 Juliana Agudelo1†
Juliana Agudelo1† Miguel Toro1†
Miguel Toro1† Luis Felipe Ramírez-Sánchez1
Luis Felipe Ramírez-Sánchez1 Luisa M. Barrientos1
Luisa M. Barrientos1 Catalina Alfonso-Parra1,2
Catalina Alfonso-Parra1,2 Frank W. Avila1*
Frank W. Avila1*- 1Max Planck Tandem Group in Mosquito Reproductive Biology, Universidad de Antioquia, Medellín, Colombia
- 2Instituto Colombiano de Medicina Tropical, Universidad CES, Sabaneta, Colombia
In insect vectors of disease, male and female molecules that mediate reproductive processes are promising targets to suppress fertility of these populations. One process, the storage of sperm in the female reproductive tract, is essential for optimal fertility in all organisms examined to date. In the dengue vector mosquito Aedes aegypti, female sperm storage has not been fully characterized, a requirement to identify sex-specific molecules that mediate this process. Aedes aegypti males deposit the ejaculate into the bursa of the female reproductive tract, and sperm enter the spermathecae—the long-term storage sites—quickly after insemination. However, the proportion of sperm received during mating that are stored in the spermathecae is unclear, and the fate of non-stored sperm unknown. We quantified sperm storage in two Ae. aegypti strains, mated in all combinations, and in two contexts (mass mated and when mating was observed) at 1-, 3- and 5-days post-mating. Sperm quantity in the spermathecae was similar at all timepoints; most females stored ~400 sperm on average. Sperm that did not enter the spermathecae remained in the bursa, where they declined in number and became more fragile to mechanical manipulation at each timepoint. Further, sperm viability in the bursa fell from 91.6% shortly after mating to 12.2% 24 h later. One day after insemination, ~50% of sperm detected in the female reproductive tract was stored in the spermathecae. When we quantified sperm storage in females mated to males that transferred reduced ejaculate quantities (but still able to induce optimal fertility in their mates), sperm detected in the spermathecae similarly declined; females stored ~50% of the sperm received even as sperm quantities transferred at mating declined. Our results suggest that sperm storage in Ae. aegypti females is influenced by ejaculate volume, and that sperm that do not enter the spermathecae remain in the bursa, where they appear to degrade. The consistent presence of sperm in the bursa, even when males transferred low sperm quantities, suggests that the putative degradation of bursa sperm may play a role in Ae. aegypti female fertility, potentially identifying a novel process in this important vector species.
Introduction
Sperm storage in the female reproductive tract (RT) is a common process in animals with internal fertilization (1, 2). In female insects, sperm received at mating are stored in specialized structures of the RT called spermathecae (3), which allows insemination and fertilization to be uncoupled, permitting the continual fertilization of eggs in the absence of additional mating opportunities. Sperm that are transferred to females during mating need to travel to and enter the sites of storage, be maintained viably within storage, and be efficiently released to fertilize eggs (4). Disrupting any of these steps impacts fertility (5, 6), highlighting the essential nature of sperm storage in reproduction. In the extensively studied insect model Drosophila melanogaster, male seminal fluid proteins (SFPs), transferred to females along with sperm during mating, are essential in mediating sperm storage in mated females (7–14). Female genes expressed from the spermathecae are also required for sperm entry into storage and sustained sperm motility there (15, 16).
Aedes aegypti mosquitoes are the primary vectors of the dengue (17), Zika (18), and yellow fever (19) viruses, among others. This invasive species has colonized large portions of the tropics and sub-tropics (20), and its territory is predicted to expand with increasing global temperatures (21). Characterizing reproductive processes important for fertility in Ae. aegypti, including sperm storage, will identify novel targets to control this vector species, potentially creating alternatives to insecticide use, or contemporary control strategies that release of genetically modified (22) or Wolbachia infected (23) Ae. aegypti adults into the field.
To date, 280 Ae. aegypti SFPs (24, 25) and hundreds of mating responsive genes in female RT tissues have been identified (26), including from the spermathecae (27, 28). However, identifying the sex-specific molecules necessary for sperm storage in Ae. aegypti requires a better understanding of this physiological process. To date, most studies have focused on factors that influence sperm quantities in the Ae. aegypti male RT (29, 30) or that influence sperm quantities transferred during mating (31). Aedes aegypti males deposit the ejaculate into the bursa of the female RT (also called the bursa copulatrix or bursa inseminalis), and sperm begin to enter the spermathecae seconds after insemination (reviewed in 32), typically filling 2.5 of the three spermathecae (on average) with no effect of male or female size influencing this number (33). About 1000-2000 sperm are transferred to females during mating, with factors such as male size and age influencing this quantity (31). However, fundamental questions regarding sperm storage in mated Ae. aegypti females remain unanswered. For example, the number of transferred sperm that are stored by Ae. aegypti females is not clear. De Jesus and Reiskind (34) reported that mass mated Ae. aegypti females store ~800 sperm in the spermathecae. However, they detected similar sperm quantities in females before and after each of 5 oviposition cycles, when sperm numbers should decline due to their release to fertilize eggs.
In Ae. aegypti, the fate of sperm that do not enter the storage organs is also unknown. Some female insects derive nutrients from non-stored sperm and/or other seminal components that enhance their fertility. For example, Lepidopteran females obtain nutrients from seminal components that aid in egg production (35), with the bursa copulatrix expressing digestion-related genes thought to aid this process (36). The bursa of Tettigoniidae bush crickets also has a digestive function, and females reabsorb sperm and other seminal components that remain there (37). In Drosophila mojavensis, the ejaculate solidifies within the female RT and contributes nutrients to developing oocytes (38). In Ae. albopictus, non-stored sperm and other seminal components become a solidified mass in the bursa, which decreases in size and is nearly absent by 48 h (39). Other female insects such as D. melanogaster physically expel non-stored sperm a few hours after mating (40, 41). The Ae. aegypti bursa has no associated musculature (42), so the expulsion of non-stored sperm is unlikely to occur. Jones and Wheeler (43) reported that nearly all Ae. aegypti females had empty bursa by 48 h post-mating. However, Spielman (42) noted that an unspecified amount of sperm remained in the bursa, and that their motility ceased by 2 d post-mating, speculating that females digest sperm that remain there.
In this study, we assessed the quantity of sperm transferred by Ae. aegypti males that females ultimately store and examined the fate of sperm that do not enter the spermathecae. To do so, we quantified sperm in the bursa and spermathecae at 1-, 3- and 5-days after insemination. Further, we performed our assays in two contexts: 1) when females were mass mated and 2) when mating was observed. Nearly 25% of Ae. aegypti females re-mate when given the opportunity (30, 44), suggesting that a significant proportion of mass mated females are likely to re-mate; the potential effect of re-insemination on sperm storage is unknown. To identify possible strain-dependent effects, we used two Ae. aegypti strains in our assays—Thai and DsRed—mated in all combinations. Next, we asked if total sperm stored was related to the quantity received at insemination, or whether females maximized sperm storage levels when males transferred reduced quantities. We used consecutive mating assays, mating a single male with five consecutive females, to reduce ejaculate volume (45), which allowed us to determine 1) the quantity of sperm transferred by the male after an initial mating and each subsequent mating, and 2) the quantity of sperm stored by the female after an initial mating and each successive mating. The first three mated females are equally fertile in this assay (46), suggesting SFP transfer is sufficient to induce optimal fertility in female Ae. aegypti, as SFP receipt is required for optimal fecundity in this species (47).
This study further elucidates the process of sperm storage in Ae. aegypti females by determining sperm quantities stored by females under optimal conditions and examines characteristics that influence this process. As sperm storage in insects is typically a multi-step process regulated by male- and female-derived molecules (5, 6, 15, 48), our results are likely to highlight areas where male SFPs, females-derived proteins, or both, potentially exert their effects to ensure the successful storage of sperm and subsequent female fertility in Ae. aegypti vector mosquitoes.
Materials and Methods
Mosquitoes
We used Thai and DsRed strain mosquitoes in our assays. Thai strain Ae. aegypti was collected in Bangkok, Thailand and has been maintained in colony since 2009. The DsRed strain contains a transgene that labels sperm with the red fluorescent protein DsRed and was created using Ae. aegypti collected in Riverside, California, USA (49). Mosquito eggs were hatched under vacuum pressure (-50 kPa) and larvae reared at a density of 200/L in type II H2O supplemented with four large (7.2-8.2 mm) Hikari Gold Cichlid food pellets (Hikari, Himeju, Japan). This feeding regimen produces adults of similar size (30, 46). Pupae were transferred to 5 ml tubes to ensure virginity, and resulting adults were separated into sex-specific cages upon eclosion. Larvae and adults were maintained in an incubator at 27°C, 70% relative humidity and a 12 h:12 h light:dark cycle. Adults had constant access to 10% sucrose. Four-to-six-day-old adults were used in all experiments. As adult size influences sperm transfer during mating (29, 31) and sperm uptake into the spermathecae (34), we estimated body size of a subset of males/females used in our assays by measuring wing lengths as in (50). Sizes of the adults used in our sperm storage assays are shown in (Table S1).
