European Blackbirds Exposed to Aircraft Noise Advance Their Chorus, Modify Their Song and Spend More Time Singing
 Javier Sierro
Javier Sierro Elodie Schloesing1
Elodie Schloesing1  Diego Gil
Diego Gil- 1Departamento de Ecología Evolutiva, Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain
- 2Grupo de Investigación en Instrumentación y Acústica Aplicada, Universidad Politécnica de Madrid, Madrid, Spain
Noise pollution has a strong impact on wildlife by disrupting vocal communication or inducing physiological stress. Songbirds are particularly reliant on vocal communication as they use song during territorial and sexual interactions. Birds living in noisy environments have been shown to change the acoustic and temporal parameters of their song presumably to maximize signal transmissibility. Also, research shows that birds advance their dawn chorus in urban environments to avoid the noisiest hours, but little is known on the consequences of these changes in the time they spent singing at dawn. Here we present a comprehensive view of the European blackbird singing behavior living next to a large airport in Madrid, using as a control a population living in a similar but silent forest. Blackbird song is composed of two parts: a series of loud low-frequency whistles (motif) and a final flourish (twitter). We found that airport blackbirds were more likely to sing songs without the twitter part. Also, when songs included a twitter part, airport blackbirds used a smaller proportion of song for the twitter than control blackbirds. Interestingly, our results show no differences in song frequency between airport and control populations. However airport blackbirds not only sang earlier but also increased the time they spent singing when chorus and aircraft traffic overlapped on time. This effect disappeared as the season progressed and the chorus and the aircraft traffic schedule were separated on time. We propose that the typical urban upshift in frequency might not be useful under the noise conditions and landscape structure found near airports. We suggest that the modifications in singing behavior induced by aircraft noise may be adaptive and that they are specific to airport acoustic habitat. Moreover, we found that adjustment of singing activity in relation to noise is plastic and possibly optimized to cope with aircraft traffic activity. In a soundscape characterized by intermittent and strong noise bursts, singing for longer could be more advantageous than modifying frequency parameters, although it is likely more costly.
Introduction
Birdsong is one of the most elaborate acoustic signals in the natural world. Like many other communication systems, birdsong is crucial for birds because it plays a key role in two very important aspects of reproduction: mate choice and territory defense (Catchpole and Slater, 2003). In temperate climates, males advertise their territory by singing from particular posts during the breeding season. Although it is common to hear birds singing throughout the day, many species concentrate their singing activity at dawn in what is known as dawn chorus (Kacelnik and Krebs, 1983; Catchpole and Slater, 2003). As a sound signal, birdsong has evolved under the selection pressures of the acoustic habitat in which the species thrives, such as the sound propagation properties of the habitat and its particular type of environmental noise (Morton, 1975; Ryan and Brenowitz, 1985; Hanna et al., 2011; Wiley, 2015). Environmental noise is present in every soundscape. Depending on its intensity it can distort the perception of the signal to a variable extent and reduce the number of receivers within the communication network (Hansen et al., 2005). Critically, worldwide urbanization leads to a remarkable increment of natural background noise together with other environmental factors that also hinder acoustic communication, such as the presence of vertical reflective surfaces or an increase in habitat fragmentation (Warren et al., 2006; Slabbekoorn and Ripmeester, 2008). Because of the important functions of song, as well as other vocal signals, increased environmental noise is likely to have a direct impact on fitness, posing a selection pressure over urban populations of birds (Slabbekoorn and Ripmeester, 2008).
Despite the altered conditions of the city soundscape, many bird species thrive in urban environments and cumulative evidence points out to the importance of behavioral plasticity in coping with noise (Ríos-Chelén et al., 2012; Shanahan et al., 2014). Regarding birdsong, studies have shown a general pattern of song modifications that appear in response to noise, such as upshifted frequency (Slabbekoorn and Peet, 2003; Nemeth and Brumm, 2009; Verzijden et al., 2010; Bermúdez-Cuamatzin et al., 2012), increased song amplitude (Cynx et al., 1998; Brumm, 2004; Brumm et al., 2009), modifications in temporal parameters of song (Slabbekoorn and den Boer-Visser, 2006; Díaz et al., 2011; Ríos-Chelén et al., 2013) or changes in the timing of the dawn chorus (Fuller et al., 2007; Arroyo Solís et al., 2013; Dominoni et al., 2016). In many bird species, it has been found that populations living near anthropogenic noise sources sing at higher frequencies, partly avoiding masking (Slabbekoorn and Peet, 2003; Nemeth and Brumm, 2009; Verzijden et al., 2010; Ríos-Chelén et al., 2012; Roca et al., 2016). Also, birds can increase the amplitude of their songs to counteract the detrimental effect of noise, thus improving the signal-to-noise ratio and the distance over which the signal can be perceived (Brumm and Todt, 2002; Brumm, 2004). On the other hand, those studies that have analyzed changes in the duration of songs in relation to increased background noise show inconclusive results: some species are found to sing shorter songs/syllables under noisy conditions (Slabbekoorn and den Boer-Visser, 2006; Verzijden et al., 2010; Francis et al., 2011), whereas other species increase song/syllable length (Nemeth and Brumm, 2009; Bermúdez-Cuamatzin et al., 2011; Ríos-Chelén et al., 2013). Thus, it is not yet clear the role that these changes of song duration play in noisy conditions, it may depend on the information that song length is coding in each species and the temporal characteristics of noise (intermittent vs. continuous; Gil and Brumm, 2013). A study with serins (Serinus serinus) showed an increase in time spent singing as noise levels increased but this relationship broke down at a 70 dBA threshold (Díaz et al., 2011). In contrast to direct song modifications, it is possible that increased time spent singing is a more efficient strategy to improve signal transmission without compromising the information coded by song length or syllable duration, although such a behavioral change is bound to be more costly in terms of energy balance (Thomas, 2002).
The high predictability of daily anthropogenic noise patterns allows birds to avoid temporal overlap with noisy hours by means of shifts in their singing activity. There is a growing body of evidence suggesting that birds advance the onset of dawn chorus when living in noisy conditions (Fuller et al., 2007; Gil et al., 2014; Dominoni et al., 2016; Dorado-Correa et al., 2016). The question remains open as to whether this shift of singing activity is a by-product of noise-induced perturbations on the birds' behavior, as suggested by some authors (Arroyo Solís et al., 2013), or an adaptive modification to counter act the effect of noise later on the day (Fuller et al., 2007; Dominoni et al., 2016). Taking an optimization approach and assuming that singing earlier has negative effects due to energy balance and predation (McNamara et al., 1987), it would be expected that birds should advance dawn chorus in order to avoid noisy conditions if this leads to improved signal transmission. In an earlier study (Gil et al., 2014), we suggested that the advance of singing activity at dawn was more relevant in those species whose dawn song was closer to the onset of aircraft traffic activity. On this respect, we should consider that the advance in dawn chorus depends on the amount of noise-chorus overlapping, which in turn is determined by at least three different factors: (1) human activity, which starts at a fixed time regardless of seasonal variations in daylight; (2) the circadian cycle of activity expressed by wild animals which, unlike humans, is synchronized with the natural cycles of sunlight; and (3) differences of the time of sunrise that varies in relation to latitude. Finally, it is yet to be confirmed whether or not an advance in the dawn chorus results in a prolonged singing activity during dawn, since such a change could carry heavier consequences in terms of time budgets.
