The Effect of Algal-Gardening Damselfish on the Resilience of the Mesoamerican Reef
 Ángela Randazzo Eisemann1,2*
Ángela Randazzo Eisemann1,2*  Jorge Luis Montero Muñoz2
Jorge Luis Montero Muñoz2  Melanie McField3 Jenny Myton4
Melanie McField3 Jenny Myton4  Jesús Ernesto Arias-González1,2*
Jesús Ernesto Arias-González1,2*- 1Laboratorio de Ecología de Ecosistemas de Arrecifes Coralinos, Centro de Investigación y de Estudios Avanzados del IPN, Mérida, Mexico
- 2Departamento de Recursos del Mar, Centro de Investigación y de Estudios Avanzados del IPN, Mérida, Mexico
- 3Healthy Reefs for Healthy People Initiative, Smithsonian Marine Station, Fort Pierce, FL, United States
- 4Coral Reef Alliance, San Francisco, CA, United States
The structures, functions, and services provided by coral reef ecosystems are deteriorating worldwide. However, not all coral reefs are affected the same way, with some showing signs of resistance and/or recovery from disturbances. Understanding the drivers and feedbacks that contribute to shifts in community structure is valuable to support resilience-based management. In this study, key community variables that influence the resilience of coral reef ecosystems were examined in 64 sites of the Mesoamerican Reef (MAR) monitored in both 2006 and 2016, as part of the Healthy Reef Initiative (HRI), using the Atlantic and Gulf Rapid Reef Assessment (AGRRA) monitoring protocol. Based on benthic cover thresholds, sites were classified into three different states: coral state (CS) with >10% live coral and <5% fleshy macroalgae; stressed coral state (SCS) with >10% live coral and >5% fleshy macroalgae and; depauperate coral state (DCS) <10% live coral. The associations between site states and the density of different fish functional groups were analyzed to determine their effects on coral reef resilience. The results highlight that territorial herbivores (algal-gardening damselfish) may play a key role in maintaining feedbacks toward macroalgae-stressed states. This supports the recommendation of reinforcing Marine Replenishment Zones (MRZ) in order to promote healthy populations of resident predator fish (like groupers and snappers), which could potentially regulate algal-gardening damselfish populations and diminish negative cascade effects on coral reefs. Collaborative and resilience-based management will continue to be promoted by the HRI partners, supporting the establishment of additional MRZs along with ongoing efforts to directly protect herbivorous fish (surgeonfish and parrotfish) and to improve water quality, through better wastewater treatment, watershed management, and coastal development plans, with the purpose of continuing to build coral reef resilience in the MAR.
Introduction
Over the last four decades, degradation of coral reef ecosystem structures and functions have been reported worldwide (Gardner et al., 2003; De’ath et al., 2012; Birkeland, 2015; Bruno and Valdivia, 2016; Hughes et al., 2017) impacting human well-being through the erosion of ecosystems services (Moberg and Folke, 1999; Birkeland, 2015; Norström et al., 2016). This global trend is due to a combination of anthropogenic stressors acting both on large scales (e.g., ocean acidification, sea level rise, and increases in seawater temperature) as well as locally (e.g., dredging, pollution, and overfishing). Of even greater concern are the cumulative interactions between these multiple stressors, leading to quicker and more severe reef degradation than previously anticipated (Pendleton et al., 2016) and the loss of coral reef resilience: the capacity of an ecological system to persist with change (through resistance and/or recovery) and to retain essential structures, processes and feedbacks (Walker et al., 2004; Folke et al., 2010).
Pelling (2011) suggested that managing ecosystems with a resilience focus facilitates the protection of important ecosystem functions in the face of external threats. Lately, resilience has become a conceptual cornerstone in the management of tropical coral reefs (Darling and Côté, 2018) and one approach to confronting the coral reef crisis is to consider coral reefs under the lens of ecological resilience. From a resilience-based management perspective and in order to overcome “wicked” resilience dynamics (sensu Glaser et al., 2018), it is key to understand changes in critical feedbacks of the system, which diminish coral reef resilience and drive the reorganization to undesirable and strongly resilient states (Walker et al., 2004; Graham et al., 2013; Glaser et al., 2018).
Loss of resilience in these ecosystems is generally associated with a reduction in coral cover and a general increase in fleshy macroalgae cover, referred to as a coral-macroalgal phase shift (Done, 1992; Bellwood et al., 2004), with higher severity of these phase shifts in the Caribbean as compared to the Indo-Pacific (Connell, 1997; Mumby et al., 2007; Bruno et al., 2009). Before 1980, the upper reef slopes of most Caribbean reefs were dominated by the fast-growing Acropora spp. (Bellwood et al., 2004). Currently, however, 80–98% of these species’ regional populations have been decimated (Kline and Vollmer, 2011). Furthermore, the Caribbean is a region that might be fundamentally predisposed to macroalgal growth, which can partly be explained by a combination of: (i) stochastic events, such as the pervasive role of disease dramatically reducing acroporid corals (Aronson and Precht, 2001) and Diadema urchins (Lessios et al., 1984); (ii) bottom-up nutrification effects, such as inadequate wastewater management, coastal development, poor land cultivation practices and even Saharan dust (Mora, 2008; Roff and Mumby, 2012; Martínez-Rendis et al., 2016; Arias-González et al., 2017); and (iii) the limited scope for top-down control, in terms of low herbivore and predator biomass and diversity (Mumby et al., 2012; Jackson et al., 2014). Moreover, these cumulative factors may help explain the low coral resilience of the region, increasing vulnerability shifting from a coral state to a stressed coral state (sensu Bellwood et al., 2004) or to a depauperated coral state (sensu Mumby, 2009).
However, not all reefs have experienced a shift in benthic community. Some “reefs of hope” (sensu Darling and Côté, 2018) in the Caribbean still harbor high coral cover, including Acropora spp., particularly in specific sites of the Mesoamerican Reef (MAR), located in the Caribbean waters of Honduras, Guatemala, Belize, and Mexico. The MAR ecoregion has an overall land and sea surface area of 464,263 km2 (Kramer and Kramer, 2002) and includes a variety of marine and coastal ecosystems, including coral reefs, seagrasses, mangroves, and their associated watersheds. This region is considered to be one of the most complex coral reef systems in the Northern Hemisphere and since 2006, has been regularly monitored by the Healthy Reefs Initiative (HRI), in order to evaluate the health of its coral reefs (Mcfield et al., 2018).
Eutrophication (Arias-González et al., 2017) and loss of herbivore controls (Mumby, 2009, 2016) have been identified as drivers of fleshy macroalgal increase in the MAR, although not always producing a clear direct relationship (Suchley et al., 2016). Another less-explored question is the role of algal-gardening damselfish in amplifying macroalgal increases. Theoretically, overfishing leads to the direct loss of larger fishes (Glaser et al., 2018) and eventually, the loss of mesopredators such as small groupers and snappers when the system is heavily fished (Mumby et al., 2012). The loss of mesopredators can result in a greater number of algal-gardening damselfishes that cultivate more turf algae (Glaser et al., 2018). Damselfish gardening activity will increase nutrient loads and this mechanism will facilitate fleshy macroalgal cover (Blanchette et al., 2019), resulting in a decrease of hard corals and structural complexity (Glaser et al., 2018). Understanding these community drivers and feedbacks is valuable to support resilience-based management strategies, to develop practical tools for managers to anticipate ecosystem shifts (Norström et al., 2016) and promote strategic interventions to prevent undesirable processes that decrease coral reef resilience (Glaser et al., 2018).
This study integrates theoretical effects and feedbacks between key benthic and fish variables in different benthic states in the MAR (Figure 1). A coral state (Figure 1A) is characterized by adequate hard coral cover and low fleshy macroalgae cover, providing the three-dimensional spatial structure supporting diverse fish communities (Gratwicke and Speight, 2005; O’Brien and Scheibling, 2018), by mediating predator-prey interactions, enhancing food availability, as well as nursery and spawning sites (Mumby, 2016). Fish are known to be functionally important as they control other organisms through predation, mediate nutrient fluxes, and can act as ecosystem engineers (Villéger et al., 2017). Moreover, key processes that have been identified to contribute to coral reefs resilience include fish herbivory (Mumby and Harborne, 2010; Roff et al., 2019) and predators’ regulatory effects (Villéger et al., 2017).
