequencing technique.
Results: At the phylum level, Firmicutes/Bacteroidetes ratio significantly increased in the BK polyomavirus group. Bacteroidetes was positively correlated with CD4/CD8 ratio. In the top 20 dominant genera, Romboutsia and Roseburia exhibited a significant difference between the two groups. No significant difference was observed in microbial alpha diversity. Beta diversity revealed a significant difference between the two groups. Nine distinguishing bacterial taxa were discovered between the two groups. We established a random forest model using genus taxa to predict BK polyomavirus infectious status, which achieved the best accuracy (80.71%) with an area under the curve of 0.82. Two genera were included in the best model, which were Romboutsia and Actinomyces.
Conclusions: BK polyomavirus-infected patients had gut microbiota dysbiosis in which the Firmicutes/Bacteroidetes ratio increased in the course of the viral infection. Nine distinguishing bacterial taxa might be potential biomarkers of BK polyomavirus infection. The random forest model achieved an accuracy of 80.71% in predicting the BKV infectious status, with Romboutsia and Actinomyces included.
Introduction
BK polyomavirus (BKV) infection is one of the most common but intractable complications following renal transplantation, which can result in renal allograft dysfunction and graft loss (Hariharan, 2006; Yi et al., 2017). Due to the existence of a buffering period from the onset of asymptomatic BKV infection to BKV-associated nephropathy, early determination of BKV infection and subsequent intervention are important for preventing the progression to BKV-associated nephropathy. Nowadays, detection of BKV almost depends on polymerase chain reaction (PCR), which is four times more sensitive than urine cytology for monitoring asymptomatic viruria, nonetheless lacking other predictive methods (Dalianis et al., 2019; Zakaria et al., 2019). Moreover, there have been no direct antiviral agents for BKV infection. Progressive reduction of immunosuppression according to the BKV viral load in the urine/blood samples or allograft biopsy results is the accepted therapeutic regimen (Hariharan, 2006; Dalianis et al., 2019; Myint et al., 2022). However, that may risk acute rejection, which challenges the treatment of BKV infection and leads to worse allograft survival (Baek et al., 2018). Thus, exploration of a new predictive method and treatment without the reduction of immunosuppression is needed.
Both innate immunity and adaptive immunity have been elucidated to play vital roles in controlling BKV infection (Ambalathingal et al., 2017). Recently, the gut microbiota, which is a complex ecosystem, has been demonstrated to be an important regulator of host immunity. Evidence suggests that gut microbial metabolites contribute to the development of both T-regulatory cells (Tregs) and the anti-inflammatory immune state. Especially the short-chain fatty acids, which are fermentation products of dietary fiber and carbohydrates by gut bacteria (Cummings, 1983), promote the conversion of naive CD4+ T lymphocytes to Tregs, contributing to immune suppression (Arpaia et al., 2013; Haase et al., 2018). Meanwhile, the metabolites can trigger immune responses against pathogens by inducing the secretion of pro-inflammatory cytokines (Kalina et al., 2002; Hosseinkhani et al., 2021). In brief, gut microbial metabolites are integral to immune homeostasis. Therefore, the dysbiosis of the gut microbiota may induce immune deficiency and subsequent infections by microbial metabolites. Several studies have demonstrated the correlation between gut microbiota dysbiosis and infections (Chan et al., 2019; Lee et al., 2019; Campisciano et al., 2020). In addition, bidirectional interactions between infections and the gut microbiota are reported that viral infections can also change the gut microbiota (Hanada et al., 2018). To date, the correlation between gut microbiota dysbiosis and posttransplant BKV infection is still unknown. Thus, this study aimed to characterize the gut microbiota in BKV-infected renal transplant recipients, compared with the controls, in order to explore the biomarkers that might be potential therapeutic targets and establish a prediction model for posttransplant BKV infection based on the gut microbiota.
Materials and Methods
Patient Cohort
Patients who received a first renal transplant within 2 years and received regular surveillance for BKV were included in the study. Exclusion criteria included the following: 1) multiorgan or pediatric transplants; 2) lacking BKV detection; 3) taking high-dose antibiotics; 4) delayed graft function early after renal transplantation; 5) concurrent human immunodeficiency virus, hepatitis virus, or Mycobacterium tuberculosis infection; 6) obese patients. BKV viral load was detected by PCR in all participants. All of them received low-dose sulfamethoxazole/trimethoprim and ganciclovir for prophylaxis after transplantation. Information about posttransplant infections, rejection, and medications was collected. A total of 48 recipients were enrolled and divided into the BKV group (n = 25) and the control group (n = 23), with a ratio of proximately 1/1. Fecal samples were collected after the diagnosis of BKV infection. The study was approved by the ethics committee of Beijing Friendship Hospital (2020-P2-212-01).
Sample Collection
Fecal samples were collected by Faeces container (Sarstedt, Germany) without preserving reagent and stored at -80°C, and processed within 2 weeks after collection.
Extraction of Genome DNA
Genome DNA was extracted from the samples using cetyltrimethylammonium bromide/sodium dodecyl sulfate (CTAB/SDS) method. DNA concentration and purity were monitored on 1% agarose gels. Extracted DNA was diluted to a concentration of 1 ng/μl using sterile water.
Amplicon Generation
In this study, 16S ribosomal RNA (rRNA) genes were amplified using the specific primer Bakt_341F (5′- CCTACGGGNGGCWGCAG-3′) and Bakt_805R (5′-GACTACHVGGGTATCTAATCC-3′) (Herlemann et al., 2011). PCR reactions were performed in 30-μl reactions, containing 15 μl of Phusion® High-Fidelity PCR Master Mix (New England Biolabs), 0.2 μl of forward and reverse primers, and 10 ng of template DNA. The thermal cycling protocol included initial denaturation at 95°C for 3 min, 25 cycles of denaturation at 95°C for 30 s, annealing at 55°C for 30 s, elongation at 72°C for 30 s, and finally 16°C for 2 min.
PCR Product Quantification and Qualification
PCR products were mixed with an equal volume of 1× loading buffer (containing SYB green). Electrophoresis was performed on 2% agarose gel. Samples with a bright main strip around 460 bp (V3+V4) were chosen for further experiments (Vasileiadis et al., 2012).
PCR Product Mixing and Purification
Since we mixed the PCR products in equi-density ratios, the mixture was purified using GeneJET Gel Extraction Kit (Thermo Scientific).
Library Preparation and Sequencing
NEB Next Ultra DNA Library Prep Kit for Illumina (NEB, USA) was used to generate sequencing libraries according to recommendations from the manufacturer, and index codes were added. The library quality was assessed on the Qubit@ 2.0 Fluorometer (Life Technologies, CA, USA) and Agilent Bioanalyzer 2100 system. Finally, libraries were sequenced on Illumina MiSeq platform, and 250-bp paired-end reads were generated.