European blackbirds (Turdus merula) belong to the thrushes family (Turdidae) and, although they are forest birds, they are also common urban dwellers (Evans et al., 2009). Blackbirds are among the earliest singers at dawn, with their first songs being heard 1 h before sunrise (Snow, 1958) although actual dawn chorus begins rather later around 30–45 min before sunrise (Ripmeester et al., 2010, pers. obs.), lasting for approximately 20–30 min (Snow, 1958). Males are territorial and will use several song posts to deliver their song (Cramp, 1992). Blackbird song comprises short phrases of few seconds that can be broke down in two parts, the motif and the twitter, although the twitter is not always present and some songs are built by entirely by motif notes. The motif is made of low frequency, high amplitude melodic notes (pure tones), whereas the twitter is a final flourish composed of a burst of short, higher frequency and low amplitude notes that cover a wider frequency spectrum (Dabelsteen, 1984). Previous studies of this species show that urban blackbirds sing higher frequency motif notes (Nemeth and Brumm, 2009; Ripmeester et al., 2010). Through an increase in the frequency of this typically low-frequency part, blackbirds may improve signal transmission by reducing masking with noise, although it has been argued that higher frequency motif notes are higher in amplitude too and that the later modification should be more relevant than frequency changes to improve transmission (Nemeth et al., 2013). Also, urban birds increase the proportion of their twitter within the song (Ripmeester et al., 2010). Even if this implies less spectral overlap with noise it might not be a functional noise-related modification since twitter elements are usually very weak in amplitude. In fact, because of the motivational role of the twitter during male-male interactions and its seasonal variation (increasing with date), it has been suggested that larger twitter proportions are due to the increased density of territories and the advanced breeding season in urban habitats (Dabelsteen, 1984; Ripmeester et al., 2010). Blackbirds have also been found to advance the onset of dawn song when living next to urban infrastructures such as cities or airports (Nordt and Klenke, 2013; Gil et al., 2014; Dominoni et al., 2016). In such cases, there is likely a compound effect of artificial lighting and anthropogenic noise, which are not always easy to tear apart. When acoustic and light pollution occur simultaneously, light seems to have a stronger impact over the advance of dawn song than noise (Nordt and Klenke, 2013; Da Silva et al., 2016), but birds also advance dawn chorus near large airports in places where artificial lighting is absent (Dominoni et al., 2014, 2016; Gil et al., 2014).
In the present study, we aimed to have a comprehensive view of the impact of aircraft traffic noise on the general singing behavior of the blackbird. Firstly, we focused on noise-induced modifications in the acoustic and temporal characteristics of song, comparing songs recorded from a blackbird population living near Madrid airport with songs of a population distant from the airport. It is important to underline the fact that both populations live in forest habitats with low urbanization of the landscape and that noise is the only important difference between sites as far as we could determine (Gil et al., 2014). We predicted that songs of airport blackbirds would be higher in frequency than those from the control population, following previous findings and to the extent that effects of road traffic noise are similar to aircraft noise. Although aircraft noise may show similar acoustic spectral characteristics, it has definitely different temporal characteristics. The temporal structure of aircraft noise events are usually louder, and more spaced over time, leaving silent intervals in between (Dominoni et al., 2016), in contrast to the continuous emission of car traffic noise (Arroyo Solís et al., 2013). Secondly, we investigated the onset and duration of dawn chorus. We predicted that, if the advance of dawn chorus allows the bird to avoid high levels of noise during rush hours, airport blackbirds should advance dawn chorus at the particular moment of the breeding season when natural singing time overlaps with aircraft traffic, whereas this advance would not be expected when the overlap is absent due to earlier sun rising hours. We believe that our set-up is suitable to investigate this prediction because strict aircraft traffic regulations and schedules lead to a predictable noise levels clearly separated on time of aircraft noise levels (http://sicaweb.cedex.es/ume-fase1.php?id=65) compared to the more diffuse and flexible dawn noise patterns in cities (cfr. Warren et al., 2006).
Materials and Methods
Study Species and Study Site
The European blackbird is a widespread and abundant species all over the Western Palearctic. Thanks to its diverse habitat use, blackbird populations are found in high densities both in urban and non-urban environments making this species an excellent model to understand ecological and behavioral adaptations to the urban life (Partecke et al., 2006; Ripmeester et al., 2010; Mendes et al., 2011; Nemeth et al., 2013). In this study, we have focused on two blackbird populations living in a riparian forest along the Jarama river near Madrid. The “airport population” is located in the proximity of two runways of the Madrid-Barajas airport running parallel to the river forest. This population is affected by two airport runways; a distant one approximately 1.5 km away from the river forest (runway number 3), and a close-by runway, hardly 200 m from the edge of the forest (runway number 4) though this distance varies along the runway. The second population is located approx. 10 km northwards within the same strip of river forest, hereafter “control population” (see Figure 1 in Gil et al., 2014). The two habitats are similar (Gil et al., 2014), and consist of meadows and a gallery forest running along the river and ponds and composed mainly of willow trees, ash trees, alder trees and poplars. Although the control study site receives a small impact of aircraft traffic, it is negligible in comparison to the airport population (see below and Results for direct data on this difference). Moreover, a previous study in the same areas showed that there were no differences in light pollution between the two sites (Gil et al., 2014). For an aerial view of the study area please see Gil et al. (2014).
Noise Measurements
Noise maps provided by the airport authorities show a clear difference in average noise levels (2008 data: http://sicaweb.cedex.es/ume-fase1.php?id=65). The blackbird song posts near the airport that we included in this study were all located within the 65–75 Lden isophones, while the control area falls entirely inside the 55–60 Lden isophones. However, these maps do not show environmental noise levels from all sound sources -total sound-, only the equivalent noise contribution by aircraft events—specific sound—(International Organization for Standardization, 2016). To obtain direct information of the anthropogenic noise impact of both sites we collected our own data in the field.
We used recordings made by autonomous recording units (Song Meter™: model SM2 Wildlife Acoustics) to monitor the acoustic habitat at each site. The recorders were deployed in the field using a custom-made harness to hang them from a tree branch at a height of 4–5 m. We collected data during 6 days from the 12th to the 18th of March 2016 in the center of each study site. Song meters were programmed to record during 3 h (5:00 a.m.–8:00 a.m.) alternating periods of 10 min of recording and 10 min of inactivity. Aircraft traffic is restricted to two of the four runways during the night time (from 23 to 7 h a.m.), and it is reestablished to all runways during day time (from 7 a.m. to 23.h). On runway 3, aircraft activity is allowed during night time but it mostly takes place after 6 a.m. and is limited to certain aircraft which do not surpass a threshold of noise. For runway 4, the aircraft activity is allowed during day time with an onset time at 7 a.m. Hence, from 5 to 8 a.m. there is an increase in noise in three steps that represent the three characteristic noise conditions of the study site. For this description we defined three time intervals regarding the state of aircraft traffic; “no traffic” is the time interval from 5 to 6 a.m. when there is nearly no traffic activity, “traffic runway 3” defining the time interval from 6 to 7 a.m., when aircraft activity occurs only in the more distant runway and “traffic runway 4” when aircrafts can take off from both runways. In the lab, we used the methodology described by Merchant et al. (2015) to measure equivalent continuous sound levels from .wav recordings using PAMGuide functions in R software version 3.3.1 (Team, 2012). We used the following technical specifications for the calibration of microphone and recorder: sensitivity of transducer = 36 mV/Pa, zero-to-peak voltage of the analog-to-digital converter = 1.414 mV and a gain of 27 dB and we averaged the sound pressure level to a resolution of 15 s via the Welch method. Based on this calibration, we used the PAMGuide function to analyse the entire 3 h recordings of each day and obtain absolute decibel levels with zero weighting setting atype as “Broadband.” We calculated the logarithmic average of broadband decibel levels per interval of aircraft activity (Figure 2). Furthermore we carried out a finer analysis per 1/3-octave band levels using the same function PAMGuide in the package but setting atype to “TOL” (Third Octave Levels) and later calculating the mean decibel levels of each frequency band of the 5 days sampled (Figure 3). We performed the description of the energy distribution of noise along the frequency spectrum in order to explore possible interactions of aircraft noise and vocal communication in birds.
High Quality Recordings and Song Analyses
By mid-February, we began to visit the study sites in order to map blackbird individuals' song-posts. We heard the first blackbirds singing during the first week of March (March the 2nd in the control site and March the 5th in the airport site). Full blackbird dawn chorus began a few days later around the 15th of March. During dawn, we identified the singing locations of individual male blackbirds by listening to neighboring males singing simultaneously. When detected, song posts were visited again over several days for confirmation. Once we mapped the song posts, we collected high quality song recordings during dawn chorus using a Marantz Professional PMD661 handheld solid state recorder and a Parabolic Microphone Telinga MK2 (sampling rate: 48 kHz, 24 bits). Each day, we targeted only one or two individuals and recorded around 15 min of song. We alternated daily between the control and the airport sites to avoid any seasonal effect in singing behavior interfering with potential population differences. In total we collected high quality recordings for 32 individuals, a balanced sample of 16 males per study site. In the lab, we selected 10 songs per male recorded with no aircraft noise in the background; songs that overlapped with aircraft noise were not amenable to acoustic analysis.