Mating Assays
Mass mating and observed mating. For mass mating, we mated females in a 2:1 male:female ratio for 24 h in an 8 L cage (30 females per cage), after which males were removed. For observed mating, a single virgin male and female were placed into an 8 L cage until a copulation occurred, defined as genitalia engagement of ≥10 s (45, 46). Adults from the same hatch were used in both mating contexts (i.e., mass mated and observed), and all mating combinations were conducted on the same day. At each post-mating timepoint (1-, 3- and 5-d), a subset of females was knocked down on ice and stored at -80°C until sperm quantification commenced.
Consecutive mating assays. Consecutive mating assays were performed as in (46). Separate assays were performed to 1) directly quantify sperm transfer to females after each mating event, and 2) to quantify sperm in the spermathecae and bursa of each female of the assay (i.e., the first mated female, second mated female, etc.). To directly quantify sperm transfer, mated females were flash frozen on dry ice shortly after uncoupling (~30 s). To assess sperm storage in each female of the assay, mated females were placed in an incubator for 24 h before being knocked down on ice. All specimens were stored at -80°C until sperm quantification commenced.
Detection of DsRed Sperm in the Female Reproductive Tract
To assess the presence of sperm in the bursa of Thai females mated to DsRed males, we mass mated females and dissected their reproductive tracts at 1-, 5-, and 7-d post-mating to detect fluorescence from DsRed sperm (49). At each timepoint, females were knocked down on ice and stored at -80°C until dissection; freezing tissue does not affect fluorescence from DsRed sperm (30, 46). We also examined the presence of fluorescence from DsRed sperm prior to the first, and after two oviposition cycles (5-, 7- and 13-d post-insemination, respectively). Thai females were mass mated to DsRed males and blood-fed 24 h later on the arm of a volunteer. Blood feeding on human subjects was approved by the Comité de Bioética Sede de Investigación Universitaria (Universidad de Antioquia) and all volunteers signed a consent form. Four days after blood-feeding, females were individually placed into 50 ml conical tubes that contained a 13 cm x 4 cm paper towel strip and 6.5 ml of type II H2O. After 48 h, females were again blood-fed to allow for a second egg-laying cycle. Immediately prior to the first, and after each oviposition cycle, a subset of females was knocked down on ice and stored at -80°C until dissection. The lower female reproductive tract (bursa, spermathecae, and common oviduct) was dissected in 1X PBS, and the presence/absence of fluorescent sperm in the bursa was determined using a Nikon Eclipse Ti-U fluorescent microscope (Nikon Instruments Inc., Tokyo, Japan).
Sperm Quantification
Sperm were quantified using a modified protocol from (29). In females from mass mated, observed mated, and consecutive mating assays (24 h), bursa and spermathecae were dissected separately. In assays to determine total sperm transferred by males, the bursa and spermathecae were dissected together. Dissected tissue was placed into a 250 μl chamber containing 100 μl of 1X PBS, ruptured with minutiae pins to release sperm, and vigorously mixed by pipetting up and down. An additional 100 μl of PBS was added and the solution re-mixed. Ten 5 μl aliquots of the sperm mixture were placed onto a glass slide and dried at 50°C for 5 min. Sperm were fixed in 70% ethanol and stained with Giemsa dye (Merck, Kenilworth, USA). Sperm heads in each drop were counted under brightfield illumination at 200X magnification using a Nikon Eclipse Ti-U microscope (Nikon Instruments Inc., Tokyo, Japan). This subsample was used to calculate total sperm.
Sperm Head Measurement
Sperm heads were measured at 200X magnification using the “multi-line” function in the NIS-Elements imaging software package (Nikon Instruments Inc., Tokyo, Japan). At each timepoint, the heads of five sperm isolated from the bursa or spermathecae were measured from each female. Only sperm heads where the distal tip could be visualized were measured. For comparison, we measured the heads of sperm dissected from the seminal vesicles (SVs) of both Thai and DsRed males. Sperm isolated from the male SVs were processed identically as sperm isolated from the spermathecae and bursa of mated females.
Sperm Viability Assays
To determine sperm viability in the spermathecae and bursa, we mass mated Thai males and females. We assessed sperm viability in the bursa and spermathecae at 0.5 – 2 h and 24 h post-mating using the LIVE/DEAD Sperm Viability Kit (ThermoFisher Scientific, Waltham, USA). Females were knocked down on ice and the lower female RT was dissected in 1X PBS. Dissected tissue was placed in 10 μl 1X PBS with 1 μl SYBR14 (1:50 in DMSO) and 1 μl Propidium Iodide, a coverslip was applied, and sperm visualized with a Nikon Eclipse Ti-U microscope and the NIS-Elements imaging software package (Nikon Instruments Inc., Tokyo, Japan); sperm were visualized within 1-2 minutes of dissection. Sperm that stained green were considered alive, and those that stained red were considered dead. We focused on a single area where sperm (typically 15-40) were displaced from the examined tissue by the weight of the coverslip; tissues where less than 10 sperm were displaced were excluded from the analysis.
Statistical Analysis
We first determined the probability distribution that fit the sperm count data for each experiment including normal, negative binomial, and Poisson distributions. Sperm count was analyzed using a negative binomial distribution based on the Akaike information criterion (AIC), where the lowest AIC value corresponds to the best fitted distribution. The sperm counts in all experiments were analyzed using general linear models (GLMs) with a negative binomial distribution. In the mass mated and observed mated experiments, a GLM was developed for each individual cross. For sperm head length data, we developed linear models (LMs) for all experiments based on the continuous nature of the data. To evaluate differences in the proportion of live sperm between replicates, time after mating (0.5 or 24 h) and female organ (bursa or spermathecae), a GLM with a binomial distribution was developed. Post hoc Tukey tests were developed for each model when significant differences were observed. To identify differences in the number of sperm in each organ for each cross between observed mated and mass mated contexts, we used data from day 1 to develop a GLM with a negative binomial distribution.
For data from consecutive mating assays, we first analyzed if there were significant differences between replications. As both replicates behaved similarly, we used the data from both replicates in one model. We analyzed the distribution of the data using normal, negative binomial, and Poisson distributions, and used the best fitted distribution based on the AIC value described above. To analyze the proportion of sperm in the spermathecae at 24 h (in relation to total sperm present in the female reproductive tract), we used a GLM with a binomial distribution. Post hoc Tukey tests were developed for each model when significant differences were observed. R statistical software version 3.6.1 coupled with R-Studio Version 1.2.1335 was used for all analyses (51).
Results
Sperm Are Detected in the Bursa for Up to Two Weeks After Insemination
We first asked if all sperm transferred to the female entered the spermathecae, or if some remained in the bursa, as older experiments gave conflicting results: Jones and Wheeler (43) reported that bursa were empty of sperm by 2 days post-mating, while Spielman (42) reported finding ample sperm in the bursa at the same timepoint. We mass mated Thai females to DsRed males and examined the bursa at 1-, 5-, and 7-d post-mating to determine if we could visualize fluorescence from DsRed male sperm (49). We detected fluorescence from the bursa of all females examined at each timepoint (N = 8 – 10; Figure S1A). We next asked if we could detect fluorescence in the bursa of Thai females after multiple egg-laying cycles—we again mass mated Thai females to DsRed males, blood-fed females, and examined the bursa for fluorescence immediately prior to, and after each of two oviposition cycles (5-, 7-, and 13-d post-mating, respectively). We observed fluorescence from the bursa in 92% of females (N = 52) prior to laying their first clutch of eggs, in 87% (N = 54) of bursa after the first oviposition cycle, and 84% (N = 43) after the second oviposition cycle (Figure S1B). Thus, we detected fluorescence from DsRed sperm in the bursa from most females for ~2 weeks after insemination.
Sperm That Remain in the Bursa Decline in Number and Display Increased Fragility
As we were able to detect sperm in the bursa two weeks after insemination, we next determined if sperm quantity changes in the bursa or spermathecae in the immediate days after insemination. We initially used Thai strain Ae. aegypti mated in two contexts: when females were mass mated and when mating was observed. In each context, we quantified sperm in the spermathecae and bursa at 1-, 3- and 5-d post-mating. In both contexts, we observed a significant interaction between sperm quantity, organ type, and timepoint (GLM observed mated: DF = 2, θ = 2.91, SE = 0.49, P = 0.0004; mass mated: DF = 2, θ = 2.02, SE = 0.27, P = 0.0093). In the spermathecae, sperm quantity was similar at each post-mating timepoint in both the observed (Figure 1A) and mass mated groups (Figure 1B). However, a significant decline in sperm quantity in the bursa was observed by day 5 in both contexts (Figures 1A, B).
