Thermal Stress Has Minimal Effects on Bacterial Communities of Thermotolerant Symbiodinium Cultures
 Erika M. Díaz-Almeyda1,2*
Erika M. Díaz-Almeyda1,2*  Tyrone Ryba1
Tyrone Ryba1  Aki H. Ohdera2,3
Aki H. Ohdera2,3  Shannon M. Collins1,4
Shannon M. Collins1,4  Natali Shafer1
Natali Shafer1  Caroline Link1
Caroline Link1  Marcela Prado-Zapata1
Marcela Prado-Zapata1  Cara Ruhnke1
Cara Ruhnke1  Meredith Moore1,5
Meredith Moore1,5  A. M. González Angel4
A. M. González Angel4  F. Joseph Pollock6
F. Joseph Pollock6  Monica Medina4
Monica Medina4- 1New College of Florida, Sarasota, FL, United States
- 2The Pennsylvania State University, University Park, PA, United States
- 3California Institute of Technology, Pasadena, CA, United States
- 4Department of Biology, University of North Texas, Denton, TX, United States
- 5Department of Biological Sciences and Chemistry, University of Southern California, Los Angeles, CA, United States
- 6The Nature Conservancy, Hawai’i & Palmyra Program, Honolulu, HI, United States
Algae in the dinoflagellate family Symbiodiniaceae are endocellular photosymbionts of corals and other cnidarians. This close relationship is disrupted when seawater temperature increases, causing coral bleaching eventually affecting entire coral reefs. Although the relationship between animal host and photosymbiont has been well-studied, little is known about the bacterial community associated with Symbiodiniaceae in culture. We compared the microbial communities of three isolates from different species of the genus Symbiodinium (formerly known as Symbiodinium clade A) with different ecophysiology, levels of interaction with the animal host, and thermal adaptations. Two species, Symbiodinium microadriaticum and Symbiodinium necroappettens, exhibit intermediate thermotolerance, with a decrease of both growth rate and photochemical efficiency with increased temperature. The third species, Symbiodinium pilosum, has high thermotolerance with no difference in growth rate or photochemical efficiency at 32°C. Microbial communities were characterized after 27 days of growth under control (26°C) and high temperature (32°C). Data shows stronger grouping of bacterial assemblages based on Symbiodinium species than temperature. Microbial communities did not group phylogenetically. We found a shared set of fifteen ASVs belonging to four genera and three families that remained in all three Symbiodiniaceae species. These included Labrenzia, Phycisphaeraceae (SM1A02), Roseovarius, and Muricauda, which are all commonly associated with corals and Symbiodiniaceae cultures. Few ASVs differed significantly by temperature within species. S. pilosum displayed significantly lower levels of microbial diversity and greater individual variability in community composition at 32°C compared to 26°C. These results suggest that bacteria associated or co-cultured with thermotolerant Symbiodinium might play an important role in thermotolerance. Further research on the functional metabolic pathways of these bacteria might hold the key to understanding Symbiodinium’s ability to tolerate thermal stress.
Introduction
The holobiont, a host and its associated microbial community, is a concept that we are beginning to understand is of crucial importance for host health, and in some cases for ecosystem health as well. Not only does the holobiont serve as a unit for natural selection, but each constitutive member can increase the plasticity of the whole by their physiological responses to the environment (Ye et al., 2019). For example, lichen microbiota may shift their metabolism to a fasting state under dry conditions, thereby increasing holobiont tolerance to drought stress (Cernava et al., 2019). Furthermore, the resilience of symbiotic microbial communities can directly affect host fitness. Certain bacteria found growing in the roots of desert plants were found to promote drought resistance in wheat (Zhang et al., 2020). While animal-microbe and plant-microbe holobionts have been intensely studied (Bronstein, 2015; Sitaraman, 2015; Hacquard, 2016; Hassani et al., 2018; Mayoral-Peña et al., 2020), microbe-microbe interactions such as phytoplankton-bacteria, mycorrhiza-helper bacteria, and symbiotic bacteriophages (Rodriguez et al., 2008; Seymour et al., 2017) are only recently being recognized as important components of the holobiont. The coral holobiont encompasses the animal (cnidarian host), its endosymbiotic photosymbionts, bacteria, archaea, fungi, and viruses (Thurber et al., 2009). Coral-associated microorganisms are critical to host fitness and survival (Glasl et al., 2016; Sweet and Bulling, 2017). Coral holobionts are highly sensitive to increases in temperature, with only a +1°C increase above long-term summer maximum temperature resulting in bleaching (i.e., disruption of symbiosis between the coral and endosymbiotic dinoflagellate), and increasingly, in coral mortality (Hoegh-Guldberg, 1999; Webster et al., 2016; McDevitt-Irwin et al., 2017; Peixoto et al., 2017).
A growing body of research has started to reveal the role of bacteria in the coral holobiont (Bourne et al., 2016; van Oppen and Blackall, 2019). They can protect the host against pathogenic microbes by preventing colonization through the physical occupation of their potential niches (Ritchie and Smith, 2004). Coral-associated bacteria are known to contribute to carbon cycling (Kimes et al., 2010), sulfur cycling (Wegley et al., 2007; Raina et al., 2009; Tandon et al., 2020), phosphorus fixation, metal homeostasis, organic remediation, secondary metabolism (Zhang et al., 2015), and production of antibiotics (Ritchie, 2006). Changes in environmental stress can alter the fitness advantages of having such functional traits represented in the microbiome. The Coral Probiotic Hypothesis proposed by Reshef et al. (2006) posits that when environmental conditions are altered, a coral’s populations of associated bacteria will undergo rapid change that effectively establishes a new, context-dependent coral holobiont. The modified bacterial community is thought to provide an advantage to cope with the new environment (i.e., warmer water). Recently, experiments involving transplantation of bacterial consortia showed improvements to coral host physiology and temperature tolerance (Doering et al., 2021). Elucidating microbial community dynamics during thermal stress and holobiont acclimatization is increasingly important in understanding the long-term persistence of coral reefs in the face of global climate change (Bellantuono et al., 2012).
For decades, dinoflagellate algae of the family Symbiodiniaceae have been recognized as the driving force behind the high productivity of coral reefs (Freudenthal, 1962; Hoegh-Guldberg et al., 2007; Roth, 2014). Symbiodiniaceae can supply up to 96% of its host’s energy budget (Muscatine, 1990) and may help with other important functions such as irradiation and thermal tolerance, with different assemblages being associated with different light and temperature conditions (Rowan et al., 1997; Rowan, 2004; Goulet et al., 2005; Oliver and Palumbi, 2011). Symbiodiniaceae therefore play a key role in coral microbiomes, and likely in their resilience to environmental disturbance as well (Trench and Blank, 1987; Warner and Suggett, 2016). Although the roles of Symbiodiniaceae in coral reefs have been fairly well-documented, these dinoflagellates harbor microbial communities of their own that remain poorly understood (Matthews et al., 2020). Taking advantage of the ability to grow these algae in culture, only three studies have aimed to identify the core microbiome of Symbiodiniaceae (Lawson et al., 2018; Camp et al., 2020; Maire et al., 2021). These studies used highly evolutionarily divergent lineages with diverse physiology, finding little overlap across isolates as well as closely related, conspecific strains. While such studies offer great insight into the similarities and differences between microbial communities of Symbiodiniaceae species with differing physiologies, comparing divergent taxa (what now includes multiple genera) of Symbiodiniaceae overgeneralized the ecology and evolution of diverse lineages (LaJeunesse et al., 2018). Comparing closely-related species with similar physiology can allow for a more detailed understanding of the ecological and evolutionary mechanisms of host-symbiont interactions.
Understanding symbiotic microbial communities holistically is critical to conservation and restoration efforts, as microbial communities may influence holobiont fitness in the face of highly stressful disturbances such as climate change (Putnam et al., 2017). In order to predict future shifts and mitigate coral reef decline, discerning shifts in host- and photosymbiont-associated microbial species during and after thermal stress is crucial. This study aims to further characterize the microbes associated with three thermotolerant photosymbionts (two intermediately thermotolerant and one highly thermotolerant) within the genus Symbiodinium (formerly known as Clade A) (Díaz-Almeyda et al., 2017). Herein, we compare three closely related species of Symbiodinium with known thermotolerance, grown under normal and high temperature conditions. We hypothesize that phylogenetically related species of Symbiodinium (e.g., Symbiodinium pilosum, Symbiodinium microadriaticum, and Symbiodinium necroappettens) with relatively similar thermotolerance will have overlapping microbial communities. We observe bacterial assemblages strongly grouping according to Symbiodinium species, with these communities remaining stable under high temperature conditions. We are yet to determine if the bacteria present in each group have redundant functions.
Materials and Methods
Sample Culture, Collection, and DNA Extraction
Algal isolates of three species of Symbiodinium, formerly known as clade A, were used for this study (Table 1). Cultures were grown in ASP-8A media [pH 8.5, 35ppt, Blank (1987)] at control (26°C) and high (32°C) temperature conditions. Cultures were grown on Percival incubators and placed on a glass shelf on top of full-spectrum fluorescent lights at 100 μmol quanta m–2 s–1 facing up, with a 12:12 light/dark cycle photoperiod. Light intensities were measured inside a 300 ml culture flask with media using a 4π sensor (Biospherical, United States). Cultures were maintained by adjusting cell density to 1 × 105 cells ml–1 every 10 days by adding fresh media, acclimated to the control temperature (26°C) for at least 2 months (Supplementary Figure 1A). To measure cell density, a sterile rubber policeman was used to scrape all cells from the flask walls to ensure collection of all cells. The culture was homogenized manually and 1 ml aliquot was collected into a 2 ml screwcap tube. To detach cell aggregations, lugol was added (200 μl) and tubes were vortexed for 5 min. Cells were then counted manually with a hemocytometer. To begin the experiment, replicate 15 ml clear glass tubes containing 8 ml of culture with 1 × 105 cells ml–1 were placed vertically on top of a glass shelf with full spectrum lights facing up, in an incubator set at 26 or 32°C (n = 5 per isolate per temperature) (Supplementary Figure 1B). Tubes were undisturbed until after 27 days of incubation, when Fv/Fm was measured for all replicates (n = 5 per isolate per temperature) and cells were collected for DNA extraction (n = 3 per isolate per temperature). Cells were collected by scraping all cells attached to the glass tube with a disposable inoculating loop and centrifuged at approximately 2,000 g for 1 min, removing the supernatant and flash freezing with liquid nitrogen immediately. DNA was isolated using the MO BIO PowerSoil DNA isolation Kit following manufacturer’s instructions. A previous study established that S. microadiaticum (strain CassKB8) and S. necroappetens (strain RT80) have intermediate thermotolerance, since they were able to grow at high temperature but not as well as at normal temperature. S. pilosum (strain RT130) grew at the same rate in normal and high temperature conditions, suggesting that it has higher thermotolerance (Díaz-Almeyda et al., 2017). S. pilosum and S. necroappetens were collected in the Caribbean and are part of the same isolate collection (formerly known as Trench collection) while S. microadriaticum was isolated in the North Pacific (formerly known as BURR collection) (Table 1).