In the lab, the first step of song analysis was a visual inspection of the spectrogram using Audacity software (FFT-window length: 512, window type: Hanning, frequency limits: 0–10 kHz). During this step we took manual measurements using the cursor over the spectrogram for song length, singing effort (ratio between the sum song length of 10 songs divided time elapsed from the first to the last song), motif length, twitter length, twitter proportion (ratio between the length of the twitter part divided by song length), and note to silence ratio within song (the ratio between the sum of note durations and song length) (see Figure 1). Every note was categorized as a motif or a twitter note. After inspection of spectrograms, wav files were loaded into R for acoustic analysis using the “seewave” package (Sueur et al., 2006). First, we applied a high pass filter under 1 kHz (ffilter function) to remove background low frequency noise. Secondly, using previous manual measurements, we cut out every song from the original recording and exported them as individual wav files using the savewav function in seewave. By default, this function normalizes the amplitude of the wave to a numeric value ranging from −1 to 1. We then cut and saved separately every single note within each song (same procedure) and performed an acoustic analysis of each note extracting dominant frequency, which is the maximum amplitude frequency at each time point (dfreq function), FFT-window length: 512, window type: Hanning, window overlap: 50%, 5% amplitude threshold for signal detection. Dominant frequency function gives a frequency contour in the form of a two-column matrix corresponding to time and maximum amplitude frequency at each window point. From this contour we obtained mean frequency and the maximum and minimum dominant frequencies of each note. Note that these measures do not represent maximum and minimum frequencies as used in most studies. We decided to use this measurement because it is a more objective measurement than a manual procedure using the cursor where the presence of background noise may introduce biases and thus enables us to compare two study populations with differential noise levels. We developed a short script in R that allowed us to batch-process the notes semi automatically, plotting and inspecting the measurements over the original spectrogram for every analysis to ensure they were correct. If loud signals (commonly other bird's song or call) overlapped temporally but not spectrally with the blackbird note, we applied a band pass excluding this sound from signal detection. We dismissed any note if there was any sound artifact interfering with the detection of dominant frequency.
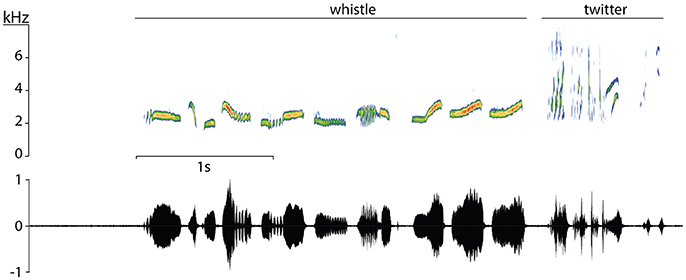
Figure 1. Song of an European blackbird. In the spectrogram (Top) the motif and twitter notes are shown with their relative amplitude represented by differences in color (blue lowest, red highest). Below the spectrogram, the oscillogram of the same song shows the relative differences in amplitude.
Automatic Recordings and Dawn Chorus Analyses
To understand how aircraft noise affects the start and the duration of dawn chorus, we deployed autonomous recording devices in the field (Bioacoustic Audio Recorders™ -BAR-, Frontier Labs). Each BAR was strapped onto a tree branch less than 5 m from the song post of a specific individual, recording its singing activity during 3 h of continuous recording from 1:30 h before sunrise to 1:30 h after sunrise (sampling rate: 48 kHz, 16 bits). The BARs were fixed in the same position for several days (3.18 ± 0.15 days per individual, mean ± SE) in order to get a representative sample of each individual's chorus characteristics. Afterwards, we retrieved the BAR and downloaded the data to an external hard drive before it was deployed back in the field to a new blackbird territory. We carried out a total of 105 automatic recordings of dawn chorus with 15 individuals in the airport and 18 in the control population from April to June.
In the lab, we monitored the automatic recordings manually using a set of Sennheiser HD 205 headphones. In every recording (1 individual, 1 day) we determined the start of dawn chorus, defined as the first blackbird song heard in the recording with high relative amplitude and continuity. Blackbird songs that presented very low relative amplitude in the recording were dismissed because they could be songs from neighboring territories. Continuity was validated by moving 5 min forward in the recording and checking whether the blackbird was still singing (Bruni et al., 2014). Note that this procedure is different to a previous study in which we took the absolute earliest song (Gil et al., 2014). If the blackbird was still singing 5 min later in the recording, we would confirm this song as the start of dawn chorus and if not, we would omit this song considering it an isolated song. Moving on in the recording we would repeat this procedure until determining the beginning of dawn chorus. Once we confirmed the start of the chorus, we continued the analysis by listening to 1 min of recording every 5 min starting from the beginning of the chorus until the end of the recording (1:30 h after sunrise). For every 5 min interval, we noted whether the blackbird was singing or not based on the 1 min that was analyzed. Dawn chorus duration was defined as the number of intervals in which a blackbird was singing uninterrupted multiplied by 5 min of interval duration. Over the 220.5 h of recording analyzed we identified a blackbird singing on 1,271 intervals and out of these, there were 196 intervals in which the blackbird that was singing was identified as the neighbor and not the territory holder, this represents 15.43% of occasions.
In order to understand how the temporal overlapping of dawn chorus with aircraft traffic activity affected song timing, we divided the season in two halves depending on whether chorus was overlapping or non-overlapping with day time traffic schedule. To this end we required a reference of what is the normal start time of blackbird dawn chorus living in a habitat with no noise impact. Therefore, we used the timing parameters of dawn chorus of our control population as the standard time of singing for blackbirds living under non-urban conditions. In the control site, blackbird chorus began 37.7 ± 1.22 min (mean ± SE) before sunrise lasting for 26.1 ± 1.98 min (mean ± SE), which is consistent with previous references (Snow, 1958; Blotzheim et al., 1993; Ripmeester et al., 2010). We estimated that natural blackbird chorus would overlap with the traffic when sunrise occurred at 7:11 a.m. or later. When sunrise occurred at 7:11, normal blackbird dawn chorus would begin at 6:34 on average, lasting for 26 min until 7:00 am and therefore not overlapping with aircraft traffic, according to our reference. Hence, when sunrise was later than 7:11, blackbird dawn chorus would overlap with aircraft traffic noise but later on the season when sunrise occurs before 7:11, blackbird dawn chorus would not overlap with high levels of aircraft traffic noise.
Statistics
All statistical analyses were carried out in R software, version 3.3.1 (Team, 2012). Firstly, we tested whether background noise differed between the study sites. We analyzed the differences of broadband dB levels between sites and its interaction with the three time intervals of aircraft traffic activity defined before. We fitted a Generalized Linear Mixed Model (glmm) to determine the effect of the study site, the time intervals (regarding the state of aircraft traffic) and its interaction on the dependent variable: dBz levels as an unbiased measure of noise. The model was fitted using a Gamma family distribution and a log link function. The diagnostic plots were visually inspected for possible deviations from heterogeneity and homoscedasticity assumptions. After this, we proceeded to model selection by computing all possible models using dredge function from the “MuMIn” package (Barton, 2011) and ranking all models by their Akaike's Information Criteria (AICc). Final model was the full model being the best model with a ΔAICc of 15.59 with the second best.
To investigate potential differences in the acoustic characteristics of the song between the two study populations, we compared the mean, the maximum and minimum dominant frequency of each note. For the analysis, we treated the motif and twitter data separately (Dabelsteen et al., 1993). The data for this analysis was nested using random factors: notes within songs, songs within individuals. Therefore, we fitted Linear Mixed Models (lmm). When the response variable was not normal, we fitted a Generalized Linear Mixed Models (glmm) with a Gamma family distribution since all frequency parameters are positive continuous variables.
