Differential effects of location and object overlap on new learning
 Benjamin Chaloupka
Benjamin Chaloupka Dagmar Zeithamova
Dagmar Zeithamova- 1Department of Psychology, University of Oregon, Eugene, OR, United States
- 2Institute of Neuroscience, University of Oregon, Eugene, OR, United States
Introduction: Overlap with prior experience facilitates learning in some cases while hindering it in others. As facilitation and interference are typically studied in separate lines of research, using distinct paradigms, it is unclear what key factors drive the opposing behavioral outcomes.
Methods: In two experiments, we tested whether both effects can be observed within a single task, depending on what overlaps between experiences. Participants completed a novel task in which they learned a grid of object-location associations, followed by a second grid that overlapped with the first in locations and/or objects. We hypothesized that overlap of locations would serve as a spatial schema, facilitating new learning, while overlap of objects would create interference.
Results: In line with our hypothesis, we found that location overlap facilitated learning of the second grid, while object overlap hindered learning of the second grid. We replicated these findings in a second experiment, additionally showing that both effects remain largely stable across two distinct grid shapes.
Discussion: These results demonstrate that the effect of overlap can be manipulated within a single task, pinpointing one factor that determines the direction of the effect and highlighting the differential roles of “what” and “where” in the organization of memory.
1 Introduction
Overlap of experiences can facilitate new learning by allowing us to integrate new experiences within an established framework. For example, imagine that you are visiting a friend's house for the first time. You are quickly able to orient yourself to the rooms in the house because you already have a framework, or schema, of how a house is laid out. You know that the kitchen and living room are likely on the ground level, the bedrooms are upstairs, and so on. In this case, your prior knowledge of houses facilitates your learning of the layout of your friend's house. However, overlap of experiences can also hinder new learning via proactive interference. Imagine that you head into your friend's kitchen to grab a spoon for a snack. You keep your utensils in the drawer next to the stove, but your friend keeps theirs in the drawer next to the dishwasher. Even after you find the utensils the first time, you keep going to the wrong drawer every time you come back to your friend's house. In this case, your prior knowledge (where you keep your utensils) interferes with your learning of the new information (where your friend keeps their utensils). How can overlap of experience facilitate new learning in some cases while hindering it in others?
Facilitation has often been studied in the context of schemas. A schema is a structured mental representation of a concept or other complex stimuli that allows us to quickly integrate new information into memory. The term “schema” was first applied to the field of cognitive psychology by Bartlett (1932), defining it as the mechanism of learning by which new experiences are overlaid upon prior experiences. Attneave (1957) demonstrated that participants with prior knowledge of two kinds of schemas–letter patterns and polygons–performed better on a paired-associates task compared to participants without prior knowledge of the schemas. Building on this work, Posner et al. (1967) showed that the degree of overlap among stimuli directly related to the rate of new learning, such that learning was faster when overlap was higher. These effects have been replicated across a plethora of contexts (van Kesteren et al., 2014; for a review, see van Kesteren et al., 2012) and modalities (King et al., 2019). While it has been thought that schemas take a long time to develop (Sommer, 2016), more recent research suggests that schemas may develop quickly, even when not called upon until the episodic memory for a specific event begins to fade (Tompary et al., 2020).
In a seminal study that brought schema research back into the spotlight, Tse et al. (2007) found that spatial schemas provide a framework that facilitates new learning in rats. In this study, rats were trained over several months to learn six flavor-location associations in an arena, until they learned the associations well and presumably developed a spatial schema of the arena and all the relevant locations in it. Once a spatial schema was formed, two of the flavor targets were replaced by new flavors and moved to neighboring locations. The rats were given a single training trial in which the new flavor-location pairs were rewarded. Despite receiving only one trial to learn the new associations, the rats successfully learned and retained the new pairs. This indicates the facilitatory effect of a spatial schema on new learning. This pattern has been replicated in humans, demonstrating that an experimentally-learned spatial schema effectively facilitates new learning (van Buuren et al., 2014). Collectively, this research demonstrates that overlap with prior memories, particularly spatial schemas, can facilitate new learning.
An equally established consequence of an overlap with prior memories is interference. Specifically, proactive interference–the memory of old information hindering the learning of new information–was first described by Maslow (1934) nearly a century ago. Later, Melton and von Lackum (1941) found that learning of consonant strings was slower when they overlapped highly with previously learned consonant strings. This was one of the first studies that demonstrated that greater stimulus overlap leads to greater interference. Wickens et al. (1963) confirmed this effect by showing that proactive interference only persists so long as the stimuli are of a similar kind; switching stimuli types (consonant strings to number strings, or vice versa) did not lead to interference. The finding that overlap with prior memories can hinder new learning has been extended to semantic categories (Wixted and Rohrer, 1993) and languages (Goggin and Wickens, 1971) and has been confirmed using a variety of tasks (Bunting, 2006; for a review, see Kliegl and Bäuml, 2021). Additionally, overlap can be detrimental to associative memory, as demonstrated in a classic AB/AC paradigm: after learning one set of paired-associates (AB), participants have greater difficulty learning a new set overlapping (AC) associations than non-overlapping (DE) associations (Postman et al., 1974; Tulving and Watkins, 1974; Wahlheim and Jacoby, 2013; Caplan et al., 2022). Taken together, these findings indicate that overlap with an existing memory makes it more difficult to encode a new one, and competition between two overlapping memories makes the retrieval of a target memory more difficult (Kuhl et al., 2007, 2011). Thus, a large body of work has demonstrated that overlap with prior memories can hinder learning.
As both the beneficial and detrimental effects of information overlap on new learning have been well documented, how can we reconcile these findings? As the schema research and interference research have proceeded largely in parallel, using distinct task paradigms, stimuli, timing, length of training, etc., it is challenging to discern which differences are critical for the distinct behavioral outcomes—facilitation vs. interference—across studies. In the present study, we ask if it is possible to observe both facilitation and interference within a single task by manipulating what overlaps across experiences. Notably, many previous studies showing the facilitatory effect of overlap did so using overlap with spatial, conceptual, and relational schemas (Tse et al., 2007; van Buuren et al., 2014). In contrast, many previous studies showing the interference effect of overlap utilized list learning or associative memory paradigms (Wickens et al., 1963; Goggin and Wickens, 1971; Postman et al., 1974; Wixted and Rohrer, 1993; Bunting, 2006). We thus developed a novel object-location association task that allowed us to test the effect of two types of overlap, spatial locations and object set, within a single task. Participants learned one grid of object-location associations, and then learned a second grid of object-location associations that overlapped with the first in objects and/or locations. Given prior work showing facilitatory effects of overlap with existing spatial schemas, we hypothesized that location overlap will facilitate new learning. In contrast, given prior work showing detrimental effect of overlap for word lists and object pairs, we hypothesized that object overlap will interfere with new learning. By manipulating these two factors together, we aim to bridge the gap between schema and interference research and reconcile the conflicting findings regarding the effect of overlap on memory. In a second experiment, we aim to replicate the findings from Experiment 1 and extend our findings by manipulating a third variable.
2 Experiment 1
2.1 Materials and methods
2.1.1 Participants
Participants were recruited from the University of Oregon human subjects pool and received course credit for their participation. Informed consent was obtained from all participants and experimental procedures were approved by Research Compliance Services at the University of Oregon. We recruited 102 in-person participants (79 female, 22 male, 1 other), age 18–25 years (M = 19.33, SD = 1.52). Four participants were excluded for failing to complete the task, and 13 were excluded for failing to learn at least one of the grids within the allotted number of trials. The target sample size was determined to be 160 based on a power analysis (α = 0.05, power = 0.80, partial η2 = 0.05 estimated for each main effect). However, the COVID-19 pandemic halted in-person data collection before a full sample could be collected. Thus, we recruited an additional 273 online participants. Due to experimenter error in pivoting to online data collection amidst the pandemic, demographic information was not collected for the online participants. Since both in-person and online participants were recruited through introductory psychology courses at the University of Oregon, we are confident that the in-person demographic information collected is representative of the overall sample. Of the online sample, 50 were excluded from analyses for failing to complete the task or failure to follow instructions, 13 were excluded for failing to learn one or both grids, and 6 were excluded for failing to pass the attention check. After exclusions, 85 in-person participants and 204 online participants were included in all analyses, for a total of 289 participants. For brevity and clarity, we report analyses from the full sample, including both in-person and online participants. However, we also analyzed the data with setting as an additional factor to verify that the pattern of results was the same for both the in-person sample and the online sample.