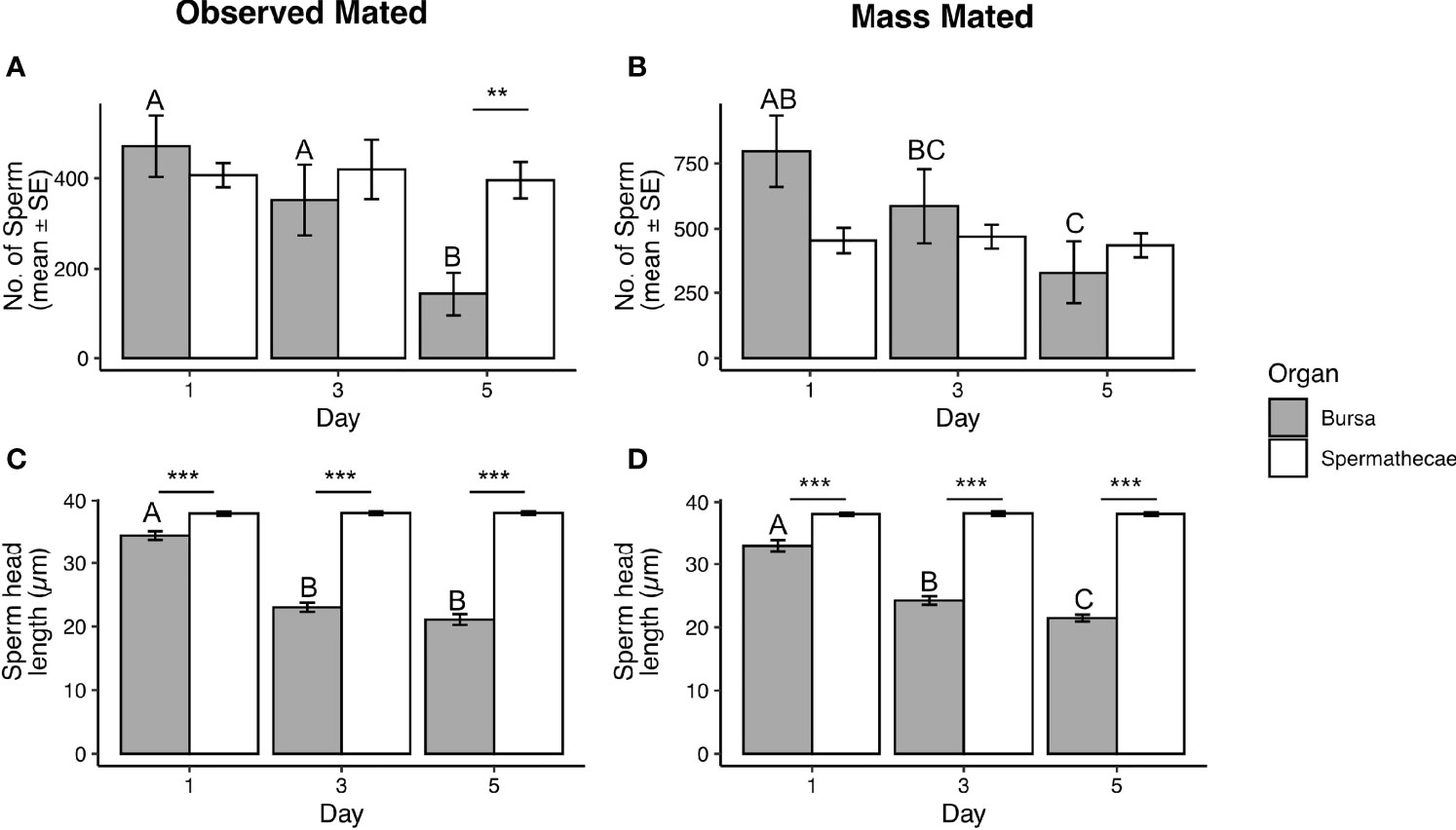
Figure 1 Total sperm detected in the spermathecae and bursa (A, B), and head lengths of sperm dissected from each organ (C, D) at 1-, 3-, and 5-d post-mating in observed mated and mass mated Thai strain females. Sample sizes in (A) N = 10 – 12; (B) N = 15 – 17; (C, D) N = 40 – 45. Different letters show significant differences (P < 0.05) for a post hoc Tukey test. **(P < 0.01), ***(P < 0.001).
The methods used to quantify sperm stain sperm nuclei (29). During processing for quantification, sperm heads often detach from tails, with sperm quantified by counting sperm heads (see Methods). We noted that sperm heads isolated from the bursa appeared to fragment into smaller pieces compared to sperm from the spermathecae (Figure S2). Therefore, we measured the lengths of sperm heads dissected from the bursa and spermathecae after processing for quantification at each time point in both mating contexts, measuring only sperm where the distal tip of the head was visible. We observed a significant interaction with organ type, timepoint, and head length in both mating contexts (LM observed mated: DF = 2, F = 117.52, P < 0.0001; mass mated: DF = 2, F = 105.26, P <0.0001). Sperm heads from the spermathecae were of similar length at each timepoint in both mating contexts (Figures 1C, D), and of similar length to sperm nuclei, which give the head its rigidity (32). However, head lengths from the bursa were present in smaller lengths at each time point in both mating contexts and were significantly shorter in size by day 3 in observed mated females (Figure 1C) and by day 5 in mass mated females (Figure 1D).
We next asked if the decline in quantity and increased fragmentation of sperm heads isolated from the bursa was specific to the Thai strain, or if these effects are also observed in another strain. Therefore, we examined sperm storage in the DsRed strain (49). As in our previous assays, we quantified sperm in the bursa and spermathecae, and measured head lengths of sperm isolated from each organ, at 1-, 3-, and 5-d post-mating in Thai and DsRed strain Ae. aegypti mated in all combinations (Female x Male: Thai x Thai, Thai x DsRed, DsRed x Thai, and DsRed x DsRed), when females were mass mated and when mating was observed.
In the observed mated group, we detected a significant interaction between sperm quantity, organ type, and timepoint in three of the four mating combinations (GLM DsRed x DsRed: DF = 2, θ = 3.08, SE = 0.48, P = 5.633e-12; DsRed x Thai: DF = 2, θ = 0.49, SE = 0.48, P < 2.2e-16; Thai x Thai: DF = 2, θ = 3.74, SE = 0.60, P = 1.387e-07); no significant interaction was detected in the Thai x DsRed combination (GLM DF = 2, θ = 2.64, SE = 0.41, P = 0.28). Sperm quantity detected in the spermathecae was similar at each timepoint (Figure 2A), but sperm quantity detected in the bursa was significantly lower by day 5 in three of the four mating combinations (Figure 2A). In the mass mated group, we observed a significant interaction between sperm quantity, organ type, and timepoint for the DsRed x Thai (GLM Df = 2, θ = 2.059, SE = 0.28, P = 0.0001) and Thai x Thai (GLM DF = 2, θ =1.84, SE = 0.25, P = 0.0048) mating combination; no significant interaction was observed in the DsRed x DsRed (GLM DF=2, θ = 3.08, SE = 0.48; P = 0.34) or Thai x DsRed (GLM DF = 2, θ = 1.87, SE = 0.25, P = 0.13) combination. Sperm quantity detected in the spermathecae was similar at each timepoint for all mating combinations (Figure 2B), but we observed a significant decline in sperm quantity by day 5 from the bursa of the DsRed x Thai and Thai x Thai groups (Figure 2B).