Then, using song as unit, we compared several temporal parameters such as the note to silence ratio within song, song length, singing effort, motif length, twitter length and twitter proportion. The analysis of twitter length and twitter proportion was restricted to the songs that actually presented a twitter section. This subset of the data included 233 songs from all 16 individuals at each site. All models had study site as fixed term: a categorical variable of two levels: “airport” and “control” (see Table 2 for model details). In this model the random term was only the individual because the measurements were taken per song and not per note. We also fitted a binomial Generalized Linear Model (glm) to investigate population differences in the number of songs with twitter vs. number of songs without twitter. For this purpose we built another data set summarizing the sum of songs with twitter and without twitter per individual. In this binomial model, because each individual was represented only once in the data set it was not necessary to fit a mixed effect model. We validated all models by visual inspection of diagnostic plots.
Finally we analyzed which characteristics explained the start and the duration of dawn chorus. For both parameters, we fitted a glmm with Poisson distributions and log link functions. The full model was similar in both cases with the following factors in the fixed term: study site (“airport” and “control”) and traffic overlap with dawn chorus (“overlap,” “no overlap”) and their full interaction. Every individual territory was sampled for several days so individuals were included as random factor to avoid pseudo-replication and account for intra individual variation. We validated all models by visual inspection of diagnostic plots and proceeded to model selection using AICc. In the model analyzing start of dawn chorus the final model was the best model since the second best was at a distance of more than 2 AICc units. The final model in the case of dawn chorus duration was the model average of the best three models.
Results
Noise
We tested differences in background noise over time, from 5 to 8 a.m., between the two study sites and the interaction between study site and time (see Table 1 for model output). Between 5 and 6 a.m., noise levels are low at both sites but significantly higher in the airport. The estimated decibel levels are very low in both cases and the difference is small; airport 57.40 dB(Z) and control 54.05 dB(Z). Nevertheless, these levels of noise are still very silent conditions. In the interval “traffic runway 3” we find a significant increase in noise regardless of the study site. This increase is probably due to the impact of aircraft traffic noise coming from runway 3 and perhaps other abiotic factors such as increased wind speed. It is important to notice that noise conditions are not significantly different between sites during this interval. After 7 a.m., the increment in noise at the airport site is markedly abrupt and significantly higher than at the control site with an estimated average noise level of 77.47 dB(Z) in the airport and 63.43 dB(Z) at the control site (see Figure 2). It is the onset of aircraft traffic under the day-time schedule that brings clear and marked differences in between study sites and, most important, it is highly fixed and sudden as it always begins at 7 a.m. throughout the entire year.
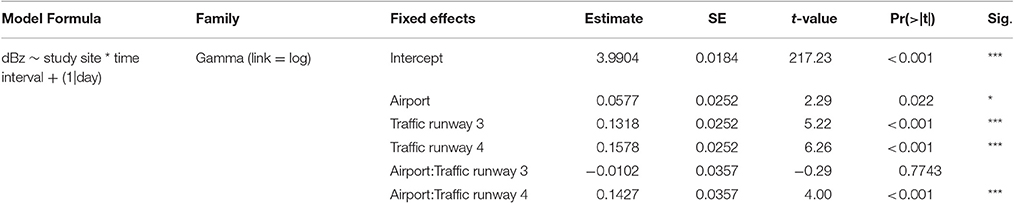
Table 1. Results from the GLMMs to understand noise differences between sites.
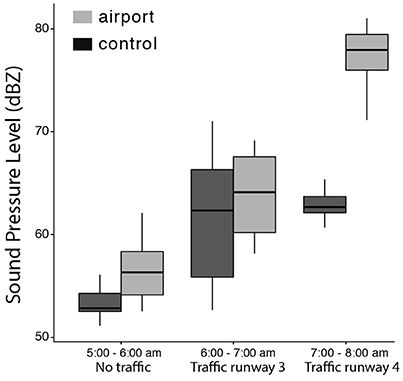
Figure 2. Noise levels at both sites during early morning. Before 6 a.m. there are some small differences between sites that disappear at 6 am. However, obvious differences between study sites arise at 7 am with the onset of day-time traffic schedule.
Most of the energy resulting from the noise produced by aircraft traffic is concentrated on the lower frequency range but it also expands to the higher frequencies typical of birdsong (Figure 3C). In Figure 3C we can see how the noise contribution coming from biotic noise, mostly bird song and other vocalization, shows a clear peak between 3 and 6 kHz in the control study site (dashed line) (Brumm and Slabbekoorn, 2005; Nemeth and Brumm, 2009). In the airport site (continuous line) this contribution is concealed because aircraft noise partially overlaps with the lower frequencies used by birds. This is specially relevant for our model species since the typical frequencies used by blackbirds in their motif notes are concentrated in the frequency range from 1.5 to 3 kHz (Dabelsteen, 1984). Finally, we can observe that the curves of noise along the spectrum are very similar for the two study sites from 5 to 7 a.m. (Figures 3A,B) and this fact is obviously different after 7 a.m. when the day-time traffic schedule begins at the airport.
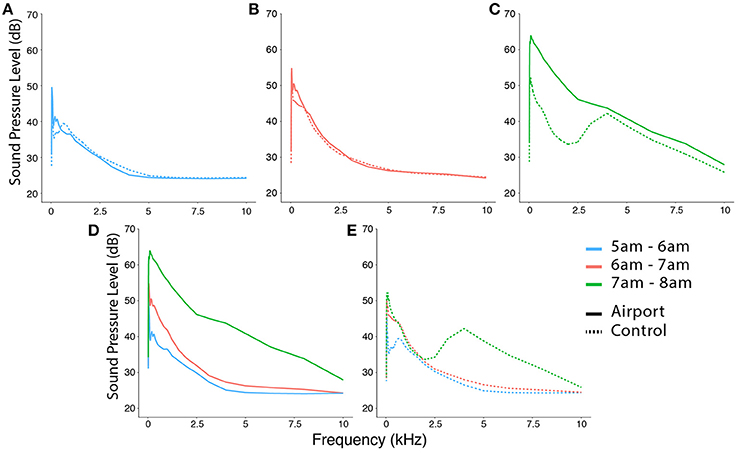
Figure 3. (A–C) show the noise distribution along the spectrum during the three time intervals defined previously: “no traffic,” “traffic runway 3,” and “traffic runway 4.” In each of these three plots, dashed lines represent the control site and continuous lines represent the airport site. We can see that both sites are very similar in (A,B) but this changes dramatically in (C) that shows the start of the noise produced by the day-time traffic. Here we can appreciate how aircraft noise overlaps at least partially with the frequencies typical of bird vocalizations whose normal representation in a quiet forest looks similar to the control curve. Finally, (D,E) show the three time intervals plotted together for each study site separately illustrating more clearly the increase of noise at dawn.
Song Characteristics
We found no differences between the airport and the control populations in any of the song frequency parameters (see Figure 4). However, regarding the temporal characteristics of song, we found significant differences between the two study populations. In the airport populations, songs without twitter part (i.e., songs composed only by a motif), were sang significantly more often than in the control (see Figure 5A). Regarding the subset of songs that included twitter, blackbirds next to the airport used shorter twitter proportions (see Figure 5B). In the airport, blackbirds also used a shorter twitter in absolute duration but this difference was not statistically significant. Other temporal parameters analyzed such as the motif duration, note to silence ratio within song, song length or singing effort presented no differences between populations (see Table 2 for model results).
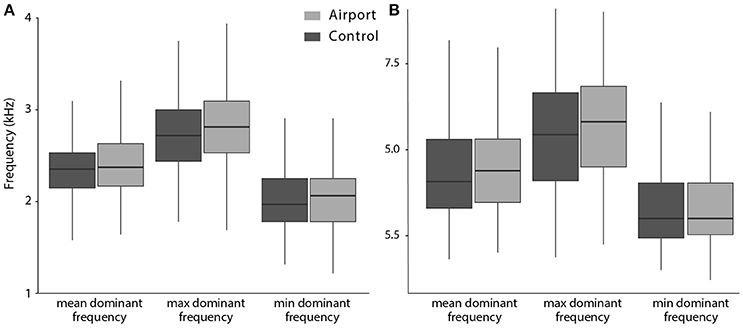
Figure 4. Boxplots showing frequency characteristics of blackbird song in the two populations of the study. (A) Shows the mean, maximum and minimum dominant frequency of motif notes and (B) shows the mean, maximum and minimum dominant frequency of twitter notes.
