 Andrey D. Poyarkov
Andrey D. Poyarkov Miroslav P. Korablev
Miroslav P. Korablev Eugenia Bragina2†
Eugenia Bragina2†- 1Laboratory of Behavior and Behavioral Ecology of Mammals, A. N. Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Moscow, Russia
- 2Wildlife Conservation Society, Bronx Zoo, New York, NY, United States
This paper provides an overview of wolf research in Russia at the beginning of the 21st century. Wolf research covered various directions, including population density estimation, management methods and minimization of human-wildlife conflicts, general and behavioral ecology, behavior, wolf population genetics and morphology, paleontology, dog domestication, helminthology and the wolves’ role in the rabies transmission. Some studies are performed with state-of-art methodology using molecular genetics, mathematical modeling, camera traps, and GPS telemetry.
Introduction
Wolf research in Russia has a long history. One of the first wolf papers (Sabaneev, 1877) summarizes Leonid P. Sabaneev’s wolf hunting experience to increase hunting efficiency. Sabaneev also provides relevant facts on wolves’ biology. After Sabaneev’s book, the main focus of wolf research was wolf hunting (Zvorykin, 1936; Manteuffel, 1949; Kozlov, 1966; Pavlov, 1982). At the same time, academic studies describing several aspects of wolf biology were published (Dinnik, 1914; Satunin, 1915; Naumov, 1967). In 1973, the Wolf Working Group (herein and after WWG) was created and chaired by Professor Dmitry I. Bibikov. WWG made a considerable contribution to wolf research. First, Wolf Working Group members studied consequences of disrupted population structure, in particular emergence of feral dog and wolf-dog hybrid populations (Ryabov, 1978, 1979). Second, wolf ontogenesis received thorough attention (Badridze, 1987, 2003) as well as population abundance in various regions (Bondarev, 2012a; Yudin, 2013). One of WWG’s accomplishments was changing public opinion on wolves, in particular a ban on poisons, and a ban on wolves killing within protected areas. The WWG held debates about best wolf monitoring practices as a basis for management decisions (Bibikov et al., 1990). The results of WWG activity are summarized in the monograph The Wolf. History, Systematics, Morphology, Ecology (Bibikov, 1985).
Reviewing all the wolf research over the past two decades is beyond the scope of our work. Rather, we describe the main directions and overall trends. We emphasize that in the Soviet, and later, Russian society, the attitude toward wolves always has been very emotional. The overwhelming majority of hunters, game managers, and rural residents have a sharply negative attitude toward wolves. We analyzed the 2019 and 2020 editions of three hunting magazines popular in Russia: Hunting and hunting ground, Hunting and Fishing, and Okhotnik. In 2019, out of 17 articles on the wolf, 13 were strongly negative, 1 was positive, and 3 offered a science-based assessment approach. In 2020, 10 publications were negative, one was positive and 4 proposed a scientific-based approach.
Assessing the role of the species in ecosystems and its economic impact is a very difficult task that requires an integrated and objective approach and long-term consistent research. Unfortunately, this task is yet to be accomplished. However, the wolf in the USSR and Russia has always had a special attitude. Often, an oversight or simply theft of livestock was covered by the alleged wolf’s predation. Unfortunately, recent evidence suggest that wolves remain “scape goats” (Zheleznov-Chukotsky, 2016).
Wolf Distribution, Number, and Population Density
The most recent (2011–2015) wolf population estimate in Russia is 39.98 ± 1.20 thousand animals, and this large number is accounted to both wolf ecological plasticity and the lack of hunting pressure as a result of expensive hunting equipment (Kolesnikov et al., 2016). Below we, first, discuss historical changes of wolf population, second, report methods of abundance estimation, and then wolf dynamics in several Russian regions namely Komi, Dagestan, and Sakha Republics (northwest, southwest, and northeast of Russia, respectively) as well as local studies from Verkhne-Kondinsky (Khanty-Mansi Autonomous Okrug) and Lazo (Primorsky Krai) Nature Reserves.
Overall, wolf population number in Russia is tightly linked to Russian history. After World War I and Russian Civil War (1914–1923), wolf population increased dramatically. Then, it was brought down by intense population control. However, during World War II (1939–1945) wolf control was stopped and the population increased again. Afterward, bounties were paid to kill wolves, and the population declined (Bibikov, 1985). In 1990s, after the Soviet Union collapsed, population control stopped and the number of wolves skyrocketed (Bragina et al., 2015).
Wolf surveys are conducted in Russia with several methods: winter track counts (Bragina et al., 2015), home range mapping (Stepanova and Okhlopkov, 2020), and camera traps surveys (Zheltukhin and Ogurtsov, 2018; Volkov, 2020). For winter track counts, established transects are followed every year, and number of tracks crossing a transect is counted; second part includes following each species daily routes to estimate daily travel distance. Two numbers provide density estimation with Formozov’s formula: number of track crossings and daily travel distance (Bragina et al., 2015). Second method requires mapping a territory of each pack and then estimating the number of animals in a pack. Latter method is labor-intensive but more precise; in fact, difference between two methods’ assessments can be disagree by 2–3 times with winter track count providing inaccurate numbers (Stepanova and Okhlopkov, 2020; Volkov, 2020). Camera trap surveys provide abundance indices rather than population numbers, for example a number of animals per camera trap-days, and are expensive as they require camera traps arrays placed representatively in all habitat types. The most widely used index for camera-trapping data is the number of focal species captures per trap day (O’Brien, 2011), also often referred to as relative abundance index (RAI). As “capture” here is each series of photos or videos of the focal species made during a given time interval. Usually, camera traps are used within protected areas. For example, a wolf pack of 10–13 individuals was observed with camera traps in Kerzhensky Nature Reserve (Nizhny Novgorod Oblast; Volkov, 2020); similar wolf abundance was reported for Central Forest Nature Reserve (Tver Oblast, Central Russia; Zheltukhin and Ogurtsov, 2018).
The northeast of European Russia (Komi Republic) was inhabited by 2 wolf subspecies, arctic and taiga wolves, in 1930–1950s, which concentrated, respectively, in the northern and southern parts of Komi Republic. Wolf abundance in open habitats was significantly higher than in forested ones, and habitat type was the only abundance driver while hunter number and moose Alces alces density did not matter. In 1980s though two large clusters were formed: one included wolves in the northern and eastern parts while one—wolves in the central, western, and southern parts of Komi Republic. The most important driver of wolf abundance was moose and reindeer population density. Wolf penetration deep into the territory and its wide dispersal are associated with the landscape transformation by humans. Forest fragmentation, the growth of mosaicism, the emergence of large areas with young forests, and the road network development contributed to the wolf range expansion in Komi Republic (Korolev, 2016).
At the Northern Caucasus, the number and distribution of carnivores, including wolves, has been impacted by land use change: forest disturbance, agriculture development, melioration, and establishment of protective forest belts (Sukhomesova, 2013). For example, in Dagestan Republic (eastern Caucasus), wolf population is estimated at 2,750 individuals (Yarovenko, 2015). Hunters and fishermen are involved in the wolf population regulation within assigned hunting grounds. Despite the population control, there has been a slight tendency toward a number increase within those hunting grounds in 2013–2016, while throughout the whole Dagestan Republic the increase of wolf population has been even more significant (Yarovenko, 2015).
Survey of the huge Sakha Republic area, > 3,000,000 km2, resulted into 0.01 wolves/1,000 ha (Stepanova and Okhlopkov, 2020). This density is 5 times lower than the desirable goal set by the Russian Ministry of Natural Resources. Nevertheless, the authors make a point that total number of wolves in Russia in 2019 is at least twice as large as it was 40 years ago, 55,000 vs. 25,000, while the annual harvest of wolves is two times lower, 23% vs. 55%. Wolf population has been increasing in Sakha Republic, though the highest number was observed in 2011–2012, then declined in 2013, increased until 2019, and declined again in 2020. An additional driver of the wolf population increase is so-called synanthropic wolves i.e., wolves feeding on livestock. Labutin (1950 cit. by Stepanova and Okhlopkov, 2020) identified three “types” of Yakutian wolves based on their feeding behavior: tundra wolves predominantly feeding on wild and domestic deer, central taiga wolves feeding on mountain hare Lepus timidus, and southern taiga wolves preferring moose and red deer Cerphus elaphus (Stepanova and Okhlopkov, 2020). Recently, the white hare population in Sakha Republic has decreased leading to increase of wolf attacks on domestic reindeer and free-grazing horses (Safronov, 2016). The authors conclude that wolf population number exceeds desirable by 2–3 times. A mass-media campaign was launched to fight the “wolf threat” in the Republic, resulting in more funding allocated to regulate wolf population.
Long-term dynamics of the game species density in the Verkhne-Kondinsky Nature Reserve (Khanty-Mansi Autonomous Okrug) provides data on wolf population density dynamics in 1970–2010 (Vorobiev, 2015). The average density was 0.05 individuals per 1,000 ha. Wolf population was surveyed in 1975, 1994, 2009. The maximum populations number was 30 individuals, the long-term average was 12 individuals. The wolf population growth was not always associated with its prey increase. For example, average wolf density was higher in 1990s than in 1980s, 0.07 vs. 0.02 individuals/1,000 ha while moose breeding in adjacent territories decreased significantly at the same time, probably forcing wolves to move toward high abundance of ungulates.
