 Pilar Herrón1,2,3,4*
Pilar Herrón1,2,3,4* Gustavo A. Castellanos-Galindo1
Gustavo A. Castellanos-Galindo1 Moritz Stäbler1
Moritz Stäbler1 Juan Manuel Díaz5,6Matthias Wolff1,2
Juan Manuel Díaz5,6Matthias Wolff1,2- 1Leibniz Centre for Tropical Marine Research (ZMT), Bremen, Germany
- 2Faculty of Biology and Chemistry, University of Bremen, Bremen, Germany
- 3CEMarin – Corporation Center of Excellence in Marine Sciences, Bogotá, Colombia
- 4Fundación Ecomares, Cali, Colombia
- 5Department of Geography, Universidad Nacional de Colombia, Bogotá, Colombia
- 6Fundación Marviva, Bogotá, Colombia
Small-scale fisheries (SSF) remain a largely under-assessed and overlooked sector by governments and researchers, despite contributing approximately 50% to global fish landings and providing food and income for millions of people. The multi-species, multi-gear and data-poor nature of SSF makes implementation of traditional single-species management approaches – like catch-quotas or size limits – particularly challenging and insufficient. A more holistic approach is thus required, which demands assessment of ecological impacts. Here we carried out an estimation of selected ecological indicators of the impact of fisheries (mean length, maximum body size, mean trophic level, trophic and spatial guilds, threatened species and landed by-catch) based on the nominal catch of different gears in three representative SSF along the Colombian Pacific using landings data collected in multiple years (2011–2017). Results showed that taxonomic, size-based, functional and conservation features of the nominal catch vary greatly with geographical location and gear type used. Overall, handlines and longlines tend to select larger sizes and higher trophic levels than nets, but they also catch a higher proportion of intrinsically vulnerable species and species of conservation concern. This challenges the idea that more selective gears have overall lower ecological impacts. In contrast, nets target a wider size range – although focusing on small or medium sized fish – and include a higher diversity of trophic and spatial guilds, which could arguably be considered a more “balanced harvest” type of fishing that retains ecosystem structure and functionality. Bottom trawls, though, exhibited a relatively high percentage of landed by-catch, an undesirable feature for any fisheries in terms of sustainability. We propose that the assessment of a suite of ecological indicators, like those implemented here, should be included as part of periodic evaluations of multi-gear and multi-species SSF in tropical coastal areas, as a practical step toward ecosystem-based fisheries management.
Introduction
Small-scale fisheries (SSF) are widely recognized for their contribution to nearly half of global landings and for the multiple socio-economic benefits they provide to coastal communities (Andrew et al., 2007; Béné et al., 2010; FAO, 2015). However, this fisheries sector remains largely under-assessed and overlooked by governments and researchers (Salas et al., 2007, 2019; FAO, 2015; Purcell and Pomeroy, 2015). Management of SSF in tropical developing countries is generally constrained by insufficient government funding, lack of political will, open access regimes, multiple and scattered landing sites and low participation of resource users in decision making (Andrew et al., 2007; Salas et al., 2007; Kato et al., 2012). Traditional management approaches like catch-quotas and size limits for target species exhibit several practical difficulties when tried to be implemented in multi-gear and multi-species tropical SSF (Salas et al., 2007; Purcell and Pomeroy, 2015). Furthermore, the establishment of catch-quotas, one of the most common management measures, depends on reliable assessments of the target stock size and condition of main target species but these type of assessments are often hindered by low quality of the data available, high uncertainties underlying length-frequency catch data and lack of knowledge of basic growth and reproduction features of target species (Froese, 2004; Cope and Punt, 2009; Ramírez et al., 2017; Herrón et al., 2018).
In the past two decades a shift in fisheries management has been observed from a single-species approach – in which the main objective was to obtain maximum sustainable yields (MSY) of target species – to a more holistic approach that also considers the impacts of fishing at the community and ecosystem level, for which two main frameworks are commonly used: the Ecosystem Approach to Fisheries – EAF (Garcia, 2003) and the Ecosystem-Based Fisheries Management – EBFM (Pikitch et al., 2004). Both frameworks take into account the undesired effects of fishing on ecosystems due to the inherent selectivity of the fisheries for a particular size range and/or taxonomic group; these effects may include impacts on biodiversity, taxonomic composition, population abundance, size structure, trophic structure and trophic dynamics of biological communities (Pauly, 1984; Jennings and Kaiser, 1998; Arias-González et al., 2004; Pikitch et al., 2004). To detect such impacts, several ecological indicators have been proposed based on empirical or model-derived evidence of their potential to adequately inform of fishing impacts. These indicators often relate to basic ecosystem’s attributes such as: species richness and diversity, biomass, relative abundance of specific target or non-target groups, size structure, trophic level, structure and dynamics of the food web (Rochet and Trenkel, 2003; Fulton et al., 2005; Jennings, 2005; Jennings and Dulvy, 2005; Link, 2005; Shin et al., 2005). Current scientific advice for fisheries management in the European Union, for example, incorporates assessment of indicators such as: mean length of the fish community, proportion of predatory fish in the community, catch-based marine trophic index, proportion of discards in the fishery, among others [“IndiSeas” project – Coll et al. (2016)]. Other approaches to holistically assess fisheries and examine fishing impacts at the ecosystem level are mass-balanced trophic models, which require knowledge of trophic relations, as well as detailed data on diet composition and fishing effort that are not always available for coastal tropical systems [but see for example: Bacalso and Wolff (2014); Rehren et al. (2018), and Tesfaye and Wolff (2018)].
Here we examine the composition of the nominal catch of the multi-gear and multi-specific SSF of the Colombian Pacific coast to assess geographic or gear-related differences in selected indicators used as proxies of the potential ecological impacts of current fishing practices. Our analyses used a unique set of landings data from recent years (from 2011 to 2017) collected at three coastal zones of the Colombian Pacific with different environmental, socio-economic and fisheries management regimes. Finally, we discuss the potential benefit of implementing a periodic monitoring of ecological indicators to assess and manage SSF under an ecosystem-based approach.
Materials and Methods
Study Area
The Colombian Pacific coast is part of the tropical eastern Pacific region and it is located in the western side of the country bordering with Panama (7° 13′ 21″N, 77°53′25″W) and Ecuador (1° 27′ 48″N, 78°51′43″W), and stretching for approximately 1,300 km (Correa and Morton, 2010) (Figure 1). The northern coastal sub-region extends for approximately 335 km of coastline south of the Panama border and is characterized by rocky and sandy shores, and relatively small mangrove forests [ca. 50 km2, Velandia and Díaz (2016)]. This sub-region has a narrow continental shelf (1–15 km) and a low human population density [6 people∗km-2, DANE (2011)]. In contrast, the central coastal sub-region of Buenaventura, which encompasses approximately 150 km of coastline south and north of the city of Buenaventura, is dominated by mangrove forests [ca. 220 km2; Mejía-Rentería et al. (2018)], alluvial plains, river deltas and estuaries. These seascapes are also the dominant ones in the remaining Colombian Pacific southern coast up to the border with Ecuador. The Buenaventura sub-region has a wider continental shelf (32–52 km) and a higher human population density [70 people∗km-2, DANE (2011)] mainly due to the presence of the main city port of the entire Colombian Pacific (Buenaventura city).
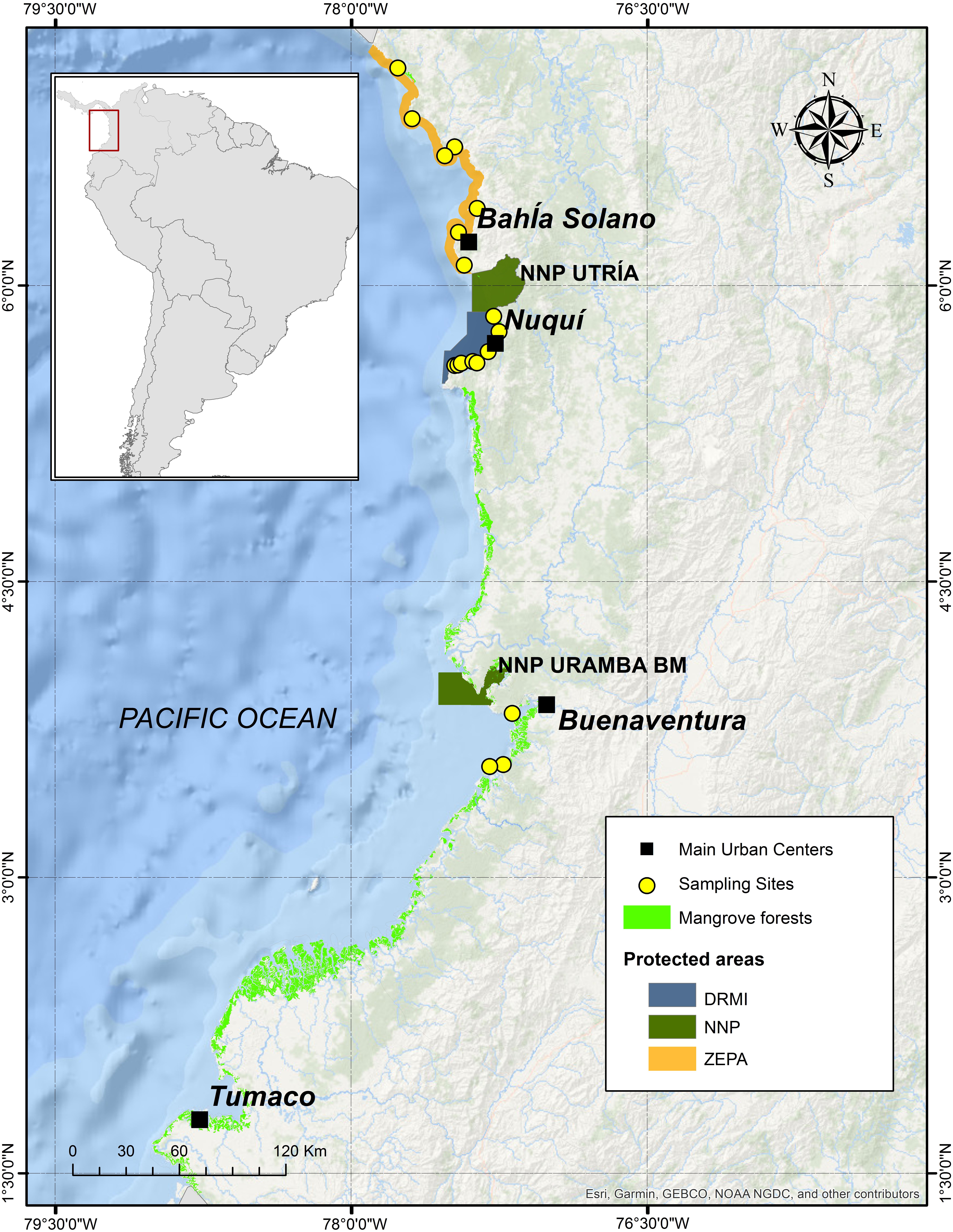
Figure 1. Colombian Pacific coast with location of the three coastal zones included in this study: ZEPA, Tribugá (DRMI + National Natural Park Utría) and Buenaventura. Location of sampled landing sites, National Natural Parks and mangrove forests within the three coastal zones are also shown.
Within the northern sub-region, there are two management areas declared in recent years: (1) an Exclusive Artisanal Fisheries Zone or ZEPA, for its Spanish acronym, and (2) a regional marine protected area (Tribugá – Integrated Regional Management District or DRMI for its Spanish acronym), declared recently by the Colombian fisheries authority and by the regional environmental authority, respectively (AUNAP, 2013; CODECHOCO, 2014) (Figure 1). These two management zones cover ca. 1,600 km2 of coastal and marine habitats (Velandia and Díaz, 2016) and complement conservation efforts by the adjacent National Natural Park Utría established in 1987 (PNN, 2006), which includes a marine area of ca. 132 km2. Current fishing practices inside the marine area of the National Park are similar to those within the DRMI (PNN, 2011) and therefore we considered the Park’s area as part of the same coastal zone, referred to hereafter as Tribugá.
Fishing Gears
At least 13 different main types of fishing gears have been reported in the Colombian Pacific SSF (Saavedra-Diaz, 2012) and eight of those are used by fishers at some or all of the three coastal zones studied here. These eight gears are: handlines, longlines (bottom), gillnets (including lobster nets), bottom trawls, purse seines, beach seines, cast nets and spear guns. Cast nets are mostly used to collect bait (such as sardines or anchovies) used in longlines or handlines and therefore the catch derived from this gear is rarely landed. Spear guns are used by a very low number of fishers while beach seines are more commonly used by family groups in the coastal communities. However, these two gears (spear guns and beach seines) contributed to <1% of the nominal catch recorded within each zone (Figure 2) and therefore were not included in further analyses. The main characteristics of the five gears that account for most of the catch are summarized in Table 1, including a sub-classification of gillnets based on the net material and on their mesh size. Given that lobster nets are a type of gillnet targeted on a specific taxonomic group (Palinuridae) and include the use of bait, we treat them here as a separate type of gear. Detailed technical specifications of these gears and how they are used in the Colombian Pacific can be found in Saavedra-Diaz (2012) and Puentes et al. (2014).
