 Imran Khan1
Imran Khan1 Din Muhammad Zahid1
Din Muhammad Zahid1 Muhammad Zubair1
Muhammad Zubair1 Syed Amir Manzoor1
Syed Amir Manzoor1 Ghulam Yasin1Mamoona Wali Mohammad2Khalid Ali Khan3,4
Ghulam Yasin1Mamoona Wali Mohammad2Khalid Ali Khan3,4 Shafeeq Ur Rahman5*
Shafeeq Ur Rahman5* Anzhen Qin4,6*
Anzhen Qin4,6*- 1Department of Forestry and Range Management, FAS & T, Bahauddin Zakariya University Multan, Multan, Pakistan
- 2Forest Education Division, Pakistan Forest Institute, Peshawar, Pakistan
- 3Unit of Bee Research and Honey Production, Research Center for Advanced Materials Science (RCAMS), King Khalid University, Abha, Saudi Arabia
- 4Applied College, King Khalid University, Abha, Saudi Arabia
- 5Water Science and Environmental Engineering Research Center, College of Chemical and Environmental Engineering, Shenzhen University, Shenzhen, China
- 6Institute of Farmland Irrigation, Chinese Academy of Agricultural Sciences, Key Laboratory of Crop Water Use and Regulation, Ministry of Agriculture and Rural Affairs, Xinxiang, China
The dry temperate forests of Pakistan exhibit rich diversity of understory floral species. These important and endemic plant species play a pivotal role in ecosystem functioning and providing livelihoods to the forest communities. South Waziristan, located in the southwest of Pakistan, is known for its rare pure stands of Pinus gerardiana Wallichex. D. Don forests. The rapidly expanding population and current state of affairs are leading to deforestation and causing disturbance to the understory vegetation of this valuable forest. The current study aims to fill the currently scanty information present in the literature regarding the diversity status of understory vegetation in P. gerardiana forests. Data of understory vegetation diversity were gathered for the northern and southern aspects of the forest. A 50 m2 sampling plot along with the placement of fifty 1x1m2 quadrats on both sites were laid out to determine species dominance, diversity, and evenness using Simpson, Shanon, Marglef, and Buzas–Gibson indices. The study depicted 612 individuals of about 29 species representing 23 families from the study site. The most abundant species in all of the sites was Salvia Glutinosa cccIV = 0.52. The southern aspect depicted the lowest diversity, but had high species dominance. Allium carolinianum DC. in this site was the most important species (IV = 0.74). The most abundant species were medicinal plants (75%), followed by edible plants (25%). Aspect had a significant effect on the understory vegetation and different categories of plants with respect to their diversity, dominance, and evenness as predicted by the various diversity indices. This study provides essential insight in to the florisitic diversity and community structure of the fragile pine nut understory vegetation in South Waziristan, which was hitherto not available. It was concluded that the southern aspect of understudy dry temperate forests depicted lower diversity and density as compared with the northern aspect. But it was observed that the southern aspect more specifically had more density and diversity of medicinal plants. It is therefore the need of the hour that conservation of these species from grazing and unsustainable harvesting must be ensured to sustain the livelihood of the inhabitants.
1. Introduction
Majestic conifers around the world mostly occupy temperate, boreal, and subtropical climates (1). Global populations of these long-lived species are facing serious threats to their existence mostly due to anthropogenic activities and climate change (2). Among these conifers, Pinus geradiana Wallichex. D. Don forests are extremely important. They are native to the Western Himalayas and are distributed in eastern Afghanistan, southern Xizang (Tibet) in China, Jammu and Kashmir in India, and northern Pakistan (Shalizi et al., 2018; Urooj, 2019). These forests are extremely valuable, having key socioeconomic importance. However, rapid exploitation of these forests is gradually leading them toward extinction from their original habitat (Urooj and Jabeen, 2015; Aziz et al., 2018; Shalizi et al., 2018). Globally, P. gerardina forests are native to South East Asia, West Asia, and in the states of Central Asia (Shalizi and Khurram, 2016; Urooj, 2019). Pakistan is home to one of the largest P. gerardiana forest stands in the world (Print et al., 2015; Urooj and Jabeen, 2015; Urooj et al., 2020). These forests are extensively located in the Sulieman and Hindu Kush mountain ranges in the west (Khyber Pakhtun Khwa and Baluchistan) and north (Kashmir and Gilgit Baltistan) of the country (Print et al., 2015; Urooj and Jabeen, 2015; Urooj, 2019). P. gerardiana forests are major economic players in the region, generating a hefty revenue of 13 million USD annually (Haq et al., 2013; Print et al., 2015; Urooj, 2019). These forests are responsible for sustaining rural livelihoods in the form of employment and businesses (Shalizi et al., 2018; Alami and Mousavi, 2020; Rahimzadeh, 2020; Urooj et al., 2020; Alami and Mousavi, 2021). Forest understory vegetation is regarded as an ecological indicator due to its quick response to changing environmental conditions (Von Oheimb and Härdtle, 2009). As such, understory vegetation can serve as an important indicator of species composition in a forest. The understory layer is known to affect the establishment, survival rate, and adaptability of plant species, including soil physicochemical properties such as nutrient content, pH, and moisture (Dzwonko and Gawroński, 2002; Tinya et al., 2009). Understory medicinal plant species are an important component of forest communities’ livelihood, playing a vital role in providing subsistence and cash income to a large part of the world’s population, particularly in developing countries (Sher et al., 2014; Muhammad et al., 2019).
Studies have depicted the importance of understory species among the people living in the forest vicinity in the north of Pakistan (Kayani et al., 2014; Sher et al., 2014, 2016). Muhammad et al. (2019) displayed the inventory of medicinal plants in the dry and moist temperate forests of Kashmir. The authors depicted a large dependence of local communities upon these medicinal plants. Moreover, majority of people in these communities were dependent upon medicinal plant collection, processing, and marketing for sustaining their livelihood (Zubair et al., 2019). Understory species are affected by structural changes in the forests resulting from anthropogenic disturbances and overexploitation (Vellend et al., 2006; Liira et al., 2007; Wyatt and Silman, 2010). Such changes in forest structure may even lead to the local extinction of certain species (Vellend et al., 2006).
Slope aspect remains pivotal in affecting plant communities in terms of their characteristics, functional groups, and aspect orientation (Zhang et al., 2022). Slope aspect is instrumental in changing solar radiation distribution patterns, thereby altering soil temperature and moisture levels (Burnett et al., 2008). Thus, slope aspect affects almost all vegetation properties, importantly, species composition, photosynthetic efficiency, species diversity, cover, nutritional dynamics, and, in turn, productivity (Kakembo et al., 2007; Ortiz-Pulido and Pavon, 2010; Sharm et al., 2010; Aimme and Normaniza, 2015).
In Pakistan, the dependence of people on pine forest resources has continued to rise in the last few decades. This is mostly due to the increasing population inhabiting previously untouched forest ecosystems (Ali et al., 2005). Pakistan, with 260 km2 of P. gerardina forest, experiences unsustainable overexploitation of resources. This not only leads to decreased production but aids deforestation. In these mountain ranges, most of the P. gerardiana forests are exploited by local people for pine nut collection and medicinal plant collection (Alami and Mousavi, 2020). Non-timber forest products (NTFPs) collected from the understory of these forests play a significant role in the socioeconomics of the forest community. But unsustainable and continuous resource exploitation coupled with deforestation mostly leading to settlements for new communities are having adverse effects on the understory vegetation that has been poorly explored by the scientific community of this region (Vellend et al., 2006; Urooj and Jabeen, 2015; Aziz et al., 2018). Hence, there is need of a study to measure the resources in the understory layer of these forests and to observe their relationship with changing land-use patterns (Hayat et al., 2021). The current study will provide crucial knowledge regarding the suitability of the ecosystem, especially in terms of habitat management.
There are already extensive studies reporting the benefits and importance of pine nut forests regarding its top story. But there are very few studies exploring the ecological, ethnological, and economic importance of the understory of Pinus spp. forests (López-Marcos et al., 2019, 2020). It is extremely crucial that the understory vegetation of pine forests are studied, and in particular P. gerardina. This would help in effective forest management and resource sustainability, thus ensuring the stable livelihood of the inhabitants dependent upon them (Mölder et al., 2008; Muhammad Azam Khan et al., 2010; Haq et al., 2011). Researching the understory vegetation of these forests can help in determining the vital role that it plays in nutrient cycling and carbon sequestration, thus influencing the overall ecosystem’s health and resilience (Ou et al., 2020; Ali and Khan, 2022). More importantly studying the diversity and distribution of understory vegetation can help in identifying invasive species, promoting biodiversity conservation, and mitigating the risk of pest and disease outbreaks (Ullah et al., 2013; Zilliox and Gosselin, 2014).