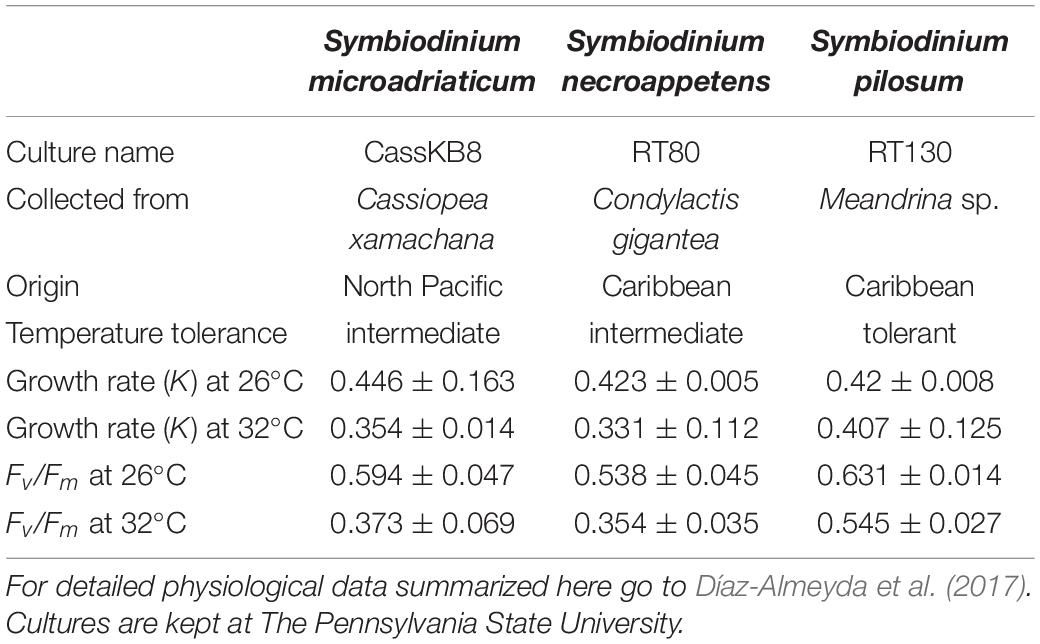
Table 1. Cultures used in this study, invertebrate species from which the culture was isolated, and geographic location where sample was collected.
16S rRNA Gene Amplification and Sequencing
Samples were amplified using specific primers pairs (Caporaso et al., 2011) for the V4 region of the 16S rRNA gene. A 25 μL PCR reaction was prepared with 10 μL 5Prime HotMaster Mix (QuantaBio), 0.5 μL forward primer 515F (GTGCCAGCMGCCGCGCGGTAA) (10 μM), 0.5 μL reverse primer 806R (GGACTACHVGGGTWTCTAAT) (10 μM), 1 μL of BSA (10 mg/ml), 1 μL template DNA (10 ng), and 12 μL PCR-grade water. PCR amplifications consisted of a 3 min denaturation at 94°C; 30 cycles of 45 s at 94°C, 60 s at 50°C and 90 s at 72°C; and 10 min at 72°C. Samples were submitted for sequencing to the Department of Energy’s Joint Genome Institute (JGI) and were processed according to their iTag sequencing protocol. Individual libraries were prepared and barcoded before being multiplexed and sequenced on an Illumina MiSeq sequencer machine in a 2 × 300 run mode.
Sequence Generation and Processing
Sequences were demultiplexed and processed according to the JGI’s iTag amplicon sequencing protocol for taxonomic identification. Initial processing of 16S rRNA gene sequence libraries was performed using the Quantitative Insights Into Microbial Ecology (QIIME2; 2019.7.0) package (Bolyen et al., 2019). Read trimming, filtering, denoising, and amplicon sequence variant (ASV) inference was performed with the DADA2 implementation of QIIME2 (Callahan et al., 2016). Taxonomy was assigned to ASVs using the Naive Bayes classifier pre-trained on the Silva database version 138 clustered at 99% sequence similarity and optimized for the 515F/806R primer pairs. Extraction blanks and template PCR controls were not included as part of our sequencing effort. We therefore applied additional filters to the reads to remove possible contaminants. Initially, ASVs were removed if they were not present in a minimum of three of the 18 samples sequenced. Additionally, low abundance ASVs totaling less than 63 reads (0.2 percent of the total reads from the sample with the fewest reads) were removed from each sample. The additional filtering removed rare ASVs, many of which were present in a single sample and highly likely to be contamination of the culture. Filtering reduced the total ASV count from 255 to 124. Phylogenetic diversity metrics were generated with alignments performed with MAFFT and trees generated with FastTree (Supplementary File 1), which was followed by alpha and beta diversity calculations.
Differential Abundance and Distance Contribution
Differential abundance testing was performed with DESeq2 [v1.32.0, Love et al. (2014)], against sample abundances normalized by the geometric mean of positive counts. Fold change values were adjusted for dispersion using the adaptive shrinkage estimator from the ashr package [v2.2-47, Stephens et al. (2022)], and ASVs differentially abundant beyond a false discovery rate of 0.01 were collected (Supplementary Table 1). Contributions of each ASV to Bray-Curtis dissimilarity were calculated for using the SIMPER function from the vegan package (v2.5-7) (Supplementary Table 2). Permutational multivariate analysis of variance (PERMANOVA, 999 permutations) was performed as well (Supplementary Table 3). Initially, samples were rarefied to 30,000 reads and pairwise comparisons were performed between 26 and 32°C (Supplementary Table 4).
Data Visualization
Data visualization, including PCoA plots of unweighted and weighted UniFrac distance results calculated in QIIME2 were generated with qiime2R (v0.99.6)1 and phyloseq [v1.36.0, McMurdie and Holmes (2013)] R packages (RStudio Team, 2020). Overlapping taxa (grouped at the genus level) between species and temperature were identified using the online tool from VIB/Ugent Bioinformatics and Evolutionary Genomics Website.2
Results
Bacterial Communities Are Specific to Symbiodinium Species
The microbial communities of each Symbiodinium sp. were characterized via sequencing of the V4 region of the 16S rRNA gene. We recovered an average of 50,623 raw reads per sample. After filtering and processing the sequences, the average number of sequences per sample was 48,507, with a minimum of 31,090 and maximum of 68,716. After removal of low abundance taxa, we identified 124 bacterial ASVs across all samples and we summarized them at the genus level by replicate (Supplementary Figure 2) and by treatment and species (Supplementary Figure 3). Samples grouped largely by Symbiodinium species when clustered by Bray-Curtis dissimilarity (Figure 1). Permutational multivariate analysis of variance (PERMANOVA) confirmed diversity composition significantly varied by species (999 permutations, q-value < 0.05), indicating Symbiodinium species harbor specific microbial communities (Supplementary Table 3).
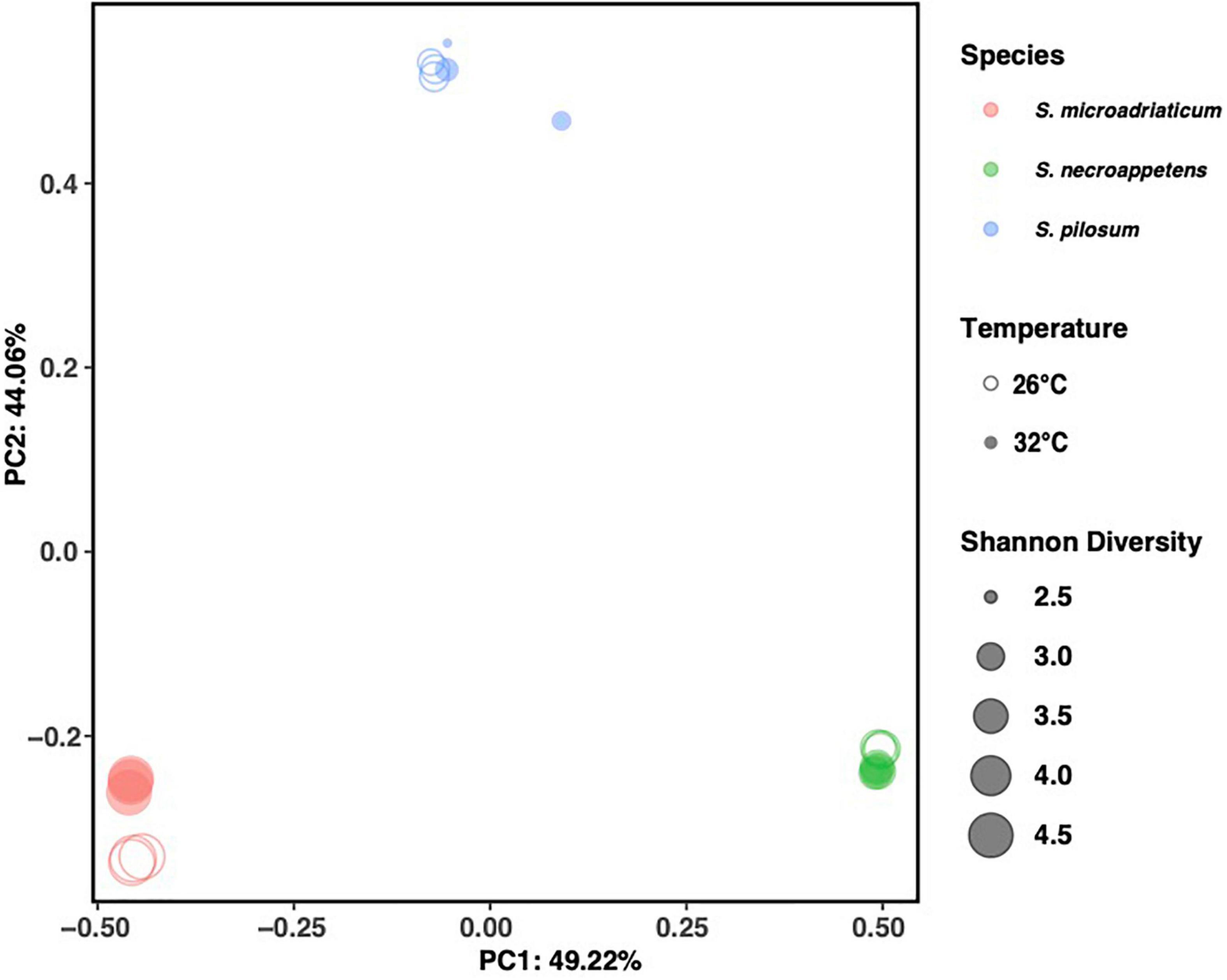
Figure 1. Principal coordinate analysis (PCoA) plot of bacterial communities, with three replicates for Symbiodinium pilosum, Symbiodinium microadriaticum, and Symbiodinium necroappettens under 26 and 32°C conditions. All data shows strong bacterial assemblages grouped on Symbiodinium species rather than temperature.