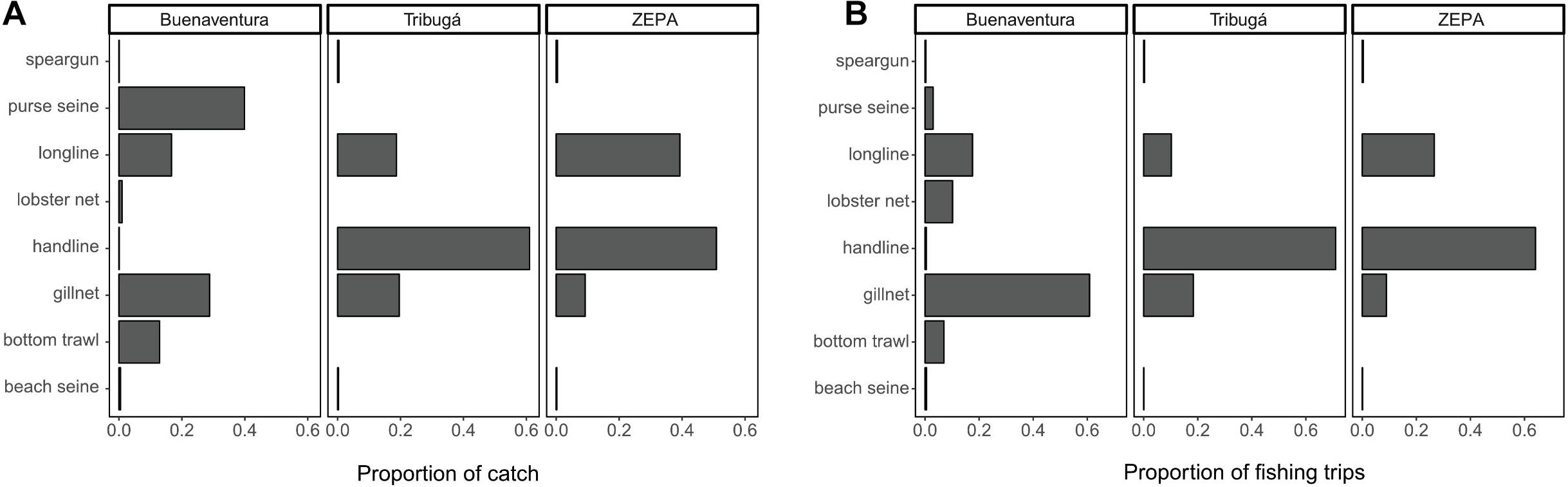
Figure 2. Relative contribution to the catch in terms of biomass (A) and to the number of sampled fishing trips (B) by each gear type used in the SSF at three coastal zones of the Colombian Pacific. Sampling periods and landing sites included in each coastal zone are described in Section “Materials and Methods.”
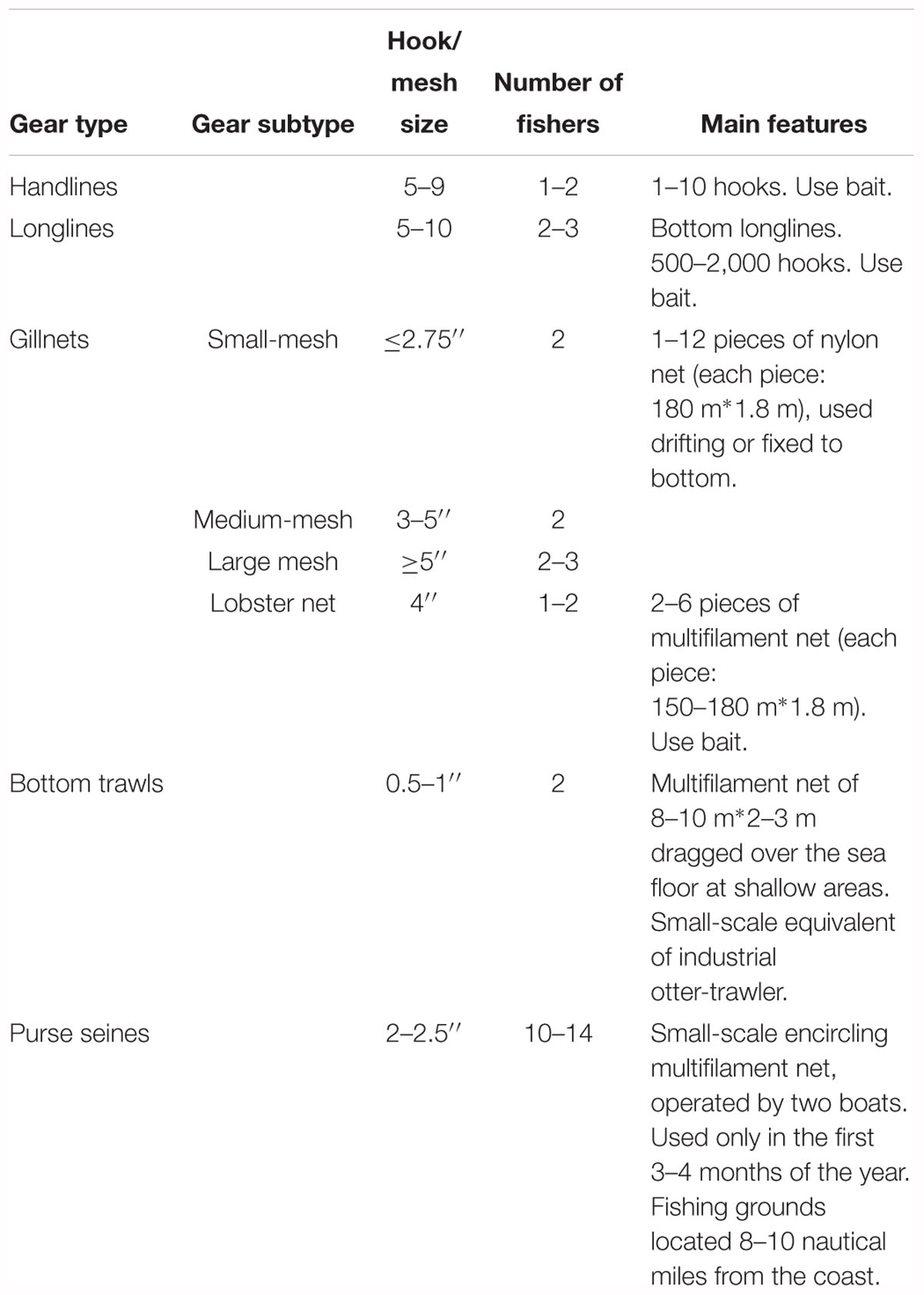
Table 1. Characteristics of gear types and subtypes that contribute most to total SSF landings at the three zones of the Colombian Pacific included in the present study.
Data Collection
In the ZEPA and Tribugá coastal zones, a community-based fisheries monitoring program was implemented from 2011 to 2016 by the regional non-government organization (NGO) MarViva Foundation1. Local observers were trained and hired to collect data at landing sites within each coastal community (Díaz et al., 2016; López-Angarita et al., 2018). Monthly visits were made by staff from the NGO to verify data quality and species identification. Data gathered through this monitoring program and used in the present study include data from nine landing sites located in ZEPA (2011–2013) and nine landing sites located in Tribugá (2011–2013 and 2016). At the Buenaventura coastal zone a similar community-based monitoring scheme was adopted by the authors of this study to collect data from August 2016 to July 2017 at three representative landing sites (Figure 1). Data gathered at landing sites included: date, common name of landed species, weight landed per taxa to the nearest 0.05 kg, catch status (e.g., whole, gutted), fishing gear type and fishing method. Also, total length of fish (or disk width in rays) and total length of invertebrates to the nearest 0.5 cm were measured in a representative sample of the catch (20–30%). All fish species were identified to the lowest taxonomic level possible following identification guides available for the region (Keen, 1971; Fisher et al., 1995; Acero, 2004; Marceniuk and Acero, 2009; Robertson and Allen, 2015).
Taking into account the collective ownership and management of the land occupied by Afro-descendent communities along the Pacific coast of Colombia (Law 70 of 1993), formal agreements with the Boards of the Community Councils Los Riscales, Los Delfines, Cupica, Juradó, Cajambre and Bazán-Bocana (in charge of the coastal areas where this study took place) were made by either MarViva Foundation or by the first author, whereby written informed consent was obtained. Additionally, meetings with fishers’ representatives (locally elected leaders of fishers associations) were held at each coastal community to explain the objectives and methods of the project prior to the beginning of field activities. Approval from an external ethical committee was not required by local legislation for research collecting fisheries data at landing sites.
Data Processing and Analyses
Considering that 80% of fish were not landed whole, but gutted (42.6%), beheaded (2.2%), gutted and beheaded (31.4%) or as trunks (3.4%), weight corrections factors based on FAO (2000) were applied to landed weight for more accurate estimates of live weight removed per taxon. For some taxonomic families of small-sized species of relatively low market value (e.g., Acanthuridae and Muraenidae) there was partial or no data available on conversion factors. We assigned a conversion factor of 1.1 to those cases, being this value the most common reported as conversion factor for gutted weight across taxa (FAO, 2000). Large sting rays (Hypanus spp.) that could not be weighted were measured and disk-widths were later converted to total weight based on literature values for the two species involved (Ehemann et al., 2017). A table with all correction factors used per taxon is included as part of the (Supplementary Table S1). Landings data converted to live weight is technically known as “nominal catch” (FAO, 2018) which does not include discarded specimens (live or dead) that are not brought to landing sites. For practical reasons we will refer here to the nominal catch as “catch.” After weight conversion was performed, as described above, relative weight per taxa (species, genus, or family) was calculated based on the catch (kg) per taxon divided by the total catch (all taxa combined) within each coastal zone.
To explore potential inter-annual differences in the catch composition of the coastal zones of Tribugá and ZEPA, we carried out cluster and non-metric multi-dimensional scaling analyses (nMDS) to compare relative weight within landing sites among years of those species that contributed to 95% of the catch at each landing site.
Size-based, functional and conservation indicators related to the composition of the catch were estimated and assessed among coastal zones and fishing gears. A list of the selected indicators is presented in Table 2, along with a brief description and the rationale behind their current global use as proxies of ecological fishing impacts. Mean total length (cm) in the catch was estimated across taxa for each gear within each coastal zone and visualized through violin plots. Maximum body size (cm), trophic level, trophic guild and spatial guild were assigned to all species registered in the landings based on data available on FishBase (Froese and Pauly, 2017), the Smithsonian Tropical Eastern Pacific Fish Guide (Robertson and Allen, 2015) and SeaLifeBase (Palomares and Pauly, 2018). Additionally, published values from local studies (Criales-Hernandez et al., 2006; Castellanos-Galindo et al., 2017) were used to assign trophic levels to some invertebrate species for which information could not be found on international databases. Trophic guilds categories used were: herbivore, invertivore, omnivore, piscivore and planktivore, while categories of spatial guilds used were: demersal, bentho-pelagic and pelagic.
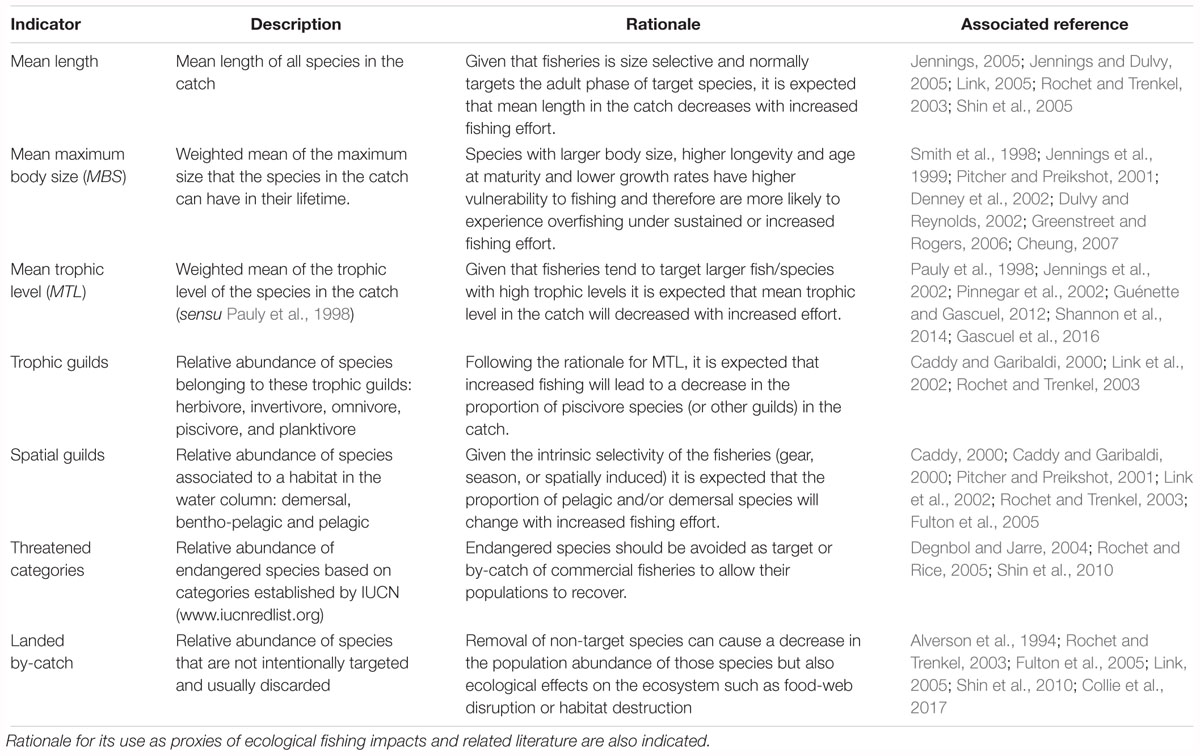
Table 2. Size-based, functional and conservation indicators used here for the assessment of the catch of SSF in the Colombian Pacific.
Conservation threat status was assigned to species based on regional and national assessments that follow the International Union for the Conservation of Nature’s (IUCN) Red List standards (IUCN, 2017). National assessments used are those carried out in Colombia in recent years for marine fish species (Chasqui et al., 2017), marine invertebrates (Ardila et al., 2002) and reptiles (Morales-Betancourt et al., 2015); the last one was included taking into account the rare occurrence of some species of sea turtles in the catch. Information on regional assessments was based on Polidoro et al. (2012). The categories used are: Not Evaluated (NE), Data Deficient (DD), Least Concern (LC), Near Threatened (NT), Vulnerable (VU), Endangered (EN), and Critically Endangered (CR). Definitions and criteria used for each category can be found at www.iucnredlist.org.