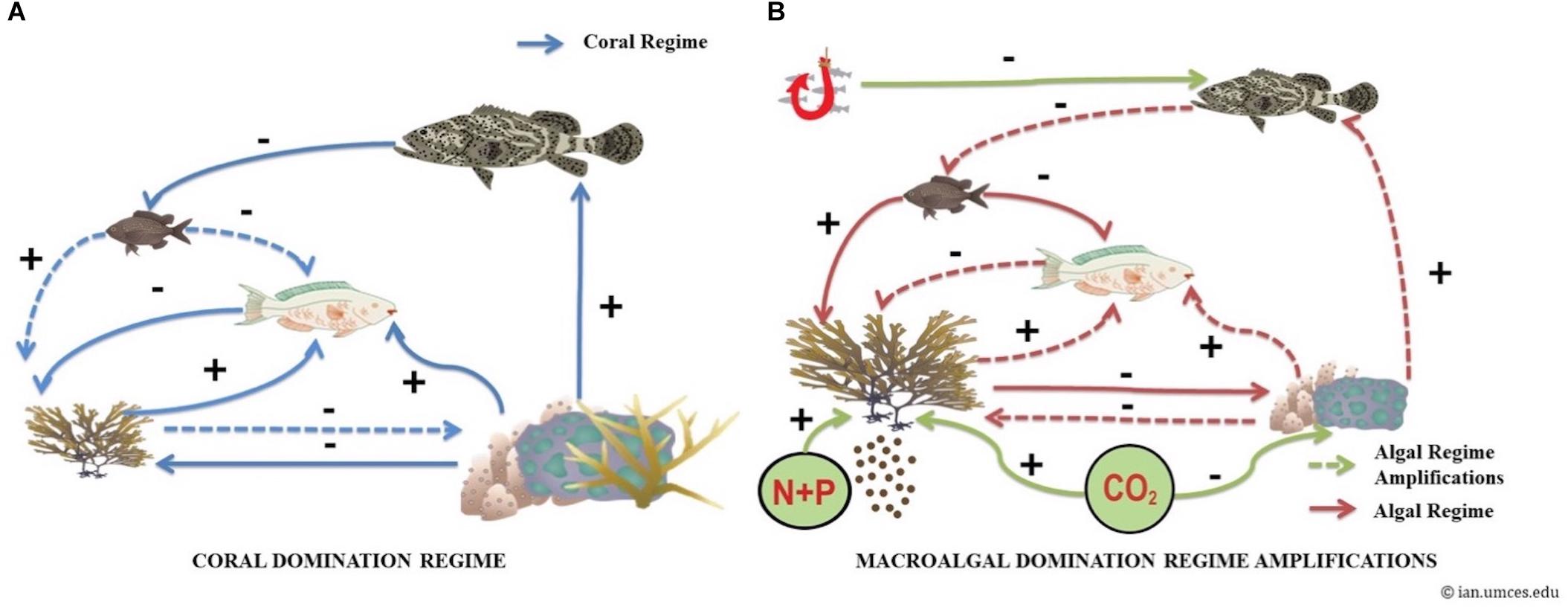
Figure 1. Feedbacks and interactions in two different coral states in the Mesoamerican Reef (MAR); (A) coral state (left) and (B) stressed coral state (right). The signs represent (+) positive (mutualism or commensalism) and (–) negative (competitive or predation) effects. The solid arrows represent strong effects and the dashed arrows weak ones. Blue arrows describe the effects and feedbacks in a healthy coral state. Green arrows describe the drivers inducing a loss of coral reef resilience in the system when levels of CO2, nutrients (Nitrogen and Phosphorus: N+P) and fishing pressure (represented by a red fishhook) increase. Red arrows describe the stressed coral state effects and feedbacks, with a positive feedback loop between macroalgae, damselfish, and herbivorous fishes, amplifying coral reef resilience loss.
A stressed coral state (Figure 1B) is driven by bottom-up feedback control, such as nutrients/sediments (Smith et al., 2010) and CO2 (Hughes et al., 2018) and/or top-down feedback control such as fishing pressure increases, provoking negative trophic cascades effects (Mumby et al., 2012). This change from a coral state to a stressed coral state (i.e., coral reef resilience loss) will weaken the positive effect of corals over fish by eroding habitat complexity, thereby diminishing the herbivores’ regulatory effects on turf and fleshy macroalgae (Suchley et al., 2016), and the predator effects on algal-gardening damselfish (Vermeij et al., 2015), which will increasingly amplify the stressed state and decrease the coral reef system’s resilience (Glaser et al., 2018). Indeed, the territorial and gardening activities of some damselfish species have been recognized to cause direct mortality to corals (Schopmeyer and Lirman, 2015) and once algal lawns become too large, they fuel the overgrowth by larger undesirable macroalgae (Schopmeyer and Lirman, 2015).
The main objective of this study, which uses HRI long term spatially expansive dataset, is to examine the association of key functional fish groups (algal-gardening herbivores, grazers, scrapers, browsers, and resident predators) and different coral states, hypothesizing that some functional fish groups may amplify reef degradation (i.e., coral reef resilience loss) in the Mesoamerican Reef (MAR).
Materials and Methods
Study Site and Data Collection
All data were collected according to the Atlantic and Gulf Rapid Reef Assessment (AGRRA) protocol (Marks, 2018), with trained survey teams coordinated by HRI. AGRRA protocol materials can be downloaded at http://www.agrra.org/training-tools/agrra-method/. Under this protocol, 300 different sites have been monitored in the MAR during five HRI surveys from 2006 to 2016. For this study 64 sampling sites were selected (Figure 2) because they fulfill two criteria: (1) they were monitored in 2006 and had live coral cover higher than 10% (positive accretion rate for the Caribbean according to Perry et al., 2013) and (2) they were monitored again in 2016. This timeframe is considered to be sufficient to study ecologically meaningful changes (Babcock et al., 2010).
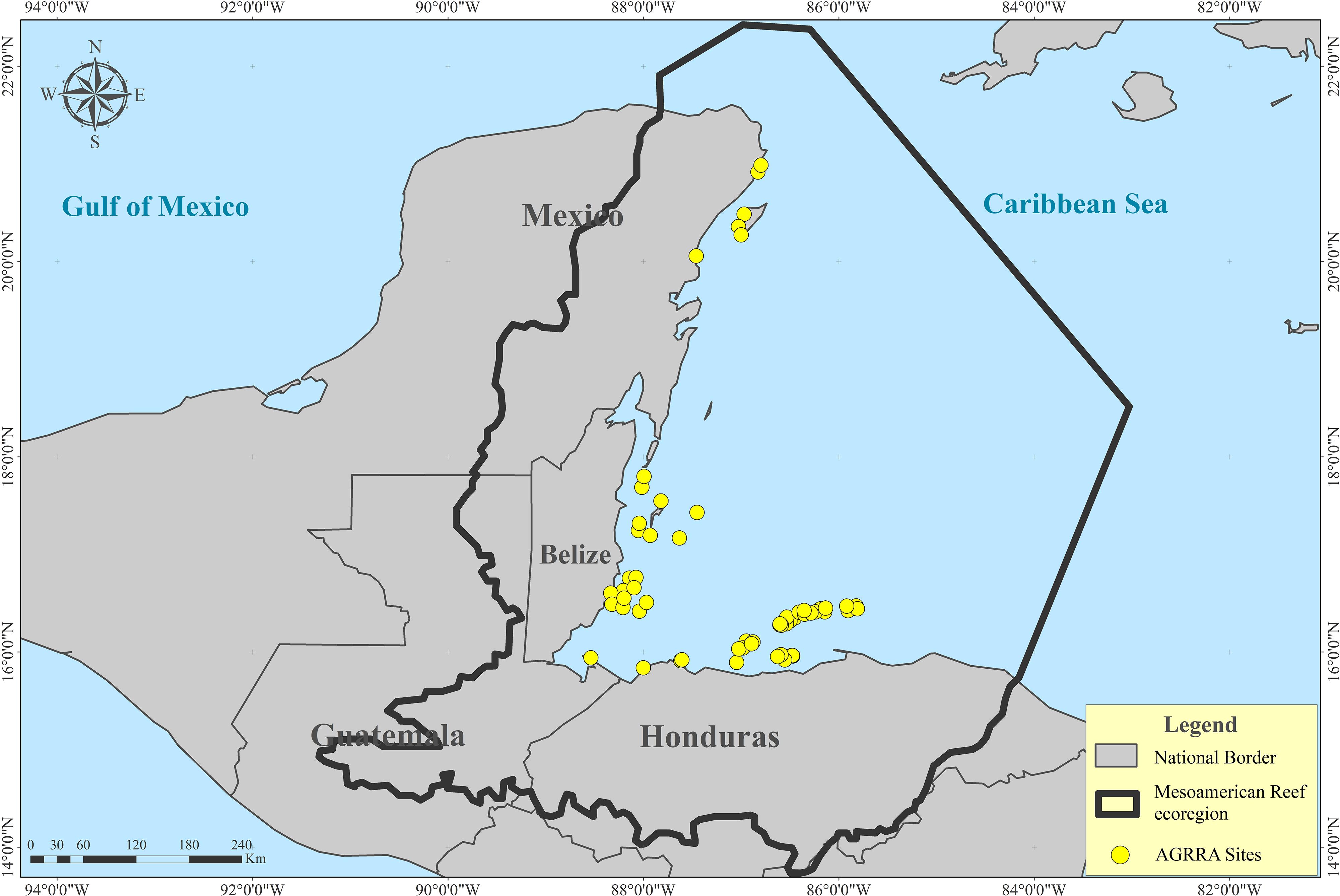
Figure 2. Sites selected (n = 64) in the Mesoamerican Reef ecoregion based on two criteria: (1) their live coral cover was higher than 10% in 2006 and (2) they were monitored both in 2006 and in 2016 by the Healthy Reefs Initiative.
