 Bram Verstockt1,2†
Bram Verstockt1,2† Clémentine Perrier1,3†Gert De Hertogh4Jonathan Cremer1,3Brecht Creyns1,3Gert Van Assche1,2Marc Ferrante1,2Jan L. Ceuppens3Séverine Vermeire1,2Christine Breynaert3*
Clémentine Perrier1,3†Gert De Hertogh4Jonathan Cremer1,3Brecht Creyns1,3Gert Van Assche1,2Marc Ferrante1,2Jan L. Ceuppens3Séverine Vermeire1,2Christine Breynaert3*- 1KU Leuven, Department of Chronic Diseases, Metabolism and Ageing, Translational Research for Gastrointestinal Disorders, Leuven, Belgium
- 2University Hospitals Leuven, Department of Gastroenterology and Hepatology, Leuven, Belgium
- 3KU Leuven, Laboratory of Clinical Immunology, Department of Microbiology and Immunology, Leuven, Belgium
- 4KU Leuven, Department of Imaging & Pathology, Translational Cell & Tissue Research, Leuven, Belgium
Background: Mucosal IL-13 Receptor alpha 2 (IL13RA2) mRNA expression is one of the best predictive markers for primary non-responsiveness to infliximab therapy in patients with inflammatory bowel disease (IBD). The objective of this study was to understand how IL-13Rα2, a negative regulator of IL-13 signaling, can contribute to IBD pathology.
Methods: IL13RA2 knockout (KO) and wild type (WT) mice were exposed to dextran sodium sulfate (DSS) in drinking water to induce colitis. Furthermore, mucosal biopsies and resection specimen of healthy individuals and IBD patients before the start of anti-tumor necrosis factor (anti-TNF) therapy were obtained for immunohistochemistry and gene expression analysis.
Results: After induction of DSS colitis, IL13RA2 KO mice had similar disease severity, but recovered more rapidly than WT animals. Goblet cell numbers and mucosal architecture were also more rapidly restored in IL13RA2 KO mice. In mucosal biopsies of active IBD patients, immunohistochemistry revealed that IL-13Rα2 protein was highly expressed in epithelial cells, while expression was restricted to goblet cells in healthy controls. Mucosal IL13RA2 mRNA negatively correlated with mRNA of several goblet cell-specific and barrier genes, and with goblet cell numbers.
Conclusions: The data suggest that IL-13Rα2 on epithelial cells contributes to IBD pathology by negatively influencing goblet cell recovery, goblet cell function and epithelial restoration after injury. Therefore, blocking IL-13Rα2 could be a promising target for restoration of the epithelial barrier in IBD.
Introduction
Crohn's disease (CD) and ulcerative colitis (UC) are chronic inflammatory disorders of the gastrointestinal tract with a relapsing and remitting course. They are thought to result from an excessive immune response toward the intestinal and colonic microbiota in genetically susceptible hosts (1). Current treatments, able to achieve and maintain clinical remission and mucosal healing, include anti-tumor necrosis factor (anti-TNF) agents. Despite an overall good response to anti-TNF therapies, 10–30% of patients are primary non-responders (2). Among several other genes, our group previously identified interleukin 13 receptor alpha 2 (IL13RA2) mRNA expression as one of the best predictive markers for anti-TNF non-responsiveness. As this holds true both in UC and in CD, two diseases mediated by different combinations of cytokines (1, 3, 4), these results point toward a role of IL-13Rα2 in a mechanism common to both diseases. In a more recent study we also found that IL13RA2 expression reflects an increased TNF burden in non-responders (5). However, how IL-13Rα2 affects the response to anti-TNF therapy is currently unknown.
The IL-13 signaling receptor complex consists of two chains (IL-13Rα1 and IL-4Rα) which, after ligand binding, activate the transcription factor STAT-6 (6). IL-13Rα2 is a transmembrane receptor which binds IL-13 with very high affinity. Due to its very short cytoplasmic region lacking signaling motifs, IL-13Rα2 is generally thought to act primarily as a “decoy” receptor, sequestering IL-13 and thus preventing excessive IL-13 signaling via the complex IL-13Rα1 and IL-4Rα (7). In rodents, IL-13Rα2 is released as a soluble factor and its expression correlates with a decreased responsiveness to IL-13 in chronic granulomatous inflammation in the liver (8), airway inflammation and airway-hyperreactivity (9), and immune and functional response to nematode infection (10). Humans do not alternatively splice the IL13Rα2 transcript and exclusively express this receptor as a transmembrane molecule (6, 11). However, it has been shown that also in humans, membrane IL-13Rα2 reduces responsiveness to IL-13 (12–14).
The present study aimed to identify how IL13RA2 expression can contribute to the pathology of IBD. First, we performed experimental studies with IL13RA2 KO and WT mice subjected to experimental colitis. Next, we determined the localization of the IL-13Rα2 protein in human colonic biopsies by immunohistochemistry. Based on the murine findings and because it is known that IL-13 influences goblet cells, we further correlated IL13RA2 expression with goblet cell specific genes, barrier genes and goblet cell numbers in IBD.
Materials and Methods
Animals and Experimental Colitis Model
IL13RA2 knock out (KO) mice backcrossed more than 10 times in the Balb/c background were obtained from Pfizer (New York, NY, USA). Both IL13RA2 KO and wild type (WT) Balb/c mice were bred in the specific pathogen free area of the animal care facility of the KU Leuven, Belgium. Acute colitis was induced by administration of dextran sodium sulfate (DSS, MP Biomedicals) in drinking water (3%) for 7 days, followed by a recovery period. Mice were sacrificed by injection of pentobarbital at day 9 or 12 for evaluation of markers of disease activity and inflammation. Colons were harvested, weighted and measured from the ileocecal junction to the anus. A macroscopic score of inflammation was given as previously described (15). One part of the most infiltrated distal colon was fixed in 6% paraformaldehyde for 3 h and then embedded in paraffin for histology evaluation.