Viral Infection Monitoring and Definition
The BKV infection status of each subject was confirmed by electronic medical records. Detection of BKV was performed by PCR (McNees et al., 2005). DNA was extracted from 200 µl of urine sample using the viral DNA/RNA extraction and purification kit (Xi’an Tianlong Science and Technology, Xi`an, China) according to the manufacturer’s instructions. PCR was performed using the ABI 7500 FAST Real-Time PCR System following the manufacturer’s recommendations and PCR fluorescence probe method with BKV nucleic acid detection kit (Beijing SinoMDgene Technology, Beijing, China). PCR reactions were performed in 25-μl reactions, containing 20-µl master mix and 5-µl template DNA. The thermal cycling protocol included initial denaturation at 95°C for 3 min, 40 cycles of denaturation at 94°C for 15 s, annealing, elongation, and signal acquisition at 60°C for 35 s, and finally 25°C for 1 min. BK viruria was defined as a positive result above the detectable level that the lower limit was 2.0E+03 copies/ml.
Disease Prediction Model
We used the random Forest (version 4.6) package of R language to build the random forest model. The sequential forward selection method was used to select the best feature set. We started with the best feature with the largest classification accurate for feature selection, and then the other feature was added one by one. Each time the model classification accuracy was evaluated, the feature with the highest accuracy was added to the model. This process was repeated until the highest accuracy for the model was achieved. The 5-fold cross-validation was used to split the data into training and test datasets in model evaluation, and the cross-validation was repeated 20 times.
Statistical Analysis
Alpha diversity and beta diversity on Bray–Curtis were measured using default parameters and QIIME2 tools. Principal coordinate analysis (PCoA) was generated to reveal the divergence between groups, and analysis of similarities (ANOSIM) was applied to test the significance of the clustering based on a distance matrix of Bray–Curtis. Linear discriminant analysis (LDA) Effect Size (LEfSe) was used to identify the distinguishing bacterial taxa between groups, limiting the log LDA score >4.0.
We used SPSS (ver. 26.0, SPSS Inc., Chicago, IL, USA) for statistical analysis. Measurement data were expressed as mean [standard deviation (SD)]. Student’s t-test and chi-square test were applied to compare the quantitative variables and the categorical variables between groups, respectively. Wilcoxon rank-sum test was used to compare the abundance of microbial species between groups. Spearman correlation analysis was applied to explore the correlation between the microbiota and the CD4/CD8 ratio. A p < 0.05 was considered to be statistically significant.
Results
Clinical Characteristics of Participants in the Study
The clinical characteristics of the BKV group and the control group were displayed in Table 1. There was no significant difference in gender, age, body mass index, concurrent diabetes, medications, concurrent infections, or clinical rejection between the two groups. The BKV viral load in urine was 2.35E+10 ± 9.98E+10 copies/ml in the BKV group. The CD4/CD8 ratio was 1.18 ± 0.71 in the BKV group, lower than that in the control group (p = 0.012).
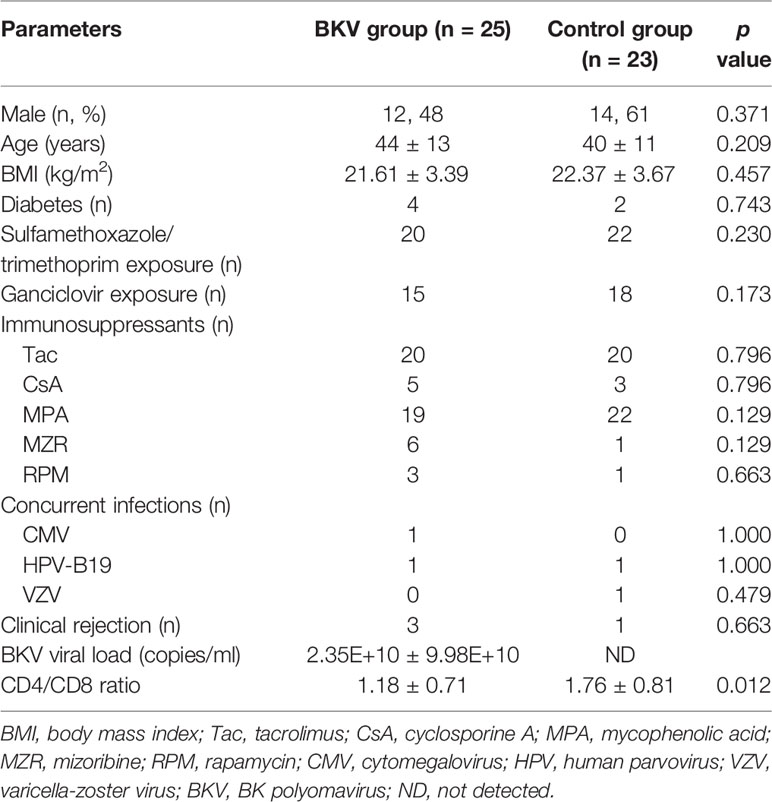
Table 1 The clinical characteristics of the BKV group and the control group.
Sample Processing and Sequencing Results
A total of 48 fecal samples were processed for 16S rRNA gene amplicon sequencing. On average, 69,683 ± 6,297 sequence reads were generated per sample in the BKV group, and 68,779 ± 6,696 sequence reads were generated per sample in the control group. There was no significant difference between the two groups (p = 0.632).
Microbial Community Structure
On average, the samples in the BKV group were characterized by comparable operational taxonomic unit (OTU) counts (mean: 218 ± 61 OTUs) compared with those in the control group (185 ± 70 OTUs), p = 0.083. There were a total of 390 shared OTUs between the two groups and 9 unique OTUs in the BKV group.
At the phylum level (Figure 1A), the abundance of Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria, and Verrucomicrobia accounted for over 99% in both groups. Firmicutes, Actinobacteria, and Verrucomicrobia were richer in the BKV group than those in the control group (63.92% vs. 58.32%, 5.08% vs. 2.69%, 3.59% vs. 2.30%), but there was no statistically significant difference (p = 0.458, p = 0.214, p = 0.469, respectively). Bacteroidetes and Proteobacteria were poorer in the BKV group than those in the control group (12.04% vs. 21.52%, 14.58% vs. 14.85%), but there was no statistically significant difference (p = 0.225 and p = 0.360, respectively). However, the Firmicutes/Bacteroidetes ratio was significantly higher in the BKV group than that in the control group (133.74 ± 306.79 vs. 26.65 ± 84.96, p = 0.046) (Figure 1C). The Spearman correlation analysis showed that Bacteroidetes was positively correlated with the CD4/CD8 ratio (r = 0.289, p = 0.046) (Figure 1D). In the top 20 dominant genera (Figure 1B), Romboutsia (2.26% in the BKV group vs. 0.16% in the control group, p = 0.022) and Roseburia (1.84% in the BKV group vs. 0.02% in the control group, p = 0.026) exhibited a significant difference between the two groups.