The monitoring was canceled in poor visibility conditions (established to <5 m). The amount of inter-observer variability was reduced by the standardized aspect of the methodology in space and time, and because local biologists were trained during an entire week training course, including lectures and testing in theory and in the field, in order to conform to the protocol. Each AGRRA site is situated in a homogeneous geomorphic zone of a reef on an insular or continental shelf, with a surface of roughly 200 m × 200 m (Marks and Lang, 2018). These sites were selected through stratified randomized sampling, based on reef area extension and habitat (e.g., cross-shelf position, orientation, depth, and slope) in order to be representative of the reef region studied (Marks and Lang, 2018). In this study, 33 sites are fore reefs, 14 are crests, 10 are patch reefs and 7 are banks. The average depth of the sites is 6.6 m with a range between 0.9 m and 19 m.
All AGRRA data are available online: http://www.agrra.org/database-and-related-products/ and MAR data is also displayed and available at www.healthyreefs.org under data explorer tool. The two AGRRA data products analyzed through this research include benthic cover collected through linear point intercept in six haphazardly chosen 10 m transects per site (each with 100 data points) and fish density collected through visual census along ten haphazardly chosen 30 m × 2 m belt transects per site. The benthic and fish data were selected for this study, as they are key variables linked to reef health (McField and Kramer, 2008) and to some extent may influence coral reef resilience in the MAR region, as proposed in Figure 1.
The averaged percentage of live hard coral cover and fleshy macroalgae cover were used to assess coral state per site. In addition, fish density averages (expressed as a number of individuals per 100 m2) were used to evaluate key fish community per site. The choice of fish variables includes prominent herbivorous fishes in the MAR, namely parrotfish (mainly scrapers), surgeonfish (grazers), chubs (browsers), and damselfish (territorial algal-gardening herbivores). In addition, the principal resident predator fish are groupers, snappers and invasive lionfish (with the latter suspected to have become increasingly functionally important since 2009). The coral, algae, and fish species included are presented in Supplementary Tables 1, 2.
Data Analysis
All data analysis was performed using R software 3.5.1 (R Core Team, 2019) and followed two steps for each site: (i) description of benthic state changes between 2006 and 2016; (ii) association of benthic states and fish key functional groups.
Regarding the first step, changes were examined in benthic cover (live coral vs. fleshy macroalgae cover). Benthic thresholds were used to characterize sites state in 2006 and 2016, using a >10% of live coral (LC), which was found to be the positive accretion rate for the Caribbean (Perry et al., 2013), and <5% of fleshy macroalgae (FMA), which characterized a healthy coral condition (Kramer et al., 2015). Sites were then separated into three categories: Coral State (CS) with >10% LC and <5% FMA, Stressed Coral State (SCS) with >10% LC and >5% FMA and, Depauperate Coral State (DCS) <10% LC.
Coral state describes a resilient state which in theory maintain the structure, functions, and feedbacks of a coral reef, while SCS is a coral state which has loss resilience, and as a consequence, the structures, functions, and feedbacks are degrading. Finally, in a DCS the structure, functions, and feedbacks have changed and are no longer similar to a coral reef. Moreover, for each coral state, LC and FMA cover mean and standard deviation (sd) were calculated. A permutational multivariate analysis of variance, function “adonis” from the “vegan” library (Oksanen et al., 2018), using 9999 permutations, was done to test coral state on both benthic variables (hard coral and fleshy macroalgae). Also, paired t-tests, function “t-test” from the “stats” library (R Core Team, 2019), were carried out comparing hard corals and macroalgae, between 2006 and 2016.
Regarding the second step aiming at exploring the association between benthic states and the density of key fish groups, sites were classified in CS, SCS, and DCS (n = 128), and the monitoring year was considered as a factor variable. For each key functional fish group, mean, and standard deviation (sd) density were calculated for coral state and stressed state (n = 122). Due to the unbalanced number of samples, a non-parametric alternative to the paired t-test was calculated to identify significative changes in fish density between Coral State and Stressed Coral State, using function “wilcox test” from the “stats” library (R Core Team, 2019).
Canonical Analysis of Principal Coordinates (CAP) was used to correlate each key functional fish group with benthic variables. CAP performs a constrained ordination analysis in two steps (Anderson and Willis, 2003). In a first step, a Principal Coordinate Analysis (PCoA) was computed, using the coefficient of percentage difference (Bray-Curtis), on the benthic data matrix, which was previously transformed using Hellinger transformation allowing to preserve Euclidian distances (Legendre and De Cáceres, 2013). In a second step a Redundancy Analysis (RDA) of the PCoA created above (acting as the response data) was run, constrained by each fish group density (Borcard et al., 2018; Oksanen et al., 2018). For the CAP analysis, we used the function capscale of the vegan library, which uses all axes with positive eigenvalues, and axes are weighted by corresponding eigenvalues so that the ordination distances are the best approximations of original dissimilarities (Borcard et al., 2018; Oksanen et al., 2018).
Finally, a classification tree model, which is a non-parametric regression model, was used to predict the coral state depending on fish variables, using rpart function in rpart library (Therneau and Atkinson, 2018). This analysis is defined by recursive binary partitioning of the dataset into subgroups that are successively more homogeneous in relation to the values of the response variable. At each partitioning step, one predictor variable is used, maximizing homogeneity in the resulting subgroups by minimizing the sum of squared errors. A cross-validation procedure is used to determine the optimum size of the tree, using only a subset of the available data, while the remaining subset is used to assess the performance of the assembled tree.
Results
Coral State Changes in the MAR
In 2006, all the 64 sites were selected based on their LC cover >10% in order to have an initial positive accretion rate. However, only 23% of the sites were in a coral state (CS), and 77% were in a stressed coral state (SCS), considering the <5% FMA threshold (Figures 3A,B). In contrast, in 2016 (Figures 3C,D), only 4% of all sites were found in the CS. There was an 8% increase in sites in SCS representing 85% of total sites and 9% of total sites moved to a DCS. Benthic variables (hard coral and fleshy macroalgae) had significant differences between the three states (p = 0.001). In summary, 27% of the sites (n = 17) have degraded (Table 1).
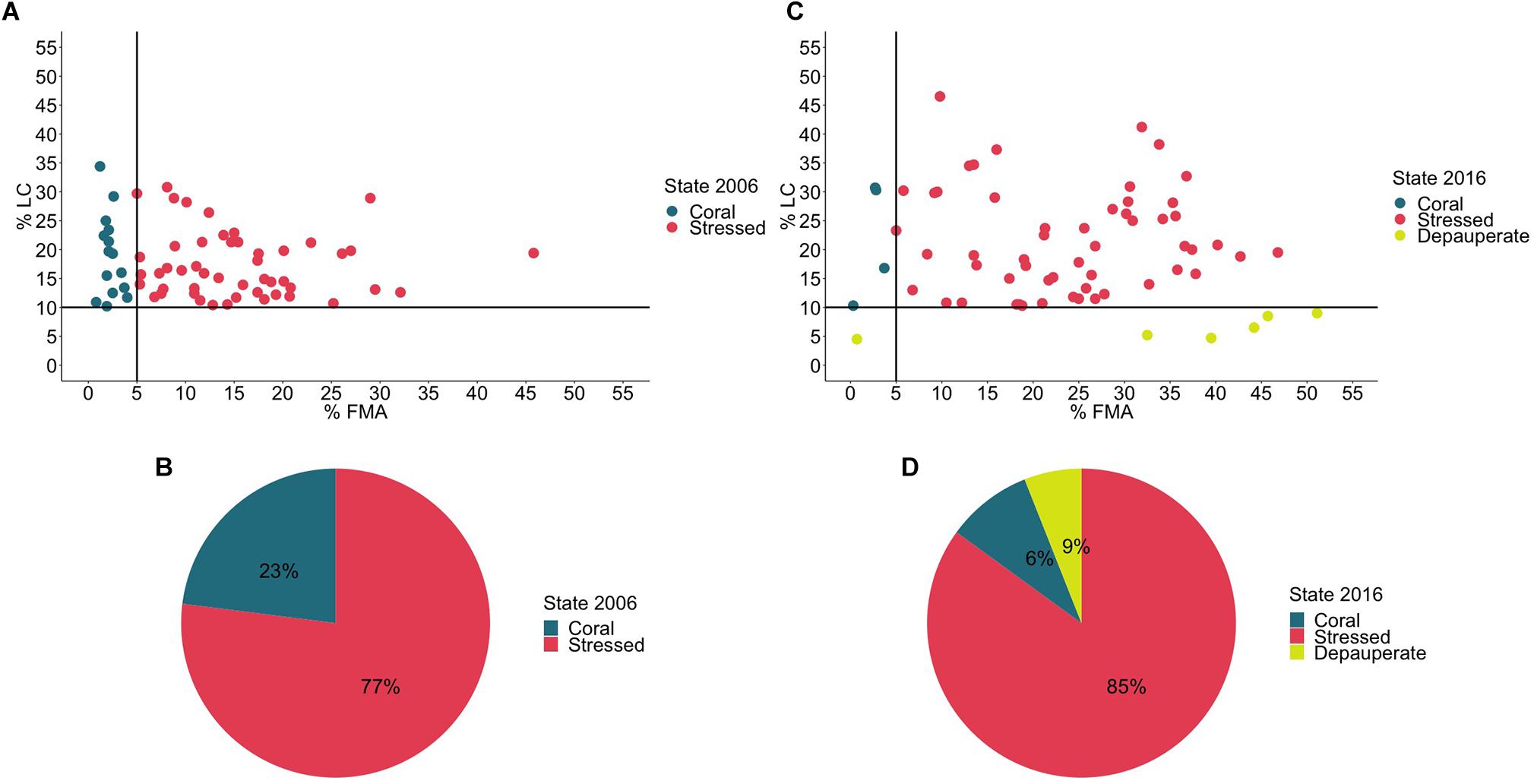
Figure 3. (A) Coral State in 2006 as a function of live coral (% LC) and fleshy macroalgae (% FMA) covers (top left). Sites in a coral state are in green and sites in a stressed coral state are in red. (B) Percentage of sites in a coral state and a stressed coral state in 2006 (bottom left). (C) Coral State in 2016 as a function of live coral (% LC) and fleshy macroalgae (% FMA) covers (top right). Sites in a coral state are in green, sites in a stressed coral state are in red and sites in a depauperate coral state are in yellow. (D) Percentage of sites in a coral state, a stressed coral state and a depauperate coral state in 2016 (bottom right).