In this study, we aim to explore the understory vegetation of P. gerardiana forests in South Waziristan Agency under different aspects (northern and southern). There are limited cross-sectional studies and systematic documentation highlighting the understory vegetation density concerning various aspects and the use of different plant species in Pakistan (Liira et al., 2007; Wyatt and Silman, 2010; Ali and Khan, 2022). The present study introduces an avenue of knowledge about the role of different aspects influencing the distribution, abundance of understory vegetation, and its diversity for this precious forest resource. Since aspects were the focus of study across which different vegetation parameters were assessed, therefore, the core objectives of the study was to assess species diversity across different aspects (northern and southern) of the understory vegetation in P. gerardiana stands in the dry temperate forest of South Waziristan, Khyber Pakhtunkhwa (KPK), Pakistan.
2. Materials and methods
2.1. Description of the study area
South Waziristan extends over an area of 6,500 km2, located about 580 km northeast of Islamabad, Pakistan, and shares a 300 km border with Afghanistan. The P. gerardiana forest is located near the Pakistan–Afghanistan border (Ullah et al., 2013). The study area is situated at 32031.5810’ N 69023.6470′ E with an altitude of 1,250–2,134 m above sea level (Figure 1). The Suleiman Mountain range consists of dry temperate forest with an area of 260 km2. The vegetation diversity is composed of dry temperate forest, dominated by P. gerardiana, Cedrus deodara (Lamb.) G.Don., Abies pindrow (Royle ex D. Don.), and Quercus incana Roxb. Some of the lower areas have the Juglans regia L. species. Among the shrub species are Alhagi maurorum Medic Medik. Berberis lycium Royle, and Viburnum grandiflorum f. foetens, while Salvia Glutinosa L, Salvia moocroftiana Wall. ex Benth., Senecio flaevus (Herb), Artemisia scoparia Maxim., Convolvulus arvensis L., Urtica dioica L., Peganum harmala L., Portulaca oleracea L., and Amaranthus viridis L. are common herb species in the study area (Richardson and Rundel, 1998; Arnold and Perez, 2001).

Figure 1. Study Site: Pinus gerardiana Forest Reserve, South Waziristan Agency, KPK.
The area receives an average annual precipitation of about 152.4 mm. Winters are usually characterized by heavy snow, whereas summers are hot with scanty rainfall (Ullah et al., 2013; Aziz et al., 2018). Summers are short, of about four months, whereas winters are long, starting from October up to the end of May. The short interval of summers provides a short time for the growth and regeneration of trees and understory plants, herbs, and shrubs. The average altitude of the P. gerardiana forest is 2,473.47 m according to the data collected through GPS from the study site. We conducted the data collection in the summer season to get easy access to the high altitudes of the forests and document the vegetation in the study site. In a similar study, the authors indicated that most of the population in the nearby settlements of the forest were dependent upon the P. gerardiana forest for nutritional and economic needs (Ullah et al., 2013). People living near or near pine nut forests are extremely dependent upon pine nut collection (Richardson and Rundel, 1998).
2.2. Sampling design and instruments
We employed the plot sampling method in the current study for the vegetation inventory. This technique considers an efficient and normalized way to deal with already gathered information on plant species, density diversity, and other vegetation qualities in a particular region or territory (Greig-Smith, 1961). Plots of a certain size were formulated throughout the sites and quadrats were placed randomly for collecting the vegetation data. A total of 12 sampling plots were established for biodiversity assessment of the research study site. Six random plots were formed at each site on both the aspects. These study sites were divided based on the two different aspects: the southern aspect at an altitude of 2,437 m and the northern aspect at 2,435 m. The instruments used for the field data collection were a 1 × 1 m quadrat, measuring tape, GPS, clippers, weight balance, pegs along with field range vegetation forms and a map of the study area, paper bags, pencil, sharpeners, and erasers.
2.3. Data collection
For each plot, 10 quadrats of 1 m2 were randomly placed for vegetation assessment during August and September 2021 (northern and southern aspects). The quadrats were placed randomly across the six plots selected per site. A total of 120 quadrats were inventoried. Species within the quadrats were quantified for diversity data. The nomenclature of the species as proposed by Peruzi et al. was followed (Peruzzi, 2020). Vegetation was analyzed quantitatively for density (D) and frequency (F). Moreover, the Global Biodiversity Information Facility (gbif.org) and the Angiosperm Phylogeny Group III classifications were used for the identification of plant species nomenclature and family names (The Angiosperm Phylogeny Group, 2009).
2.4. Data analysis
The data were subjected to biodiversity analysis. Some of the basic quantitative analysis including vegetation frequency, density, and richness was estimated using Curtis and McIntosh (1950); the relative values for species frequency (RF), density (RD), cover percent (CP), and important value index (IVI) were also estimated and presented. Species distribution and abundance was calculated following Wiegert (1989), and the important value (IV) index was used for the determination of species dominance (Wang and Shangguan, 2010). The species richness, dominance, evenness, and heterogeneity were determined through various indices including Simpson (1949), Margalef (1958), Buzas and Gibson (2010), and Shannon (1949) index calculations at plot level. The Sorensen–Dice index (Sorensen, 1948) was used for the calculation of similarities in various plant species composition among the different sampling sites. Furthermore, the species richness and evenness were visualized through plotting the rank abundance curve. Rank abundance was plotted to determine the accumulation of the species in the study area (Baker et al., 2006; Society and Press, 2016). Sample efficacy, species richness in a specific habitat in comparison with species richness in numerous habitats, was determined by plotting an equal basis rarefaction curve (Hall and Bawa, 2012; Hurlbert, 2012).
3. Results
3.1. Global vegetation description
The research site was composed of pure stands of P. gerardiana. A total of 612 plant specimens representing 29 species from 23 families were identified in the P. gerardiana forest understory (Tables 1, 2). Regarding the composition of species, about 70% of the plants were identified as herbs. The shrubs found on the study site accounted for 20%, whereas fern species accounted for about 10%. The survey was conducted on two different aspects, the southern (southwest and southeast) and northern (northwest and northeast) aspects. The southern aspect exhibited slightly higher species richness, with 15 observed species, compared with the northern aspect, which had 14 observed species (Table 1). On the northern aspect, the most abundant plant species identified was Allium carolinianum DC., while on the southern aspect, Salvia nubicola (Darshel) emerged as the dominant species.
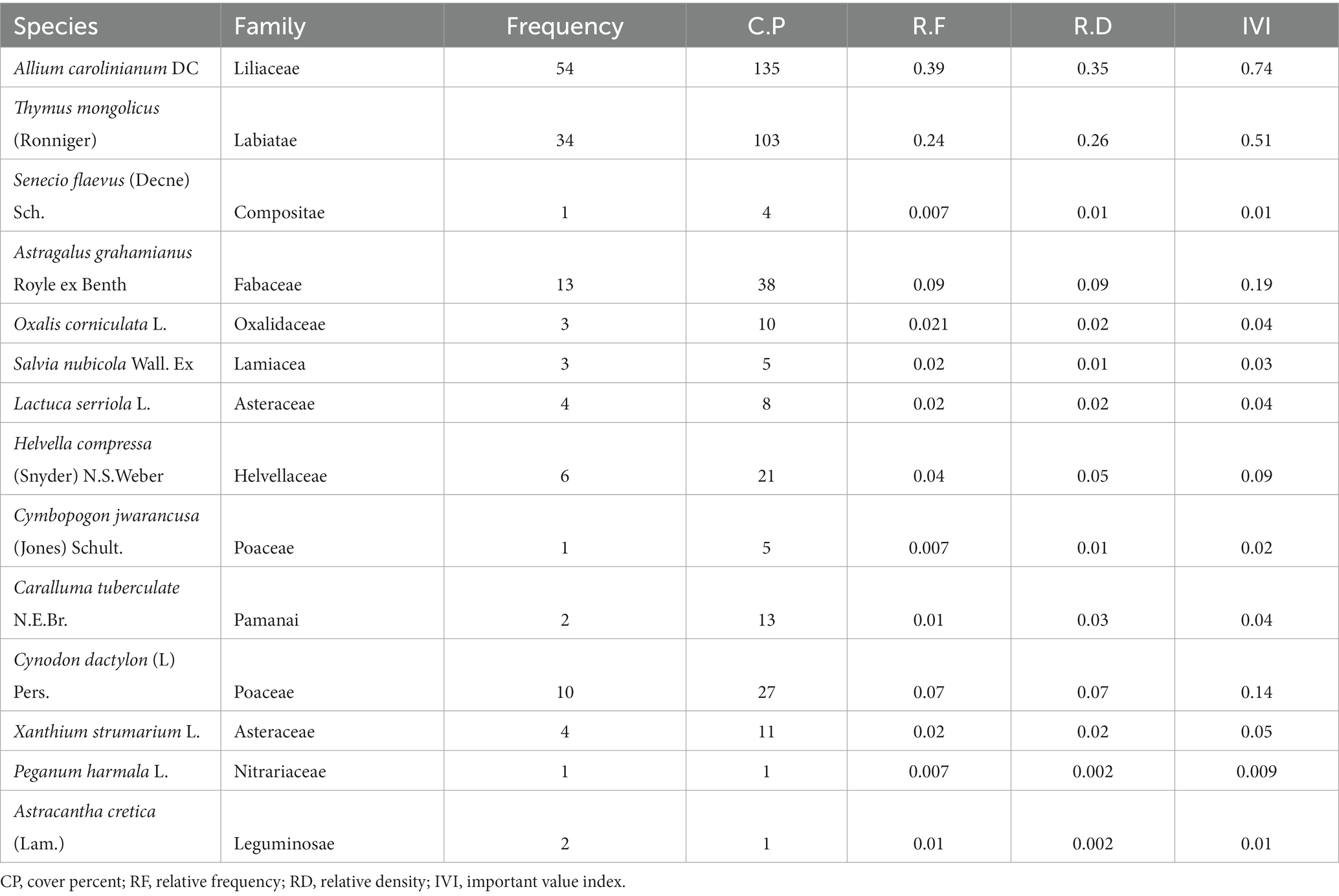
Table 1. Species importance value index for the understorey vegetation of Pinus gerardiana forest on the northern aspect.