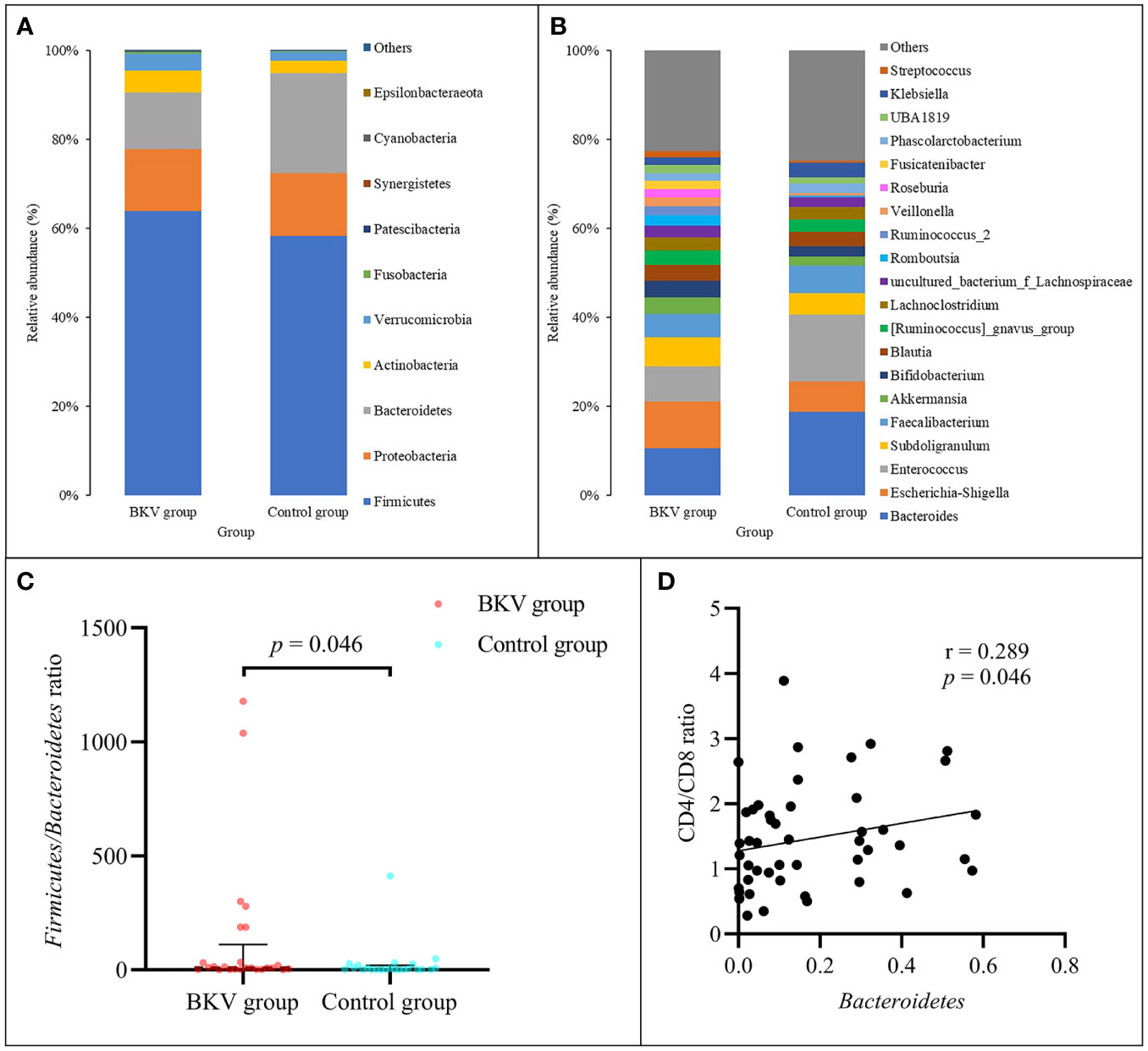
Figure 1 Abundance of bacterial taxa between the BKV group and the control group. (A) Top 10 dominant phyla. (B) Top 20 dominant genera. (C) The Firmicutes/Bacteroidetes ratio was significantly higher in the BKV group than that in the control group. (D) Spearman correlation analysis showed that Bacteroidetes was positively correlated with the CD4/CD8 ratio.
Analysis of Microbial Diversity
As shown in Table 2 and Figure 2, no significant difference was observed in the microbial alpha diversity.

Table 2 Microbial diversity in the BKV group and the control group.