Interaction with a larger carnivore results into wolves being displaced by Amur tigers Panthera tigris (Miquelle et al., 2005), according to habitat suitability modeling for both species (Voloshina et al., 2014). Study period spans 1960–1989 and includes 566 wolf and 2,543 tiger locations within and around Lazo Nature Reserve, Russian Far East. In 1960s and 1970s, out of 19 WolrdClim variables (Hijmans et al., 2005) main environmental variables impacting wolf presence was annual precipitation and precipitation of wettest quarter; in 1980s, when wolf population declined, it was precipitation of coldest quarter. Amur tiger presence was mostly impacted by temperature metrics: maximum temperature of warmest month in 1960s and annual mean temperature in 1970s. Only in 1980s, annual precipitation started playing the most important role for tigers. Overall, habitat quality has been improving for tigers and declining for wolves across the study period.
Wolf Management
In spite of fundamental ecological and societal importance of wolf population dynamic and management, there is currently no strategy nor action plan for population management other than encouragement for wolf killing and bounties for wolf pelts. At the same time, killing wolves can exacerbate the situation instead of improving it, while other methods of population control are more effective (Bondarev and Kotlov, 2006, 2007, 2008; Suvorov and Kirienko, 2008; Bondarev, 2012a, 2013). Both A.Y. Bondarev and A.P. Suvorov, active members of WWG in the 1980s, embraced the ideas of population self-regulation within the system “ungulates—predators. Using buffer zone hypotesis (Mech, 1977, 1979)”. Below, we discuss disadvantages of opportunistic killing and advantages of other methods.
First, we believe that an approach to wolf population control should be region- and habitat-specific. Intensive control is required in areas where wolves have a significant impact on livestock and game mammals e.g., in forest-steppe, regions of wild ungulates active exploitation, and remote livestock husbandry (Bondarev, 2012a). In other words, it is important to focus efforts on control of steppe synanthropic wolves. At the same time, mountain taiga wolves should not be controlled since they feed mainly on wild ungulates in slightly disturbed habitats (Suvorov and Kirienko, 2008). Also, the polar wolf of the Yenisei Territory (Central Siberia) should not be managed with aircrafts and snowmobiles, since this led to the total population extirpation in the past (Suvorov, 2016a). Similar concerns about complete extirpation of wolves with high-speed vehicles are raised in European Russia e.g., Belgorod Oblast (Chervonny and Gorbacheva, 2014).
In our opinion, the most effective strategy to control wolves is removing cubs from the dens because it reduces population number while maintaining the population structure at the same time. This way, a wolf population ages, and older wolves produce fewer cubs. In the south-western Altai Kray, this strategy successfully and cheaply prevents wolf immigration from numerous Kazakhstan population, excludes feral dogs from the wild, and, mostly important, significantly reduces wolf damage on livestock and wild ungulates maintaining the highest population density of roe deer and moose in the region (Bondarev, 2012a). If removing cubs is not possible, for example because of protected area regime or proximity to a state border, this procedure should be applied to adjacent areas while preserving breeding pairs. When a mating pair is killed, the boundaries of their home range and buffer zones are erased, the intrapopulation structure and spatial predator-prey relationships are disrupted. The destruction of wolf packs and consequent population size decrease facilitates wolf-dog hybridizations leading to a quick population recover. Therefore, such control does more harm than good for wild ungulates (Suvorov and Kirienko, 2008). Stable breeding pairs will prevent wolf immigration and reduce the damage from wolves, since the pairs need several times less food than large packs. This approach, in spite of being labor-intensive, keeps population spatial structure due to preserving adult breeding pairs as well as denning and territorial fidelity inherent to wolves (Bondarev and Kotlov, 2007).
Another popular approach for wolf population control is professional brigades of wolf hunters (Bondarev, 2013; Suvorov, 2016b). This idea was also discussed at the WWG meetings in the 1980s and was recognized as useful and cost-efficient as compared to the aviation use (Suvorov, 2016b).
In spite of proven and efficient management tools described above, there is a big hunting lobby promoting other approaches: increasing the hunting season length, bounties for wolf pelts, and use of currently prohibited foot traps and snares (Budlyansky and Sinilov, 2019). Advocates of this approach consider wolves a “pest species,” and do not take into account wolves’ population structure, social organization, and self-regulation mechanisms. According to this point of view, wolves should be hunted all year around instead of the current season of 5.5 months/year, and bounties of 10,000–20,000 rubles ($100–200) should be paid for each killed wolf from the federal budget. Currently, the main obstacle preventing evidence-based scientific approach to wolf management is a requirement #138 signed on April 30, 2010 directing to keep maximum wolf density in hunting grounds below 0.05 individuals/1,000 ha (Budlyansky and Sinilov, 2019)/While this document indicates the upper limit of wolf density, nothing is said about the lower one, leaving a loophole for hunting outside of the hunting period.
One of the hunting lobby’s arguments is assumption that wolves have high impact on ungulates. For example, Bersenev et al. (2012) provide calculations of wolves’ predation pressure: “Every year wolves kill about 34 thousand moose, 123 thousand roe deer, 20 thousand red deer, and 140 thousand reindeer. Lost profit for the hunting economy as a result of the annual feeding of one wolf is 0.6 moose + 2.5 reindeer (or another species replacing it) + 0.37 red deer + 1.85 roe deer + 0.7 wild boar + 49.7 hares + farm animals weighing 77.6 kg.” They conclude that “at present, the necessity to control for wolf number in the Russian Federation is obvious. With a 50% reduction of the wolf population, the positive economic effect from the ungulate number increase alone will be at least 4,000,000,000 rubles ($40,000,000) annually.” This calculation does not account for (1) compensatory as opposed additive impact of carnivore on ungulates and (2) species other than ungulates in wolf diet, especially important during summer time (Kolpashchikov, 2016; Suvorov, 2016a).
We should mention that in spite of the strong hunting lobby, some ecologists do recognize importance of wolves for ecosystems and promote wolf conservation (Suvorov, 2016a,b). For example, there are no more than 30 wolves in Samara region (53,600 km2), and it is not clear how many of those belong to local population as opposed to migrants and hybrids with dogs. For such a large territory, this is an extremely small number. In the neighboring Orenburg region, there are 200 wolves, and it was suggested that wolves should be included in the local Red List (Rigina and Vinogradov, 2007a,b).
Wolf-Ungulates Relationship
Below we describe 3 studies of wolf-ungulate relationships. Neither study found that wolves limit ungulate density. First study was conducted in the ecosystem with multiple ungulate species while second one in a system where moose is wolves’ exclusive prey, and the 3rd study was theoretical.
In Belgorod region, wolves prey on moose, red deer, wild boar, and roe deer (Chervonny and Gorbacheva, 2014). In 1964–2011, wolf number declined from 103 to 12 individuals, and then to 0 in 2013. The highest population density of > 100 animals was observed in 1964 and 1973, and also more than 70 wolves were observed in 1990s. For the rest of the study period, there were 25–30 individuals in 2–3 packs, and less than half suitable wolf habitats were occupied. Ratio of ungulate/wolf number varied from 20 to 285, with 20 ungulates per 1 wolf observed only at the beginning of the study period when it resulted in a sharp decline of wolves. For most of the study period, number of wolves was too low to impact ungulates. Local wolf extinction was caused by excessive hunting and increase of the snowmobiles use for hunting (Chervonny and Gorbacheva, 2014).
In Karelia Republic, at the Russian northwest, wolves predominantly hunt moose. In 1961–2020, wolf/moose ratio always was higher than 1–23, usually much higher (Danilov et al., 2020). Wolves did not consume more than 6–7% of the winter moose population and mostly ate calves and females. In winter, wolves killed 35–80% of the moose population while in the summer it was only 10–17%. The nature and efficiency of wolf hunting moose barely depended on a pack size (Danilov, 2017). In 2018, the highest number of wolves was killed in Karelia (250), and the total number of this carnivore in the region has recently decreased to 300–350 individuals (Danilov et al., 2020). The high wolf harvest rate results from cash bounties for wolf pelts and reduced payment to hunt ungulates.
Agent-based modeling of wolf-moose system with AnyLogic software also provided evidence for moose driving the wolf population, not wolves limited moose (Elufereva and Limanova, 2020). In spite of erroneous assumption that wolves breed twice as opposed to once a year (Mech, 1970; Packard, 2003), the author concluded that wolf number followed their prey density. The authors fitted a large number of models varying the wolf hunting parameters (in a pack or alone), food availability (moose density), temperature, human hunting pressure on wolf and moose as well as presence of infection (Elufereva and Limanova, 2020). At a low moose number, the wolf disappeared from suboptimal habitats without being pursued by hunters. When hunters were introduced into the model with a high probability of wolf extermination, the extinction of species occured. After introducing an infection into the model that killed 50% of any species, populations recovered in 2–3 years (Elufereva and Limanova, 2020).
