The current and future seasonal geographic distribution of largehead hairtail Trichiurus japonicus in the Beibu Gulf, South China Sea
 Pengli Xiong1,2
Pengli Xiong1,2  Youwei Xu1,2
Youwei Xu1,2  Mingshuai Sun1,2 Xingxing Zhou1,2
Mingshuai Sun1,2 Xingxing Zhou1,2  Peiwen Jiang1,2
Peiwen Jiang1,2  Zuozhi Chen1,2*
Zuozhi Chen1,2*  Jiangtao Fan1,2*
Jiangtao Fan1,2*- 1South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou, China
- 2Key Laboratory of Open-Sea Fishery Development, Ministry of Agriculture and Rural Affairs, Guangzhou, China
Largehead hairtail Trichiurus japonicus is a major commercial fish species in the Beibu Gulf of the northwestern South China Sea. Despite much effort to protect the fishery resource, the current stock of T. japonicus is overexploited. As the impacts of climate change unfold globally, seasonal changes in the distribution of largehead hairtail in the Beibu Gulf have not yet been clarified. Maximum entropy model based on mixed layer depth and salinity were projected onto seasonal habitat changes of T. japonicus in the Beibu Gulf under a current scenario and three different Representative Concentration Pathways (126, 370, 585) to evaluate geographic distribution changes under the different climate-change scenarios. The current geographic distribution results showed variation with seasonality, as the wintering population shifts toward the northeast. Under each of three SSP scenarios, there is higher risk to habitat suitability in the 2090s as compared with that in the 2050s. The disadvantage to T. japonicus distribution is greatest in winter under each of the three climate change scenarios, both in the short- and long-term. Potential suitable habitat distributions have a minor range extension in Representative Concentration Pathway 370–2050 winter, but in the rest of the scenes and years they contract to south of the Beibu Gulf. The overall results indicate that seasonal differences in suitable habitat should be considered to ensure effective planning of future management strategies for T. japonicus.
1 Introduction
Climate change is affecting the distributions of marine organisms on regional and global scales. It is expected that climate-driven changes in various environmental factors will alter the composition and distribution of marine biota to various extents (Davis et al., 2020; Hu et al., 2022). The Intergovernmental Panel on Climate Change predicts that by the end of the 21st century the average global temperature would increase by 1°C to 4°C (IPCC, 2007). Sea surface temperatures in China’s coastal waters rose at a rate of 0.27°C per decade from 1980 to 2020 (MNR, 2021). Changes in the distributions of marine species as a consequence of global warming are now documented in Chinese waters. For instance, a tropical threadfin bream, Nemipterus mesoprion, was newly found in the Beibu Gulf (Cai et al., 2020), and the flatfish Tosarhombus octoculatus, is previously recorded in the waters off Kochi Prefecture, Japan, has moved southward into the South China Sea (SCS) (Jiang et al., 2015).And some studies related to the impact of climatic change on species’ distribution reach similar conclusions.By the 2050s, some suitable fish habitats, such as for Japanese scad Decapterus maruadsi and Japanese jack mackerel Trachurus japonicus, are expected to shift northward under anticipated climate-change conditions (Hu et al., 2022). The warming ocean will drive a substantial shift in Japanese anchovy habitat in the China seas (Liu et al., 2020). Such changes are expected to lead to decreases in species richness in southern areas of China’s seas and increases in the northern areas. Thus, with climate change already affecting all oceans, it is necessary to consider how fish distributions might change in Chinese waters to promote adaptive conservation and management strategies (Zhang et al., 2020b).
The largehead hairtail Trichiurus japonicus is an economically important fishery species with a wide distribution along Chinese coast (Chen et al., 2019). The Chinese domestic annual marine catch of hairtails (Trichiurus spp.) in 2020 was 903,345 tons, which accounted for ~14% of the total fish production in China, and the species ranked top in fish catches of both the northern SCS and nationwide (MARA, 2021). The importance of the hairtail fishery has resulted in considerable conservation measures. For example, to protect fish spawning stocks and juveniles, the SCS fishery was closed for 2 months yearly (in June and July) from 1999 to 2008, and the closure has been prolonged to 3.5 months yearly (May to August) since 2017 (Zhang et al., 2022a). Despite these efforts, the current stock of T. japonicus in the Beibu Gulf is overexploited, and its length frequency now tends to simplification and miniaturization (Shi et al., 2020; Zhang et al., 2022b). Fully understanding the habitat environments of T. japonicus in the Beibu Gulf and its likely responses to climate-induced environmental changes is critical for sustainable exploitation and management of the fishery.