Evaluation of Inflammation in Mice by Histology
Five μm-thick longitudinal and transverse sections of colons were stained with hematoxylin and eosin (H&E). Three sections per mouse were blindly and independently scored by a post-doctoral researcher (CP) and by an experienced IBD pathologist (GDH). The following five characteristics were scored cumulatively. Architectural disturbance, based on irregularity of the mucosal surface, crypt distortion and branching, loss of glands, and fraying of the muscularis mucosae (0: none, 1: focal and mild, 2: multifocal, or diffuse, 3: severe); goblet cell depletion (0: normal, 1: focal, 2: multifocal, and 3: generalized); epithelial defects (0: none, 1: focal erosion, 2: multifocal erosion, and 3: ulceration); neutrophil infiltration (0: none, 1: in the lamina propria, 2: in the lamina propria with crypt abscesses, 3: infiltration in the mucosa or in the submucosa, or in the muscularis propria or in the subserosa, to a maximum of 12 if several areas were infiltrated); mononuclear cell infiltration (0: within normal limits, 1: slightly increased in lamina propria, 2: dense infiltrate in the lamina propria, 3: cell aggregates in the mucosa or in the submucosa or in the muscularis propria or in the subserosa, to a maximum of 12 if several areas were infiltrated).
Immunohistochemistry for Human IL-13Rα2 and IL-13Rα1 and Quantification of Goblet Cells in Mucosal Biopsies of IBD Patients
Immunohistochemical staining was performed on 5 μm-thick step sections prepared from paraffin formalin-fixed endoscopic-derived mucosal biopsies and resection specimens from IBD patients and controls. Endogenous peroxidase activity was blocked in deparrafined sections by incubating the slides for 20 min in a 0.3% solution of H2O2 in methanol. Epitope retrieval was performed by heating the slides for 30 min in Tris/EDTA buffer (pH 9) at 98°C. Sections were then incubated with the anti-human IL-13Rα2 mouse monoclonal antibody clone ab55275 (Abcam plc, Cambridge, UK) at a concentration of 1 μg/ml for 30 min. IL-13Rα1 protein was localized using anti-human IL-13Rα1 rabbit polyclonal antibody ab79277 (Abcam) at a concentration of 10 μg/ml. The Dako REALTM EnvisionTM Detection System kit (Dako Belgium NV, Heverlee, Belgium) was used for visualization of bound primary antibody according to the manufacturer's instructions. Formalin-fixed, paraffin-embedded surgical biopsies of an ovarian serous adenocarcinoma served as positive controls for IL-13Rα2 (16). The primary antibody was omitted in the negative controls.
In parallel, H&E stained sections of mucosal biopsies were used to quantify goblet cells. Numbers of goblet cell were counted manually on images of the biopsies. Each picture covers a total surface of 0.32 mm2. The total length of the epithelium on the picture was measured using ImageJ, and data were expressed as number of goblet cells per μm of epithelium.
Gene Expression Study in Mucosal Biopsies of IBD Patients
Publically available microarray data (Affymetrix Human Genome U133 Plus 2.0 Arrays, GSE14580, GSE12251, and GSE16879) of inflamed mucosal biopsies of IBD patients prior to their first infliximab administration were analyzed. Patient characteristics at baseline were previously described (17, 18). Raw data were preprocessed {normalization robust multichip average (RMA) metho(19)}, , and quality-control evaluation was performed [arrayQualityMetrics (20)] using Bioconductor (http://www.bioconductor.org) in R version 3.5.0 (R Development Core Team, Vienna, Austria), whereafter outliers were detected and removed based on cluster analysis (21). Microarray data were used to quantify mRNA expression of goblet cell specific and barrier genes.
Statistical Analysis
Murine data analysis was performed with GraphPad Prism 5 (La Jolla, CA, USA). Data are represented as medians, and P-values were obtained using two-tailed Mann-Whitney U testing. Differences were considered statistically significant at p < 0.05. Human data analysis was performed in R (R Development Core Team, Vienna, Austria). Correlations between IL13RA2 expression and goblet cell related genes and numbers were analyzed using bivariate two-tailed correlation tests.
Results
IL13Rα2 KO Mice Are Equally Affected in Acute DSS Colitis as WT Mice, but Recover Faster
Acute colitis was induced in IL13RA2 KO and WT mice by DSS administration. At day 9, weight loss was most severe in both strains, reflecting the time point of strongest inflammation. At day 12 (recovery phase), most mice had recovered their initial weight despite persistent inflammatory features in the colon. Both strains developed colitis and equally lost weight (p = 0.07 at both day 9 and 12) (Figure 1A). The inflammation of the colon, as evaluated by the macroscopic score, the colon length, the colon weight and the colon weight to length ratio, was similar in IL13RA2 KO and WT mice at day 9 (p = 0.10, 0.34, 0.18, and 0.41, respectively) (Figures 1B–E). In contrast, at day 12, colons of IL13RA2 KO mice were less inflamed than colons of WT mice, with significant differences in the macroscopic score of inflammation (p = 0.01), the colon length (p < 0.001) and colon weight (p = 0.01), reflecting that inflammation resolved more rapidly in IL13RA2 KO mice.
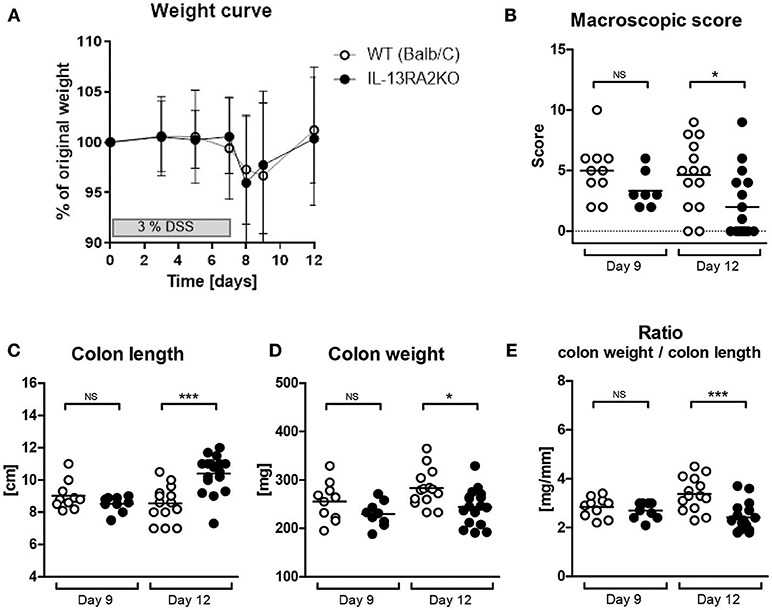
Figure 1. Acute colitis in WT and IL13RA2 KO mice. Wild type (WT) and IL13RA2 knock out (KO) mice were exposed for 7 days to 3% DSS in drinking water to induce acute colitis. Data are pooled from two independent experiments including 24 WT mice (day 9: n = 10 and day 12: n = 14) and 26 IL13RA2 KO mice (day 9: n = 9 and day 12: n = 17). WT mice are shown in white, IL13RA2 KO mice in black. Weight curve (A); Macroscopic score of inflammation of the colon at day 9 and day 12 after start of DSS exposure (B); Length of colon (C); Weight of colon (D); Ratio colon weight on length (E). Data are represented as medians (*p < 0.05, ***p ≤ 0.001, NS, not significant).