2.1.2 Procedure
2.1.2.1 Overview
Participants completed a novel grid learning task in which they learned grids of object-location associations. Each 8 x 8 grid contained eight objects in various locations throughout the grid (see Figure 1). Each participant sequentially learned two object-location grids, where the second grid would overlap with the first in the objects, locations, both, or neither. By measuring how long it took participants to learn each grid, we could evaluate if overlap in each of these factors was facilitatory or detrimental to new learning. The grid task was administered using PsychoPy (RRID:SCR_006571) for in-person participants and Pavlovia (RRID:SCR_023320) for online participants.
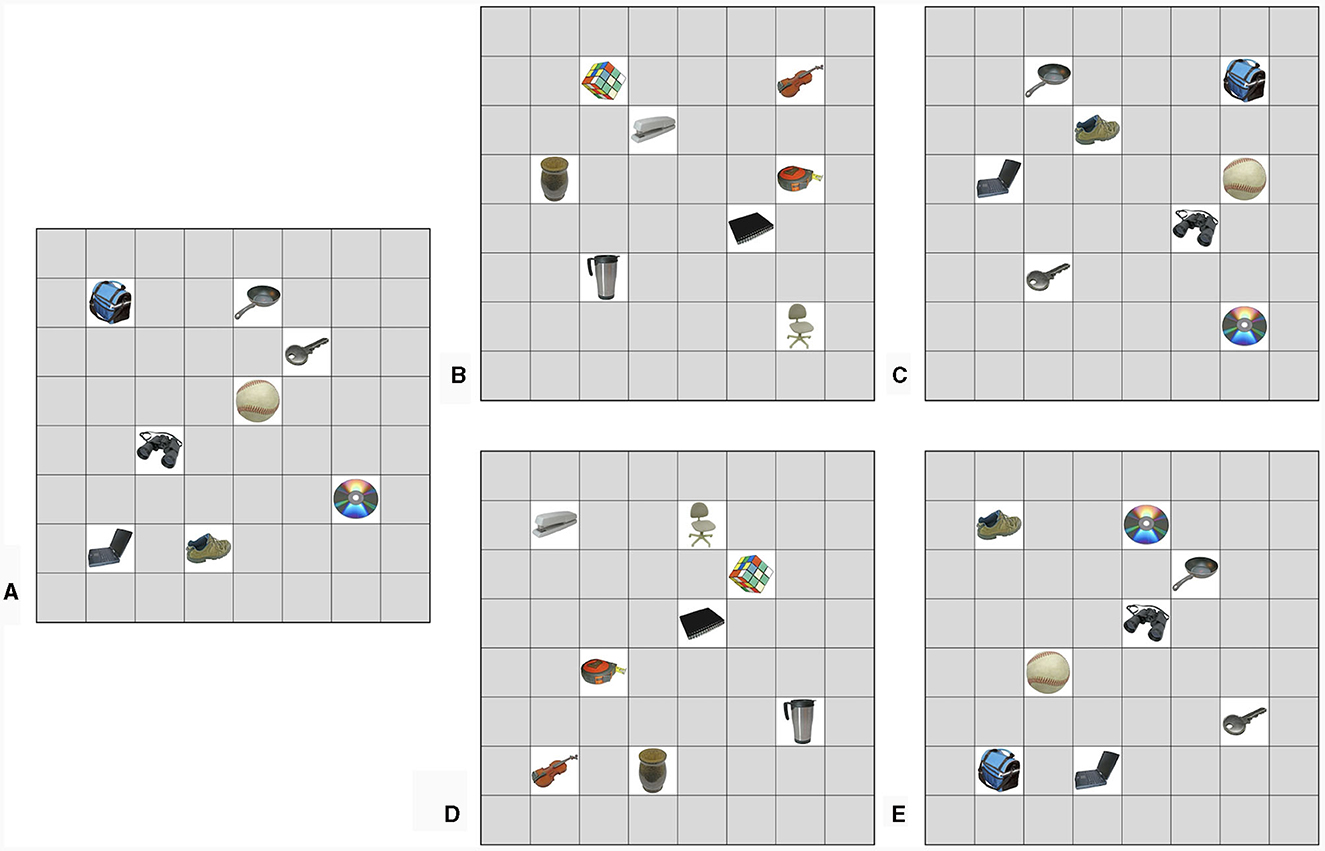
Figure 1. The grids of object-location associations used in Experiment 1. (A) Example starting grid. (B) No overlap condition; different stimulus set and locations. (C) Object overlap condition; same stimulus set in different locations. (D) Location overlap condition; different stimulus set but same locations. (E) Full overlap condition; same stimulus set and locations. Images used in this figure are unaltered images from the BOSS stimulus sets (Brodeur et al., 2010, 2014) used under the Creative Commons Attribution License (https://creativecommons.org/licenses/by-sa/3.0/).
2.1.2.2 Stimuli
We combined two sets of objects and two sets of locations to create eight distinct grids to be used in the study. To create each location set, we pseudo-randomly selected locations from the grid, with the constraint that no location was selected at the border (edge of the grid) and no two locations were immediate neighbors vertically or horizontally (diagonal contact ok). Each object set was then randomly placed within each location set, for a total of four object-location grids. The objects in each of these grids were then randomly shuffled (with no object in the same location across the two grids) to create a second version of each object-location grid, for a total of eight possible grids used in the study across participants. All grids were constructed in Microsoft Excel (RRID:SCR_016137).
Participants were randomly assigned one of these grids to be their starting grid, and the starting grid was roughly counterbalanced across participants and conditions. Their second grid was chosen from the remaining grids based on the condition that the participant was randomly assigned to (see Figure 1). In the no overlap control group, the second grid used a new set of objects and a new set of locations (Figure 1B). In the object overlap group, the second grid contained the same set of objects, but placed in a new set of locations (Figure 1C). In the location overlap group, the second grid contained a new set of objects, but in the same locations as the first grid (Figure 1D). In the full overlap group, the second grid used the same set of objects and the same set of locations, but the objects were shuffled so that the grids were not identical (Figure 1E).
2.1.2.3 Grid learning
First, participants learned their starting grid. The grid learning procedure is depicted in Figure 2A and was the same for Grid 1 and Grid 2 (Figure 2B). The grid was displayed on screen for 8 s followed by 4 s of a fixation cross. Then, a blank grid appeared with the eight objects displayed beneath the grid. Participants attempted to recreate the grid by dragging and dropping the objects to their correct locations within the blank grid. The test was self-paced. Participants submitted each attempt by pressing the spacebar. Following each attempt, participants received feedback in the form of the number of correctly placed objects (out of eight). If all eight objects were correctly placed, they moved on to the next phase of the task. Otherwise, they repeated this process until they correctly placed all eight objects with a maximum of eight attempts. We recorded the number of trials it took each participant to learn the grid. The duration of each grid (8 s) and the maximum number of trials to learn the grid (8 attempts) was based on pilot data collection to ensure that the majority of participants need more than 1 attempt but fewer than 8 attempts to learn. Participants who failed to learn the grid after eight attempts were categorized as “non-learners” and excluded from analysis. Prior to the actual task, participants received instructions and practiced the task using a simplified training grid with just two objects (not used in the main experiment).