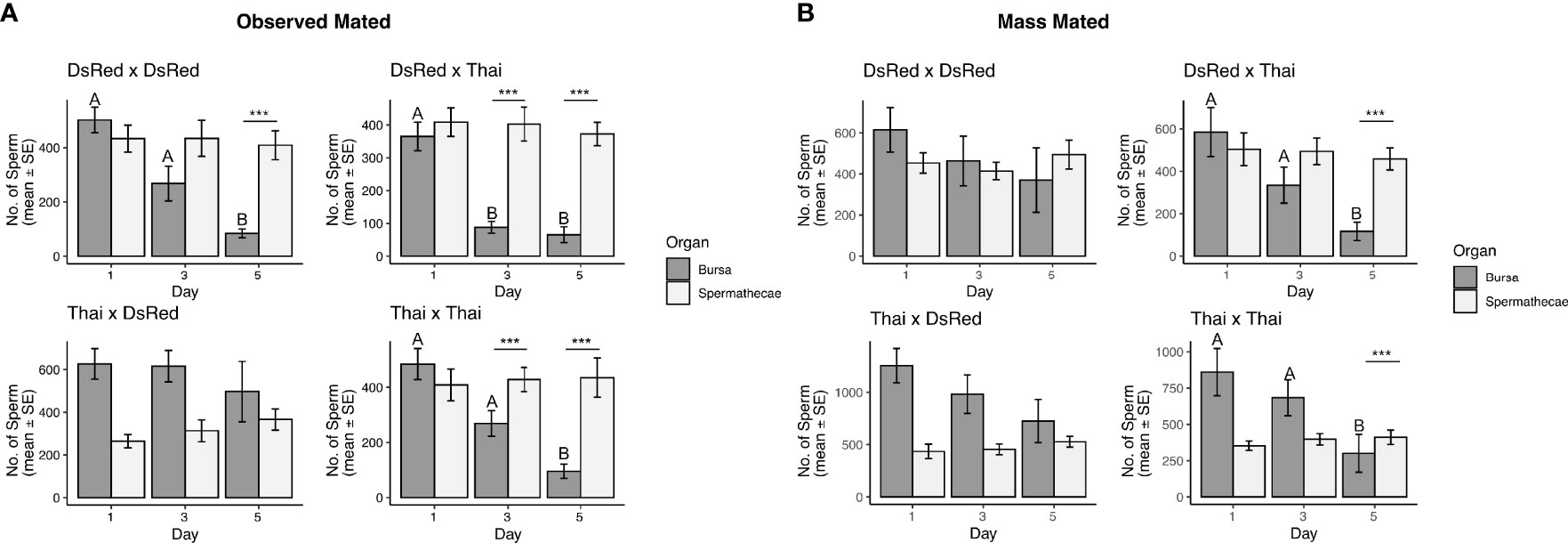
Figure 2 Total sperm detected in the spermathecae and bursa in observed mated (A) and mass mated (B) Ae. aegypti females at 1-, 3-, and 5-d post-mating. The mating combination (Female x Male) is shown above each graph. Sample sizes for each tissue and all mating combinations: (A) N = 12 – 14, (B) N = 15 – 17. Different letters show significant differences (P < 0.05) for a post hoc Tukey test. ***(P < 0.001).
When we examined the head lengths of sperm dissected from the female RT and processed for quantification, we observed a significant interaction between sperm head length, organ type, and timepoint in all mating combinations in both observed mated (LM DsRed x DsRed: DF = 2, F = 80.64, P < 2.2e-16; DsRed x Thai: DF = 2, F = 28.62, P = 3.086e-12; Thai x DsRed: DF = 2, F = 60.42, P < 2.2e-16; Thai x Thai: DF = 2, F = 104.72, P < 2.2e-16) and mass mated individuals (LM DsRed x DsRed: DF = 2, F = 71.98, P < 2.2e-16; DsRed x Thai: DF = 2, F = 58.30, P < 2.2e-16; Thai x DsRed: DF = 2, F = 49.78, P < 2.2e-16; Thai x Thai: DF = 2, F = 98.4, P < 2.2e-16). The head lengths of sperm isolated from the spermathecae were of similar size in both mating contexts at all timepoints examined (Figures 3A, B; Figure S3; Table S2). Further, head lengths of sperm from the spermathecae were similar to head lengths of sperm dissected directly from the male seminal vesicles (Table S2). In contrast, head lengths of sperm isolated from the bursa were found in significantly smaller fragments at each post-mating timepoint, in both mating contexts, and in all mating combinations (Figures 3A, B; Figure S3; Table S2).
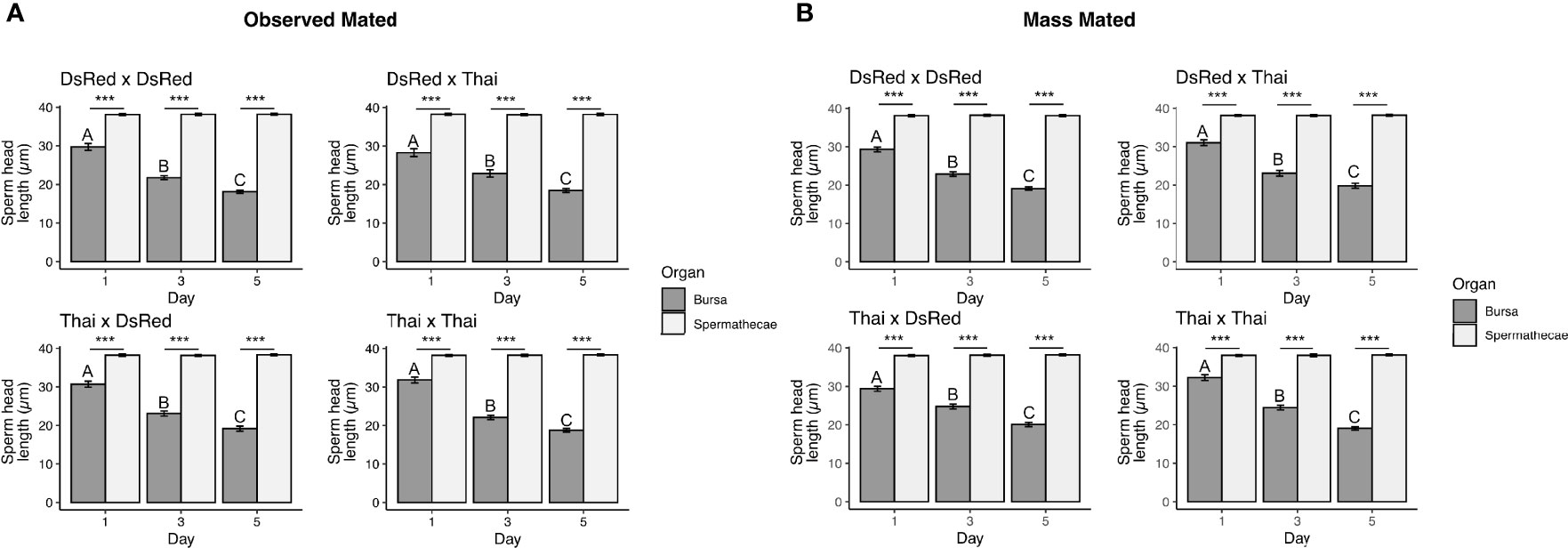
Figure 3 Head lengths of sperm dissected from the spermathecae and bursa of observed mated (A) and mass mated (B) Ae. aegypti females at 1-, 3-, and 5-d post-mating. The mating combination (Female x Male) is shown above each graph. Sample sizes for each tissue and all mating combinations: (A) N = 55 – 60, (B) N = 70 – 75. Different letters show significant differences (P < 0.05) for a post hoc Tukey test. ***(P < 0.001).
Sperm Viability in the Bursa Substantially Declines by 24 h After Insemination
The decline in sperm quantity detected in the bursa, and the increased fragmentation in sperm observed there in most females, suggested that sperm that are not stored in the spermathecae and remain in the bursa die in the days following insemination. Therefore, we examined sperm viability in bursa from mass mated Thai adults shortly after mating (0.5 – 2 h) and 24 h after mating was initiated. As a control, we also measured sperm viability from the spermathecae—where sperm are expected to maintain high viability—at the same timepoints.
We observed significant differences in the proportion of live sperm between time points for each organ (GLM SE = 0.48, Z = -7.60, P = 2.78e-14). Sperm from the spermathecae and bursa were motile at the early timepoint, and sperm viability in the spermathecae and bursa were also similar (post hoc Tukey test: P = 0.934; Table 1). At 24 h post-insemination, sperm in the spermathecae were motile but showed a marginally significant decline in viability at 24 h (post hoc Tukey test: P = 0.042; Table 1). However, sperm in the bursa from most females displayed no motility 24 h post-insemination, and the decline in sperm viability in the bursa between time points was much greater than in the spermathecae, with only 12.2% of bursa sperm viable by 24 h (post hoc Tukey test: P < 0.001; Table 1).

Table 1 Proportion of live sperm in the bursa and spermathecae at the indicated timepoint.
Mass Mated and Observed Mated Females Contain Similar Spermathecal Sperm Quantities
Our experimental design allowed us to assess if total sperm present in the spermathecae and bursa differed between mating contexts (i.e., mass mated or observed mated). A proportion of Ae. aegypti females re-mate shortly after an initial mating when given the opportunity (44). Thus, re-insemination would increase not only sperm quantity in the female RT, but also SFP concentration in the female RT; SFPs mediate sperm storage in insects (5).