An additional classification of the taxa registered in the catch was made based on their current use or importance to fishers and markets. Three categories were considered for this purpose: “commercial,” for those species of commercial interest, “local use” for those species that are not sold to external markets but are locally consumed or used as bait, and “by-catch” for those species that are not intentionally targeted and are usually discarded before reaching the landing site. However, when the size of the individuals was not so small (approximately >25 cm) or when fishers did not carry out the sorting process of the catch while they were on-board, some of that by-catch made it to the landing site and we will refer to that portion of the catch as “landed by-catch.” In the case of bottom trawlers, fishers generally brought the last haul completely unsorted and separated from the rest of the catch, so we could use that haul to estimate landed by-catch. The classification of species in the above mentioned categories was based on Díaz et al. (2016) and on interviews made to local fishers of the coastal zone of Buenaventura by the first author (unpublished data).
Mean trophic level (MTL) of the catch for each gear category (g) at each coastal zone was estimated using the formula described by Pauly et al. (1998):
Where Wig is the biomass (total weight) of species i caught by gear g, and TLi is the trophic level of species i for n species. In a similar way we estimated mean maximum body size (MBS) per gear type at each zone, replacing TL in the previous formula for MBS. Generalized linear models (GLMs), using a logarithmic link function and a quasi-poisson distribution, were used to assess differences in mean length, MTL and mean MBS among gear types and zones. When statistical differences were detected within either factor or their interaction, pairwise comparisons were carried out using the “emmeans” R Package, based on least-square means and adjusted p-values following Tukey tests (R Core Team, 2017; Russell, 2018).
Results
A total of 40,035 one-day fishing trips were sampled accounting for 1,823.2 tons of estimated biomass in the catch and 515,243 specimens measured. The proportion of the catch contributed by each fishing gear differed among ZEPA, Tribugá and Buenaventura (Figure 2). Hook-based gears contributed the most to the catch in ZEPA and Tribugá, while net-based gears dominated in Buenaventura. The relative contribution made by each gear to the total biomass was similar to the proportion of fishing trips per gear in ZEPA and Tribugá, but not so in Buenaventura, where a very large biomass contribution was made by purse seine nets despite the relatively low number of fishing trips recorded for that gear type (Figure 2). Total biomass (kg), number and estimated percentage of fishing trips sampled per gear type at each zone are presented in Table 3.
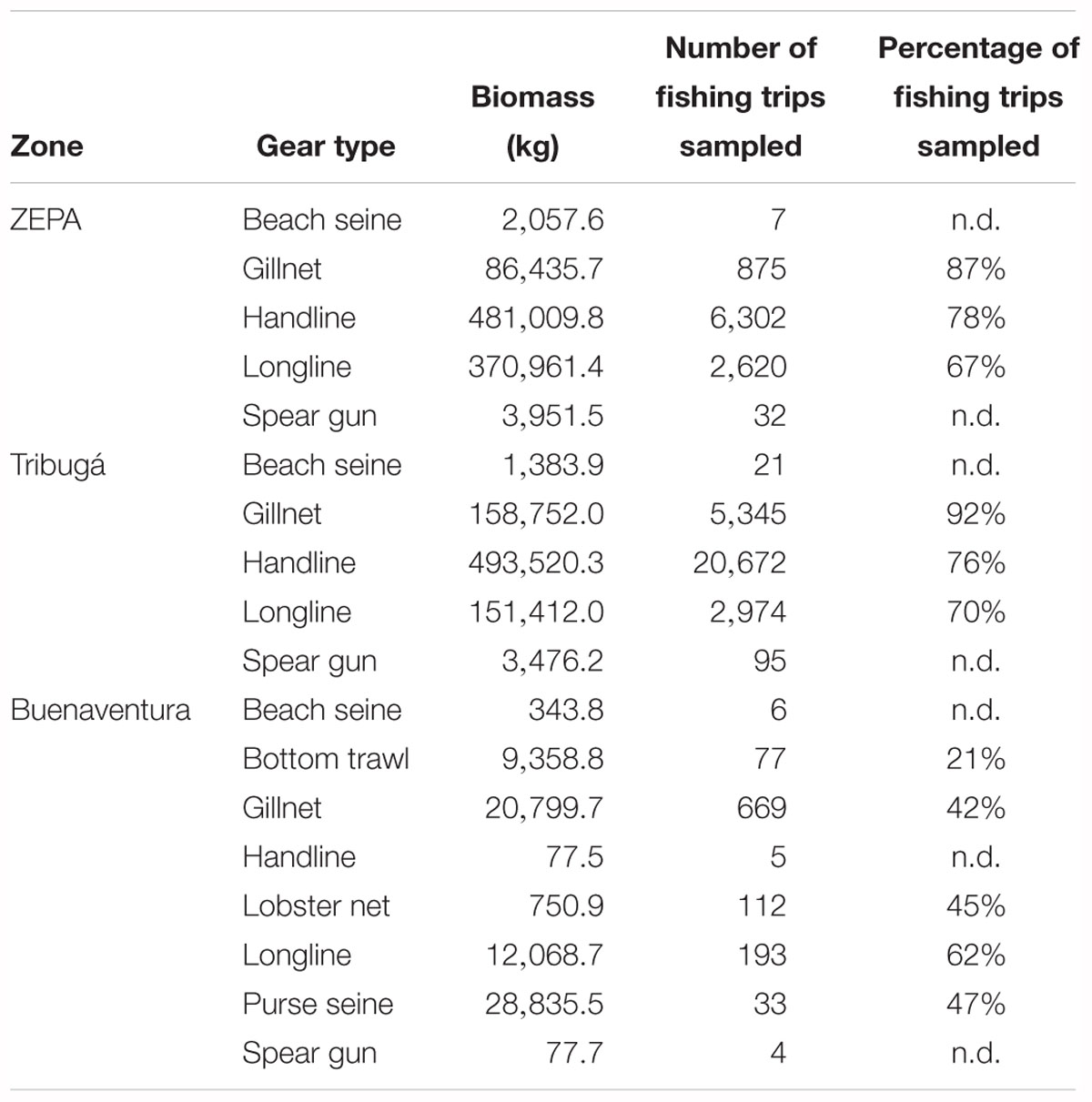
Table 3. Total biomass in the nominal catch (i.e., live weight converted from landed weight, as described in section “Materials and Methods”) of SSF, number of fishing trips sampled and estimated percentage of trips sampled per gear type at landing sites of the three coastal zones of the Colombian Pacific included in the present study (n.d., no data available on total number of trips).
Taxonomic Composition of the Catch
179+ species belonging to 80 families were identified as part of the catch of the SSF of the three coastal zones of the Colombian Pacific. However, this number of species is probably an under-estimation of the richness of the catch considering that 66 common names of mostly rare species (i.e., low relative abundance in the catch) were not assigned to any taxonomic category and 31 of them were only identified to genus or family level, resulting in a total of 276 different common names registered in the catch of the three coastal zones.
95% of the biomass in the catch was accounted for by 24 families and 72 species (Figure 3 and Supplementary Figure S1). Mackerels, tunas and bonitos (Scombridae) contributed between 20 and 30% of the annual catch at all zones, indicating an overall importance of this family in the SSF of the entire Colombian Pacific coast. Jacks (Carangidae), cusk eels (Ophidiidae), groupers (Serranidae) and snappers (Lutjanidae) were also important in the landings of ZEPA and Tribugá while in Buenaventura, catfishes (Ariidae), whiptail stingrays (Dasyatidae) and drums or croakers (Sciaenidae) followed Scombridae in the relative abundance ranking (Supplementary Figure S1). A higher number of species (41) accounted for 95% of the catch in Tribugá than in Buenaventura and ZEPA (35 species each) (Figure 3). The distribution of the relative abundance of species shows a more even pattern in the catch of Tribugá than that of ZEPA, where two dominant species (Thunnus albacares – Scombridae and Brotula clarkae – Ophidiidae) contributed to 35% of the catch. Invertebrate species, mainly shrimps (Penaeidae) and lobsters (Palinuridae), were abundant in Buenaventura, but not so in ZEPA or Tribugá. Additionally, several shark species were relatively abundant in ZEPA and Tribugá compared to Buenaventura (Figure 3). A complete list of the taxa recorded in the catch of each zone, with their absolute and relative weight, is included in the (Supplementary Table S2).
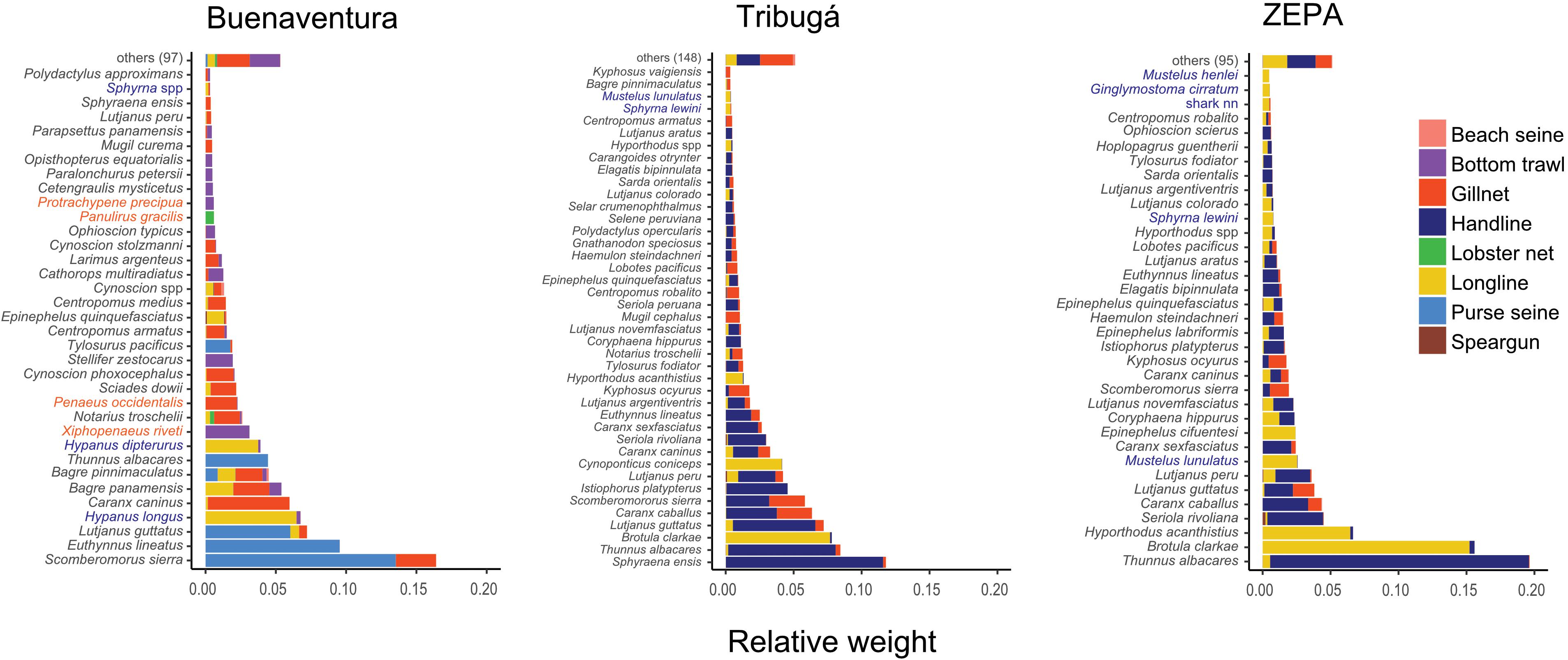
Figure 3. Relative weight of taxa in the catch of the SSF at three coastal zones of the Colombian Pacific: ZEPA, Tribugá, and Buenaventura. Taxonomic identity is indicated for those taxa that contributed 95% to total catch and relative catch contribution by each gear type is illustrated. Elasmobranch species (sharks and rays) are shown in blue font, crustaceans in orange and bony fish in black.
Results from the cluster and the nMDS analyses showed that there were not distinctive inter-annual differences related to species composition of the catch at the landing sites sampled in ZEPA and Tribugá (Supplementary Figure S2). Based on records of daily fishing activity in Tribugá and ZEPA collected by MarViva during their monitoring program (Díaz et al., 2016) and the information available from the Colombian fisheries authority2, there was no evidence of changes in fishing effort made by the small-scale fleet in those areas during the past 10 years. We thus used the combined catch data for all years of each of these coastal zones for subsequent analyses.
Size-Based Indicators of the Catch
Overall, most specimens at all zones were <100 cm of total length (Figure 4) with longlines in ZEPA capturing on average larger size classes (Table 4), even though the largest specimens were caught by handlines in Tribugá (e.g., the sailfish species Istiophorus platypterus reaching >400 cm TL; Supplementary Figure S3). In contrast, bottom trawls in Buenaventura exhibited a high relative abundance of small-sized individuals with a narrow unimodal distribution of length. The catch of this gear was composed mainly of the target small shrimps species Pacific seabob – Xiphopenaeus riveti and titi shrimp – Protrachypene precipua, and other non-target small-sized invertebrates and juvenile fish of several species (Figures 3, 4). Lobster nets and gillnets in Buenaventura had most of their catch toward the lower side of the overall length range observed in this study (Figure 4 and Table 4). Results from the GLM conducted with the entire catch (all species included) showed that mean length in the catch within the same gear type was statistically different among zones, with ZEPA showing higher mean length in the catch than Tribugá and Buenaventura for gillnets and longlines, and also for handlines when compared to Tribugá (p < 0.001 in all cases). Mean length was also statistically different among gears within the same zone: in Buenaventura, mean length of purse seines was higher than that of all other gears whereas mean length of bottom trawls was lower than all the other gears (p < 0.001 in all cases). In Tribugá and ZEPA, longlines had a significantly higher mean length in their catch compared to handlines and gillnets (p < 0.001 in all cases).