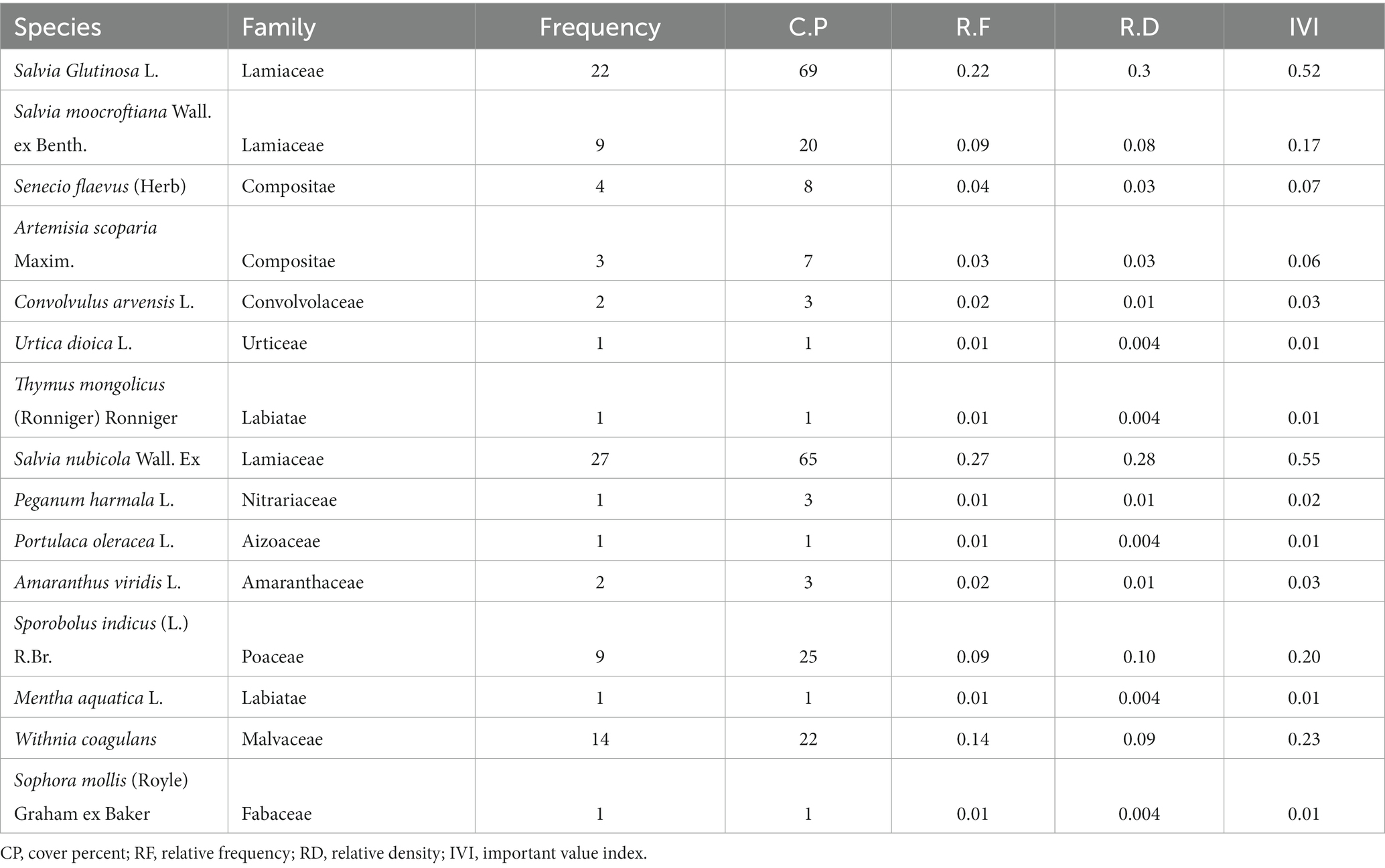
Table 2. Species importance value index for understorey vegetation of P. gerardiana forest on the southern aspect.
When assessing the importance value (IV) index of species, variations between the northern and southern aspects were observed. The highest IV value of 0.74 was recorded for A. carolinianum DC. on the northern aspect, while S. glutinosa L, Jupiters Distaff (Rashal), had the highest IV value of 0.5 on the southern aspect. The majority of the species fell within the range of 0.02–0.06 IV. Notably, three species, namely, A. carolinianum, T. mongolicus, S. glutinosa, and S. nubicola showed high IV values at both sites (Tables 1, 2). Overall, the data indicate differences in species abundance and IV values between the northern and southern aspects, highlighting the specific characteristics and composition of each site. The study showed a higher presence of medicinal plants on the southern aspect (80%) as compared with the northern aspect (70%).
3.2. Vegetation composition on the northern aspect
The values of frequency (F) and density (D) calculated among the various aspects showed different results. During the sampling in the northern and southern aspects, the most dominant species found was A. carolinianum (RF = 39%; RD = 135) followed by T. mongolicus (F = 25%; D = 103) present on the northern aspect, wherein 70% of plant species were classified as highly important medicinal plant species. The second most important plant present on the northern aspect was T. mongolicus (RF = 25%; RD = 103). The northern aspect also contained significant numbers of A. grahamianus (RF = 9%; RD = 38). However, P. harmala L (RF = 0.26%; RD = 1) was also found in the area, being an endangered and endemic medicinal plant. The most important and widely used species was T.mongolicus (F = 27%; D = 103) and P. harmala (RF = 0.72%; RD = 1). Few plants were normally used by the livestock. These plants include C. dactylon (RF = 7.2%; RD = 27), C. tuberculata, (RF = 1.44%; RD = 13), and X. strumarium (RF = 3%; RD = 11). The remaining species, i.e., S. flaevus (Decne) Sch. (RF = 0.72% RD = 18) was noted in few numbers on the northern aspect.
3.3. Vegetation composition on the southern aspect
The frequency and density of vegetation on the southern aspect were higher when compared with the northern aspect. The southern aspect represented species mostly utilized by local communities for medicinal purposes and livestock grazing. The most abundant of them were S. nubicola Wall. Ex. (RF = 27.5%; RD = 65), S. glutinosa L (RF = 22%; RD = 69), and S. moocroftiana Wall. ex Benth. (RF = 9.4%; RD = 20). Plants with medicinal characteristics (64%) also existed on the southern aspect; the most important and widely used species were S. glutinosa L (RF = 27.5%; D = 65), S. nubicola (RF = 27.5% RD = 65), W. coagulans (RF = 14.5%; RD = 22), and S. moocroftiana Wall. ex Benth. (RF = 9.4%; RD = 20), while other plant species were quite rare, with minimum frequencies and densities. Among all the species on the southern aspect 13% were used as fodder for livestock whereas the remaining (86%) species including herbs and shrubs were used for medicinal purposes. The total understory vegetation on the northern and southern aspects was characterized as 70% medicinal and 25% edible plants. The southern aspect consisted of the bulk of medicinal plant species. Nonetheless, edible plants were not very dominant on the southern aspect, and hence were sparsely scattered on the sampled sites of the southern aspect (southwest and southeast). Among all these species, S. glutinosa L. is also used for making flavoring (RF = 22% RD = 69) and had considerable representation.
3.4. Rank abundance curve and rarefaction analyses
Figure 2 represents the abundance rank and relative abundance measured on a log scale. The northern aspect was dominated by 14 species contributing to the bulk of the biomass. The curves of the southern and northern aspects differ in their slopes, showing good evenness of the species at the southern site. The northern aspect showed more species abundance than the southern aspect. The species on the northern aspect were quite diverse as compared with the southern aspect. A rarefaction curve depicts how many samples are needed for analyzing population vegetation diversity (Figure 3). The northern aspect curve reaches asymptote after a specific point steeply going in upward direction showing prominent effect. Conversely, the southern aspect curve of the area where sampling was done was full of unique species used for medicinal purposes and it was found that the site could contain more species if further sampling had been conducted.