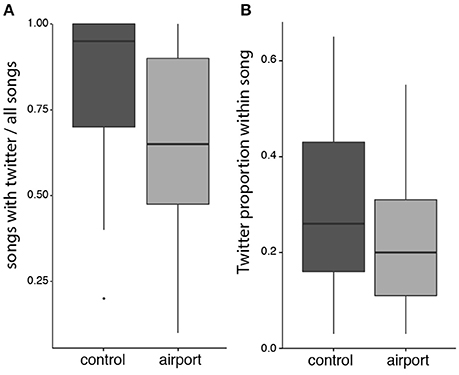
Figure 5. Boxplots comparing (A) the proportion of songs with twitter out of the total number of songs analyzed per male, and (B) the proportion of twitter length within the song for the subset of songs that present twitter notes.
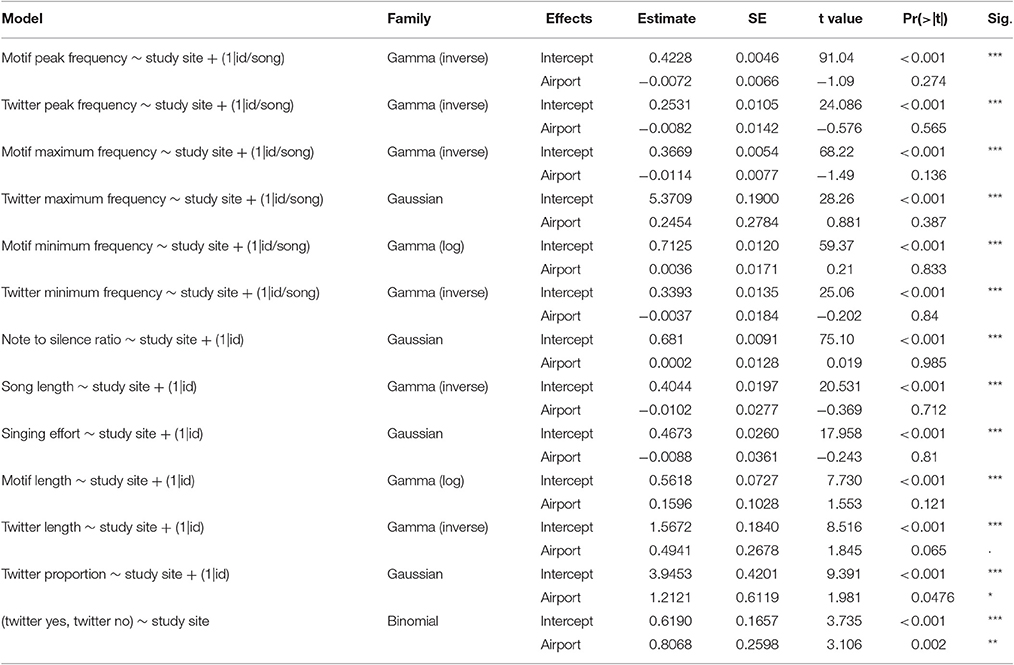
Table 2. Results of GLMMs modeling song characteristics.
Dawn Chorus
The final model investigating the factors that influence the onset of dawn chorus shows that airport blackbirds significantly advanced their chorus but only when it overlapped temporally with the day-time traffic schedule. Onset of dawn chorus in the control population was not different whether it overlapped or not with traffic activity. The model estimates a dawn chorus advance of 12.8 min in the airport when there was an overlap between the dawn chorus and aircraft traffic (see Figure 6B and Table 3 for model results).
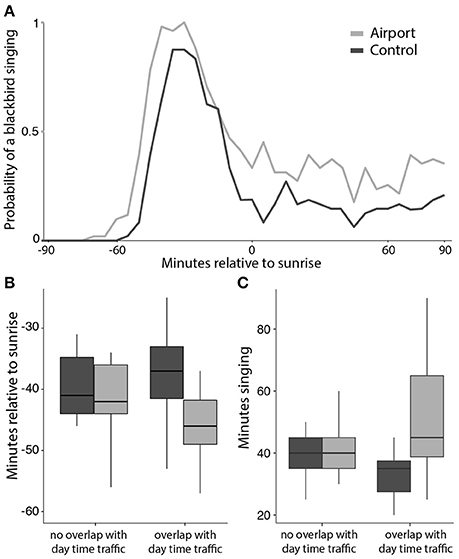
Figure 6. Characterization of the temporal parameters of dawn chorus. Curves in (A) show the probability of hearing a blackbird singing at dawn. All recordings were standardized in relation to sunrise time = 0, negative numbers meaning minutes before sunrise. Boxplots in show the differences in onset of dawn chorus in minutes relative to sunrise (B) and the duration of the chorus (C), depending on whether the chorus overlapped with day time traffic or not.
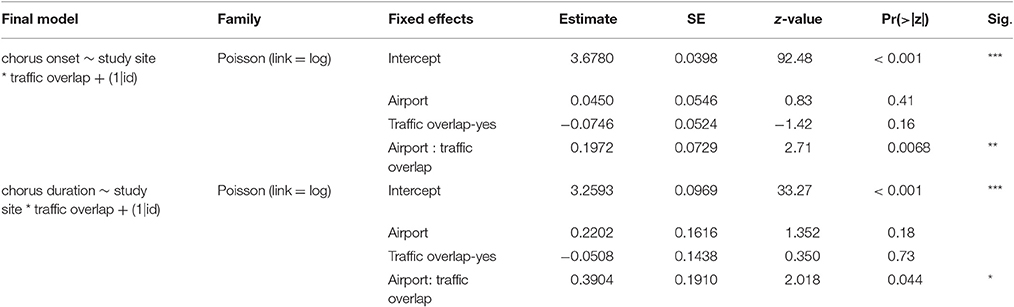
Table 3. Results of blackbird dawn chorus GLMMs.
A visual inspection of the average probability of hearing a blackbird singing along the early morning showed a very clear peak of singing activity that corresponded to dawn chorus and this was followed by sporadic singing activity (see Figure 6A). We tested whether the duration of dawn chorus was dependent on aircraft noise and/or study site. Similar to the effect found on the onset of dawn chorus, the duration of the dawn chorus was not systematically longer at the airport but blackbirds enlarged the duration of their chorus when it overlapped with day time aircraft traffic. The model estimates a difference of 24.1 min between sites, airport birds singing a longer dawn chorus. When dawn chorus did not overlap with traffic activity, the duration was similar at both sites (see Figure 6C and Table 3).
Discussion
We examined differences in song and singing behavior in two blackbird populations differing in the impact of aircraft noise perturbation. Our data show several differences in song design and timing that suggest adaptive changes to the local noise environment. Firstly, blackbirds living near the airport sang more often songs without twitter (only motif notes) and, furthermore, when they did include a twitter part in their songs, the twitter proportion was smaller. This may suggest that airport blackbirds emphasized the loudest part of their songs (motifs), disregarding the fainter part (twitter), in an effort to reduce masking by noise. However, airport blackbirds did not use higher frequencies than those in the control population. This is in contrast to previous findings on this and similar species living near other anthropogenic sources of noise (Slabbekoorn and Peet, 2003; Nemeth and Brumm, 2009; Ripmeester et al., 2010). Additionally, blackbirds living next to the airport advanced the timing of their singing activity at dawn when their dawn chorus overlapped temporally with aircraft activity (early on the season). Interestingly, this advance in chorus onset was coupled with a significant increase in chorus duration. Later on the season, when the timing of the blackbird chorus fell well before the onset of day-time traffic, we found no differences in timing between the control and airport populations. Therefore, blackbirds seem to optimize the timing of their singing activity by adjusting their chorus when it is indeed beneficial but this modification is not constant throughout the season. This may imply that selection for early singers in noisy environments does not lead to an invariant advance but rather favours a highly plastic response in their singing behavior.