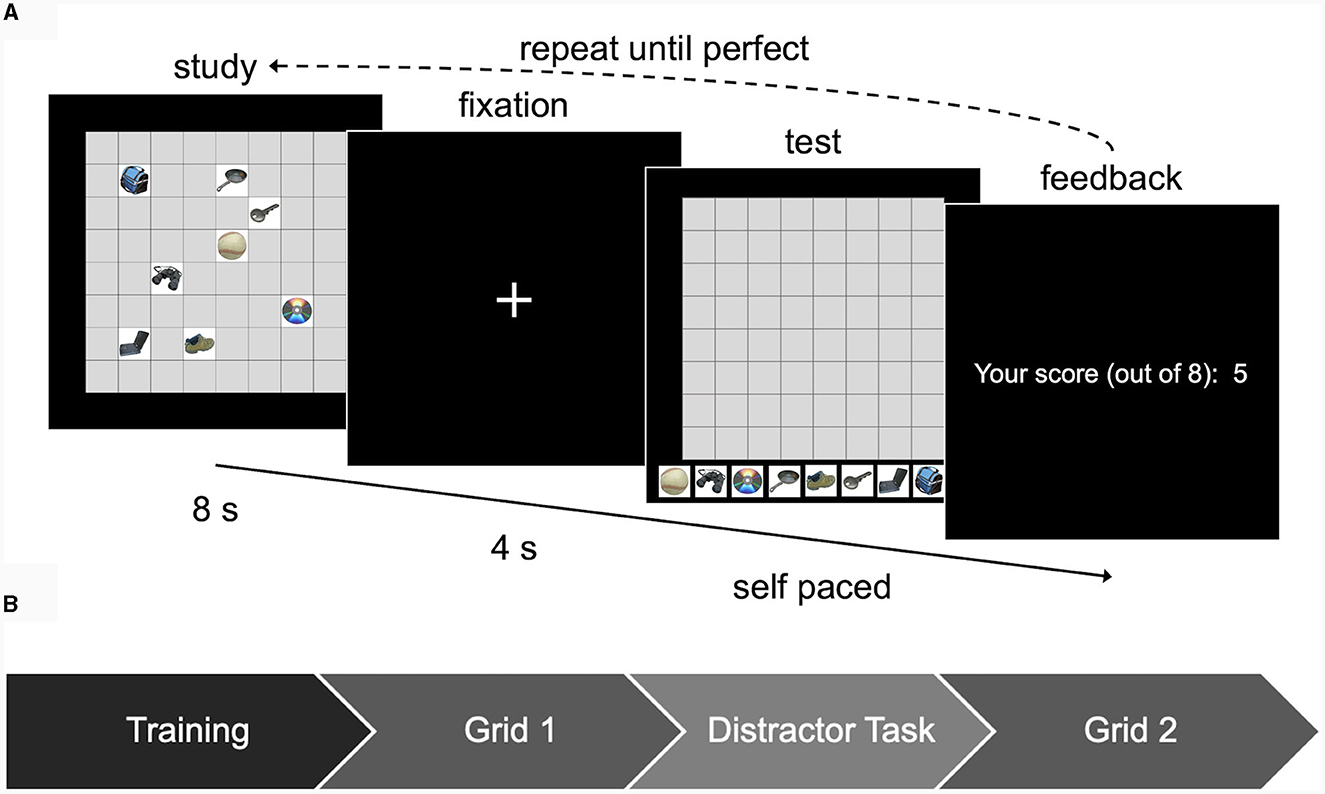
Figure 2. Grid learning task used in Experiment 1. (A) Participants learned each of the two grids using this process. (B) Overview of the timeline of the grid task. Images used in this figure are unaltered images from the BOSS stimulus sets (Brodeur et al., 2010, 2014) used under the Creative Commons Attribution License (https://creativecommons.org/licenses/by-sa/3.0/).
2.1.2.4 Math distractor task
Next, participants completed a math distractor task. On each trial, participants were shown a simple arithmetic expression and asked to evaluate if the expression was true or false using the “T” and “F” keys, respectively. For example, “3 + 5 = 9” required a response of “F.” Each arithmetic problem was displayed on screen for 4 s during which participants made their response. They then received feedback on screen for 2 s indicating if they were correct, incorrect, or too slow to respond. Participants completed 20 trials for a total of 2 min. This served as an attention check and working memory flush, preventing participants from actively rehearsing the previously learned grid. To ensure engagement, participants with less that 75% accuracy on this task were excluded from analyses.
2.1.2.5 Second grid learning
Following the math distractor task, participants learned their second grid using the same procedure as the first. Participants were randomly assigned to one of four groups, dictating whether Grid 2 overlaps with the previously learned Grid 1 in objects, locations, both objects and locations, or neither objects nor location (Figure 1). In addition to recording the number of trials it took for each participant to learn Grid 2, we also calculated the difference in the number of trials to learn the two grids by subtracting the number of trials to learn the second grid from the number of trials to learn the first grid (Grid 1 minus Grid 2). Thus, a difference score of 0 indicated that they took the same number of trials to learn both grids, a positive score meant that they learned the second grid faster (in fewer trials) than the first, and a negative number indicated that they learned the second grid slower (needing more trials) than the first.
2.1.2.6 Additional cognitive measures
In addition to the grid task, only the in-person participants also completed a short battery of cognitive tasks (visual working memory capacity measure: Adam et al., 2015; reading span: first developed by Daneman and Carpenter, 1980; adapted from Unsworth et al., 2005; and Raven's advanced progressive matrices: Verguts and Boeck, 2002) for a pilot project not discussed in this paper. The reading span task was administered using PsychoPy, while the visual working memory task and Raven's advanced progressive matrices were administered using MATLAB. Given that less than a third of the final sample had these measures collected, they were not used in any subsequent analyses.
2.2 Results
First, we measured the number of trials to learn the first grid (M = 4.23, SD = 1.68). Since this grid was learned prior to experimental manipulation, we predicted no significant differences between groups. To test this, we ran a 2x2 (object overlap x location overlap) factorial ANOVA with the number of trials to learn the first grid as the dependent variable (Figure 3A). As expected, we found no main effect of object overlap, F(1, 285) = 0.02, p = 0.882, = 0.00, no main effect of location overlap, F(1, 285) = 2.13, p = 0.146, = 0.01, and no interaction, F(1, 285) = 0.62, p = 0.431, = 0.00, confirming that there were no between-group differences prior to experimental manipulation.
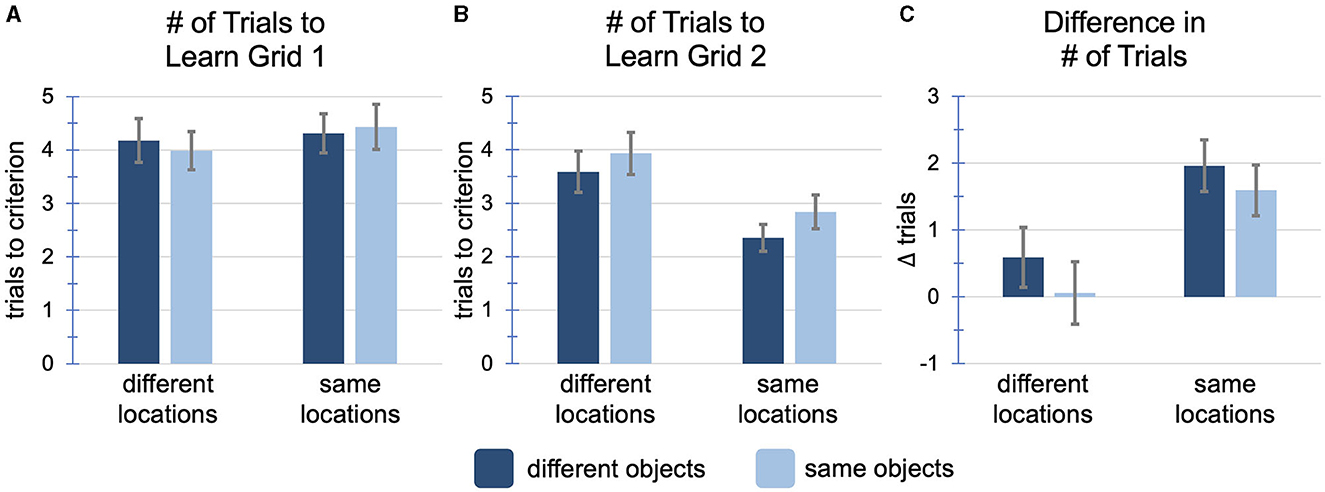
Figure 3. Results of Experiment 1. (A) The number of trials to learn the first grid by condition, prior to any manipulation. There was no main effect of object overlap (p = 0.882), no main effect of location overlap (p = 0.146), and no interaction (p = 0.431). (B) The number of trials to learn the second grid by condition, after manipulation. There was a main effect of object overlap (p = 0.016), a main effect of location overlap (p < 0.001), and no interaction (p = 0.667). (C) The difference between the number of trials to learn the two grids (grid 1–grid 2). A positive number indicates faster learning of grid 2. There was a main effect of object overlap (p = 0.038), a main effect of location overlap (p < 0.001), and no interaction (p = 0.703). Error bars represent 95% confidence intervals for all plots.