Wolf Diet and Prey Preferences
Wolf diet was analyzed in European Russia, Kaluzhskiye Zaseki Nature Reserve, where wolf prey include red deer, wild boar, moose, European bison, and roe deer, and Russian Far East, in Bolshekhetskirsky and Sikhote-Alin Nature Reserves where prey species include red deer, wild boar, roe deer. The latter reserves protect an interesting ecosystem where wolves interact with Amur tigers. Prey species composition was calculated as occurrence rate of various species in feces identified based on hair cuticula characteristics (Teerink, 1991; Rozhnov et al., 2011). This method does not account for prey species body mass.
In forest habitats of Kaluzhskiye Zaseki Nature Reserve, wild board and roe deer are the main prey species for wolves (38.7% and 30.6%, respectively, N = 87) in spite of moose, European bison, and red deer also being available within a pack’s home range. However, wolves prefer wild boar during the snowless period and roe deer in the snow period (Hernandez-Blanco and Litvinova, 2003).
In Bolshekhetskirsky Nature Reserve ungulates also provide the most of wolves’ diet—92.5%. However, the most common species is red deer making 66.3% of wolf diet in the snow period and less in snowless period. In comparison with protected lands, livestock carrions (42–55%) and small- and medium-sized mammals (33–42%) dominate wolf diet on agricultural fields surrounding Bolshekhetskirsky Nature Reserve (Tkachenko, 2010). Interestingly, this study spans a period of 1989–2005, and since the mid-1990s, wolves were forced out of the reserve by Amur tigers and barely reappeared in some winter seasons.
Influence of wolves and tigers on other carnivores was also studied in the Sikhote-Alin mountains (Salkina and Eremin, 2017) by analysis of carnivore occurrences (more than 4,000 data points). Wolves negatively impacted domestic dogs, common raccoon dogs, European badgers, European lynx, and Amur leopard cats but did not affect red foxes. Amur tiger population increase correlated with the wolf population decrease. The authors suggest that in the absence of wolves, their niche can be occupied by feral dogs. Therefore, wolf control during low population density periods is not desired (Salkina and Eremin, 2017).
Space Use
In this section, we describe spatial structure of a pack’s home range, habitat selection, individual movements, home range sizes and individual daily travel distances.
A wolf pack’s home range in forested habitats of European Russia (Voronezh Nature Reserve), calculated as a minimal convex polygon, is 146–167 km2 (Hernandez-Blanco et al., 2005). Average minimal convex polygon of individual wolf home ranges in steppe zone of Asian Russia (Daurian Nature Reserve) is much larger, 832 km2, exceeding wolf home rages in European and North American forested habitats by 2–4 folds, perhaps due to low cost of moving through the snowless landscape (Kirilyuk et al., 2019). Large distances of movements from a place of birth were recorded for 2 year old wolves of both sexes (Kirilyuk et al., 2020). Interestingly, daily activity of 17 GPS-collared wolves in the Daurian steppe (Kirilyuk et al., 2021) revealed that during invasions of neighboring packs and, also, long-distance departure from the habitat, wolves move over long distances and for longer time. In summer, activity is higher than in winter.
A concept of wolf pack home range structure proposed by Hernandez-Blanco et al. (2003a,b, 2005) identifies 3 spatial subunits: home core, vital space, and spatial shell. Home core includes den sites and rendezvous-sites. Vital space is used by adult animals to hunt with their offspring. Spatial shell is a peripheral part of the home site usually used by yearlings from the rut season until the mid-summer. In case there are so-called buffer zones between neighboring packs, they are shaped by the spatial shells from the neighboring home sites. This structure of the space use by wolves also impacts ungulates’ spatial distribution (Kazmin et al., 2001). Home ranges of neighboring packs are separated with so called buffer zones shaped by spatial shells. Continuation of the territory use through generations is provided through the replication effect when two females, an old and young one, breed at the same time within a pack’s home range.
Wolf movement analysis reveals that wolves have strong preferences for using roads and moving along ravines and rivers as opposed to crossing them (Melnik et al., 2007). At the same time, terrain characteristics of a route depend on a route’s type. Melnik et al. (2007) identify several route types: search-hunting, marking, search-social and linear. On search-hunting and search-social routes, wolves more often cross terrain isoclines, make turns, loops and temporarily separate from group members than on marking and linear routes. Among all habitat types, wolves choose those preferred by ungulates (Melnik et al., 2007).
Experimental Research on Wolf Behavior
Yachmennikova et al. (2009) described so-called “Transformational” period in ontogenesis of wolf pups: a dramatic increase followed by a decrease in the number of behavioral pattern types between 75th and 115th day. Ontogenesis of wolf behavior (Yachmennikova and Poyarkov, 2010, 2011) was studied with software Theme 5.0 from Noldus designed specially to reveal hidden temporal patterns (Magnusson, 2000). The activity type of each animal was registered each minute with the time slice method. Eighteen types of activity were recorded. The sequential stream of activity of four animals was analyzed to find time patterns that are repetitive events not randomly following each other within the critical time interval. Often, pattern types including various activities were observed (1,300–13,500 types). Although most patterns consist of 2–3 elements, complex sequences of 15–16 behavioral acts are also noted. Over the linear sequence of initial events, a complex non-linear and clearly ordered organization of the other type is revealed.
Ontogenetic development of wolf pups was studied to identify conditions critical for successful reintroduction. Badridze (2017) found that foot massage of mother’s mammary glands and opportunity to suck is dramatically important for small pups. If pups cannot exercise these behaviors, they become overstimulated, and development of their manipulative behavior is impeded leading to challenges after reintroduction. Reintroduction success also depends on food hiding behavior (Badridze, 2010b), hunting behavior (Badridze, 2010a), and avoiding of humans and livestock (Badridze et al., 1992). Successful reintroduction of 4 wolf groups led to establishing 4 wolf packs. In 3 years after the reintroduction, feral dogs disappeared from the area and roe deer travel distance increased. Wolf hunting success decreased during first 6 months after the reintroduction and then remain stable (Badridze, 2017).
Morphology of Wolves
Morphological approach, and craniometric measurements in particular, represent an effective tool for studying the polymorphism and differentiation of mammal populations (Palmeirim, 1998; Gauthier et al., 2003). In Russia, the morphological research traditions are strong; therefore, the morphological variability of the gray wolf has always been the subject of a wide range of studies. The results of these studies performed up to the half of the 1980s were summarized in the monograph “The Wolf. History, Systematics, Morphology, Ecology” by Bibikov (1985). To our knowledge, few studies on wolf morphometry, including craniometry, have been published over the past two decades.
Wolf craniometric variability in the Russian Caucasus (Republics of Kabardino-Balkaria, North Ossetia, Dagestan, and Adygea) and Transcaucasia (Azerbaijan and Georgia) was analyzed with 67 skulls (Tembotova and Kononenko, 2007a,b). The authors performed a comparative study of the sexual and geographic variation. In all regions of the study area, they found a high sexual dimorphism in size. The largest wolf skulls were found in Azerbaijan. Wolves from Russian Caucasus had intermediate skull sizes, while the smallest skulls were found in Georgia.
Studies on morphometric body parameters of wolves from a huge area of Western Siberia detected a significant geographic variation of the wolf sizes (Bondarev, 2012b). However, neither Allen’s (Allen, 1877) nor Bergman’s (Bergman, 1847) rules were confirmed. The largest wolves of Siberia inhabit areas of the northern and middle taiga followed by wolves of tundra, while the smallest dwell in the forest-steppe zone (Bondarev, 2012b). Despite the huge dataset, the author did not find taxonomic differences in wolves of the Western Siberia.
Large geographic variation of craniometric parameters and sexual dimorphism in skull size was found at the Russian Far East and Kamchatka based on 410 skulls (Yudin, 2013). Patterns of variability showed the complex population structure and no direct i.e., clinal variation. A possible reason for the significant variation of the population craniometric measurements is the geographic isolation and direct adaptive response of the populations to the environmental conditions (Yudin, 2013).
Morphometric study of 363 skulls and 242 carcasses of polar and forest wolves from Yenisei Siberia (Krasnoyarsk Region and the Republic of Khakassia) drew the same conclusion about the wolf population structure (Suvorov, 2017a,b). There also was no clinal variability; craniometric differences between polar and forest wolves were not found. The largest Russian wolves live in the northern and temperate forests while smallest live in the forest-steppe zone. Overall, wolf body and skull dimensions are shaped by environmental conditions, which, in turn, vary significantly across the study area (Suvorov, 2017a,b).
Wolf craniometric variability was studied in details in the center of European Russia (Korablev N. P. et al., 2021). With 326 skulls collected over 65 years from Tver, Smolensk, Yaroslavl, and Vologda regions, the authors analyzed various drivers of population craniometric polymorphism. A high craniometric variability and sexual dimorphism of skull sizes has been found. Polymorphism is mostly determined by temporal and spatial trends, sex and age, in order of the effect size. For example, female skulls are 3.6% smaller due to sexual selection as well as differences in male and female diets. Temporal polymorphism i.e., a weak increase of morphological parameters with time is likely driven by increase of moose and wild boar population abundance, leading to prevalence of both species in wolves’ diet. Stochastic drivers are also at play e.g., high total mortality and disruption of the population social structure as a result of hunting pressure.