Comparison of community composition with SIMPER identified less than 20 ASVs driving 85% of the observed dissimilarity between species (Supplementary Table 2). Additionally, OM182_clade, which accounted for the greatest percentage of dissimilarity between S. microadriaticum and S. necroappetens, was absent from S. microadriaticum, while present in both S. necroappetens and S. pilosum (Figure 2). Labrenzia accounted for 18.9% of dissimilarity between S. pilosum and both other Symbiodinium species. These findings were further corroborated by differential abundance analysis with DESeq2. We found the aforementioned taxa to be differentially abundant between Symbiodinium species (Supplementary Table 1).
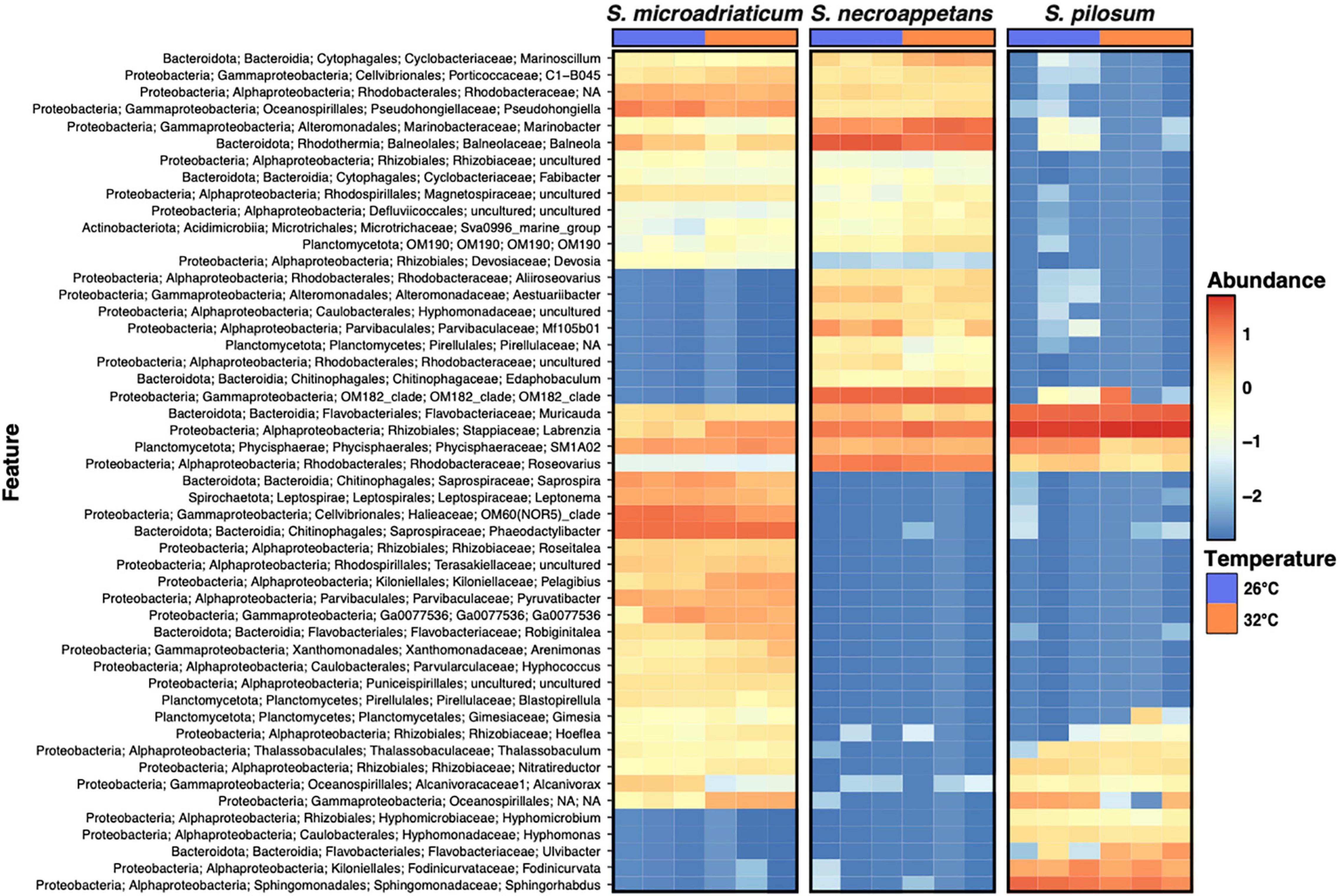
Figure 2. Heatmap of relative abundances calculated in log10 for the top 50 most abundant ASVs.
A closer inspection of these ASVs revealed that they belong to a small subset of the genera found across the three species, including Muricauda, Balneola, Labrenzia, Phaeodactylibacter, Sphingorhabdus, and OM182_clade, each with >8% influence in respective species comparisons (Figure 3 and Supplementary Table 2). Notably, four genera (Muricauda, Labrenzia, Roseovarius, and the uncultured SM1A02 group of the family Phycisphaeraceae) were commonly found across the three Symbiodinium species at both 26 and 32°C, with each dinoflagellate taxa associating with presumably a unique strain represented by a single ASV.
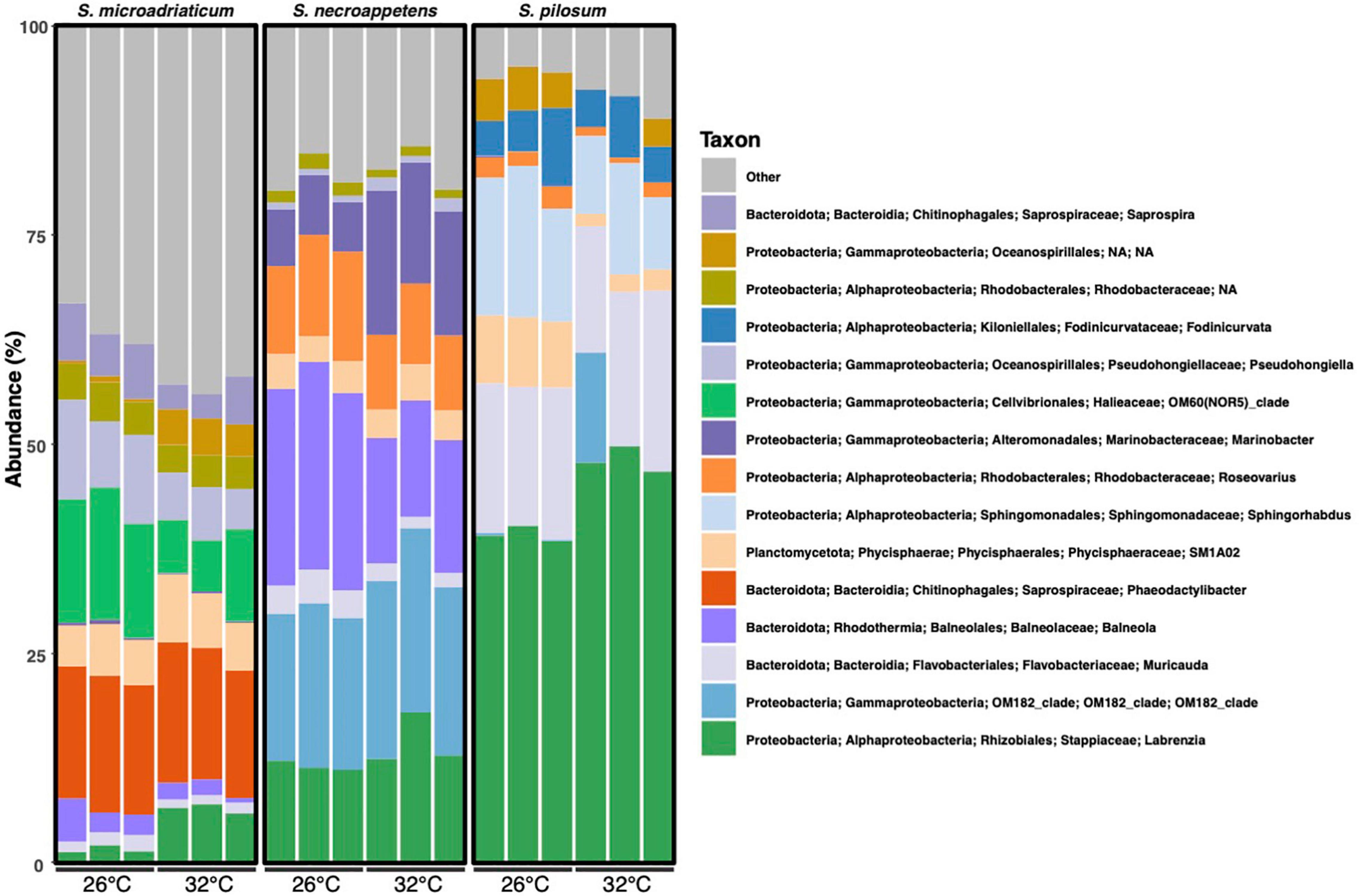
Figure 3. Bacterial community composition (relative abundance in percentage) of Symbiodinium species in culture under 26 and 32°C at the genus level, based on the 16S rRNA gene v4 region amplification. The relative bacterial abundance is similar within Symbiodinium species, independent of temperature (n = 18).
