Potential alternative tree species to Fraxinus excelsior in European forests
 Mathieu Lévesque
Mathieu Lévesque José Ignacio Bustamante Eduardo1†
José Ignacio Bustamante Eduardo1†  Valentin Queloz
Valentin Queloz- 1Silviculture Group, Department of Environmental Systems Science, Institute of Terrestrial Ecosystems, ETH Zurich, Zurich, Switzerland
- 2Forest Health and Biotic Interactions, Swiss Federal Research Institute for Forest, Snow, and Landscape Research WSL, Birmensdorf, Switzerland
Common ash (Fraxinus excelsior L.) is a keystone tree species in Europe. However, since the 1990s, this species has been experiencing widespread decline and mortality due to ash dieback [Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz and Hosoya]. Besides H. fraxineus, emerald ash borer (Agrilus planipennis Fairmaire), an invasive alien pest already devastating ash trees in western Russia, is spreading westward and becoming an emerging threat to the remaining European ash populations. While efforts to control ash dieback continue to be a priority, it is becoming crucial to compensate for the loss of ash and its ecosystem services by elaborating restoration strategies, including the search for alternative native and non-native tree species. This review summarizes available knowledge on potential alternative tree species to common ash to help forest managers to cope with ash dieback. Although using natural regeneration and promoting tree species diversity can reduce the impacts of ash dieback in European forests, our review indicates that no native species alone or in combination can fully replace the ecological niche of common ash and its associated ecosystem services (e.g., biodiversity and timber). To fill this gap, forest managers have considered using non-native ash species that are tolerant to both H. fraxineus and A. planipennis and have similar ecological and forestry values as common ash. Of the 43 ash species reviewed, few non-native ash species (e.g., Fraxinus mandshurica Rupr. and Fraxinus platypoda Oliv.) have similar ecological characteristics to common ash and are tolerant to H. fraxineus and A. planipennis. However, the performance of non-native ash species in European forests, their invasiveness potential, and the risk of hybridization with native ash species are still unknown. With the current state of knowledge, it is thus too early to recommend the use of non-native ash species as a suitable option to deal with ash dieback. The priority should be the conservation, regeneration, and breeding of tolerant common ash populations to H. fraxineus, as well as the use of the natural regeneration of other native tree species. Our review highlights the need for controlled experimental plantations to better understand the regeneration ecology and invasiveness potential of non-native ash species prior to their utilization in natural forests.
1. Introduction
Common ash (Fraxinus excelsior L.) is one of the most abundant broadleaved tree species in Europe, extending from the boreal forests of southern Scandinavia to southern Europe (Wardle, 1961; Fraxigen, 2005; Dobrowolska et al., 2011; Thomas, 2016). Common ash has a high physiological plasticity and occurs on various sites but prefers fertile soils with pH above 5.5 (Wardle, 1961; Fraxigen, 2005). It is widely present in moist and riparian lowland forests, dry calcareous sites, and up to 1,600–1,800 m a.s.l. (Wardle, 1961; Fraxigen, 2005; Kollas et al., 2014). Common ash is found in various mixed forest associations in Europe (Leuschner and Ellenberg, 2017), and it is a keystone species for a wide range of taxa (Pautasso et al., 2013; Mitchell et al., 2014; Littlewood et al., 2015). For example, in the UK, common ash is part of 61 plant community types and has over 950 species associated with it (Mitchell et al., 2014; Broome and Mitchell, 2017). The bark of ash is rich in nutrients and has a high pH, creating an essential niche for fungi, mosses, and insects (Mitchell et al., 2016). Littlewood et al. (2015) reported that at least 36 invertebrate species depend exclusively on ash, and 38 species are highly associated. In addition to its keystone role for biodiversity conservation (Hultberg et al., 2020), common ash is also essential in stabilizing riverbanks and slopes, producing a rich and easily decomposable litter, slowing wildfires (Enderle et al., 2019), and for medicinal purposes (Spadaro and Raimondo, 2022).
Besides its ecological functions, common ash is a valuable broadleaved species with excellent wood properties (Dobrowolska et al., 2011). Ash wood is malleable yet strong, so it is appreciated for its elasticity, hardness, and pressure resistance (Niemz et al., 2014). Historically, ash was used to construct weapon handles, agricultural tools, and carriages (Pratt, 2017). Today, ash wood is still widely used for tool handles, sports equipment, flooring, and veneers (Beck et al., 2016). After the drastic abundance reduction of elms (Ulmus spp.) in Europe due to the Dutch elm disease (Ophiostoma ulmi and novo-ulmi Brasier), the abundance and ecological importance of ash has increased, particularly in floodplain forests where ash and elm were dominating (Leuschner and Ellenberg, 2017). Unfortunately, the numerous ecosystem services provided by common ash are now severely compromised by the invasive fungal pathogen Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz and Hosoya (Baral et al., 2014), causing unprecedented tree dieback and mortality rates across Europe (Coker et al., 2019).
First observed in Poland in 1992 (Kowalski and Łukomska, 2005), ash dieback is a disease causing a progressive crown reduction through branch dieback and root lesions triggering secondary infections by Armillaria spp. (Cleary et al., 2016; Landolt et al., 2016; Marçais et al., 2022). Hymenoscyphus fraxineus was first described scientifically in 2006 as its anamorphic stage Chalara fraxinea Kowalski (2006). Four years later, Chalara fraxinea was found to have a sexual stage first believed to be Hymenoscyphus albidus and shortly after revised to Hymenoscyphus pseudoalbidus (Queloz et al., 2011), and finally taxonomically renamed Hymenoscyphus fraxineus (Baral et al., 2014). The fungus is highly virulent such that most ash trees are now infected in Europe (Marçais et al., 2022), with mortality rates up to 100% in natural forests (George et al., 2022). In its native range in Asia, the causal agent H. fraxineus is found in the leaf litter of Fraxinus mandshurica Rupr. and Fraxinus chinensis Roxb. [synonym Fraxinus rhynchophylla Hance (Wallander, 2008)] (Rigling et al., 2016; Inoue et al., 2019). Dieback symptoms are not present in the native range of the fungus, while inoculation experiments have shown different levels of infection in F. mandshurica in Europe (Gross et al., 2015a; Drenkhan et al., 2017). Over the past 20 years, research on ash dieback in Europe has primarily focused on the causal agent and finding genotypes and populations resistant to H. fraxineus to reduce the risk of common ash being lost from its current distribution (Enderle et al., 2019). This research direction was motivated by the ineffectiveness of forest protection or phytosanitary measures in Europe to prevent the spread of and the impacts caused by this virulent pathogen.
In addition to the search for solutions to control ash dieback, the emerald ash borer (EAB) (Agrilus planipennis Fairmaire), an invasive phloem-feeding beetle, is increasingly occupying the scientific agenda. EAB is native to the forests of Northeast Asia and was accidentally introduced in Europe prior 2003 (Valenta et al., 2017). In its native range, this insect is a minor pest of Fraxinus mandshurica, Fraxinus chinensis, and Fraxinus lanuginosa Koidz. and only attacks stressed trees (Valenta et al., 2017). However, outside its native range, EAB is a devastating pest and attacks other ash species and vigorous trees. For example, since its first detection in 2002 in North America, EAB has killed tens of millions of ash trees and devastated thousands of hectares of ash forests in the United States and Canada (Herms and McCullough, 2014). Today, 20 years after the EAB was first detected, six ash species in North America are now endangered or critically endangered (Herms and McCullough, 2014). In Europe, Fraxinus americana L. and Fraxinus pennsylvanica Marshall were until recently considered as potential alternative non-native species to replace common ash (Broome et al., 2019). However, these two North American ash species have a limited tolerance to ash dieback (Nielsen et al., 2017). Further, since 2003, many F. pennsylvanica plantations in the region of Moscow have been infested with EAB and have facilitated the establishment of this species in western Russia (Valenta et al., 2017; Musolin et al., 2021; Volkovitsh et al., 2021). The latest studies indicate that EAB is spreading rapidly westward and is now present in St. Petersburg (Selikhovkin et al., 2022) as well as in Ukraine and Belarus (Drogvalenko et al., 2019; Musolin et al., 2021). It is expected that EAB will reach Central Europe within the next 5–10 years (Valenta et al., 2017) and could potentially decimate the rest of the ash populations that have until now survived ash dieback.
In its natural range, EAB populations only infest stressed trees and are controlled by parasitoids (Valenta et al., 2017). Furthermore, higher larval performance and egg-laying preference of EAB on drought-stressed F. mandshurica indicate that traits in the phloem may be linked to resistance to EAB (Showalter et al., 2018). In Europe, efforts to prevent the introduction and stop the spread of EAB remain a priority for preserving native ash species, especially since trees weakened by ash dieback may be attractive hosts for the beetle. Infestation experiments with EAB show a high susceptibility of all three native European ash species (F. excelsior, Fraxinus angustifolia Vahl, Fraxinus ornus L.), suggesting a bleak future for the genus Fraxinus in Europe if EAB continues to expand (Musolin et al., 2021).
Given the severe and ongoing ash dieback across Europe and the potential future threat of EAB, experts have called for the search for resistant ash trees and replacement tree species to counteract the progressive disappearance of ash and ensure the provision of ecosystem services (Littlewood et al., 2015; Leuthardt et al., 2017; Marzano et al., 2019; Agostinelli et al., 2021). In order to sustain ash-specific ecosystem services in forests severely impacted by ash dieback, one forest management option that could be envisaged is the use of non-native ash species that have similar ecological and silvicultural attributes to common ash and exhibit resistance against H. fraxineus and EAB. However, it is not yet clear which non-native ash species could be suitable alternatives to common ash because a detailed assessment of their ecological similarities and characteristics (i.e., regeneration, growth, and invasion potential) is still missing.
The objectives of the present literature review were to (1) examine and collate published literature on ash dieback research in Europe; (2) identify native and non-native tree species that could replace common ash to ensure the provisioning and continuity of ecosystem services and sustain biodiversity associated with ash; and (3) discuss the benefits, risks, and limitations of non-native ash species in European forests.