Population Genetics
The rapid development of molecular genetic techniques gave rise to population genetic studies of wolves in Europe (Randi, 2011; De Groot et al., 2016; Hindrikson et al., 2017). However, in Russia, this area of species biology is still poorly understood. So far, there are only a few publications on the wolf population genetics in Russia, most of them with Siberian wolves.
Based on the study of 97 individual wolves with 6 microsatellite markers, the taxonomic status of the forest-steppe and mountain-taiga wolves of Altai was clarified: the populations inhabiting various biotopes belong to the same subspecies C. lupus altaicus (Vorobyevskaya and Baldina, 2011).
High genetic diversity was found among 163 individuals from Siberian regions including Altai Krai, Altay Republic, Tyva Republic, Republic Buryatia, Republic Khakassia, Krasnoyarsk Krai, and Zabaykalsky Krai (Bondarev et al., 2013). Expected heterozygosity He was 0.72–0.82 and a mean number of alleles Na was 9.83–12.67 with an average He and Na of 0.65 and 7.67, respectively. The mountain-taiga populations of Altai and Sayan mountains had the highest genetic diversity, as did steppe wolves at south-west of Altai Krai, perhaps due to prey abundance in these areas. Plain-taiga populations of Krasnoyarsk Krai, Evenkia and Salairskiy Kryazh in Altay Krai had the smallest genetic diversity. In general, described genetic structure coincided with landscape zones.
Talala et al. (2020) broadened a geographical span of 2 above-described papers (Vorobyevskaya and Baldina, 2011; Bondarev et al., 2013) with samples from Yakutia Republic thus analyzing 270 individuals and 7 microsatellite loci. Relatively high genetic diversity of Siberian wolves was confirmed: He 0.60–0.71 and Na 4.50–5.83 with an average He and Na of 0.68 and 5.18, respectively. Siberian wolf populations are connected by the active gene flow. However, geographical distribution of previously described subspecies (Canis l. sibiricus, C. l. altaicus, C. l. turuchanensis, Canis l. var. orientalis) was confirmed by the genetic data.
A detailed genetic study of wolves in European Russia genotyped 101 wolves and 32 dogs at 11 microsatellite loci from Tver and Pskov regions (Korablev M. P. et al., 2021). The study’s goal was assessment of the spatial and temporal dynamics of wolf population structure and genetic diversity over 30 years during population number increase, and assessment of wolf-dog hybridization rate. The authors found that the studied area is inhabited by a single wolf population confirming previously reported data (Pilot et al., 2006; Sastre et al., 2011). This population has high genetic diversity (He = 0.79 and Na = 10.00) which is more than most of European populations have (Korablev M. P. et al., 2021). The studied wolves represent a highly polymorphic part of the continuous Canis lupus lupus population with a relatively low rate of hybridization (around 3%) and, hence, can be viewed as a natural reservoir of the subspecies’ gene pool.
Wolf Helminthology
Wolf helminths have been studied in several Russian regions. Two methods are mainly used: (1) a complete helminthological examination of wolf carcasses with the Scriabin method (Skryabin, 1928) and (2) the identification of eggs and cysts with microscopes (Esaulova et al., 2015). Eighty seven species of wolf helminths found in the Holarctic: 29 species of cestodes 4, 37 nematodes, and 17 trematodes. Out of 87, 68 were found in Russia (Konyaev and Bondarev, 2011). Overall, helminthological studies highlight importance wolves as a reservoir for helminths and potential source of infection for domestic and synanthropic animals.
Several papers cover wolf helminths in Russian regions. For example, 15 species of wolf helminth was found in Volgograd region: 7 nematodes, 6 cestodes, and 2 trematodes. Toxascaris leoninae (64.7%), Dirofilaria immitis (47%) and Toxacara canis (35.2%) are the most common nematodes. Taenia pisiformis (41.1%) and Echinococcus granulosus (23.5%) are the most common cestodes (Shinkarenko and Kolesnikov, 2011).
In Dagestan, 18 species of wolf helminths were found including trematodes, cestodes, and nematodes. Notably, helminth species composition differed between mountain, foothill and lowland Dagestan wolves. Single-host helminths (nematodes) and two-host (cestodes) were the most common (Bittirov et al., 2010).
In the North-West Caucasus 17 species of wolf helminths were identified (Itin and Kravchenko, 2016). In Kabardino-Balkaria Republic, also part of the North Caucasus, 12 species of wolf cestodes and nematodes were found (Kabardiev and Bittirov, 2020) including, as well as in Dagestan (Bittirov et al., 2010), dangerous to humans Echinococcus granulosus and Dipylidium caninum with prevalence of 80.0 and 40.0%, respectively.
Only 6 species of wolf helminth species were found in Kaluzhskiye Zaseki Nature Reserve, and 3 species—in Kalmykia (Esaulova et al., 2018), perhaps because the search for helminths was done with wolf feces rather than with carcass surveys.
In Kirov region, 19 species of helminths were identified including 1 trematode, 7 cestodes, and 11 nematodes. These were helminths such as Alaria alata, Dipylidium caninum, Taenia hydatigena, Taenia krabbei, Taenia pisiformis, Tetratirotaenia polyacantha, Echinococcus granulosus, Echinococcus multilocularis, Crenosoma vulpis, Thominx aerophilus, Capillaria plica, Capillaria putoria, Trichinella nativa, Uncinaria stenocephala, Toxascaris leonina, Toxocara canis, Toxocara canis, Ancylostoma caninum, Strongiloides vulpis, Molineus patens. The most common species was the trematode Alaria alata (73%), the cestodes Taenia hydatigena and Echinococcus granulosus, and the nematodes Uncinaria stenocephala and Trichinella native (Maslennikova, 2012).
In Ivanovo region, wolves were infested with 12 species of helminths (Andreyanov et al., 2009; Kryuchkova et al., 2011; Abalikhin et al., 2013). One species of trematodes, Alaria alata, was found in the intestines of animals, with prevalence of 93.8% of infected animals and infection intensity (number of helminths in each infected individual) of 8–1,056 specimens.
There are several papers where researchers focused on one or more species of the same genus, as a rule, especially dangerous helminth diseases. For example, Andreyanov (2020) described reservoirs of alveococcus infection in game carnivore species at hunting grounds of Vladimir, Nizhny Novgorod, Moscow, Tver, Oryol, and Bryansk regions of Central Russia as well as the Karelia Republic in 2007–2018. These regions have high human population density as well as high number of hunters and pet owners. A complete or partial helminthological autopsy was conducted with Scriabin method (Skryabin, 1928) for 262 animals, including 193 common red foxes, 28 feral dogs, 16 raccoon dogs, 16 feral cats, 6 wolves, 2 brown bears, and 1 lynx. Cestodes of Echinococcus multilocularis was found in 46 foxes (23.8%), 3 raccoon dogs (18.7%), 3 wolves (50%), and 1 feral dog (3.6%). The peak tapeworm prevalence in carnivores was noted for foxes and raccoon dogs.
Trichinosis is another wolf’s helminthosis dangerous for humans. Across Russia, trichinosis prevalence is 18.5% (Tulov et al., 2013), for example 20% in Altay Krai and Altay Republic (Malkina and Konyaev, 2013). In all cases, wolves had T. nativa.
Rabies
In spite of rare occurrence of rabies in wolves as compared to red fox, wolves still carry this disease. For example, in 2013, 19 cases of rabies were registered in wolves in Russia (Novikova and Petrova, 2015). In Central Russia, in Lipetsk, Moscow, Tver’ and Yaroslavl’ regions, there was 5 cases of rabies in wolves, or 0.5% of all cases (N = 1,089 animals). In Tver’ region, the most often rabies hosts are raccoon dogs (39%), and perhaps wolves contract rabies while preying on raccoon dogs (Nesterchuk et al., 2019).
In Yakutia, rabies also has been reported in wolves, although polar foxes are the main rabies reservoir there (Zakharova, 2019).
Recently, rabies rates have increased in many regions of Russia. The most cases are observed in the south of Western Siberia and the central and southern parts of European Russia (Poleshchuk et al., 2012, 2019). Number of rabies cases have increased in Transbaikalia, Buryatia, and Krasnoyarsk Regions (Sidorova et al., 2007; Botvinkin et al., 2019). Meltsov et al. (2020) provide a detailed analysis of the rabies situation in Irkutsk region, the only Russian region free from rabies, including prevalence of rabies in red foxes and wolves. Rabies transmission can be curbed with peroral regular vaccination. Meltsov et al. (2020) identified the areas of the most likely rabies penetration into Irkutsk region, and initiated peroral vaccination of foxes and wolves in these areas up to 50 km depth from a point of possible penetration. This strategy has been highly efficient. However, Meltsov et al. (2020) warn about the risk of rabies transmission through pets.
In adjacent to Irkutsk region Buryatia Republic, two outbreaks of rabies have been reported in wild animals in 2011–2013 and 2017–2019. The most affected species was red fox. Only few cases of rabies have been reported among wolves. Both outbreaks of rabies in Buryatia took place during the years of red fox peak numbers and low numbers of wolves (Shchepin et al., 2019). Rabies was also registered in Transbaikalia (Botvinkin et al., 2019).