A large body of evidence shows that increased frequencies and increased amplitude are two typical urban song modifications (Slabbekoorn and Peet, 2003; Ríos-Chelén et al., 2012; Brumm and Zollinger, 2013). Many authors have argued that these modifications enhance signal transmission in an urban environment and therefore that they are adaptive, but there is some debate as to whether higher frequencies are a consequence of increases in amplitude (Brumm and Slabbekoorn, 2005; Slabbekoorn and den Boer-Visser, 2006; Warren et al., 2006; Nemeth and Brumm, 2010). Data show that in some species higher-frequency songs are louder (Nemeth et al., 2013), and simulations suggest that the benefit of singing higher is much smaller than that of singing louder (Nemeth and Brumm, 2010). Nevertheless, these patterns need not be universal to all species. Adding a new layer of complexity, experimental studies of signal degradation under urban and rural environments have shown that great tit (Parus major) urban songs suffered less degradation in both habitats contradicting the Acoustic Adaptation Hypothesis (Mockford et al., 2011). Regardless of the reduced signal degradation of urban songs, that study showed that rural songs prevail in rural environments suggesting that urban modifications may compromise important information of song (Mockford et al., 2011). In contrast to these findings, we found no differences in song frequency in blackbirds living near an airport. A main difference with the previous literature is that we focused on the impact of noise produced by aircraft traffic near a large airport where noise bursts during aircraft events reach very high dB levels, up to 110 dB. Unlike the continuous noise levels of noise found in the city (Dooling and Popper, 2007; Arroyo Solís et al., 2013) in the airport environment that we studied we commonly encounter silent intervals between aircraft events when noise levels may drop below 60 dB(Z) resembling those of a quiet forest (personal observation). It is possible that blackbirds do indeed increase song frequency during noise bursts produced by aircraft and return to the use of normal frequencies soon afterwards. Unfortunately, since we could only conduct song analyses when background noise levels were low, we cannot test this hypothesis. But our data do show a lack of differences in song frequency between areas when aircraft noise is absent. Alternatively, it may be argued that there is a particular noise amplitude threshold over which any modification of song becomes irrelevant (Brumm et al., 2009; Díaz et al., 2011). In addition, during silent conditions, it is likely that higher frequency songs typical of the cities entail other costs because forest songs of lower frequency may provide relevant information and perhaps better sound transmission through the foliage of the forest (Gil and Gahr, 2002; Slabbekoorn et al., 2007; Mockford et al., 2011).
Regarding the temporal parameters of song, we found that blackbirds in the airport population were more likely to drop the twitter of their songs than control blackbirds and, when they did sing twitters, the twitter proportion of these songs was significantly smaller. In contrast, a previous study in cities has shown that urban blackbirds use a larger twitter proportions than rural birds (Ripmeester et al., 2010). It has been argued that these differences could be due to an advanced breeding season and increased territory density in the city and not related to noise (Ripmeester et al., 2010). These confounding factors are unlikely to be a bias in our study because: (1) breeding season is improbably different between the two sites (see Results section) plus we alternated recording days to avoid biases due to date. In addition, given the similarity between both study sites in habitat and vegetation (Gil et al., 2014), we do not expect to find differences in density, although we do not have the data to support this claim.
A possible adaptive acoustic explanation for the patterns found may be in relation to the amplitude of the different parts that make up the blackbird song. Although twitter notes are higher in frequency than motif notes and therefore suffer less spectral masking by anthropogenic noise they are much weaker in amplitude and would present a much lower signal to noise ratio under aircraft noise than motif notes (Dabelsteen, 1984). In fact, we were obliged to drop analyses of blackbird song under aircraft noise conditions because twitter notes were nearly fully concealed by noise in the spectrogram. On the other hand, some researchers have previously suggested that increased tonality of sound elements is adaptive under noisy conditions (Dubois and Martens, 1984; Hanna et al., 2011; Mockford et al., 2011). Following this argument and given that the twitter part of the song is nearly lost completely it may pay off for blackbirds to favor the motif rather than the twitter of song.
We also found that birds living near the airport advanced the onset of dawn chorus and increased the duration of the chorus in comparison with the control population. Our results suggest that these changes in dawn chorus timing are related to the increased noise levels due to the onset of aircraft traffic in the runway closest to the forest at 7 a.m. The reason why we find aircraft traffic as a likely cause of this modifications is because (1) noise increases to significant higher levels after 7 a.m. compared to the control population and (2) blackbirds near the airport do not show this modifications later on the season when the blackbird dawn chorus does not overlap with day-time aircraft traffic. When blackbird dawn chorus occurs entirely before 7 a.m. both populations show similar chorus characteristics. Although the start and duration of dawn chorus may vary along the season in relation to their breeding stage (Thomas, 1999; Bruni et al., 2014) this should not be a confounding factor in this study since recordings were made simultaneously at both sites. In addition, the distance between study sites is small and the habitat is nearly identical within the same strip of forest so we find unlikely that breeding stage could be a bias in these results.
We present the first evidence that the advance of dawn chorus in this context is not systemic but plastic as the difference with the control population disappears when dawn chorus does not interfere with aircraft traffic noise. Moreover, this advance is possibly not due to an early awakening of the birds by noise since the advance occurs mostly when dawn chorus begins during the interval from 6 to 7 a.m. in the morning when noise levels are similar at both sites. Our model shows that noise levels during this interval are not significantly different between airport and control sites because airport traffic is still restricted by the night schedule. Therefore, both airport and control blackbirds are under similar noise levels but still the onset of dawn chorus at the airport is estimated to start 12.8 min earlier.
Perhaps more relevant than singing earlier is the finding that blackbirds increased the duration of dawn chorus near the airport when dawn chorus overlapped temporally with aircraft traffic. The model estimates that airport blackbirds sing an average of 24.1 min more than control blackbirds, almost twice as long. This novel finding further supports the adaptive perspective of the modifications of dawn chorus under noisy conditions because, in this case, the debate of whether birds are awaken or not by noise is no longer relevant. It seems intuitive that singing for longer increases the communicative efficiency of song as suggested by previous evidence (Díaz et al., 2011; Gil et al., 2014). This increase in the duration of dawn chorus under intermittent noise conditions may provide a compensation for those songs that have been masked during strong bursts of aircraft noise, ensuring that songs are being heard without compromising information coded on other temporal parameters such as song rate or song length (Wasserman and Cigliano, 1991; Galeotti et al., 1997; Linhart et al., 2012). However, it also seems plausible that doubling the normal time allocated for song at dawn may carry higher costs than the advance of dawn chorus. Spending more time singing will likely have negative effects on fitness by increased metabolic costs associated with song or by taking this time away from other vital tasks (e.g., foraging; Thomas, 1999, 2002).
Until now, there have been several attempts to explain the mechanism underlying noise-dependent advance of dawn chorus onset. An experimental study (Arroyo Solís et al., 2013) suggested that the advance of dawn chorus could be due to a short-term response (e.g., birds are waken up by increased noise levels) but two other correlative studies (Gil et al., 2014; Dominoni et al., 2016) implied that there must be another mechanism underlying the advance of chorus because this modification happens before noise levels increase at dawn. Our results support the second hypothesis since the advance of dawn chorus in the airport occurred mostly when noise levels were similar to control at the onset. Perhaps the two mechanisms, a direct awakening by noise or a plastic adaptive response to anthropogenic noise, may be associated within the same process but at different steps. Birds may respond with short-term advances of their singing activity when suddenly noise levels increase at dawn. In such cases, there could be selection for those individuals that sing earlier than usual but, if noise levels follow a strict and periodic daily pattern, birds may develop a long-term response that optimizes their singing activity advancing their chorus in order to avoid noisy hours.