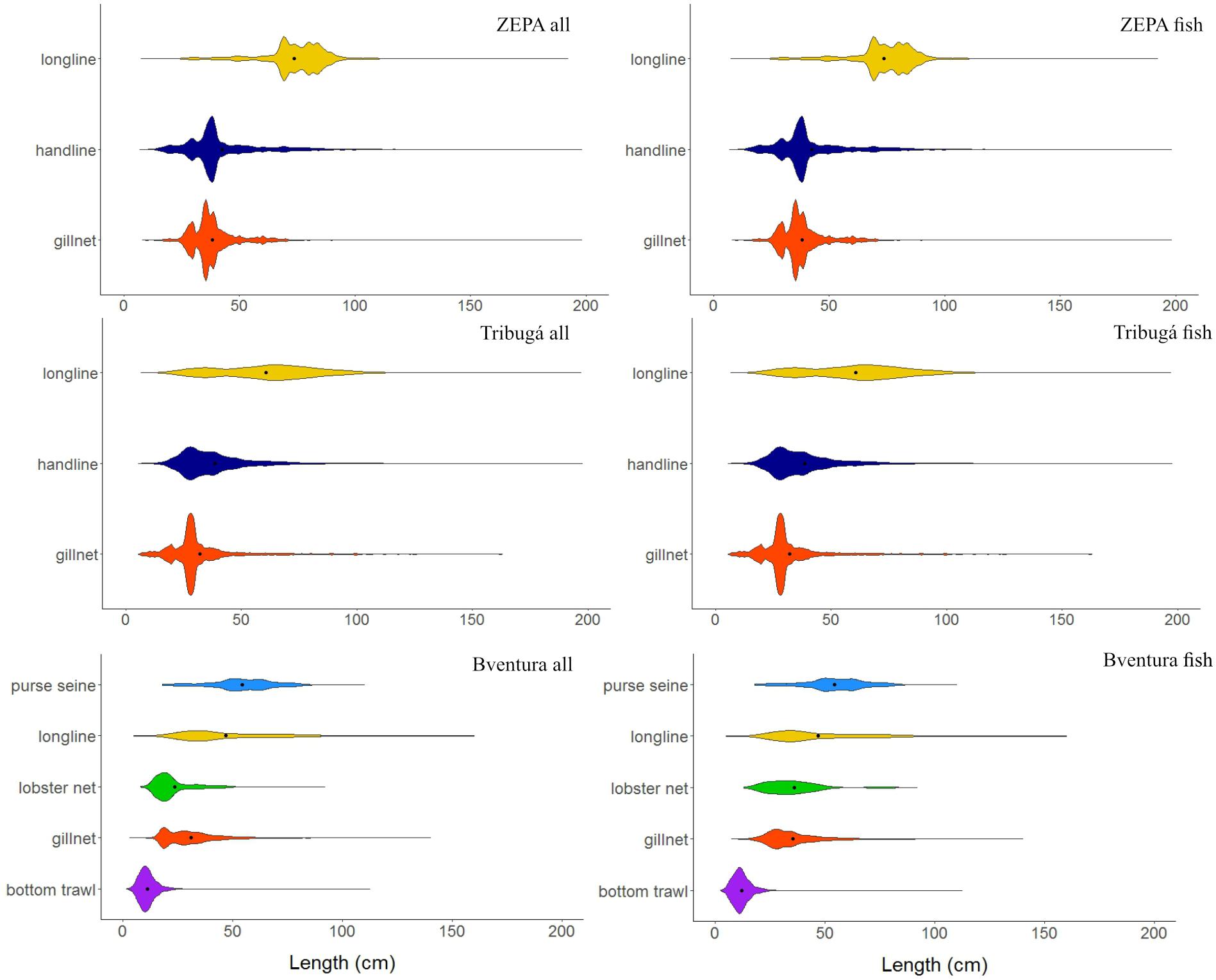
Figure 4. Length distribution of the entire catch (up to 200 cm) and of the fish portion of the catch of SSF per gear type at three coastal zones of the Colombian Pacific: ZEPA, Tribugá, and Buenaventura (abbreviation: Bventura). Black dots indicate mean values. A similar plot of the length distribution of the catch up to the maximum length observed at each zone is included as part of the Supplementary Figure S3.
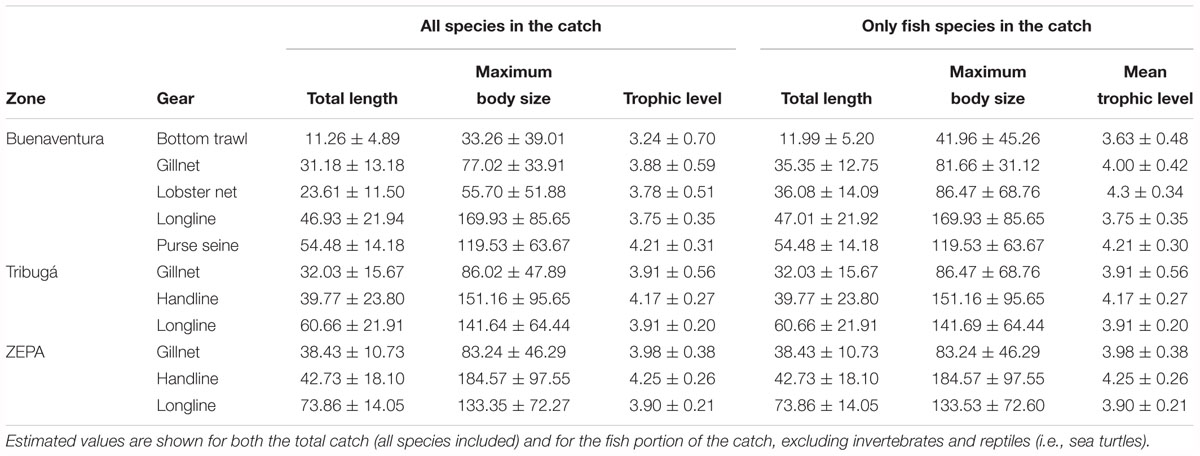
Table 4. Mean estimated values ± standard deviation for total length, maximum body size and trophic level of the catch at the three geographical zones studied here: Buenaventura, Tribugá, and ZEPA.
Handlines and longlines showed the largest maximum body size (MBS) of the species in the catch, with mean values above 130 cm in all cases (Figure 5 and Table 4). Mean MBS of the entire catch of handlines was statistically higher (p < 0.001) in ZEPA than in Tribugá. In the case of longlines, Buenaventura showed higher mean MBS than ZEPA and Tribugá, related to the high relative abundance of stingrays (Hypanus spp.) in the catch of longlines of that central coastal zone, although the mean MBS was only statistically different when compared to ZEPA (p = 0.02). On the other hand, gillnets appear to target species of medium MBS at all three zones with mean values close to 80 cm and statistical differences found between Tribugá and Buenaventura (p = 0.01). Bottom trawls had a significantly smaller mean MBS than all other gears except for lobster nets (p < 0.01 in all cases).
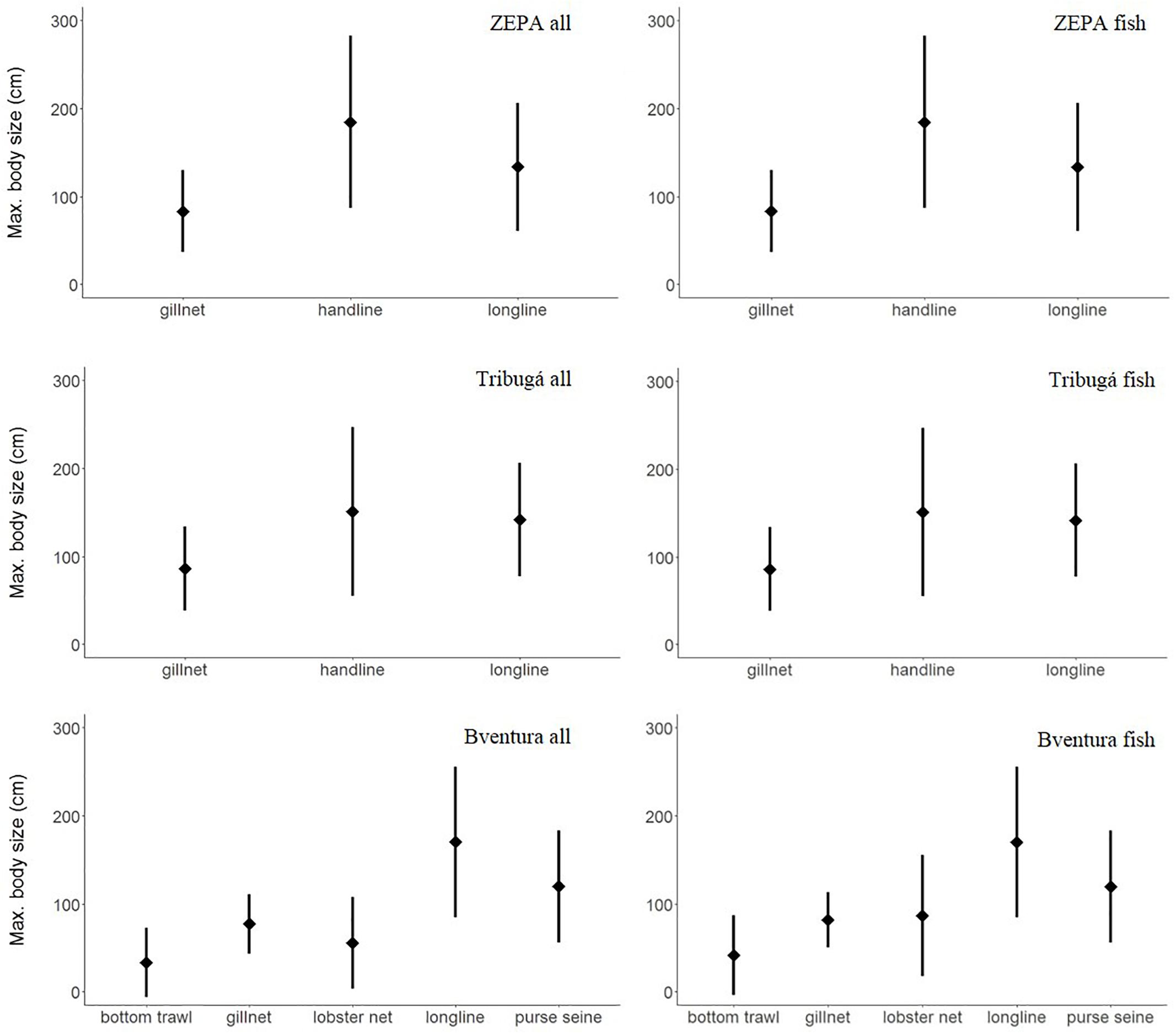
Figure 5. Weighted mean and standard deviation of maximum body size of the entire catch and of the fish portion of the catch of SSF per gear type at three coastal zones of the Colombian Pacific: ZEPA, Tribugá, and Buenaventura (abbreviation: Bventura).
Paired comparisons based on the entire catch within each coastal zone revealed that in Buenaventura, longlines had higher MBS than all other gears, except for purse seines (p < 0.001 in all cases), while bottom trawl had lower MBS than all other gears except for lobster nets (p < 0.01 in all cases); none of the other paired comparisons was statistically significant in Buenaventura. In ZEPA, handlines exhibited a significantly higher MBS than longlines and gillnets (p < 0.001 in both cases). In Tribugá mean MBS of gillnets was lower than that of handlines and longlines (p < 0.001 in both cases), but mean MBS values of the two hook-based gears, i.e., longlines and handlines, were not significantly different between each other.
For bottom trawls, gillnets and lobster nets in Buenaventura invertebrates accounted for 34, 8, and 56% of the catch, respectively. To assess the influence of shrimp and other small invertebrates on the estimates of size-based indicators, we estimated mean length and mean MBS for the fish portion of the catch, i.e., excluding all invertebrates and other non-fish species (i.e., sea turtles) from the data set prior to analyses. As expected, values of both size-based indicators, especially of MBS, increased for bottom trawls, lobster nets and gillnets in Buenaventura (Table 4) but had no effect in other gears of that coastal zone nor in the estimates derived from Tribugá and ZEPA, where invertebrates and reptiles accounted for only 0.03 and 0.01% of the catch, respectively. GLMs conducted for the fish portion of the catch showed the same statistical differences in total length among zones and/or gears observed previously for the whole catch, except for the difference between gillnets and lobster nets in Buenaventura which was not significant this time (p = 0.98). In contrast, the results of the pairwise comparisons of MBS values based on the fish portion of the catch showed that differences among zones or gears previously observed for the entire catch were no longer significant. In particular, mean MBS of the fish caught with gillnets was not statistically different between Buenaventura and Tribugá (p = 0.12) and within Buenaventura mean MBS of longlines and lobster nets were not statistically different (p = 0.34) from each other.
Functional Indicators of the Catch
Mean trophic level of the entire catch (all species included) was very similar across gears and zones, with mean values lying above 3.5 for all cases except for bottom trawls in Buenaventura that exhibited the lowest mean value, while handlines and purse seines exhibited the highest values (Figure 6 and Table 4). Statistically significant differences among gears within the same coastal zone were only found between MTL of handlines and longlines within ZEPA (p < 0.01).