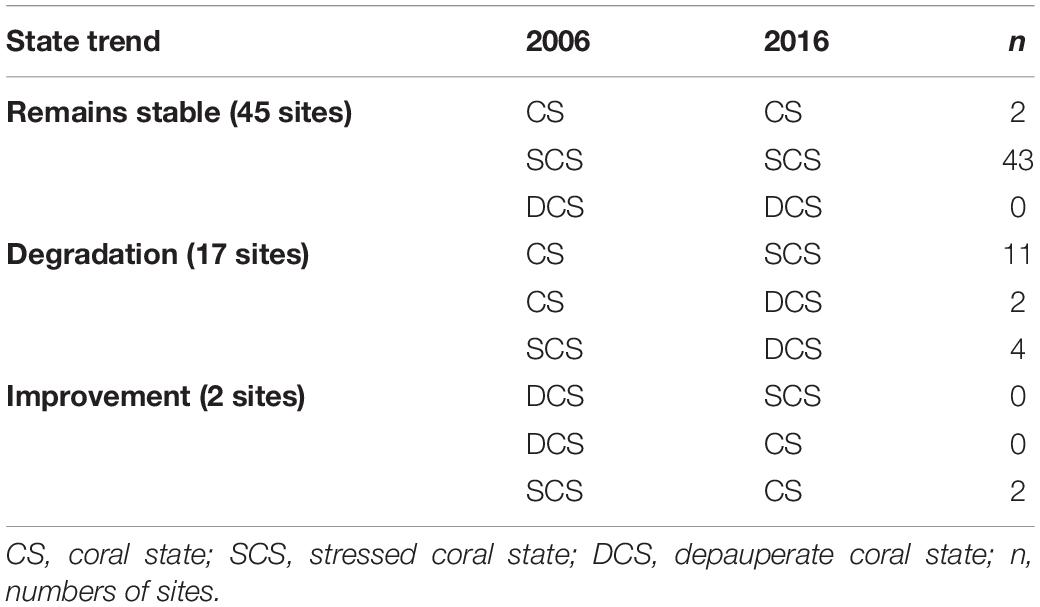
Table 1. Summary of state trends between 2006 and 2016 and the number of sites belonging to each trend.
In the 64 MAR sites included in this analysis, coral cover average was similar for both benthic states (Table 2), varying between 17.3% (±5.60) to 22% (±10.14). Yet a significative difference was found between hard corals in 2006 and 2016 (t = 2.0635, df = 63, p = 0.04319), with a mean increase of 3% in coral cover. Additionally, at sites belonging to a stressed coral state, mean fleshy macroalgae cover increased from 15.53% (±8.16) in 2006 to 23.76% (±10.44) in 2016 (Table 2). Effectively, for fleshy macroalgae cover a significant mean increase of 11% in the 10 years period was found (t = 7.2911, df = 63, p = 6.203e–10).
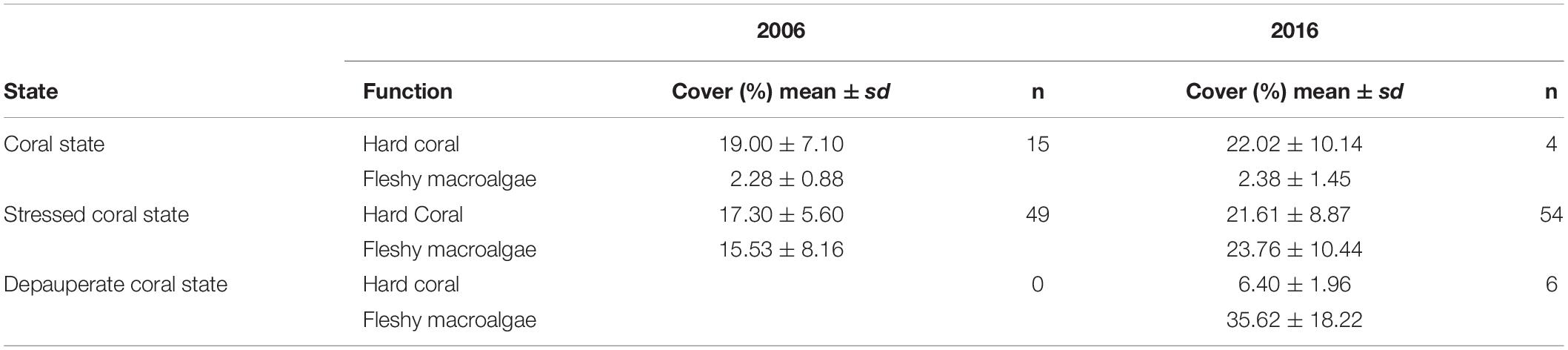
Table 2. The mean and the standard deviation (sd) of hard coral cover and fleshy macroalgae in 2006 and 2016 in the number of sites (n) belonging to each benthic state.
Association Between Benthic States and Key Functional Fish Groups in the MAR
Significative differences between coral and stressed coral states were found for algal-gardening damselfish (p = 0.004564) and parrotfish (p = 0.01818) and these groups were on average less abundant in sites classified as “coral state” than in a “stressed coral state” (Table 3). Other key functional groups were far less abundant, and exhibited similar density patterns in both states, as in the case of chubs, lionfish, and groupers (Table 3). However, fish density variability is generally high, particularly for herbivores belonging to parrotfish and surgeonfish families as these groups can either be seen solitary or forming schools.
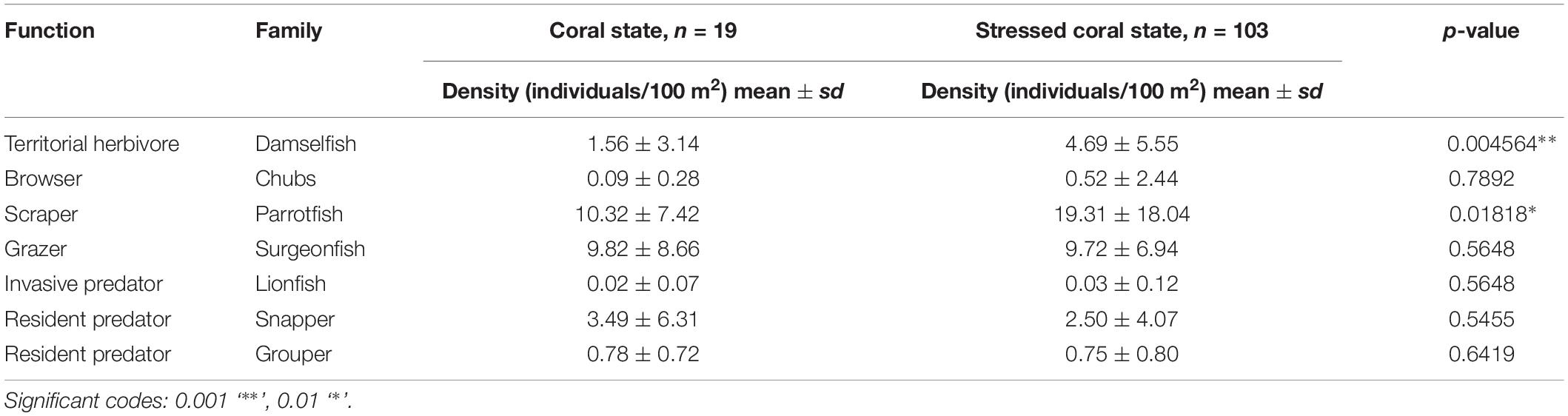
Table 3. Mean and standard deviation (sd) of key fish density in each coral state, present at an n number of sites for both years combined.