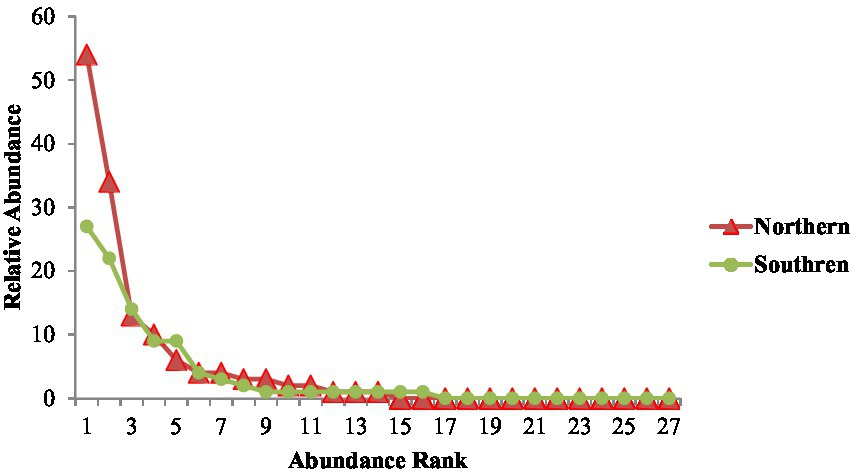
Figure 2. Species wise rank abundance curve of the vegetation on the northern and southern aspects of P. gerardiana forest.
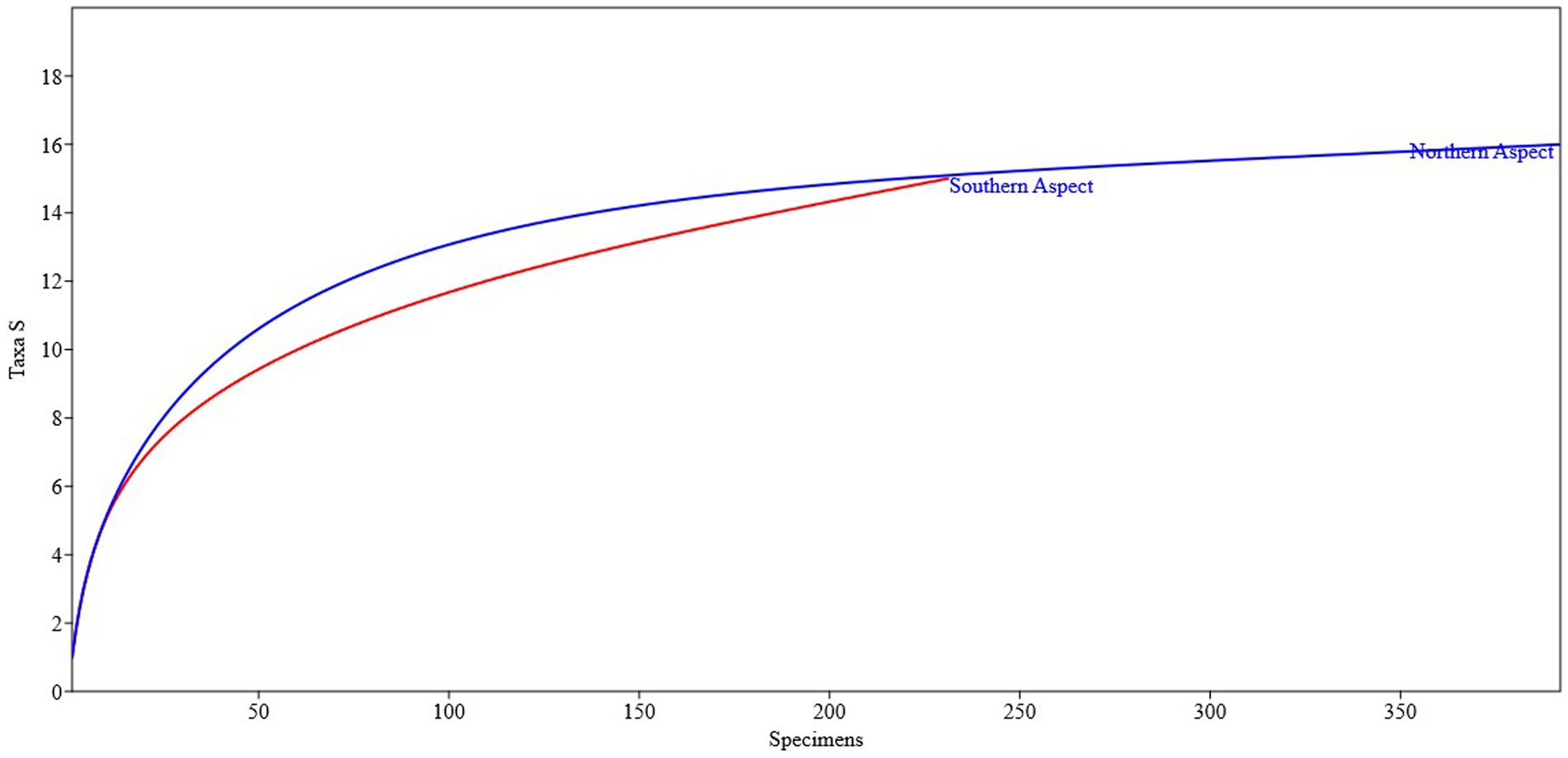
Figure 3. Plot-based rarefaction curves for the northern and southern aspects of P. gerardiana.
3.5. Biodiversity analysis
It was observed that the Shannon diversity index was at the maximum on the northern aspect of the forest (Table 3). This greater value illustrates the fact that the plant communities were more diverse, integrating more richness and evenness, whereas the lowest value of the Shannon index was on the southern aspect of the study site. Comparatively, this site was low in diversity and the number of species as compared to northern aspect. To determine the abundance of the species and its dominance in the aspect, Simpson’s index was calculated from both the aspects in the study site.
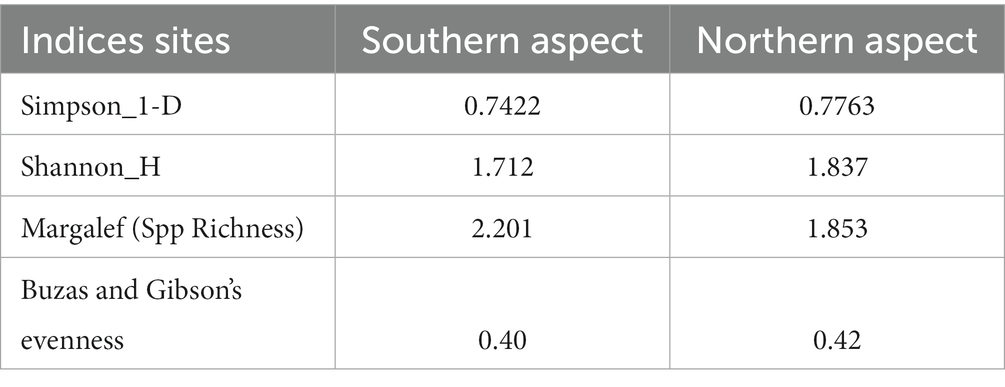
Table 3. Comparison of diversity indices on the northern and southern aspects of P. gerardiana.
The result presented that the southern aspect was highly dominated by understory vegetation. Moreover, the site also showed dominancy of fewer plants due to the sparse vegetation. For observing the species richness and evenness, the Margalef’s and Buza’s and Gibson’s diversity indices showed similar results when compared with the other indices used during findings. The main outcome of the findings depicted in those different indices values and aspects was the high biodiversity on the northern aspect of the sampling site. The Sorensen-Dice index presented least similarity in the northern aspect and southern aspect in the study site (Table 4). The decrease in similarity inferred that there were less species shared among the different aspects of the sampling sites. The index describes that the species prevalence in both the sites were rare.
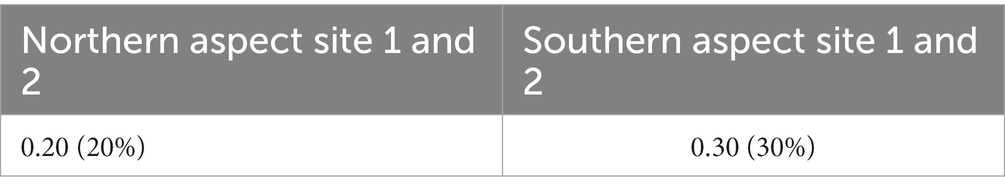
Table 4. Sørensen–Dice coefficients showing similarities between the northern and southern aspects of P. gerardiana forest.
3.6. Principal component analysis
Principal component analysis (PCA) was conducted to analyze the effect of aspect on the types of plant diversity and plant density. The results showed PC1 (Principal component 1) to account for 100% of the total variance. Density showed the highest loading in the first component. The experimental sites were separated along principal component 1 (PC1) where the southern aspect had higher PC1 scores, while the northern aspect displayed a lower density (Figure 4).