Symbiodinium microadriaticum harbored the greatest number of unique microbial genera (37 at 26°C and 36 at 32°C), whereas S. pilosum harbored the least, with seven species specific genera at both 26 and 32°C (Figure 4). No additional genera were shared between S. pilosum and S. necroappetens beyond the aforementioned four taxa, whereas S. microadriaticum and S. necroappetens shared 13 to 14 genera at 26 and 32°C, respectively.
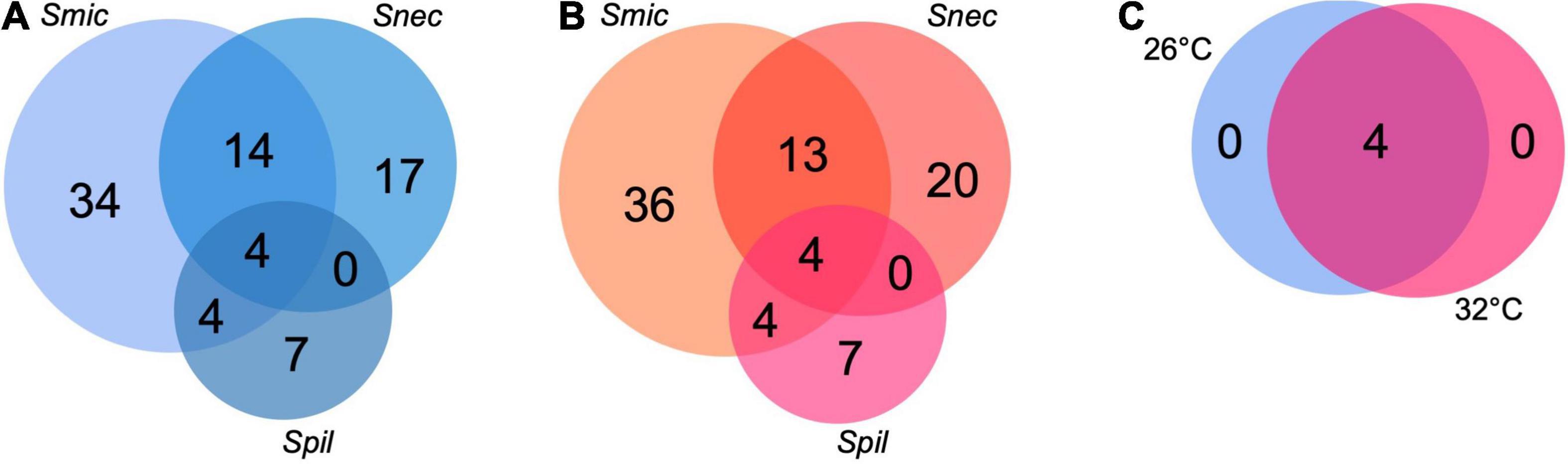
Figure 4. Shared number of genera present in each core microbiome for two temperatures, independent of Symbiodinium species (A,B). Combined core microbiome identified at 26°C and 32°C, across the three species of Symbiodinium (C). To be considered part of the core microbiome at each temperature, the OTU must be present in all replicates (n = 18). See Supplementary Figures 2, 3 for details.
While the four shared genera were often found to exhibit high relative abundance within the samples, other taxa were represented in greater abundance within each Symbiodinium species. In S. microadriaticum, Phaeodactylibacter (15.21–16.75%) and usually a member of the OM60(NOR5) clade of Halieaceae (6.08–15.80%) were the dominant genera. In S. necroappetens, OM182_clade of Gammaproteobacteria (17.51–21.96%), Balneola (13.89–24.78%), Marinobacter (5.89–17.18%), and Labrenzia (11.14–18.01%) were the dominant genera. In S. pilosum, Muricauda (15.09–21.61%), Sphingorhabdus (8.60–18.06%), and Labrenzia (38.32–49.74%) were the dominant genera (Figure 3).
Diversity of Intermediately-Thermotolerant Symbiodinium Remains Relatively Stable Under Elevated Temperatures
Shannon Diversity Index (H’) values were calculated for each sample (Figure 5). S. microadriaticum had the highest average H’ with no significant differences between temperatures (4.67 at 26°C and 4.78 at 32°C), followed by S. necroappetens (H’ = 3.60 at 26°C and 3.61 at 32°C). S. pilosum had the lowest average H’ of 3.04 at 26°C and 2.67 at 32°C. One-way ANOVA analysis showed significant differences between species [F(5,12) = 218.3432, p < 0.0001] but only S. pilosum had a significant change in H’ between the two temperatures (Tukey HSD, p = 0.0053).
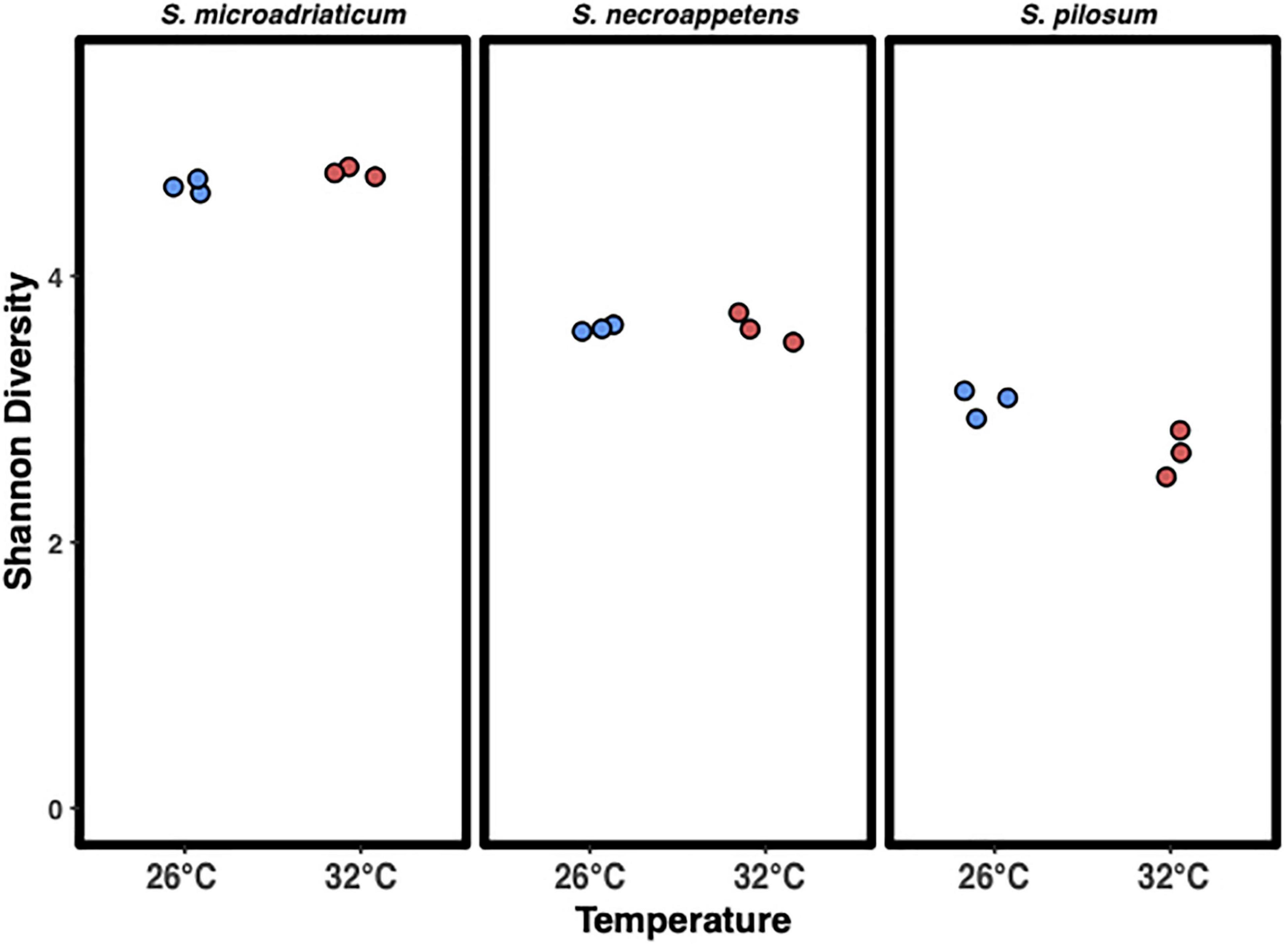
Figure 5. Shannon diversity index of microbial communities associated with Symbiodinium in culture. Blue 26°C and red 32°C. There was a significant effect of species and temperature on the Shannon diversity index at the p < 0.0001 level for the three conditions with Symbiodinium pilosum being the only species different by temperature [F(5,12) = 154.07, p < 0.0001].
Our samples did not cluster well by temperature (q-value > 0.05). We did, however, find ASVs that were significantly differentially abundant between 26 and 32°C. Seventeen ASVs were differentially abundant in cultures of S. microadriaticum. Two of the differentially abundant taxa belonged to the shared genera Labrenzia and Muricauda, and increased by 8- and 2-fold (log2), respectively. In S. necroappetens, both bacterial genera exhibited differential abundance, with Labrenzia increasing 5-fold (log2) and Muricauda decreasing in relative abundance by 5-fold (log2). Nine ASVs were differentially abundant in S. pilosum, although none belonged to the four genera commonly found between the three dinoflagellate species.