2. Materials and methods
Our literature search was conducted using electronic bibliographic databases, namely Google Scholar, Web of Science, and Scopus, and included published scientific literature (articles and book chapters), doctoral and master theses, and reports. The following terms were searched in titles, keywords, and abstracts: “ash dieback” OR “Hymenoscyphus fraxineus” OR “Hymenoscyphus pseudoalbidus” OR “Chalara fraxinea” AND “Fraxinus” AND “tree regeneration” AND “tree replacement.” The search yielded 541 records published between 1992 and June 2022, of which 200 were retained after evaluating abstracts and titles. We used the following evaluation criteria for selection: (1) the work focused on ash dieback; (2) the studies were not limited to identifying or confirming the presence of ash dieback; and (3) the abstracts were available in English. For publication records that met these criteria, we categorized them by country and research themes such as forest management, regeneration, replacement tree species, and non-native ash species.
3. Results and discussion
3.1. Ash dieback in Europe: Impacts and research directions
The health status of ash stands has deteriorated rapidly since the arrival of the ash dieback disease in Europe, with mortality rates of up to 70% in forests and 85% in plantations, across Europe (Coker et al., 2019; Enderle et al., 2019). The most recent data from the European forest vitality monitoring program of the International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP Forests) Forests,1 indicate mortality of up to 100% in some locations [for country-specific infestation levels and mortality rates see George et al. (2022)]. Ash dieback and mortality rates are exceptionally high for small-diameter trees, on flat and wet sites, and where ash density is high (Enderle et al., 2018; Grosdidier et al., 2020; Klesse et al., 2021). However, a small fraction of individuals resistant or less susceptible to the disease suggests the possibility that common ash may survive (Queloz et al., 2017; Evans, 2019). Model simulations also suggest that climate change and H. fraxineus may interact to affect the future spatial distribution of F. excelsior as both species may extend their range poleward, hence reducing ash dieback prevalence in temperate European regions (Goberville et al., 2016). Still, it is unclear how the pathogen and common ash will coevolve and influence ash mortality.
In the last three decades, research on ash dieback has explored various research fields and yielded many scientific achievements (Supplementary Table 1). Early research about ash dieback focused essentially on the identification of the causal agent of the new disease and its pathogenicity (Kowalski, 2006; Kowalski and Holdenrieder, 2009a,b; Queloz et al., 2011; Gross et al., 2014a), and the disease cycle (Gross et al., 2012). Following the identification of the causal agent of ash dieback, research was conducted on the origin of the new pathogen species with the use of microsatellite markers and populations from impacted regions in Europe (Gross et al., 2014b; Burokiene et al., 2015; Haňáčková et al., 2015). In parallel, the native range of H. fraxineus in eastern Asia was identified (Zhao et al., 2013; Drenkhan et al., 2017), and its pathogenicity in its natural environment was investigated (Cleary et al., 2016). These early research directions have yielded fundamental information on the pathogen, hosts, and disease cycle that have stimulated further research to cope with ash dieback, such as the search for mycoviruses as biological control of H. fraxineus (e.g., Schoebel et al., 2014; Shamsi et al., 2022). At present, whether the use of mycoviruses can be successfully used as biocontrol of H. fraxineus remains unclear because this method relies strongly on the spread and hypovirulence of mycoviruses within H. fraxineus populations, which the fungal host population may hinder (Gross et al., 2014b; Burokiene et al., 2015). Further work investigating virus-mediated controls of ash dieback in Europe is thus necessary (Shamsi et al., 2022) before using mycoviruses for biocontrol of ash dieback. More recently, studies have investigated the potential of fungal endophytes for biocontrol of ash dieback (e.g., Schlegel et al., 2016; Kosawang et al., 2018; Halecker et al., 2020). These works have identified promising endophytes, but future studies will be needed to assess their full potential as biocontrol agents.
Besides fundamental research on H. fraxineus to cope with ash dieback, the research agenda in many European countries has also focused on the identification of resistant ash genotypes and the initiation of breeding programs (McKinney et al., 2014; Cleary et al., 2017; Skovsgaard et al., 2017). The establishment of breeding programs has been motivated by the fact that in situ conservation of viable populations of healthy ash may be limited because of the very low percentage of resistant genotypes to H. fraxineus (1–5% of the ash trees), as well as population fragmentation and isolation between resistant trees (Skovsgaard et al., 2017). Although it remains mainly unknown which mechanisms are responsible for the genetic variation in resistance, there is robust evidence that variation in resistance is transmitted from parents to offspring (Marçais et al., 2022). Particularly, the moderate levels of narrow-sense heritability (0.3–0.5) for resistance reported in the literature (McKinney et al., 2011; Pliûra et al., 2011; Kjær et al., 2012; Muñoz et al., 2016; Plumb et al., 2020) indicates good perspective for breeding programs because resistant ash trees can be identified, tested in plantation trials and selected for seed production (Marçais et al., 2022). Selections of resistant ash genotypes from planted trials will also be more effective for breeding programs than selecting healthy individuals growing in heterogenous forest environments where heritability may be significantly less (Kjær et al., 2017; Plumb et al., 2020). However, it should be noted that the heritability at the mature stage is relevant for the selection of candidate mature and resistant ash individuals in the forests for breeding, but this heritability is not yet known (Kjær et al., 2017).
To date, many European countries have initiated breeding programs, and most of them focus on F. excelsior (Douglas et al., 2013; Vasaitis and Enderle, 2017; Marçais et al., 2022) with a few exceptions where F. angustifolia is also tested, for example, in Slovenia (Hauptman et al., 2016) and Austria (Unger et al., 2021). In Sweden, several seed orchards with selected plus-tree clones have been monitored for disease susceptibility since 2006 (Stener, 2018; Liziniewicz et al., 2022). Researchers have reported a stable resistance of resistant F. excelsior clones over a 15-year period, promising results for the continuation of breeding programs. Besides, a breeding trial in Denmark has recently yielded the first commercial seed lot of resistant ash (Kjær et al., 2017; Marçais et al., 2022). Despite the scientific advances in breeding programs in the last two decades, it is not yet known if it will be feasible to breed common ash to have resistance to both H. fraxineus and A. planipennis, and further research is needed to fully evaluate the viability and feasibility of future breeding programs (Plumb et al., 2020). Therefore, breeding for pathogen or pest resistance should be considered as a mid- to long-term silvicultural option to cope with ash dieback (Skovsgaard et al., 2017).
3.2. Forest management to cope with ash dieback
Managing forests impacted by ash dieback involves several challenges, including harvesting timber before depreciation, removing dead/dying and dangerous trees, retaining healthy trees, promoting regeneration, and maintaining ecosystem services (Skovsgaard et al., 2017). Interestingly, before the advent of ash dieback, ash’s high regeneration capacity and colonization potential under a wide range of ecological conditions and its “invasive” potential were sometimes a concern for forest managers (Rysavy and Roloff, 1994; Marigo et al., 2000; Fraxigen, 2005; Dobrowolska et al., 2011). Today, tree regeneration and the health of ash populations have shown a severe and widespread decline across Europe, impacting recruitment rates and forest succession (Diaz-Yanez et al., 2020). Although ash dieback resistance levels may improve with subsequent generations (Harper et al., 2016), it seems unlikely that the natural regeneration of healthy ash alone will be sufficient to maintain stable populations in ash-dominated forests or ash typical sites (Turczanski et al., 2021). Steep declines in ash populations and shifts in species composition can already be observed in ash stands with high rates of dieback and mortality of young ash trees (Lygis et al., 2014; Coker et al., 2019). With mortality rates of over 80% reported in young ash stands in Europe (Coker et al., 2019) and the high susceptibility of planting material to ash dieback in nurseries (Chavez et al., 2015), establishing reforestation programs with common ash becomes a challenge. Therefore, the regeneration of sites impacted by ash dieback, either naturally or through planting, is an important issue. From a silvicultural point of view, the natural regeneration of healthy ash trees should be the priority to preserve the genetic diversity of ash populations resistant to ash dieback, and planting should be the exception if we want to minimize the costs and risks associated with artificial regeneration.
European silvicultural programs are attempting to implement the most current knowledge in their strategies to minimize the impacts of ash dieback. Silvicultural strategies include (1) monitoring, safe elimination of infested plant materials, and alternating fungicides in nurseries (Hrabětová et al., 2017; Noble et al., 2019); (2) identifying and breeding ash dieback-resistant genotypes to H. fraxineus to build immunity in native populations (Plumb et al., 2020); (3) monitoring the health status of ash stands (Chandelier et al., 2011); (4) creating and maintaining mixed stands with vigorous ash individuals (Rosenvald et al., 2015; Short and Hawe, 2019); (5) adapting silvicultural regimes to minimize the impact of ash dieback by adjusting the intensity of thinnings based on dieback level and site conditions (Short and Hawe, 2019); and (6) focusing conservation effort on sites with low infection rates such as dry and warm sites and sites with low ash density (Grosdidier et al., 2020; Klesse et al., 2021). Along with these silvicultural strategies, efforts have been made to identify species that can potentially replace European ash (Broome et al., 2019).
3.3. Potential replacement species to ash in forests strongly impacted by ash dieback
The search for replacement species is particularly a priority for ash-dominated moist sites where ash mortality is high. In theory, replacement native and non-native species should have similar ecological niche and silvicultural characteristics to common ash. For example, potential species should have a broad ecological range, be present along a wide elevational gradient, tolerate a variety of soil moisture and nutrient conditions, and be present throughout forest succession. Ideally, replacement species should also fulfill most ecosystem services and ecological functions specific to common ash, such as habitats for ash-associated species and timber (Broome et al., 2019). Importantly, criteria for selecting replacement species may vary across biogeographic zones and European countries since forest communities, forest management, and silvicultural practices are not uniform across the range of common ash. Table 1 summarizes the native and non-native species reported in forest inventories conducted in forests impacted by ash dieback in selected European countries.