Inflammation was microscopically quantified on H&E stained colon sections (Figure 2A). Microscopically, the score of inflammation was not different between IL13RA2 KO and WT mice at day 9 (p = 0.82), but it was significantly lower in IL13RA2 KO mice at day 12 (p = 0.02) (Figure 2B). At day 12, more goblet cells were present in the epithelium of IL13RA2 KO mice (arrows). More specifically, the characteristics that were different between the two strains of mice on day 12 were the architectural changes (p = 0.009), the goblet cell depletion (p = 0.01) and the infiltration of neutrophils (p = 0.03) (Figure 2C). Taken together, these data demonstrate that the absence of IL-13Rα2 allows a more rapid recovery of the mucosa and in particular for the regeneration of goblet cells and normalization of the architecture of the colonic mucosa. IL-13Rα2 thus seems to negatively regulate epithelial/mucosal healing.
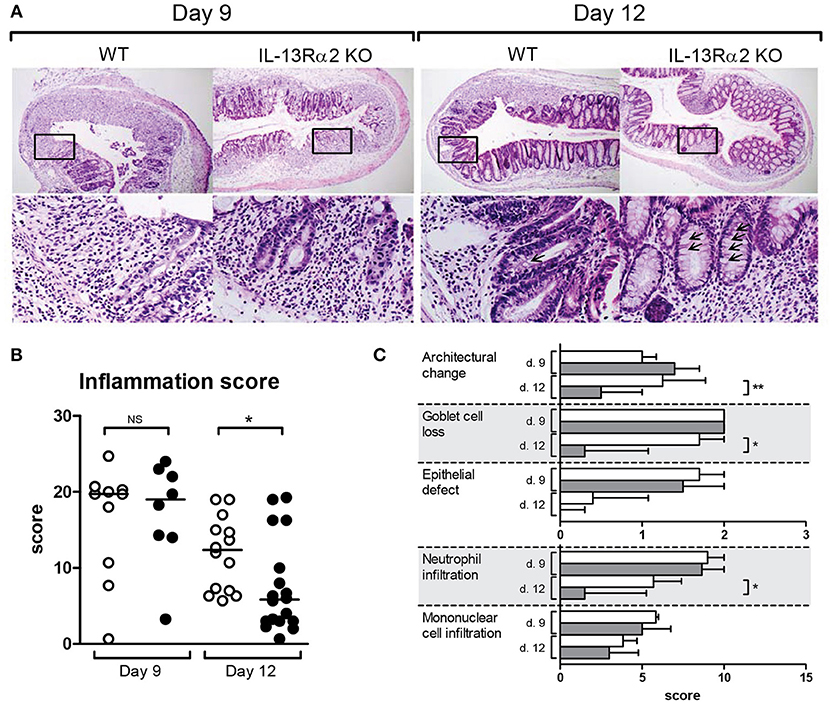
Figure 2. Microscopic score of inflammation in mice with DSS colitis. Wild type (WT) and IL13RA2 knock out (KO) mice were exposed for 7 days to 3% DSS in drinking water to induce acute colitis. Data are pooled from two independent experiments including 24 WT mice (day 9: n = 10 and day 12: n = 14) and 26 IL13RA2 KO mice (day 9: n = 9 and day 12: n = 17). WT mice are shown in white, IL13RA2 KO mice in black. Representative pictures of haemotoxylin and eosin (HandE) stained cross sections of colon of mice. The lower pictures represent the area in the rectangle at higher magnification (A); Inflammatory score evaluated on histological sections (B); Details of the inflammatory score (C). Data are represented as medians (*p < 0.05, **p ≤ 0.01, NS, not significant).
IL-13Rα2 Is Expressed in Human Epithelial Cells
Using mucosal gene expression gene signaling analysis, we previously reported that IL13RA2 is highly expressed in the mucosa of IBD patients, and more so in anti-TNF non-responders (5, 17, 18). We therefore determined the cellular localization of the IL-13Rα2 protein by immunohistochemistry on mucosal surface biopsies and full thickness resection specimen of IBD patients and healthy controls. In normal colon, IL-13Rα2 was localized in the cytoplasm and on the membrane of the goblet cells (Figure 3A). In inflamed CD and UC biopsies, there was increased staining intensity for IL-13Rα2 in all epithelial cells (Figures 3B,C). The staining was very intense at the base of the crypts and became slightly less intense in cells at the tip of the villi. We did not observe any staining in lymphoid aggregates, nor in cells surrounding fibrotic areas in resection specimen of CD and UC patients (Figures 3D,E). Immune cells, such as macrophages and monocytes, as well as fibroblasts did not express IL13RA2, even in the deeper layer of the tissues. In resection specimens of patients with colorectal carcinoma, IL13RA2 was also expressed in epithelial cells surrounding the tumor, although the intensity of the staining was slightly weaker than in CD and UC patients. The IL13RA2 expression in these patients suggests that it is not specific to IBD, but might be a consequence of chronic inflammation.