The fish groups that are associated significantly to the changes in benthic variables are in order of importance (Table 4): algal-gardening damselfish (9%), parrotfish (6%), and surgeonfish (5%). Algal-gardening damselfish (DAMS) and parrotfish (PARR) densities had a positive correlation with fleshy macroalgae cover (FMA) and were related to stressed coral state and the year 2016. On the other hand, snappers (SNAP) and surgeonfish (SURG) density had a positive correlation with coral cover (LC) and were related to coral state and the year 2006 (Figure 4). Depauperate state was not correlated with any fish density. Moreover, benthic cover variation is better explained by the state than by the year (Table 4).
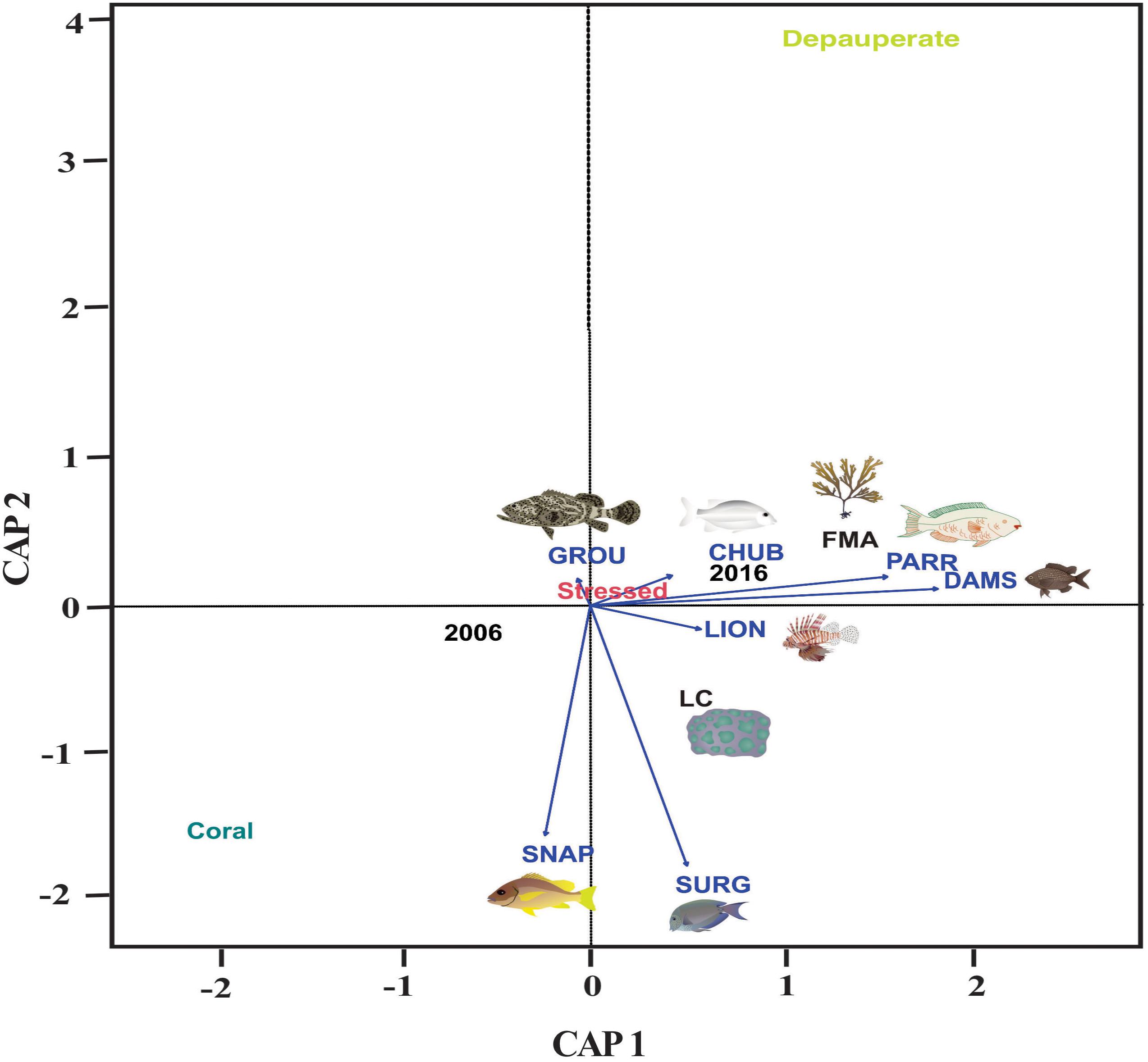
Figure 4. Canonical Analysis of Principal Coordinates of benthic cover dissimilarities represented by live coral (LC) and fleshy macroalgae (FMA) explained by key functional fish density, integrating the benthic state: coral, stressed or depauperate; and the monitoring year: 2006 or 2016. DAMS, algal-gardening damselfish; CHUB, chubs; PARR, parrotfish; SURG, surgeonfish; LION, lionfish; GROU, groupers; SNAP, snappers.
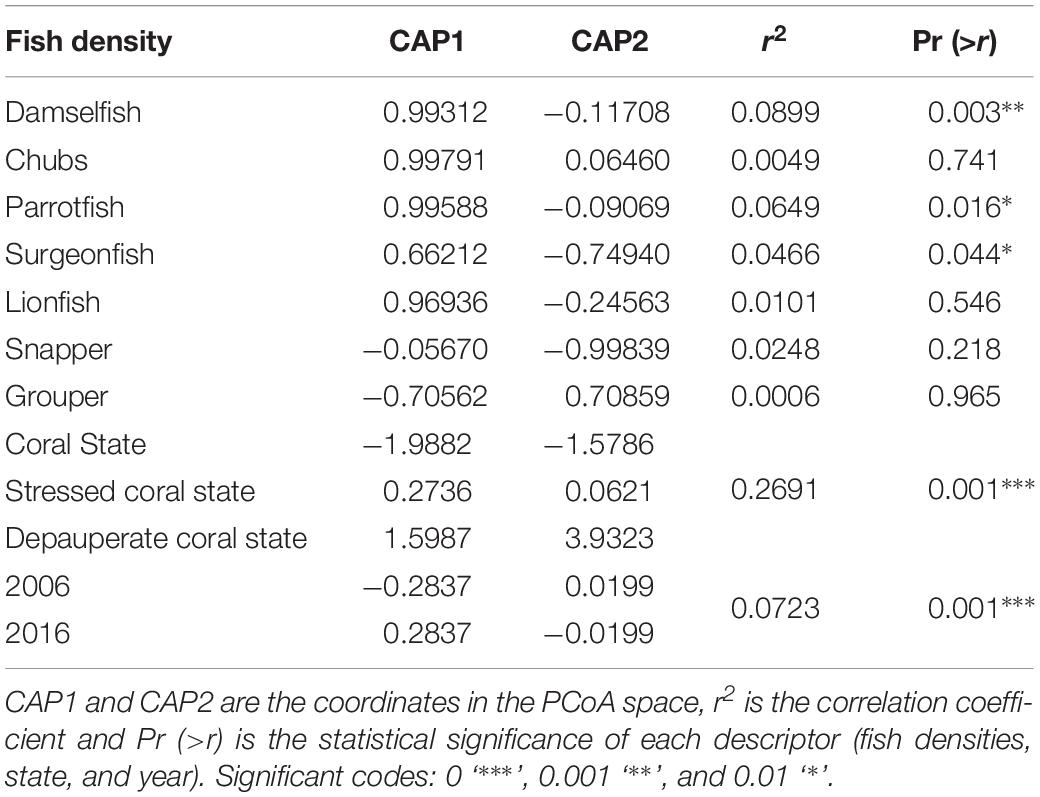
Table 4. Results of the Canonical Analysis of Principal Coordinates used to correlate each key functional fish group with benthic variables.
The more parsimonious classification tree model (Figure 5) includes just three functional herbivorous fish groups: algal-gardening damselfishes, scraper parrotfishes, and grazer surgeonfishes. It did not include chubs and resident predators (lionfish, groupers, and snappers), in order to enhance the model predictions. Higher values of damselfish predicted with a high level of accuracy (between 0.81 and 1.00 of accuracy) that a site belonged to a stressed coral state (Figure 5). Nevertheless, the prediction of a coral state with this model is not good (between 0.55 and 0.57 of accuracy). This could be related to the lower number of coral state sites (n = 19 out of 122).