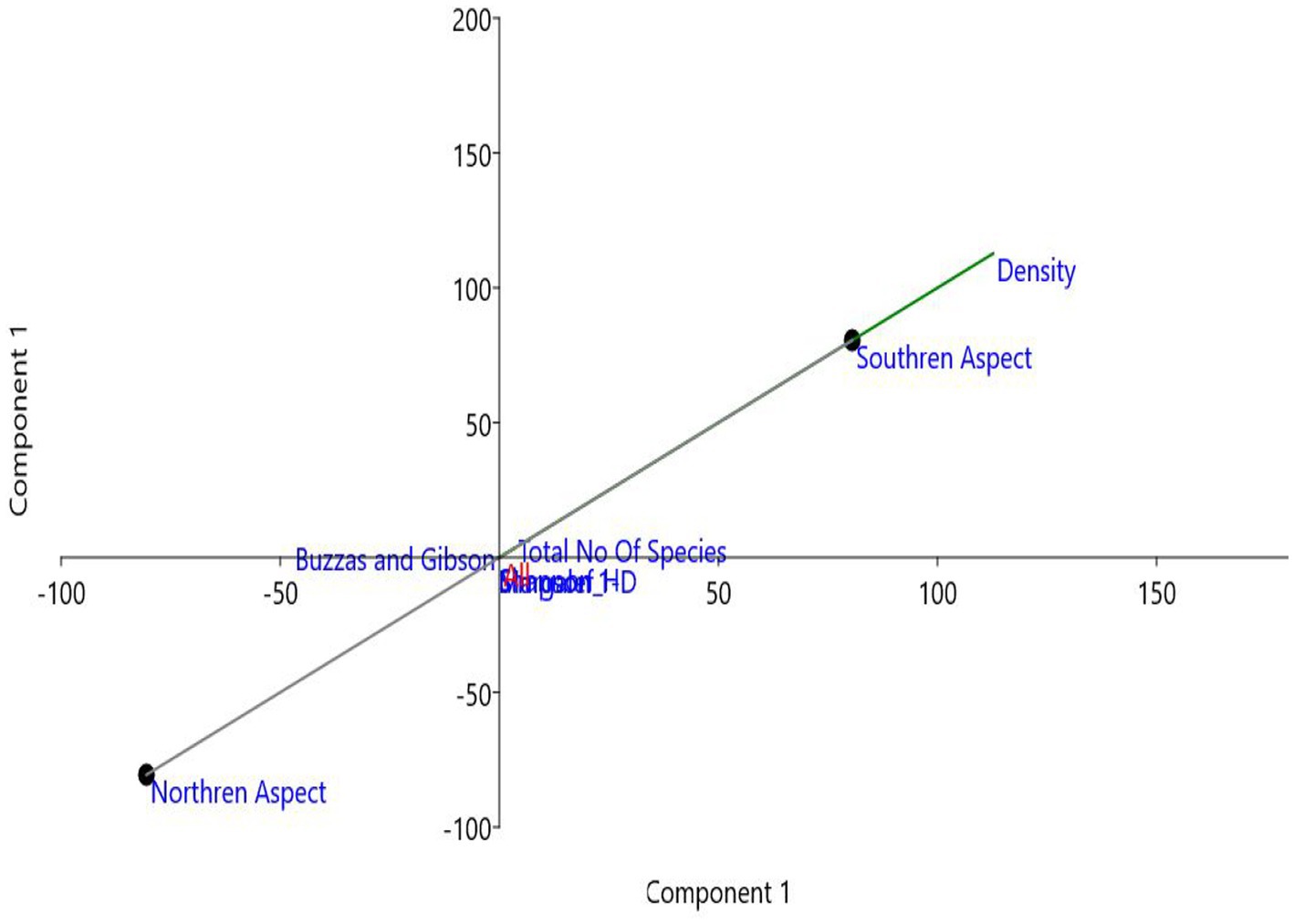
Figure 4. Principal component analysis (PCA) on the basis of the northern and southern aspects of P. gerardiana forest.
4. Discussion
The results showed the understory portion of the forest to have considerable biodiversity. Our research study depicted that the P. gerardiana forest of South Waziristan Agency has diverse understory vegetation. The understory is enriched with different medicinal plants that are used for various purposes including food and fodder. These plants play a key role in the livelihood of the local communities using them in herbal medicines as well as the functioning of the forest (Chidumayo and Gumbo, 2010). Most of the plants were used for medicinal need. The current research study was similar to the research work of Qasim et al. (2013) depicting that most of the plants in the Himalayan range were being used for medicinal purposes. Vegetation density came out to be moderate in the understudied area. Qasim et al. (2013) observed similar results depicting low species diversity in P. gerardiana stands. Results depicted homogenous variance in understory species composition on the northern and southern aspects of the study sites. Similarly, various researchers have observed variations in understory composition and structure according to the aspects (Mishra et al., 2004; Mölder et al., 2008).
The maximum biodiversity was found on the southern aspect of the study site. The area was open in canopy cover which provided a suitable environment for different medicinal plants (Qasim et al., 2013; Khel et al., 2022). Similar observations were found by Adnan and Hölscher (2012) at Ayubia National Park of Khyber Pakhtoon Khwa, where they perceived that the decrease in understory plants, herbs, and shrubs was due to the dense canopy cover of overstorey vegetation. Zilliox and Gosselin (2014) concluded that northern aspects showed minimum abundance of understory vegetation because of the closed canopy and less availability of sunlight, moisture, and nutrients for the understory plants and grasses. Similar observations were found by Whitmore (1978), wherein forest cover remained instrumental in affecting the vegetation underneath it. Similar outcomes were perceived by Sagar et al. (2008), in which they exhibited that the areas with sparse canopy cover had high understory plant cover. It was observed that the evergreen forest closed canopy had less understory biodiversity. The study noted that understory vegetation is completely dependent on the overstory of the forest trees including pure and mixed forests.
Aspect and canopy affected the diversity, density, and cover of understory vegetation of the study site, as endorsed in various similar studies (Khan et al., 2012). Overall diversity of the understory vegetation in the study area was on the lower side. This is coherent with similar studies conducted in these areas (Khan et al., 2011; Zubair et al., 2021; Ali et al., 2022). Studies in the pine nut forests of Pakistan and Afghanistan have usually depicted lower understory vegetation diversity (Khan et al., 2011; Rahman et al., 2017; Luintel et al., 2018; Ali et al., 2022; Prakash et al., 2023). However, the pine nut forests of China, India, and other regions have shown higher diversity of understory plant species compared with understory vegetation diversity of pine forests of the area under study (Huo et al., 2014; Su and Lu, 2020; Aqeel et al., 2023; Prakash et al., 2023). The study depicted a lower density and frequency of plant species in both study areas. This is mostly related to the vegetation damaging effects caused by overgrazing, unsustainable medicinal/edible plant collection, and deforestation, as reported in various studies (Shinwari and Qaiser, 2011; Shalizi et al., 2018; Hussein et al., 2021; Khan et al., 2021).
The majority of these plants are available year round and the local communities use them for their subsistence (Farooq et al., 2012; ANG et al., 2014; Aziz et al., 2016). The higher composition of species diversity and dominance on the southern aspect compared with the northern aspect was shown by the rank abundance curve highlighting the richness of S. glutinosa and S. nubicola species. Similar results were observed by Ali et al. (2007), which depicted that there was a slight decrease in the species abundance of northern aspects. Promis et al. (2012) observed similar results, stating that the areas with dense vegetation showed less understory plants. The aspects with a decrease in grazing patterns and partially closed forest cover (southern aspects) had high understory vegetation including medicinal plants. Similar results were observed by Zubair et al. (2021), stating that the spots with medium canopy cover resulted in high understory vegetation. The research study results were obtained by appropriate samples collection as justified in the rarefaction curves. The curves were used in data to overcome difficulties and interference often experienced during counting and relating to taxon richness. (Gotelli and Colwell, 2001). The availability of plants across time is a critical factor for observing richness and abundance (Wulf and Naaf, 2009; Mestre et al., 2017). Moreover, plants grown under conditions of ample light and space are more abundant compared with plants dispersed under dense forests (Walker et al., 1999). Likewise, species tend to be more diverse and richer in their flora when the available forest condition is either termed as open canopy or undisturbed partial canopy cover(Hamilton, 2004; Promis et al., 2012).
The P. gerardiana forests of Pakistan are rapidly degrading due to the overexploitation of pine nuts, unsustainable collection of medicinal/edible plant species, and continuous deforestation (Urooj et al., 2020; Khan et al., 2021). These factors not only alter the structure and functioning of the forest but also increase soil erosion events and reduce nutrients availability (Mölder et al., 2008; Qasim et al., 2013; Hussein et al., 2021). Our study depicts the lower density, cover, and diversity of important medicinal/edible plant species on the northern and southern aspects. This indicates a crucial restoration hotspot that harbors endangered species whose recovery could benefit from effective forest landscape rehabilitation and conservation practices (Höhl et al., 2020; De Matos et al., 2021).