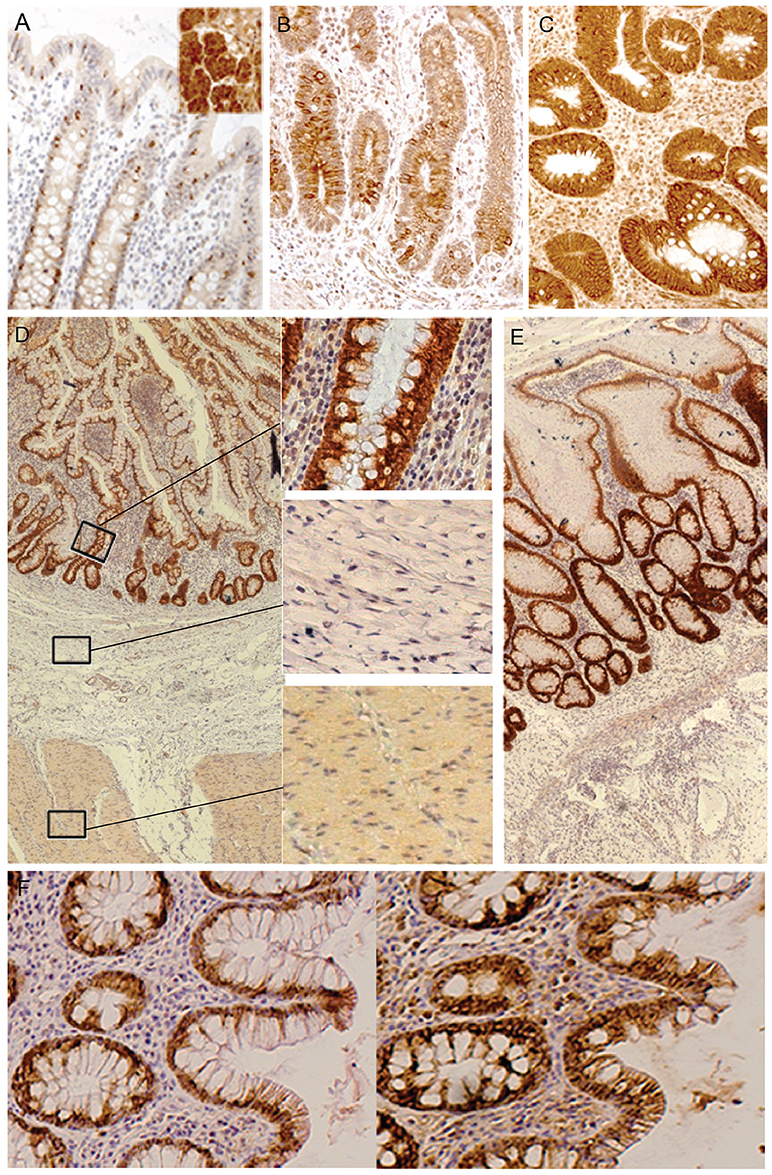
Figure 3. Immunohistochemical detection of IL-13Rα2 and IL-13Rα1 in human tissues. Staining of IL-13Rα2 protein with specific antibody in human tissues: normal colon and in the insert an ovarian serous adenocarcinoma as positive control (16) (negative controls where the first antibody was omitted are not shown) (A); Ileum of a CD non-responder before therapy (B); Colon of an UC non-responder before therapy (C); Resection specimen of CD (D); Resection specimen of UC (E); Sequential staining of colon of UC non-responders for IL-13Rα2 (left) and IL-13Rα1 (right) (F). The black bar corresponds to 100 μm.
As IL-13 signals via the complex IL-13Rα1/IL-4Rα, we analyzed whether IL-13Rα1 was also expressed in epithelial cells of IBD patients. In patients with strong inflammation, IL-13Rα1 was co-expressed in epithelial cells together with IL-13Rα2 (Figure 3F). As expected, IL-13Rα1 was also expressed in lymphoid cells in the lamina propria (data not shown).
Expression of IL13RA2 mRNA Negatively Correlates With Expression of Goblet Cell Related Genes and Goblet Cell Numbers in the Mucosa of IBD Patients
In follow up of our murine findings on goblet cell recovery in the absence of IL-13Rα2, we analyzed the correlation between IL13RA2 expression and the expression of various goblet cell related genes. Mucosal expression of ATOH1, a transcription factor specific for the development of goblet cells, SPDEF, a key promotor of goblet cell differentiation and regulator of secretory gene products including mucin 2, and RELMβ, a key goblet cell mediator, were negatively correlated with IL13RA2 expression (Figures 4A–C). Additionally, IL13RA2 mRNA expression correlated inversely with gene expression of key components of the intestinal mucus layer (22), including FCGBP and CLCA1 (r = −0.5, p = 4.0 × 10−4; r = −0.4, p = 2.0 × 10−3, respectively) (Supplementary Figures 1A,B). To strengthen these findings, the number of goblet cells per mm epithelium was quantified in mucosal biopsies of IBD patients, and correlated negatively with the expression of IL13RA2 (r = −0.33, p = 0.03) (Figure 4D). Thus, the expression of IL13RA2 prior to therapy is associated with lower numbers of goblet cells, lower expression of transcription factors for goblet cell development and goblet cell function. This suggests that IL-13Rα2 negatively regulates development of goblet cells during inflammation, most likely by counteracting the effects of IL-13.
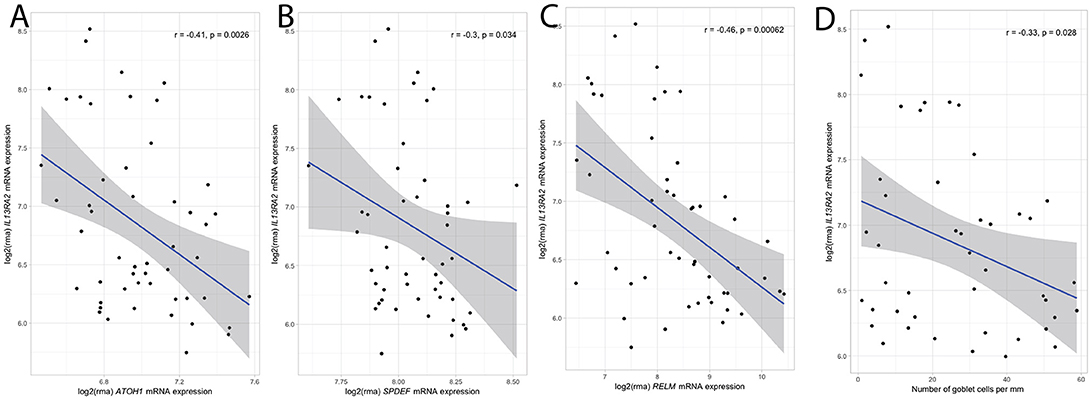
Figure 4. Correlation between IL13RA2 mRNA and mRNA of goblet cell related genes. Correlations between expression of IL13RA2 with the expression of ATOH1 (A), SPDEF (B), RELMβ (C) in mucosal biopsies of IBD patients before anti-TNF therapy. Correlations between expression of IL13RA2 with the number of goblet cells per mm in IBD patients prior to start of anti-TNF therapy (D).