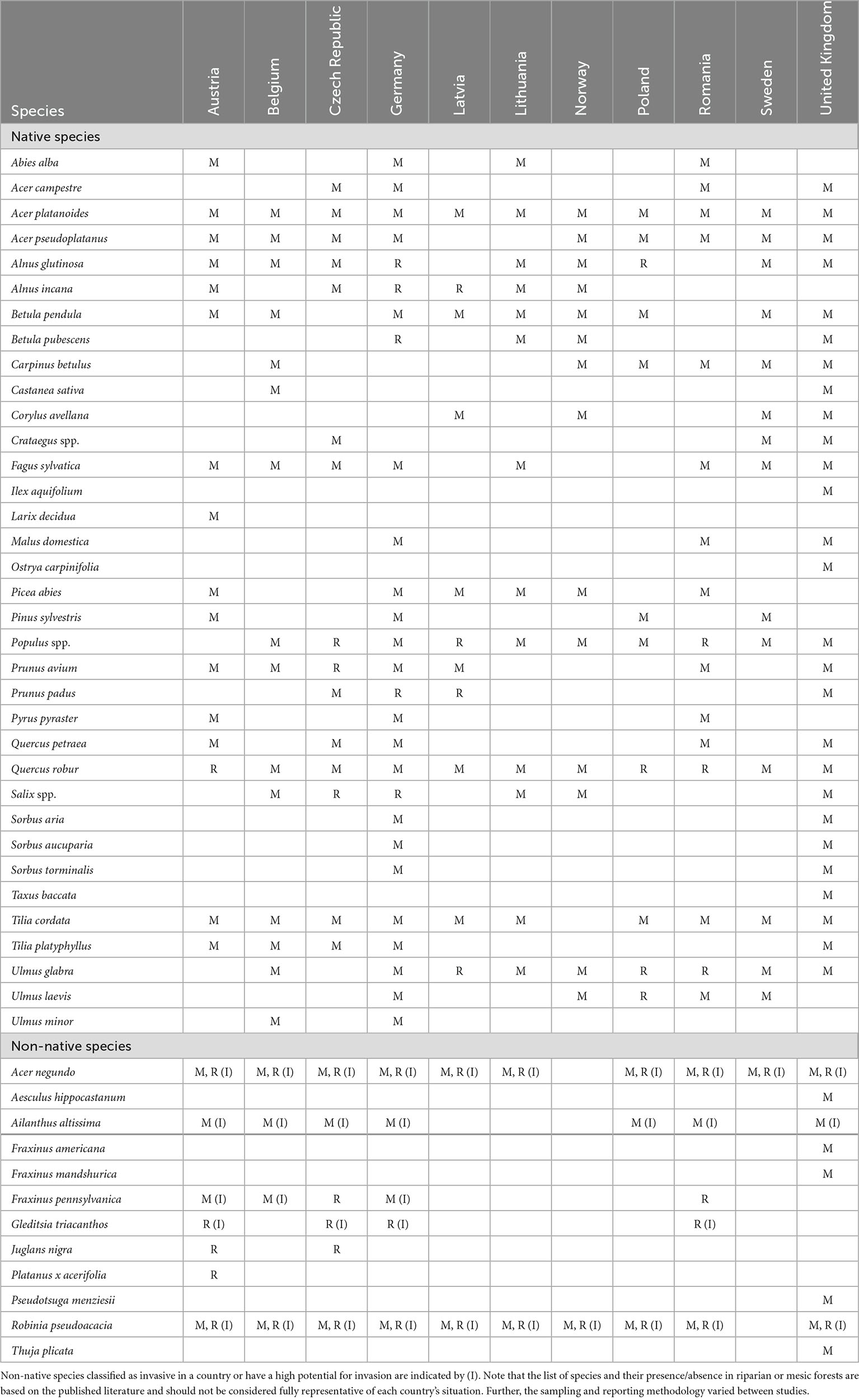
Table 1. According to the literature review, tree species that have been recorded in riparian (R) and mesic (M) forests impacted by ash dieback in some European countries.
Of the list of species presented in Table 1, many species have little or no ecological similarity to common ash (e.g., conifers) and therefore are not logical candidates to replace ash. Further, five non-native species reported in Table 1 [Acer negundo L., Ailanthus altissima (Mill.) Swingle, Fraxinus pennsylvanica, Gleditsia triacanthos L., and Robinia pseudoacacia L.] are classified as invasive in many European countries or have a high potential for invasion. As a result, the threat to the native ash species and their associated biodiversity can be exacerbated if these non-native tree species are successful invaders and occupy the niche of the native ash species (Ennos et al., 2019). Three of the 12 non-native species reported in some European countries (Table 1) are closely related taxa (Fraxinus americana, Fraxinus mandshurica, and Fraxinus pennsylvanica) and could potentially support biodiversity associated with common ash. However, F. americana and F. pennsylvanica show a limited resistance to ash dieback (Nielsen et al., 2017), and F. pennsylvanica also has a good invasiveness potential in riparian forests in Europe (Schmiedel et al., 2013; Drescher and Prots, 2016). The Asian ash species (F. mandshurica) shows some resistance to H. fraxineus and A. planipennis, but it is not yet known if this species can become invasive outside its native range.
Table 2 lists the tree species with the most ecological similarities to common ash that could potentially serve as replacement species for ash. The list contains 15 native tree species and is based on the work of Mitchell et al. (2016) and Broome et al. (2019). The species are characterized by their similarities to common ash based on their litter characteristics, ecosystem functions, and tolerance to site conditions. Results presented in Table 2 highlight that no native species alone can replace the ecological niche and ecosystem functions of common ash and that a combination of native species may potentially, although only partly, substitute ash.
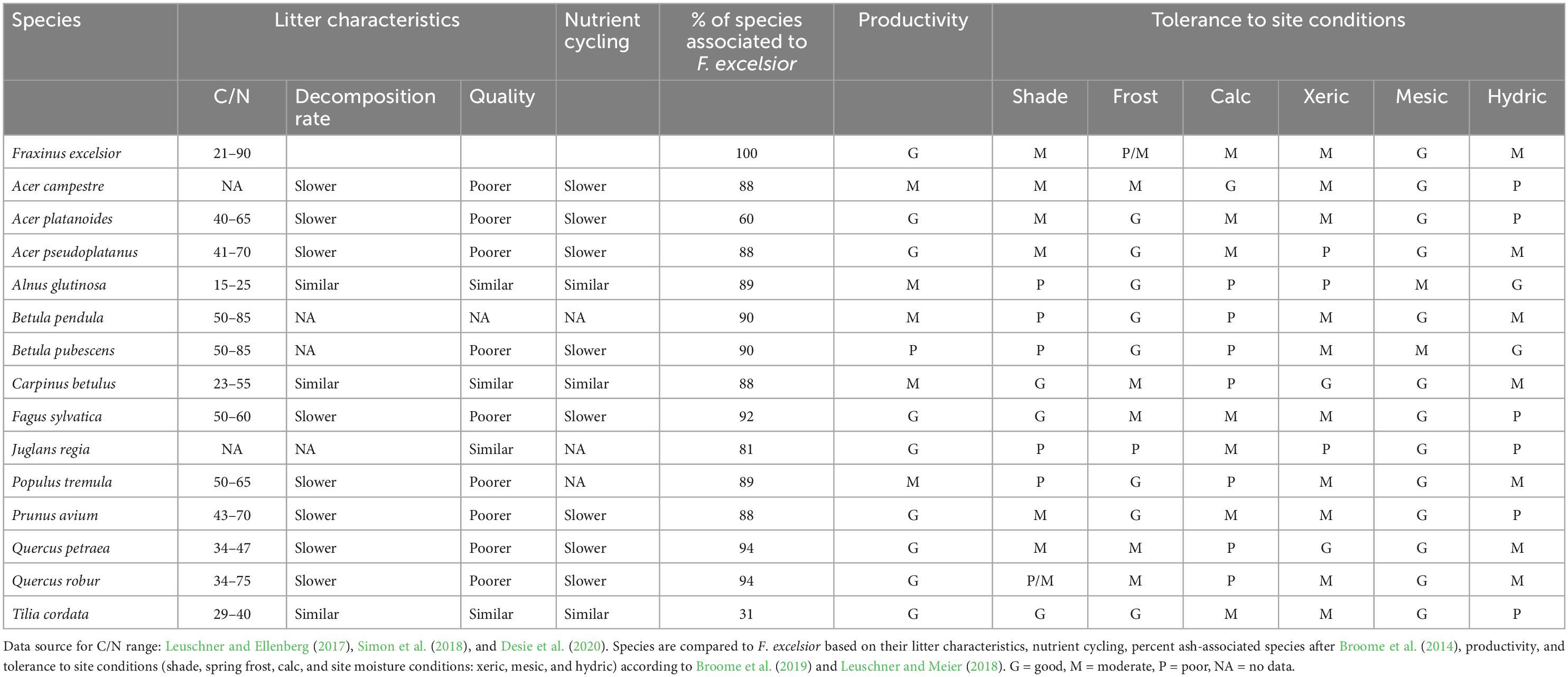
Table 2. Potential alternative native species to Fraxinus excelsior after Mitchell et al. (2016) and Broome et al. (2019).
For example, a recent study reported that up to nine tree species would be needed to maintain all common ash-dependent species (Hultberg et al., 2020). The species studied included Acer platanoides L., Alnus spp., Betula spp., Corylus avellana L., Fagus sylvatica L., Juniperus communis L., Picea abies (L.) H. Karst., Pinus sylvestris L., Populus tremula L., Prunus spp., Quercus spp., Salix spp., Sorbus aucuparia L., Ulmus spp., and Tilia spp. Of these species, Quercus spp., Populus tremula, Acer pseudoplatanus, and Fagus sylvatica were estimated to support 95% of the organisms associated with common ash (Hultberg et al., 2020). The dominance of Acer pseudoplatanus over Fagus sylvatica and Quercus robur L. after ash dieback was also demonstrated after running simulations with eight broadleaved species under different ash survival scenarios (Evans, 2019). Acer pseudoplatanus abundance increased after ash dieback under all scenarios and was only limited under dry conditions (Evans, 2019).