We compared total sperm present in the bursa and spermathecae when females were mass mated and when mating was observed, focusing on sperm quantities 24 h after insemination due to the decline in sperm quantity in the bursa observed at day 3 and 5. We found that sperm quantities in the bursa were significantly increased in mass mated Thai females compared to observed mated females (GLM Thai x Thai: Df = 1, θ = 3.22, SE = 0.84, P = 0.009; Thai x DsRed: DF = 1, θ = 2.99, SE = 0.75, P = 0.002) but no effect was observed for DsRed females (GLM DsRed x DsRed DF = 1, θ = 4.56, SE = 1.19, P = 0.25; DsRed x Thai: DF = 1, θ = 3.24, SE = 0.83, P = 0.10). Further, we detected a small, but significant increase of sperm stored in the spermathecae in mass mated compared to observed mated females in only one mating combination: Thai x DsRed (DF = 1, θ = 4.38, SE = 1.14, P = 0.006). No significant increases in sperm stored in the spermathecae was detected in any other mating combination when females were mass mated (GLM Thai x Thai: DF = 1, θ = 9.21, SE = 2.52, P = 0.53; DsRed x Thai: DF = 1, θ = 3.61, SE 0.90, P = 0.28; DsRed x DsRed: DF = 1, θ = 5.46, SE = 1.39, P = 0.77).
Sperm Storage by Aedes aegypti Females Is Influenced by Sperm Quantity Received at an Initial Mating
In observed mated females, ~50% of the sperm detected in the RT at 24 h was stored in the spermathecae (Figures 1A, 2A). We asked whether this proportion was constant, or whether females maximize sperm storage when males transfer sperm in lower quantities. To generate males that transferred reduced sperm quantities, we utilized consecutive mating assays (46), where a single male is mated to five consecutive females. Although SFP quantity declines with each successive mating (45), sperm quantity transferred after successive mating events is unknown; 4-6 day-old Thai strain males contain ~5000 mature sperm in their seminal vesicles (30), the organ that contains mature sperm transferred in the ejaculate (52).
We observed significant differences in the number of sperm transferred and female order in the mating sequence (first mated, second mated, etc.; GLM DF = 4, θ = 1.66, SE = 0.01, P < 2.2e-16)—males transferred the largest sperm quantity at the first mating and less sperm with each successive mating (Figure 4). Although males transferred sperm in decreasing quantities, the quantity of sperm transferred in the first, second and third mating (Figure 4) often exceeded the average quantity females stored (Figures 1, 2A, B). Therefore, we assessed the quantity of sperm ultimately stored in the spermathecae and the quantity that remained in the bursa in each female of the assay 24 h after insemination. Females stored the largest amount of sperm after the initial mating, and reduced quantities with each successive mating (Figure 5A). Sperm were always observed in the bursa, even after a fifth consecutive mating (Figure 5A). Finally, we examined the proportion of sperm in the female RT detected in the spermathecae at 24 h when we successfully quantified sperm from the bursa and spermathecae from the same female. We found that the proportion of sperm detected in the spermathecae did not significantly differ in the first three mated females (Figure 5B), with ~50% of the sperm detected found in the spermathecae. However, the proportion of sperm detected in the spermathecae significantly declined in the fourth and fifth mated female, with a larger proportion of sperm remaining in the bursa (Figure 5B).
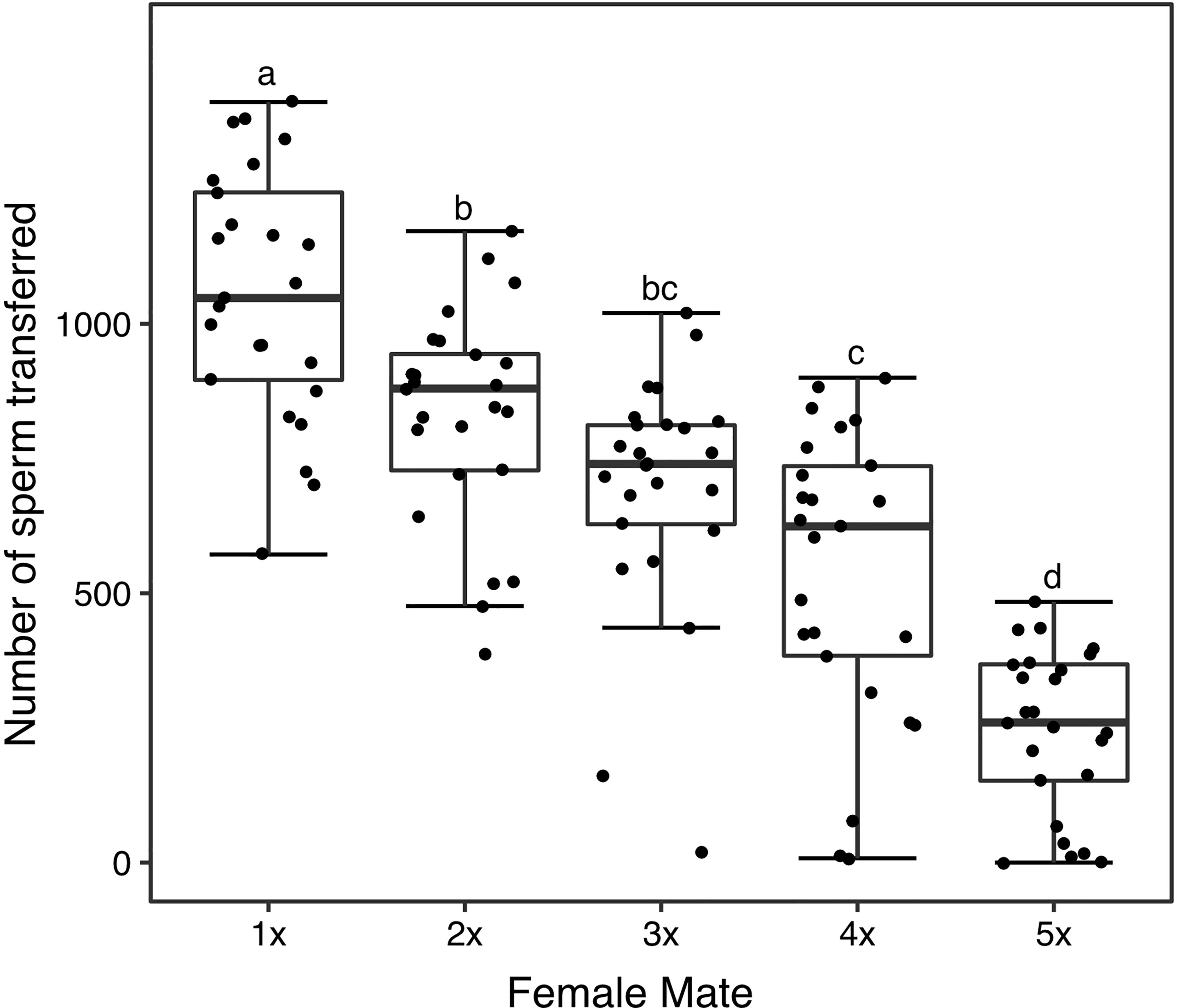
Figure 4 Sperm transfer to each female of the consecutive mating assays (1 = first mated female, 2 = second, etc.). N = 25 for each female. The middle horizontal line in the box plots represents the median, the lower and upper margins of the box the 25th and 75th quartile, and the whiskers extend to the minimum and maximum of the data (excluding outliers, shown as points outside the whiskers). Different letters show significant differences (P < 0.05) for a post hoc Tukey test.