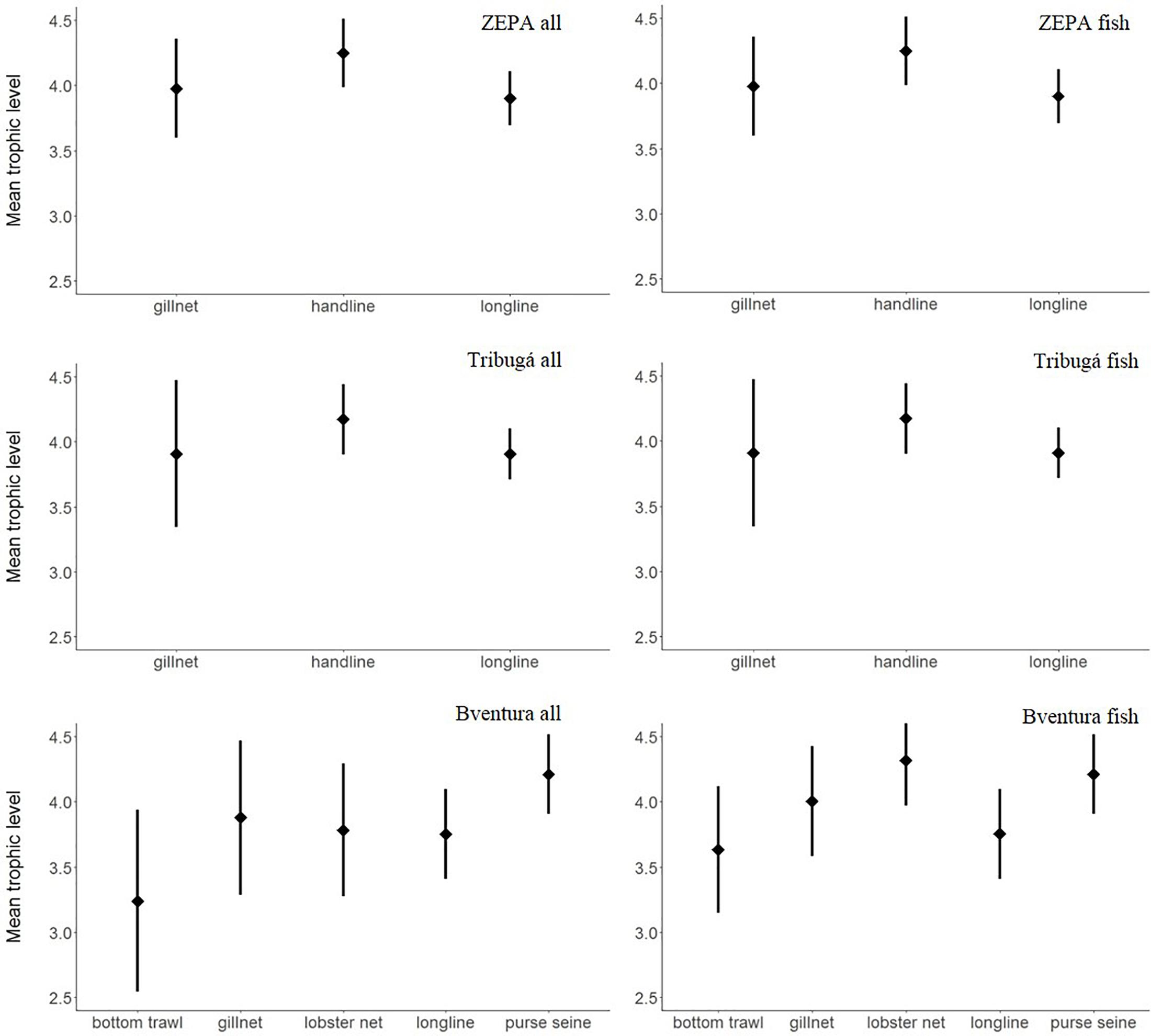
Figure 6. Weighted mean and standard deviation of trophic level of the entire catch and of the fish portion of the catch of SSF per gear type at three coastal zones of the Colombian Pacific: ZEPA, Tribugá, and Buenaventura (abbreviation: Bventura).
Following the rationale explained above for size-based indicators and considering the general positive relationship between a species body size and its trophic level (Romanuk et al., 2011), we also estimated MTL for the fish portion of the catch only. Similarly to the findings related to mean length and MBS, there was an increase – although relatively smaller – in the estimated values of MTL for bottom trawls, gillnets and lobster nets in Buenaventura (Figure 6 and Table 4). The small increase resulted in the difference between MTL of gillnets from Buenaventura and gillnets from Tribugá being no longer significant (p = 0.24).
Results of the relative abundance of trophic guilds corroborated that high trophic level guilds (piscivores and invertivores) are dominant in the catch of most gears across zones, except for bottom trawls, the gear that showed the highest diversity of trophic guilds (Figure 7). Also worth noting is the higher relative abundance of invertivores in the catch of longlines in Buenaventura compared to that of Tribugá and ZEPA for the same gear, where piscivores accounted for more than 90% of the catch.
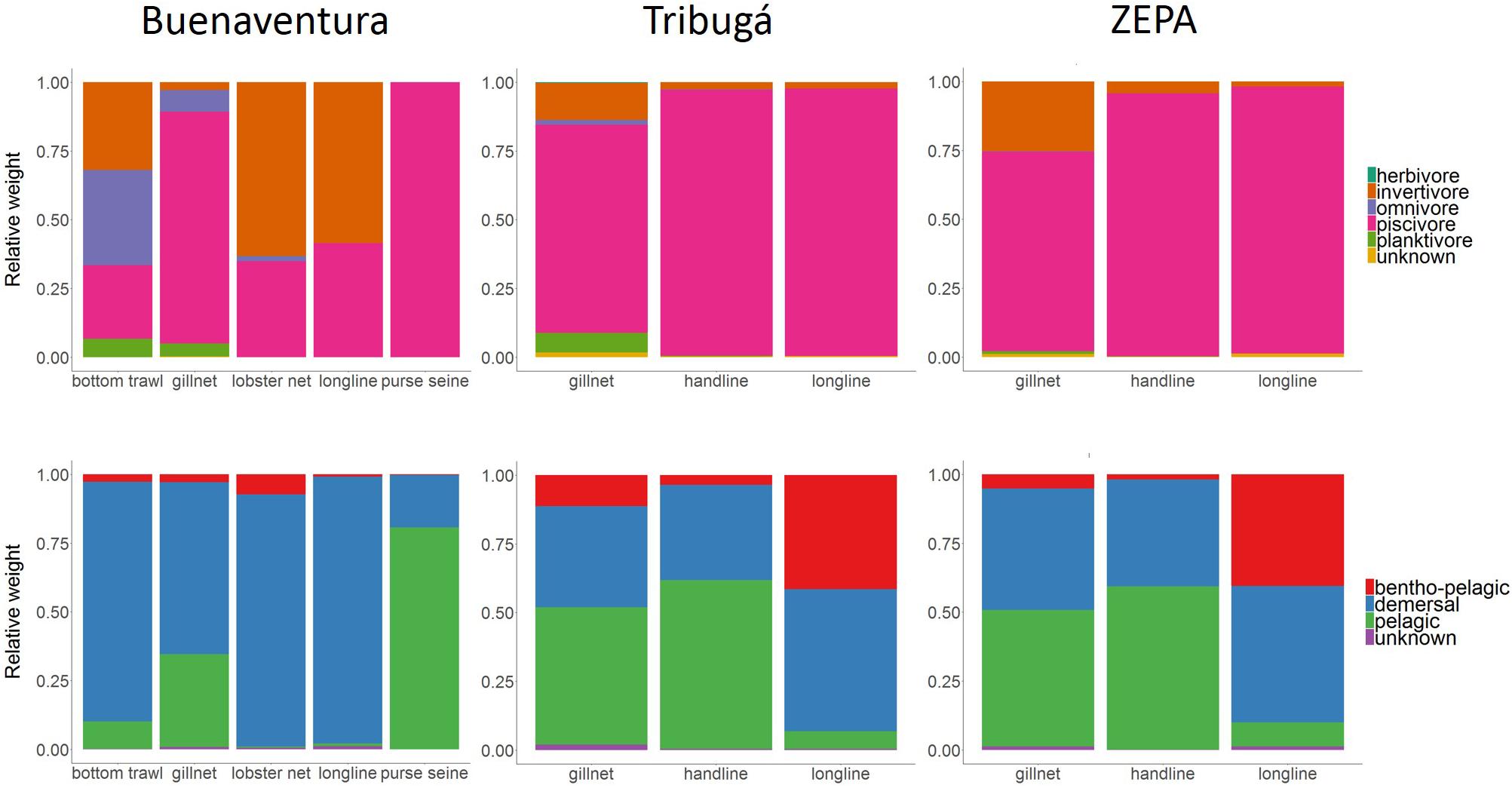
Figure 7. Proportion of trophic guilds and spatial guilds per gear type in the catch of SSF at three coastal zones of the Colombian Pacific: ZEPA, Tribugá, and Buenaventura.
In terms of spatial guilds, demersal species were dominant in Buenaventura for all gears except for purse seines, contrasting with the results from ZEPA and Tribugá where pelagic species had a higher relative abundance in the catch of gillnets and handlines while longlines caught more demersal and bentho-pelagic species (Figure 7). The overall proportions of species belonging to different spatial guilds was similar between ZEPA and Tribugá for the same type of gear: gillnets, handlines or longlines.
Conservation Indicators of the Catch
Based on the regional assessment of IUCN’s Red List, the three coastal zones have Least Concern (LC) as the predominant category of the biomass in the catch (54–73%), while threatened categories (Vulnerable – VU, Endangered – EN and Critically Endangered – CR) represented less than 1% of the biomass. The relative weight of species classified as Near Threatened (NT) was higher in ZEPA than in the other two zones with handlines being the gear that contributed most to that difference (Table 5). When the same analysis was based on the national assessment (Colombian’s red lists assessments), Not Evaluated (NE) and Near threatened (NT) were the dominant categories in the catch of all zones – with ZEPA exhibiting the highest relative abundance of NT species – while Data Deficient (DD) and LC had overall low values. Based on the national assessments, the relative weight of species under category VU was higher in the catch of ZEPA and Buenaventura, mostly due to the presence of species caught with longlines (e.g., stingrays). Overall, the relative abundance of threatened or near threatened categories in the catch was higher when based on national assessments than when the analysis was based on IUCN’s regional assessments.
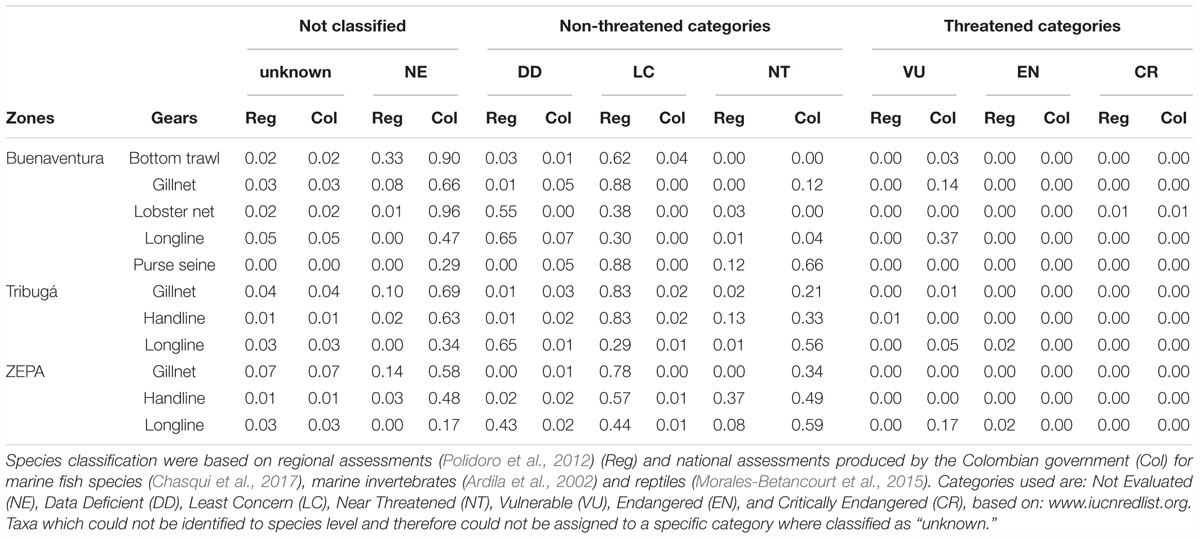
Table 5. Proportions of threatened or non-threatened taxa in the catch of SSF of the Colombian Pacific.
Landed by-catch species, those that are not commercialized or locally used, were only a conspicuous proportion of the catch of bottom trawls where they accounted for >30% of the catch (Table 6). For the rest of the gears, landed by-catch was below 3% and more than 75% of the catch corresponded to commercially important species. In ZEPA and Tribugá, 20% of the catch of gillnets is locally consumed or used as bait, instead of sold to local or external markets.
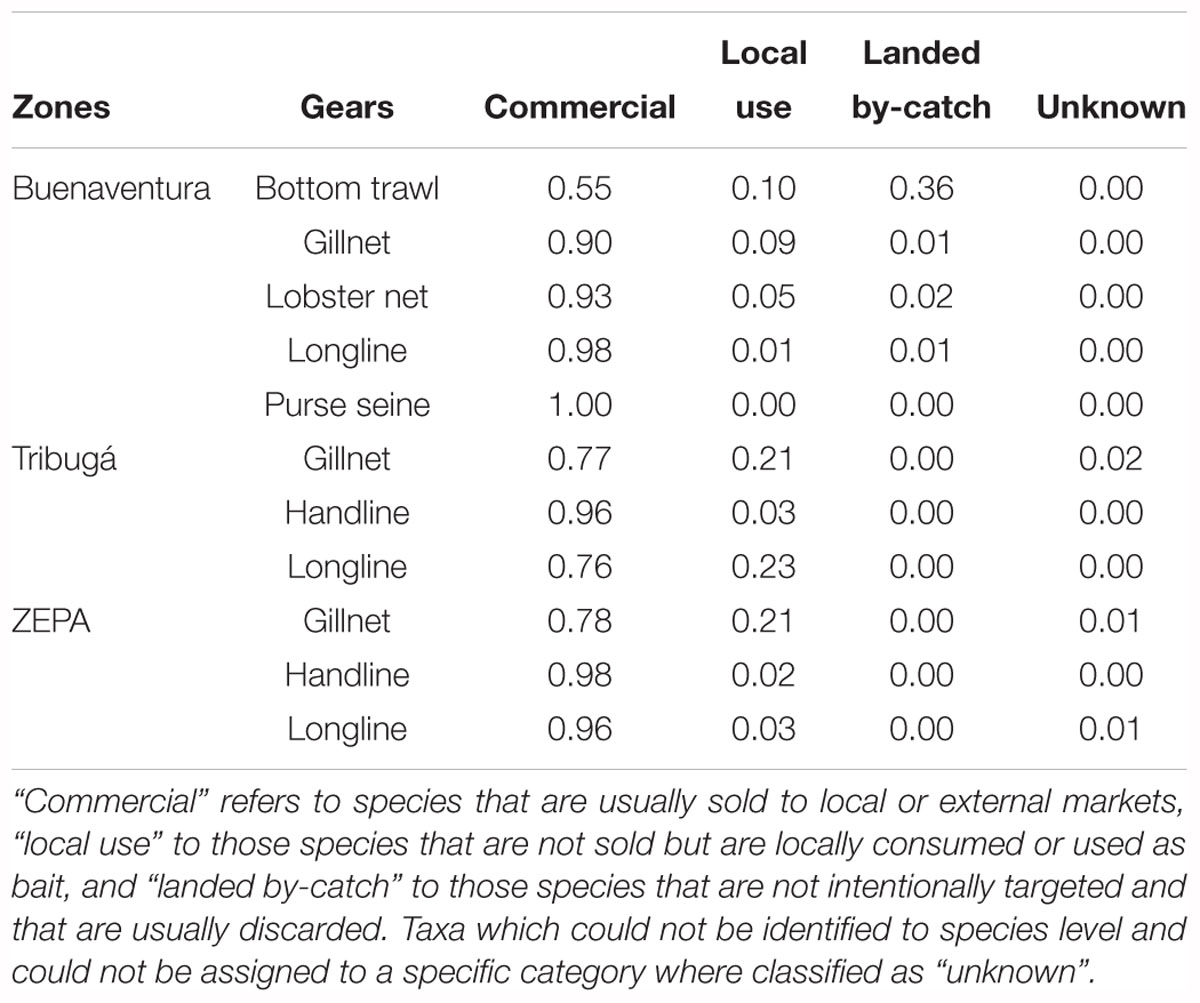
Table 6. Proportions of categories related to fishers’ use of the species registered in the catch of SSF in the Colombian Pacific.