In general, these murine and human findings suggest a link between IL13RA2 and mucosal barrier function. Based on previous findings demonstrating an intestinal epithelial barrier dysfunction in IBD, with up- (CDN1, DSG3) or downregulation (CLDN8, OCLDN, MEP1A, and MAGI1) of particular genes in active IBD compared to healthy individuals (23), we studied the correlation between IL13RA2 expression and these barrier genes. Although causality cannot be proven, we found a significant positive correlation between IL13RA2 and CDN1 (r = 0.38, p = 0.006) and DSG3 expression (r = 0.61, p = 2.2 × 10−6), whereas IL13RA2 correlated negatively with CLDN8 (r = −0.38, p = 0.006), OCLDN (r = −0.44, p = 0.001), MEP1A (r = −0.35, p = 0.01), MAGI1 (r = −0.43, p = 0.002), and MARVELD2 (r = −0.32, p = 0.02).
Discussion
Previously, IL13RA2 mRNA expression has been identified as one of the best predictive markers for anti-TNF non-responsiveness (5). However, how IL-13Rα2 interferes with the effect of anti-TNF therapy is currently unknown. The present study aimed to unravel the effects of mucosal IL-13Rα2 expression in IBD. We first hypothesized that IL-13Rα2 downregulates the inflammatory process, as it has been shown to inhibit several IL-13-mediated effects in mice (8–10, 24). IL-17A/IFNγ-mediated colitis in IL10 KO mice is regulated indirectly by soluble IL-13Rα2 (25), which blocks the inhibitory function of IL-13 on Th17 cells, and thus promotes Th17 activities (26). However, we did not notice any difference in macro- or microscopic inflammation between WT and IL13RA2 KO mice in this acute DSS colitis model, arguing against a role for IL13RA2 in the inflammatory process. Still it might be important to study other models of IBD. The DSS model was chosen for our study because DSS triggers inflammation by disruption of the epithelial barrier (27), and is as such an appropriate model to study mucosal healing. Importantly, using this model we indeed did observe a more rapid epithelial recovery in IL13RA2 KO mice, including more goblet cells at day 12. These findings suggest that IL13RA2 expression is not influencing susceptibility to colitis, but instead influences the recovery after removal of the trigger in mice, and that in particular effects on goblet cells are important. Indeed, also in human genome-wide association studies in IBD already pointed toward distinct mechanisms between disease susceptibility and disease outcome, once the disease has settled (28).
Besides its effects on inflammation, IL-13 has been shown to promote airway epithelial cell proliferation (29–31), and to prevent apoptosis (32), features that are crucial for epithelial cell recovery. IL-13 has an established role in the differentiation of goblet cells in the airways (30, 33, 34), and overexpression of IL-13 in the gut results in villus blunting, goblet cell hyperplasia and increased epithelial cell proliferation (35). The role of IL-13 in goblet cell hyperplasia is further supported by studies showing that stimulation of primary lung epithelial cells by IL-13 causes an increase in the population of goblet cells (33). These effects are likely to be mediated via IL-13Rα1 and STAT6. Co-expressed IL-13Rα2 can act as a negative regulator of IL-13 effects, reducing signaling via IL-13Rα1, and thus reducing the above-mentioned beneficial effects of IL-13 in the gut. It has already been demonstrated that IL13RA2 expression is inversely correlated with STAT-6 phosphorylation (36–38). As STAT-6 is key in goblet cell hyperplasia and intestinal mucosal repair (39, 40), the observed goblet cell hyperplasia in IL13RA2 KO mice can easily be explained.
Interestingly, when we studied biopsies of patients with IBD, we confirmed an inverse correlation between IL13RA2 expression and total number of goblet cells, as well as between IL13RA2 expression and the expression of several goblet cell specific and barrier genes. We therefore hypothesize that the increased IL13RA2 expression dampens one's ability to restore the mucosal barrier and hence impairs mucosal healing. Although goblet cell hyperplasia and mucus production contribute to inflammation in asthma (41), these mechanisms are protective in the gut (42, 43). Translated to IBD patients this may explain why some patients achieve mucosal healing and others do not, after receiving the same anti-TNF agent, neutralizing one of the driving forces and upstream regulators of IL13RA2 expression (5). Additionally, blocking IL-13Rα2 could be a promising agent for restoration of the epithelial barrier in IBD.
We cannot exclude that IL-13Rα2 also influences disease by other mechanisms. Besides IL-13, another ligand for IL-13Rα2, chitinase 3-like 1 (CHI3L1/YKL-40) in complex with TMEM219 (the chitosome) or Galectin-3 (Gal-3), can regulate a variety of cellular and tissue responses (14, 44, 45). CHI3L1, secreted by macrophages and neutrophils, plays essential roles in the pathogen clearance and generation of the host tolerance (46). The CHI3L1-IL13RA2 axis is known to play a critical role in cell death, inflammasome activation, Th1/Th2 cytokine balance, and Erk, Akt, and Wnt/β-catenin signaling (45). Recent data suggested that IL-13Rα2 is an α-receptor subunit that can interact with TMEM219 to mediate anti-apoptotic effector responses, whereas IL-13Rα2 interactions with Gal-3 would augment apoptotic responses and Wnt/β-catenin activation (45). Additional research is necessary to further unravel the significance of these effects of IL-13RA2, also in IBD.
In conclusion, IL13RA2 expression correlates negatively with numbers of goblet cells, as well as with the expression of goblet cell transcription factors and goblet cell function, suggesting that high IL13RA2 expression impairs goblet cell development. The role of IL-13Rα2 is further supported by murine data showing a more rapid recovery of IL13RA2 KO mice compared to WT mice after DSS induced colitis, with a more rapid restoration of goblet cells. In anti-TNF therapy non-responder patients in whom there is increased IL13RA2 expression (5, 17, 18), the latter probably hampers the renewal of the epithelium and the differentiation of goblet cells that would otherwise be induced by IL-13 via the IL-13Rα1/IL-4Rα complex. Neutralization of TNF by anti-TNF therapies decreases the inflammation via several mechanisms (47), but it is conceivable that this can be efficient only if the epithelium recovers properly to prevent entry of commensal bacteria and/or bacterial products contributing to the inflammation.