Other studies have also recommended including Picea abies and Pinus sylvestris in mixtures with broadleaved species to buffer fluctuations in wood prices following ash dieback (Hultberg et al., 2020; Petucco et al., 2020). While this measure may reduce potential economic risks, it is likely not universally applicable as it depends on existing site conditions and the regional forest socioeconomic context. In addition, the inclusion of conifers will not fill the ecological niche of common ash and will not ensure ecosystem services directly related to ash. Although native broadleaved and conifer species may partly replace common ash (Broome et al., 2019), native species alone or in combination may not fulfill all ecosystem services specific to common ash (e.g., habitats for ash-dependent species, ash timber). Altogether, our literature review highlights the need also to consider non-native ash species that exhibit resistance against ash dieback and EAB and share ecological and forestry similarities with common ash as potential alternative species if we want to support the ecological functions and biodiversity associated with native European ash species.
3.4. Are non-native ash species an alternative to ash species native to Europe?
The perspective of replacing native ash species with closely related non-native species could be a potential solution to the economic and ecological challenges caused by ash dieback in Europe (Marzano et al., 2019). At least four criteria should be considered when selecting non-native ash species. First, non-native ash species should be from the same section as the native European ash species (Wallander, 2008), as closely related species are likely to host the same associated organisms (Cleary et al., 2016). Second, non-native ash species must have similarities in ecological requirements and growth form. Third, alternative species must be resistant to H. fraxineus and EAB. Fourth, species must not be invasive and cause potential problems in European forests. It is not yet clear which non-native ash species meet each of these criteria because there are many ash species, and only some of them have been studied outside their native range.
The genus Fraxinus comprises 43 recognized species (three in Europe and western Asia, 20 in eastern Asia, and 20 in North America) distributed throughout the Northern Hemisphere ranging from temperate rainforests to xerophytic scrub forests (Wallander, 2008 and Supplementary Table 2). Fraxinus excelsior and F. angustifolia belong to the section Fraxinus, which includes F. nigra and F. mandshurica. The sections Sciadanthus, Pauciflora, and Melioides are closely related to the section Fraxinus, while the sections Ornus, to which F. ornus belongs, and Dipetalae are more phylogenetically distant. Outside of the section Fraxinus, very few species share the same ecology or growth form as common ash since most have a bush or small tree form. Despite the different growth forms, the most decisive factor in selecting a non-native ash species remains its resistance to H. fraxineus and EAB, as well as its potential invasiveness and possible impacts on native ecosystems.
Of the list of non-native ash species presented in Table 3 that could potentially replace F. excelsior, F. mandshurica is an interesting candidate. This species has good timber quality and is one of the most valuable hardwood species in China (Han-ying and Dai-bin, 2003). F. mandshurica has similar characteristics than common ash (e.g., height and pollination) and shows some resistance to H. fraxineus and A. planipennis. In its native range, F. mandshurica is host to ash dieback pathogen and emerald ash borer, infesting exclusively stressed trees. Laboratory resistance experiments showed a low degree of susceptibility of F. mandshurica to H. fraxineus (Nielsen et al., 2017) and agreed with the good resistance to the pathogen recorded in experimental plantations in Estonia, Sweden, and Switzerland (Drenkhan et al., 2014; Cleary et al., 2016; Queloz et al., 2017). In Estonia, typical symptoms of ash dieback were reported on 40-year-old F. mandshurica planted in parks, and H. fraxineus was isolated from the shoots of trees showing these symptoms (Drenkhan and Hanso, 2010). However, F. mandshurica was very little affected and showed only mild symptoms, including leaf wilting, minor shoot and twig diebacks, and sometimes the presence of bark necrosis. To avoid a situation analogous to common ash dieback in introduced populations of F. mandshurica, experiments with inoculation of the endemic species Hymenoscyphus albidus, the European sister species of H. fraxineus, were conducted, and H. albidus caused no symptoms to inoculated F. mandshurica (Gross et al., 2015a).
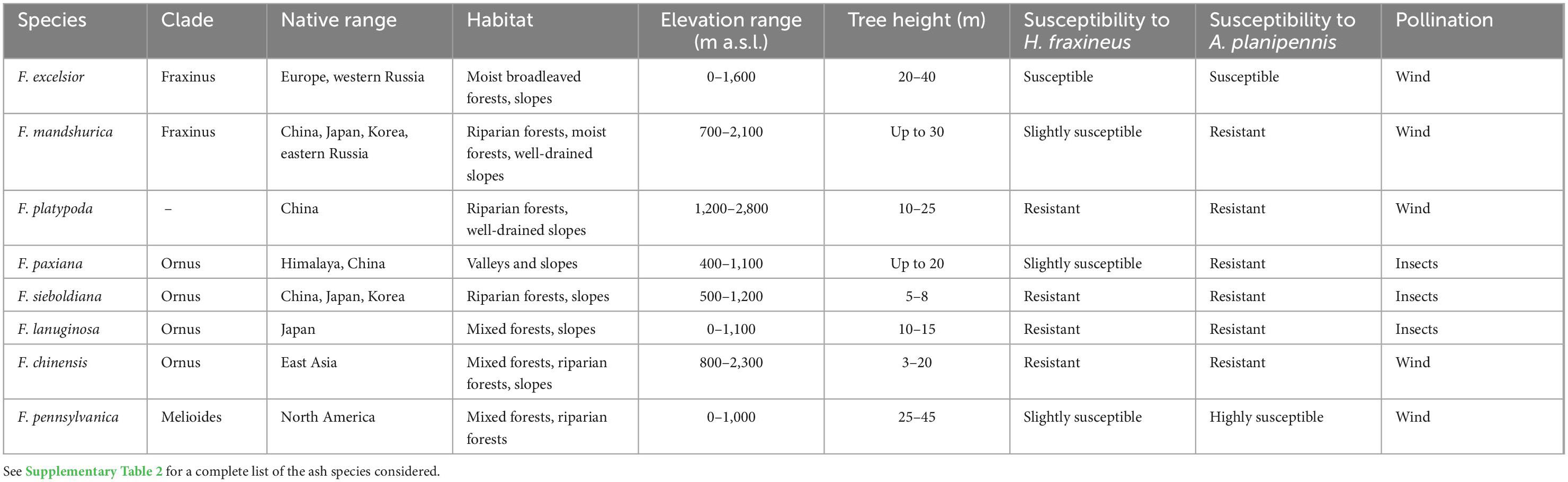
Table 3. Characteristics of Fraxinus excelsior (as comparison) and non-native ash species having the highest replacement potential identified in the literature.
Although F. mandshurica was widespread in East Asia in the past, this species has suffered from deforestation and overexploitation and is now classified as a species vulnerable to extinction in China (Barstow et al., 2018). Conservation efforts for F. mandshurica focus on implementing propagation and regeneration programs in its native range (Liu et al., 2021) and reducing herbaceous competition to stimulate regeneration (Wang et al., 2013). In Switzerland, this species was introduced into the Arboretum Vallon de l’Aubonne about a decade ago. Work by Queloz et al. (2017) suggests that H. fraxineus is a frequent pathogen on F. mandshurica and that it is capable, to some extent, of inducing necrotic lesions on the twigs and stems of this species. In addition, monitoring of the plantings in the arboretum shows that F. mandshurica is sensitive to late frosts and droughts (personal communication of Sylvain Meier). In the United States (Minnesota), plantations with F. mandshurica conducted in 2011–2012 (Looney et al., 2015) showed survival rates of 22–58% eight years after planting (Palik et al., 2021). Regarding the ability of F. mandshurica to naturally regenerate outside of its range and its invasiveness potential, our literature review reports no studies on this topic. Therefore, it is premature to conclude that F. mandshurica could be used as a replacement species for common ash in Europe. Further studies, including controlled experimental plantations, are needed to better understand F. mandshurica’s ecology and invasiveness potential outside its natural range before its utilization in European forests.
Another non-native ash species with good potential as a replacement species is Fraxinus platypoda Oliv. This valuable timber species occurs primarily in riparian forests but is also found in other environments in its native range in Japan and China (Sakio, 2020). F. platypoda can regenerate easily and abundantly through a high production of seeds and vegetative shoots, which is increased after natural disturbances (Sakio et al., 2002; Sakio, 2020). However, F. platypoda regeneration is sensitive to browsing, and it is also unclear how global warming may influence seed production (Sakio, 2020). F. platypoda also attains a similar height to F. excelsior and is resistant to winter frost (Sakio, 1997). In its native range, F. platypoda hosts a non-virulent common ash fungus, Hymenoscyphus linearis, which is closely related to H. fraxineus. Hybridization of the two fungi appears possible, and it is a potential risk that should not be ignored (Gross et al., 2015b). Thus, if this species is introduced, appropriate phytosanitary measures must be taken to avoid such a threat. Despite this limitation and the small range of F. platypoda, its good regeneration potential and germination plasticity make this species a possible alternative to common ash in Europe. Further, the study of Nielsen et al. (2017) done on specimens of F. platypoda growing in an arboretum in Denmark indicated that this species showed no disease symptoms after inoculation with H. fraxineus. Arboretum assessments with F. platypoda in Switzerland confirm these results (Queloz et al., 2017). Until now, this species has been little studied, and it is thus too early to give clear recommendations about its use in Europe.
In addition to F. mandshurica and F. platypoda, other Asian species deserve attention as alternatives to F. excelsior, and Fraxinus paxiana Lingelsh. is one of them. This species has low susceptibility to H. fraxineus and occurs at similar elevations as common ash (Zhi and Green, 1996; Nielsen et al., 2017). Until now, little is known about the regeneration ecology of F. paxiana outside its natural range. With a broader distribution range, Fraxinus chinensis has an ecological niche similar to that of F. excelsior and occurs in mixed and riparian forests and at various elevations. This species is a host for H. fraxineus in its native range, but it is less susceptible to leaf infection than F. excelsior or F. mandshurica (Nielsen et al., 2017).