The Maximum Entropy model (MaxEnt) is the most important statistical method for ecological modelling that is widely used to predict the geographic distribution of a species based on areas of suitable habitat (Harte and Newman, 2014). Based on machine learning and maximum entropy theory, MaxEnt model use occurrence records and associated environmental data to infer the ecological needs of a species and then predict its potential distribution in the area studied (Phillips et al., 2006; Phillips and Dudík, 2008; Zhao et al., 2021). Compared with other models, MaxEnt shows accurate prediction capabilities in simulations and evaluations with presence-only data (Mamun et al., 2018), and is relatively robust even with small sample sizes (Zhao et al., 2022).
This study used a MaxEnt model that had been optimized by the ‘kuenm’ package to forecast the potential geographic distribution of T. japonicus in the Beibu Gulf under the current scenario and three future climate scenarios (SSP126, SSP370 and SSP585), in the 2050s (short-term) and 2090s (long-term). 2050s was defined as short-term change and 2090s was defined as long-term change in the present study.The aim was to project the potential suitable habitat of T. japonicus in the Beibu Gulf under current climatic conditions in different seasons, and to explore the important climatic factors limiting this species’ distribution. On the basis of the changes of mixed layer depth and salinity, we predict the potential suitable habitat for T. japonicus according to future global climate-change scenarios, and we discuss the changes in the habitat suitability distributions during three periods (current, 2050s, and 2090s). The present results may provide implications for investigations of variations in fishery resources in the context of climate change. And the study could provide important information for planning future preventive measures of T.japonicus fishery to cope with the impact of global warming.
2 Materials and methods
2.1 Fishery data
Catch data were collected through four independent surveys from 2014–2015 and used to identify fishing locations for T. japonicus in the Beibu Gulf, and the occurrence records were then used as presence points in MaxEnt models. The surveys were conducted between 18°00′–21°15′ N and 107°15′–109°30′ E (Figure 1). The fishery data were provided by the South China Sea Fisheries Research Institute, Chinese Academy