In summary, the rabies situation in Russia is becoming challenging. Recently, there has been a shift from natural (autochthonous) foci and carriers of the disease to synanthropic ones, with feral dogs and cats as a main source of infection. Large-scale vaccination of wild, feral and domestic animals is needed as an effective measure to curb rabies (Novikova and Petrova, 2015; Meltsov et al., 2020).
Wolf Paleontology
Paleontological studies is a special section of our review. There are few of them but usually it is high-quality research published in high-ranked journals. Usually, these studies are co-authored by big international teams including Russian specialists who found some paleontological remains on Russian territory. For example, Lee et al. (2015) analyzed mDNA of 14 canid remains aged 17,000–360,000 years before present. One of these samples was identified as a separate species named Canis cf. variabilis. All the samples had affinity with pre-domesticated as well as modern dogs leading Lee et al. (2015) to question an opinion about European origin of domestic dogs.
Ní Leathlobhair et al. (2018) analyzed ancient wolves to look into the origin of American dog. American dogs arrived from Siberia through Beringia about 9 thousand years ago. Genetic analysis reveals that American dogs are closer to Siberian wolves than to American ones. Sinding et al. (2020) came to similar conclusions regarding sled dogs. Present-day sled dogs are closely related to Siberian 9,500-year-old dogs and 33,000-year-old Pleistocene Siberian wolves, but not modern American wolves. The importance of the Beringia wolf expansion during the last glacial maximum is demonstrated by Loog et al. (2019): “Our results suggest that contemporary wolf populations trace their ancestry to an expansion from Beringia at the end of the Last Glacial Maximum and that this process was most likely driven by Late Pleistocene ecological fluctuations that occurred across the Northern Hemisphere. This study provides direct ancient genetic evidence that long-range migration has played an important role in the population history of a large carnivore, and provides insight into how wolves survived the wave of megafaunal extinctions at the end of the last glaciation. Moreover, because Late Pleistocene gray wolves were the likely source from which all modern dogs trace their origins, the demographic history described in this study has fundamental implications for understanding the geographical origin of the dog.”
Canid remains from the Yana paleolithic site (Republic of Yakutia) shed light on the early wolf domestication (Nikolskiy et al., 2018). Most remains belongs to “intermediate” between wolves and dogs canids with worn, partially missing teeth and various bone pathologies, often small in body size. As modern experiments show, such pathologies accompany tolerance to people. Perhaps, those canids used human settlements as a food source in spite of human-wildlife conflict risk due to their tolerance to humans, at the cost of accumulating various anomalies. Also, some evidence of wolf totemic cult was found at the Yana site. Thus, self-domestication of Yana wolves could be considered one of the first steps of wolf domestication (Nikolskiy et al., 2018).
Various wolves from Siberia aged 14–50 thousand years reveal two lines of wolves: Pleistocene wolves morphologically close to modern Siberian wolves, and shorter-faced “Paleolithic dogs” morphologically intermediate between wolves and dogs (Ramos-Madrigal et al., 2021). Both lineages do not form a monophyletic group. Pleistocene wolves are represented by several extinct lines that could be ancestral forms of Arctic and some Asian dog breeds (Ramos-Madrigal et al., 2021). Remains of Pleistocene wolves aged 40,000–50,000 years were found in 2 caves of Altay mountains, Razboynichya and Fanatikov. A new wolf species was identified based on its gracile skull shape and smaller brain size and named Canis subtilis (Ovodov and Martynovich, 2011).
Conclusion
Papers on Russian wolves have been published in sources of varying accessibility and varying scientific significance. A brief analysis of the sources we listed shows an extremely uneven distribution of works depending on a study subject. Table 1 shows the distribution of works depending on the subject (corresponding to our review) by journals, monographs and compilation. We divided publications into journals indexed in Web of Science by quartiles, Scopus (if the journal is not cited in Web of Science but is cited in Scopus), other journals, Monographs or chapters in monographs, compilation combining thematic publications and Ph.D. dissertations thesis (Table 1). The distribution of works showed a sharp skew in the direction of hard-to-reach and low ranked sources. Only 24 studies (22%) are published in the international Web of Science or Scopus databases, while the remaining 85 studies (78%) are difficult to access for international scientific community preventing the integration of Russian wolf specialists to the world scientific community. Only paleontological research stand out being published in high-ranked journals, primarily due to large highly professional scientific teams in which Russian specialists take an active part. Such research areas as regional distribution, abundance, population density, relationship with ungulates, trophic ecology and management generally do not reach international scientific databases making results unavailable for the international scientific community.
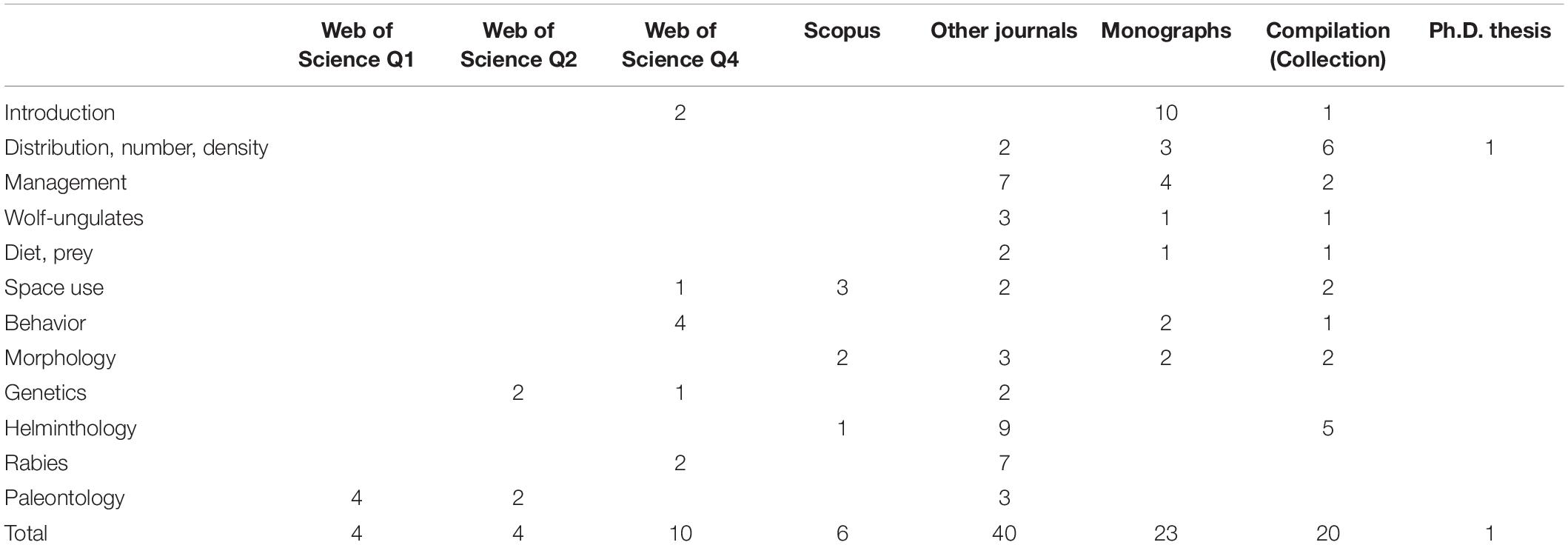
Table 1. Distribution of publications by significance level in databases.
Among all the countries overlapping with wolf, the Russian part of the wolf range is the largest one. Russian wolves live almost everywhere, with minor exceptions such as large cities and some Arctic and Pacific islands. Wolves occupy various habitats and geographical zones. Our knowledge about wolves has many gaps to fill; the lack of deep understanding is the main obstacle to the comprehensive assessment of wolf impacts on diverse ecosystems.
Author Contributions
AP, MK, and JH-B collected literature and wrote the text (partially). EB carried out the general revision of the work. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abalikhin, B. G., Kryuchkova, E. N., and Egorov, S. V. (2013). Same aspects of ecology and parasitofauna of the wolf in the conditions of the Ivanovo region. Rus. Parasit. J. 3, 41–44.
Allen, J. A. (1877). The influence of physical conditions in the genesis of species. Radic Rev. 1, 108–140.
Andreyanov, O. N. (2020). Examining Echinococcus multilocularis infection in some Midland Russia predatory animal species. Rus. J. Infect. Immun. 10, 193–196. doi: 10.15789/2220-7619-eem-1182
Andreyanov, O. N., Safiullin, R. T., Gorokhov, V. V., and Kryuchkova, E. N. (2009). Parasitofauna of canid predators in the Central Non-Chernozem region of Russia. Vet. Med. 6, 37–40.
Badridze, J. K. (2010a). Formation of hunting behavior of the wolf (Canis lupus). Zool. Zhurnal 89, 862–870.
Badridze, J. K. (2017). Wolf Problems Associated with the Reintroduction of Large Carnivorous Mammals. M. Modern. 136.
Badridze, J. K., Gurilelidze, Z., Todua, G., Badridze, N., and Butkhuzi, L. (1992). The Reintroduction of Captive- Raise Large Mammals into Their Natural Habitat: Problems and Method. Tbilisi: Institute of Zoology of the Academy of Science, 12.
Badridze, Y. K. (1987). Feeding Behavior of the Wolf (Questions of Ontogenesis). Tbilisi: Metsniereba, 86.