Like other species in the Ornus section, Fraxinus lanuginosa and Fraxinus sieboldiana Blume are both resistant to H. fraxineus (Nielsen et al., 2017). Both species thrive in moist forests and well-drained soils but are both sensitive to late frosts. In its native range, F. lanuginosa shows no evidence of crown or leaf damage caused by H. fraxineus (Nielsen et al., 2017). F. sieboldiana frequently co-occurs with Quercus mongolica Turchaninov in Korea (Gab-Cheul and Gab-Tae, 2005). This species produces abundant regeneration after fires and can grow rapidly and vigorously (Goto et al., 1996). The wood of F. lanuginosa is strong and hard, and it is used for making sports equipment such as baseball bats (Muto and Koizumi, 2007). In contrast to F. excelsior, F. sieboldiana is a large shrub or small (5–8 m), strongly branched ash species little suitable as timber species.
Fraxinus pennsylvanica is a widely distributed ash species in North America and was introduced to Europe in the 18th century, where it is used for ornamental purposes and timber production (Schmiedel et al., 2013). In the native range of F. pennsylvanica in North America, none of the currently described Hymenoscyphus species has been found until now (Kowalski and Bilański, 2019). This is not the case in Europe where, for example, H. fraxineus and H. pusillus occur on F. pennsylvanica (Kowalski and Bilański, 2019). Several studies have reported some degree of resistance of F. pennsylvanica to H. fraxineus and less susceptibility to the pathogen than F. excelsior (Drenkhan and Hanso, 2010; Gross and Sieber, 2016; Nielsen et al., 2017). Nevertheless, F. pennsylvanica is highly susceptible to EAB, and this exotic pest is already causing severe damage to F. pennsylvanica in European Russia and Ukraine (Orlova-Bienkowskaja et al., 2020). Moreover, F. pennsylvanica has been classified as an invasive species in many European riparian forests with negative impacts on local flora diversity (Schmiedel et al., 2013; Drescher and Prots, 2016), although a recent study suggests that it might become a rescue species for the fauna associated to F. excelsior (Floren et al., 2022). To date, there is no robust scientific evidence supporting the use of F. pennsylvanica as a potential alternative species to F. excelsior in European forests.
As evidenced by the invasiveness of F. pennsylvanica in many European forests (Schmiedel and Tackenberg, 2013; Schmiedel et al., 2013; Drescher and Prots, 2016), there are several attributes of non-native ash species that could promote their establishment, growth, and invasive character. All ash species have wind-dispersed seeds, mature early, and are strong competitors in the early stages of forest succession. In addition, although most ash species prefer mesic environments, they often possess broad ecological plasticity that can favor their establishment in various environments. Although some non-native ash species share similarities to common ash regarding their habitats and growth forms (Table 3 and Supplementary Table 2), little is known about the ecological behavior, adaptability, and invasiveness of non-native ash species outside their natural range. It is also unknown whether non-native Asian ash species can hybridize with native European ash populations and what is the risk of genomic invasion. In Europe, interspecific hybridization frequently occurs between F. excelsior and F. angustifolia (Fernández-Manjarrés et al., 2006; Douglas et al., 2013; Thomasset et al., 2014), but further work is necessary to determine whether natural hybridization can also occur between Asian and European ash species and how climate change may influence their hybridization potential. Thus, it is a high priority that additional studies and controlled planting experiments be conducted to fill these knowledge gaps before using non-native ash species for reforestation in European forests. Further, before introducing and recommending non-native ash species as potential replacement species for European ash species, it is important to have comprehensive risk assessments (Ennos et al., 2019).
4. Conclusion and perspectives
The abundance and vitality of common ash in Europe have declined rapidly since the first report of H. fraxineus in Poland in the early 1990s. Most ash trees in Europe show dieback symptoms, and high mortality rates have been reported. Despite research efforts over the past three decades and scientific advances, there is still no effective control solution against ash dieback. Furthermore, the future of ash and the ecosystem services provided by this species seem to be increasingly threatened by the upcoming arrival of A. planipennis in Central Europe. From a practical point of view, searching for replacement species for common ash seems to be a valid option to ensure the provision of ecosystem services where ash plays an essential role. However, our literature review indicates that few studies have addressed this topic. Furthermore, existing studies (e.g., Mitchell et al., 2016; Broome et al., 2019) have focused on tree regeneration and species composition of forests impacted by ash dieback. These studies indicate that some native species may, to some extent, complementarily cover the ecological niche and sustain biodiversity associated with ash (Table 3). Since few native species possess similar characteristics to common ash and any of them have similar timber characteristics, the search for replacement species should additionally include non-native ash species. However, special attention should be given to avoid introducing new invasive tree species.
Our literature review showed that a few non-native ash species, such as F. mandshurica, F. platypoda, and F. chinensis, possess similar stature and habitats to common ash while exhibiting resistance to H. fraxineus and A. planipennis (Table 3 and Supplementary Table 2). However, the ecology of these species and their invasiveness potential outside their native range are still unknown. No studies have addressed these aspects, and no controlled and carefully monitored experimental plantations with non-native ash species exist in Europe. A few experimental plantations with F. mandshurica exist in the United States (e.g., Looney et al., 2015; Palik et al., 2021), but these plantation trials have focused on the survival of young non-native ash trees and have not provided information on the invasive potential of F. mandshurica. In Europe, some Asian ash species are present in arboreta (e.g., L’Arboretum du Vallon de l’Aubonne in Switzerland, Arboretum in Hørsholm in Denmark) and parks (e.g., Drenkhan and Hanso, 2010). Still, no long-term continuous and rigorous monitoring has been carried out. Due to the lack of research on non-native ash species in Europe, it is not possible to determine whether F. mandshurica or other non-native ash species represent valid substitutes for common ash and what risks they could pose when introduced in European forests. Thus, establishing experimental plantations where non-native ash trees are monitored over a long period would allow us to learn more about their ecology and potential benefits and impacts on forest ecosystems. These plantations should be established on sites with contrasting edaphic and climatic conditions and monitored rigorously to better inform about species’ regeneration ecology and minimize the risk of invasion.
Author contributions
ML and JIBE performed the literature review and wrote the first draft. VQ provided expertise and feedback during the drafting process. ML and VQ acquired the funding. ML supervised the project. All authors contributed to the final version and approved the submitted version.
Funding
This work was supported by Swiss Federal Office for the Environment (FOEN). Project « Espèces d’arbres de remplacement pour le frêne commun en Suisse », grant number: 00.5053.PZ/7368828B9. Open access funding provided by ETH Zurich.
Acknowledgments
We thank Joana Meyer and Robert Jenni from the Swiss Federal Office for the Environment (FOEN) for the constructive feedback during the drafting process.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2023.1048971/full#supplementary-material
Footnotes
References
Agostinelli, M., Nguyen, D., Witzell, J., and Cleary, M. (2021). Mycobiome of Fraxinus excelsior with different phenotypic susceptibility to ash dieback. Front. For. Glob. Change 4:580514. doi: 10.3389/ffgc.2021.580514
Baral, H. O., Queloz, V., and Hosoya, T. (2014). Hymenoscyphus fraxineus, the correct scientific name for the fungus causing ash dieback in Europe. IMA Fungus 5, 79–80. doi: 10.5598/imafungus.2014.05.01.09
Barstow, M., Oldfield, S., Westwood, M., Jerome, D., Beech, E., and Rivers, M. (2018). The red list of Fraxinus. Richmond, VA: Botanic Gardens Conservation International.
Beck, P., Caudullo, G., Tinner, W., and de Rigo, D. (2016). “Fraxinus excelsior in Europe: distribution, habitat, usage and threats,” in European atlas of forest tree species, eds J. San-Miguel-Ayanz, D. de Rigo, G. Caudullo, T. Houston Durrant, and A. Mauri (Luxembourg: Publ. Off), e0181c0180.
Broome, A., and Mitchell, R. (2017). Ecological impacts of ash dieback and mitigation methods. Monmouthshire: Forestry Commission.
Broome, A., Mitchell, R. J., and Harmer, R. (2014). Ash dieback and loss of biodiversity: Can management make broadleaved woodlands more resilient? Q. J. For. 108, 241–248.
Broome, A., Ray, D., Mitchell, R., and Harmer, R. (2019). Responding to ash dieback (Hymenoscyphus fraxineus) in the UK: Woodland composition and replacement tree species. For. Int. J. For. Res. 92, 108–119. doi: 10.1093/forestry/cpy040
Burokiene, D., Prospero, S., Jung, E., Marciulyniene, D., Moosbrugger, K., Norkute, G., et al. (2015). Genetic population structure of the invasive ash dieback pathogen Hymenoscyphus fraxineus in its expanding range. Biol. Invasion. 17, 2743–2756. doi: 10.1007/s10530-015-0911-6
Chandelier, A., Delhaye, N., and Helson, M. (2011). First report of the ash dieback pathogen Hymenoscyphus pseudoalbidus (Anamorph Chalara fraxinea) on Fraxinus excelsior in Belgium. Plant Dis. 95, 220–220. doi: 10.1094/PDIS-07-10-0540
Chavez, V. A., Parnell, S., and Bosch, F. V. D. (2015). Designing strategies for epidemic control in a tree nursery: The case of ash dieback in the UK. Forests 6, 4135–4145. doi: 10.3390/f6114135
Cleary, M., Nguyen, D., Marciulyniene, D., Berlin, A., Vasaitis, R., and Stenlid, J. (2016). Friend or foe? Biological and ecological traits of the European ash dieback pathogen Hymenoscyphus fraxineus in its native environment. Sci. Rep. 6:21895. doi: 10.1038/srep21895
Cleary, M., Nguyen, D., Stener, L., Stenlid, J., and Skovsgaard, J. (2017). “Ash and ash dieback in Sweden: A review of disease history, current status, pathogen and host dynamics, host tolerance and management options in forests and landscapes,” in Dieback of European Ash (Fraxinus spp.): Consequences and guidelines for sustainable management, eds R. Vasaitis and R. Enderle (Uppsala: Swedish University of Agricultural Sciences), 195–208.