of Fisheries Science and were split into four quarters: summer (July-August), autumn (October-November), winter (January-February), and spring (April-May). The survey vessel, with weight 242 tons and length 36.8 m, was equipped with a bottom trawl; the main dimensions of the net were 80.4 × 60.54 m, total length 60.58 m, and net cage mesh size 40 mm. Fishing locations with a 0.25° spatial resolution were divided into four grids of 0.05°, because a larger dataset for presence data allows for a greater range of features in MaxEnt. We removed presence points that were outside the study area and those in fishing locations without T. japonicus.
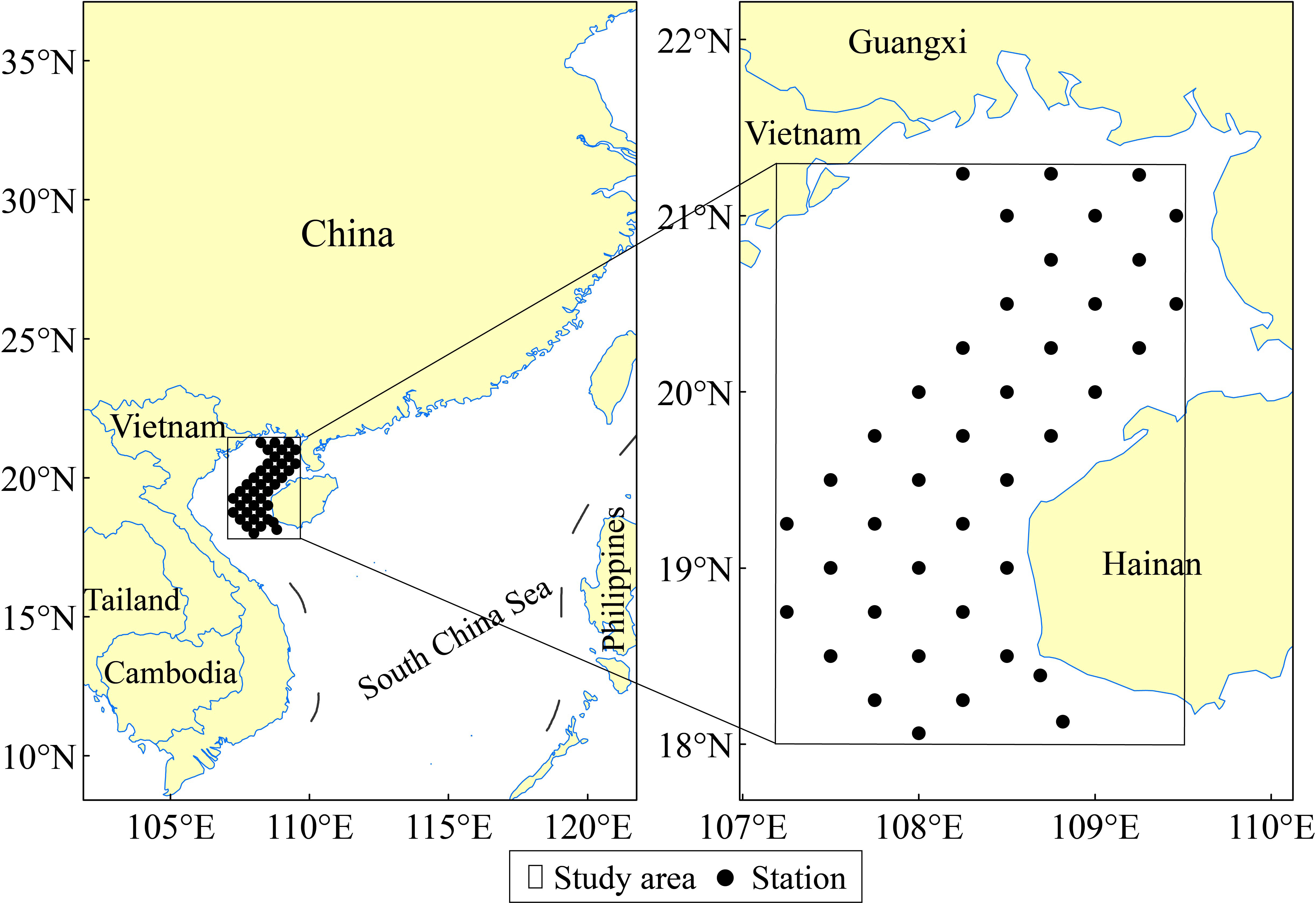
Figure 1 Maps showing the study area and stations where T. japonicus were sampled in the Beibu Gulf, South China Sea.
2.2 Collection and screening of environmental data
Environmental variables employed in prediction models for marine fishes include chlorophyll a, mixed layer depth, salinity, water temperature and depth (Montiel et al., 2019; Zhang et al., 2020a). Distance from land and sea surface temperature are known to influence marine fish in different ways, among which sea temperature is usually the dominant driver (Ma et al., 2019; Tian, 2020). The feeding activity of Trichiurus spp. in the Beibu Gulf was distributed from the middle part of the upper water layer to the bottom (Yan et al., 2012). Adults and juveniles of T. japonicus have opposing, complementary vertical diurnal feeding migration: large adults usually feed near the surface during the daytime and migrate to the bottom at night, while juveniles and small adults form schools near the bottom during the daytime and then loose feeding aggregations near the surface at night (Fishbase). In addition, the temperature difference between the surface and bottom water was identified as an important environmental factor in the reproductive process and early growth of T. japonicus (Yan et al., 2012). Therefore, we also selected the temperature in the 10–m and 20–m water layers, the bottom temperature, and the temperature difference between the surface and bottom water as model predictors. These satellite-derived oceanographic variables were obtained from official platforms (Table 1).
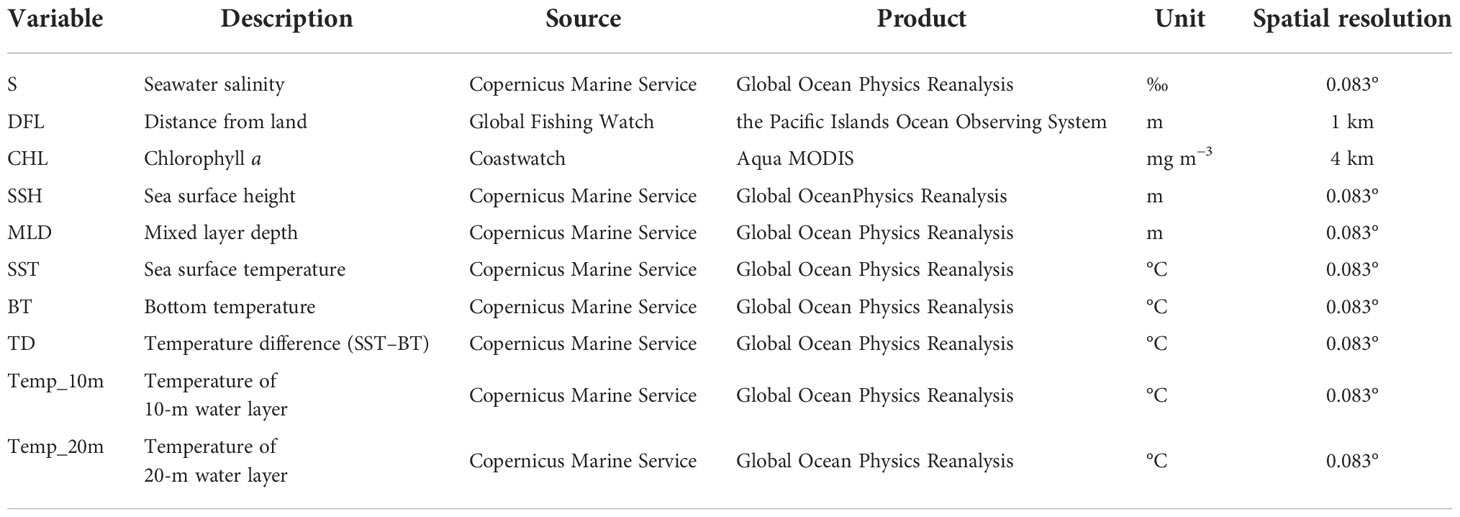
Table 1 Candidate factors for construction of the MaxEnt model for prediction of Trichiurus japonicus geographic distribution.