Badridze, Y. K. (2003). Questions of Behavior Ontogenesis, Problems and Methods of Reintroduction. Tbilisi: Tbilisi State University, 116.
Bergman, C. (1847). About the relationship between the heat economy of animals and their size. Göttingen Stud. 3, 595–708.
Bersenev, A. E., Kulpin, A. A., and Gubar, Y. P. (2012). “The wolf and the number of hunting species of ungulates. State of the habitat and fauna of hunting animals in Eurasia,” in Materials of the IV All-Russian Scientific-Practical Conference “The State of the Habitat and Fauna of Hunting Animals in Russia. (Moscow: Russian State Agrarian University), 31–40.
Bibikov, D. I. (ed.) (1985). The Wolf. History, Systematics, Morphology, Ecology. Moscow: Nauka Publishers.
Bibikov, D. I., Ovsyanikov, N. G., and Poyarkov, A. D. (1990). Meeting of the commission on large predators of the WTO of the academy of sciences of the USSR. Zool. J. 69, 158–160.
Bittirov, A. K., Kazancheva, L. K., Atalaev, M. M., and Sarbasheva, M. M. (2010). Wild Carnivorus bio and geohelminthoses in the north Caucasus region and dehelmintization method perfection. Vestn. Krasgau 2010, 112–117.
Bondarev, A. Y. (2012a). On the principles of regulation of the number of wolves. Altai State Agrar. Univ. Bull. 9, 70–71.
Bondarev, A. Y. (2012b). Morphological characteristics of the wolf (Canis lupus) of the western and central Siberia. Vestn. Altai Gos. Agrar. Univ. 87, 30–34.
Bondarev, A. Y., and Kotlov, A. A. (2006). “Management of wolf populations as an element of rational environmental management,” in Agrarian Science – Agriculture: Intern. Scientific-Practical Conference (Barnaul: AGAU Publ.), 362–365.
Bondarev, A. Y., and Kotlov, A. A. (2007). “From the extermination of the wolf to its populations management,” in Modern Problems of Nature Management, Hunting and Animal Husbandry: Intern. Scientific-Practical Conference. - Kirov, -“VNIOZ RASCHN”, 54–55.
Bondarev, A. Y., and Kotlov, A. A. (2008). On monitoring the reproduction of the gray wolf and regulation of its number and location. Izvestia. Orenburg. State Agrar. Univ. 4, 180–183.
Bondarev, A. Y., Vorobevskaya, E. A., and Politov, D. V. (2013). On the genetic differentiation of the Siberian wolf. Vestn. Altai Gos. Agrar. Univ. 107, 49–57. doi: 10.1111/eva.13257
Botvinkin, A. D., Zarva, I. D., Yakovchits, N. V., Adelshin, R. V., Melnikova, O. V., Andaev, E. I., et al. (2019). Epidemiological analysis of rabies outbreaks in the Trans-Baikal Region after transboundary drift of infection. Epidemiol. Infect. Dis. 9, 15–24. doi: 10.18565/epidem.2019.9.3.15-24
Bragina, E. V., Ives, A. R., Pidgeon, A., Kuemmerle, T., Baskin, L., Gubar, Y. P., et al. (2015). Rapid declines of large mammal populations after the collapse of the Soviet Union. Conserv. Biol. 29, 844–853. doi: 10.1111/cobi.12450
Budlyansky, M. A., and Sinilov, A. M. (2019). Dynamics of number and improvement of the mechanism of legal regulation of wolf population in the Irkutsk region. Proc. Lower Volga Agro Univ. Comp. 2, 245–252. doi: 10.32786/2071-9485-2019-02-30
Chervonny, V. V., and Gorbacheva, A. A. (2014). Features of the relationship between the wolf and wild ungulates in the Belgorod region. Sci. Statements Ser. Nat. Sci. 3, 62–71.
Danilov, P. I. (2017). Okhotnich’i zveri Karelii (ekologiya, resursy, upravlenie, okhrana). Petrozavodsk, 385 p. (Hunting Mammals of Kareliy (Ecology, Resorses, Manegment and Conservation). Petrozavodsk: Karelian Scientific Center of the Russian Academy of Sciences Editorial and Publising Departament.
Danilov, P. I., Tirronen, K. F., and Panchenko, D. V. (2020). Moose (Alces alces L.) and wolf Canis lupus L.): monitoring of populations and relationship in the European north of Russia. Bull. Moscow Soc. Nat. Biol. Ser. 125, 13–24. doi: 10.2981/wlb.1997.003
De Groot, G. A., Nowak, C., Skrbinšek, T., Andersen, L. W., Aspi, J., Fumagalli, L., et al. (2016). Decades of population genetic research reveal the need for harmonization of molecular markers: the grey wolf Canis lupus as a case study. Mamm. Rev. 46, 44–59. doi: 10.1111/mam.12052
Elufereva, Y., and Limanova, N. (2020). Modeling population interaction in the natural habitat of the elk and wolf. Bull. Sci. Pract. 6, 288–297. doi: 10.33619/2414-2948/55
Esaulova, N. V., Poyarkov, A. D., Karnaukhov, A. S., Kuksin, A. N., Istomov, S. V., Korablev, M. P., et al. (2015). “On the parasite fauna of the snow leopard (Panthera uncia) and other carnivorous mammals in the ecosystems of the Altai-Khangai-Sayan mountainous country,” in “New Knowledge About Parasites”. Materials of the 5th Interregional Conference “Parasitological Research in Siberia and the Far East” Novosibirsk “Garamond, 43–44.
Esaulova, N. Y., Poyarkov, A. D., Eranadez-Blanco, K. A., and Chistopolova, M. D. (2018). “Endoparasite fauna of wolves in different geographic zones of the Russian Federation. Modern problems of general and applied parasitology,” in Materials of the XII Scientific and Practical Conference in Memory of Professor V.A. Romashova November 1-2, 2018. (Voronezh: Publishing and Printing Center), 35–38.
Gauthier, O., Landry, P. A., and Lapointe, F. J. (2003). Missing data in craniometrics: a simulation study. Acta Theriol. 48, 25–34. doi: 10.1007/BF03194263
Hernandez-Blanco, J. A., and Litvinova, E. M. (2003). “Wolf diet in the south cluster of Kaluzhskiye zaseki Reserve,” in Proceedings of Kaluzhskiye Zaseki Nature Reserve, Tom 1. (Kaluga: Polygraph Inform), 268–274.
Hernandez-Blanco, J. A., Litvinova, E. M., and Poyarkov, A. D. (2003a). “Spatio-ethological organization of Wolf (Canis lupus lupus L., 1758) pack in Kaluzhskiye Zaseki Reserve: pack composition and Home range structure,” in Proceedings of Kaluzhskiye Zaseki Nature Reserve, Tom 1. (Kaluga: Polygraph Inform), 268–274.
Hernandez-Blanco, J. A., Litvinova, E. M., and Poyarkov, A. D. (2003b). “Spatio-ethological organization of Wolf (Canis lupus lupus L., 1758) pack in Kaluzhskiye zaseki Reserve: social interation dynamics and home range use,” in Proceedings of Kaluzhskiye Zaseki Nature Reserve, Tom 1. (Kaluga: Polygraph Inform), 243–267.
Hernandez-Blanco, J. A., Poyarkov, A. D., and Krutova, V. I. (2005). Wolf (Canis lupus lupus) pack organization at the Voronezh Biosphere Reserve. Zool. Zhurnal 84, 80–93.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., and Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978. doi: 10.1002/joc.1276
Hindrikson, M., Remm, J., Pilot, M., Godinho, R., Stronen, A. V., Baltrunaité, L., et al. (2017). Wolf population genetics in Europe: a systematic review, meta-analysis and suggestions for conservation and management. Biol. Rev. 92, 1601–1629. doi: 10.1111/brv.12298
Itin, G. S., and Kravchenko, V. M. (2016). Specific structure of helminthocenoses of wild carnivorous mammals in the landscape-geographical zones of the North-West Caucasus. Theory Pract. Anim. Parasit. Dis. 17, 194–198.
Kabardiev, S. S., and Bittirov, A. M. (2020). Fauna of helminths of the class Cestoda and Nematoda in a wolf (Canis lupus) in Kabardino-Balkaria. Int. Vet. Bull. 2020, 67–74.
Kazmin, V. D., Poyarkov, A. D., and Hernandez-Blanco, J. A. (2001). Habitat, wolf Home range inner structure and ungulates distribution in Voronezh Nature Reserve. Zapovednoye Delo 8, 29–41.
Kirilyuk, A. V., Kirilyuk, V. E., and Ke, R. (2020). Long-distance dispersal of wolves in the Dauria ecoregion. Mamm. Res. 65, 639–646. doi: 10.1007/s13364-020-00515-w
Kirilyuk, A. V., Kirilyuk, V. E., and Minaev, A. N. (2021). Daily activity patterns of wolves in open habitats in the Dauria ecoregion, Russia. Nat. Conserv. Res. 6, 95–109. doi: 10.24189/ncr.2021.049
Kirilyuk, V. E, Kirilyuk, A. V., and Minaev, A. N. (2019). Wolf Home range and movements in Daurian steppe. Nat. Conserv. Res. 4, 91–105. doi: 10.24189/ncr.2019.068
Kolesnikov, V. V., Piminov, V. N., Ekonomov, A. V., Makarova, D. S., Strelnikov, D. P., Sinitsyin, A. A., et al. (2016). Perennial changes and distribution of resources the main species of hunting animals of Russian. Agric. Sci. Eur. North East 6, 56–61.