Coker, T. L., Rozsypálek, J., Edwards, A., Harwood, T. P., Butfoy, L., and Buggs, R. J. (2019). Estimating mortality rates of European ash (Fraxinus excelsior) under the ash dieback (Hymenoscyphus fraxineus) epidemic. Plants People Planet 1, 48–58. doi: 10.1002/ppp3.11
Desie, E., Vancampenhout, K., Nyssen, B., van den Berg, L., Weijters, M., van Duinen, G.-J., et al. (2020). Litter quality and the law of the most limiting: Opportunities for restoring nutrient cycles in acidified forest soils. Sci. Total Environ. 699:134383. doi: 10.1016/j.scitotenv.2019.134383
Diaz-Yanez, O., Mola-Yudego, B., Timmermann, V., Tollefsrud, M. M., Hietala, A. M., and Oliva, J. (2020). The invasive forest pathogen Hymenoscyphus fraxineus boosts mortality and triggers niche replacement of European ash (Fraxinus excelsior). Sci. Rep. 10:5310. doi: 10.1038/s41598-020-61990-4
Dobrowolska, D., Hein, S., Oosterbaan, A., Wagner, S., Clark, J., and Skovsgaard, J. P. (2011). A review of European ash (Fraxinus excelsior L.): Implications for silviculture. For. Int. J. For. Res. 84, 133–148. doi: 10.1093/forestry/cpr001
Douglas, G. C., Pliura, A., Dufour, J., Mertens, P., Jacques, D., Fernandez-Manjares, J., et al. (2013). “Common Ash (Fraxinus excelsior L.),” in Forest tree breeding in Europe: Current state-of-the-art and perspectives, ed. L. E. Pâques (Dordrecht: Springer Netherlands), 403–462. doi: 10.1007/978-94-007-6146-9_9
Drenkhan, R., and Hanso, M. (2010). New host species for Chalara fraxinea. New Dis. Rep. 22, 16–16. doi: 10.5197/j.2044-0588.2010.022.016
Drenkhan, R., Sander, H., and Hanso, M. (2014). Introduction of Mandshurian ash (Fraxinus mandshurica Rupr.) to Estonia: Is it related to the current epidemic on European ash (F. excelsior L.)? Eur. J. For. Res. 133, 769–781. doi: 10.1007/s10342-014-0811-9
Drenkhan, R., Solheim, H., Bogacheva, A., Riit, T., Adamson, K., Drenkhan, T., et al. (2017). Hymenoscyphus fraxineus is a leaf pathogen of local Fraxinus species in the Russian Far East. Plant Pathol. 66, 490–500. doi: 10.1111/ppa.12588
Drescher, A., and Prots, B. (2016). Fraxinus pennsylvanica – an invasive tree species in middle Europe: Case studies from the Danube basin. Contributii Botanice 51, 55–69.
Drogvalenko, A. N., Orlova-Bienkowskaja, M. J., and Bienkowski, A. O. (2019). Record of the emerald ash borer (Agrilus planipennis) in Ukraine is confirmed. Insects 10:338. doi: 10.3390/insects10100338
Enderle, R., Metzler, B., Riemer, U., and Kändler, G. (2018). Ash dieback on sample points of the national forest inventory in south-western Germany. Forests 9:25. doi: 10.3390/f9010025
Enderle, R., Stenlid, J., and Vasaitis, R. (2019). An overview of ash (Fraxinus spp.) and the ash dieback disease in Europe. CAB Rev. 14, 1–12. doi: 10.1079/PAVSNNR201914025
Ennos, R., Cottrell, J., Hall, J., and O’Brien, D. (2019). Is the introduction of novel exotic forest tree species a rational response to rapid environmental change? – A British perspective. For. Ecol. Manag. 432, 718–728. doi: 10.1016/j.foreco.2018.10.018
Evans, M. R. (2019). Will natural resistance result in populations of ash trees remaining in British woodlands after a century of ash dieback disease? R. Soc. Open Sci. 6:190908. doi: 10.1098/rsos.190908
Fernández-Manjarrés, J. F., Gérard, P. R., Dufour, J., Raquin, C., and Frascaria-Lacoste, N. (2006). Differential patterns of morphological and molecular hybridization between Fraxinus excelsior L. and Fraxinus angustifolia Vahl (Oleaceae) in eastern and western France. Mol. Ecol. 15, 3245–3257. doi: 10.1111/j.1365-294X.2006.02975.x
Floren, A., Horchler, P. J., and Müller, T. (2022). The impact of the neophyte tree Fraxinus pennsylvanica [Marshall] on beetle diversity under climate change. Sustainability 14:1914. doi: 10.3390/su14031914
Fraxigen (2005). Ash species in Europe: Biological characteristics and practical guidelines for sustainable use. Oxford: University of Oxford.
Gab-Cheul, C., and Gab-Tae, K. (2005). Vegetation structure of mountain ridge from Bubong to Poamsan in Baekdudaegan, Korea. Korean J. Environ. Ecol. 19, 83–89.
George, J.-P., Sanders, T. G. M., Timmermann, V., Potočić, N., and Lang, M. (2022). European-wide forest monitoring substantiate the neccessity for a joint conservation strategy to rescue European ash species (Fraxinus spp.). Sci. Rep. 12:4764. doi: 10.1038/s41598-022-08825-6
Goberville, E., Hautekèete, N.-C., Kirby, R. R., Piquot, Y., Luczak, C., and Beaugrand, G. (2016). Climate change and the ash dieback crisis. Sci. Rep. 6:35303. doi: 10.1038/srep35303
Goto, Y., Yoshitake, T., Okano, M., and Shimada, K. (1996). Seedling regeneration and vegetative resprouting after fires in Pinus densiflora forests. Vegetatio 122, 157–165. doi: 10.1007/BF00044698
Grosdidier, M., Scordia, T., Ioos, R., Marçais, B., and Thrall, P. (2020). Landscape epidemiology of ash dieback. J. Ecol. 108, 1789–1799. doi: 10.1111/1365-2745.13383
Gross, A., and Sieber, T. N. (2016). Virulence of Hymenoscyphus albidus and native and introduced Hymenoscyphus fraxineus on Fraxinus excelsior and Fraxinus pennsylvanica. Plant Pathol. 65, 655–663. doi: 10.1111/ppa.12450
Gross, A., Holdenrieder, O., and Woodward, S. (2015a). Pathogenicity of Hymenoscyphus fraxineus and Hymenoscyphus albidus towards Fraxinus mandshurica var. japonica. For. Pathol. 45, 172–174. doi: 10.1111/efp.12182
Gross, A., Hosoya, T., Zhao, Y. J., and Baral, H. O. (2015b). Hymenoscyphus linearis sp. nov: another close relative of the ash dieback pathogen H. fraxineus. Mycol. Prog. 14, 1–15. doi: 10.1007/s11557-015-1041-3
Gross, A., Holdenrieder, O., Pautasso, M., Queloz, V., and Sieber, T. N. (2014a). Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol. Plant Pathol. 15, 5–21. doi: 10.1111/mpp.12073
Gross, A., Hosoya, T., and Queloz, V. (2014b). Population structure of the invasive forest pathogen Hymenoscyphus pseudoalbidus. Mol. Ecol. 23, 2943–2960. doi: 10.1111/mec.12792
Gross, A., Zaffarano, P. L., Duo, A., and Grünig, C. R. (2012). Reproductive mode and life cycle of the ash dieback pathogen Hymenoscyphus pseudoalbidus. Fungal Genet. Biol. 49, 977–986. doi: 10.1016/j.fgb.2012.08.008
Halecker, S., Wennrich, J. P., Rodrigo, S., Andree, N., Rabsch, L., Baschien, C., et al. (2020). Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 45:100918. doi: 10.1016/j.funeco.2020.100918
Haňáčková, Z., Koukol, O., Havrdová, L., and Gross, A. (2015). Local population structure of Hymenoscyphus fraxineus surveyed by an enlarged set of microsatellite markers. For. Pathol. 45, 400–407. doi: 10.1111/efp.12185
Han-ying, S., and Dai-bin, L. (2003). Influence of main site factors on Fraxinus mandshurica (Oleaceae) plantation. J. For. Res. 14, 83–86. doi: 10.1007/BF02856770
Harper, A. L., McKinney, L. V., Nielsen, L. R., Havlickova, L., Li, Y., Trick, M., et al. (2016). Molecular markers for tolerance of European ash (Fraxinus excelsior) to dieback disease identified using Associative Transcriptomics. Scientific Reports 6, 19335. doi: 10.1038/srep19335
Hauptman, T., Ogris, N., de Groot, M., Piškur, B., and Jurc, D. (2016). Individual resistance of Fraxinus angustifolia clones to ash dieback. For. Pathol. 46, 269–280. doi: 10.1111/efp.12253
Herms, D. A., and McCullough, D. G. (2014). Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 59, 13–30. doi: 10.1146/annurev-ento-011613-162051
Hrabětová, M., Černý, K., Zahradník, D., and Havrdová, L. (2017). Efficacy of fungicides on Hymenoscyphus fraxineus and their potential for control of ash dieback in forest nurseries. For. Pathol. 47:e12311. doi: 10.1111/efp.12311
Hultberg, T., Sandström, J., Felton, A., Öhman, K., Rönnberg, J., Witzell, J., et al. (2020). Ash dieback risks an extinction cascade. Biol. Conserv. 244:108516. doi: 10.1016/j.biocon.2020.108516
Inoue, T., Okane, I., Ishiga, Y., Degawa, Y., Hosoya, T., and Yamaoka, Y. (2019). The life cycle of Hymenoscyphus fraxineus on Manchurian ash, Fraxinus mandshurica, in Japan. Mycoscience 60, 89–94. doi: 10.1016/j.myc.2018.12.003
Kjær, E. D., McKinney, L. V., Nielsen, L. R., Hansen, L. N., and Hansen, J. K. (2012). Adaptive potential of ash (Fraxinus excelsior) populations against the novel emerging pathogen Hymenoscyphus pseudoalbidus. Evol. Appl. 5, 219–228. doi: 10.1111/j.1752-4571.2011.00222.x
Kjær, E., McKinney, L., Hansen, L., Olrik, D., Lobo, A., Thomsen, I., et al. (2017). “Genetics of ash dieback resistance in a restoration context – experiences from Denmark,” in Dieback of European Ash (Fraxinus spp.): Consequences and guidelines for sustainable management, eds R. Vasaitis and R. Enderle (Uppsala: Swedish University of Agricultural Sciences), 106–114.