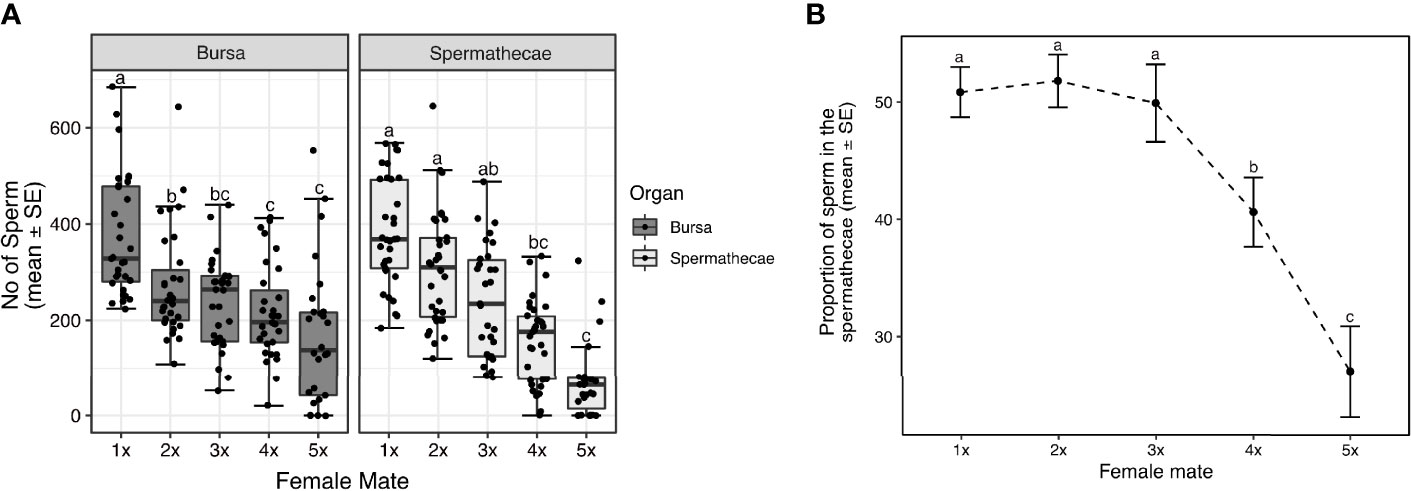
Figure 5 Sperm stored in the spermathecae, and sperm that remain in the bursa by each female of the consecutive mating assays 24 h after insemination (A). The proportion of sperm detected in the female RT that are stored in the spermathecae by each female of the assay (B). Sample sizes: (A) spermathecae: N = 27 – 34; bursa: N = 26 – 31, (B) N = 21 – 30. The middle horizontal line in the box plots represents the median, the lower and upper margins of the box the 25th and 75th quartile, and the whiskers extend to the minimum and maximum of the data (excluding outliers, shown as points outside the whiskers). Different letters show significant differences (P < 0.05) for a post hoc Tukey test.
Discussion
Sperm storage is an essential process in reproduction that requires both male- and female-specific molecules for females to efficiently store and utilize sperm (1, 5). In the insect model D. melanogaster, sex-specific molecules mediate sperm storage parameters (5, 6, 15, 16). Elimination of individual seminal proteins required for sperm storage severely impacts female fertility (7–11, 13, 14), suggesting that molecules that mediate sperm storage offer intriguing targets to suppress vector populations. Early studies showed that in female Ae. aegypti, sperm are transferred to the bursa of the female reproductive tract, where they quickly aggregate at, and orient towards, the entrance of the spermathecae (53), accumulating into the spermathecae minutes after insemination (42). However, since these studies, other questions regarding this process in Ae. aegypti have remained unanswered. In our study, we examined three such areas: 1) the quantity of transferred sperm females ultimately store, 2) the fate of non-stored sperm, and 3) the influence of ejaculate volume on sperm storage in Ae. aegypti females.
We found that although males transfer large sperm quantities during mating under optimal conditions—1000-2000 sperm (30, 31)—Ae. aegypti females only stored a proportion of this amount, with most females storing 300-500 sperm. Studies that have examined lifetime fecundity of large bodied females regularly given a blood meal show that females laid ~200 eggs on average under laboratory (34) and semi-field conditions (54). This suggests that the range of sperm stored detected in our assays is sufficient for Ae. aegypti females to maintain a high reproductive output throughout their lifetimes. As we observed that ejaculate volume influenced the number of sperm that are ultimately stored, the large sperm quantities transferred by male Ae. aegypti may be a response to this characteristic—larger sperm quantities transferred during mating would effect larger sperm quantities to accumulate into the spermathecae in their mates. Larger ejaculate volumes would also suggest larger doses of SFPs, which may better induce female sperm storage.
Sperm that did not enter the spermathecae remained in the bursa, where they became more fragile to mechanical manipulation and had mostly died there 24 h after insemination. This result aligns with that of Spielman (42), who observed non-motile sperm in the bursa 2 d after insemination, and hypothesized that sperm that remain in the bursa were degraded. Our observation of fluorescence from transgenic DsRed sperm in the bursa for two weeks likely results from the ability of the DsRed protein to maintain conformational stability under a variety of conditions (55). It is also possible that females release stored sperm into the bursa under unfavorable conditions. Over time, an increase in reactive oxygen species occurs in the male seminal vesicles (56) and female spermathecae (48) in mosquitoes, which may impact sperm function. Although “sperm dumping” is associated with re-mating in Drosophila (40, 57), it is unknown if singly mated females also dump unhealthy sperm from storage and will require further investigation.
SFPs often associate with sperm in the female RT (58, 59). It is possible that SFP-sperm interaction in the bursa sperm is necessary for some SFPs to exert their effects in the mated female—sperm stored in the spermathecae shed their outer coat within 24 h of storage, a process that is completed by ~20 h after insemination, a time that correlates with sperm ability to fertilize eggs (60). The modification of stored sperm observed in Ae. aegypti may impede sperm-SFP interaction in the spermathecae. It is also possible that females benefit from non-stored sperm. Some female insects obtain nutrients from seminal components transferred that can enhance fertility (35, 37, 38). Although mating induces the transcription of genes that code for digestion-related enzymes (e.g., proteases and lipases) in the Ae. aegypti female RT (26, 27), the influence of sperm degradation in the bursa in Ae. aegypti fertility, and the proteins responsible, remain to be determined.
Our results also suggest that males deplete SFPs more rapidly than sperm. Alfonso-Parra etal. (45) found that when males were mated to five consecutive females, SFPs were nearly depleted by the fifth mating. Ramírez-Sanchez etal. (46) similarly found that fecundity and fertility of the first three mated females does not differ, but is impacted after a third consecutive mating. Further, half of fifth mated females re-mated 7 d after the initial mating (46). Re-mating inhibition is mediated by SFP receipt in this species (61); some females re-mate shortly after an initial mating, but the proportion of re-mating females is reduced with time and refractoriness is complete by 24 h (44). Further, complete female refractoriness is observed by injection of 4% of one male equivalent (61). These results suggest that males are SFP-depleted by a fifth mating. However, we found that fourth and fifth mated females contained ample sperm quantities in their RTs, but that the proportion of sperm stored in the spermathecae, which did not differ in the first three mated females, declined significantly after a third consecutive mating. This result supports a role for Ae. aegypti SFPs in promoting sperm accumulation into the female storage organs, as is observed in other insects (5). A reduction in SFPs may also impact sperm motility, often important for sperm accumulation into storage (62); SFPs are required for sperm motility in several insects, including Culex pipiens mosquitoes (63).
Surprisingly, we found that mass mating, which leads to a large proportion of females being re-inseminated, did not have a significant impact on sperm storage. When mass mated, females are often inseminated quickly after males are introduced (64), and a subset of females are likely to re-mate; nearly 25% of Thai strain females re-mate shortly after an initial mating when given the opportunity (30, 44). In our mating assays, we found that Thai females had significantly more sperm in the bursa compared to observed mated females, suggesting some females had re-mated in this context. However, a second insemination—and second dose of SFPs—had only a small effect on the number of sperm ultimately stored, suggesting that the influence of ejaculate volume is limited, and that the spermathecae is restricted in the number of sperm it can maintain.