Of main interest, we measured the number of trials to learn the second grid (M = 3.15, SD = 1.58) as well as the within-subject difference in the number of trials to learn the second grid in comparison to the first grid. We hypothesized that object overlap would cause interference, resulting in a greater number of trials to learn the second grid, while location overlap would cause facilitation, resulting in fewer trials to learn the second grid. To test this hypothesis, we ran a 2 x 2 (object overlap x location overlap) factorial ANOVA with the number of trials to learn the second grid as the dependent variable (Figure 3B). Consistent with our hypothesis, we found a main effect of object overlap, F(1, 285) = 5.91, p = 0.016, = 0.02, driven by fewer trials required to learn the second grid when objects did not overlap (M = 2.92, SD = 1.51) compared to when objects did overlap (M = 3.40, SD = 1.63). We also found a main effect of location overlap, F(1, 285) = 45.81, p < 0.001, = 0.14, driven by fewer trials required to learn the second grid when locations did overlap (M = 2.58, SD = 1.25) compared to when locations did not overlap (M = 3.76, SD = 1.67). These results indicate that overlap with prior experience can have dissociable effects, depending on the type of overlap. There was no interaction, F(1, 285) = 0.19, p = 0.667, = 0.00, indicating that the main effects were independent and additive.
To ensure that these effects were not driven by individual differences in grid learning ability, we calculated the difference in the number of trials to learn the two grids by subtracting the number of trials to learn the second grid from the number of trials to learn the first grid (Figure 3C). A score of 0 would indicate that a participant took the same number of trials to learn both grids, a positive score that they learned the second grid faster than the first, and a negative score would indicate that they learned the second grid slower than the first. Averaged across conditions, participants showed practice effects, learning the second grid about one trial faster than the first (M = 1.08, SD = 1.98), with overall 63% of participants improving from the first to the second grid. However, the magnitude of the practice effect varied across conditions. A 2 x 2 factorial ANOVA with the difference score as the dependent variable replicated a main effect of object overlap, F(1, 285) = 4.33, p = 0.038, = 0.015, driven by greater improvement across grids when objects did not overlap (M = 1.33, SD = 1.94) compared to when objects did overlap (M = 0.81, SD = 1.99). When objects did not overlap, 68% of participants improved, learning the second grid in fewer trials than the first, while only 57% of participants improved when objects did overlap, = 3.61, p = 0.057. We also found a main effect of location overlap, F(1, 285) = 45.62, p < 0.001, = 0.14, driven by greater improvement across grids when locations did overlap (M = 1.79, SD = 1.70) compared to when locations did not overlap (M = 0.31, SD = 1.97). When locations did overlap, 77% of participants improved, learning the second grid in fewer trials, while only 48% of the participants improved when locations did not overlap, χ2(1, N = 289) = 26.62, p < 0.001. There was no interaction, F(1, 285) = 0.15, p = 0.703, = 0.00, confirming that the main effects were independent and additive.
To verify that these results were not affected by setting, we also conducted all the analyses with setting (in-person, online) as an additional between-subjects factor. In all analyses, we found a main effect of setting (all p's < 0.04). Online participants learned the first grid about 1 trial faster and second grid about 0.5 trials faster. Importantly, setting did not interact with any other factor and the main effects of location and object overlap remained the same.
2.3 Discussion
In Experiment 1, we wanted to know if location overlap and object overlap would have opposing effects, and if we could observe both effects within a single task. In line with our hypotheses, we found in an object-location associative learning task that location overlap facilitates new learning while object overlap hinders new learning. Additionally, our results indicate that these effects are independent and additive. While we found significant main effects of both facilitation and interference, the magnitude of the interference effect was rather small, with at 0.01–0.02. Thus, we conducted a second experiment to replicate these findings. Given that observing changes in the number of trials to criterion, each constrained between 1 and 8, provides a relatively coarse measure of learning, we reduced the exposure time and increased the maximum number of attempts to ten in the second experiment. If we again observe both facilitation and interference effects in a separate sample, we can conclude that they are robust, even if the effect size of the interference effect is small.
Additionally, our findings led us to inquire as to how these effects would hold up to context shifts, operationalized as a change of the shape of the grid. On one hand, when the context, or the shape of one's environment changes, hippocampal place cell representations undergo remapping, as observed in both rodents (Lever et al., 2002; Wills et al., 2005) and more recently humans (Wanjia et al., 2021). Across two different contexts, the same relative locations are represented by a different set of hippocampal neurons. However, it is possible that the learned spatial pattern may itself become the context, or schema. For example, people may remember the configuration of locations from the first grid, utilizing that information when it overlaps in the second grid. In this case, overlap of the spatial pattern may supersede a shift in grid shape and the facilitatory effect of location overlap would generalize.
Interference effects are also modulated by shifts in context. Specifically, interference effects caused by stimulus overlap can be eliminated by shifting contexts or stimulus types (Goggin and Wickens, 1971; Wixted and Rohrer, 1993; Bunting, 2006; Unsworth et al., 2013). Stimulus overlap may no longer be detrimental to new learning when the context changes. To explore these questions in Experiment 2, we created two distinct grid shapes to operationalize a change in the environment. We then explicitly tested whether changing the grid shape eliminates the facilitatory effect of location overlap and/or the interference effect of object overlap, or if these effects are preserved.
3 Experiment 2
3.1 Materials and methods
3.1.1 Participants
Participants were recruited from the University of Oregon human subjects pool and received course credit for their participation. Informed consent was obtained from all participants and experimental procedures were approved by Research Compliance Services at the University of Oregon. All participants completed the task online. We recruited 534 participants; 102 participants were excluded for failing to complete the task, 4 were excluded for failing to learn at least one of the grids within the allotted number of trials, 21 were excluded for failing to pass the attention check, and 17 were excluded for learning the first grid on the first trial (see below). The remaining 390 participants were used in the following analyses. In Experiment 1, we conducted a power estimate using a medium effect size since there was no directly related prior research from which to base the estimate on. However, the actual effect size for object overlap was smaller. In Experiment 2, we used the effect size from Experiment 1 to motivate our power analysis. The target sample size was determined to be 387 based on a power analysis (α = 0.05, power = 0.80, partial η2 = 0.02 estimated for each main effect, based on the results from Experiment 1). Due to experimenter error in pivoting to online data collection amidst the pandemic, demographic information was not collected for these participants. Since all participants were recruited through introductory psychology courses at the University of Oregon, we are confident that the demographic information reported in Experiment 1 is representative of the overall sample.
3.1.2 Procedure
3.1.2.1 Overview
The task procedure was largely the same as in Experiment 1. The grid task was administered online using Pavlovia (RRID:SCR_023320).
3.1.2.2 Stimuli
Here, we manipulated a third factor: grid shape. We used two distinct grid shapes. To construct the shapes, we started with an 8 x 8 grid and then eliminated two squares on each side. For the first shape, we removed the two center squares on each edge, creating an “x”-like shape. For the second shape, we removed each corner square as well as one adjacent square immediately counterclockwise to each corner, creating a “t”-like shape (see Figure 4). As in Experiment 1, we constructed two unique grids for each grid shape, object set, and location set to allow for the full overlap condition, resulting in 16 total grids. As in Experiment 1, all grids were constructed in Microsoft Excel (RRID:SCR_016137). Participants were randomly assigned one of these grids to be their starting grid, and the starting grid was roughly counterbalanced across participants and conditions. Their second grid was chosen from the remaining grids based on the condition that the participant was randomly assigned to.
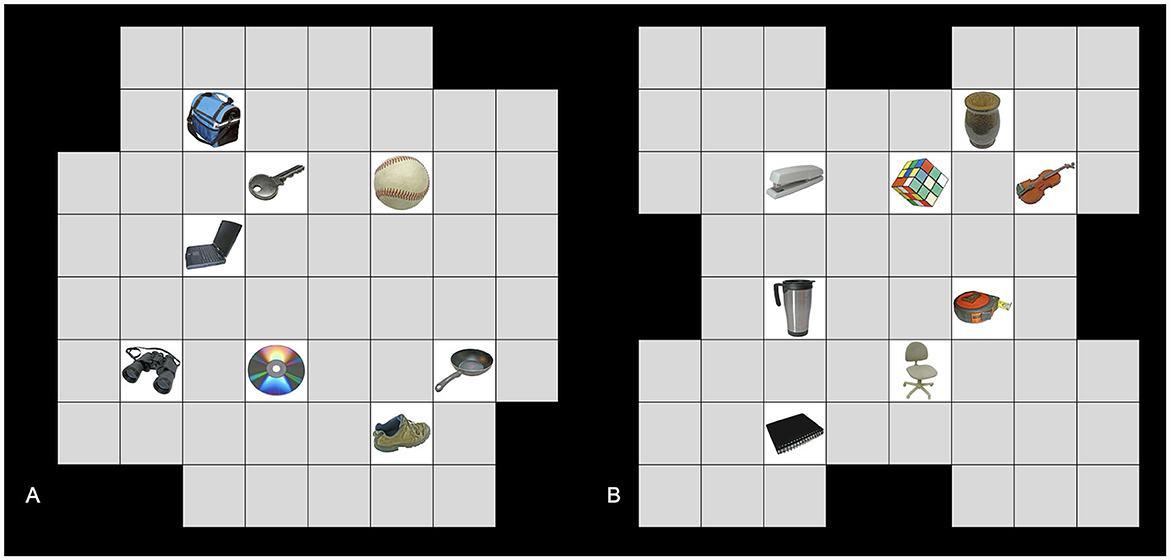
Figure 4. Example grids used in Experiment 2. (A) An example grid with one set of objects, one set of locations, and the “t” grid shape. (B) An example grid with the other set of objects, the other set of locations, and the “x” grid shape. Images used in this figure are unaltered images from the BOSS stimulus sets (Brodeur et al., 2010, 2014) used under the Creative Commons Attribution License (https://creativecommons.org/licenses/by-sa/3.0/).