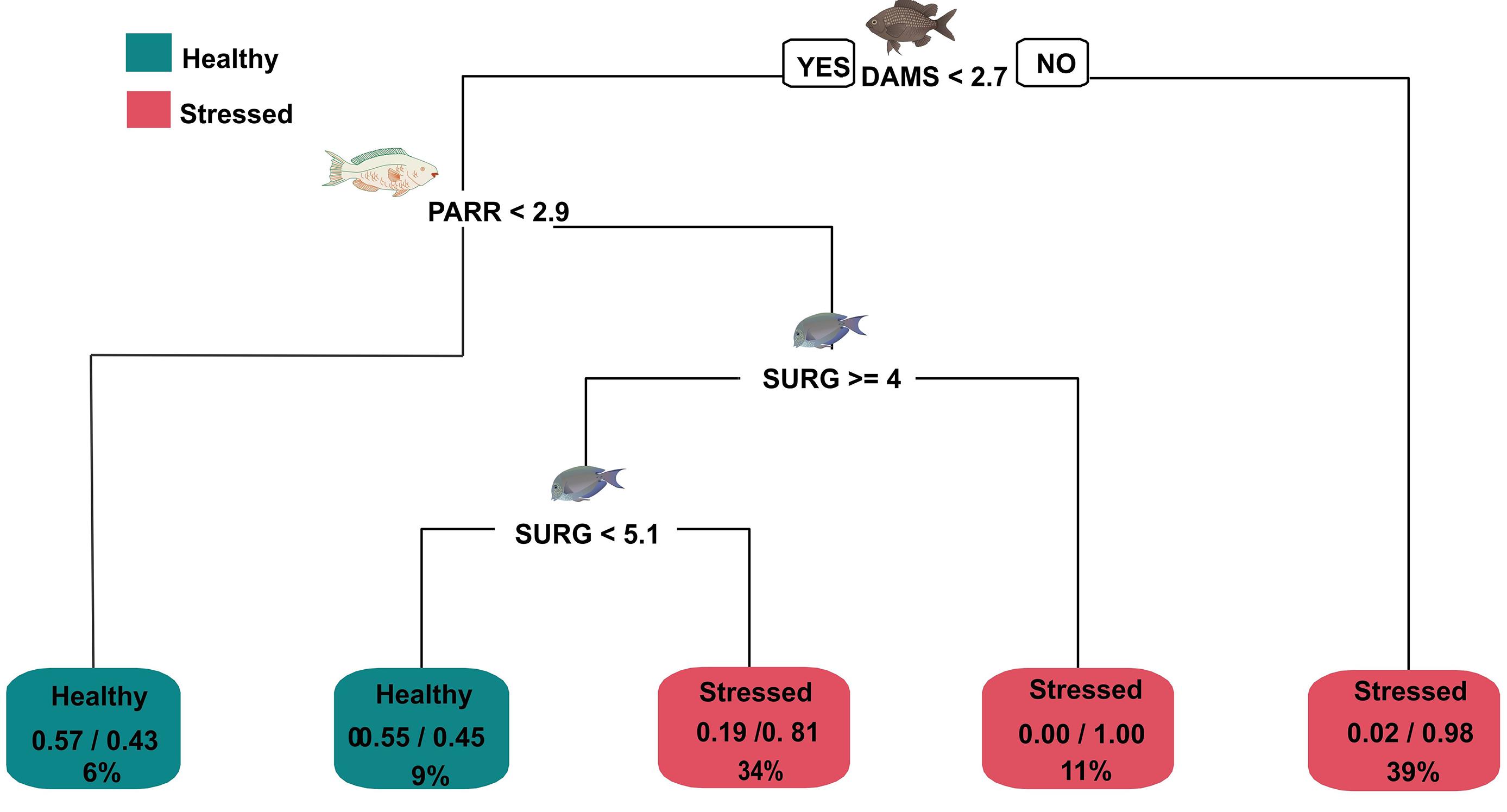
Figure 5. Classification tree model to predict the coral state (coral or stressed) explained by key functional fish density (number of individuals per 100 m2). DAMS, algal-gardening damselfish; PARR, parrotfish; SURG, surgeonfish. Numbers in the nodes correspond to fish density values when this condition is true, one must follow the left path of the classification tree model. The proportion between 0 and 1 in each rectangle correspond to the number of sites predicted in a coral state (left) and a stressed coral state (right) during the cross-validation. The percentage in the rectangle is the proportion of sites characterized by one of the states and the density fish conditions used in the construction of the classification model. The node error is 0.16, which means that 19/122 sites were not predicted in the state they really belonged.
Discussion
In contrast to the global context of coral reef ecosystem degradation, slight improvements in reef health have been reported in the MAR over the last decade when the full set of over 300 monitoring sites are included (2006–2016), linked with coral cover and herbivorous fish increases (Mcfield et al., 2018). These results are actually an encouraging example of the potential benefits of science-based management. Since the signing of the Tulum agreement in 1997, this high-level political collaboration has been reinforced by the growing number of partner institutions within HRI, cooperating with science-based reef management since 2004 (Mcfield et al., 2018). The one reef health indicator that consistently shows negative changes, is the fleshy macroalgal cover which increased two-fold from 12 to 23% over this decade and is a widely used characteristic of a stressed coral state (Mcfield et al., 2018).
Developing research on the drivers and feedbacks that maintain undesirable -less resilient- coral states is a field of opportunity to improve coral reef management (Nyström et al., 2012). Our study uses benthic thresholds established by previous studies (Perry et al., 2013; Kramer et al., 2015), to distinguish a desirable coral state from undesirable SCS or DCS. We highlight that most of the desirable coral state in 2006 have moved toward a stressed coral state, but have not changed still to a depauperated coral state and coral cover has even increased on average, which can be related to the management actions undertaken in the MAR. Three well documented anthropogenic drivers (Hughes et al., 2018) that push a coral state to lose resilience are pollution and overfishing (at the local scale), and climate change (at the global scale). However, less is known about the precise fish community drivers and feedbacks which favor fleshy macroalgal increases.
To improve our understanding of fish community effects on coral state changes, it is then fundamental to systematically follow benthic and fish community key variables over time and space as it is done in the MAR by HRI, and couple monitoring with additional experimentation and modeling. Despite the limitations of our analyses, which does not include all potential drivers of resilience loss in coral reefs such as nutrient enrichment, overfishing, and climate change stressors, some patterns have emerged. Our findings highlight differential associations between herbivores and resident predators to states with different degrees of coral reef resilience (a more resilient coral state and a less resilient stressed coral state), suggesting that some key functional fish play a more important role than others in influencing coral reef resilience.
For instance, algal-gardening damselfish density was correlated with fleshy macroalgae cover and predicted with accuracy in sites characterized as stressed. Moreover, snappers’ density was correlated to live coral cover, but other predators’ density had a very low correlation, independently of the site category (coral or stressed coral state). In the functional absence of other predators, snappers are likely playing an important role in regulating territorial algal-gardening damselfish and diminish a stress factor on the coral system. In addition, surgeonfish were also correlated to live coral cover, suggesting that this herbivorous group could be more capable than other herbivores of breaking algal-gardening damselfish territorial defenses, providing better control of turf algae and ultimately of fleshy macroalgae, associated with a stress state.
The abundance of damselfish has been increasing in Caribbean reefs (Ceccarelli et al., 2001), possibly due to the low abundance of damselfish predators, including groupers and snappers (Robertson, 1996; Mumby et al., 2012; Vermeij et al., 2015; Rivera-Sosa et al., 2018). Consequently, when predator abundance is low, it may lead to decreasing reef health (Vermeij et al., 2015). Overall, our results emphasize that territorial herbivores (algal-gardening damselfish) may play a key role in maintaining feedbacks toward fleshy macroalgae-dominated states (Arnold et al., 2010). There are multiple mechanisms through which algal-gardening damselfish can amplify macroalgal increases. These territorial fish can actively bite and remove living coral tissue, cultivating dense algal lawns on coral skeletons (Kaufman, 1977), and are important mediators for interactions between corals, algae, and herbivores (Precht et al., 2010). This gardening activity has many feedbacks which amplify fleshy macroalgal abundance and domination, by provoking changes in coral composition (Precht et al., 2010), promoting coral predation (Schopmeyer and Lirman, 2015), increasing nutrient loads (Blanchette et al., 2019), acting as reservoirs for microbes related to coral diseases (Casey et al., 2014), decreasing coral recruitment and deterring predators and herbivores (Aronson and Precht, 2001; Eurich et al., 2018).
The high abundance of these territorial algal-gardening damselfish may be an ecological indicator of coral reef’s ongoing degradation. Consequently, our study suggests that it is important to continue improving our understanding of all potential reef health stressors (e.g., the multiple effects of nutrification, fishing, and CO2 increase) which may provoke a loss of coral reef resilience, but it is also vital to pay attention to community interactions and feedbacks, which are direct consequences of the stressors affecting coral reefs and may amplify “wicked” resilient regimes dominated by, for example, fleshy macroalgae. Additional studies are needed to investigate interactive effects between damselfish and their coral hosts to determine potential thresholds for algal farming activities (Schopmeyer and Lirman, 2015).