Discussion
Our results showed clear differences in the catch composition among the three coastal zones and, particularly, between the northern rocky-dominated coast (Tribugá and ZEPA) and the central estuarine and mangrove-dominated coast (Buenaventura) of the Colombian Pacific. Some of the observed differences were related to the interaction between gear type and geographical location of the coastal zones.
Despite being in a traditionally data-poor tropical SSF context, our data sets, produced by a non-government organization and by an academic research project, included higher sampling frequency, sample size and geographic coverage than normal government fisheries data (Ramírez et al., 2017; Herrón et al., 2018). Community-based fishing monitoring schemes (as those followed in the present study) are therefore useful and likely more effective and less expensive ways of monitoring fisheries resources in typical SSF like the ones evaluated here. Overcoming some limitations in these schemes like the correct differentiation of certain species and common names within certain taxonomic groups, e.g., groupers, sharks, is something that will require further attention in the future (Castellanos-Galindo et al., 2018).
Size-Based and Functional Indicators
In the management areas established in the northern coastal zones, a higher selectivity of fishing gears has been promoted based on the assumption that gillnets tend to catch a higher proportion of immature fish and have higher by-catch rates than hook-based gears (Vieira et al., 2016; Ramírez-Luna and Chuenpagdee, 2019). Our results confirmed a lower mean length in the catch of gillnets when compared to longlines and handlines in ZEPA and Tribugá. However, fisheries selectivity is influenced not only by the gear used but also by spatial and temporal patterns of resource distribution (Maunder et al., 2014; Sampson, 2014). Therefore, the observed differences reflect not only the inherent selectivity of gears but also the location of the fishing grounds used by each gear. Particularly in ZEPA, longline fishers use deeper grounds located at greater distances from the shore, whereas gillnets tend to fish in areas closer to shore (Velandia and Díaz, 2016). A higher abundance of larger/older individuals in deeper habitats has been widely reported for many fish species and has been attributed to ontogenetic changes, although recent evidence indicate that this “deepening” could also be associated to increased fishing pressure in shallower areas (Frank et al., 2018). Distance to shore and depth of fishing grounds could also explain the higher mean length observed in the catch of purse seines in Buenaventura compared to other gears in the same zone (Figure 4). Purse seines are used at fishing grounds located further offshore (8–10 nautical miles) than other gears (unpublished data) and target mostly pelagic species (Figure 3). Bottom trawls – which exhibited the lowest mean length in the catch of all zones even when invertebrates are removed – have the smallest mesh size of all nets (0.5″) and are used in near-shore, shallow waters (unpublished data), targeting mainly two small-sized shrimp species (X. riveti and P. precipua). Continued monitoring of mean length in the catch complemented by spatial analyses of fishing grounds could provide more information regarding the factors explaining the observed differences and the potential long-term impacts of different gears on the size-structure of fish and invertebrate communities.
Our results of maximum body size (MBS) in the catch indicate that longlines and handlines are targeting larger body-sized species that are more vulnerable to overfishing due to their life history characteristics (Jennings et al., 1998; Cheung, 2007) (e.g., sailfish, tunas, and sharks, Figure 3), while bottom trawls are targeting species that could potentially withstand more fishing pressure and/or recover more rapidly (e.g., shrimps, other small invertebrates and small-sized fish species). Particularly in Buenaventura, longlines had a significantly higher MBS than the rest of the gears, probably linked to the fact that large-sized stingrays of the genus Hypanus were an important part of the catch of this gear (Figures 3, 5).
Targeting a relative high proportion of small-sized specimens has been suggested as a way of improving overall yields while maintaining the structure of the natural ecosystem, under the concept of “balanced harvest” (Kolding et al., 2015b), an approach that contradicts traditional management measures like imposing size limits for target species to avoid fishing immature individuals thus preventing growth and recruitment overfishing (Beverton, 1992; Myers and Mertz, 1998; Froese, 2004). Despite being more aligned to the principles of EBFM, critics of the balanced harvest approach have also argued that there are many practical difficulties of implementing such harvest scheme, particularly a drastic shift in consumers’ seafood preferences toward new species and sizes (Charles et al., 2015; Froese et al., 2015; Garcia et al., 2015).
Similarly to size-based indicators, MTL of the catch has been used as an indicator of ecological fishing impacts as it is expected to decrease with increasing fishing pressure [Pauly et al., 1998; Jennings et al., 2002; Pinnegar et al., 2002; Gascuel et al., 2016, but see Sethi et al. (2010)]. However, MTL has been criticized as an indicator of ecosystem condition since it can be largely influenced by external economic factors, such as market demands (for species and sizes) and by environmental variability that alters the dynamics of primary productivity and the recruitment of planktivore species (Caddy et al., 1998; Caddy and Garibaldi, 2000; Branch et al., 2010). Nevertheless, MTL may still be a suitable indicator for the state of a fishery system, if fishing pattern and external factors remain constant over time and only fishing effort increases (Shannon et al., 2014). Our estimates of MTL were fairly similar across gears and coastal zones and showed that SSF in the Colombian Pacific are extracting mainly high trophic level species of the system. This is corroborated by the high proportion of piscivores and invertivores in the catch of most gears across all zones, with the exception of bottom trawls that exhibited the highest diversity of trophic guilds in the catch (Figure 7).
These results go in line with a worldwide pattern of fishing that has focused on high trophic levels (Kolding et al., 2015a). MTL values observed here (overall mean: 3.9) are higher than MTL values reported in tropical SSF of the Western Indian Ocean [2.3–3.6, Rehren et al. (2018); Tuda et al. (2016)], the Caribbean [3.3–3.5, Arias-González et al. (2004)], the Indo-Pacific [2.4–3.7, Bacalso and Wolff (2014)] and other localities in the tropical eastern Pacific [2.5–2.9, Zetina-Rejon et al. (2003) and Díaz-Uribe et al. (2007)]. However, values of trophic level per species used in this study correspond to the adult phase of the species (FishBase, Froese and Pauly, 2017) and do not necessarily correspond to the actual trophic level of the size classes harvested per species. This can impose biases in the estimates of mean MTL of the catch (Caddy et al., 1998; Reed et al., 2016). In the future, local studies on the diet composition of target species should be conducted and used to estimate trophic levels per size class of main target species.
Differences observed in the proportion of spatial guilds across zones and gears seem best explained by location and habitat type. In ZEPA and Tribugá, coastal zones characterized by narrow continental shelves and few estuaries, pelagic and bentho-pelagic species dominated the catch (Figure 7). In contrast, fishing gears in the mangrove-dominated and estuarine area of Buenaventura caught mainly demersal species, except for purse seines, the only gear that operates further off-shore. Therefore, observed differences in proportions of spatial guilds do not seem to offer at this point an unequivocal indication of potential geographical or gear-based differences in fishing impacts but future assessments of temporal trends of this indicator might indicate changes in fishing effort or in the natural abundance of the resources (Caddy, 2000; Pitcher and Preikshot, 2001; Link et al., 2002).
Conservation Indicators
Based on regional assessments of the threatened status of species (Polidoro et al., 2012), most of the catch of SSF in the Colombian Pacific does not currently face major extinction risks, which could be interpreted as a sign of a sustainable fishery. However, the diagnosis is different when the national assessments are used (Ardila et al., 2002; Morales-Betancourt et al., 2015; Chasqui et al., 2017), since a large proportion of the biomass in the catch corresponds to Nearly Threatened (NT) species (Table 5). Based on the national red lists, longlines’ catch is conformed partly by species classified as Vulnerable (VU) in Buenaventura (37%) and in ZEPA (17%), mainly attributed to the presence of rays, stingrays and sharks. However, national assessments of commercially important species have generally been based on stock assessments with limited landings time-series or with poor spatial coverage. This could impose biases and is a common situation in data-limited tropical SSFs assessments (Costello et al., 2012; Ramírez et al., 2017; Herrón et al., 2018). On the other hand, the high proportion of Not Evaluated (NE) species in the catch of SSF, based on national assessments (Table 5), highlights the need to collect data on the status of natural populations based also on fishery-independent surveys.
By-catch and discards have also been considered to be meaningful indicators of the potential ecosystem impacts of fishing (Fulton et al., 2005; Link, 2005). They are increasingly being monitored and regulated in fisheries of developed countries [e.g., Landings:Discards ratio from the IndiSeas project, Coll et al. (2016)]. The high proportion of landed by-catch of bottom trawls observed here (36%) suggests a higher ecosystem impact of this fishing gear compared to other gears currently used. Bottom trawling has long been identified as a fishing method that can cause a variety of ecological impacts, such as: reduced abundance of non-target species, reduced diversity of the benthic community, sediment resuspension, disruption of nutrients cycles, changes in primary productivity, destruction of habitat and changes in trophic dynamics of the demersal and benthic communities (Collie et al., 2000, 2017; Olsgard et al., 2008; Dell et al., 2013). Fisheries authorities in Colombia banned the use of bottom trawls more than 10 years ago (INCODER, 2004) but fishers continue to use it since there is low enforcement capacity and high market demand for the main target species (small-sized shrimp species). On the other hand, a recent study on the effects of small-scale bottom trawling in similar estuarine environments in Brazil found that observed differences in the structure of macrofaunal communities seemed to be more related to natural variability than to the degree of trawling impact (Ortega et al., 2018). These authors discussed whether those communities could be adapted to a highly dynamic and frequently disturbed estuarine environment, which could also be the case of the benthic communities in Buenaventura that have sustained a bottom trawl fishery for more than 30 years. Specific studies on the dynamics of the catch of bottom trawls involving on-board monitoring and surveys of natural benthic communities will provide valuable inputs for management decisions regarding the continuation of the ban currently established on this gear or, perhaps, a transition toward fishing effort regulation.
ConcLusion
Analyses of the catch through the lens of ecological indicators provide alternative paths for the assessment and monitoring of SSF that complement the traditional single-species assessment methods and provide insights into potential ecological impacts of fishing. Observed differences in taxonomic composition of the catch and in the proportion of gears used among coastal zones most likely reflect the deep knowledge of small-scale fishers about the temporal and spatial distribution of resources (Saavedra-Diaz, 2012; Purcell et al., 2018). Hook-based gears (handlines and longlines) tend to catch larger sizes and higher trophic levels than nets, but they also include a higher proportion of species that are more vulnerable to fishing impacts and/or have higher conservation concerns. These findings challenge the generalized notion that more selective gears have overall lower ecological impacts. In contrast, net-based gears catch wider size ranges – although tend to focus on small-size classes – and include a wider representation of species, trophic and spatial guilds, which could arguably be considered a more “balanced harvest” type of fishing that retains ecosystem functionality (Garcia et al., 2015). Using the data presented here, a preliminary snap-shot assessment of the gears (Supplementary Table S3) suggests that there is not one ideal or “green” fishing gear since each gear harvests a specific size and/or functional component of the system and therefore will affect that component more severely than other gears. The rapid assessment also shows that the same type of gear can have different ecological impacts when used in different environmental contexts, e.g., the differences in the proportion of trophic and spatial guilds in the catch of longlines in Buenaventura compared to that in ZEPA.
Ecological indicators to assess the impacts of fisheries are most useful when assessed on a temporal timeframe and used simultaneously, taking into account that no single indicator can adequately inform on its own about the status or trends of a complex ecological system (Link et al., 2002; Shin et al., 2010; Coll et al., 2016). Additionally, the criteria to assess the degree of ecological impact of the gears must be aligned with fisheries management and conservation objectives that sometimes have conflicting long-term goals (Link, 2002). For example, targeting large individuals is usually considered a sound fisheries management measure on the basis of avoiding juveniles in the catch and allowing individuals to reproduce prior to being harvested. However, fish species that attain large body sizes are generally those that are more fecund (Barneche et al., 2018) and more vulnerable to overfishing compared to small-sized fish species, potentially facing higher extinction risks (Jennings et al., 1998; Cheung et al., 2005; Cheung, 2007).
In order to better inform management decisions related to ecological impacts imposed by different fishing gears, medium to long-term monitoring of the relative effort of each gear and of the metrics associated to ecological indicators is needed. We propose that simple ecological indicators, such as those used in this study, be included as part of annual assessments of multi-gear SSF in tropical countries where data and management capacities are limited. In this way, a systematic evaluation of the potential impacts of fishing at the community and ecosystem level could be developed and facilitate the transition toward EBFM.
Data Availability
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
PH, GC-G, and MW conceived and designed the analyses. PH and JD collected the data. PH, GC-G, and MS performed the analyses and produced the final figures. All authors discussed the results and contributed to the writing of the final manuscript.