Kolpashchikov, L. A. (2016). “Impact of the wolf (Canis lupus L., 1758) on the wild reindeer population of Taimyr,” in Large Predator in Holarctic. eds N. K. Zheleznov-Chukotsky and T. Zheleznova (Moscow: U Nikitskich vorot), 74–78.
Konyaev, S. V., and Bondarev, A. Y. (2011). Helminths of a wolf (Canis lupus L.) in Holarctic. Theory Pract. Anim. Parasit. Dis. 12, 246–248.
Korablev, M. P., Korablev, N. P., and Korablev, P. N. (2021). Genetic diversity and population structure of the grey wolf (Canis lupus Linnaeus, 1758) and evidence of wolf × dog hybridisation in the centre of European Russia. Mamm. Biol. 101, 91–104. doi: 10.1007/s42991-020-00074-2
Korablev, N. P., Korablev, P. N., Korablev, M. P., Korablev, A. P., Sedova, N. A., and Zinoviev, A. V. (2021). Variability of craniometric characteristics of Canis lupus, Carnivora, Canidae in the center of European Russia. Nat. Conserv. Res. 6, 50–67. doi: 10.24189/ncr.2021.008
Korolev, A. N. (2016). The dynamics of the range of the wolf (Canis lupus Linnaeus 1758) in the taiga zone of the northeast of European Russia. Bull. Inst. Biol. Komi Ural Branch Russ. Acad. Sci. 2, 2–11.
Kryuchkova, E. N., Buslaev, S. V., and Abalikhin, B. G. (2011). “Parasitofauna of predators of the canine family on the territory of the Central Non-Chernozem region of the Russian Federation,” in Actual Problems and Prospects of Development of the Agro-Industrial Complex. Vol. 79, ed. V. F Petrov (Ivanovo: Sciences Problems Publishing), 99–100.
Lee, E. J., Merriwether, D. A., Kasparov, A. K., Nikolskiy, P. A., Sotnikova, M. V., Pavlova, E. Y., et al. (2015). Ancient DNA Analysis of the oldest canid species from the Siberian arctic and genetic contribution to the domestic dog. PLoS One 10:e0125759. doi: 10.1371/journal.pone.0125759
Loog, L., Thalmann, O., Sinding, M., Schuenemann, V. J., Perri, A., Germonpré, M., et al. (2019). Ancient DNA suggests modern wolves trace their origin to a Late Pleistocene expansion from Beringia. Mol. Ecol. 29, 1596–1610. doi: 10.1111/mec.15329
Magnusson, M. S. (2000). Discovering hidden time patterns in behavior: T-patterns and their detection. Behav. Res. Methods Instruments Comput. 32, 93–110. doi: 10.3758/bf03200792
Malkina, A. V., and Konyaev, S. V. (2013). “Distribution of trichinellosis in the territory Altay Region and Republic of Altay,” In problems of biology and biological educations in pedagogical universities,” in Materials of the VIII All-Russian Scientific and Practical Conference with International Participation (Novosibirsk, March 29-30, 2013).
Manteuffel, P. A. (1949). The Wolf and Its Extermination In Ch. ex. on Hunting Affairs Under the Council of Ministers of the RSFSR. - Physical Culture and Sport, Leningrad “Pec. Dvor”. 56.
Maslennikova, O. V. (2012). To the helmintofauna of the wolf of the European North East of Russia. Modern Prob. Nat. Manage. Hunt. Anim. Husb. 1:360.
Mech, L. D. (1970). The Wolf: The Ecology and Behaviour of an Endangered Species. New York, NY: Natural History press.
Mech, L. D. (1977). Wolf-pack buffer zone prey reservoirs. Science 197, 320–321. doi: 10.1126/science.198.4314.320
Melnik, K. S., Hernandez-Blanco, J. A., Litvinova, E. M., Chistopolova, M. D., Ogurtsov, S. V., and Poyarkov, A. D. (2007). Biotope selectivity and movement features of wolf Canis lupus (Mammalia, Canidae) within the family home range in Kaluzhskie Zaseki reserve. Bull. Moscow Soc. Nat. Biol. Ser. 112, 3–14.
Meltsov, I. V., Ablov, A. M., Shkolnikova, E. N., Koplik, M. E., Minchenko, P. A., Desyatova, T. V., et al. (2020). Lessons learnt from measures taken to prevent rabies introduction and spread into a long rabies free territory (case study of the Irkutsk Oblast). Vet. Sci. Today 3, 154–161. doi: 10.29326/2304-196x-2020-3-34-154-161
Miquelle, D. G., Stephens, P. A., Smirnov, E. N., Goodrich, J. M., Zaumyslova, O. Y., and Myslenkov, A. E. (2005). “Tigers and wolves in the Russian Far East: competitive exclusion, functional redundancy and conservation implications,” in Large Carnivores and The Conservation of Biodiversity, eds J. C. Ray, J. Berger, K. H. Redford, and R. Steneck (Washington, DC: Island Press), 179–207.
Nesterchuk, S. L., Makarova, E. A., and Ostapenko, V. A. (2019). Epizootological features of animal rabies in the Central Federal District of Russia. Trop. Issues Zool. Ecol. Conserv. Issue 2, 102–112.
Ní Leathlobhair, M., Perri, A. R., Irving-Pease, E. K., Witt, K. E., Linderholm, A., Haile, J., et al. (2018). The evolutionary history of dogs in the Americas. Science 361, 81–85. doi: 10.1126/science.aao4776
Nikolskiy, P. A., Sotnikova, M. V., Nikol’skii, A. A., and Pitulko, V. V. (2018). Predomestication and Wolf-human relationships in the arctic Siberia of 30, 000 years ago: evidence from the yana palaeolithic site. High Anthropol. Schl. Univ. 1, 231–262.
Novikova, I. V., and Petrova, O. G. (2015). Modern epizootology of animal rabies. Agrar. Bull. Urals 10, 19–24.
O’Brien, T. (2011). “Abundance, density and relative abundance: a conceptual framework,” in Camera Traps in Animal Ecology, eds A. F. O’Connell, J. D. Nichols, and K. U. Karanth (Berlin: Springer), 71–96. doi: 10.1007/978-4-431-99495-4_6
Ovodov, N. D., and Martynovich, N. V. (2011). Pleistocene gray wolves of Altai and Kuznetsk Alatau. Baikalskij Zool. Žurnal 3, 10–20.
Packard, J. M. (2003). “Wolf behavior: reproductive, social, and intelligent,” in Wolves. Behavior, Ecology, and Conservation, eds D. Mech and L. Boitani (Chicago, IL: University Chicago press), 39–66.
Palmeirim, J. M. (1998). Analysis of skull measurements and measurers: can we use data obtained by various observers? J. Mammal. 79, 1021–1028. doi: 10.2307/1383111
Pilot, M., Jędrzejewski, W., Branicki, W., Sidorovich, V. E., Jędrzejewska, B., Stachura, K., et al. (2006). Ecological factors influence population genetic structure of European grey wolves. Mol. Ecol. 15, 4533–4553. doi: 10.1111/j.1365-294X.2006.03110.x
Poleshchuk, E. M., Sidorov, G. N., and Berezina, E. S. (2012). Animal rabies in Russia in 2007–2011. Russ. Vet. J. 6, 8–12. doi: 10.1038/laban0107-8b
Poleshchuk, E. M., Sidorov, G. N., Nashatyreva, D. N., Gradoboyeva, E. A., Pakskina, N. D., and Popova, I. V. (2019). “Rabies in the Russian federation,” in Information and Analysis Newsletter. (Omsk: Izdatelsky Tsentr KAN), 110.
Ramos-Madrigal, J., Sinding, M. H. S., Carøe, C., Mak, S. S. T., Niemann, J., Samaniego Castruita, J. A., et al. (2021). Genomes of Pleistocene Siberian wolves uncover multiple extinct wolf lineages. Curr. Biol. 31, 198–206. doi: 10.1016/j.cub.2020.10.002
Randi, E. (2011). Genetics and conservation of wolves Canis lupus in Europe. Mamm. Rev. 41, 99–111. doi: 10.1111/j.1365-2907.2010.00176.x
Rigina, E. Y., and Vinogradov, A. V. (2007a). Formation of the mammalian fauna MAMMALIA of the Samara region in the Pleistocene. Ecol. Anim S Russ. Ecol. Dev. 2007, 48–54.
Rigina, E. Y., and Vinogradov, A. V. (2007b). “Regulation of mammal hunting in Samara region N. Modern problems of nature management, hunting and animal husbandry,” in International Scientific and Practical Conference Kirov, VNIOZ RASCHN, 359–360.
Rozhnov, V. V., Chernova, O. F., and Perfilova, T. V. (2011). Species Diagnostics of Deers - Prey of the Amur Tiger (by the Microstructure of Guard Hairs From the Predator Feces). Rockford, IL: KMK press.