To keep the same resolution and achieve better accuracy, all environmental data were resampled using bilinear interpolation to 0.5° resolution. It is well documented that high collinearity of predictor variables can negatively affect a model’s prediction accuracy and the estimate of a variable’s effect (Xiao et al., 2019). Multicollinearity of variables were assessed with Pearson’s correlation analysis to confirm independence of variables in the study. Combined the results of the contribution rate of pre-experimental and Pearson’s results, compare with any two highly correlated environmental variables (r > 0.85), the variable that had a more vital contribution to the model was retained (Appendix A). variables were retained for prediction of suitable habitat of T. japonicus in four seasons: spring (S, DFL, CHL, MLD, SSH, TD, Temp_20m), summer (S, DFL, CHL, SSH, MLD, SST, BT, Temp_10m), autumn (S, DFL, MLD, TD), and winter (S, DFL, CHL, MLD, TD, Temp_20m).
2.3 Species distribution modeling
In this study, MaxEnt 3.4.4 was used to predict T. japonicus geographic distribution (Phillips et al., 2017). The feature class (FC) and regularization multiplier (RM) greatly influence the accuracy of the MaxEnt model (Phillips and Dudík, 2008). For both parameters, default settings based on MaxEnt outputs are sometimes unreliable, making the results difficult to interpret (Zhu and Qiao, 2016; Sill and Dawson, 2021). To arrive at an appropriate model, the FC and RM settings were adjusted using the kuenm R package (Cobos et al., 2019). For each seasonal model, we created 1,160 candidate models by combining 40 values of RM (0.1–4.0, at intervals of 0.1) and all 29 possible combinations of the five FCs (linear = l, quadratic = q, product = p, threshold = t, and hinge = h).
The selection of the final model was based on three main criteria: the partial ROC, omission rate, and delta Akaike Information Criterion (delta AICc). Evaluation of the models in terms of statistical significance rely on the partial ROC (p < 0.05) (Cobos et al., 2019). For each model in this study, the omission rate was >10%, suggesting overfitting of the model (Angeles-Gonzalez et al., 2021). The values of AICc and Delta_AICcwere used to search for the most parsimonious model with fewer fit parameters (Angeles-Gonzalez et al., 2021; Rodriguez-Burgos et al., 2022). Model with the lowest AICc value (i.e. ΔAICc = 0) is considered the best model out of the current suite of models (Muscarella et al., 2014).
To reduce the uncertainty of a MaxEnt model, 75% of the occurrence data are selected for training, and the remaining 25% are selected for testing. We selected the bootstrap method, with the average of 10 replicates used to determine the probable distribution of T. japonicus. Maximization of the sum of specificity and sensitivity was used to create habitat suitability maps using the binary choice of suitable or unsuitable (Lin et al., 2022). In MaxEnt models, the receiver operating characteristic (ROC) curve and the area under the ROC curve (AUC) are used to evaluate the precision of the model. Values of AUC range from 0 to 1, wherein 1 equals optimal performance and 0.5 denotes low predictive performance.
The value of the map of the MaxEnt model output was used to define habitat appropriateness. T. japonicus habitat appropriateness was categorized as follows: Unsuitable = a value of 0–0.25, indicating that the environment is not suitable for T. japonicus in theory; Marginally suitable = a value of 0.25–0.50, indicating that habitat elements are insufficient to allow persistent T. japonicus presence; Moderately suitable = a value of 0.50–0.70, indicating that the habitat characteristics support T. japonicus occurrence on a broad scale; or Highly suitable = a value of 0.70–1.0, indicating that the habitat characteristics enable T. japonicus presence at an ideal level.