Funding
MarViva Foundation received financial support from USAID (Cooperation Agreement #49-2012 of their BIOREDD+ Program), AUNAP (Cooperation Agreement #65-2012), the European Union (Cooperation Agreement #75-2013), and INVEMAR (Cooperation Agreement #009-2015). PH received financial support from CEMarin (Call # 5 of 2015) and from Rufford Foundation (RSG 19522-1).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the local communities and fishers from the municipalities of Juradó, Bahía Solano, Nuquí, and Buenaventura in the Colombian Pacific coast for allowing and supporting data collection at different landing sites between 2011 and 2017. We thank to C. A. Puentes for producing the map for Figure 1.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00127/full#supplementary-material
Footnotes
References
Acero, A. (2004). Systematics and Biogeography of the Tropical Sea Catfishes of the New World (Siluriformes: Ariidae). Ph.D. thesis, University of Arizona, Tucson, AZ.
Alverson, D. L., Freeberg, M. H., Murawski, S. A., and Pope, J. G. (1994). A Global Assessment of Fisheries Bycatch and Discards. Rome: Food & Agriculture Organization of the United Nations.
Andrew, N. L., Béné, C., Hall, S. J., Allison, E. H., Heck, S., and Ratner, B. D. (2007). Diagnosis and management of small-scale fisheries in developing countries. Fish Fish. 8, 227–240. doi: 10.1111/j.1467-2679.2007.00252.x
Ardila, N., Navas, G. R., and Reyes, J. (2002). Libro Rojo de Invertebrados Marinos de Colombia. Bogotá: INVEMAR.
Arias-González, J. E., Nuñez-Lara, E., González-Salas, C., and Galzin, R. (2004). Trophic models for investigation of fishing effect on coral reef ecosystems. Ecol. Modell. 172, 197–212. doi: 10.1016/j.ecolmodel.2003.09.007
AUNAP (2013). Resolución 899 de 2013. Por la cual se Establecen una Zona Exclusiva de Pesca Artesanal (ZEPA), una Zona Especial de Manejo Pesquero (ZEMP), en el Departamento del Chocó y se Adoptan Otras Medidas. Bogotá: Autoridad Nacional de Acuicultura y Pesca - AUNAP.
Bacalso, R. T. M., and Wolff, M. (2014). Trophic flow structure of the Danajon ecosystem (Central Philippines) and impacts of illegal and destructive fishing practices. J. Mar. Syst. 139, 103–118. doi: 10.1016/j.jmarsys.2014.05.014
Barneche, D. R., Robertson, D. R., White, C. R., and Marshall, D. J. (2018). Fish reproductive energy output increases disproportionately with body size. Science 360, 642–645. doi: 10.1126/science.aao6868
Béné, C., Hersoug, B., and Allison, E. H. (2010). Not by rent alone: analysing the pro-poor functions of small-scale fisheries in developing countries. Dev. Policy Rev. 28, 325–358. doi: 10.1111/j.1467-7679.2010.00486.x
Beverton, R. (1992). Patterns of reproductive strategy parameters in some marine teleost fishes. J. Fish Biol. 41, 137–160. doi: 10.1111/j.1095-8649.1992.tb03875.x
Branch, T. A., Watson, R., Fulton, E. A., Jennings, S., McGilliard, C. R., Pablico, G. T., et al. (2010). The trophic fingerprint of marine fisheries. Nature 468, 431–435. doi: 10.1038/nature09528
Caddy, J. (2000). Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. ICES J. Mar. Sci. 57, 628–640. doi: 10.1006/jmsc.2000.0739
Caddy, J., Csirke, J., Garcia, S., and Grainger, R. (1998). How pervasive is” fishing down marine food webs”? Science 282, 1383–1383. doi: 10.1126/science.282.5393.1383a
Caddy, J. F., and Garibaldi, L. (2000). Apparent changes in the trophic composition of world marine harvests: the perspective from the FAO capture database. Ocean Coast. Manag. 43, 615–655. doi: 10.1016/S0964-5691(00)00052-1
Castellanos-Galindo, G. A., Cantera, J., Valencia, N., Giraldo, S., Peña, E., Kluger, L. C., et al. (2017). Modeling trophic flows in the wettest mangroves of the world: the case of Bahía Málaga in the Colombian Pacific coast. Hydrobiologia 803, 13–27. doi: 10.1007/s10750-017-3300-6
Castellanos-Galindo, G. A., Chong-Montenegro, C., Baos, E. R. A., Zapata, L. A., Tompkins, P., Graham, R. T., et al. (2018). Using landing statistics and fishers’ traditional ecological knowledge to assess conservation threats to Pacific goliath grouper in Colombia. Aquat. Conserv. Mar. Freshw. Ecosyst. 28, 305–314. doi: 10.1002/aqc.2871
Charles, A., Garcia, S., and Rice, J. (2015). Balanced harvesting in fisheries: economic considerations. ICES J. Mar. Sci. 73, 1679–1689. doi: 10.1093/icesjms/fsv161
Chasqui, L., Polanco, A., Acero, A., Mejía- Falla, P. A., Navia, A., Zapata, L. A., et al. (2017). Libro Rojo de Peces Marinos de Colombia. Santa Marta: Ministerio de Ambiente y Desarrollo Sostenible, INVEMAR.
Cheung, W. L. (2007). Vulnerability of Marine Fishes to Fishing: from Global Overview to the Northern South China Sea. Doctoral dissertation, University of British Columbia, Vancouver.
Cheung, W. W., Pitcher, T. J., and Pauly, D. (2005). A fuzzy logic expert system to estimate intrinsic extinction vulnerabilities of marine fishes to fishing. Biol. Conserv. 124, 97–111. doi: 10.1016/j.biocon.2005.01.017
CODECHOCO (2014). Acuerdo 011 de 2014 del Consejo Directivo de Codechoco -Corporación Autónoma Regional Para el Desarrollo Sostenible del Chocó. Quibdó: CODECHOCO.
Coll, M., Shannon, L., Kleisner, K., Juan-Jordá, M., Bundy, A., Akoglu, A., et al. (2016). Ecological indicators to capture the effects of fishing on biodiversity and conservation status of marine ecosystems. Ecol. Indic. 60, 947–962. doi: 10.1016/j.ecolind.2015.08.048
Collie, J., Hiddink, J. G., van Kooten, T., Rijnsdorp, A. D., Kaiser, M. J., Jennings, S., et al. (2017). Indirect effects of bottom fishing on the productivity of marine fish. Fish Fish. 18, 619–637. doi: 10.1111/faf.12193
Collie, J. S., Hall, S. J., Kaiser, M. J., and Poiner, I. R. (2000). A quantitative analysis of fishing impacts on shelf-sea benthos. J. Anim. Ecol. 69, 785–798. doi: 10.1046/j.1365-2656.2000.00434.x
Cope, J. M., and Punt, A. E. (2009). Length-based reference points for data-limited situations: applications and restrictions. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 1, 169–186. doi: 10.1577/C08-025.1
Correa, I., and Morton, R. (2010). Pacific Coast of Colombia, Encyclopedia of the World’s Coastal Landforms. Berlin: Springer, 193–198. doi: 10.1007/978-1-4020-8639-7_29
Costello, C., Ovando, D., Hilborn, R., Gaines, S. D., Deschenes, O., and Lester, S. E. (2012). Status and solutions for the world’s unassessed fisheries. Science 338, 517–520. doi: 10.1126/science.1223389
Criales-Hernandez, M. I., Duarte, L. O., García, C. B., and Manjarrés, L. (2006). Ecosystem impacts of the introduction of bycatch reduction devices in a tropical shrimp trawl fishery: insights through simulation. Fish. Res. 77, 333–342. doi: 10.1016/j.fishres.2005.10.005
DANE (2011). Estimación y Proyección de Población Nacional, Departamental y Municipal Total por Área 1985-2020, Proyecciones de Población. Available at: www.dane.gov.co/index.php/estadisticas-por-tema/demografia-y-poblacion/proyecciones-de-poblacion
Degnbol, P., and Jarre, A. (2004). Review of indicators in fisheries management–a development perspective. Afr. J. Mar. Sci. 26, 303–326. doi: 10.2989/18142320409504063
Dell, Q., Griffiths, S., Tonks, M., Rochester, W., Miller, M., Duggan, M., et al. (2013). Effects of trawling on the diets of common demersal fish by-catch of a tropical prawn trawl fishery. J. Fish Biol. 82, 907–926. doi: 10.1111/jfb.12026
Denney, N. H., Jennings, S., and Reynolds, J. D. (2002). Life–history correlates of maximum population growth rates in marine fishes. Proc. R. Soc. Lond. B Biol. Sci. 269, 2229–2237. doi: 10.1098/rspb.2002.2138
Díaz, J. M., Guillot, L., and Velandia-Diaz, M. C. (2016). La Pesca Artesanal en la Costa Norte del Pacífico Colombiano: un Horizonte Ambivalente. Bogotá: Fundación Marviva.
Díaz-Uribe, J. G., Arreguín-Sánchez, F., and Cisneros-Mata, M. A. (2007). Multispecies perspective for small-scale fisheries management: a trophic analysis of La Paz Bay in the Gulf of California, Mexico. Ecol. Modell. 201, 205–222. doi: 10.1016/j.ecolmodel.2006.09.015
Dulvy, N. K., and Reynolds, J. D. (2002). Predicting extinction vulnerability in skates. Conserv. Biol. 16, 440–450. doi: 10.1046/j.1523-1739.2002.00416.x
Ehemann, N., Pérez-Palafox, X., Mora-Zamacona, P., Burgos-Vázquez, M., Navia, A., Mejía-Falla, P., et al. (2017). Size–weight relationships of batoids captured by artisanal fishery in the southern Gulf of California, Mexico. J. Appl. Ichthyol. 33, 1051–1054. doi: 10.1111/jai.13421
FAO (2000). Conversion Factors: Landed Weight to Live Weight. Food and Agricultural Organization of the United Nations. Rome: FAO, 378.
FAO (2015). Voluntary Guidelines for Securing Sustainable Small-Scale Fisheries in the Context of Food Security and Poverty Eradication. Rome: Food and Agriculture Organization of the United Nations, 34.
FAO (2018). Handbook of Fishery Statistical Standards. Coordinating Working Party on Fishery Statistics (CWP). Rome: Food and Agriculture Organization of the United Nations.
Fisher, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K., and Niem, V. (1995). Guía FAO Para la Identificación de Especies Para los Fines de la Pesca: Pacífico Centro-Oriental. Vol. I, II and III. Roma: FAO.
Frank, K. T., Petrie, B., Leggett, W. C., and Boyce, D. G. (2018). Exploitation drives an ontogenetic-like deepening in marine fish. Proc. Natl. Acad. Sci. U.S.A. 115, 6422–6427. doi: 10.1073/pnas.1802096115
Froese, R. (2004). Keep it simple: three indicators to deal with overfishing. Fish Fish. 5, 86–91. doi: 10.1111/j.1467-2979.2004.00144.x
Froese, R., and Pauly, D. (2017). FishBase, World Wide Web Electronic Publication. Available at: www.fishbase.org
Froese, R., Walters, C., Pauly, D., Winker, H., Weyl, O. L., Demirel, N., et al. (2015). A critique of the balanced harvesting approach to fishing. ICES J. Mar. Sci. 73, 1640–1650. doi: 10.1093/icesjms/fsv122
Fulton, E. A., Smith, A. D., and Punt, A. E. (2005). Which ecological indicators can robustly detect effects of fishing? ICES J. Mar. Sci. 62, 540–551. doi: 10.1016/j.icesjms.2004.12.012
Garcia, S., Rice, J., and Charles, A. (2015). Balanced harvesting in fisheries: a preliminary analysis of management implications. ICES J. Mar. Sci. 73, 1668–1678. doi: 10.1093/icesjms/fsv156
Garcia, S. M. (2003). The Ecosystem Approach to Fisheries: Issues, Terminology, Principles, Institutional Foundations, Implementation and Outlook. Rome: Food & Agriculture Organization of the United Nations.
Gascuel, D., Coll, M., Fox, C., Guénette, S., Guitton, J., Kenny, A., et al. (2016). Fishing impact and environmental status in European seas: a diagnosis from stock assessments and ecosystem indicators. Fish Fish. 17, 31–55. doi: 10.1111/faf.12090
Greenstreet, S. P., and Rogers, S. I. (2006). Indicators of the health of the North Sea fish community: identifying reference levels for an ecosystem approach to management. ICES J. Mar. Sci. 63, 573–593. doi: 10.1016/j.icesjms.2005.12.009
Guénette, S., and Gascuel, D. (2012). Shifting baselines in European fisheries: the case of the Celtic Sea and Bay of Biscay. Ocean Coast. Manag. 70, 10–21. doi: 10.1016/j.ocecoaman.2012.06.010
Herrón, P., Mildenberger, T. K., Diaz, J. M., and Wolff, M. (2018). Assessment of the stock status of small-scale and multi-gear fisheries resources in the tropical Eastern Pacific region. Reg. Stud. Mar. Sci. 24, 311–323. doi: 10.1016/j.rsma.2018.09.008
INCODER (2004). Resolución 00695 de 2004 - Instituto Colombiano de Desarrollo Rural - INCODER, Diario Oficial No. 45.545. 11 de Mayo de 2004. Bogotá: INCODER.
IUCN (2017). The IUCN Red List of Threatened Species. Version 2017-3. International Union for Conservation of Nature. Gland: IUCN.
Jennings, S. (2005). Indicators to support an ecosystem approach to fisheries. Fish Fish. 6, 212–232. doi: 10.1111/j.1467-2979.2005.00189.x
Jennings, S., and Dulvy, N. K. (2005). Reference points and reference directions for size-based indicators of community structure. ICES J. Mar. Sci. 62, 397–404. doi: 10.1016/j.icesjms.2004.07.030
Jennings, S., Greenstreet, S., Hill, L., Piet, G., Pinnegar, J., and Warr, K. J. (2002). Long-term trends in the trophic structure of the North Sea fish community: evidence from stable-isotope analysis, size-spectra and community metrics. Mar. Biol. 141, 1085–1097. doi: 10.1007/s00227-002-0905-7
Jennings, S., and Kaiser, M. J. (1998). “The effects of fishing on marine ecosystems,” in Advances in Marine Biology, eds J. H. S. Blaxter, A. J. Southward, and P. A. Tyler (Cambridge, MA: Academic Press), 201–352.