Discussion
We compared the bacterial communities of closely related Symbiodinium isolates that have been previously characterized with similar thermotolerance in culture (Díaz-Almeyda et al., 2017). Similar to previous studies, we found microbial communities of Symbiodinium clustered primarily by species (Lawson et al., 2018; Camp et al., 2020; Maire et al., 2021). Of the three Symbiodinium species included in our study, two intermediately thermotolerant species (S. necroappetens and S. microadriaticum) had greater microbial diversity than the highly thermotolerant species (S. pilosum). The community composition of S. necroappetens remained relatively stable at 32°C, while that of S. microadriaticum clustered separately by temperature, and the community composition of S. pilosum became more dispersed at 32°C (Figure 1). Relatively few ASVs differed by temperature, and there were no ASVs that consistently increased or decreased in abundance across all species, although we detail why some of them may be notable. More evidence is needed to determine whether differences in the relative abundance of any thermally differing taxa influence Symbiodinium thermotolerance (or vice-versa). Our results demonstrate that even closely related Symbiodinium species with similar physiology are host to unique assemblages of microbes which may differ in their responses to increased temperature.
Symbiodinium Species Share Few Associated Bacterial Genera
All three Symbiodinium species shared members of Muricauda (Flavobacteriaceae), Labrenzia (Rhodobacteraceae), Roseovarius (Rhodobacteraceae), and a member of Phycisphaeraceae (SM1A02). These findings are consistent with previous studies of Symbiodiniaceae-associated microbial communities (Lawson et al., 2018; Camp et al., 2020). Importantly, although microbial communities grouped by Symbiodinium species, they did not group phylogenetically by Symbiodinium species. Rather, they grouped by location and collection of origin (Supplementary Figure 4). S. necroappetens and S. microadriaticum are more closely related phylogenetically than S. pilosum (LaJeunesse et al., 2015). However, S. necroappetens had a microbial community more similar to S. pilosum than to S. microadriaticum. S. necroappetens was isolated by Robert K. Trench at least 20 years ago (LaJeunesse, 2001), S. pilosum was isolated at least 27 years ago (Iglesias-Prieto and Trench, 1994), and both were isolated from the Caribbean by the same laboratory with protocols which included adding antibiotics and using a low nutrient culture media to remove bacteria (Schoenberg and Trench, 1980). This may explain the low bacterial diversity we observed in S. pilosum. In contrast, S. microadriaticum was isolated at least 35 years ago by a different laboratory using f/2 media (Taguchi and Kinzie, 2001). It is therefore unknown whether grouping of bacterial communities is due to long-term culturing artifacts or true symbiotic associations, as these differing protocols may have resulted in very different founder effects, leading to the differences we observed. Maire et al. (2021) found in situ samples of Symbiodiniaceae differed greatly from corresponding cultures, suggesting that co-culturing may have influenced our results rather than true symbiosis. The same study also found the presence of intracellular bacteria in Symbiodiniaceae to be common, diverse, and highly distinct from extracellular bacteria. We are unable to separately characterize the responses of intracellular and extracellular communities to thermal stress because we did not distinguish these communities. With these considerations, our results provide insight into the potential connection between Symbiodinium thermotolerance and associated bacterial communities, and point toward future studies on in situ Symbiodinium that distinguish between closely- and loosely-associated bacteria.
Microbial Community Response to Temperature Stress May Correlate With Host Thermotolerance
Bacterial diversity of S. microadriaticum and S. necroappetens remained stable at high temperatures while that of S. pilosum was significantly lower at 32°C. Stable H’ values across different environmental conditions can indicate a lack of substantive change in community composition, or that the combined richness and evenness of the community is relatively stable (Kimbro and Grosholz, 2006; Willis, 2019). The flexibility that we observed in S. pilosum may be related to its higher thermotolerance, as acclimating to rapid changes in environmental conditions has been associated with dramatic shifts in bacterial community composition (Vargas et al., 2020; Voolstra and Ziegler, 2020). In contrast to our results, Camp et al. (2020) found that the microbiome of Durusdinium trenchii, a thermotolerant Symbiodiniaceae, remained stable at high temperatures.
The bacterial communities of S. microadriaticum and S. pilosum also clustered by temperature. S. necroappetens samples clustered together but not separately by temperature, most likely due to experiencing a much smaller change with temperature. These results support the idea that microbiome responses to heat stress are unique to Symbiodinium species, perhaps regardless of dinoflagellate thermotolerance. Our results also suggest that increased thermotolerance may correlate with lower levels of microbial diversity and greater individual variability in community composition. According to the “Anna Karenina principle,” the increased prevalence of environmental stressors may reduce a host’s ability to regulate its microbiome composition (Zaneveld et al., 2017). While our results did not offer evidence for this principle, if well-supported with future research, the Anna Karenina principle has strong implications for coral reefs in the context of climate change, as a strong link between rising ocean temperatures and abundance of coral pathogens within communities of coral-associated microbes has been described (Tout et al., 2015), and microbial communities without functional redundancy can be subject to environmental stressors such as ocean warming (Voolstra and Ziegler, 2020). Further research using controlled studies to investigate this relationship may find a link between the thermotolerance of a microbial community and instability of the holobiont under high temperatures. The potential benefits of greater flexibility and diversity in microbiome community composition for thermotolerance should be further explored in future studies using larger sample sizes and other Symbiodinium species. Additionally, while thermotolerance is known to be affected by other factors such as genetics, environmental factors, and host-symbiont type mismatches (Brown et al., 2002; Mieog et al., 2009), the degree of contribution (positive or negative) by certain microbial taxa, their potential symbiotic interactions with Symbiodinium, and their relationships to Symbiodinium thermotolerance remain unexplored and should be considered for future studies.
Notable ASVs and Their Potential Roles in Symbiodinium Thermotolerance
No ASVs consistently increased with temperature across all Symbiodinium species. Rather, different ASVs were differentially abundant by temperature for each Symbiodinium species. While some or all of these differences may be a result of long-term culturing, we offer insights on the potential importance of key ASVs for Symbiodinium thermotolerance.
We found Labrenzia to be particularly abundant across all Symbiodinium species, but especially in S. pilosum, the most thermotolerant species. Additionally, Labrenzia were not differentially abundant between temperature treatments in S. pilosum but were significantly less abundant at 32°C in the two intermediately thermotolerant Symbiodinium. Labrenzia are known for their ability to produce dimethylsulfoniopropionate, which scavenges reactive oxygen species and thus may play a role in increasing Symbiodinium stress tolerance (Diaz et al., 2016; Curson et al., 2017). It is also plausible that these bacteria could be directly reducing pathogenic microbes, which might opportunistically take advantage of heat stressed Symbiodinium. An oyster-colonizing Labrenzia sp. exhibited broad antimicrobial activity and reduced the growth of a pathogenic Roseovarius (Amiri Moghaddam et al., 2018), and labrenzbactin, a siderophore produced by a species of Labrenzia, has been shown to exhibit antimicrobial activity in vitro (Sharma et al., 2019). However, its effect on Symbiodinium-specific pathogens has not yet been explored. The increased prevalence of Labrenzia in S. pilosum and its relatively stable abundance with temperature stress may be related to thermotolerance, or it may be a result of having lower levels of diversity in general. Research on the ecological and physiological roles of Labrenzia for Symbiodinium fitness is necessary to discern whether these bacteria are generally mutualistic, parasitic, or commensal partners in heat-stressful conditions.
Roseovarius were also present in all three host species, in much lower relative abundances than Labrenzia, and was highest in S. necroappetens. Other studies have similarly found Roseovarius in relatively low abundance with Symbioiniaceae (Camp et al., 2020; Maire et al., 2021). Notably, Pootakham et al. (2019) found that Roseovarius were one of the two most prevalent genera associated with heat-stressed corals. Members of Roseovarius have also been implicated in coral diseases such as black band disease (Ng et al., 2015). However, we did not find that Roseovarius significantly increased in relative abundance with increased temperature in Symbiodinium (Figure 2). Our results therefore suggest that Roseovarius may not play a similar role in heat stressed Symbiodinium, or perhaps only in more thermosensitive species. Further research on the functional ecology of Roseovarius with Symbiodinium under heat stress is necessary to investigate these hypotheses.
Muricauda was also shared by all three Symbiodinium species we cultured and were differentially abundant by temperature in the two intermediately thermotolerant Symbiodinium, but not in S. pilosum. Several studies have demonstrated that Muricauda are intimately associated with Symbiodiniaceae (Han et al., 2016; Lawson et al., 2018). In the first comparison of intracellular, closely-associated, and loosely-associated bacterial communities, Maire et al. (2021) found Muricauda present in most in situ samples of different Symbiodiniaceae species, but consistently in less than 1% abundance. We found Muricauda in greater than 1% abundance in all Symbiodinium, but especially in S. pilosum, in which Muricauda consistently accounted for over 15% of each sample. This discrepancy may be the result of culturing, as cultured samples have previously been found to differ substantially from in situ Symbiodiniaceae samples (Maire et al., 2021). Regardless of its potentially low abundance in wild Symbiodiniaceae, Muricauda is one of 12 bacterial genera ubiquitously associated with various reef-building corals across the globe, suggesting its functional importance not only to Symbiodiniaceae but also to the coral holobiont (Bernasconi et al., 2019). Furthermore, Muricauda may serve a functional role in Symbiodinium heat resistance. A close relative to Muricauda lutaonensis enhanced thermal and light stress resistance of cultured Symbiodiniaceae (Motone et al., 2020). We did not find significant differences in Muricauda by temperature for any Symbiodinium species, but in situ sequencing may indicate whether they play a key role in Symbiodinium thermotolerance.
Sphinghorabdus were one of the dominant taxa in S. pilosum but were not present in greater than 1% abundance in the intermediately thermotolerant host species (Figure 3). Sphinghorabdus have been isolated from Gorgonian coral and have been successfully cultivated (Keller-Costa et al., 2017; Silva et al., 2018), but otherwise are not well-known, so further research may reveal their importance for Symbiodinium thermotolerance.