2.4 Future projection of suitable habitat distribution
For future scenarios, three SSP scenarios from CMIP6 were selected for projecting changes in the potential distribution of T. japonicus in the Beibu Gulf under predicted future climate change. These scenarios are described as follows: (1) the SSP126 scenario is the low-end of the future scenario range, with radiative forcing stabilizing at 2.6 W/m2 in 2100; (2) the SSP370 scenario is a moderately stable scenario in which radiative forcing stabilizes at 7.0 W/m2 by 2100; and (3) the SSP585 scenario is considered a high radiative forcing scenario, whereby radiative forcing stabilizes at 8.5 W/m2 in 2100. The environmental variables in CMIP6 were extracted from the BCC-CSM2-MR climate model (Beijing Climate Center). To reduce bias yielded in the global climate model (GCM) projections, the delta method, a widely used bias-correction technique, was applied. Because we could not obtain all the future environmental factors selected in this study, the future modeling was carried out with only the MLD and salinity changes. The parameters of the future models remain the same as for the present models. The distribution models were projected onto climate conditions for comparison of potential suitable habitats under the present climate and three climatic scenarios in the future.
2.5 Changes in suitable habitat distribution and core distributional shifts
To examine the change trend in suitable habitat for T. japonicus in the Beibu Gulf, habitat loss or gain was calculated by combining the future binary maps and the current binary map using the Python-based ArcGIS toolkit SDM toolbox (Brown, 2014; Cong et al., 2020). A shifting map of distribution under each climate-change scenario was generated with the habitat range expansion, stable habitat, and habitat range contraction, respectively. Furthermore, for the sake of further exploring the dynamic migration paths, the centroids of future and current suitable areas were calculated and compared using the centroid changes tool in the SDM toolbox (Brown, 2014).
3 Results
3.1 Model parameter optimization
For each season, we selected the best-performing MaxEnt model based on the evaluation metrics. The best combination of parameters of RM and FC in the four seasons-spring, summer, autumn, winter-were 1.5*l, 2*t, 0.1*q, and 1.4*pt, respectively (Table 2).
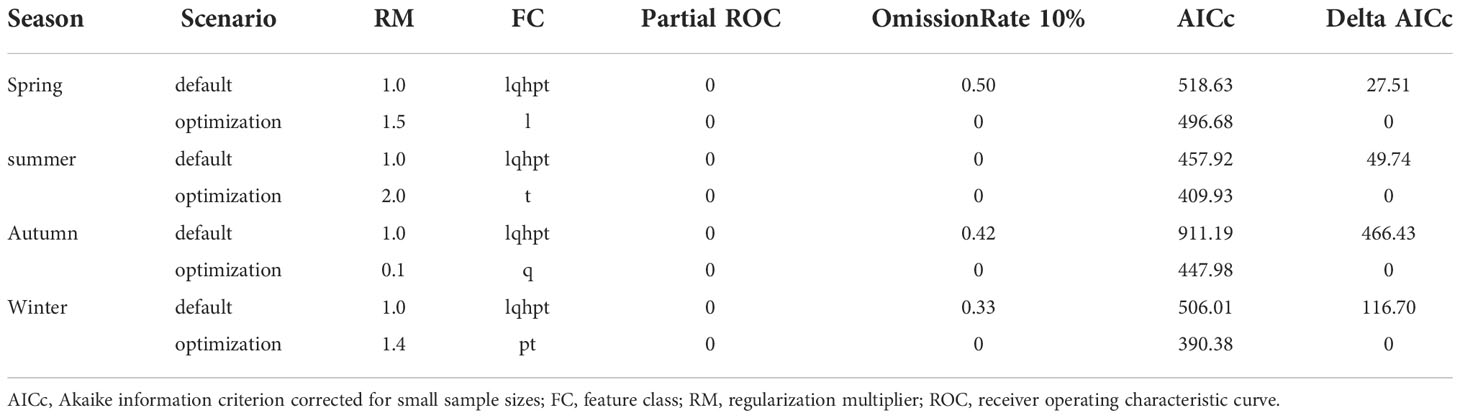
Table 2 MaxEnt model performance under default and optimized settings.
3.2 Current geographic distribution
MaxEnt performed well in generating the potential distribution of T. japonicus, with a mean AUC of 0.767 across the four seasons; logic thresholds of 0.463, 0.558, 0.430, and 0.437 were achieved for spring, summer, autumn, and winter, respectively. The current habitat suitability map for T. japonicus in the Beibu Gulf is shown in Figure 2. The results reveal significant seasonal variation in habitat selection by T. japonicus, manifested as relatively high suitable habitat area in spring and winter, and low suitable habitat area in summer and autumn.