3.1.2.3 Grid learning
The learning procedure was similar but not identical to Experiment 1. The grids were displayed on screen for 6 s instead of 8 s, and the maximum number of attempts to learn each grid was increased from 8 to 10. We also included an additional exclusion criterion based on the data from the online sample from Experiment 1. We noticed that several participants learned the first grid in a single trial. While it is possible for participants with exceptional memory abilities to achieve such a feat, it is unlikely and had not occurred in the in-person sample. We suspected that participants may be taking a picture of the grid and using that to reconstruct the grid. Thus, we excluded from analyses any participants who learned the first grid in a single trial but verified that the pattern of the results would be the same regardless. Besides these changes, the rest of the task remained the same as in Experiment 1.
3.2 Results
First, we measured the number of trials to learn the first grid (M = 3.85, SD = 1.57). Since this grid was learned prior to experimental manipulation, we predicted no significant differences between groups. To test this, we ran a 2 x 2 x 2 (object overlap x location overlap x grid shape) factorial ANOVA with the number of trials to learn the first grid as the dependent variable (Figure 5A; Table 1). As expected, we found no significant main effects or interactions, confirming that there were no between-group differences prior to experimental manipulation.
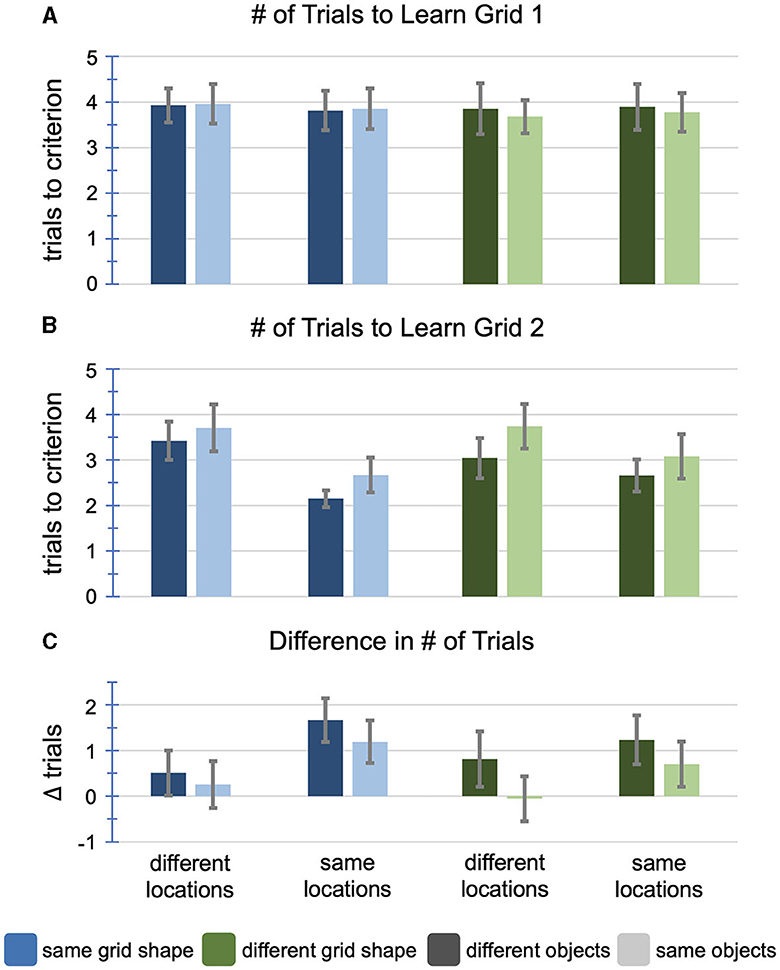
Figure 5. Results of Experiment 2. (A) The number of trials to learn the first grid by condition, prior to any manipulation. There were no significant differences. (B) The number of trials to learn the second grid by condition, after manipulation. There was a main effect of object overlap (p = 0.002), a main effect of location overlap (p < 0.001), and an interaction between location overlap and grid shape (p = 0.039). (C) The difference between the number of trials to learn the two grids (grid 1–grid 2), accounting for individual differences. There was a main effect of object overlap (p = 0.005), and a main effect of location overlap (p < 0.001). Error bars represent 95% confidence intervals for all plots.

Table 1. Experiment 2 ANOVA table with the number of trials to learn grid 1 as the dependent variable and location overlap, object overlap, and grid shape overlap as independent variables.
Next, we measured the number of trials to learn the second grid (M = 3.07, SD = 1.58). We hypothesized that object overlap would cause interference, resulting in a greater number of trials to learn the second grid, while location overlap would cause facilitation, resulting in fewer trials to learn the second grid. We did not have a strong prediction regarding the effect of grid shape. However, we were interested in the interaction between object overlap and grid shape. If a change in grid shape resulted in a release from proactive interference, we would expect to see an interaction between object overlap and grid shape. However, if proactive interference persists across grid shapes, then we would not expect to find an interaction. We were also interested in the interaction between location overlap and grid shape. If a change in grid shape causes spatial remapping, we would expect to see an interaction between location overlap and grid shape. However, if the facilitatory effect of location overlap persists across grid shapes, then it suggests that participants remember the overall configuration of locations rather than the locations with respect to the grid edges. To test these hypotheses, we ran a 2 x 2 x 2 (object overlap x location overlap x grid shape) factorial ANOVA with the number of trials to learn the second grid as the dependent variable (Figure 5B; Table 2). Consistent with Experiment 1, we found a main effect of object overlap, driven by fewer trials required to learn the second grid when objects did not overlap (M = 2.82, SD = 1.39) compared to when objects did overlap (M = 3.34, SD = 1.72). We also found a main effect of location overlap, driven by fewer trials required to learn the second grid when locations did overlap (M = 2.60, SD = 1.24) compared to when locations did not overlap (M = 3.48, SD = 1.73). We found no main effect of grid shape. However, we found a significant interaction between location overlap and grid shape, such that the magnitude of the facilitatory effect of location overlap was larger when the grid shape remained the same, but smaller when the grid shape changed.
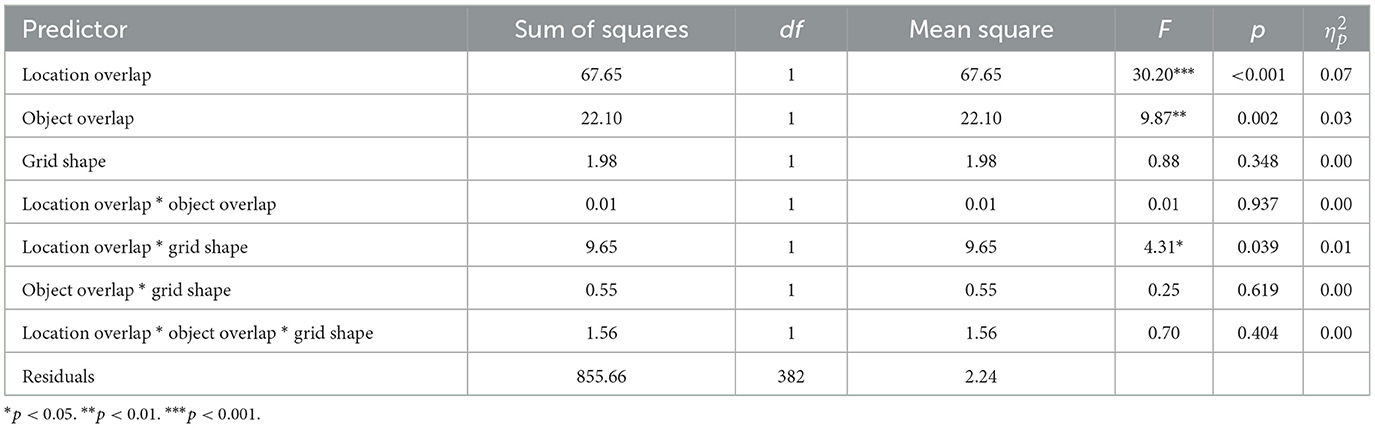
Table 2. Experiment 2 ANOVA table with the number of trials to learn grid 2 as the dependent variable and location overlap, object overlap, and grid shape overlap as independent variables.