In S. pilosum, a member of Phycisphaeraceae (SM1A02) accounted for the greatest amount of community dissimilarity between temperature treatments, displaying greater abundance at 26°C. This may be due to strong competitive pressure from the highly dominant Labrenzia in thermally-stressed S. pilosum, as ASVs from Phycisphaeraceae did not significantly decrease in abundance in the other two Symbiodinium species. Kellogg (2019) found three OTUs belonging to Phycisphaerales to be bacterial associates across seven deep-sea corals (Eloe et al., 2011; Allers et al., 2013). Sequences similar to those three OTUs were not associated with tropical corals, possibly suggesting a role specific to cold-water corals (Kellogg, 2019). Contrarily, members of Phycisphaerales were associated with tropical stony corals, and in greater abundance under highly variable environmental conditions by Ziegler et al. (2017). Together with our results, these findings suggest that the observed decrease in Phycisphaeraceae with temperature may not be indicative of thermotolerance, but rather a consequence of the increase in relative abundance of Labrenzia in our most thermotolerant host.
In total, no ASVs were indicative of thermotolerance across all Symbiodinium species. Rather, we found ASVs that were differentially abundant by temperature differed by Symbiodinium species. These differences may be explained by functional redundancy of thermotolerance within and between bacterial communities (Stilianos et al., 2018). While Symbiodinium-associated bacterial communities may differ substantially, even among similar Symbiodinium species, these communities are likely to converge on key functional traits. For example, members of Rhodobacteraceae have generally been found to associate with other algae and provide vitamins and other amino acids to their hosts (Rosales et al., 2020), and play important roles in marine sulfur and nitrogen cycles (Chen et al., 2011). In microbial communities, many species fulfilling similar functions often coexist (Louca et al., 2018). Since all our cultures contained diverse members of Rhodobacteraceae, functional stability with stress could theoretically be maintained despite each culture containing different representatives of this family. For example, soil microbial communities showed high functional stability in long-term drought (Yuste et al., 2014). Future research on the relationship between phylogenetic diversity and functional stability may evidence similar consistencies in functional stability of dinoflagellate microbial communities under long-term increases in heat stress driven by climate change.
Additionally, high rates of horizontal gene transfer in long-term co-cultures could result in changes in the functional roles of different taxa, including those that may affect Symbiodinium thermotolerance (Cairns et al., 2018). This could have led to the development of similar thermotolerance-related functions in different taxa, leading to dissimilar patterns in bacterial community change with heat stress across cultures. While shifts in the functional roles of different community members have not been well-documented in long-term co-cultures, research on the human gut microbiome suggests that high rates of horizontal gene transfer and selective pressure have promoted functional redundancy in gut microbial communities despite wide diversity in microbiome community composition (Tian et al., 2020). More similar to our study system, biofilm bacterial communities of the kelp Ecklonia radiata commonly had horizontal gene transfers between bacterial members of the same class or order, and these transfers often included genes involved in stress response (Song et al., 2021). More research on Symbiodiniaceae is necessary to determine whether functional redundancy may be important for thermotolerance rather than any one bacterial taxa in particular.
Conclusion
In conclusion, we show that closely related species of Symbiodinium cultures with similar thermotolerance have specific bacterial assemblages that remain relatively stable under high temperature conditions. While differences in relative abundance of key bacterial genera were not consistent across Symbiodinium species, significant differences in certain genera by temperature treatment within Symbiodinium species suggest functional redundancy within bacterial communities. Future studies should focus on isolating symbiotic bacteria and assessing their interactions with Symbiodinium and with each other. Experimenting with axenic Symbiodinium isolates in culture with specific bacterial isolates can allow us to determine the effects of the specific microbes, which species might be essential, and whether those species can influence the physiology of Symbiodinium. Since multiple strains of Symbiodinium can be cultured and their physiology is well-known, this phylogenetic group can aid in characterizing the role of microbes in functional and physiological diversity of their symbionts. Additionally, metagenomes of bacteria associated with Symbiodinium can inform about the metabolic capabilities to hypothesize functional roles and if the interactions between these bacteria and Symbiodinium are mutualistic, neutral or pathogenic.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJNA758393.
Author Contributions
ED-A and MMe contributed to conception and design of the study. All authors contributed to the data analysis. AHO, TR, and FP performed the statistical analysis. ED-A wrote the first draft of the manuscript. All authors wrote sections of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This project was funded by CONACYT (Mexico) Ph.D. award 305321, which funded ED-A’s studies. The Pennsylvania State University MMe start up funds, funded equipment for experiments. Joint Genome Institute provided sequencing. NSF (OCE 1442206) funded reagents for experiments.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Mary Alice Coffroth (SUNY Buffalo) and Todd E. LaJeunesse for providing the algal cultures used in this study. We thank our reviewers for their insightful comments that helped improve this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.764086/full#supplementary-material
Supplementary Figure 1 | Cultures were grown on top of a glass shelf under lights facing up to prevent shading. (A) Cultures in flasks were acclimated for 2 months at 100 μM quanta m2 s–1 in a 12:12 hr/light:dark cycle. Cell densities were adjusted to 1 × 105 cells/ml before starting experiments. (B) Cultures were moved to their experimental conditions with the previous light and circadian rhythm into their respective temperatures in glass tubes. Cultures started at 1 × 105 cells/ml, Fv/Fm was measured for n = 5 on day 27, cells for DNA extraction were collected for n = 3 on day 27.
Supplementary Figure 2 | Shared number of genera present in each species core microbiome of Symbiodinium under two temperatures (26 and 32°C). Each treatment had three replicates per Symbiodinium species per temperature.
Supplementary Figure 3 | Shared number of genera present in each species core microbiome of Symbiodinium (A–C) over two temperatures (26 and 32°C). The shared core microbiome at 26 and 32°C, independent of Symbiodinium species is also represented (D). To be considered part of the core microbiome at each temperature, the genera must be present in all samples (n = 18).
Supplementary Figure 4 | Heatmap of relative abundances of bacteria. At the top, a Euclidean distance cladogram shows samples grouped by Symbiodinium species.
Supplementary Table 1 | Differential abundances calculated with DESeq2.
Supplementary Table 2 | Differential abundances calculated with SIMPER.
Supplementary Table 3 | Permanova comparisons per species (A) and per treatments (B).
Supplementary Table 4 | Number of observed features per sample after alpha rarefaction. Reads were rarefied using the Qiime2 alpha-diversity function. Rarefaction was performed to a maximum depth of 30,000 reads over 20 steps. Feature tables were generated over 10 iterations. The variability in observed features for each iteration are presented as box plots for each sampling depth.
Footnotes
References
Allers, E., Wright, J. J., Konwar, K. M., Howes, C. G., Beneze, E., Hallam, S. J., et al. (2013). Diversity and population structure of marine group a bacteria in the Northeast subarctic Pacific Ocean. ISME J. 7, 256–268. doi: 10.1038/ismej.2012.108
Amiri Moghaddam, J., Dávila-Céspedes, A., Kehraus, S., Crüsemann, M., Köse, M., Müller, C. E., et al. (2018). Cyclopropane-containing fatty acids from the marine bacterium labrenzia sp. 011 with antimicrobial and GPR84 activity. Mar. Drugs 16:369. doi: 10.3390/md16100369
Bellantuono, A. J., Hoegh-Guldberg, O., and Rodriguez-Lanetty, M. (2012). Resistance to thermal stress in corals without changes in symbiont composition. Proc. Biol. Sci. 279, 1100–1107. doi: 10.1098/rspb.2011.1780
Bernasconi, R., Stat, M., Koenders, A., and Huggett, M. J. (2019). Global networks of Symbiodinium-bacteria within the coral holobiont. Microb. Ecol. 77, 794–807. doi: 10.1007/s00248-018-1255-4