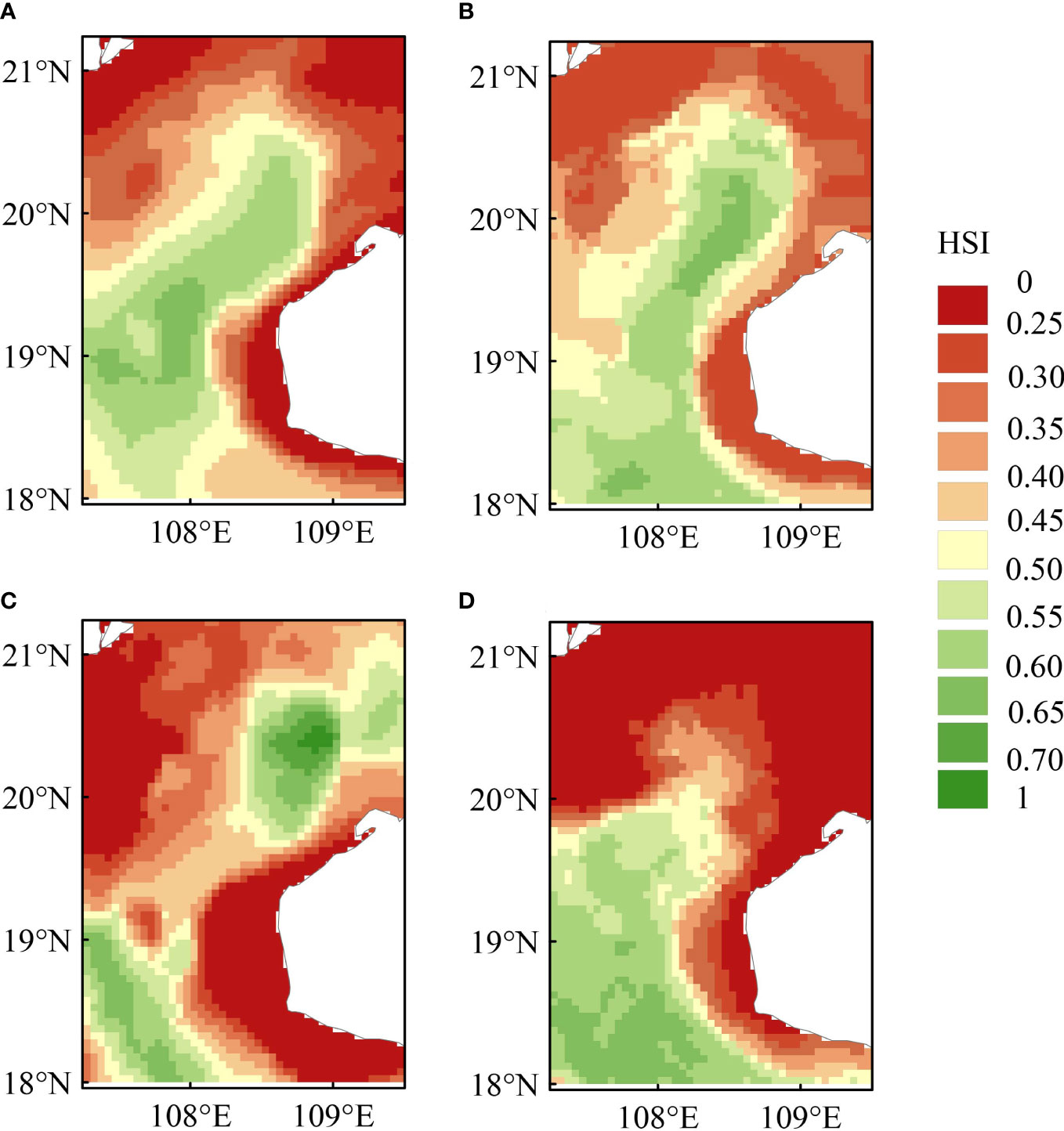
Figure 2 The probability of occurrence of Trichiurus japonicus in the Beibu Gulf, South China Sea, in different seasons: (A) spring; (B) summer; (C) autumn; (D) winter.
3.3 Jackknife test and response curves of the environmental variables
The jackknife test illustrated the contribution percentage of environmental variables of the predictive geographic distribution model (Figure 3). The importance of each variable and its relative contribution to the MaxEnt models differed between seasons. The results of the jackknife test in the models of the current scenario indicate the temperature difference between surface and bottom water contributed most in the spring model; the bottom temperature contributed most in the summer and autumn models; and mixed layer depth contributed most in the winter model.