To ensure that these effects were not driven by individual differences in grid learning ability, we calculated the difference in the number of trials to learn the two grids by subtracting the number of trials to learn the second grid from the number of trials to learn the first grid, as in Experiment 1. Averaged across conditions, participants showed practice effects, learning the second grid nearly one trial faster than the first (M = 0.78, SD = 1.90). Overall, 56% of participants improved from the first grid to the second grid. However, the magnitude of the practice effect varied across conditions. We ran a 2 x 2 x 2 (object overlap x location overlap x grid shape) factorial ANOVA with the difference score as the dependent variable (Figure 5C; Table 3). We replicated the main effect of object overlap, driven by greater improvement across grids when objects did not overlap (M = 1.05, SD = 1.96) compared to when objects did overlap (M = 0.47, SD = 1.78). When objects did not overlap, 61% of participants improved, learning the second grid in fewer trials than the first, while only 50% of participants improved when objects did overlap, = 4.58, p = 0.032. We also replicated the main effect of location overlap, driven by greater improvement across grids when locations did overlap (M = 1.23, SD = 1.75) compared to when locations did not overlap (M = 0.37, SD = 1.94). When locations did overlap, 66% of participants improved, learning the second grid in fewer trials, while only 46% of the participants improved when locations did not overlap, = 15.34, p < 0.001. Controlling for individual differences, we no longer found a significant interaction between location overlap and grid shape, suggesting that the facilitatory effect of location overlap persisted across grid shapes. The magnitude of the effect was still numerically larger when the grid shape remained the same (d = 0.57) compared to when the grid shape changed (d = 0.32; Figure 5C), but post-hoc comparisons between location overlap and no location overlap for both the same grid shape and different grid shapes reached significance. We again found no main effect of grid shape and no other interactions. These comparisons replicated findings from Experiment 1 that location overlap leads to facilitation of learning the second grid while object overlap hinders learning of the second grid. In addition, they demonstrate that these effects remain largely stable across grid shapes.
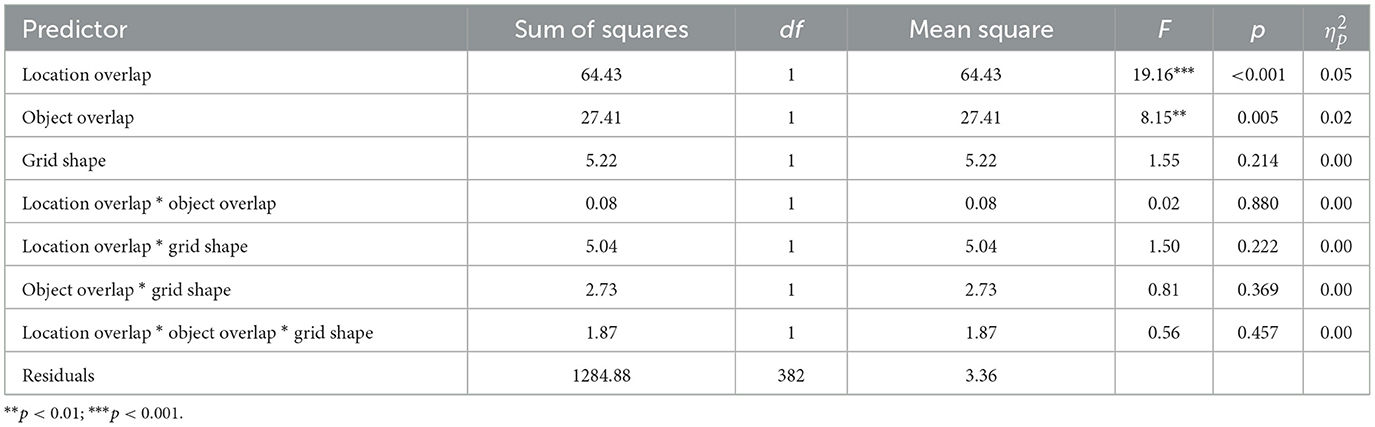
Table 3. Experiment 2 ANOVA table with the difference in the number of trials to learn the two grids as the dependent variable and location overlap, object overlap, and grid shape overlap as independent variables.
3.3 Discussion
In Experiment 2, we aimed to replicate the results of Experiment 1 and test the degree to which the effect of location and content overlap are modulated when the overall shape of the grid is altered. First, the results largely replicated Experiment 1. Location overlap provided clear facilitation of learning the second grid. The interference effect driven by object overlap remained relatively small with at 0.02–0.03, but its replication across two studies indicates it is statistically reliable. Furthermore, in both Experiment 1 and Experiment 2, about 10% fewer participants improved across grids when objects overlapped. Thus, the interference effects can be seen in both quantitative (mean number of trials) and qualitative (improvement or not) measures of learning.
Experiment 2 also explored to what degree these effects are modulated by or robust to a change of the overall grid shape. We found that the shift in grid shape did not eliminate the proactive interference observed due to object overlap. This indicates that changing the grid shape was not comparable to shifts in context used in prior studies. When controlling for individual variability, we also found that the shift in grid shape did not eliminate the facilitatory effect of location overlap. This suggests that the change in grid shape did not induce remapping in our participants, since the spatial pattern was similarly beneficial even in a new grid shape. Thus, we replicated our findings from Experiment 1 while also demonstrating that these effects generalize across grid shapes.
4 General discussion
Prior work has shown both beneficial and detrimental effects of information overlap on new learning. Here, we tested whether both consequences of overlap can be observed in a single task by manipulating two types of overlap: spatial (locations) and content (objects) in an object-location associative learning task. Across two experiments, we demonstrated that location overlap facilitates new learning while object overlap hinders new learning. When controlling for individual variability by focusing on the change in the number of trials one needs to learn two grids, we found in Experiment 2 that these effects were largely preserved even when the grid shape changed. Taken together, our findings identify a key factor that determines the behavioral consequences of overlap and indicate a differential role of what information and where information in memory organization. Associating a second object with one location was easy for participants; associating a second location with one object presented a challenge.
Critically, our study is one of the first to dissociate the effects of these two distinct types of information overlap on new learning within the same task. The AB/AC task consisting of overlapping word-word or image-image associations is a classic paradigm for demonstrating interference resulting from overlap (for a review, see Kliegl and Bäuml, 2021). Recently, this paradigm was used to examine both memory enhancement and impairment by manipulating context using a background scene (Cox et al., 2021). In the present study, we modified this paradigm to use object-location associations and demonstrated that the detrimental effect of overlap is not universal. Consistent with prior work, when the overlapping (A) items were objects, new learning was hindered. This effect is consistent with interference theory: overlapping objects (what information) caused proactive interference. Here, we newly demonstrate that the detrimental effect of content overlap generalizes to a situation that requires associating content with their spatial locations. However, when the overlapping (A) information involved locations (where information), new learning was facilitated rather than impaired. Thus, it is possible to manipulate the direction of the effect of overlap (facilitation or interference) within a single task by varying the type of information–content or spatial context–that overlaps across experiences.