Klesse, S., Abegg, M., Hopf, S. E., Gossner, M. M., Rigling, A., and Queloz, V. (2021). Spread and severity of ash dieback in Switzerland – Tree characteristics and landscape features explain varying mortality probability. Front. For. Glob. Change 4:645920. doi: 10.3389/ffgc.2021.645920
Kollas, C., Körner, C., and Randin, C. F. (2014). Spring frost and growing season length co-control the cold range limits of broad-leaved trees. J. Biogeogr. 41, 773–783. doi: 10.1111/jbi.12238
Kosawang, C., Amby, D. B., Bussaban, B., McKinney, L. V., Xu, J., Kjaer, E. D., et al. (2018). Fungal communities associated with species of Fraxinus tolerant to ash dieback, and their potential for biological control. Fungal Biol. 122, 110–120. doi: 10.1016/j.funbio.2017.11.002
Kowalski, T. (2006). Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 36, 264–270. doi: 10.1111/j.1439-0329.2006.00453.x
Kowalski, T., and Bilański, P. (2019). Hymenoscyphus pusillus, a new species on leaves of Fraxinus pennsylvanica in Poland. For. Pathol. 49:e12481. doi: 10.1111/efp.12481
Kowalski, T., and Holdenrieder, O. (2009a). Pathogenicity of Chalara fraxinea. For. Pathol. 39, 1–7. doi: 10.1111/j.1439-0329.2008.00565.x
Kowalski, T., and Holdenrieder, O. (2009b). The teleomorph of Chalara fraxinea, the causal agent of ash dieback. For. Pathol. 39, 304–308. doi: 10.1111/j.1439-0329.2008.00589.x
Kowalski, T., and Łukomska, A. (2005). The studies on ash dying (Fraxinus excelsior L.) in the Włoszczowa Forest Unit stands. Acta Agrobotanica 58, 429–440. doi: 10.5586/aa.2005.068
Landolt, J., Gross, A., Holdenrieder, O., and Pautasso, M. (2016). Ash dieback due to Hymenoscyphus fraxineus: What can be learnt from evolutionary ecology? Plant Pathol. 65, 1056–1070. doi: 10.1111/ppa.12539
Leuschner, C., and Ellenberg, H. (2017). Ecology of central european forests. Vegetation ecology of central europe, Vol. I., Berline: Springer. doi: 10.1007/978-3-319-43042-3
Leuschner, C., and Meier, I. C. (2018). The ecology of central European tree species: Trait spectra, functional trade-offs, and ecological classification of adult trees. Perspect. Plant Ecol. Evol. Syst. 33, 89–103. doi: 10.1016/j.ppees.2018.05.003
Leuthardt, F., Queloz, V., Rigling, D., Lüchinger, R., Wegmann, S., and Zumbrunnen, T. (2017). Strategie des bundes und der kantone zum eschentriebsterben. Bern: Bundesamt für Umwelt BAFU.
Littlewood, N. A., Nau, B. S., Pozsgai, G., Stockan, J. A., Stubbs, A., and Young, M. R. (2015). Invertebrate species at risk from ash dieback in the UK. J. Insect Conserv. 19, 75–85. doi: 10.1007/s10841-014-9745-2
Liu, L., Cao, Y., Zhan, Y., and Qi, F. (2021). A rapid and efficient protocol for adventitious shoot regeneration and genetic transformation of Manchurian ash (Fraxinus mandshurica Rupr.) using hypocotyl explants. [preprint] Res. Square doi: 10.21203/rs.3.rs-60987/v1
Liziniewicz, M., Tolio, B., and Cleary, M. (2022). Monitoring of long-term tolerance of European ash to Hymenoscyphus fraxineus in clonal seed orchards in Sweden. For. Pathol. 52:e12773. doi: 10.1111/efp.12773
Looney, C. E., D’Amato, A. W., Palik, B. J., and Slesak, R. A. (2015). Overstory treatment and planting season affect survival of replacement tree species in emerald ash borer threatened Fraxinus nigra forests in Minnesota, USA. Can. J. For. Res. 45, 1728–1738. doi: 10.1139/cjfr-2015-0129
Lygis, V., Bakys, R., Gustiene, A., Burokiene, D., Matelis, A., and Vasaitis, R. (2014). Forest self-regeneration following clear-felling of dieback-affected Fraxinus excelsior: Focus on ash. Eur. J. For. Res. 133, 501–510. doi: 10.1007/s10342-014-0780-z
Marçais, B., Kosawang, C., Laubray, S., Kjær, E., and Kirisits, T. (2022). “Chapter 13 – ash dieback,” in Forest microbiology, eds F. O. Asiegbu and A. Kovalchuk (Massachusetts, MA: Academic Press), 215–237. doi: 10.1016/B978-0-323-85042-1.00022-7
Marigo, G., Peltier, J.-P., Girel, J., and Pautou, G. (2000). Success in the demographic expansion of Fraxinus excelsior L. Trees 15, 1–13. doi: 10.1007/s004680000061
Marzano, M., Woodcock, P., and Quine, C. P. (2019). Dealing with dieback: Forest manager attitudes towards developing resistant ash trees in the United Kingdom. For. Int. J. For. Res. 92, 554–567. doi: 10.1093/forestry/cpz022
McKinney, L. V., Nielsen, L. R., Collinge, D. B., Thomsen, I. M., Hansen, J. K., and Kjær, E. D. (2014). The ash dieback crisis: Genetic variation in resistance can prove a long-term solution. Plant Pathol. 63, 485–499. doi: 10.1111/ppa.12196
McKinney, L. V., Nielsen, L. R., Hansen, J. K., and Kjær, E. D. (2011). Presence of natural genetic resistance in Fraxinus excelsior (Oleraceae) to Chalara fraxinea (Ascomycota): An emerging infectious disease. Heredity 106, 788–797. doi: 10.1038/hdy.2010.119
Mitchell, R. J., Beaton, J. K., Bellamy, P. E., Broome, A., Chetcuti, J., Eaton, S., et al. (2014). Ash dieback in the UK: A review of the ecological and conservation implications and potential management options. Biol. Conserv. 175, 95–109. doi: 10.1016/j.biocon.2014.04.019
Mitchell, R. J., Hewison, R. L., Hester, A. J., Broome, A., and Kirby, K. J. (2016). Potential impacts of the loss of Fraxinus excelsior (Oleaceae) due to ash dieback on woodland vegetation in Great Britain. N. J. Bot. 6, 2–15. doi: 10.1080/20423489.2016.1171454
Muñoz, F., Marçais, B., Dufour, J., and Dowkiw, A. (2016). Rising out of the ashes: Additive genetic variation for crown and collar resistance to Hymenoscyphus fraxineus in Fraxinus excelsior. Phytopathology 106, 1535–1543. doi: 10.1094/PHYTO-11-15-0284-R
Musolin, D. L., Selikhovkin, A. V., Peregudova, E. Y., Popovichev, B. G., Mandelshtam, M. Y., Baranchikov, Y. N., et al. (2021). North-westward expansion of the invasive range of emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) towards the EU: From Moscow to saint Petersburg. Forests 12:502. doi: 10.3390/f12040502
Muto, G., and Koizumi, A. (2007). Mechanical properties of aodamo (Fraxinus lanuginosa), white ash (Fraxinus americana), and sugar maple (Acer saccharum) as baseball bats. Res. Bull. Hokkaido Univ. For. 64, 113–122.
Nielsen, L. R., McKinney, L. V., Hietala, A. M., and Kjær, E. D. (2017). The susceptibility of Asian, European and North American Fraxinus species to the ash dieback pathogen Hymenoscyphus fraxineus reflects their phylogenetic history. Eur. J. For. Res. 136, 59–73. doi: 10.1007/s10342-016-1009-0
Niemz, P., Clauss, S., and Michel, F. (2014). Physical and mechanical properties of common ash (Fraxinus excelsior L.). Wood Res. 59, 671–682.
Noble, R., Woodhall, J. W., Dobrovin-Pennington, A., Perkins, K., Somoza-Valdeolmillos, E., Gómez, H. L., et al. (2019). Control of Hymenoscyphus fraxineus, the causal agent of ash dieback, using composting. For. Pathol. 49:e12568. doi: 10.1111/efp.12568
Orlova-Bienkowskaja, M. J., Drogvalenko, A. N., Zabaluev, I. A., Sazhnev, A. S., Peregudova, E. Y., Mazurov, S. G., et al. (2020). Current range of Agrilus planipennis Fairmaire, an alien pest of ash trees, in European Russia and Ukraine. Ann. For. Sci. 77:29. doi: 10.1007/s13595-020-0930-z
Palik, B. J., D’Amato, A. W., Slesak, R. A., Kastendick, D., Looney, C., and Kragthorpe, J. (2021). Eighth-year survival and growth of planted replacement tree species in black ash (Fraxinus nigra) wetlands threatened by emerald ash borer in Minnesota, USA. For. Ecol. Manag. 484:118958. doi: 10.1016/j.foreco.2021.118958
Pautasso, M., Aas, G., Queloz, V., and Holdenrieder, O. (2013). European ash (Fraxinus excelsior) dieback – A conservation biology challenge. Biol. Conserv. 158, 37–49. doi: 10.1016/j.biocon.2012.08.026
Petucco, C., Lobianco, A., and Caurla, S. (2020). Economic evaluation of an invasive forest pathogen at a large scale: The case of ash dieback in France. Environ. Model. Assess. 25, 1–21. doi: 10.1007/s10666-019-09661-1
Pliûra, A., Lygis, V., Suchockas, V., and Bartkevicius, E. (2011). Performance of twenty four European Fraxinus excelsior populations in three Lithuanian progeny trials with a special emphasis on resistance to Chalara fraxinea. Balt. For. 17, 17–34.