Ethics Statement
All patients included in the analysis had given written consent to participate in the Institutional Review Board approved IBD Biobank (B322201213950/S53684), collecting serum, biopsies and clinical characteristics among other items. All murine studies were approved by the local ethical committee for animal experimentation of the KU Leuven(P013/2011).
Author Contributions
BV: contributed to the acquisition of data, analysis and interpretation of data, drafting of the manuscript, and statistical analysis. CP: study concept and design, acquisition data, analysis and interpretation of data, and drafting of the manuscript. GDH: acquisition data, analysis and interpretation of data, and critical revision of the manuscript for important intellectual content. JC: acquisition data, analysis, and interpretation of data. BC: interpretation of data. GVA: study concept and design, interpretation of data, and critical revision of the manuscript for important intellectual content. MF and SV: acquisition of the data, interpretation of data, and critical revision of the manuscript for important intellectual content. JLC: study concept and design, interpretation of data, and critical revision of the manuscript for important intellectual content. CB: study concept and design, acquisition of data, analysis and interpretation of data, material support, drafting of the manuscript, and study supervision. All authors agreed with the final version of the manuscript. CB: guarantor of the manuscript.
Funding
BV is a doctoral fellow and GVA, SV, and MF are Senior Clinical Investigators of the Research Foundation Flanders (FWO), Belgium. BV has also received research grants by the Belgium Week of Gastroenterology, the Belgian IBD Research and Development (BIRD), the European Crohn's and Colitis Organization (ECCO) and the IBD Patient's Association Flanders (CCV VZW). This work was supported by a grant from the Swiss Science Research Foundation (PBLAP3-129427/1) to Clementine Perrier, a grant from the Fund for Scientific Research-Flanders (FWO-Flanders) Belgium (FWO project nr.G.0440.06), a GOA grant from the Research Council, KU Leuven (GOA/11/015) to SV, and a grant from the Agency for Innovation by Science and Technology in Flanders (IWT) to CB. CB is also supported by the Clinical Research Fund KOF (University Hospitals Leuven, Leuven, Belgium) and received previously research grants by the Belgium Week of Gastroenterology and the Flemish Organisation for Gastroenterology (VVGE).
Conflict of Interest Statement
BV received financial support for research from Pfizer; lectures fees from Abbvie, Ferring, Takeda Pharmaceuticals, Janssen and R Biopharm; consultancy fees from Janssen. GDH received consultancy fees from Centocor and Takeda. GVA received financial support for research from Abbott and Ferring Pharmaceuticals; lecture fees from Janssen, MSD and Abbott; consultancy fees from PDL BioPharma, UCB Pharma, Sanofi-Aventis, Abbott, Abbvie, Ferring, Novartis, Biogen Idec, Janssen Biologics, NovoNordisk, Zealand Pharma A/S, Millenium/Takeda, Shire, Novartis, and Bristol Mayer Squibb. MF received financial support for research from Takeda and Janssen; lecture fees from Ferring, Boehringer- Ingelheim, Chiesi, Merck Sharpe andamp; Dohme, Tillotts, Janssen Biologics, AbbvieTakeda, Mitsubishi Tanabe, Zeria; consultancy fees from Abbvie, Boehringer-Ingelheim, Ferring, Merck Sharpe andamp; Dohme, and Janssen Biologics. SV received financial support for research from MSD, Abbvie, Janssen and UCB Pharma; lecture fees from Abbott, Abbvie, Merck Sharpe andamp; Dohme, Ferring Pharmaceuticals and UCB Pharma; consultancy fees from Pfizer, Ferring Pharmaceuticals, Shire Pharmaceuticals Group, Merck Sharpe andamp; Dohme, and AstraZeneca Pharmaceuticals.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Thomas Wynn (NIAID/NIH) for the critical reflections and revision of the manuscript for important intellectual content. Vera Ballet, Eline Vandeput, Sophie Organe, Nooshin Ardeshir Davani, Helene Blevi, Tamara Coopmans, and Willem-Jan Wollants for an excellent job in maintaining the Biobank database. We would like to thank Marion Kasaian at Pfizer who provided the original breeding pairs of IL13RA2 KO mice.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2018.02983/full#supplementary-material
Supplementary Figure 1. Correlation between IL13RA2 mRNA and goblet specific cell genes. Correlations between expression of IL13RA2 with the expression of FCGBP (A) and CLCA1 (B), backbone of the colonic mucus layer.
Abbreviations
ATOH1, atonal BHLH transcription factor 1; CD, Crohn's disease; CHI3L1, chitinase 3 like 1; CLCA1, chloride channel accessory 1; DSS, dextran sodium sulfate; FCGBP, FC fragment of IgG binding protein; GAL3, galectin 3; H&E, haemotoxylin and eosin; IBD, inflammatory bowel disease; IL, interleukin; KO, knock out; RELMβ, resistin like beta; SPDEF, SAMpointed domain containing ETS transcription factor; TMEM219, transmembrane protein 219; TNF, tumor necrosis factor; UC, ulcerative colitis;WT, wild type; YKL-40, chitinase 3 like 1.