In conclusion, short-term noise-induced modifications of song regarding spectral and temporal characteristics may not follow a linear response but depend on the amplitude and temporal features of anthropogenic noise, as suggested by some authors (Brumm et al., 2009; Díaz et al., 2011; Gil et al., 2014). Anthropogenic noise includes an ample array of sources that may lead to different adaptive responses of song depending on the acoustic features and the temporal regime of noise. This is supported by our results that found airport blackbirds showing different song modifications than those found on other studies for their city counterparts (Nemeth and Brumm, 2009; Ripmeester et al., 2010). In relation to long-term noise-induced changes in singing behavior, our results agree with former evidence that birds advance the onset of chorus in locations where background levels rise at dawn. Finally, we provide evidence that anthropogenic noise may induce birds to increase the time singing at dawn, suggesting higher fitness costs in relation to daily energy expenditure.
Author Contributions
DG directed the research and provided most of the conceptual framework behind this study. JS participated in the development of hypotheses and predictions, carried out the field work and data collection, performed acoustic and statistical analyses and wrote the first draft. ES helped out during field work and IP provided technical knowledge on acoustics and noise measurements.
Funding
This study has been funded by a research grant from the Spanish Ministerio de Economía y Competitividad (CGL2014-55577R) to DG.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Arroyo Solís, A., Castillo, J., Figueroa, E., López Sánchez, J., and Slabbekoorn, H. (2013). Experimental evidence for an impact of anthropogenic noise on dawn chorus timing in urban birds. J. Avian Biol. 44, 288–296. doi: 10.1111/j.1600-048X.2012.05796.x
Bermúdez-Cuamatzin, E., Ríos-Chelén, A. A., Gil, D., and Garcia, C. M. (2011). Experimental evidence for real-time song frequency shift in response to urban noise in a passerine bird. Biol. Lett. 7, 36–38. doi: 10.1098/rsbl.2010.0437
Bermúdez-Cuamatzin, E., Ríos-Chelén, A. A., Gil, D., and Garcia, C. M. (2012). Experimental evidence for real-time song frequency shift in response to urban noise in a passerine bird. Biol. Lett. 8:320. doi: 10.1098/rsbl.2011.1225
Blotzheim, U. N. G., Bauer, K., and Bezzel, E. (1993). Handbuch der Vögel Mitteleuropas. Wiesbaden: AULA-Verlag.
Brumm, H. (2004). The impact of environmental noise on song amplitude in a territorial bird. J. Anim. Ecol. 73, 434–440. doi: 10.1111/j.0021-8790.2004.00814.x
Brumm, H., Schmidt, R., and Schrader, L. (2009). Noise-dependent vocal plasticity in domestic fowl. Anim. Behav. 78, 741–746. doi: 10.1016/j.anbehav.2009.07.004
Brumm, H., and Slabbekoorn, H. (2005). Acoustic communication in noise. Adv. Stud. Behav. 35, 151–209. doi: 10.1016/S0065-3454(05)35004-2
Brumm, H., and Todt, D. (2002). Noise-dependent song amplitude regulation in a territorial songbird. Anim. Behav. 63, 891–897. doi: 10.1006/anbe.2001.1968
Brumm, H., and Zollinger, S. A. (2013). “Avian vocal production in noise,” in Animal Communication and Noise, ed H. Brumm (Berlin; Heidelberg: Springer), 187–227.
Bruni, A., Mennill, D. J., and Foote, J. R. (2014). Dawn chorus start time variation in a temperate bird community: relationships with seasonality, weather, and ambient light. J. Ornithol. 155, 877–890. doi: 10.1007/s10336-014-1071-7
Catchpole, C. K., and Slater, P. J. B. (2003). Bird Song: Biological Themes and Variations. Cambridge: Cambridge University Press.
Cramp, S. (1992). Handbook of the Birds of Europe, the Middle East and North Africa: Tyrant Flycatchers to Thrushes v.5: The Birds of the Western Palearctic: Tyrant Flycatchers to Thrushes. Vol. 5. Oxford: Oxford University Press.
Cynx, J., Lewis, R., Tavel, B., and Tse, H. (1998). Amplitude regulation of vocalizations in noise by a songbird, Taeniopygia guttata. Anim. Behav. 56, 107–113. doi: 10.1006/anbe.1998.0746
Dabelsteen, T. (1984). An analysis of the full song of the blackbird Turdus merula with respect to message coding and adaptations for acoustic communication. Ornis Scandinavica 15, 227–239. doi: 10.2307/3675931
Dabelsteen, T., Larsen, O. N., and Pedersen, S. B. (1993). Habitat-induced degradation of sound signals: quantifying the effects of communication sounds and bird location on blur ratio, excess attenuation, and signal-to-noise ratio in blackbird song. J. Acoust. Soc. Am. 93, 2206–2220. doi: 10.1121/1.406682
Da Silva, A., Valcu, M., and Kempenaers, B. (2016). Behavioural plasticity in the onset of dawn song under intermittent experimental night lighting. Anim. Behav. 117, 155–165. doi: 10.1016/j.anbehav.2016.05.001
Díaz, M., Parra, A., and Gallardo, C. (2011). Serins respond to anthropogenic noise by increasing vocal activity. Behav. Ecol. 22, 332–336. doi: 10.1093/beheco/arq210
Dominoni, D. M., Carmona-Wagner, E. O., Hofmann, M., Kranstauber, B., and Partecke, J. (2014). Individual-based measurements of light intensity provide new insights into the effects of artificial light at night on daily rhythms of urban-dwelling songbirds. J. Anim. Ecol. 83, 681–692. doi: 10.1111/1365-2656.12150
Dominoni, D. M., Greif, S., Nemeth, E., and Brumm, H. (2016). Airport noise predicts song timing of European birds. Ecol. Evol. 6, 6151–6159. doi: 10.1002/ece3.2357
Dooling, R. J., and Popper, A. N. (2007). The Effects of Highway Noise on Birds. The California Department of Transportation Division of Environmental Analysis 74, Sacramento, CA.
Dorado-Correa, A. M., Rodríguez-Rocha, M., and Brumm, H. (2016). Anthropogenic noise, but not artificial light levels predicts song behaviour in an equatorial bird. R. Soc. Open Sci. 3:160231. doi: 10.1098/rsos.160231
Dubois, A., and Martens, J. (1984). A case of possible vocal convergence between frogs and a bird in Himalayan torrents. J. Ornithol. 125, 455–463. doi: 10.1007/BF01640137
Evans, K. L., Gaston, K. J., Frantz, A. C., Simeoni, M., Sharp, S. P., McGowan, A., et al. (2009). Independent colonization of multiple urban centres by a formerly forest specialist bird species. Proc. R. Soc. Lond. B Biol. Sci. 276, 2403–2410. doi: 10.1098/rspb.2008.1712
Francis, C. D., Ortega, C. P., and Cruz, A. (2011). Different behavioural responses to anthropogenic noise by two closely related passerine birds. Biol. Lett. 7, 850–852. doi: 10.1098/rsbl.2011.0359
Fuller, R. A., Warren, P. H., and Gaston, K. J. (2007). Daytime noise predicts nocturnal singing in urban robins. Biol. Lett. 3, 368–370. doi: 10.1098/rsbl.2007.0134
Galeotti, P., Saino, N., Sacchi, R., and MÃLler, A. P. (1997). Song correlates with social context, testosterone and body condition in male barn swallows. Anim. Behav. 53, 687–700. doi: 10.1006/anbe.1996.0304
Gil, D., and Brumm, H. (eds.). (2013). “Acoustic communication in the urban environment: patterns, mechanisms, and potential consequences of avian song adjustments,” in Avian Urban Ecology: Behavioural and Physiological Adaptations (Oxford: Oxford University Press), 69–83.