Blank, R. J. (1987). Cell architecture of the dinoflagellate Symbiodinium sp. inhabiting the Hawaiian stony coral Montipora verrucosa. Mar. Biol. 94, 143–155. doi: 10.1007/BF00392906
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Bourne, D. G., Morrow, K. M., and Webster, N. S. (2016). Insights into the coral microbiome: underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 70, 317–340. doi: 10.1146/annurev-micro-102215-095440
Brown, B., Dunne, R., Goodson, M., and Douglas, A. (2002). Experience shapes the susceptibility of a reef coral to bleaching. Coral Reefs 21, 119–126. doi: 10.1007/s00338-002-0215-z
Cairns, J., Ruokolainen, L., Hultman, J., Tamminen, M., Virta, M., Hiltunen, T., et al. (2018). Ecology determines how low antibiotic concentration impacts community composition and horizontal transfer of resistance genes. Commun. Biol. 1:35. doi: 10.1038/s42003-018-0041-7
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Camp, E. F., Kahlke, T., Nitschke, M. R., Varkey, D., Fisher, N. L., Fujise, L., et al. (2020). Revealing changes in the microbiome of Symbiodiniaceae under thermal stress. Environ. Microbiol. 22, 1294–1309. doi: 10.1111/1462-2920.14935
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U.S.A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Cernava, T., Aschenbrenner, I. A., Soh, J., Sensen, C. W., Grube, M., and Berg, G. (2019). Plasticity of a holobiont: desiccation induces fasting-like metabolism within the lichen microbiota. ISME J. 13, 547–556. doi: 10.1038/s41396-018-0286-7
Chen, Y., Patel, N. A., Crombie, A., Scrivens, J. H., and Murrell, J. C. (2011). Bacterial flavin-containing monooxygenase is trimethylamine monooxygenase. Proc. Natl. Acad. Sci. U.S.A. 108, 17791–17796. doi: 10.1073/pnas.1112928108
Curson, A. R. J., Liu, J., Bermejo Martinez, A., Green, R. T., Chan, Y., Carrión, O., et al. (2017). Dimethylsulfoniopropionate biosynthesis in marine bacteria and identification of the key gene in this process. Nat. Microbiol. 2:17009. doi: 10.1038/nmicrobiol.2017.9
Diaz, J. M., Hansel, C. M., Apprill, A., Brighi, C., Zhang, T., Weber, L., et al. (2016). Species-specific control of external superoxide levels by the coral holobiont during a natural bleaching event. Nat. Commun. 7:13801. doi: 10.1038/ncomms13801
Díaz-Almeyda, E. M., Prada, C., Ohdera, A. H., Moran, H., Civitello, D. J., Iglesias-Prieto, R., et al. (2017). Intraspecific and interspecific variation in thermotolerance and photoacclimation in Symbiodinium dinoflagellates. Proc. Biol. Sci. 284:20171767. doi: 10.1098/rspb.2017.1767
Doering, T., Wall, M., Putchim, L., Rattanawongwan, T., Schroeder, R., Hentschel, U., et al. (2021). Towards enhancing coral heat tolerance: a “microbiome transplantation” treatment using inoculations of homogenized coral tissues. Microbiome 9:102. doi: 10.1186/s40168-021-01053-6
Eloe, E. A., Shulse, C. N., Fadrosh, D. W., Williamson, S. J., Allen, E. E., and Bartlett, D. H. (2011). Compositional differences in particle-associated and free-living microbial assemblages from an extreme deep-ocean environment. Environ. Microbiol. Rep. 3, 449–458. doi: 10.1111/j.1758-2229.2010.00223.x
Freudenthal, H. D. (1962). Symbiodinium gen. nov. and Symbiodinium microadriaticum sp. nov., a Zooxanthella: Taxonomy, Life Cycle, and Morphology. J. Protozool. 9, 45–52. doi: 10.1111/j.1550-7408.1962.tb02579.x
Glasl, B., Herndl, G. J., and Frade, P. R. (2016). The microbiome of coral surface mucus has a key role in mediating holobiont health and survival upon disturbance. ISME J. 10, 2280–2292. doi: 10.1038/ismej.2016.9
Goulet, T. L., Cook, C. B., and Goulet, D. (2005). Effect of short-term exposure to elevated temperatures and light levels on photosynthesis of different host-symbiont combinations in the Aiptasia pallida/Symbiodinium symbiosis. Limnol. Oceanogr. 50, 1490–1498. doi: 10.4319/lo.2005.50.5.1490
Hacquard, S. (2016). Disentangling the factors shaping microbiota composition across the plant holobiont. New Phytol. 209, 454–457. doi: 10.1111/nph.13760
Han, J., Zhang, L., Wang, S., Yang, G., Zhao, L., and Pan, K. (2016). Co-culturing bacteria and microalgae in organic carbon containing medium. J. Biol. Res. 23:8. doi: 10.1186/s40709-016-0047-6
Hassani, M. A., Durán, P., and Hacquard, S. (2018). Microbial interactions within the plant holobiont. Microbiome 6:58. doi: 10.1186/s40168-018-0445-0
Hoegh-Guldberg, O. (1999). Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 50, 839–866. doi: 10.1071/MF99078
Hoegh-Guldberg, O., Mumby, P. J., Hooten, A. J., Steneck, R. S., Greenfield, P., Gomez, E., et al. (2007). Coral reefs under rapid climate change and ocean acidification. Science 318, 1737–1742. doi: 10.1126/science.1152509
Iglesias-Prieto, R., and Trench, R. K. (1994). Acclimation and adaptation to irradiance in symbiotic dinoflagellates. I. Responses of the photosynthetic unit to changes in photon flux density. Mar. Ecol. Prog. Ser. 113, 163–175. doi: 10.3354/meps113163
Keller-Costa, T., Eriksson, D., Gonçalves, J. M. S., Gomes, N. C. M., Lago-Lestón, A., and Costa, R. (2017). The gorgonian coral Eunicella labiata hosts a distinct prokaryotic consortium amenable to cultivation. FEMS Microbiol. Ecol. 93:fix148. doi: 10.1093/femsec/fix143
Kellogg, C. A. (2019). Microbiomes of stony and soft deep-sea corals share rare core bacteria. Microbiome 7:90. doi: 10.1186/s40168-019-0697-3
Kimbro, D. L., and Grosholz, E. D. (2006). Disturbance influences oyster community richness and evenness, but not diversity. Ecology 87, 2378–2388. doi: 10.1890/0012-9658(2006)87[2378:diocra]2.0.co;2
Kimes, N. E., Van Nostrand, J. D., Weil, E., Zhou, J., and Morris, P. J. (2010). Microbial functional structure of Montastraea faveolata, an important Caribbean reef-building coral, differs between healthy and yellow-band diseased colonies. Environ. Microbiol. 12, 541–556. doi: 10.1111/j.1462-2920.2009.02113.x
LaJeunesse, T. C. (2001). Investigating the biodiversity, ecology, and phylogeny of endosymbiotic dinoflagellates in the genus Symbiodinium using the ITS region: in search of a “species” level marker. J. Phycol. 37, 866–880. doi: 10.1046/j.1529-8817.2001.01031.x
LaJeunesse, T. C., Lee, S. Y., Gil-Agudelo, D. L., Knowlton, N., and Jeong, H. J. (2015). Symbiodinium necroappetens sp. nov.(Dinophyceae): an opportunist ‘zooxanthella’ found in bleached and diseased tissues of Caribbean reef corals. Eur. J. Phycol. 50, 223–238. doi: 10.1080/09670262.2015.1025857
LaJeunesse, T. C., Parkinson, J. E., Gabrielson, P. W., Jeong, H. J., Reimer, J. D., Voolstra, C. R., et al. (2018). Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 28, 2570–2580. doi: 10.1016/j.cub.2018.07.008
Lawson, C. A., Raina, J., Kahlke, T., Seymour, J. R., and Suggett, D. J. (2018). Defining the core microbiome of the symbiotic dinoflagellate, Symbiodinium. Environ. Microbiol. Rep. 10, 7–11. doi: 10.1111/1758-2229.12599
Louca, S., Polz, M. F., Mazel, F., Albright, M. B., Huber, J. A., O’Connor, M. I., et al. (2018). Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2, 936–943. doi: 10.1038/s41559-018-0519-1
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Maire, J., Girvan, S. K., Barkla, S. E., Perez-Gonzalez, A., Suggett, D. J., Blackall, L. L., et al. (2021). Intracellular bacteria are common and taxonomically diverse in cultured and in hospite algal endosymbionts of coral reefs. ISME J. 15, 2028–2042. doi: 10.1038/s41396-021-00902-4
Matthews, J. L., Raina, J., Kahlke, T., Seymour, J. R., van Oppen, M. J. H., and Suggett, D. J. (2020). Symbiodiniaceae-bacteria interactions: rethinking metabolite exchange in reef-building corals as multi-partner metabolic networks. Environ. Microbiol. 22, 1675–1687. doi: 10.1111/1462-2920.14918
Mayoral-Peña, Z., Álvarez-Martínez, R., Fornoni, J., and Garrido, E. (2020). “The Extended Microbiota: How Microbes Shape Plant-Insect Interactions,” in Evolutionary Ecology of Plant-Herbivore Interaction, eds J. Núñ;ez-Farfán and P. Valverde (Berlin: Springer), 135–146. doi: 10.1007/978-3-030-46012-9_7
McDevitt-Irwin, J. M., Baum, J. K., Garren, M., and Vega Thurber, R. L. (2017). Responses of Coral-Associated Bacterial Communities to Local and Global Stressors. Front. Mar. Sci. 4:262. doi: 10.3389/fmars.2017.00262
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Mieog, J. C., Olsen, J. L., Berkelmans, R., Bleuler-Martinez, S. A., Willis, B. L., and van Oppen, M. J. H. (2009). The roles and interactions of symbiont. host and environment in defining coral fitness. PLoS One 4:e6364. doi: 10.1371/journal.pone.0006364
Motone, K., Takagi, T., Aburaya, S., Miura, N., Aoki, W., and Ueda, M. (2020). A zeaxanthin-producing bacterium isolated from the algal phycosphere protects coral endosymbionts from environmental stress. mBio 11:e1019-19. doi: 10.1128/mBio.01019-19
Muscatine, L. (1990). “The role of symbiotic algae in carbon and energy flux in reef corals,” in Ecosystems of the World: Corals Reef, Vol. 25, Z. Dubinsky, (Amsterdam: Elsevier Science Publishing Company), 75–87.