We therefore recommend the integration of all territorial algal-gardening damselfish in future AGRRA monitoring (Stegastes planifrons, S. diencaeus, S. fuscus, and Microspathodon crysurus) as it would not require additional cost, as well as to create in situ experiments to assess the density of algal-gardening damselfish provoking benthic shifts in the MAR, in order to both define a threshold for remaining in a safe operating system (sensu Norström et al., 2016), and to advance our understanding of predator-prey interactions. Monitoring and experimental studies coupled with feedbacks and interactions modeling will lead to a more integrated understanding of coral reef resilience loss driven by communities in the MAR and will continue to contribute to adaptive management based on rigorous science. Moreover, in further studies, it will be interesting to go beyond fish communities’ interactions and integrate as well other key functional groups of the MAR, such as is the case of sea urchins and crabs, which are also known to play an important role in regulating fleshy macroalgae (Mcfield et al., 2018).
Finally, our research strengthens the recommendation made by HRI partners for the reinforcement of Marine Replenishment Zones (Mcfield et al., 2018), in order to promote a healthy population of resident predator fish (like groupers and snappers), which may in turn regulate algal-gardening damselfish populations to levels that can maintain the reef’s ecological integrity and reduce negative top-down cascade effects. In a context where reefs are affected by a cumulation of stressors and feedbacks, this recommendation may go hand-in-hand with other recommendations put forward by previous studies promoting the preservation of herbivorous assemblages (parrotfish and surgeonfish) and good water quality to maintain desirable reef conditions. All these actions reinforced by collaborative management may contribute to building overall coral reef resilience in the MAR.
Author Contributions
ÁRE and JA-G conceived the idea. MM provided the ecological data. ÁRE, JMM, and MM carried out the data analysis and interpretation of the data. ÁRE wrote the manuscript. JMM, MM, JM, and JA-G commented and revised the manuscript.
Funding
This work was funded by the Consejo Nacional de Ciencia y Tecnologiìa (CONACYT#721071/600297), the Coral Reef Alliance (CORAL), as well as the Summit Foundation and the Oak Foundation, towards data collection that is coordinated with the Healthy Reefs Initiative.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the reviewers for their valuable comments to improve this manuscript. We also thank J. B. Raina, A. Schaffer, R. Watt-Pringle, and the LEEAC team for their valuable comments on the manuscript. We thank J. E. Mendoza for his help in the preparation of Figure 2. We are grateful to the CONACYT, the Oak Foundation, and the CORAL for providing financial support. Our special thanks to the 72 partner organizations collaborating within the Healthy Reefs Initiative and the individuals who have assisted in the data collection, the HRI team for organizing the field monitoring expeditions (Ian Drysdale, Ana Giro, Marisol Rueda, Melina Soto, Roberto Pott, and Nicole Craig), and the AGRRA team (Ken Marks, Judith Lang, Patricia Kramer, and Lynette Roth).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00414/full#supplementary-material
References
Anderson, M. J., and Willis, T. J. (2003). Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84, 511–525. doi: 10.1890/0012-96582003084
Arias-González, J. E., Fung, T., Seymour, R. M., Garza-Pérez, R., Acosta-González, G., Bozec, Y.-M., et al. (2017). A coral-algal phase shift in mesoamerica not driven by changes in herbivorous fish abundance. PLoS One 12:e0174855. doi: 10.1371/journal.pone.0174855
Arnold, S. N., Steneck, R. S., and Mumby, P. J. (2010). Running the gauntlet: inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 414, 91–105. doi: 10.3354/meps08724
Aronson, R. B., and Precht, W. F. (2001). White band diseases and the changing face of Caribbean coral reefs. Hydrobiologia 460, 25–38. doi: 10.1007/978-94-017-3284-0_2
Babcock, R. C., Shears, N. T., Alcala, A. C., Barrett, N. S., Edgar, G. J., Lafferty, K. D., et al. (2010). Decadal trends in marine reserves reveal differential rates of change in direct and indirect effects. Proc. Natl. Acad. Sci. U.S.A. 107, 18256–18261. doi: 10.1073/pnas.0908012107
Bellwood, D. R., Hughes, T. P., Folke, C., and Nystrom, M. (2004). Confronting the coral reef crisis. Nature 429, 827–833. doi: 10.1038/nature02691
Blanchette, A., Ely, T., Zeko, A., Sura, S. A., Turba, R., and Fong, P. (2019). Damselfish Stegastes nigricans increase algal growth within their territories on shallow coral reefs via enhanced nutrient supplies. J. Exp. Mar. Biol. Ecol. 513, 21–26. doi: 10.1016/j.jembe.2019.02.001
Borcard, D., Gillet, F., and Legendre, P. (2018). Numerical Ecology with R. Berlin: Springer, doi: 10.1007/978-0-387-78171-6
Bruno, J. F., Sweatman, H., Precht, W. F., Selig, E. R., and Schutte, V. G. W. (2009). Assessing evidence of phase shifts from coral to macroalgal dominance on coral reefs. Ecology 90, 1478–1484. doi: 10.1890/08-1781.1
Bruno, J. F., and Valdivia, A. (2016). Coral reef degradation is not correlated with local human population density. Sci. Rep. 6:29778. doi: 10.1038/srep29778
Casey, J. M., Ainsworth, T. D., Choat, J. H., and Connolly, S. R. (2014). Farming behaviour of reef fishes increases the prevalence of coral disease associated microbes and black band disease. Proc. Biol. Sci. 281:20141032. doi: 10.1098/rspb.2014.1032
Ceccarelli, D. M., Jones, G. P., and McCook, L. J. (2001). Territorial damselfishes as determinants of the structure of benthic communities on coral reefs. Oceanogr. Mar. Biol. 39, 355–389. doi: 10.1016/j.chemphys.2005.03.003
Connell, J. H. (1997). Disturbance and recovery of coral assemblages. Coral Reefs 16, S101–S113. doi: 10.1007/s003380050246
Darling, E. S., and Côté, I. M. (2018). Seeking resilience in marine ecosystems. Science 359, 986–987. doi: 10.1126/science.aas9852
De’ath, G., Fabricius, K. E., Sweatman, H., and Puotinen, M. (2012). The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. U.S.A. 109, 17995–17999. doi: 10.1073/pnas.1208909109
Done, T. J. (1992). Phase shifts in coral reef communities and their ecological significance. Hydrobiologia 247, 121–132. doi: 10.1007/BF00008211
Eurich, J. G., Mccormick, M. I., and Jones, G. P. (2018). Habitat selection and aggression as determinants of fine-scale partitioning of coral reef zones in a guild of territorial damselfishes. Mar. Ecol. Prog. Ser. 587, 201–215. doi: 10.3354/meps12458
Folke, C., Carpenter, S. R., Walker, B., Scheffer, M., Chapin, T., and Rockström, J. (2010). Resilience thinking: integrating resilience, adaptability and transformability. Ecol. Soc. 15:20. doi: 10.5751/ES-03610-150420
Gardner, T. A., Coté, I. M., Gill, J. A., Grant, A., and Watkinson, A. R. (2003). Long-term region-wide declines in caribbean corals. Science 301, 958–960. doi: 10.1126/science.1086050
Glaser, M., Plass-Johnson, J. G., Ferse, S. C. A., Neil, M., Satari, D. Y., Teichberg, M., et al. (2018). Breaking resilience for a sustainable future: thoughts for the anthropocene. Front. Mar. Sci. 5:34. doi: 10.3389/fmars.2018.00034
Graham, N. A. J., Bellwood, D. R., Cinner, J. E., Hughes, T. P., Norström, A. V., and Nyström, M. (2013). Managing resilience to reverse phase shifts in coral reefs. Front. Ecol. Environ. 11, 541–548. doi: 10.1890/120305
Gratwicke, B., and Speight, M. R. (2005). The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. J. Fish Biol. 66, 650–667. doi: 10.1111/j.1095-8649.2005.00629.x
Hughes, T. P., Barnes, M. L., Bellwood, D. R., Cinner, J. E., Cumming, G. S., Jackson, J. B. C., et al. (2017). Coral reefs in the Anthropocene. Nature 546, 82–90. doi: 10.1038/nature22901
Hughes, T. P., Kerry, J. T., Baird, A. H., Connolly, S. R., Dietzel, A., Eakin, C. M., et al. (2018). Global warming transforms coral reef assemblages. Nature 556, 492–507. doi: 10.1038/s41586-018-0041-2
Jackson, J. B. C., Donovan, M. K., Cramer, K. L., and Lam, W. (2014). Status and Trends of Caribbean Coral Reefs: 1970-2012. Global Coral Reef Monitoring Network. Gland: IUCN.
Kaufman, L. (1977). “The Three Spot Damselfish: Effects on Benthic Biota of Caribbean Coral Reefs,” in Third International Coral Reef Symposium, ed. D. L. Taylor (Miami, FL: Rosenstiel School of Marine and Atmospheric Science), 559–564.