Jennings, S., Reynolds, J. D., and Mills, S. C. (1998). Life history correlates of responses to fisheries exploitation. Proc. R. Soc. Lond. B Biol. Sci. 265, 333–339. doi: 10.1098/rspb.1998.0300
Jennings, S., Reynolds, J. D., and Polunin, N. V. (1999). Predicting the vulnerability of tropical reef fishes to exploitation with phylogenies and life histories. Conserv. Biol. 13, 1466–1475. doi: 10.1046/j.1523-1739.1999.98324.x
Kato, Y., Davis, A., and Ruddle, K. (2012). Appropriate Management for Small-Scale Tropical Fisheries. SPC Traditional Marine Resource Management and Knowledge Information Bulletin #30, 13. Available at: https://spccfpstore1.blob.core.windows.net/digitallibrary-docs/files/31/310e0ee58324c593947d1b9fad8461a4.pdf?sv=2015-12-11&sr=b&sig=eN%2B9MESvIliQliPVF%2BRE0PPKqVLvnatYHwNZFtz9mUg%3D&se=2019-09-04T09%3A19%3A29Z&sp=r&rscc=public%2C%20max-age%3D864000%2C%20max-stale%3D86400&rsct=application%2Fpdf&rscd=inline%3B%20filename%3D%22Trad30_03_Kato.pdf%22
Keen, A. M. (1971). Sea Shells of Tropical West America: Marine Mollusks from Baja California to. Palo Alto, CA: Stanford University Press.
Kolding, J., Bundy, A., van Zwieten, P. A., and Plank, M. J. (2015a). Fisheries, the inverted food pyramid. ICES J. Mar. Sci. 73, 1697–1713. doi: 10.1093/icesjms/fsv225
Kolding, J., Jacobsen, N. S., Andersen, K. H., and van Zwieten, P. A. (2015b). Maximizing fisheries yields while maintaining community structure. Can. J. Fish. Aquat. Sci. 73, 644–655. doi: 10.1139/cjfas-2015-0098
Link, J. S. (2005). Translating ecosystem indicators into decision criteria. ICES J. Mar. Sci. 62, 569–576. doi: 10.1016/j.icesjms.2004.12.015
Link, J. S., Brodziak, J. K., Edwards, S. F., Overholtz, W. J., Mountain, D., Jossi, J. W., et al. (2002). Marine ecosystem assessment in a fisheries management context. Can. J. Fish. Aquat. Sci. 59, 1429–1440. doi: 10.1139/f02-115
López-Angarita, J., Tilley, A., Díaz, J. M., Hawkins, J. P., Cagua, E. F., and Roberts, C. M. (2018). Winners and losers in area-based management of a small-scale fishery in the Colombian pacific. Front. Mar. Sci. 5:23. doi: 10.3389/fmars.2018.00023
Marceniuk, A. P., and Acero, P. (2009). A new species of Cathorops (Siluriformes; Ariidae) from Mesoamerica, with redescription of four species from the eastern Pacific. Bull. Mar. Sci. 85, 245–280.
Maunder, M., Crone, P., Valero, J., and Semmens, B. (2014). Selectivity: theory, estimation, and application in fishery stock assessment models. Fish. Res. 158, 11–14. doi: 10.1016/j.fishres.2014.03.017
Mejía-Rentería, J. C., Castellanos-Galindo, G. A., Cantera-Kintz, J. R., and Hamilton, S. E. (2018). A comparison of Colombian Pacific mangrove extent estimations: implications for the conservation of a unique Neotropical tidal forest. Estuar. Coast. Shelf Sci. 212, 233–240. doi: 10.1016/j.ecss.2018.07.020
Morales-Betancourt, M. A., Lasso, C. A., Páez, V. P., and Bock, B. C. (2015). Libro Rojo de Reptiles de Colombia Bogotá. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt.
Myers, R., and Mertz, G. (1998). The limits of exploitation: a precautionary approach. Ecol. Appl. 8, S165–S169. doi: 10.2307/2641375
Olsgard, F., Schaanning, M. T., Widdicombe, S., Kendall, M. A., and Austen, M. C. (2008). Effects of bottom trawling on ecosystem functioning. J. Exp. Mar. Biol. Ecol. 366, 123–133. doi: 10.1016/j.jembe.2008.07.036
Ortega, I., Colling, L. A., and Dumont, L. F. C. (2018). Response of soft-bottom macrobenthic assemblages to artisanal trawling fisheries in a subtropical estuary. Estuar. Coast. Shelf Sci. 207, 142–153. doi: 10.1016/j.ecss.2018.04.007
Palomares, M. L. D., and Pauly, D. (2018). SeaLifeBase. World Wide Web Electronic Publication. Available at: www.sealifebase.org
Pauly, D. (1984). Fish Population Dynamics in Tropical Waters: a Manual for use with Programmable Calculators, Vol. 8. Penang: WorldFish.
Pauly, D., Christensen, V., Dalsgaard, J., Froese, R., and Torres, F. (1998). Fishing down marine food webs. Science 279, 860–863. doi: 10.1126/science.279.5352.860
Pikitch, E., Santora, C., Babcock, E., Bakun, A., Bonfil, R., Conover, D., et al. (2004). Ecosystem-based fishery management. Science 305, 346–347. doi: 10.1126/science.1098222
Pinnegar, J. K., Jennings, S., O’Brien, C., and Polunin, N. (2002). Long-term changes in the trophic level of the Celtic Sea fish community and fish market price distribution. J. Appl. Ecol. 39, 377–390. doi: 10.1046/j.1365-2664.2002.00723.x
Pitcher, T. J., and Preikshot, D. (2001). RAPFISH: a rapid appraisal technique to evaluate the sustainability status of fisheries. Fish. Res. 49, 255–270. doi: 10.1111/jfb.12122
PNN (2006). Plan de Manejo Parque Nacional Natural Utria. Bahia Solano: Parques Nacionales Naturales de Colombia, 223.
PNN (2011). Plan de Manejo Recursos Hidrobiologicos y Pesqueros Parque Nacional Natural Utría Parques Nacionales Naturales de Colombia - Dirección Territorial Pacifico. Chocó: El Valle, 32.
Polidoro, B. A., Brooks, T., Carpenter, K. E., Edgar, G. J., Henderson, S., Sanciangco, J., et al. (2012). Patterns of extinction risk and threat for marine vertebrates and habitat-forming species in the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser. 448, 93–104. doi: 10.3354/meps09545
Puentes, V., Polo, C. J., Roldán, A. M., and Zuluaga, A. P. (2014). Artes y Métodos de Pesca en Colombia. Serie Recursos Pesqueros de Colombia. Autoridad Nacional de Acuicultura y Pesca – AUNAP. Bogotá: Conservación Internacional, 216.
Purcell, S. W., Fraser, N., Tagica, S., Lalavanua, W., and Ceccarelli, D. M. (2018). Discriminating catch composition and fishing modes in an artisanal multispecies fishery. Front. Mar. Sci. 5:243. doi: 10.3389/fmars.2018.00243
Purcell, S. W., and Pomeroy, R. S. (2015). Driving small-scale fisheries in developing countries. Front. Mar. Sci. 2:44. doi: 10.3389/fmars.2015.00044
R Core Team (2017). A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Ramírez, J. G., Lleonart, J., Coll, M., Reyes, F., and Puentes, G. M. (2017). Improving stock assessment and management advice for data-poor small-scale fisheries through participatory monitoring. Fish. Res. 190, 71–83. doi: 10.1016/j.fishres.2017.01.015
Ramírez-Luna, V., and Chuenpagdee, R. (2019). Exclusive Fishing Zone for Small-Scale Fisheries in Northern Chocó, Colombia: Pre-and Post-implementation, Viability and Sustainability of Small-Scale Fisheries in Latin America and the Caribbean. Berlin: Springer, 197–217. doi: 10.1007/978-3-319-76078-0_9
Reed, J., Shannon, L., Velez, L., Akoglu, E., Bundy, A., Coll, M., et al. (2016). Ecosystem indicators—Accounting for variability in species’ trophic levels. ICES J. Mar. Sci. 74, 158–169. doi: 10.1093/icesjms/fsw150
Rehren, J., Wolff, M., and Jiddawi, N. (2018). Holistic assessment of Chwaka Bay’s multi-gear fishery–Using a trophic modeling approach. J. Mar. Syst. 180, 265–278. doi: 10.1016/j.jmarsys.2018.01.002
Robertson, D. R., and Allen, G. R. (2015). Shorefishes of the Tropical Eastern Pacific: Online Information System. Balboa: Smithsonian Tropical Research Institute.
Rochet, M.-J., and Rice, J. C. (2005). Do explicit criteria help in selecting indicators for ecosystem-based fisheries management? ICES J. Mar. Sci. 62, 528–539. doi: 10.1016/j.icesjms.2005.01.007
Rochet, M.-J., and Trenkel, V. M. (2003). Which community indicators can measure the impact of fishing? A review and proposals. Can. J. Fish. Aquat. Sci. 60, 86–99. doi: 10.1139/f02-164
Romanuk, T. N., Hayward, A., and Hutchings, J. A. (2011). Trophic level scales positively with body size in fishes. Glob. Ecol. Biogeogr. 20, 231–240. doi: 10.1111/j.1466-8238.2010.00579.x
Russell, L. (2018). emmeans: Estimated Marginal Means, aka Least-Squares Means., R package version 1.2.3.
Saavedra-Diaz, L. M. (2012). Towards Colombian Small-Scale Marine Fisheries Management: Hacia un Manejo de la Pesca Marina Artesanal en Colombia. Ph.D thesis. University of New Hampshire, Durham, NC, 367.
Salas, S., Barragan-Paladines, M. J., and Chuenpagdee, R. (2019). Viability and Sustainability of Small-Scale Fisheries in Latin America and the Caribbean. Cham: Springer. doi: 10.1007/978-3-319-76078-0
Salas, S., Chuenpagdee, R., Seijo, J. C., and Charles, A. (2007). Challenges in the assessment and management of small-scale fisheries in Latin America and the Caribbean. Fish. Res. 87, 5–16. doi: 10.1016/j.fishres.2007.06.015
Sampson, D. B. (2014). Fishery selection and its relevance to stock assessment and fishery management. Fish. Res. 158, 5–14. doi: 10.1016/j.fishres.2013.10.004
Sethi, S. A., Branch, T. A., and Watson, R. (2010). Global fishery development patterns are driven by profit but not trophic level. Proc. Natl. Acad. Sci. U.S.A 107, 12163–12167. doi: 10.1073/pnas.1003236107
Shannon, L., Coll, M., Bundy, A., Gascuel, D., Heymans, J. J., Kleisner, K., et al. (2014). Trophic level-based indicators to track fishing impacts across marine ecosystems. Mar. Ecol. Prog. Ser. 512, 115–140. doi: 10.3354/meps10821
Shin, Y.-J., Rochet, M.-J., Jennings, S., Field, J. G., and Gislason, H. (2005). Using size-based indicators to evaluate the ecosystem effects of fishing. ICES J. Mar. Sci. 62, 384–396. doi: 10.1016/j.icesjms.2005.01.004
Shin, Y. J., Shannon, L. J., Bundy, A., Coll, M., Aydin, K., Bez, N., et al. (2010). Using indicators for evaluating, comparing, and communicating the ecological status of exploited marine ecosystems. 2. Setting the scene. ICES J. Mar. Sci. 67, 692–716. doi: 10.1093/icesjms/fsp294
Smith, S. E., Au, D. W., and Show, C. (1998). Intrinsic rebound potentials of 26 species of Pacific sharks. Mar. Freshw. Res. 49, 663–678. doi: 10.1071/MF97135
Tesfaye, G., and Wolff, M. (2018). Modeling trophic interactions and the impact of an introduced exotic carp species in the Rift Valley Lake Koka, Ethiopia. Ecol. Modell. 378, 26–36. doi: 10.1016/j.ecolmodel.2018.04.003
Tuda, P. M., Wolff, M., and Breckwoldt, A. (2016). Size structure and gear selectivity of target species in the multispecies multigear fishery of the Kenyan South Coast. Ocean Coast. Manag. 130, 95–106. doi: 10.1016/j.ocecoaman.2016.06.001
Velandia, M. C., and Díaz, J. M. (2016). Atlas Marino-Costero del Pacífico Norte Colombiano. Bogotá: Fundación MarViva.
Vieira, C. A., Díaz-Granados, M. A., and Díaz, J. M. (2016). “Ordenamiento y manejo pesquero en la costa norte del Pacifico Chocoano,” in La Pesca Artesanal en el Norte del Pacífico Colombiano: Un Horizonte Ambivalente, eds J. M. Diaz, L. Guillot, and M. C. Velandia-Diaz (Bogotá: Fundación MarViva),45–57.
Keywords: Colombia, ecological indicators, ecosystem approach to fisheries, gear-based management, tropical fisheries, catch composition
Citation: Herrón P, Castellanos-Galindo GA, Stäbler M, Díaz JM and Wolff M (2019) Toward Ecosystem-Based Assessment and Management of Small-Scale and Multi-Gear Fisheries: Insights From the Tropical Eastern Pacific. Front. Mar. Sci. 6:127. doi: 10.3389/fmars.2019.00127
Received: 06 November 2018; Accepted: 01 March 2019;
Published: 19 March 2019.
Edited by:
Beatrice Irene Crona, The Royal Swedish Academy of Sciences, SwedenReviewed by:
Konstantinos Tsagarakis, Hellenic Centre for Marine Research (HCMR), GreeceMichele Gristina, National Research Council (CNR), Italy
Copyright © 2019 Herrón, Castellanos-Galindo, Stäbler, Díaz and Wolff. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pilar Herrón, cGlsYXJoZXJyb25AZ21haWwuY29t