Ng, J. C., Chan, Y., Tun, H. M., Leung, F. C., Shin, P. K., and Chiu, J. M. (2015). Pyrosequencing of the bacteria associated with platygyra carnosus corals with skeletal growth anomalies reveals differences in bacterial community composition in apparently healthy and diseased tissues. Front. Microbiol. 6:1142. doi: 10.3389/fmicb.2015.01142
Oliver, T. A., and Palumbi, S. R. (2011). Do fluctuating temperature environments elevate coral thermal tolerance? Coral Reefs 30, 429–440. doi: 10.1007/s00338-011-0721-y
Peixoto, R. S., Rosado, P. M., Leite, D. C., de, A., Rosado, A. S., and Bourne, D. G. (2017). Beneficial Microorganisms for Corals (BMC): Proposed Mechanisms for Coral Health and Resilience. Front. Microbiol. 8:341. doi: 10.3389/fmicb.2017.00341
Pootakham, W., Mhuantong, W., Yucca, T., Putchim, L., Jomchai, N., Sonthirod, C., et al. (2019). Heat-induced shift in coral microbiome reveals several members of the Rhodobacteraceae family as indicator species for thermal stress in Porites lutea. Microbiol. Open 8:e935. doi: 10.1002/mbo3.935
Putnam, H. M., Barott, K. L., Ainsworth, T. D., and Gates, R. D. (2017). The vulnerability and resilience of reef-building corals. Curr. Biol. 27, R528–R540. doi: 10.1016/j.cub.2017.04.047
Raina, J.-B., Tapiolas, D., Willis, B. L., and Bourne, D. G. (2009). Coral-Associated bacteria and their role in the biogeochemical cycling of sulfur. Appl. Environ. Microbiol. 75, 3492–3501. doi: 10.1128/AEM.02567-08
Reshef, L., Koren, O., Loya, Y., Zilber-Rosenberg, I., and Rosenberg, E. (2006). The coral probiotic hypothesis. Environ. Microbiol. 8, 2068–2073. doi: 10.1111/j.1462-2920.2006.01148.x
Ritchie, K. B. (2006). Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar. Ecol. Prog. Ser. 322, 1–14. doi: 10.3354/meps322001
Ritchie, K. B., and Smith, G. W. (2004). “Microbial Communities of Coral Surface Mucopolysaccharide Layers,” in Coral Health and Disease, eds E. Rosenberg and Y. Loya (Berlin: Springer), 259–264. doi: 10.1007/978-3-662-06414-6_13
Rodriguez, R. J., Henson, J., Van Volkenburgh, E., Hoy, M., Wright, L., Beckwith, F., et al. (2008). Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2, 404–416. doi: 10.1038/ismej.2007.106
Rosales, S. M., Clark, A. S., Huebner, L. K., Ruzicka, R. R., and Muller, E. M. (2020). Rhodobacterales and Rhizobiales are associated with stony coral tissue loss disease and its suspected sources of transmission. Front. Microbiol. 11:681. doi: 10.3389/fmicb.2020.00681
Roth, M. S. (2014). The engine of the reef: photobiology of the coral-algal symbiosis. Front. Microbiol. 5:422. doi: 10.3389/fmicb.2014.00422
Rowan, R., Knowlton, N., Baker, A., and Jara, J. (1997). Landscape ecology of algal symbionts creates variation in episodes of coral bleaching. Nature 388, 265–269. doi: 10.1038/40843
Schoenberg, D. A., and Trench, R. K. (1980). Genetic variation in Symbiodinium (=Gymnodinium) microadriaticum Freudenthal, and specificity in its symbiosis with marine invertebrates. I. Isoenzyme and soluble protein patterns of axenic cultures of Symbiodinium microadriaticum. Proc. R. Soc. Lond. B Biol. Sci. 207, 405–427. doi: 10.1098/rspb.1980.0031
Seymour, J. R., Amin, S. A., Raina, J.-B., and Stocker, R. (2017). Zooming in on the phycosphere: the ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2:17065. doi: 10.1038/nmicrobiol.2017.65
Sharma, A. R., Zhou, T., Harunari, E., Oku, N., Trianto, A., and Igarashi, Y. (2019). Labrenzbactin from a Coral-Associated Bacterium Labrenzia Sp. J. Antibiot. 72, 634–639. doi: 10.1038/s41429-019-0192-x
Silva, S. G., Lago-Lestón, A., Costa, R., and Keller-Costa, T. (2018). Draft genome sequence of Sphingorhabdus sp. Strain EL138, a metabolically versatile alphaproteobacterium isolated from the gorgonian coral eunicella labiata. Genome Announc. 6:e00142-18. doi: 10.1128/genomeA.00142-18
Sitaraman, R. (2015). Pseudomonas spp. as models for plant-microbe interactions. Front. Plant Sci. 6:787. doi: 10.3389/fpls.2015.00787
Song, W., Wemheuer, B., Steinberg, P. D., Marzinelli, E. M., and Thomas, T. (2021). Contribution of horizontal gene transfer to the functionality of microbial biofilm on a macroalgae. ISME J. 15, 807–817. doi: 10.1038/s41396-020-00815-8
Stephens, M., Carbonetto, P., Dai, C., Gerard, D., Lu, M., Sun, L., et al. (2022). ashr: Methods for adaptive shrinkage, using Empirical Bayes. URL: http://CRAN.R-project.org/package=ashr
Stilianos, L., Polz, M. F., Florent, M., Albright Michaeline, B. N., Huber, J. A., O’Connor, M. I., et al. (2018). Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2, 936–943. doi: 10.1038/s41559-018-0519-1
Sweet, M. J., and Bulling, M. T. (2017). On the importance of the microbiome and pathobiome in coral health and disease. Front. Mar. Sci. 4:9. doi: 10.3389/fmars.2017.00009
Taguchi, S., and Kinzie, R. A. (2001). Growth of zooxanthellae in culture with two nitrogen sources. Mar. Biol. 138, 149–155. doi: 10.1007/s002270000435
Tandon, K., Lu, C. Y., Chiang, P. W., Wada, N., Yang, S. H., Chan, Y. F., et al. (2020). Comparative genomics: Dominant coral-bacterium Endozoicomonas acroporae metabolizes dimethylsulfoniopropionate (DMSP). ISME J. 14, 1290–1303. doi: 10.1038/s41396-020-0610-x
Thurber, R. V., Willner-Hall, D., Rodriguez-Mueller, B., Desnues, C., Edwards, R. A., Angly, F., et al. (2009). Metagenomic analysis of stressed coral holobionts. Environ. Microbiol. 11, 2148–2163. doi: 10.1111/j.1462-2920.2009.01935.x
Tian, L., Wang, X. W., Wu, A. K., Fan, Y., Friedman, J., Dahlin, A., et al. (2020). Deciphering functional redundancy in the human microbiome. Nat. Commun. 11:6217. doi: 10.1038/s41467-020-19940-1
Tout, J., Siboni, N., Messer, L. F., Garren, M., Stocker, R., Webster, N. S., et al. (2015). Increased seawater temperature increases the abundance and alters the structure of natural Vibrio populations associated with the coral Pocillopora damicornis. Front. Microbiol. 6:432. doi: 10.3389/fmicb.2015.00432
Trench, R. K., and Blank, R. J. (1987). Symbiodinium microadriaticum Freudenthal, S. goreauii sp. nov., S. kawagutii sp. nov. and S. pilosum sp. nov.: Gymnodinioid dinoflagellate symbionts of marine invertebrates. J. Phycol. 23, 469–481. doi: 10.1111/j.1529-8817.1987.tb02534.x
van Oppen, M. J., and Blackall, L. L. (2019). Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 17, 557–567. doi: 10.1038/s41579-019-0223-4
Vargas, S., Leiva, L., and Wörheide, G. (2020). Short-Term Exposure to High-temperature Water Causes a Shift in the Microbiome of the Common Aquarium Sponge Lendenfeldia chondrodes. Microb. Ecol. 81, 213–222. doi: 10.1007/s00248-020-01556-z
Voolstra, C. R., and Ziegler, M. (2020). Adapting with microbial help: microbiome flexibility facilitates rapid responses to environmental change. Bioessays 42:e2000004. doi: 10.1002/bies.202000004
Warner, M. E., and Suggett, D. J. (2016). “The photobiology of Symbiodinium spp.: Linking physiological diversity to the implications of stress and resilience,” in The Cnidaria, Past, Present and Future: The World of Medusa and her Sisters, eds S. Goffredo and Z. Dubinsky (Dordrecht: Springer Inc), 489–509. doi: 10.1007/978-3-319-31305-4_30
Webster, N. S., Negri, A. P., Botté, E. S., Laffy, P. W., Flores, F., Noonan, S., et al. (2016). Host-associated coral reef microbes respond to the cumulative pressures of ocean warming and ocean acidification. Sci. Rep. 6:19324. doi: 10.1038/srep19324
Wegley, L., Edwards, R., Rodriguez-Brito, B., Liu, H., and Rohwer, F. (2007). Metagenomic analysis of the microbial community associated with the coral Porites astreoides. Environ. Microbiol. 9, 2707–2719. doi: 10.1111/j.1462-2920.2007.01383.x
Willis, A. D. (2019). Rarefaction, alpha diversity, and statistics. Front Microbiol. 10:2407. doi: 10.3389/fmicb.2019.02407
Ye, S., Badhiwala, K. N., Robinson, J. T., Cho, W. H., and Siemann, E. (2019). Thermal plasticity of a freshwater cnidarian holobiont: detection of trans-generational effects in asexually reproducing hosts and symbionts. ISME J. 13, 2058–2067. doi: 10.1038/s41396-019-0413-0
Yuste, J. C., Fernandez-Gonzalez, A. J., Fernandez-Lopez, M., Ogaya, R., Penuelas, J., Sardans, J., et al. (2014). Strong functional stability of soil microbial communities under semiarid Mediterranean conditions and subjected to long-term shifts in baseline precipitation. Soil Biol. Biochem. 69, 223–233. doi: 10.1016/j.soilbio.2013.10.045
Zaneveld, J. R., McMinds, R., and Thurber, R. V. (2017). Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2:17121. doi: 10.1038/nmicrobiol.2017.121
Zhang, L., Zhang, W., Li, Q., Cui, R., Wang, Z., Wang, Y., et al. (2020). Deciphering the Root Endosphere Microbiome of the Desert Plant Alhagi sparsifolia for Drought Resistance-Promoting Bacteria. Appl. Environ. Microbiol. 86:e02863-19. doi: 10.1128/AEM.02863-19
Zhang, Y., Ling, J., Yang, Q., Wen, C., Yan, Q., Sun, H., et al. (2015). The functional gene composition and metabolic potential of coral-associated microbial communities. Sci. Rep. 5:16191. doi: 10.1038/srep16191
Keywords: microbiome, holobiont, thermotolerance, microbial communities, photosymbionts
Citation: Díaz-Almeyda EM, Ryba T, Ohdera AH, Collins SM, Shafer N, Link C, Prado-Zapata M, Ruhnke C, Moore M, González Angel AM, Pollock FJ and Medina M (2022) Thermal Stress Has Minimal Effects on Bacterial Communities of Thermotolerant Symbiodinium Cultures. Front. Ecol. Evol. 10:764086. doi: 10.3389/fevo.2022.764086
Received: 25 August 2021; Accepted: 13 April 2022;
Published: 19 May 2022.
Edited by:
John Everett Parkinson, University of South Florida, United StatesReviewed by:
Samantha R. Coy, Rice University, United StatesJustin Maire, The University of Melbourne, Australia
Copyright © 2022 Díaz-Almeyda, Ryba, Ohdera, Collins, Shafer, Link, Prado-Zapata, Ruhnke, Moore, González Angel, Pollock and Medina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erika M. Díaz-Almeyda, ediazalmeyda@ncf.edu