Conclusions
Although the importance of sperm storage has been established in a multitude of insects, this physiological process has not been fully dissected in mosquitoes. The results of this study expands upon our knowledge of sperm storage in Ae. aegypti. We show that Ae. aegypti females store only a proportion of the sperm they receive at mating, with the non-stored set remaining in the bursa where they appear to slowly degrade, effects that did not differ between geographically distinct Ae. aegypti strains. Additionally, females stored similar proportions of sperm even after mating with males that transferred sperm in decreasing quantities, leaving a similar proportion of sperm in the bursa. This suggests that the putative degradation of bursal sperm may play a role in the fertility of this mosquito vector, although more study is required to determine a female benefit of this process. Further exploration of female sperm storage parameters will continue to elucidate this reproductive process in Ae. aegypti, allowing for the identification of sex-specific molecules that mediate this process, and give us a better understanding of how male and female coordinate their activities to ensure successful reproduction in this vector species.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
FA, JA, and MT conceived and designed the experiments. FA analyzed the data and wrote the manuscript. JA, MT, LR-S, LB, CA-P, and FA performed the experiments. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the COLCIENCIAS, Universidad de Antioquia and Max Planck Society cooperation grant 566-1-2014 to FA.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank all members of the Avila laboratory for their support in the mating assays, Laura Harrington for generously providing the mosquito strains used in this study, Carolina Camargo for statistical consulting, and Ruta N Medellín for laboratory support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fitd.2022.816556/full#supplementary-material
References
1. Talevi R, Gualtieri R. Molecules Involved in Sperm-Oviduct Adhesion and Release. Theriogenology (2010) 73:796–801. doi: 10.1016/j.theriogenology.2009.07.005
3. Pascini TV, Martins GF. The Insect Spermatheca: An Overview. Zoology (Jena). (2017) 121:56–71. doi: 10.1016/j.zool.2016.12.001
4. Bloch Qazi MC, Heifetz Y, Wolfner MF. The Developments Between Gametogenesis and Fertilization: Ovulation and Female Sperm Storage in Drosophila Melanogaster. Dev Biol (2003) 256:195–211. doi: 10.1016/S0012-1606(02)00125-2
5. Avila FW, Sirot LK, LaFlamme BA, Rubinstein CD, Wolfner MF. Insect Seminal Fluid Proteins: Identification and Function. Annu Rev Entomol (2011) 56:21–40. doi: 10.1146/annurev-ento-120709-144823
6. Hopkins BR, Avila FW, Wolfner MF. “Insect Male Reproductive Glands and Their Products,”. Encyclopedia Reprod (Elsevier) (2018)6:, 137–44. doi: 10.1016/B978-0-12-809633-8.20547-5
7. Neubaum DM, Wolfner MF. Mated Drosophila Melanogaster Females Require a Seminal Fluid Protein, Acp36DE, to Store Sperm Efficiently. Genetics (1999) 153:845–57. doi: 10.1093/genetics/153.2.845
8. Bloch Qazi MC, Wolfner MF. An Early Role for the Drosophila Melanogaster Male Seminal Protein Acp36DE in Female Sperm Storage. J Exp Biol (2003) 206:3521–8. doi: 10.1242/jeb.00585
9. Ravi Ram K, Wolfner MF. Sustained Post-Mating Response in Drosophila Melanogaster Requires Multiple Seminal Fluid Proteins. PloS Genet (2007) 3:2428–38. doi: 10.1371/journal.pgen.0030238
10. Wong A, Albright SN, Giebel JD, Ravi Ram K, Ji S, Fiumera AC, et al. A Role for Acp29AB, a Predicted Seminal Fluid Lectin, in Female Sperm Storage in Drosophila Melanogaster. Genetics (2008) 180:921–31. doi: 10.1534/genetics.108.092106
11. Avila FW, Wolfner MF. Acp36DE is Required for Uterine Conformational Changes in Mated Drosophila Females. Proc Natl Acad Sci (2009) 106:15796–800. doi: 10.1073/pnas.0904029106
12. Avila FW, Wolfner MF. Cleavage of the Drosophila Seminal Protein Acp36DE in Mated Females Enhances its Sperm Storage Activity. J Insect Physiol (2017) 101:66–72. doi: 10.1016/j.jinsphys.2017.06.015
13. Avila FW, Ravi Ram K, Bloch Qazi MC, Wolfner MF. Sex Peptide is Required for the Efficient Release of Stored Sperm in Mated Drosophila Females. Genetics (2010) 186:595–600. doi: 10.1534/genetics.110.119735
14. LaFlamme BA, Ravi Ram K, Wolfner MF. The Drosophila Melanogaster Seminal Fluid Protease “Seminase” Regulates Proteolytic and Post-Mating Reproductive Processes. PloS Genet (2012) 8:e1002435. doi: 10.1371/journal.pgen.1002435
15. Schnakenberg SL, Matias WR, Siegal ML. Sperm-Storage Defects and Live Birth in Drosophila Females Lacking Spermathecal Secretory Cells. PloS Biol (2011) 9:e1001192. doi: 10.1371/journal.pbio.1001192
16. Sun J, Spradling AC. Ovulation in Drosophila is Controlled by Secretory Cells of the Female Reproductive Tract. Elife (2013) 2:1–16. doi: 10.7554/elife.00415
18. Alfonso-Parra C, Avila F. Molecular Responses to the Zika Virus in Mosquitoes. Pathogens (2018) 7:E49. doi: 10.3390/pathogens7020049
19. Chippaux J-P, Chippaux A. Yellow Fever in Africa and the Americas: A Historical and Epidemiological Perspective. J Venom. Anim. Toxins Incl. Trop Dis (2018) 24:20. doi: 10.1186/s40409-018-0162-y
20. Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, et al. The Global Distribution of the Arbovirus Vectors Aedes Aegypti and Ae. Albopictus. Elife (2015) 4:e08347. doi: 10.7554/elife.08347
21. Messina JP, Brady OJ, Golding N, Kraemer MUG, Wint GRW, Ray SE, et al. The Current and Future Global Distribution and Population at Risk of Dengue. Nat Microbiol (2019) 4:1508–15. doi: 10.1038/s41564-019-0476-8
22. Qsim M, Ashfaq UA, Yousaf MZ, Masoud MS, Rasul I, Noor N, et al. Genetically Modified Aedes Aegypti to Control Dengue: A Review. Crit Rev Eukaryot. Gene Expr. (2017) 27:331–40. doi: 10.1615/CritRevEukaryotGeneExpr.2017019937
23. O’Neill SL, Ryan PA, Turley AP, Wilson G, Retzki K, Iturbe-ormaetxe I, et al. Scaled Deployment of Wolbachia to Protect the Community From Dengue and Other Aedes Transmitted Arboviruses. Gates Open Res (2018) 2:36. doi: 10.12688/gatesopenres.12844.2
24. Sirot LK, Hardstone MC, Helinski MEH, Ribeiro JMC, Kimura M, Deewatthanawong P, et al. Towards a Semen Proteome of the Dengue Vector Mosquito: Protein Identification and Potential Functions. PloS Negl Trop Dis (2011) 5:1–11. doi: 10.1371/journal.pntd.0000989
25. Degner EC, Ahmed-Braimah YH, Borziak K, Wolfner MF, Harrington LC, Dorus S. Proteins, Transcripts, and Genetic Architecture of Seminal Fluid and Sperm in the Mosquito Aedes Aegypti. Mol Cell Proteomics (2018) 18:S6–S22. doi: 10.1074/mcp.ra118.001067
26. Alfonso-Parra C, Ahmed-Braimah YH, Degner EC, Avila FW, Villarreal SM, Pleiss JA, et al. Mating-Induced Transcriptome Changes in the Reproductive Tract of Female Aedes Aegypti. PloS Negl Trop Dis (2016) 10:e0004451. doi: 10.1371/journal.pntd.0004451
27. Camargo C, Ahmed-Braimah YH, Amaro IA, Harrington LC, Wolfner MF, Avila FW. Mating and Blood-Feeding Induce Transcriptome Changes in the Spermathecae of the Yellow Fever Mosquito Aedes Aegypti. Sci Rep (2020) 10:14899. doi: 10.1038/s41598-020-71904-z
28. Pascini TV, Ramalho-Ortigão M, Ribeiro JM, Jacobs-Lorena M, Martins GF. Transcriptional Profiling and Physiological Roles of Aedes Aegypti Spermathecal-Related Genes. BMC Genomics (2020) 21:143. doi: 10.1186/s12864-020-6543-y
29. Ponlawat A, Harrington LC. Age and Body Size Influence Male Sperm Capacity of the Dengue Vector Aedes Aegypti (Diptera: Culicidae). J Med Entomol (2007) 44:422–6. doi: 10.1093/jmedent/44.3.422
30. Agudelo J, Alfonso-Parra C, Avila FW. Male Age Influences Re-Mating Incidence and Sperm Use in Females of the Dengue Vector Aedes Aegypti. Front Physiol (2021) 12:691221. doi: 10.3389/fphys.2021.691221
31. Ponlawat A, Harrington LC. Factors Associated With Male Mating Success of the Dengue Vector Mosquito, Aedes Aegypti. Am J Trop Med Hyg (2009) 80:395–400. doi: 10.4269/ajtmh.2009.80.395
32. Degner EC, Harrington LC. A Mosquito Sperm’s Journey From Male Ejaculate to Egg: Mechanisms, Molecules, and Methods for Exploration. Mol Reprod Dev (2016) 83:897–911. doi: 10.1002/mrd.22653
33. Carrasquilla MC, Lounibos LP, Honorio NA, Murr S. Spermathecal Filling in Aedes Aegypti and Aedes Albopictus: Effects of Female and Male Body Sizes and Species. J Med Entomol (2019) 56:334–40. doi: 10.1093/jme/tjy158
34. De Jesus CE, Reiskind MH. The Importance of Male Body Size on Sperm Uptake and Usage, and Female Fecundity in Aedes Aegypti and Aedes Albopictus. Parasitol Vectors (2016) 9:447. doi: 10.1186/s13071-016-1734-8
35. Boggs CL, Gilbert LE. Male Contribution to Egg Production in Butterflies: Evidence for Transfer of Nutrients at Mating. Science (1979) 206:83–4. doi: 10.1126/science.206.4414.83
36. Meslin C, Plakke MS, Deutsch AB, Small BS, Morehouse NI, Clark NL. Digestive Organ in the Female Reproductive Tract Borrows Genes From Multiple Organ Systems to Adopt Critical Functions. Mol Biol Evol (2015) 32:1567–80. doi: 10.1093/molbev/msv048
37. Simmons L. Nuptial Feeding in Tettigoniids Male Costs and the Rates of Fecundity Increase. Behav Ecol Sociobiol (1990) 27:43–7.