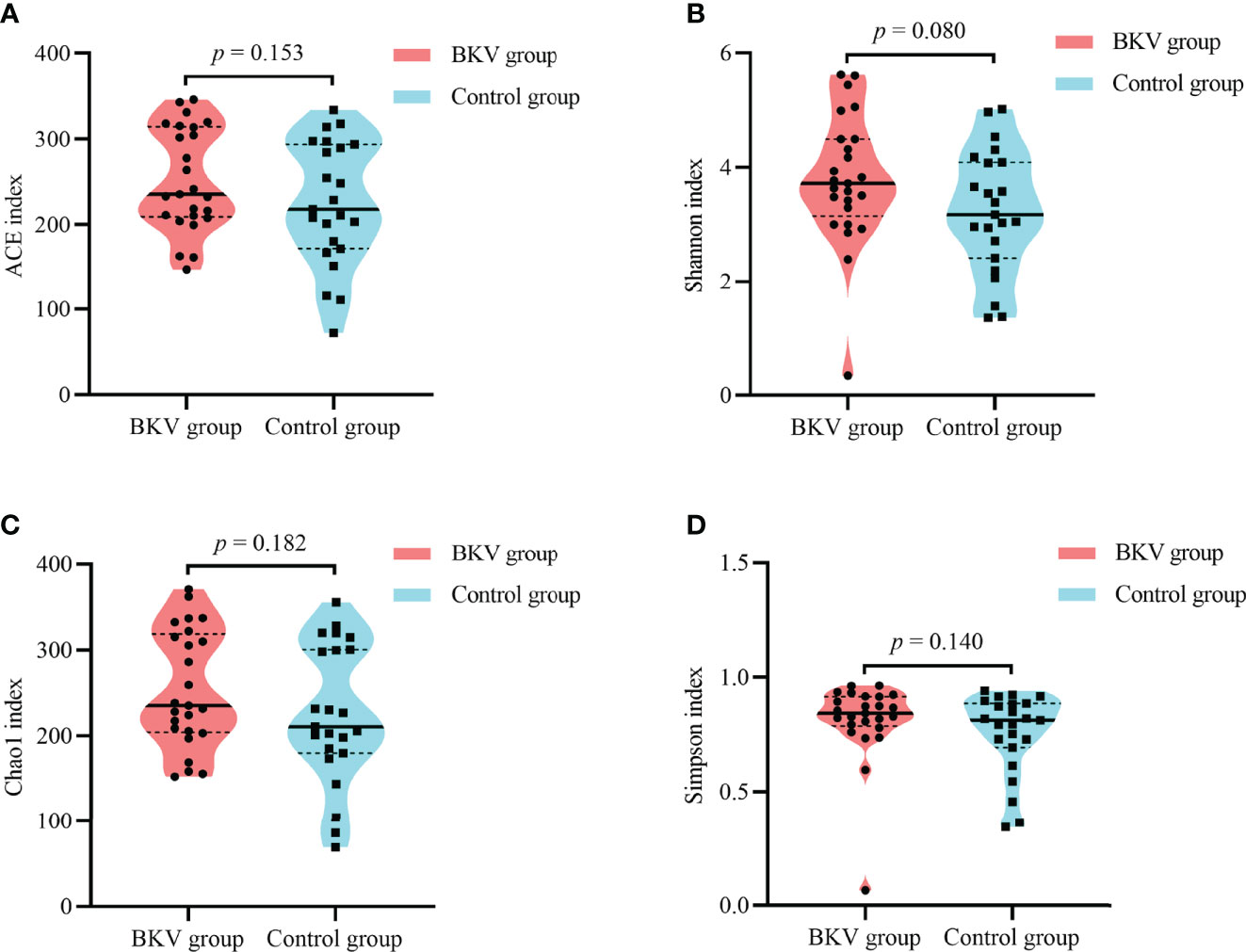
Figure 2 Alpha diversity indices between the BKV group and the control group. (A) ACE index. (B) Shannon index. (C) Chao1 index. (D) Simpson index. No significant difference was observed in microbial diversity. ACE, abundance-based coverage estimator.
Clustering of Microbial Community
The PCoA plot was applied to evaluate the beta diversity based on Bray–Curtis distance analysis, which showed that the microbiota in the BKV group and the control group were clustered closely with some overlap (Figure 3A). The first principal component (PC1) and the second principal component (PC2) accounted for 14.64% and 12.12% of total variations, respectively. ANOSIM at the OTU level was conducted (Figure 3B). The R value was 0.059, while the p value was 0.028. The difference in microbiota in the inter-group was greater than that in the intra-group, indicating that the grouping was significantly meaningful.
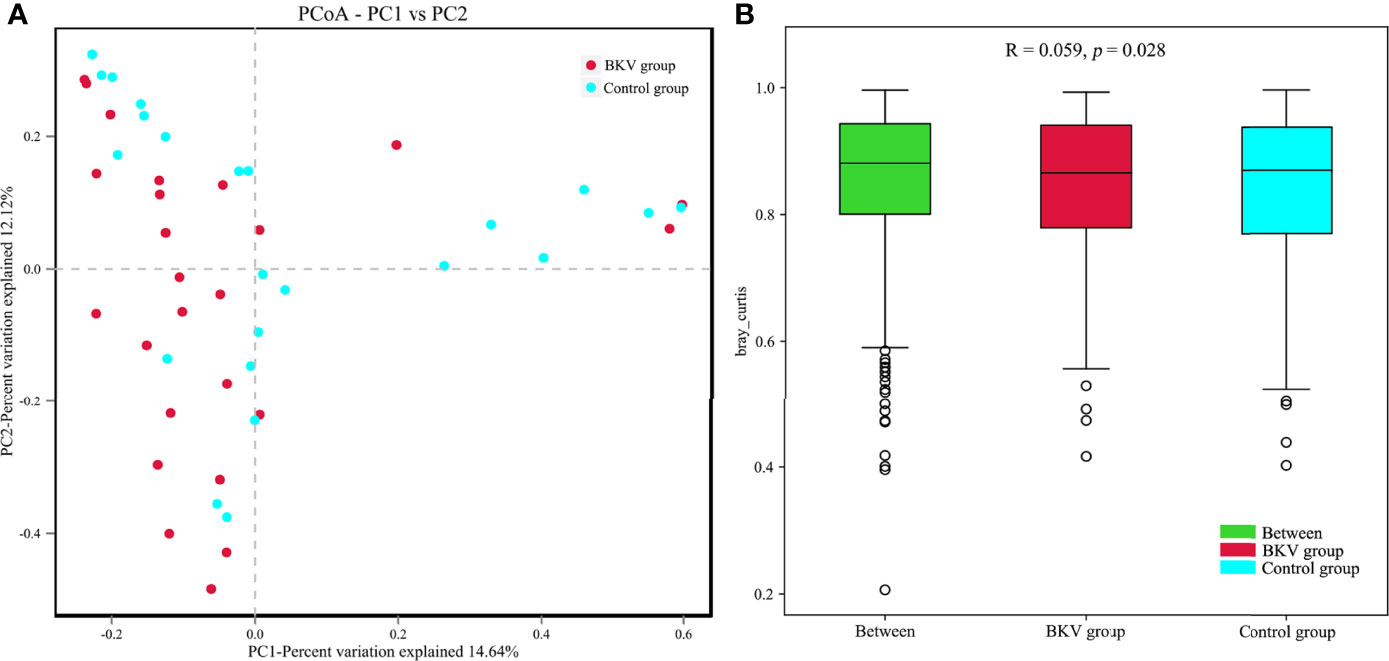
Figure 3 Comparative analysis between the BKV group and the control group. (A) PCoA was performed based on the Bray–Curtis distance. Clustering patterns of the BKV and control groups were identified by red and blue colors, respectively. PC1 and PC2 explained 14.64% and 12.12% of total variations, respectively. (B) ANOSIM at the OTU level was conducted. The difference in microbiota in the inter-group was bigger than that in the intra-group.
Difference in Bacterial Taxa Between the BKV Group and the Control Group
LEfSe was used to explore the biomarkers between the BKV group and the control group. We discovered 9 distinguishing bacterial taxa between the two groups, with a log LDA score >4.0. The abundance of the class Clostridia, order Clostridiales, family Peptostreptococcaceae, Veillonellaceae, genus Romboutsia, and species uncultured bacterium of genus Romboutsia was higher, while the abundance of family Enterococcaceae, genus Enterococcus, and species uncultured bacterium of genus Enterococcus was lower in the BKV group than that in the control group. The increase and decrease of bacterial taxa abundance in the BKV group were represented by different colors (Figure 4).