Plumb, W. J., Coker, T. L., Stocks, J. J., Woodcock, P., Quine, C. P., Nemesio-Gorriz, M., et al. (2020). The viability of a breeding programme for ash in the British Isles in the face of ash dieback. Plants People Planet 2, 29–40. doi: 10.1002/ppp3.10060
Pratt, J. (2017). “Management and use of ash in britain from the prehistoric to the present: some implications for its preservation,” in Dieback of European Ash (Fraxinus spp.): Consequences and Guidelines for Sustainable Management, eds R. Vasaitis and R. Enderle (Uppsala: Swedish University of Agricultural Sciences), 1–14.
Queloz, V., Grünig, C. R., Berndt, R., Kowalski, T., Sieber, T. N., and Holdenrieder, O. (2011). Cryptic speciation in Hymenoscyphus albidus. For. Pathol. 41, 133–142. doi: 10.1111/j.1439-0329.2010.00645.x
Queloz, V., Hopf, S., Schoebel, C., Rigling, D., and Gross, A. (2017). “Ash dieback in Switzerland: history and scientific achievements,” in Dieback of European Ash (Fraxinus spp.) - consequences and guidelines for sustainable management, eds R. Vasaitis and R. Enderle (Uppsala: Swedish University of Agricultural Sciences), 68–78.
Rigling, D., Hilfiker, S., Schöbel, C., Meier, F., Engesser, R., Scheidegger, C., et al. (2016). Le dépérissement des pousses du frêne. Biologie, symptômes et recommandations pour la gestion. Birmensdorf: Institut fédéral de recherches WSL, 8.
Rosenvald, R., Drenkhan, R., Riit, T., and Lõhmus, A. (2015). Towards silvicultural mitigation of the European ash (Fraxinus excelsior) dieback: the importance of acclimated trees in retention forestry. Can. J. For. Res. 45, 1206–1214. doi: 10.1139/cjfr-2014-0512
Rysavy, T., and Roloff, A. (1994). Ursachen der vereschung in mischbeständen und vorschläge zu ihrer vermeidung. For. Holz 49, 392–396.
Sakio, H. (1997). Effects of natural disturbance on the regeneration of riparian forests in a Chichibu Mountains, Central Japan. Plant Ecol. 132, 181–195. doi: 10.1023/A:1009775923208
Sakio, H. (2020). “Fraxinus platypoda,” in Long-Term Ecosystem Changes in Riparian Forests, ed. H. Sakio (Singapore: Springer), 23–37. doi: 10.1007/978-981-15-3009-8_2
Sakio, H., Kubo, M., Shimano, K., and Ohno, K. (2002). Coexistence of three canopy tree species in a riparian forest in the Chichibu Mountains, central Japan. Folia Geobot. 37, 45–61. doi: 10.1007/BF02803190
Schlegel, M., Dubach, V., von Buol, L., and Sieber, T. N. (2016). Effects of endophytic fungi on the ash dieback pathogen. FEMS Microbiol. Ecol. 92:fiw142. doi: 10.1093/femsec/fiw142
Schmiedel, D., and Tackenberg, O. (2013). Hydrochory and water induced germination enhance invasion of Fraxinus pennsylvanica. For. Ecol. Manag. 304, 437–443. doi: 10.1016/j.foreco.2013.04.027
Schmiedel, D., Huth, F., and Wagner, S. (2013). Using data from seed-dispersal modelling to manage invasive tree species: The example of Fraxinus pennsylvanica Marshall in Europe. Environ. Manag. 52, 851–860. doi: 10.1007/s00267-013-0135-4
Schoebel, C. N., Zoller, S., and Rigling, D. (2014). Detection and genetic characterisation of a novel mycovirus in Hymenoscyphus fraxineus, the causal agent of ash dieback. Infect. Genet. Evol. 28, 78–86. doi: 10.1016/j.meegid.2014.09.001
Selikhovkin, A. V., Musolin, D. L., Popovichev, B. G., Merkuryev, S. A., Volkovitsh, M. G., and Vasaitis, R. (2022). Invasive populations of the emerald ash borer Agrilus planipennis Fairmaire, 1888 (Coleoptera: Buprestidae) in Saint Petersburg, Russia: A hitchhiker? Insects 13:191. doi: 10.3390/insects13020191
Shamsi, W., Kondo, H., Ulrich, S., Rigling, D., and Prospero, S. (2022). Novel RNA viruses from the native range of Hymenoscyphus fraxineus, the causal fungal agent of ash dieback. Virus Res. 320:198901. doi: 10.1016/j.virusres.2022.198901
Short, I., and Hawe, J. (2019). Ash dieback in Ireland - A review of European management options and case studies in remedial silviculture. Irish For. 75, 44–72.
Showalter, D. N., Villari, C., Herms, D. A., and Bonello, P. (2018). Drought stress increased survival and development of emerald ash borer larvae on coevolved Manchurian ash and implicates phloem-based traits in resistance. Agric. For. Entomol. 20, 170–179. doi: 10.1111/afe.12240
Simon, J., Dörken, V. M., Arnold, A., and Adamczyk, B. (2018). Environmental conditions and species identity drive metabolite levels in green leaves and leaf litter of 14 temperate woody species. Forests 9:775. doi: 10.3390/f9120775
Skovsgaard, J. P., Wilhelm, G. J., Thomsen, I. M., Metzler, B., Kirisits, T., Havrdová, L., et al. (2017). Silvicultural strategies for Fraxinus excelsior in response to dieback caused by Hymenoscyphus fraxineus. Forestry Int. J. For. Res. 90, 455–472. doi: 10.1093/forestry/cpx012
Spadaro, V., and Raimondo, F. M. (2022). The “manna” extracted from the ash trees still cultivated in Sicily from mythical food to pharmaceutical and nutraceutical resource. Arch. Food Nutr. Sci. 6, 038–048. doi: 10.29328/journal.afns.1001036
Stener, L.-G. (2018). Genetic evaluation of damage caused by ash dieback with emphasis on selection stability over time. For. Ecol. Manag. 409, 584–592. doi: 10.1016/j.foreco.2017.11.049
Thomas, P. A. (2016). Biological flora of the british isles: Fraxinus excelsior. J. Ecol. 104, 1158–1209. doi: 10.1111/1365-2745.12566
Thomasset, M., Hodkinson, T. R., Restoux, G., Frascaria-Lacoste, N., Douglas, G. C., and Fernández-Manjarrés, J. F. (2014). Thank you for not flowering: conservation genetics and gene flow analysis of native and non-native populations of Fraxinus (Oleaceae) in Ireland. Heredity 112, 596–606. doi: 10.1038/hdy.2013.141
Turczanski, K., Dyderski, M. K., and Rutkowski, P. (2021). Ash dieback, soil and deer browsing influence natural regeneration of European ash (Fraxinus excelsior L.). Sci. Total Environ. 752:141787. doi: 10.1016/j.scitotenv.2020.141787
Unger, G. M., Konrad, H., Schwanda, K., Cech, T. L., Hoch, G., Fera, A., et al. (2021). “Ash in distress: conservation and resistance breeding programme for Fraxinus excelsior in Austria,” in Perspectives for Forest and Conservation Management in Riparian Forests, eds M. Sallmannshofer, S. Schüler, and M. Westergren (Ljubljana: Slovenian Forestry Institute), 146–150.
Valenta, V., Moser, D., Kapeller, S., and Essl, F. (2017). A new forest pest in Europe: A review of Emerald ash borer (Agrilus planipennis) invasion. J. Appl. Entomol. 141, 507–526. doi: 10.1111/jen.12369
Vasaitis, R., and Enderle, R. (2017). Dieback of European ash (Fraxinus spp.) - Consequences and Guidelines for Sustainable Management. Uppsala: SLU Service/Repro.
Volkovitsh, M. G., Bieńkowski, A. O., and Orlova-Bienkowskaja, M. J. (2021). Emerald ash borer approaches the borders of the European Union and Kazakhstan and is confirmed to infest European ash. Forests 12:691. doi: 10.3390/f12060691
Wallander, E. (2008). Systematics of Fraxinus (Oleaceae) and evolution of dioecy. Plant Syst. Evol. 273, 25–49. doi: 10.1007/s00606-008-0005-3
Wang, J., Wu, L., Zhao, X., Fan, J., Zhang, C., and Gadow, K.v. (2013). Influence of ground flora on Fraxinus mandshurica seedling growth on abandoned land and beneath forest canopy. Eur. J. For. Res. 132, 313–324. doi: 10.1007/s10342-012-0676-8
Wardle, P. (1961). Biological flora of the british isles: Fraxinus Excelsior L. J. Ecol. 49, 739–751. doi: 10.2307/2257236
Zhao, Y.-J., Hosoya, T., Baral, H.-O., Hosaka, K., and Kakishima, M. (2013). Hymenoscyphus pseudoalbidus, the correct name for Lambertella albida reported from Japan. Mycotaxon 122, 25–41. doi: 10.5248/122.25
Keywords: Agrilus planipennis, ash dieback, common ash, emerald ash borer, Hymenoscyphus fraxineus, invasive species, non-native tree species
Citation: Lévesque M, Bustamante Eduardo JI and Queloz V (2023) Potential alternative tree species to Fraxinus excelsior in European forests. Front. For. Glob. Change 6:1048971. doi: 10.3389/ffgc.2023.1048971
Received: 20 September 2022; Accepted: 11 January 2023;
Published: 02 February 2023.
Edited by:
Thomas Campagnaro, University of Padua, ItalyReviewed by:
Tadeusz Kowalski, University of Agriculture in Krakow, PolandDusanka Vujanovic, BioSense Institute, Serbia
Arnaud Dowkiw, Institut National de Recherche pour l’Agriculture, l’Alimentation et l’Environnement (INRAE), France
Copyright © 2023 Lévesque, Bustamante Eduardo and Queloz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mathieu Lévesque,  mathieu.levesque@usys.ethz.ch
mathieu.levesque@usys.ethz.ch
†These authors have contributed equally to this work and share first authorship