Many previous studies have indicated direct links between diversity, composition, and the structure of understory vegetation on the functioning and provisioning of the forest (Ou et al., 2020). The quantitative inventory of the understory vegetation clearly translates as a warning of potential irreversible degradation (Xie et al., 2021). Building on the results of such vegetation surveys, it is important that we engage government departments and non-governmental organizations to take part in conservation practices utilizing wider and systematic formal monitoring activities, since forest communities have minimum input in the management and conservation of forest resources (Tabassum, 2020). This emphasized the need to understand policy and practice of forest landscape restoration (FLR) in meeting local needs and livelihoods for sustainable management of forest resources, though important, yet is important (Mansourian et al., 2020; Tabassum, 2020). Various global studies have indicated that different resource management endeavors with minimum integration of vegetation surveys in terms of monitoring usually lead to poor management and low rehabilitation success (Chazdon, 2019; Mansourian et al., 2020). Through this study, there is an opportunity for considering the incorporation of vegetation surveys with respect to aspects and canopy cover to measure the diversity of understory vegetation for better management and conservation of forest ecosystems.
5. Conclusion and policy implication
The research conducted in the P. gerardiana forest of South Waziristan Agency, Pakistan, has illuminated the intricate vegetation dynamics influenced by aspectual variations. Our findings underscore that the understory vegetation is rich in biodiversity, with a marked representation of both edible and medicinal plants. While both the northern and southern aspects of the study site showcased diverse species, the southern aspect stood out with a higher prevalence of medicinal plants. Notably, the vegetation diversity was found to be marginally higher on the southern aspect when compared with the northern aspect. This is particularly significant as it hints at the influence of aspect on species richness and abundance. For instance, species such as A. carolinianum DC. and S. nubicola (Darshel) emerged as dominant on the northern and southern aspects, respectively. Furthermore, the biodiversity indices, including the Shannon diversity index and Simpson’s index, provided a deeper understanding of species richness and evenness across the two aspects. Such differences in vegetation dynamics between the northern and southern aspects could be attributed to various ecological factors, which may include light availability, soil properties, moisture content, and other microclimatic conditions. It is also worth noting that the region, though rich in biodiversity, is vulnerable to anthropogenic pressures, such as livestock grazing. The southern aspect, in particular, displayed signs of being more affected by these practices. Sustainable management practices and conservation efforts are paramount to ensure the protection and preservation of these ecologically valuable landscapes. In light of our findings, it becomes evident that understanding the role of aspect in vegetation dynamics can provide valuable insights for biodiversity conservation and sustainable forest management. Further studies across different forest reserves in Pakistan would amplify our knowledge about the intricate relationships between aspect and vegetation, aiding in informed conservation strategies for the country’s rich biodiversity.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
IK and DZ conceptualized and wrote the main manuscript. MZ, GY, and SM helped to perform the lab analysis. MM, MZ, GY, and SM reviewed and edited the manuscript in the present form. KAK provided resources and funds. SR and AQ supervised and provided funding and approved the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the scientific and Technological Project of Henan Province (222102110175) and scientific and Technological Project of Xinxiang City (GG2021024). The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University Saudi Arabia for funding this work through Large Groups Project under grant number RGP2/447/44. The authors acknowledge the support of the Unit Bee Research and Honey Production at King Khalid University, Abha, Saudi Arabia.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adnan, M., and Hölscher, D. (2012). Diversity of medicinal plants among different Forest-use types of the Pakistani Himalaya. Econ. Bot. 66, 344–356. doi: 10.1007/s12231-012-9213-4
Aimme, H. N., and Normaniza, O. (2015). The effects of plant density of Melastoma malabathricum on the erosion rate of slope soil at different slope orientations. Int. J. Sediment. Res. 30, 131–141. doi: 10.1016/j.ijsrc.2015.03.003
Alami, K., and Mousavi, S. Y. (2020). Afghan Chehelghoza (Pinus gerardiana L.) pine nut diet enhances the learning and memory in male rats. Nutr. Diet. Suppl. 12, 277–288. doi: 10.2147/nds.s278350
Alami, K., and Mousavi, S. (2021). Effect of afghan chehelghoza (Pinus gerardiana L.) on food consumption, body, and brain weight of male rats. J. Complement. Med. Res. 12:5. doi: 10.5455/jcmr.2021.12.01.02
Ali, J., Benjaminsen, T. A., Hammad, A. A., and Dick, Ø. B. (2005). The road to deforestation: an assessment of forest loss and its causes in Basho Valley, northern Pakistan. Glob. Environ. Chang. 15, 370–380. doi: 10.1016/j.gloenvcha.2005.06.004
Ali, M. S., Ibrahim, S. A., Ahmed, S., and Lobkovsky, E. (2007). Guaiane sesquiterpene lactones from Salvia nubicola (Lamiaceae). Chem. Biodivers. 4, 98–104. doi: 10.1002/cbdv.200790011
Ali, F., and Khan, N. (2022). Do environmental variables and overstory communities affect the spatial pattern of understory vegetation? Lessons from Monotheca buxifolia (Falc.) a. DC. Forests in Pakistan. Acta Bot. Bras. 36, 41–48.
Ali, F., Khan, N., Abd_Allah, E. F., and Ahmad, A. (2022). Species diversity, growing stock variables and carbon mitigation potential in the Phytocoenosis of Monotheca buxifolia forests along altitudinal gradient across Pakistan. Appl. Sci. 12:1292. doi: 10.3390/app12031292
Ang, H. Y., Subramani, T., Yeap, S. K., Omar, A. R., Ho, W. Y., Abdullah, M. P., et al. (2014). Immunomodulatory effects of Potentilla indica and Dendrophthoe pentandra on mice splenocytes and thymocytes. Exp. Ther. Med. 7, 1733–1737. doi: 10.3892/etm.2014.1657
Aqeel, M., Ran, J., Hu, W., Irshad, M. K., Dong, L., Akram, M. A., et al. (2023). Plant-soil-microbe interactions in maintaining ecosystem stability and coordinated turnover under changing environmental conditions. Chemosphere 318:137924. doi: 10.1016/j.chemosphere.2023.137924
Arnold, J. E. M., and Pérez, M. R. (2001). Can non-timber forest products match tropical forest conservation and development objectives? Ecol. Econ. 39, 437–447. doi: 10.1016/S0921-8009(01)00236-1
Aziz, M. A., Adnan, M., Khan, A. H., Rehman, A. U., Jan, R., and Khan, J. (2016). Ethno-medicinal survey of important plants practiced by indigenous community at Ladha subdivision, South Waziristan agency, Pakistan. J. Ethnobiol. Ethnomed. 12:53. doi: 10.1186/s13002-016-0126-7
Aziz, M. A., Adnan, M., Khan, A. H., Abdella, K. A., Alqarawi, E. F., Abd-Allah, M. N. O., et al. (2018). Innovative approach for the management of medicinal plants: a case study of plant pine nuts (Pinus gerardiana wall.). Pak. J. Bot. 50, 1637–1644.
Baker, J. D., Harting, A. L., and Johanos, T. C. (2006). Use of discovery curves to assess abundance of Hawaiian monk seals. Mar. Mamm. Sci. 22, 847–861. doi: 10.1111/j.1748-7692.2006.00061.x
Burnett, B. N., Meyer, G. A., and McFadden, L. D. (2008). Aspect related microclimatic influences on slope forms and processes, northeastern Arizona. J.GeoPhysical Res: Earth Surface 113, 1–18. doi: 10.1029/2007JFO00789
Buzas, M. A., and Gibson, T. G. (2010). Species diversity: benthonic foraminifera in Western North Atlantic. American Assoc. Advan. Sci. 163, 72–75.
Chazdon, R. L. (2019). Towards more effective integration of tropical forest restoration and conservation. Biotropica 51, 463–472. doi: 10.1111/btp.12678
Chidumayo, E. N., and Gumbo, D. J.. The dry forests and woodlands of Africa: Managing for products and services. (2010). England: Routledge.
Curtis, J., and McIntosh, R. (1950). The interrelations of certain analytic and synthetic phytosociological characters. Ecology 31, 434–455. doi: 10.2307/1931497
De Matos, T. P. V., De Matos, V. P. V., De Mello, K., and Valente, R. A. (2021). Protected areas and forest fragmentation: sustainability index for prioritizing fragments for landscape restoration. Geol. Ecol. Landscapes 5, 19–31. doi: 10.1080/24749508.2019.1696266
Dzwonko, Z., and Gawroński, S. (2002). Effect of litter removal on species richness and acidification of a mixed oak-pine woodland. Biol. Conserv. 106, 389–398. doi: 10.1016/S0006-3207(01)00266-X
Farooq, S., Barki, A., Khan, M. Y., and Fazal, H. (2012). Ethnobotanical studies of the flora of tehsil Birmal in South Waziristan agency, Pakistan. Pakistan J. Weed Sci. Res. 18, 277–291.