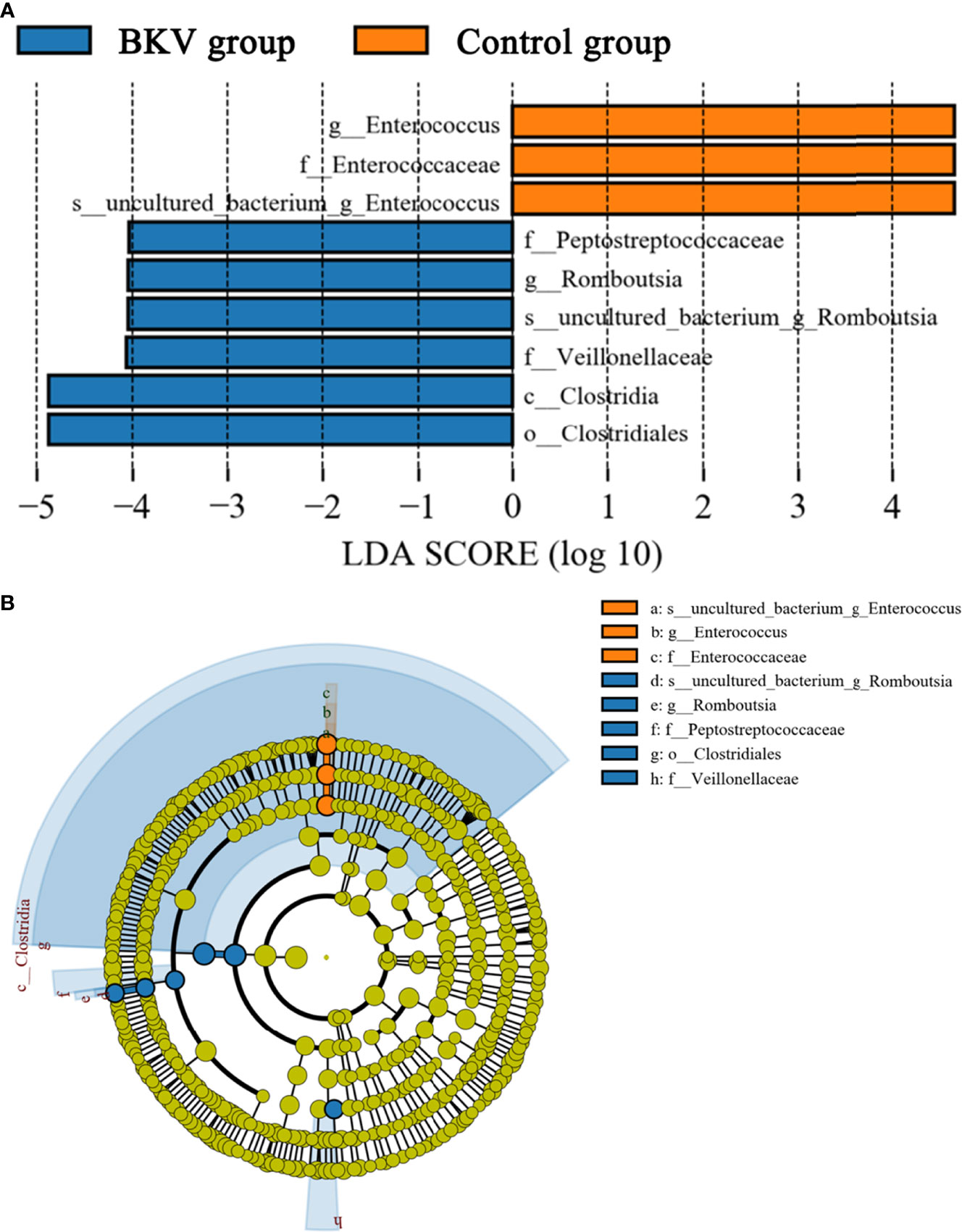
Figure 4 LEfSe analysis between the BKV group and the control group. (A) The LEfSe analysis demonstrated a significant difference in gut microbiota between the BKV group and the control group, with a log LDA score >4.0. The increase and decrease of bacterial taxa abundance in the BKV group were represented by blue and orange colors, respectively. (B) The cladogram demonstrated relationships among those taxa.
Prediction Model of BKV Infection
We established a random forest model using genus taxa to predict the BKV infectious status. The random forest model achieved the best accuracy (80.71%) to distinguish BKV-infected patients and those non-infected. The best model achieved an area under the curve (AUC) of 0.82 (Figure 5). Two genera were included in the best model, which were Romboutsia and Actinomyces.