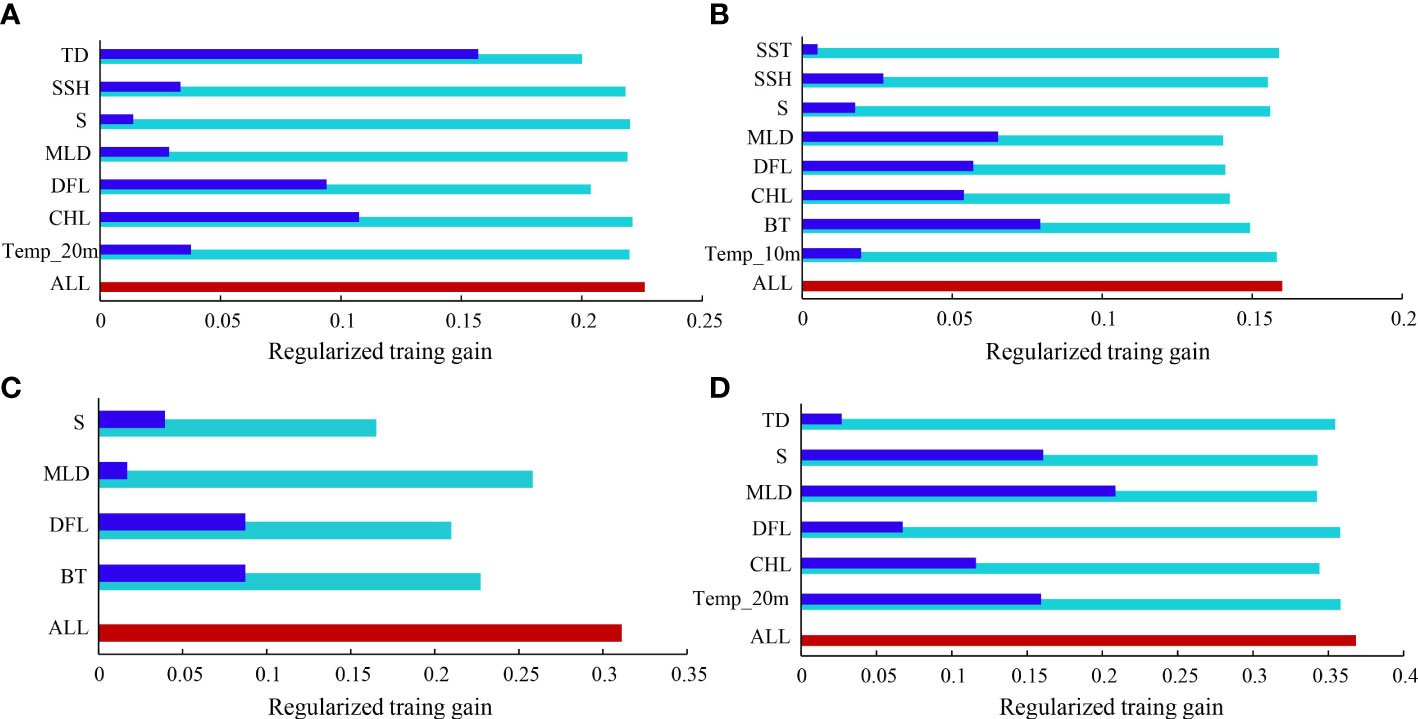
Figure 3 Jackknife test of environmental variable importance for training data in the current scenario model of Trichiurus japonicus distribution in the Beibu Gulf in different seasons: (A) spring; (B) summer; (C) autumn; (D) winter. Colors of the bars represent the contribution when the model is executed only with the variable, without the variable, or with all variables.
The MaxEnt model provided a response curve to show the relationship between the presence probability of T. japonicus and the environmental variables (Figure 4). Generally, when the presence probability is >0.50, the corresponding environmental factor is suitable for the species’ growth (Zhao et al., 2021). The probability of occurrence increased with the increase in TD in spring, and the optimum range was above 5.8 (with a probability occurrence value of >0.50). In relation to bottom water temperature, the largest probability of occurrence in summer was between 16.2 m and 20 m. In autumn, the BT was negatively correlated with habitat suitability, such that suitability was <50% when BT exceeded 23.8°C. The highest probability of T. japonicus occurrence was in areas experiencing a winter MLD of 28–35 m, and the optimum range was above 25.1 m. As the result in Table 3, Contribution (%) of S and MLD to the building of the Maxent models in winter greater than in the other seasons.