Gotelli, N. J., and Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 4, 379–391. doi: 10.1046/j.1461-0248.2001.00230.x
Greig-Smith, P. (1961). The vegetation of Wisconsin. An ordination of plant communities. JSTOR 49:463. doi: 10.2307/2257277
Hamilton, A. C. (2004). No TitleMedicinal plants, conservation and livelihoods. Biodivers. Conserv. 13, 1477–1517. doi: 10.1023/B:BIOC.0000021333.23413.42
Hall, P., and Bawa, K. (2012). Methods to assess the impact of extraction of non-timber tropical forest products on plant populations. Econ. Bot. 47, 234–247. doi: 10.1007/BF02862289
Haq, F., Ahmad, H., and Alam, M. (2011). Traditional uses of medicinal plants of Nandiar Khuwarr catchment (district Battagram), Pakistan. J. Med. Plant Res. 5, 39–48.
Haq, M. A., Alam, M. J., and Hasnain, A. (2013). Gum Cordia: a novel edible coating to increase the shelf life of Chilgoza (Pinus gerardiana). LWT Food Sci. Technol. 50, 306–311. doi: 10.1016/j.lwt.2012.05.008
Hayat, W., Khan, S., Iqbal, A., Ahmad, S., and Abbasi, A. M. (2021). Protection is better than management to maintain tree species: a case study of lesser-Himalayan moist-temperate forests of Pakistan. Trees, For. People 6:100149
Höhl, M., Ahimbisibwe, V., Stanturf, J. A., Elsasser, P., Kleine, M., and Bolte, A. (2020). Forest landscape restoration—what generates failure and success? Forests 11:938. doi: 10.3390/f11090938
Huo, H., Feng, Q., and Su, Y. (2014). The influences of canopy species and topographic variables on understory species diversity and composition in coniferous forests. Sci. World J. 2014, 1–8. doi: 10.1155/2014/252489
Hurlbert, S. H. (2012). The Nonconcept of Species Diversity: A Critique and Alternative Parameters. Plant world 52, 577–586. doi: 10.2307/1934145
Hussein, E. A., El-Ghani, M. M. A., Hamdy, R. S., and Shalabi, L. F. (2021). Do anthropogenic activities affect floristic diversity and vegetation structure more than natural soil properties in hyper-arid desert environments? Diversity 13:157. doi: 10.3390/d13040157
Kakembo, V., Rowntree, K., and Palmer, A. R. (2007). Topo- graphic controls on the invasion of Pteronia incana (blue bush) onto hillslopes in Ngqushwa (formerly Peddie) district, eastern cape, South Africa. Catena 70, 185–199. doi: 10.1016/j.catena.2006.08.005
Kayani, S., Ahmad, M., Zafar, M., Sultana, S., Khan, M. P. Z., Ashraf, M. A., et al. (2014). Ethnobotanical uses of medicinal plants for respiratory disorders among the inhabitants of Gallies – Abbottabad, northern Pakistan. J. Ethnopharmacol. 156, 47–60. doi: 10.1016/j.jep.2014.08.005
Khan, A., Ahmed, M., Khan, A., Siddiqui, M. F., Shah, M., and Hazrat, A. (2021). Quantitative description, present status and future trend of conifer forests growing in the indus kohistan region of khyber pakhtunkhwa, Pakistan. Pak. J. Bot. 53, 1343–1353. doi: 10.30848/PJB2021-4(10)
Khan, N., Ahmed, M., Shaukat, S. S., Wahab, M., and Siddiqui, M. F. (2011). Structure, diversity, and regeneration potential of Monotheca buxifolia (Falc.) a. DC. Dominated forests of lower Dir District, Pakistan. Front. Agric. China 5, 106–121. doi: 10.1007/s11703-011-1062-x
Khan, S. M., Page, S., Ahmad, H., Shaheen, H., and Harper, D. M. (2012). Vegetation dynamics in the Western Himalayas, diversity indices and climate change. Sci. Technol. Dev. 31, 232–243.
Khel, J., Shah, I. A., Badshah, L., Burni, T., Uza, N. U., and Khan, A. R. (2022). Floristic inventory and ecological evaluation of plants of Jani Khel, Bannu, Khyber Pakhtunkhwa, Pakistan. PAB 11, 881–890. doi: 10.19045/bspab.2022.110090
Liira, J., Sepp, T., and Parrest, O. (2007). The forest structure and ecosystem quality in conditions of anthropogenic disturbance along productivity gradient. For. Ecol. Manag. 250, 34–46. doi: 10.1016/j.foreco.2007.03.007
López-Marcos, D., Turrion, M.-B., Bravo, F., and Martinez-Ruiz, C. (2019). Understory response to overstory and soil gradients in mixed versus monospecific Mediterranean pine forests. Eur. J. For. Res. 138, 939–955. doi: 10.1007/s10342-019-01215-0
López-Marcos, D., Turrión, M.-B., Bravo, F., and Martínez-Ruiz, C. (2020). Can mixed pine forests conserve understory richness by improving the establishment of understory species typical of native oak forests? Ann. For. Sci. 77, 1–13. doi: 10.1007/s13595-020-0919-7
Luintel, H., Bluffstone, R. A., and Scheller, R. M. (2018). The effects of the Nepal community forestry program on biodiversity conservation and carbon storage. PLoS One 13:e0199526. doi: 10.1371/journal.pone.0199526
Mansourian, S., Parrotta, J., Balaji, P., Bellwood-Howard, I., Bhasme, S., Bixler, R. P., et al. (2020). Putting the pieces together: integration for forest landscape restoration implementation. L. Degrad. Dev. 31, 419–429. doi: 10.1002/ldr.3448
Margalef, R. (1958). Temporal succession and spatial heterogeneity in natural phytoplankton. Digital. csic.es. 23–29.
Mestre, L., Toro-Manríquez, M., Soler, R., Huertas-Herrera, A., Martínez-Pastur, G., and Lencinas, M. V. (2017). The influence of canopy-layer composition on understory plant diversity in southern temperate forests. For. Ecosyst. 4, 1–13. doi: 10.1186/s40663-017-0093-z
Mishra, B. P., Tripathi, O. P., Tripathi, R. S., and Pandey, H. N. (2004). Effects of anthropogenic disturbance on plant diversity and community structure of a sacred grove in Meghalaya, Northeast India. Biodivers. Conserv. 13, 421–436. doi: 10.1023/B:BIOC.0000006509.31571.a0
Mölder, A., Bernhardt-Römermann, M., and Schmidt, W. (2008). Herb-layer diversity in deciduous forests: raised by tree richness or beaten by beech? For. Ecol. Manag. 256, 272–281. doi: 10.1016/j.foreco.2008.04.012
Muhammad, Z., Akash, J., Din, M. Z., Syed, B. H., and Kashif, A. (2019). Vegetation resource inventory of NWFPs in the state of Azad Jammu and Kashmir. Fresenius Environ. Bull. 28, 8591–8600.
Muhammad Azam Khan, M. A., Hussain, M., and Ghulam, G. M. (2010). An ethnobotanical inventory of himalayan region poonch valley Azad Kashmir (Pakistan). Ethnobot. Res. Appl. 8:107, 107–124. doi: 10.17348/era.8.0.107-123
Ortiz-Pulido, R., and Pavon, N. P. (2010). Influence of slope orientation on sex ratio and size distribution in a dioecious plant Bursera fagaroides var. purpusii (Brandeg.) McVaugh and Rzed. (Burseraceae). Plant Ecol. 208, 271–277. doi: 10.1007/s11258-009-9704-2
Ou, Z., Pang, S., He, Q., Peng, Y., Huang, X., and Shen, W. (2020). Effects of vegetation restoration and environmental factors on understory vascular plants in a typical karst ecosystem in southern China. Sci. Rep. 10:12011. doi: 10.1038/s41598-020-68785-7
Peruzzi, L. (2020). Using scientific names guarantees universality of communication in science… but are plant biologists aware of it? Plant Biosyst. 154, 1000–1002. doi: 10.1080/11263504.2020.1736203
Prakash, O., Samant, S. S., and Yadava, A. K. (2023). Assessment of natural population and regeneration dynamics of Pinus gerardiana in dry temperate region of north western Himalaya. Trop. Ecol. 64, 26–36. doi: 10.1007/s42965-021-00176-z
Print, I., Khan, H., Akbar, M., Zaman, M., Ali, A., and Hyder, S. (2015). Diameter size class distributions of Pinus gerardiana wall. Ex D. Don from Gohar Abad Valley district Diamer, Gilgit-Baltistan, Pakistan Forest education division Pakistan Forest Institute, Peshawar, Pakistan: J. Biodivers, Environ.Sci. 6, 50–56.