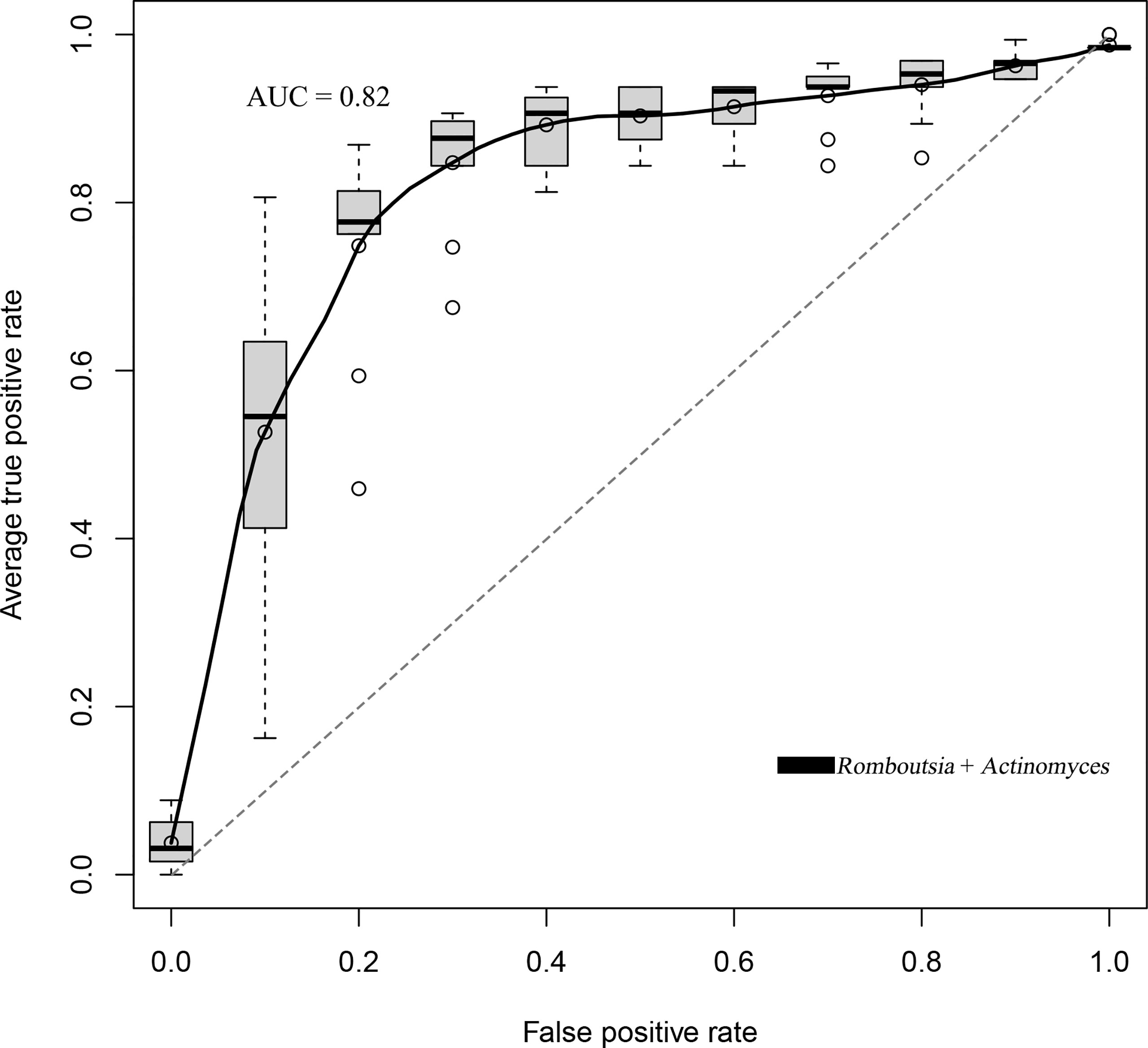
Figure 5 The ROC curve of the random forest model that trades off the rate of true positives against the rate of false positives. The best accuracy of the random forest model was 80.71% with an AUC of 0.82.
Discussion
The gut microbiota exhibited various physiological functions and plays a pivotal role in both the metabolism and immune system. A recent study demonstrated that the changes in the gut microbiota after renal transplantation might be associated with posttransplant infections (Lee et al., 2019). However, the correlation between microbiota dysbiosis and posttransplant BKV infection was rarely studied. Here, we discovered a gut microbiota dysbiosis in BKV-infected renal transplant recipients, which had not been reported previously.
In this study, the alpha diversity of the gut microbiota in BKV-infected patients was comparable to that in controls. However, the beta diversity revealed a significant difference between the two populations. At the phylum level, we found an obvious increase in the Firmicutes/Bacteroidetes ratio in the course of BKV infection. The phyla Bacteroidetes and Firmicutes contained the most abundant components of the human gut microbiota (Qin et al., 2010). Similarly, the elevated ratio had also been reported in several infectious diseases, including HBV infection (Zhu et al., 2019), HIV infection (González-Hernández et al., 2019), and Clostridium difficile infection (Bishara et al., 2013). Also, the alteration in this ratio was observed in obesity (Crovesy et al., 2020). An opposite situation was observed in some autoimmune diseases, such as type 1 diabetes (Zhou et al., 2020), Sjögren’s syndrome (Moon et al., 2020), and systemic lupus erythematosus (van der Meulen et al., 2019), in which the dysbiosis was characterized by a decrease in the Firmicutes/Bacteroidetes ratio. In the top 20 dominant genera, we found a significant increase in both Romboutsia and Roseburia in BKV-infected patients, which exhibited an anti-inflammatory effect and negatively correlated with inflammatory bowel disease (Schirmer et al., 2019; Qiu et al., 2020). Interestingly, we also found a lower CD4/CD8 ratio in BKV-infected patients, which indicated a higher risk of infection. Correlation analysis demonstrated that Bacteroidetes was positively correlated with CD4/CD8 ratio. Thus, this dysbiosis might contribute to BKV infection by immune modulation functions of the microbiota. However, it was still not known whether the microbial community was altered as a consequence of the infection process or the dysbiosis contributed to the infection.
Butyrate was a short-chain fatty acid, which was known to contribute to anti-inflammatory and immunosuppressive properties by affecting the differentiation, maturation, and function of dendritic cells and macrophages generated from human monocytes (Millard et al., 2002). It could induce extrathymic Treg differentiation, which was positively correlated with BK viremia (Arpaia et al., 2013; Furusawa et al., 2013; Karantanos et al., 2019). A recent study also demonstrated the impact of Tregs on immunity by Treg/IL-10/Th17 axis (Hui et al., 2019). In this study, we discovered 9 distinguishing bacterial taxa that were biomarkers of BKV infection that might be potential therapeutic targets. The abundance of the class Clostridia, order Clostridiales, family Peptostreptococcaceae, Veillonellaceae, genus Romboutsia, and species uncultured bacterium of genus Romboutsia was higher, while the abundance of family Enterococcaceae, genus Enterococcus, and species uncultured bacterium of genus Enterococcus was lower in the BKV group. Interestingly, the order Clostridiales (within the class Clostridia) includes many butyrate producers, such as the family Peptostreptococcaceae, Veillonellaceae, and genus Romboutsia (within the family Peptostreptococcaceae) (Esquivel-Elizondo et al., 2017; Liu et al., 2019; Chen et al., 2021), which might be correlated with BKV infection by their immunosuppressive properties. Inversely, the genus Enterococcus (within the family Enterococcaceae), a group of pro-inflammatory bacteria, declined in the feces of BKV-infected patients.
In this study, we established a random forest model to predict the BKV infectious status. We found that the inclusion of the genus taxa achieved the best accuracy (80.71%), which was better than that of other taxonomic categories of the gut microbiota. Moreover, only the combination of Romboutsia and Actinomyces achieved the best classification accuracy for the patients from the two groups. Either single feature or other feature combinations could not achieve a good classification effect for the model. We considered that this model could be clinically used as a non-invasive supplementary diagnostic method of BKV infection, and the efficiency could be improved with the continual enrollment of clinical samples in further studies.
Conclusion
BKV-infected patients had a gut microbiota dysbiosis that the Firmicutes/Bacteroidetes ratio increased in the course of the viral infection. Nine distinguishing bacterial taxa might be potential biomarkers of BKV infection. The random forest model achieved an accuracy of 80.71% in predicting the BKV infectious status, with Romboutsia and Actinomyces included.