Kline, D. I., and Vollmer, S. V. (2011). White band disease (Type I) of endangered caribbean acroporid corals is caused by pathogenic bacteria. Sci. Rep. 1:7. doi: 10.1038/srep00007
Kramer, P., McField, M., Álvarez-Filip, L., Drysdale, I., Ruedas-Flores, M., Giro-Petersen, A., et al. (2015). 2015 Mesoamerican Reef Report Card. Available at: http://www.healthyreefs.org/cms/front-page-2/mar-en-small/
Kramer, P. A., and Kramer, P. (2002). Ecoregional Conservation Planning for the Mesoamerican Caribbean Reef. Gland: World Wildlife Fund.
Legendre, P., and De Cáceres, M. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. Lett. 16, 951–963. doi: 10.1111/ele.12141
Lessios, H. A., Robertson, D. R., and Cubit, J. D. (1984). Spread of diadema mass mortality through the caribbean. Science 226, 335–337. doi: 10.1126/science.226.4672.335
Marks, K. W. (2018). AGRRA Database, version (2018-03). AGRRA, Florida, USA. Available at: www.agrra.org/data-explorer/explore-summary-products (accessed January 2018).
Marks, K. W., and Lang, J. (2018). AGRRA Summary Products, version (2018-03). AGRRA, Florida, USA. Available at: www.agrra.org/data-explorer/explore-summary-products (accessed January 2018).
Martínez-Rendis, A., Acosta González, G., Hernández-Stefanoni, J. L., and Arias González, J. E. (2016). Quantifying the reefscape transformation of a coastal Caribbean coral reef during a phase shift and the associated coastal landscape change. Mar. Ecol. 37, 697–710. doi: 10.1111/maec.12334
McField, M., and Kramer, P. (2008). Healthy Reefs for Healthy People: A Guide to Indicators of Reef Health and Social Well-being in the Mesoamerican Reef Region. With contributions by M. Gorrez and M. McPherson. Miami, FL: Franklin Trade Graphics. Available at: http://www.healthyreefs.org/cms/wp-content/uploads/2012/12/2008-Report-Card.pdf
Mcfield, M., Kramer, P., Álvarez-Filip, L., Drysdale, I., Rueda-Flores, M., Giró-Petersen, A., et al. (2018). 2018 Mesoamerican Reef Report Card. Available at: http://www.healthyreefs.org/cms/wp-content/uploads/2012/12/2018-MAR-Report-Card-Web.pdf
Moberg, F., and Folke, C. (1999). Ecological goods and services of coral reef ecosystems. Ecol. Econ. 29, 215–233. doi: 10.1016/s0921-8009(99)00009-9
Mora, C. (2008). A clear human footprint in the coral reefs of the Caribbean. Proc. R. Soc. B 275, 767–773. doi: 10.1098/rspb.2007.1472
Mumby, P. J. (2009). Phase shifts and the stability of macroalgal communities on Caribbean coral reefs. Coral Reefs 28, 761–773. doi: 10.1007/s00338-009-0506-8
Mumby, P. J. (2016). Stratifying herbivore fisheries by habitat to avoid ecosystem overfishing of coral reefs. Fish Fish. 17, 266–278. doi: 10.1111/faf.12078
Mumby, P. J., and Harborne, A. R. (2010). Marine reserves enhance the recovery of corals on caribbean reefs. PLoS One 5:e8657. doi: 10.1371/journal.pone.0008657
Mumby, P. J., Hastings, A., and Edwards, H. J. (2007). Thresholds and the resilience of Caribbean coral reefs. Nature 450, 98–101. doi: 10.1038/nature06252
Mumby, P. J., Steneck, R. S., Edwards, A. J., Ferrari, R., Coleman, R., Harborne, A. R., et al. (2012). Fishing down a Caribbean food web relaxes trophic cascades. Mar. Ecol. Prog. Ser. 445, 13–24. doi: 10.3354/meps09450
Norström, A. V., Nyström, M., Jouffray, J., Folke, C., and Graham, N. A. J. (2016). Guiding coral reef futures in the Anthropocene. Front. Ecol. Environ. 14, 490–498. doi: 10.1002/fee.1427
Nyström, M., Norström, A. V., Blenckner, T., de la Torre-Castro, M., Eklöf, J. S., Folke, C., et al. (2012). Confronting feedbacks of degraded marine ecosystems. Ecosystems 15, 695–710. doi: 10.1007/s10021-012-9530-6
O’Brien, J. M., and Scheibling, R. E. (2018). Turf wars: competition between foundation and turf-forming species on temperate and tropical reefs and its role in regime shifts. Mar. Ecol. Prog. Ser. 590, 1–17. doi: 10.3354/meps12530
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2018). vegan: Community Ecology Package. R package version 2.5-1. Available at: https://CRAN.R-project.org/package=vegan
Pelling, M. (2011). Adaptation to Climate Change: from Resilience to Transformation. Abingdon: Routledge.
Pendleton, L. H., Hoegh-guldberg, O., Langdon, C., and Comte, A. (2016). Multiple stressors and ecological complexity require a new approach to coral reef research. Front. Mar. Sci. 3:36. doi: 10.3389/fmars.2016.00036
Perry, C. T., Murphy, G. N., Kench, P. S., Smithers, S. G., Edinger, E. N., Steneck, R. S., et al. (2013). Caribbean-wide decline in carbonate production threatens coral reef growth. Nat. Commun. 4, 1402–1407. doi: 10.1038/ncomms2409
Precht, W. F., Aronson, R. B., Moody, R. M., and Kaufman, L. (2010). Changing patterns of microhabitat utilization by the threespot damselfish, stegastes planifrons, on caribbean reefs. PLoS One 5:e10835. doi: 10.1371/journal.pone.0010835
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rivera-Sosa, A., Muñiz-Castillo, A. I., McField, M., and Arias-González, J. E. (2018). Unusual “Knob-Like Chimney” growth forms on acropora species in the caribbean. Front. Mar. Sci. 5:41. doi: 10.3389/fmars.2018.00041
Robertson, D. R. (1996). Interspecific competition controls abundance and habitat use of territorial caribbean damselfishes. Ecology 77, 885–899. doi: 10.2307/2265509
Roff, G., Bejarano, S., Priest, M., Marshell, A., Chollett, I., Steneck, R., et al. (2019). Seascapes as drivers of herbivore assemblages in coral reef ecosystems. Ecol. Monogr. 89, 1–18. doi: 10.1002/ecm.1336
Roff, G., and Mumby, P. J. (2012). Global disparity in the resilience of coral reefs. Trends Ecol. Evol. 27, 404–413. doi: 10.1016/j.tree.2012.04.007
Schopmeyer, S. A., and Lirman, D. (2015). Occupation dynamics and impacts of damselfish territoriality on recovering populations of the threatened staghorn coral, Acropora cervicornis. PLoS One 10:e0141302. doi: 10.1371/journal.pone.0141302
Smith, J. E., Hunter, C. L., and Smith, C. M. (2010). The effects of top–down versus bottom–up control on benthic coral reef community structure. Oecologia 163, 497–507. doi: 10.1007/s00442-009-1546-z
Suchley, A., McField, M. D., and Alvarez-Filip, L. (2016). Rapidly increasing macroalgal cover not related to herbivorous fishes on Mesoamerican reefs. PeerJ 4:e2084. doi: 10.7717/peerj.2084
Therneau, T., and Atkinson, E. (2018). An Introduction to Recursive Partitioning Using the RPART Routines. Available at: https://cran.r-project.org/web/packages/rpart/vignettes/longintro.pdf.
Vermeij, M. J. A., DeBey, H., Grimsditch, G., Brown, J., Obura, D., DeLeon, R., et al. (2015). Negative effects of gardening damselfish Stegastes planifrons on coral health depend on predator abundance. Mar. Ecol. Prog. Ser. 528, 289–296. doi: 10.3354/meps11243
Villéger, S., Brosse, S., Mouchet, M., Mouillot, D., and Vanni, M. J. (2017). Functional ecology of fish: current approaches and future challenges. Aquat. Sci. 79, 783–801. doi: 10.1007/s00027-017-0546-z
Keywords: coral reefs, resilience, herbivores, predators, damselfish, management
Citation: Randazzo Eisemann Á, Montero Muñoz JL, McField M, Myton J and Arias-González JE (2019) The Effect of Algal-Gardening Damselfish on the Resilience of the Mesoamerican Reef. Front. Mar. Sci. 6:414. doi: 10.3389/fmars.2019.00414
Received: 26 September 2018; Accepted: 04 July 2019;
Published: 26 July 2019.
Edited by:
Angel Borja, Centro Tecnológico Experto en Innovación Marina y Alimentaria (AZTI), SpainReviewed by:
Gabriel Grimsditch, United Nations Environment Programme, KenyaMaria C. Uyarra, Centro Tecnológico Experto en Innovación Marina y Alimentaria (AZTI), Spain
Copyright © 2019 Randazzo Eisemann, Montero Muñoz, McField, Myton and Arias-González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ángela Randazzo Eisemann, angela.randazzo@cinvestav.mx; Jesús Ernesto Arias-González, earias@cinvestav.mx