Promis, A., Gärtner, S., Reif, A., and Cruz, G. (2012). Effects of canopy gaps on forest floor vascular and non-vascular plant species composition and diversity in an uneven-aged Nothofagus betuloides forest in Tierra del Fuego, Chile. Community Ecol. 13, 145–154. doi: 10.1556/ComEc.13.2012.2.3
Qasim, M., Khan, M. Z., Naz, A., and Khalid, S. (2013). An insight of ecosystem capitals and Services of the Kaghan Valley: the Himalayan region of Pakistan. Nat. Res. Forum 4, 163–169. doi: 10.4236/nr.2013.42021
Rahimzadeh, A. (2020). Socio-economic and environmental implications of the decline of Chilgoza pine nuts of Kinnaur, Western Himalaya. Conserv. Soc. 18, 315–326. doi: 10.4103/cs.cs_19_17
Rahman, I. U., Khan, N., and Ali, K. (2017). Classification and ordination of understory vegetation using multivariate techniques in the Pinus wallichiana forests of Swat Valley, northern Pakistan. Sci. Nat. 104, 1–10. doi: 10.1007/s00114-017-1431-2
Richardson, D. M., and Rundel, P. W. (1998). “An introduction” in Ecol. Biogeogr. Pinus. ed. D. M. Richardson (UK: Cambridge Univ. Press Cambridge), 3–46.
Sagar, R., Singh, A., and Singh, J. S. (2008). Differential effect of woody plant canopies on species composition and diversity of ground vegetation: a case study. Trop. Ecol. 49, 189–197.
Shalizi, M. N., and Khurram, S. (2016). Socio-economic importance of chilgoza pine forest of Afghanistan: a survey based assessment. Asian J. Sci. Technol. 7, 3556–3559.
Shalizi, M. N., Khurram, S., Groninger, J. W., Ruffner, C. M., and Burney, O. T. (2018). Indigenous knowledge and stand characteristics of a threatened tree species in a highly insecure area: Chilgoza pine in Afghanistan. For. Ecol. Manag. 413, 1–8. doi: 10.1016/j.foreco.2018.01.053
Shannon, C. E., The mathematical theory of communication, (and recent contributions to the mathematical theory of communication), W. Weaver. University of illinois Press Champaign, IL, USA. (1949).
Sharm, C. M., Baduni, N. P., Gairola, S., Ghildiyal, S. K., and Suyal, S. (2010). Effects of slope aspects on forest compositions, community structure and soil properties in natural temperate forest of Garhwal Himalaya. Ecol. Res. 21, 331–337. doi: 10.1007/s11676-010-0079-y
Sher, H., Aldosari, A., Ali, A., and De Boer, H. J. (2014). Economic benefits of high value medicinal plants to Pakistani communities: An analysis of current practice and potential. J. Ethnobiol. Ethnomed. 10, 1–16.
Sher, H., Bussmann, R. W., Hart, R., and De Boer, H. J. (2016). Traditional use of medicinal plants among Kalasha, Ismaeli and Sunni groups in Chitral District, Khyber Pakhtunkhwa province, Pakistan. J. Ethnopharmacol. 188, 57–69. doi: 10.1016/j.jep.2016.04.059
Shinwari, Z. K., and Qaiser, M. (2011). Efforts on conservation and sustainable use of medicinal plants of Pakistan. Pak. J. Bot. 43, 5–10.
Society, T. A., and Press, C. (2016). On the Relative Abundance of Species. Am. Nat. 94, 25–36. doi: 10.1086/282106
Sorensen, T. A. (1948). A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biol. Skar. 5, 1–34.
Su, C., and Lu, C. (2020). Distribution of eight species of large-seeded pines and their primary animal seed-dispersers in China: match or mismatch? Authorea Prepr. 2, 101–111.
Tabassum, S. (2020). Deforestation in Pakistan CPEC an auxiliary exacerbation. J. Contemp. Stud. 9, 59–72.
The Angiosperm Phylogeny Group (2009). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161, 105–121. doi: 10.1111/j.1095-8339.2009.00996.x
Tinya, F., Márialigeti, S., Király, I., Németh, B., and Ódor, P. (2009). The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in O{double acute}rség, Western Hungary. Plant Ecol. 204, 69–81. doi: 10.1007/s11258-008-9566-z
Ullah, M., Khan, M. U., Mahmood, A., Malik, R. N., Hussain, M., Wazir, S. M., et al. (2013). An ethnobotanical survey of indigenous medicinal plants in Wana district South Waziristan agency, Pakistan. J. Ethnopharmacol. 150, 918–924. doi: 10.1016/j.jep.2013.09.032
Urooj, R. (2019). Status, distribution and dynamics of Chilgoza pine (Pinus Gerardiana Wall) Forest in Suleiman Mountain range, Pakistan. Middle East J. Bus. 14, 19–21. doi: 10.5742/mejb.2019.93606
Urooj, R., and Jabeen, A. (2015). Present status of Pinus Gerardiana Wall. In Pakistan: a review. Middle East J. Bus. 10, 46–48. doi: 10.5742/mejb.2015.92719
Urooj, R., Sherani, S. H., Ehtasham, L., Younas, K., and Izbel, U. (2020). Geospatial analysis of chilghoza forest in Sulaiman mountainous range. SN Appl. Sci. 2, 1–9. doi: 10.1007/s42452-020-03737-4
Vellend, M., Verheyen, K., Jacquemyn, H., Kolb, A., van Calster, H., Peterken, G., et al. (2006). Extinction debt of forest plants persists for more than a century following habitat fragmentation. Ecology 87, 542–548. doi: 10.1890/05-1182
Von Oheimb, G., and Härdtle, W. (2009). Selection harvest in temperate deciduous forests: impact on herb layer richness and composition. Biodivers. Conserv. 18, 271–287. doi: 10.1007/s10531-008-9475-4
Walker, B., Kinzig, A., and Langridge, J. (1999). Plant attribute diversity, resilience, and ecosystem function: the nature and significance of dominant and minor species. Ecosystems 2, 95–113. doi: 10.1007/s100219900062
Wang, Y., and Shangguan, T. (2010). Discussion on calculating method of important values. J. Shanxi Univ. (Natural Sci. Ed.) 33, 312–316.
Wiegert, R. G. (1989). Documenting Cmpleteness, species-area relations, and the species-abundance distribution of a regional Flora. Ecolog. Society America 70, 16–22.
Wulf, M., and Naaf, T. (2009). Herb layer response to broadleaf tree species with different leaf litter quality and canopy structure in temperate forests. J. Veg. Sci. 20, 517–526. doi: 10.1111/j.1654-1103.2009.05713.x
Wyatt, J. L., and Silman, M. R. (2010). Centuries-old logging legacy on spatial and temporal patterns in understory herb communities. For. Ecol. Manag. 260, 116–124. doi: 10.1016/j.foreco.2010.04.010
Xie, H., Tang, Y., Yu, M., and Wang, G. G. (2021). The effects of afforestation tree species mixing on soil organic carbon stock, nutrients accumulation, and understory vegetation diversity on reclaimed coastal lands in eastern China. Glob. Ecol. Conserv. 26:e01478. doi: 10.1016/j.gecco.2021.e01478
Zhang, Q., Fang, R., Deng, C., Zhao, H., Shen, M. H., and Wang, Q. (2022). Slope aspect effects on plant community characteristics and soil properties of alpine meadows on eastern Qinghai-Tibetan plateau. Ecol. Indic. 143:109400. doi: 10.1016/j.ecolind.2022.109400
Zilliox, C., and Gosselin, F. (2014). Tree species diversity and abundance as indicators of understory diversity in French mountain forests: variations of the relationship in geographical and ecological space. For. Ecol. Manag. 321, 105–116. doi: 10.1016/j.foreco.2013.07.049
Zubair, M., Jamil, A., Hussain, S. B., Ul Haq, A., Hussain, A., Zahid, D. M., et al. (2021). Diversity of medicinal plants among different tree canopies. Sustain. For. 13, 1–13. doi: 10.3390/su13052640
Keywords: understory vegetation, diversity indices, medicinal plants, aspect vegetation diversity, Pinus gerardiana
Citation: Khan I, Zahid DM, Zubair M, Manzoor SA, Yasin G, Mohammad MW, Khan KA, Rahman SU and Qin A (2023) Influence of aspect on vegetation dynamics: insights into the understory vegetation diversity of the dry temperate forests of South Waziristan Agency, Pakistan. Front. For. Glob. Change. 6:1199601. doi: 10.3389/ffgc.2023.1199601
Edited by:
Ngaba Mbezele Junior Yannick, Southwest University, ChinaReviewed by:
Mazhar Rafique, The University of Haripur, PakistanAhmad Hussain, Karakoram International University, Pakistan
Carolina Martínez-Ruiz, University of Valladolid, Spain
Copyright © 2023 Khan, Zahid, Zubair, Manzoor, Yasin, Khan, Mohammad, Rahman and Qin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shafeeq Ur Rahman, bWFsaWtzaGFmZWVxMTU1OUBnbWFpbC5jb20=; Anzhen Qin, cWluYW56aGVuQGNhYXMuY24=