References
1. De Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. (2016) 13:13–27. doi: 10.1038/nrgastro.2015.186
2. Flamant M, Roblin X. Inflammatory bowel disease: towards a personalized medicine. Therap Adv Gastroenterol. (2018) 11:1756283X17745029. doi: 10.1177/1756283X17745029
3. Fuss IJ, Heller F, Boirivant M, Leon F, Yoshida M, Fichtner-Feigl S, et al. Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. J Clin Invest. (2004) 113:1490–7. doi: 10.1172/JCI19836
4. Peluso I, Pallone F, Monteleone G. Interleukin-12 and Th1 immune response in Crohn's disease: pathogenetic relevance and therapeutic implication. World J Gastroenterol. (2006) 12:5606–10. doi: 10.3748/wjg.v12.i35.5606
5. Verstockt B, Verstockt S, Creyns B, Tops S, Van Assche G, Gils A, et al. Mucosal IL13RA2 predicts non-response to anti-TNF therapy in Crohn's disease. Aliment Pharmacol Therapeut. (in press). doi: 10.1111/apt.15126
6. Tabata Y, Khurana Hershey GK. IL-13 receptor isoforms: breaking through the complexity. Curr Allergy Asthma Rep. (2007) 7:338–45. doi: 10.1007/s11882-007-0051-x
7. Kawakami K, Taguchi J, Murata T, Puri R. The interleukin-13 receptor alpha2 chain: an essential component for binding and internalization but not for interleukin-13-induced signal transduction through the STAT6 pathway. Blood (2001) 97:2673–9. doi: 10.1182/blood.V97.9.2673
8. Mentink-Kane MM, Cheever AW, Thompson RW, Hari DM, Kabatereine NB, Vennervald BJ, et al. IL-13 receptor alpha 2 down-modulates granulomatous inflammation and prolongs host survival in schistosomiasis. Proc Natl Acad Sci USA. (2004) 101:586–90. doi: 10.1073/pnas.0305064101
9. Wilson MS, Elnekave E, Mentink-Kane MM, Hodges MG, Pesce JT, Ramalingam TR, et al. IL-13Ralpha2 and IL-10 coordinately suppress airway inflammation, airway-hyperreactivity, and fibrosis in mice. J Clin Invest. (2007) 117:2941–51. doi: 10.1172/JCI31546
10. Morimoto M, Zhao A, Sun R, Stiltz J, Madden KB, Mentink-Kane M, et al. IL-13 receptor alpha2 regulates the immune and functional response to Nippostrongylus brasiliensis infection. J Immunol. (2009) 183:1934–9. doi: 10.4049/jimmunol.0804299
11. O'toole M, Legault H, Ramsey R, Wynn TA, Kasaian MT. A novel and sensitive ELISA reveals that the soluble form of IL-13R-alpha2 is not expressed in plasma of healthy or asthmatic subjects. Clin Exp Allergy (2008) 38:594–601. doi: 10.1111/j.1365-2222.2007.02921.x
12. Badalyan V, Thompson R, Addo K, Borthwick LA, Fisher AJ, Ort T, et al. TNF-alpha/IL-17 synergy inhibits IL-13 bioactivity via IL-13Ralpha2 induction. J Allergy Clin Immunol. (2014) 134:975–8 e975. doi: 10.1016/j.jaci.2014.05.019
13. Chandriani S, Depianto DJ, N'diaye EN, Abbas AR, Jackman J, Bevers J, et al. Endogenously expressed IL-13Ralpha2 attenuates IL-13-mediated responses but does not activate signaling in human lung fibroblasts. J Immunol. (2014) 193:111–9. doi: 10.4049/jimmunol.1301761
14. Lee CM, He CH, Nour AM, Zhou Y, Ma B, Park JW, et al. IL-13Ralpha2 uses TMEM219 in chitinase 3-like-1-induced signalling and effector responses. Nat Commun. (2016) 7:12752. doi: 10.1038/ncomms12752
15. Perrier C, De Hertogh G, Cremer J, Vermeire S, Rutgeerts P, Van Assche G, et al. Neutralization of membrane TNF, but not soluble TNF, is crucial for the treatment of experimental colitis. Inflamm Bowel Dis. (2013) 19:246–53. doi: 10.1002/ibd.23023
16. Kioi M, Kawakami M, Shimamura T, Husain SR, Puri RK. Interleukin-13 receptor alpha2 chain: a potential biomarker and molecular target for ovarian cancer therapy. Cancer (2006) 107:1407–18. doi: 10.1002/cncr.22134
17. Arijs I, Li K, Toedter G, Quintens R, Van Lommel L, Van Steen K, et al. Mucosal gene signatures to predict response to infliximab in patients with ulcerative colitis. Gut (2009) 58:1612–9. doi: 10.1136/gut.2009.178665
18. Arijs I, Quintens R, Van Lommel L, Van Steen K, De Hertogh G, Lemaire K, et al. Predictive value of epithelial gene expression profiles for response to infliximab in Crohn's disease. Inflamm Bowel Dis. (2010) 16:2090–8. doi: 10.1002/ibd.21301
19. Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, et al. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics (2003) 4:249–64. doi: 10.1093/biostatistics/4.2.249
20. Kauffmann A, Gentleman R, Huber W. arrayQualityMetrics–a bioconductor package for quality assessment of microarray data. Bioinformatics (2009) 25:415–6. doi: 10.1093/bioinformatics/btn647
21. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics (2008) 9:559. doi: 10.1186/1471-2105-9-559
22. Birchenough GM, Johansson ME, Gustafsson JK, Bergstrom JH, Hansson GC. New developments in goblet cell mucus secretion and function. Mucosal Immunol. (2015) 8:712–9. doi: 10.1038/mi.2015.32
23. Vancamelbeke M, Vanuytsel T, Farre R, Verstockt S, Ferrante M, Van Assche G, et al. Genetic and transcriptomic bases of intestinal epithelial barrier dysfunction in inflammatory bowel disease. Inflamm Bowel Dis. (2017) 23:1718–29. doi: 10.1097/MIB.0000000000001246
24. Wood N, Whitters MJ, Jacobson BA, Witek J, Sypek JP, Kasaian M, et al. Enhanced interleukin (IL)-13 responses in mice lacking IL-13 receptor alpha 2. J Exp Med. (2003) 197:703–9. doi: 10.1084/jem.20020906
25. Wilson MS, Ramalingam TR, Rivollier A, Shenderov K, Mentink-Kane MM, Madala SK, et al. Colitis and intestinal inflammation in IL10−−/−− mice results from IL-13Ralpha2-mediated attenuation of IL-13 activity. Gastroenterology (2011) 140:254–64. doi: 10.1053/j.gastro.2010.09.047
26. Newcomb DC, Zhou W, Moore ML, Goleniewska K, Hershey GK, Kolls JK, et al. A functional IL-13 receptor is expressed on polarized murine CD4+ Th17 cells and IL-13 signaling attenuates Th17 cytokine production. J Immunol. (2009) 182:5317–21. doi: 10.4049/jimmunol.0803868