Limitations
This study has limitations. Firstly, the sample size is limited. Secondly, the metagenome analysis is unused, which can provide more detailed information to explore the correlation between gut microbiota dysbiosis and BKV infection. Thirdly, a longitudinal study and metabolomics analysis are needed to explore and further understand the correlation between gut microbiota dysbiosis and the development of BKV infection.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The name of the repository and accession number can be found below: NCBI; PRJNA801362.
Ethics Statement
This study was conducted according to the ethical guidelines of the Helsinki Declaration and approved by the ethics committee of Beijing Friendship Hospital. The patients/participants provided their written informed consent to participate in this study. None of the organs were procured from executed prisoners. All of the organs were procured after informed consent and allocated by the China Organ Transplant Response System.
Author Contributions
JZ participated in design of the work, analysis and interpretation of data, drafting and revising the work. HQ and MC participated in acquisition and analysis of data and revising the work. YY participated in interpretation of data and revising the work. JL participated in conception and design of the work, interpretation of data, revising the work, acquisition of the funding, and supervision of the study. All authors contributed to the article and approved the submitted version.
Funding
The study was funded by the Natural Science Foundation of Beijing Municipality (No. 7192043).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to Beijing Friendship Hospital, Capital Medical University, for providing data for this study.
Abbreviations
ACE, abundance-based coverage estimator; ANOSIM, analysis of similarities; AUC, area under the curve; BKV, BK polyomavirus; LDA, linear discriminant analysis; LEfSe, linear discriminant analysis effect size; OTU, operational taxonomic unit; PCoA, principal coordinate analysis; PCR, polymerase chain reaction; rRNA, ribosomal RNA; SD, standard deviation; Tregs, T-regulatory cells.
References
Ambalathingal, G. R., Francis, R. S., Smyth, M. J., Smith, C., Khanna, R. (2017). BK Polyomavirus: Clinical Aspects, Immune Regulation, and Emerging Therapies. Clin. Microbiol. Rev. 30, 503–528. doi: 10.1128/CMR.00074-16
Arpaia, N., Campbell, C., Fan, X., Dikiy, S., van der Veeken, J., deRoos, P., et al. (2013). Metabolites Produced by Commensal Bacteria Promote Peripheral Regulatory T-Cell Generation. Nature 504, 451–455. doi: 10.1038/nature12726
Baek, C. H., Kim, H., Yu, H., Yang, W. S., Han, D. J., Park, S. K. (2018). Risk Factors of Acute Rejection in Patients With BK Nephropathy After Reduction of Immunosuppression. Ann. Transplant. 23, 704–712. doi: 10.12659/AOT.910483
Bishara, J., Farah, R., Mograbi, J., Khalaila, W., Abu-Elheja, O., Mahamid, M., et al. (2013). Obesity as a Risk Factor for Clostridium Difficile Infection. Clin. Infect. Dis. 57, 489–493. doi: 10.1093/cid/cit280
Campisciano, G., de Manzini, N., Delbue, S., Cason, C., Cosola, D., Basile, G., et al. (2020). The Obesity-Related Gut Bacterial and Viral Dysbiosis Can Impact the Risk of Colon Cancer Development. Microorganisms 8, 431. doi: 10.3390/microorganisms8030431
Chan, S., Isbel, N. M., Hawley, C. M., Campbell, S. B., Campbell, K. L., Morrison, M., et al. (2019). Infectious Complications Following Kidney Transplantation-A Focus on Hepatitis C Infection, Cytomegalovirus Infection and Novel Developments in the Gut Microbiota. Medicina (Kaunas) 55, 672. doi: 10.3390/medicina55100672
Chen, Z., Radjabzadeh, D., Chen, L., Kurilshikov, A., Kavousi, M., Ahmadizar, F., et al. (2021). Association of Insulin Resistance and Type 2 Diabetes With Gut Microbial Diversity: A Microbiome-Wide Analysis From Population Studies. JAMA Netw. Open 4, e2118811. doi: 10.1001/jamanetworkopen.2021.18811
Crovesy, L., Masterson, D., Rosado, E. L. (2020). Profile of the Gut Microbiota of Adults With Obesity: A Systematic Review. Eur. J. Clin. Nutr. 74, 1251–1262. doi: 10.1038/s41430-020-0607-6
Cummings, J. H. (1983). Fermentation in the Human Large Intestine: Evidence and Implications for Health. Lancet 1, 1206–1209. doi: 10.1016/S0140-6736(83)92478-9
Dalianis, T., Eriksson, B. M., Felldin, M., Friman, V., Hammarin, A. L., Herthelius, M., et al. (2019). Management of BK-Virus Infection - Swedish Recommendations. Infect. Dis. (Lond.) 51, 479–484. doi: 10.1080/23744235.2019.1595130
Esquivel-Elizondo, S., Ilhan, Z. E., Garcia-Peña, E. I., Krajmalnik-Brown, R. (2017). Insights Into Butyrate Production in a Controlled Fermentation System via Gene Predictions. mSystems 2, e00051–e00017. doi: 10.1128/mSystems.00051-17
Furusawa, Y., Obata, Y., Fukuda, S., Endo, T. A., Nakato, G., Takahashi, D., et al. (2013). Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 504, 446–450. doi: 10.1038/nature12721
González-Hernández, L. A., Ruiz-Briseño, M. D. R., Sánchez-Reyes, K., Alvarez-Zavala, M., Vega-Magaña, N., López-Iñiguez, A., et al. (2019). Alterations in Bacterial Communities, SCFA and Biomarkers in an Elderly HIV-Positive and HIV-Negative Population in Western Mexico. BMC Infect. Dis. 19, 234. doi: 10.1186/s12879-019-3867-9
Haase, S., Haghikia, A., Wilck, N., Müller, D. N., Linker, R. A. (2018). Impacts of Microbiome Metabolites on Immune Regulation and Autoimmunity. Immunology 154, 230–238. doi: 10.1111/imm.12933
Hanada, S., Pirzadeh, M., Carver, K. Y., Deng, J. C. (2018). Respiratory Viral Infection-Induced Microbiome Alterations and Secondary Bacterial Pneumonia. Front. Immunol. 9. doi: 10.3389/fimmu.2018.02640