Inspired by research on spatial and conceptual schemas, we hypothesized that location overlap would facilitate new learning. We indeed found a strong facilitation of subsequent learning by location overlap, despite our task differing in several ways from traditional schema research. While further research is needed to determine the mechanisms underpinning our findings, here we speculate that given the fixed relationship of the locations to each other in space, participants may form a generalized representation, or schema, of the location information. The object information can then be mapped onto this representation. During learning of the second grid in the location overlap condition, participants benefit from being able to “slot” new objects into the same spatial schema, similarly to how method of loci benefits memory for new material by associating it with familiar spatial locations (Ross and Lawrence, 1968; Wang and Thomas, 2000). This interpretation aligns with other work showing that spatial schemas can facilitate rapid new learning, as demonstrated in rodents by Tse et al. (2007, 2011) and in humans by van Buuren et al. (2014). The present study expands on this prior work by showing that memories of spatial configurations, which may be early formations of spatial schemas, also facilitate new learning even when all objects are swapped out. This argument is strengthened by the results of Experiment 2. The facilitatory effect of location overlap was preserved across two distinct grid shapes, suggesting that participants encoded the configural organization of the target locations rather than individual locations with respect to the grid border. Thus, the spatial pattern itself may serve as the context, or schema.
The work on spatial schemas served as a key motivation for our study and our predictions regarding facilitation driven by location overlap. However, one concern regarding interpretation of our data in the schema framework is the speed at which a spatial schema can be learned. In prior work (Tse et al., 2007; van Buuren et al., 2014), participants learned the spatial schemas over multiple sessions spanning days or weeks. In a more extreme case, Sommer (2016) utilized a 302-day study design in which participants learned a semantic schema. Despite the long-term acquisition phases used in these studies, more recent work has suggested that schemas may actually be learned quite quickly (Tompary et al., 2020). However, we typically rely on episodic memories until those memories fade, at which time we begin to rely on schemas and generalized memories. This leads us to believe that in the present study, participants may be acquiring a spatial schema while learning the first grid, and this potential schema facilitates their learning of the second grid when the locations, and thus the spatial schema, overlap. Of course, one may also consider the benefits of remembering the spatial configuration of locations without considering such a configuration to be a spatial schema. Future research is needed to determine if the current findings are best understood in terms of spatial schema or rather reflect some other process. Nonetheless, these findings provide further evidence that spatial information holds a unique role in memory organization (Arnold et al., 2011; Robin et al., 2016; Sheldon and Chu, 2017).
One limitation of the current paradigm is the location-emphasizing test response modality, asking participants to drag and drop provided objects to their location. Although this does not explain the opposing effects of content and location overlap, it may have led to extra-large benefit of location overlap (greatly reducing the number of locations that need to be considered) that may become more modest with a different type of object-location association test. Thus, future research should replicate the current findings with a wider range of memory probes. Another finding that should be revisited in future studies is the preservation of the facilitation and interference effects across a change in the overall grid shape. Participants may have been less affected by the change in grid shape in the current study because no location adjacent to the border was used for object placement, making it more likely that participants encode the overall spatial configuration of locations with limited reference to the grid border. It is possible that a stronger interaction with grid shape would be observed if border locations were used, especially if a location were to touch the border in one grid but not the other.
The interference and facilitation effects of object and location overlap observed here may also inform conflicting theories regarding how overlapping events are represented in the hippocampus, a key memory structure, to support memory. Facilitatory effects of overlap are thought to stem from incorporating new memories into existing memory representations through memory integration (Zeithamova et al., 2012) and schema-related memory (Preston and Eichenbaum, 2013; Gilboa and Marlatte, 2017; Guo and Yang, 2020). As such, increased similarity of neural representations leads to better memory of overlapping events (Tse et al., 2007; Schlichting et al., 2014; Takeuchi et al., 2022). In contrast, interference research has proposed an opposite representational strategy—minimizing similarity of neural representations—leading to better memory of overlapping events. Specifically, hippocampal pattern separation (Lohnas et al., 2018; for a review, see Yassa and Stark, 2011) or repulsion (Favila et al., 2016; Chanales et al., 2017, 2021) improves memory by representing similar events by distinct neural codes to minimize interference. The current findings suggests that both integration and separation may benefit memory depending on behavioral consequences of overlap, leading to intriguing questions for future research regarding how the hippocampus will respond when faced with both the positive and negative effects of overlap in the same task.
In summary, we developed a novel paradigm where both positive and negative effects of information overlap on new learning can be demonstrated within the same task. We demonstrated that a key factor determining the behavioral consequences of overlap is the nature of the overlap: location overlap led to facilitation, object overlap led to interference. Additionally, these effects persisted across two distinct grid shapes. These findings help reconcile prior conflicting research on how overlap affects memory and offer a new paradigm to study both facilitation and interference within the same task. More broadly, the opposing effects of object overlap and location overlap demonstrate qualitative differences in the processing of content and spatial information, informing theories on the distinct roles of what and where in the organization of memory.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: Open Science Framework https://osf.io/jypnk/.
Ethics statement
The studies involving humans were approved by Research Compliance Services at the University of Oregon. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
BC: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Software, Validation, Visualization, Writing—original draft. DZ: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing—review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Institute of Neurological Disorders and Stroke Grant R01-NS112366 awarded to DZ.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adam, K. C. S., Mance, I., Fukuda, K., and Vogel, E. K. (2015). The contribution of attentional lapses to individual differences in visual working memory capacity. J. Cogn. Neurosci. 27, 1601–1616. doi: 10.1162/jocn_a_00811
Arnold, K. M., McDermott, K. B., and Szpunar, K. K. (2011). Imagining the near and far future: the role of location familiarity. Mem. Cognit. 39, 954–967. doi: 10.3758/s13421-011-0076-1
Attneave, F. (1957). Transfer of experience with a class-schema to identification-learning of patterns and shapes. J. Exp. Psychol. 54, 81–88. doi: 10.1037/h0041231
Bartlett, F. C. (1932). Remembering: A Study in Experimental and Social Psychology. Cambridge: Cambridge University Press.
Brodeur, M. B., Dionne-Dostie, E., Montreuil, T., and Lepage, M. (2010). The bank of standardized stimuli (BOSS), a new set of 480 normative photos of objects to be used as visual stimuli in cognitive research. PLoS ONE 5:e10773. doi: 10.1371/journal.pone.0010773
Brodeur, M. B., Guérard, K., and Bouras, M. (2014). Bank of standardized stimuli (BOSS) phase II: 930 new normative photos. PLoS ONE 9:e106953. doi: 10.1371/journal.pone.0106953
Bunting, M. (2006). Proactive interference and item similarity in working memory. J. Exp. Psychol. Learn. Mem. Cogn. 32, 183–196. doi: 10.1037/0278-7393.32.2.183