Gil, D., and Gahr, M. (2002). The honesty of bird song: multiple constraints for multiple traits. Trends Ecol. Evol. (Amst). 17, 133–141. doi: 10.1016/S0169-5347(02)02410-2
Gil, D., Honarmand, M., Pascual, J., Pérez-Mena, E., and Garcia, C. M. (2014). Birds living near airports advance their dawn chorus and reduce overlap with aircraft noise. Behav. Ecol. 26, 435–443. doi: 10.1093/beheco/aru207
Hanna, D., Blouin-Demers, G., Wilson, D. R., and Mennill, D. J. (2011). Anthropogenic noise affects song structure in red-winged blackbirds (Agelaius phoeniceus). J. Exp. Biol. 214, 3549–3556. doi: 10.1242/jeb.060194
Hansen, I. J. K., Otter, K. A., Van Oort, H., and Holschuh, C. I. (2005). Communication breakdown? Habitat influences on black-capped chickadee dawn choruses. Acta Ethol. 8, 111–120. doi: 10.1007/s10211-005-0007-x
International Organization for Standardization (2016). International Organization for Standardization, 2016. ISO 1996-1:2016 Acoustics — Description, Measurement and Assessment of Environmental Noise — Part 1: Basic Quantities and Assessment Procedures. Geneva: International Organization for Standardization
Kacelnik, A., and Krebs, J. R. (1983). The dawn chorus in the great tit (Parus major): proximate and ultimate causes. Behaviour 83, 287–308. doi: 10.1163/156853983X00200
Linhart, P., Slabbekoorn, H., and Fuchs, R. (2012). The communicative significance of song frequency and song length in territorial chiffchaffs. Behav. Ecol. 23, 1338–1347. doi: 10.1093/beheco/ars127
McNamara, J., Mace, R., and Houston, A. (1987). Optimal daily routines of singing and foraging in a bird singing to attract a mate. Behav. Ecol. Sociobiol. 20, 399–405. doi: 10.1007/BF00302982
Mendes, S., Colino-Rabanal, V. J., and Peris, S. J. (2011). Bird song variations along an urban gradient: the case of the European blackbird (Turdus merula). Landsc. Urban Plan. 99, 51–57. doi: 10.1016/j.landurbplan.2010.08.013
Merchant, N. D., Fristrup, K. M., Johnson, M. P., Tyack, P. L., Witt, M. J., Blondel, P., et al. (2015). Measuring acoustic habitats. Methods Ecol. Evol. 6, 257–265. doi: 10.1111/2041-210X.12330
Mockford, E. J., Marshall, R. C., and Dabelsteen, T. (2011). Degradation of rural and urban great tit song: testing transmission efficiency. PLoS ONE 6:e28242. doi: 10.1371/journal.pone.0028242
Morton, E. S. (1975). Ecological sources of selection on avian sounds. Am. Nat. 109, 17–34. doi: 10.1086/282971
Nemeth, E., and Brumm, H. (2009). Blackbirds sing higher-pitched songs in cities: adaptation to habitat acoustics or side-effect of urbanization? Anim. Behav. 78, 637–641. doi: 10.1016/j.anbehav.2009.06.016
Nemeth, E., and Brumm, H. (2010). Birds and anthropogenic noise: are urban songs adaptive? Am. Nat. 176, 465–475. doi: 10.1086/656275
Nemeth, E., Pieretti, N., Zollinger, S. A., Geberzahn, N., Partecke, J., Miranda, A. C., et al. (2013). Bird song and anthropogenic noise: vocal constraints may explain why birds sing higher-frequency songs in cities. Proc. R Soc. B 280:20122798. doi: 10.1098/rspb.2012.2798
Nordt, A., and Klenke, R. (2013). Sleepless in town–drivers of the temporal shift in dawn song in urban European blackbirds. PLoS ONE 8:e71476. doi: 10.1371/journal.pone.0071476
Partecke, J., Schwabl, I., and Gwinner, E. (2006). Stress and the city: urbanization and its effects on the stress physiology in European blackbirds. Ecology 87, 1945–1952. doi: 10.1890/0012-9658(2006)87[1945:SATCUA]2.0.CO;2
Ríos-Chelén, A. A., Salaberria, C., Barbosa, I., Macías Garcia, C., and Gil, D. (2012). The learning advantage: bird species that learn their song show a tighter adjustment of song to noisy environments than those that do not learn. J. Evol. Biol. 25, 2171–2180. doi: 10.1111/j.1420-9101.2012.02597.x
Ríos-Chelén, A. A., Quirós-Guerrero, E., Gil, D., and Garcia, C. M. (2013). Dealing with urban noise: vermilion flycatchers sing longer songs in noisier territories. Behav. Ecol. Sociobiol. 67, 145–152. doi: 10.1007/s00265-012-1434-0
Ripmeester, E. A., Kok, J. S., van Rijssel, J. C., and Slabbekoorn, H. (2010). Habitat-related birdsong divergence: a multi-level study on the influence of territory density and ambient noise in European blackbirds. Behav. Ecol. Sociobiol. 64, 409–418. doi: 10.1007/s00265-009-0857-8
Roca, I. T., Desrochers, L., Giacomazzo, M., Bertolo, A., Bolduc, P., Deschesnes, R., et al. (2016). Shifting song frequencies in response to anthropogenic noise: a meta-analysis on birds and anurans. Behav. Ecol. 27, 1269–1274. doi: 10.1093/beheco/arw060
Ryan, M. J., and Brenowitz, E. A. (1985). The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am. Nat. 126, 87–100. doi: 10.1086/284398
Shanahan, D. F., Strohbach, M. W., Warren, P. S., and Fuller, R. A. (2014). “The challenges of urban living,” in Avian Urban Ecology, eds D. Gil and H. Brumm (Oxford: Oxford University Press), 3–20.
Slabbekoorn, H., and den Boer-Visser, A. (2006). Cities change the songs of birds. Curr. Biol. 16, 2326–2331. doi: 10.1016/j.cub.2006.10.008
Slabbekoorn, H., and Peet, M. (2003). Ecology: birds sing at a higher pitch in urban noise. Nature 424, 267–267. doi: 10.1038/424267a
Slabbekoorn, H., and Ripmeester, E. A. (2008). Birdsong and anthropogenic noise: implications and applications for conservation. Mol. Ecol. 17, 72–83. doi: 10.1111/j.1365-294X.2007.03487.x
Slabbekoorn, H., Yeh, P., and Hunt, K. (2007). Sound transmission and song divergence: a comparison of urban and forest acoustics. Condor 109, 67–78. doi: 10.1650/0010-5422(2007)109[67:STASDA]2.0.CO;2
Team, R. C. (2012). R: A Language and Environment for Statistical Computing. Vienna: R Development Core Team.
Thomas, R. J. (1999). Two tests of a stochastic dynamic programming model of daily singing routines in birds. Anim. Behav. 57, 277–284. doi: 10.1006/anbe.1998.0956
Thomas, R. J. (2002). The costs of singing in nightingales. Anim. Behav. 63, 959–966. doi: 10.1006/anbe.2001.1969
Verzijden, M. N., Ripmeester, E. A. P., Ohms, V. R., Snelderwaard, P., and Slabbekoorn, H. (2010). Immediate spectral flexibility in singing chiffchaffs during experimental exposure to highway noise. J. Exp. Biol. 213, 2575–2581. doi: 10.1242/jeb.038299
Warren, P. S., Katti, M., Ermann, M., and Brazel, A. (2006). Urban bioacoustics: it's not just noise. Anim. Behav. 71, 491–502. doi: 10.1016/j.anbehav.2005.07.014
Wasserman, F., and Cigliano, J. (1991). Song output and stimulation of the female in white-throated sparrows. Behav. Ecol. Sociobiol. 29, 55–59. doi: 10.1007/BF00164295
Keywords: birdsong, anthropogenic noise, avian communication, urban ecology, behavioral plasticity
Citation: Sierro J, Schloesing E, Pavón I and Gil D (2017) European Blackbirds Exposed to Aircraft Noise Advance Their Chorus, Modify Their Song and Spend More Time Singing. Front. Ecol. Evol. 5:68. doi: 10.3389/fevo.2017.00068
Received: 23 February 2017; Accepted: 13 June 2017;
Published: 30 June 2017.
Edited by:
Michaela Hau, Max Planck Institute for Ornithology, GermanyReviewed by:
Davide M. Dominoni, Netherlands Institute of Ecology (NIOO-KNAW), NetherlandsSharon A. Gill, Western Michigan University, United States
Copyright © 2017 Sierro, Schloesing, Pavón and Gil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Sierro, sierro.2.8@gmail.com