Hariharan, S. (2006). BK Virus Nephritis After Renal Transplantation. Kidney Int. 69, 655–662. doi: 10.1038/sj.ki.5000040
Herlemann, D. P., Labrenz, M., Jürgens, K., Bertilsson, S., Waniek, J. J., Andersson, A. F. (2011). Transitions in Bacterial Communities Along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 5, 1571–1579. doi: 10.1038/ismej.2011.41
Hosseinkhani, F., Heinken, A., Thiele, I., Lindenburg, P. W., Harms, A. C., Hankemeier, T. (2021). The Contribution of Gut Bacterial Metabolites in the Human Immune Signaling Pathway of Non-Communicable Diseases. Gut Microbes 13, 1–22. doi: 10.1080/19490976.2021.1882927
Hui, W., Yu, D., Cao, Z., Zhao, X. (2019). Butyrate Inhibit Collagen-Induced Arthritis via Treg/IL-10/Th17 Axis. Int. Immunopharmacol. 68, 226–233. doi: 10.1016/j.intimp.2019.01.018
Kalina, U., Koyama, N., Hosoda, T., Nuernberger, H., Sato, K., Hoelzer, D., et al. (2002). Enhanced Production of IL-18 in Butyrate-Treated Intestinal Epithelium by Stimulation of the Proximal Promoter Region. Eur. J. Immunol. 32, 2635–2643. doi: 10.1002/1521-4141(200209)32:9<2635::AID-IMMU2635>3.0.CO;2-N
Karantanos, T., Kim, H. T., Tijaro-Ovalle, N. M., Li, L., Cutler, C., Antin, J. H., et al. (2019). Reactivation of BK Virus After Double Umbilical Cord Blood Transplantation in Adults Correlates With Impaired Reconstitution of CD4(+) and CD8(+) T Effector Memory Cells and Increase of T Regulatory Cells. Clin. Immunol. 207, 18–23. doi: 10.1016/j.clim.2019.06.010
Lee, J. R., Huang, J., Magruder, M., Zhang, L. T., Gong, C., Sholi, A. N., et al. (2019). Butyrate-Producing Gut Bacteria and Viral Infections in Kidney Transplant Recipients: A Pilot Study. Transpl. Infect. Dis. 21, e13180. doi: 10.1111/tid.13180
Liu, S., Li, E., Sun, Z., Fu, D., Duan, G., Jiang, M., et al. (2019). Altered Gut Microbiota and Short Chain Fatty Acids in Chinese Children With Autism Spectrum Disorder. Sci. Rep. 9, 287. doi: 10.1038/s41598-018-36430-z
McNees, A. L., White, Z. S., Zanwar, P., Vilchez, R. A., Butel, J. S. (2005). Specific and Quantitative Detection of Human Polyomaviruses BKV, JCV, and SV40 by Real Time PCR. J. Clin. Virol. 34, 52–62. doi: 10.1016/j.jcv.2004.12.018
Millard, A. L., Mertes, P. M., Ittelet, D., Villard, F., Jeannesson, P., Bernard, J. (2002). Butyrate Affects Differentiation, Maturation and Function of Human Monocyte-Derived Dendritic Cells and Macrophages. Clin. Exp. Immunol. 130, 245–255. doi: 10.1046/j.0009-9104.2002.01977.x
Moon, J., Choi, S. H., Yoon, C. H., Kim, M. K. (2020). Gut Dysbiosis Is Prevailing in Sjögren's Syndrome and is Related to Dry Eye Severity. PloS One 15, e0229029. doi: 10.1371/journal.pone.0229029
Myint, T. M., Chong, C. H. Y., Wyld, M., Nankivell, B., Kable, K., Wong, G. (2022). Polyoma BK Virus in Kidney Transplant Recipients: Screening, Monitoring and Management. Transplantation 106, e76–e89. doi: 10.1097/TP.0000000000003801
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K. S., Manichanh, C., et al. (2010). A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 464, 59–65. doi: 10.1038/nature08821
Qiu, X., Zhao, X., Cui, X., Mao, X., Tang, N., Jiao, C., et al. (2020). Characterization of Fungal and Bacterial Dysbiosis in Young Adult Chinese Patients With Crohn's Disease. Therap. Adv. Gastroenterol. 13, 1756284820971202. doi: 10.1177/1756284820971202
Schirmer, M., Garner, A., Vlamakis, H., Xavier, R. J. (2019). Microbial Genes and Pathways in Inflammatory Bowel Disease. Nat. Rev. Microbiol. 17, 497–511. doi: 10.1038/s41579-019-0213-6
van der Meulen, T. A., Harmsen, H. J. M., Vila, A. V., Kurilshikov, A., Liefers, S. C., Zhernakova, A., et al. (2019). Shared Gut, But Distinct Oral Microbiota Composition in Primary Sjögren's Syndrome and Systemic Lupus Erythematosus. J. Autoimmun. 97, 77–87. doi: 10.1016/j.jaut.2018.10.009
Vasileiadis, S., Puglisi, E., Arena, M., Cappa, F., Cocconcelli, P. S., Trevisan, M. (2012). Soil Bacterial Diversity Screening Using Single 16S rRNA Gene V Regions Coupled With Multi-Million Read Generating Sequencing Technologies. PloS One 7, e42671. doi: 10.1371/journal.pone.0042671
Yi, S. G., Knight, R. J., Lunsford, K. E. (2017). BK Virus as a Mediator of Graft Dysfunction Following Kidney Transplantation. Curr. Opin. Organ Transplant. 22, 320–327. doi: 10.1097/MOT.0000000000000429
Zakaria, Z. E., Elokely, A. M., Ghorab, A. A., Bakr, A. I., Halim, M. A., Gheith, O. A., et al. (2019). Screening for BK Viremia/Viruria and the Impact of Management of BK Virus Nephropathy in Renal Transplant Recipients. Exp. Clin. Transplant. 17, 83–91. doi: 10.6002/ect.MESOT2018.O17
Zhou, H., Zhao, X., Sun, L., Liu, Y., Lv, Y., Gang, X., et al. (2020). Gut Microbiota Profile in Patients With Type 1 Diabetes Based on 16S rRNA Gene Sequencing: A Systematic Review. Dis. Markers 2020, 3936247. doi: 10.1155/2020/3936247
Keywords: gut microbiota, BK polyomavirus, infection, renal transplantation, microbial dysbiosis
Citation: Zhang J, Qin H, Chang M, Yang Y and Lin J (2022) Gut Microbiota Dysbiosis in BK Polyomavirus-Infected Renal Transplant Recipients: A Case-Control Study. Front. Cell. Infect. Microbiol. 12:860201. doi: 10.3389/fcimb.2022.860201
Received: 22 January 2022; Accepted: 14 April 2022;
Published: 27 May 2022.
Edited by:
Juan Carlos Rodriguez Diaz, Hospital General Universitario de Alicante, SpainReviewed by:
Bangzhou Zhang, Xiamen University, ChinaAshraf Kariminik, Islamic Azad University Kerman, Iran
Ramin Yaghobi, Shiraz University of Medical Sciences, Iran
Copyright © 2022 Zhang, Qin, Chang, Yang and Lin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Lin, bfhlinjun@hotmail.com