38. Markow TA, Ankney PF. Drosophila Males Contribute to Oogenesis in a Multiple Mating Species. Science (1984) 224:302–3. doi: 10.1126/science.224.4646.302
39. Oliva CF, Damiens D, Vreysen MJB, Lemperière G, Gilles J. Reproductive Strategies of Aedes Albopictus (Diptera: Culicidae) and Implications for the Sterile Insect Technique. PloS One (2013) 8(11):e78884. doi: 10.1371/journal.pone.0078884
40. Manier MK, Belote JM, Berben KS, Novikov D, Stuart WT, Pitnick S. Resolving Mechanisms of Competitive Fertilization Success in Drosophila Melanogaster. Science (2010) 328:354–7. doi: 10.1126/science.1187096
41. Lee K-M, Daubnerová I, Isaac RE, Zhang C, Choi S, Chung J, et al. A Neuronal Pathway That Controls Sperm Ejection and Storage in Female Drosophila. Curr Biol (2015) 25:790–7. doi: 10.1016/j.cub.2015.01.050
42. Spielman A. The Mechanics of Copulation in Aedes Aegypti. Biol Bull (1964) 127:324–44. doi: 10.2307/1539230
43. Jones JC, Wheeler RE. Studies on Spermathecal Filling in Aedes Aegypti (Linnaeus). I. Description. Biol Bull (1965a) 129:134–50. doi: 10.2307/1539773
44. Degner EC, Harrington LC. Polyandry Depends on Postmating Time Interval in the Dengue Vector Aedes Aegypti. Am J Trop Med Hyg (2016) 94:780–5. doi: 10.4269/ajtmh.15-0893
45. Alfonso-Parra C, Avila FW, Deewatthanawong P, Sirot LK, Wolfner MF, Harrington LC. Synthesis, Depletion and Cell-Type Expression of a Protein From the Male Accessory Glands of the Dengue Vector Mosquito Aedes Aegypti. J Insect Physiol (2014) 70:117–24. doi: 10.1016/j.jinsphys.2014.07.004
46. Ramírez-Sánchez LF, Camargo C, Avila FW. Male Sexual History Influences Female Fertility and Re-Mating Incidence in the Mosquito Vector Aedes Aegypti (Diptera: Culicidae). J Insect Physiol (2020) 121:104019. doi: 10.1016/j.jinsphys.2020.104019
47. Villarreal SM, Pitcher S, Helinski MEH, Johnson L, Wolfner MF, Harrington LC. Male Contributions During Mating Increase Female Survival in the Disease Vector Mosquito Aedes Aegypti. J Insect Physiol (2018) 108:1–9. doi: 10.1016/j.jinsphys.2018.05.001
48. Shaw WR, Teodori E, Mitchell SN, Baldini F, Gabrieli P, Rogers DW, et al. Mating Activates the Heme Peroxidase HPX15 in the Sperm Storage Organ to Ensure Fertility in Anopheles Gambiae. Proc Natl Acad Sci (2014) 111:5854–9. doi: 10.1073/pnas.1401715111
49. Smith RC, Walter MF, Hice RH, O’Brochta DA, Atkinson PW. Testis-Specific Expression of the Beta2 Tubulin Promoter of Aedes Aegypti and its Application as a Genetic Sex-Separation Marker. Insect Mol Biol (2007) 16:61–71. doi: 10.1111/j.1365-2583.2006.00701.x
50. Heuvel MJ. The Effect of Rearing Temperature on the Wing Length, Thorax Length, Leg Length and Ovariole Number of the Adult Mosquito, Aedes Aegypti (L.). Trans R Entomol Soc London (1963) 115:197–216. doi: 10.1111/j.1365-2311.1963.tb00819.x
51. RStudio_Team. RStudio: Integrated Development for R (2020). Available at:(http://www.rstudio.com/).
52. Foster WA, Lea AO. Renewable Fecundity of Male Aedes Aegypti Following Replenishment of Seminal Vesicles and Accessory Glands. J Insect Physiol (1975) 21:1085–90. doi: 10.1016/0022-1910(75)90120-1
53. Jones JC, Wheeler RE. Studies on Spermathecal Filling in Aedes Aegypti (Linnaeus). I. Experimental. Biol Bull (1965b) 129:134–50. doi: 10.2307/1539773
54. Day JF, Edman JD, Scott TW. Reproductive Fitness and Survivorship of Aedes Aegypti (Diptera: Culicidae) Maintained on Blood, With Field Observations From Thailand. J Med Entomol (1994) 31:611–7. doi: 10.1093/jmedent/31.4.611
55. Verkhusha VV, Kuznetsova IM, Stepanenko OV, Zaraisky AG, Shavlovsky MM, Turoverov KK, et al. High Stability of Discosoma DsRed as Compared to Aequorea EGFP. Biochemistry (2003) 42:7879–84. doi: 10.1021/bi034555t
56. Kumari S, Tevatiya S, Rani J, Das De T, Chauhan C, Sharma P, et al. A Testis-Specific Heme Peroxidase HPX12 Regulates Male Fertility in the Mosquito Anopheles Stephensi. . Sci Rep. 12(2597) doi: 10.1038/s41598-022-06531-x
57. Snook RR, Hosken DJ. Sperm Death and Dumping in Drosophila. Nature (2004) 428:939–41. doi: 10.1038/nature02455
58. Peng J, Chen S, Büsser S, Liu H, Honegger T, Kubli E. Gradual Release of Sperm Bound Sex-Peptide Controls Female Postmating Behavior in Drosophila. Curr Biol (2005) 15:207–13. doi: 10.1016/j
59. Singh A, Buehner NA, Lin H, Baranowski KJ, Findlay GD, Wolfner MF. Long-Term Interaction Between Drosophila Sperm and Sex Peptide is Mediated by Other Seminal Proteins That Bind Only Transiently to Sperm. Insect Biochem Mol Biol (2018) 102:43–51. doi: 10.1016/j.ibmb.2018.09.004
60. Noble JM, Degner EC, Harrington LC, Kourkoutis LF. Cryo-Electron Microscopy Reveals That Sperm Modification Coincides With Female Fertility in the Mosquito Aedes Aegypti. Sci Rep (2019) 9:18537. doi: 10.1038/s41598-019-54920-6
61. Helinski MEH, Deewatthanawong P, Sirot LK, Wolfner MF, Harrington LC. Duration and Dose-Dependency of Female Sexual Receptivity Responses to Seminal Fluid Proteins in Aedes Albopictus and Ae. Aegypti Mosquitoes. J Insect Physiol (2012) 58:1307–13. doi: 10.1016/j.jinsphys.2012.07.003
62. Daigneault BW, Miller DJ. Transient Receptor Potential Polycystin-2 (TRPP2) Regulates Motility and Intracellular Calcium of Porcine Sperm. Andrologia (2021) 53:e14124. doi: 10.1111/and.14124
63. Stephens K, Cardullo RA, Thaler CD. Culex Pipiens Sperm Motility is Initiated by a Trypsin-Like Protease From Male Accessory Glands. Mol Reprod Dev (2018) 85:440–8. doi: 10.1002/mrd.22980
Keywords: Aedes aegypti, sperm storage, sperm transfer, spermathecae, bursa
Citation: Agudelo J, Toro M, Ramírez-Sánchez LF, Barrientos LM, Alfonso-Parra C and Avila FW (2022) Putative Degradation of Non-Stored Sperm in the Female Reproductive Tract of the Dengue Vector Mosquito Aedes aegypti. Front. Trop. Dis 3:816556. doi: 10.3389/fitd.2022.816556
Received: 16 November 2021; Accepted: 28 March 2022;
Published: 29 April 2022.
Edited by:
Baldwyn Torto, International Centre of Insect Physiology and Ecology (ICIPE), KenyaReviewed by:
Rajnikant Dixit, National Institute of Malaria Research (ICMR), IndiaEunice Owino, University of Nairobi, Kenya
Copyright © 2022 Agudelo, Toro, Ramírez-Sánchez, Barrientos, Alfonso-Parra and Avila. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frank W. Avila, grupotandem.mosquito@udea.edu.co
†These authors have contributed equally to this work