Caplan, J. B., Hennies, N., and Sommer, T. (2022). Competition between associations in memory. J. Cogn. Neurosci. 34, 2144–2167. doi: 10.1162/jocn_a_01900
Chanales, A. J. H., Oza, A., Favila, S. E., and Kuhl, B. A. (2017). Overlap among spatial memories triggers repulsion of hippocampal representations. Curr. Biol. 27, 2307–2317.e5. doi: 10.1016/j.cub.2017.06.057
Chanales, A. J. H., Tremblay-McGaw, A. G., Drascher, M. L., and Kuhl, B. A. (2021). Adaptive repulsion of long-term memory representations is triggered by event similarity. Psychol. Sci. 32, 705–720. doi: 10.1177/0956797620972490
Cox, W. R., Dobbelaar, S., Meeter, M., Kindt, M., and Van Ast, V. A. (2021). Episodic memory enhancement versus impairment is determined by contextual similarity across events. Proc. Natl. Acad. Sci. 118:e2101509118. doi: 10.1073/pnas.2101509118
Daneman, M., and Carpenter, P. A. (1980). Individual differences in working memory and reading. J. Verb. Learn. Verbal Behav. 19, 450–466. doi: 10.1016/S0022-5371(80)90312-6
Favila, S. E., Chanales, A. J. H., and Kuhl, B. A. (2016). Experience-dependent hippocampal pattern differentiation prevents interference during subsequent learning. Nat. Commun. 7, 11066. doi: 10.1038/ncomms11066
Gilboa, A., and Marlatte, H. (2017). Neurobiology of schemas and schema-mediated memory. Trends Cogn. Sci. 21, 618–631. doi: 10.1016/j.tics.2017.04.013
Goggin, J., and Wickens, D. D. (1971). Proactive interference and language change in short-term memory. J. Verbal Learn. Verbal Behav. 10, 453–458. doi: 10.1016/S0022-5371(71)80046-4
Guo, D., and Yang, J. (2020). Interplay of the long axis of the hippocampus and ventromedial prefrontal cortex in schema-related memory retrieval. Hippocampus 30, 263–277. doi: 10.1002/hipo.23154
King, B. R., Dolfen, N., Gann, M. A., Renard, Z., Swinnen, S. P., and Albouy, G. (2019). Schema and motor-memory consolidation. Psychol. Sci. 30, 963–978. doi: 10.1177/0956797619847164
Kliegl, O., and Bäuml, K. -H. T. (2021). Buildup and release from proactive interference – cognitive and neural mechanisms. Neurosci. Biobehav. Rev. 120, 264–278. doi: 10.1016/j.neubiorev.2020.10.028
Kuhl, B. A., Dudukovic, N. M., Kahn, I., and Wagner, A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. Nat. Neurosci. 10, 908–914. doi: 10.1038/nn1918
Kuhl, B. A., Rissman, J., Chun, M. M., and Wagner, A. D. (2011). Fidelity of neural reactivation reveals competition between memories. Proc. Natl. Acad. Sci. 108, 5903–5908. doi: 10.1073/pnas.1016939108
Lever, C., Wills, T., Cacucci, F., Burgess, N., and O'Keefe, J. (2002). Long-term plasticity in hippocampal place-cell representation of environmental geometry. Nature 416, 90–94. doi: 10.1038/416090a
Lohnas, L. J., Duncan, K., Doyle, W. K., Thesen, T., Devinsky, O., and Davachi, L. (2018). Time-resolved neural reinstatement and pattern separation during memory decisions in human hippocampus. Proc. Natl. Acad. Sci. 115, E7418–E7427. doi: 10.1073/pnas.1717088115
Maslow, A. H. (1934). The effect of varying time intervals between acts of learning with a note on proactive inhibition. J. Exp. Psychol. 17, 141–144. doi: 10.1037/h0069853
Melton, A. W., and von Lackum, W. J. (1941). Retroactive and proactive inhibition in retention: evidence for a two-factor theory of retroactive inhibition. Am. J. Psychol. 54, 157. doi: 10.2307/1416789
Posner, M. I., Goldsmith, R., and Welton, K. E. (1967). Perceived distance and the classification of distorted patterns. J. Exp. Psychol. 73, 28–38. doi: 10.1037/h0024135
Postman, L., Stark, K., and Burns, S. (1974). Sources of proactive inhibition on unpaced tests of retention. Am. J. Psychol. 87, 33. doi: 10.2307/1421998
Preston, A. R., and Eichenbaum, H. (2013). Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 23, R764–R773. doi: 10.1016/j.cub.2013.05.041
Robin, J., Wynn, J., and Moscovitch, M. (2016). The spatial scaffold: the effects of spatial context on memory for events. J. Exp. Psychol. Learn. Mem. Cogn. 42, 308–315. doi: 10.1037/xlm0000167
Ross, J., and Lawrence, K. A. (1968). Some observations on memory artifice. Psychon. Sci. 13, 107–108. doi: 10.3758/BF03342433
Schlichting, M. L., Zeithamova, D., and Preston, A. R. (2014). CA1 subfield contributions to memory integration and inference. Hippocampus 24, 1248–1260. doi: 10.1002/hipo.22310
Sheldon, S., and Chu, S. (2017). What versus where: investigating how autobiographical memory retrieval differs when accessed with thematic versus spatial information. Q. J. Exp. Psychol. 70, 1909–1921. doi: 10.1080/17470218.2016.1215478
Sommer, T. (2016). The emergence of knowledge and how it supports the memory for novel related information. Cereb. Cortex 27, 1906–1921. doi: 10.1093/cercor/bhw031
Takeuchi, T., Tamura, M., Tse, D., Kajii, Y., Fernández, G., and Morris, R. G. M. (2022). Brain region networks for the assimilation of new associative memory into a schema. Mol. Brain 15, 24. doi: 10.1186/s13041-022-00908-9
Tompary, A., Zhou, W., and Davachi, L. (2020). Schematic memories develop quickly, but are not expressed unless necessary. Sci. Rep. 10, 16968. doi: 10.1038/s41598-020-73952-x
Tse, D., Langston, R. F., Kakeyama, M., Bethus, I., Spooner, P. A., Wood, E. R., et al. (2007). Schemas and memory consolidation. Science 316, 76–82. doi: 10.1126/science.1135935
Tse, D., Takeuchi, T., Kakeyama, M., Kajii, Y., Okuno, H., Tohyama, C., et al. (2011). Schema-dependent gene activation and memory encoding in neocortex. Science 333, 891–895. doi: 10.1126/science.1205274
Tulving, E., and Watkins, M. J. (1974). On negative transfer: effects of testing one list on the recall of another. J. Verbal Learn. Verbal Behav. 13, 181–193. doi: 10.1016/S0022-5371(74)80043-5
Unsworth, N., Brewer, G. A., and Spillers, G. J. (2013). Focusing the search: proactive and retroactive interference and the dynamics of free recall. J. Exp. Psychol. Learn. Mem. Cogn. 39, 1742–1756. doi: 10.1037/a0033743
Unsworth, N., Heitz, R. P., Schrock, J. C., and Engle, R. W. (2005). An automated version of the operation span task. Behav. Res. Methods 37, 498–505. doi: 10.3758/BF03192720
van Buuren, M., Kroes, M. C. W., Wagner, I. C., Genzel, L., Morris, R. G. M., and Fernandez, G. (2014). Initial investigation of the effects of an experimentally learned schema on spatial associative memory in humans. J. Neurosci. 34, 16662–16670. doi: 10.1523/JNEUROSCI.2365-14.2014
van Kesteren, M. T. R., Rijpkema, M., Ruiter, D. J., Morris, R. G. M., and Fernández, G. (2014). Building on prior knowledge: schema-dependent encoding processes relate to academic performance. J. Cogn. Neurosci. 26, 2250–2261. doi: 10.1162/jocn_a_00630
van Kesteren, M. T. R., Ruiter, D. J., Fernández, G., and Henson, R. N. (2012). How schema and novelty augment memory formation. Trend. Neurosci. 35, 211–219. doi: 10.1016/j.tins.2012.02.001
Verguts, T., and Boeck, P. D. (2002). The induction of solution rules in Raven's progressive matrices test. Eur. J. Cogn. Psychol. 14, 521–547. doi: 10.1080/09541440143000230
Wahlheim, C. N., and Jacoby, L. L. (2013). Remembering change: the critical role of recursive remindings in proactive effects of memory. Mem. Cogn. 41, 1–15. doi: 10.3758/s13421-012-0246-9
Wang, A. Y., and Thomas, M. H. (2000). Looking for long-term mnemonic effects on serial recall: the legacy of simonides. Am. J. Psychol. 113, 331. doi: 10.2307/1423362
Wanjia, G., Favila, S. E., Kim, G., Molitor, R. J., and Kuhl, B. A. (2021). Abrupt hippocampal remapping signals resolution of memory interference. Nat. Commun. 12, 4816. doi: 10.1038/s41467-021-25126-0
Wickens, D. D., Born, D. G., and Allen, C. K. (1963). Proactive inhibition and item similarity in short-term memory. J. Verbal Learn. Verbal Behav. 2, 440–445. doi: 10.1016/S0022-5371(63)80045-6
Wills, T. J., Lever, C., Cacucci, F., Burgess, N., and O'Keefe, J. (2005). Attractor dynamics in the hippocampal representation of the local environment. Science 308, 873–876. doi: 10.1126/science.1108905
Wixted, J. T., and Rohrer, D. (1993). Proactive interference and the dynamics of free recall. J. Exp. Psychol. Learn. Mem. Cogn. 19, 1024–1039. doi: 10.1037/0278-7393.19.5.1024
Yassa, M. A., and Stark, C. E. L. (2011). Pattern separation in the hippocampus. Trend. Neurosci. 34, 515–525. doi: 10.1016/j.tins.2011.06.006
Keywords: learning, memory, schema, interference, facilitation, overlap
Citation: Chaloupka B and Zeithamova D (2024) Differential effects of location and object overlap on new learning. Front. Cognit. 2:1325246. doi: 10.3389/fcogn.2023.1325246
Received: 20 October 2023; Accepted: 27 December 2023;
Published: 29 January 2024.
Edited by:
David Clewett, University of California, Los Angeles, United StatesReviewed by:
Yu Liu, University of Minnesota, United StatesBenjamin Jacob Tamber-Rosenau, University of Houston, United States
Copyright © 2024 Chaloupka and Zeithamova. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dagmar Zeithamova, dasa@uoregon.edu