Safronov, V. M. (2016). “Assessment of the predatory activity of the wolf (Canis lupus L., 1758) in Yakutia,” in Large Predator in Holarctic. eds N. K. Zheleznov-Chukotsky and T. Zheleznova (Moscow: U Nikitskich vorot), 90–93.
Salkina, G. P., and Eremin, D. Y. (2017). Influence of the number of tigers and wolves on some species of carnivorous mammals in the southeast of Sikhote-Alin. Bull. Texas S. Univ. 22, 984–988.
Sastre, N., Vila, C., Salinas, M., Bologov, V. V., Urios, V., Sanchez, A., et al. (2011). Signatures of demographic bottlenecks in European wolf populations. Conserv. Genet. 12, 701–712. doi: 10.1007/s10592-010-0177-6
Satunin, K. A. (1915). Mammals of the Caucasian Kray. Tiflis: Office of the Viceroy in Caucasius, 410.
Shchepin, S. G., Andrievskaya, Y. G., Kozulina, N. N., Demina, E. A., Sutula, V. I, Perepletkin, D. O., et al. (2019). Estimation of the number and distribution characteristics of the fox (Vulpes vulpes) and wolf (Canis lupus) before and after the detection of rabies epizootics in the Republic of Buryatia. Baikal Zool. J. 3, 119–127.
Shinkarenko, A. N., and Kolesnikov, P. V. (2011). Helminth fauna of a wolf at the territory of the Volgograd Region. Volgograd State Agricultural Academy. Theory Pract. Anim. Parasit. Dis. 12, 564–565.
Sidorova, D. G., Sidorov, G. N., Poleschuk, E. M., and Kolychev, N. M. (2007). Rabies in East Siberia in XX – beginning XXI. Acta Biomed. Sci. 3S, 168–172.
Sinding, M.-H., Gopalakrishnan, S., Ramos Madrigal, J., de Manuel, M., Pitulko, V. V., Kuderna, L., et al. (2020). Arctic-adapted dogs emerged at the Pleistocene–Holocene transition. Science 368, 1495–1499. doi: 10.1126/science.aaz8599
Skryabin, K. I. (1928). The Method of Complete Helmintological Dissectus of Vertebrate Including Humans. Moscow: Moscow State University, 46.
Stepanova, V. V., and Okhlopkov, I. M. (2020). The current status of the number and density of the wolf (Canis lupus Linnaeus, 1758) population in Yakutia. Nat. Resour. Arctic Subarctic 25, 121–133. doi: 10.31242/2618-9712-2020-25-4-10
Sukhomesova, M. V. (2013). Features of the Bioresource Potential of Carnivorous Mammals of the Northern Macroslope of the Central Caucasus. Ph.D. thesis. Vladikavkaz: Polygraphic section of the CPI KBSU.
Suvorov, A. P. (2016a). “In defense of the polar wolf (Canis lupus L., 1758) of the Yenisei North,” in Large Predator in Holarctic, eds N. K. Zheleznov-Chukotsky and T. Zheleznova (Moscow: U Nikitskich vorot), 78–84.
Suvorov, A. P. (2016b). “About irrational “exterminatory” regulation of the number of wolves,” in Large Predator in Holarctic, eds N. K. Zheleznov-Chukotsky and T. Zheleznova (Moscow: U Nikitskich vorot), 84–89.
Suvorov, A. P. (2017a). Geographic variability of parameters of the body of the wolf of the Yenisei Siberia. Vestn. Krasgau 7, 119–125.
Suvorov, A. P. (2017b). Geographical variability of the skull of the wolf in the Yenisei Siberia. Vestn. Krasgau. 12, 178–184.
Suvorov, A. P., and Kirienko, N. N. (2008). Features of self-regulation in wolf populations of the Yenisei Siberia. Bull. Krasgau 2008, 106–110.
Talala, M. S., Bondarev, A. Y., Zakharov, E. S., and Politov, D. V. (2020). Genetic differentiation of the wolf Canis lupus L. populations from Siberia at microsatellite loci. Russ. J. Genet. 1, 59–68. doi: 10.1134/S1022795420010123
Teerink, B. (1991). Hair of Western-European mammals: Atlas and Identification Key. New York, NY: Cambridge University Press, 224.
Tembotova, F. A., and Kononenko, E. P. (2007a). “Variability of craniometric features of the wolf (Canidae, Carnivora) in the Caucasus report 1. Sex-associated variation,” in Mammals of Mountain Areas. Materials of the International Conference, eds V. V. Rozhnov and F. A. Tembotova (Moscow: KMK publishing), 323–330.
Tembotova, F. A., and Kononenko, E. P. (2007b). “Variability of craniometric features of the wolf (Canidae, Carnivora) in the Caucasus report 2. Geographic variation,” in Mammals of Mountain Areas. Materials of the International Conference, eds V. V. Rozhnov and F. A. Tembotova (Moscow: KMK publishing), 331–339.
Tkachenko, K. N. (2010). Nutrition of wolf (Canis lupus Linnaeus, 1758, Carnivora: Canidae) in the Bolshekhekhtsirskii Nature Reserve. Amur. Zool. J. 2, 276–281.
Tulov, A. V., Zverjanovskii, M. I., Yanagida, T., Konyaev, S. V., Andreyanov, O. N., Malkina, A. V., et al. (2013). The species and genetic diversity of trichinella from members of the canine family (Canidae) in Russia. Actual Issues Vet. Biol. 17, 35–41.
Volkov, A. E. (2020). Forest roads and glades as places for organizing monitoring of large carnivorous mammals using camera traps (on the example of the Kerzhensky Reserve). Biota Environ. 4, 68–93.
Voloshina, I. V., Myslenkov, A. I., Sieber, A., and Radeloff, V. C. (2014). “Habitat modeling on Amur tiger (Panthera tigris) and grey wolf (Canis lupus) in the south of the Sikhote-Alin mountains,” in Distribution, Migration and Other Movements of Wildlife Proceedings of the International Conference (Vladivostok, 25–27 November 2014), eds A. P. Saveljev and I. V. Seryodkin (Vladivostok: Reya), 40–52.
Vorobiev, V. N. (2015). Long-term dynamics of the number of hunting game animals of the Verkhne-Kondinsky reserve (Khanty-Mansiysk Autonomous Okrug - Yugra). Fauna Urals Siberia 2015, 107–118.
Vorobyevskaya, E. A., and Baldina, S. N. (2011). Altai wolf phylogeography (Canis lupus L.) studied by microsatellite markers. Moscow Univ. Biol. Sci. Bull. 66, 53–54. doi: 10.3103/S0096392511020131
Yachmennikova, A. F., and Poyarkov, A. D. (2011). A new approach to study organization of wolves’ activity (Canis lupus) in time sequences. Biol. Bull. 38, 156–164. doi: 10.1134/s1062359011020154
Yachmennikova, AA, and Poyarkov, A. D. (2010). “New approach to study of social behavior and activity in wolf group in ontogeny,” in Proceedings of the 2nd Canine Science Forum (Viena: Viena Univerity Press).
Yachmennikova, AA, Poyarkov, A. D., Blidchenko, EY., and Badridze, Y. K. (2009). “Integrated schedule of ontogenesis stages of wolf (Canis lupus),” in Material of International Union of Game Biologist XXIX Congres (Moscow: MAI Print), 31–32.
Yarovenko, A. Y. (2015). “Review of information on wolves and their numbers in the Republic of Dagestan,” in Modern Trends in the Development of Science and Technology. Collection of Scientific Papers Based on the Materials of the II International Scientific and Practical Conference, ed. E. P. Tkacheva (Belgorod: Epicentre), 104–107.
Zakharova, O. I. (2019). Rabies of wild carnivores in the tundra zone of Yakutia. Sci.Pract. J. Bull. IrGSKhA 91, 118–124.
Zheleznov-Chukotsky, N. K. (2016). “The myth and truth about the wolf (Canis lupus L., 1758) retrospective and modernity,” in Large Predator in Holarctic. eds N. K. Zheleznov-Chukotsky and T. Zheleznova (Moscow: U Nikitskich vorot), 95–104.
Zheltukhin, A. S., and Ogurtsov, S. S. (2018). Camera Traps for Monitoring of the Forest Mammals and Birds. Tver: Tver press, 54.
Keywords: wolf, Canis lupus, Russia, population management, behavioral ecology, population genetic and morphology
Citation: Poyarkov AD, Korablev MP, Bragina E and Hernandez-Blanco JA (2022) Overview of Current Research on Wolves in Russia. Front. Ecol. Evol. 10:869161. doi: 10.3389/fevo.2022.869161
Received: 03 February 2022; Accepted: 29 March 2022;
Published: 12 July 2022.
Edited by:
Gyaneshwer Chaubey, Banaras Hindu University, IndiaReviewed by:
Katarzyna Bojarska, Polish Academy of Sciences (PAN), PolandDiego Ortega-Del Vecchyo, National Autonomous University of Mexico, Mexico
Copyright © 2022 Poyarkov, Korablev, Bragina and Hernandez-Blanco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrey D. Poyarkov, cG95YXJrb3ZAeWFuZGV4LnJ1
†These authors have contributed equally to this work