27. Izcue A, Coombes JL, Powrie F. Regulatory lymphocytes and intestinal inflammation. Annu Rev Immunol. (2009) 27:313–38. doi: 10.1146/annurev.immunol.021908.132657
28. Verstockt B, Smith KG, Lee JC. Genome-wide association studies in Crohn's disease: past, present and future. Clin Transl Immunol. (2018) 7:e1001. doi: 10.1002/cti2.1001
29. Booth BW, Sandifer T, Martin EL, Martin LD. IL-13-induced proliferation of airway epithelial cells: mediation by intracellular growth factor mobilization and ADAM17. Respir Res. (2007) 8:51. doi: 10.1186/1465-9921-8-51
30. Semlali A, Jacques E, Koussih L, Gounni AS, Chakir J. Thymic stromal lymphopoietin-induced human asthmatic airway epithelial cell proliferation through an IL-13-dependent pathway. J Allergy Clin Immunol. (2010) 125:844–50. doi: 10.1016/j.jaci.2010.01.044
31. Taniguchi K, Yamamoto S, Aoki S, Toda S, Izuhara K, Hamasaki Y. Epigen is induced during the interleukin-13-stimulated cell proliferation in murine primary airway epithelial cells. Exp Lung Res. (2011) 37:461–70. doi: 10.3109/01902148.2011.596894
32. Singhera GK, Macredmond R, Dorscheid DR. Interleukin-9 and−13 inhibit spontaneous and corticosteroid induced apoptosis of normal airway epithelial cells. Exp Lung Res. (2008) 34:579–98. doi: 10.1080/01902140802369372
33. Laoukili J, Perret E, Willems T, Minty A, Parthoens E, Houcine O, et al. IL-13 alters mucociliary differentiation and ciliary beating of human respiratory epithelial cells. J Clin Invest. (2001) 108:1817–24. doi: 10.1172/JCI200113557
34. Kondo M, Tamaoki J, Takeyama K, Nakata J, Nagai A. Interleukin-13 induces goblet cell differentiation in primary cell culture from Guinea pig tracheal epithelium. Am J Respir Cell Mol Biol. (2002) 27:536–41. doi: 10.1165/rcmb.4682
35. Wu D, Ahrens R, Osterfeld H, Noah TK, Groschwitz K, Foster PS, et al. Interleukin-13 (IL-13)/IL-13 receptor alpha1 (IL-13Ralpha1) signaling regulates intestinal epithelial cystic fibrosis transmembrane conductance regulator channel-dependent Cl-secretion. J Biol Chem. (2011) 286:13357–69. doi: 10.1074/jbc.M110.214965
36. Hsi LC, Kundu S, Palomo J, Xu B, Ficco R, Vogelbaum MA, et al. Silencing IL-13Ralpha2 promotes glioblastoma cell death via endogenous signaling. Mol Cancer Ther. (2011) 10:1149–60. doi: 10.1158/1535-7163.MCT-10-1064
37. Papageorgis P, Ozturk S, Lambert AW, Neophytou CM, Tzatsos A, Wong CK, et al. Targeting IL13Ralpha2 activates STAT6-TP63 pathway to suppress breast cancer lung metastasis. Breast Cancer Res. (2015) 17:98. doi: 10.1186/s13058-015-0607-y
38. Shibasaki N, Yamasaki T, Kanno T, Arakaki R, Sakamoto H, Utsunomiya N, et al. Role of IL13RA2 in Sunitinib resistance in clear cell renal cell carcinoma. PLoS ONE (2015) 10:e0130980. doi: 10.1371/journal.pone.0130980
39. Khan WI, Blennerhasset P, Ma C, Matthaei KI, Collins SM. Stat6 dependent goblet cell hyperplasia during intestinal nematode infection. Parasite Immunol. (2001) 23:39–42. doi: 10.1046/j.1365-3024.2001.00353.x
40. Cosin-Roger J, Ortiz-Masia D, Calatayud S, Hernandez C, Esplugues JV, Barrachina MD. The activation of Wnt signaling by a STAT6-dependent macrophage phenotype promotes mucosal repair in murine IBD. Mucosal Immunol. (2016) 9:986–98. doi: 10.1038/mi.2015.123
41. Oh CK, Geba GP, Molfino N. Investigational therapeutics targeting the IL-4/IL-13/STAT-6 pathway for the treatment of asthma. Eur Respir Rev. (2010) 19:46–54. doi: 10.1183/09059180.00007609
42. Finkelman FD, Shea-Donohue T, Morris SC, Gildea L, Strait R, Madden KB, et al. Interleukin-4- and interleukin-13-mediated host protection against intestinal nematode parasites. Immunol Rev. (2004) 201:139–55. doi: 10.1111/j.0105-2896.2004.00192.x
43. Inagaki-Ohara K, Sakamoto Y, Dohi T, Smith AL. gammadelta T cells play a protective role during infection with Nippostrongylus brasiliensis by promoting goblet cell function in the small intestine. Immunology (2011) 134:448–58. doi: 10.1111/j.1365-2567.2011.03503.x
44. He CH, Lee CG, Dela Cruz CS, Lee CM, Zhou Y, Ahangari F, et al. Chitinase 3-like 1 regulates cellular and tissue responses via IL-13 receptor alpha2. Cell Rep. (2013) 4:830–41. doi: 10.1016/j.celrep.2013.07.032
45. Zhou Y, He CH, Yang DS, Nguyen T, Cao Y, Kamle S, et al. Galectin-3 Interacts with the CHI3L1 Axis and Contributes to Hermansky-Pudlak Syndrome Lung Disease. J Immunol. (2018) 200:2140–53. doi: 10.4049/jimmunol.1701442
46. Dela Cruz CS, Liu W, He CH, Jacoby A, Gornitzky A, Ma B, et al. Chitinase 3-like-1 promotes Streptococcus pneumoniae killing and augments host tolerance to lung antibacterial responses. Cell Host Microbe (2012) 12:34–46. doi: 10.1016/j.chom.2012.05.017
Keywords: IL13RA2, goblet cells, anti-TNF non-responsiveness, IBD, inflammatory bowel disease, infliximab, microarray
Citation: Verstockt B, Perrier C, De Hertogh G, Cremer J, Creyns B, Van Assche G, Ferrante M, Ceuppens JL, Vermeire S and Breynaert C (2018) Effects of Epithelial IL-13Rα2 Expression in Inflammatory Bowel Disease. Front. Immunol. 9:2983. doi: 10.3389/fimmu.2018.02983
Received: 24 October 2018; Accepted: 04 December 2018;
Published: 18 December 2018.
Edited by:
Eric Cox, Ghent University, BelgiumReviewed by:
Britta Siegmund, Charité Universitätsmedizin Berlin, GermanyHiroshi Nakase, Sapporo Medical University, Japan
Copyright © 2018 Verstockt, Perrier, De Hertogh, Cremer, Creyns, Van Assche, Ferrante, Ceuppens, Vermeire and Breynaert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christine Breynaert, christine.breynaert@uzleuven.be
†These authors share first authorship