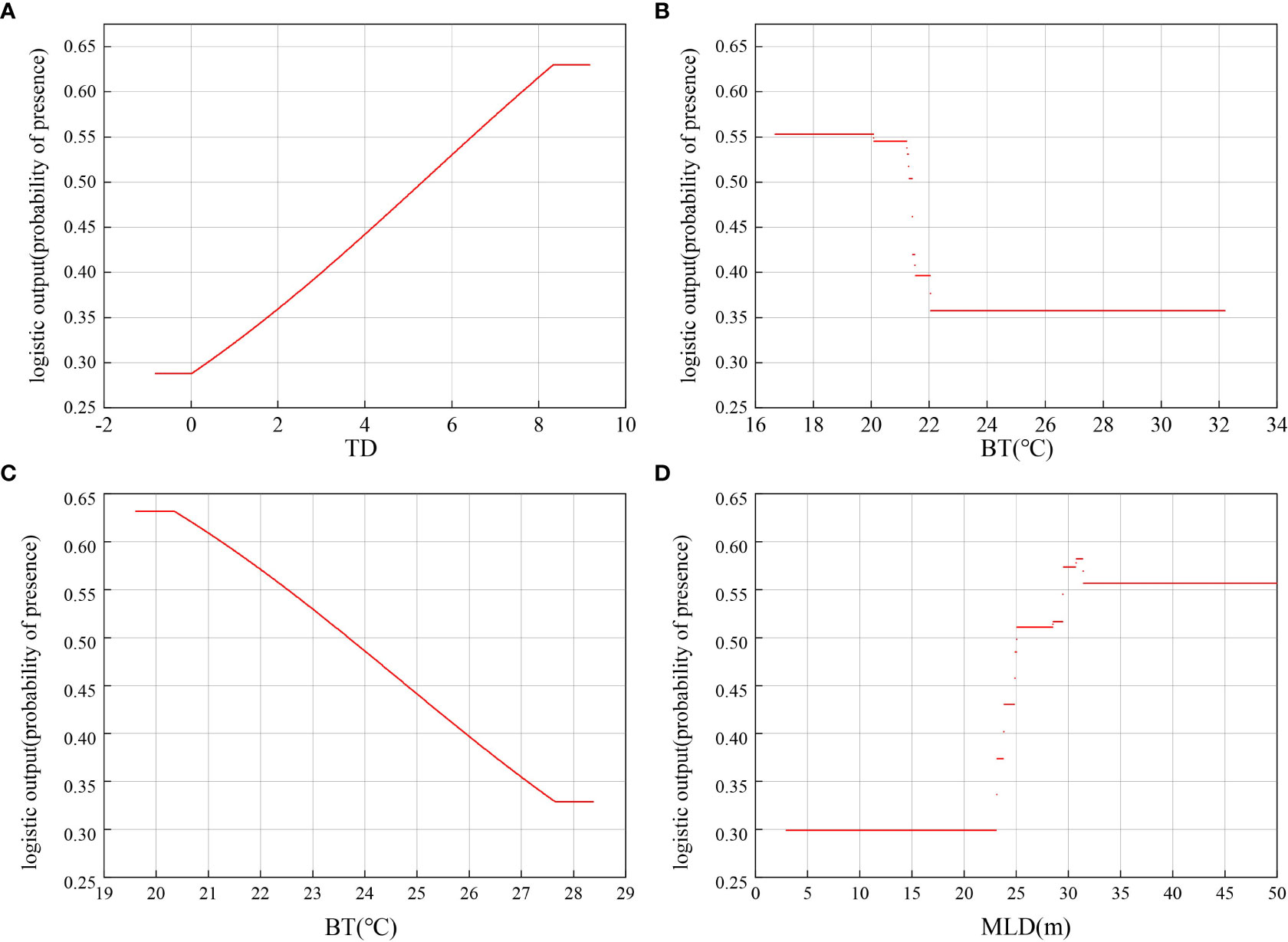
Figure 4 Relationship between the top environmental predictors and the probability of presence of Trichiurus japonicus in the Beibu Gulf under MaxEnt modeling of the current scenario: (A) spring temperature difference between surface and bottom water (TD); (B) summer bottom water temperature (BT); (C) autumn BT; (D) winter mixed layer depth (MLD).

Table 3 Estimates of relative contributions of the S and MLD to the Maxent model for each season, the best run of T. japonicus.
3.4 Changes in future potential suitable habitat
Based on MaxEnt modeling, T. japonicus in the Beibu Gulf shows both range expansion and contraction under the future climate scenarios; in Figures 5-7, blue and red represent estimated range reduction and expansion, respectively.
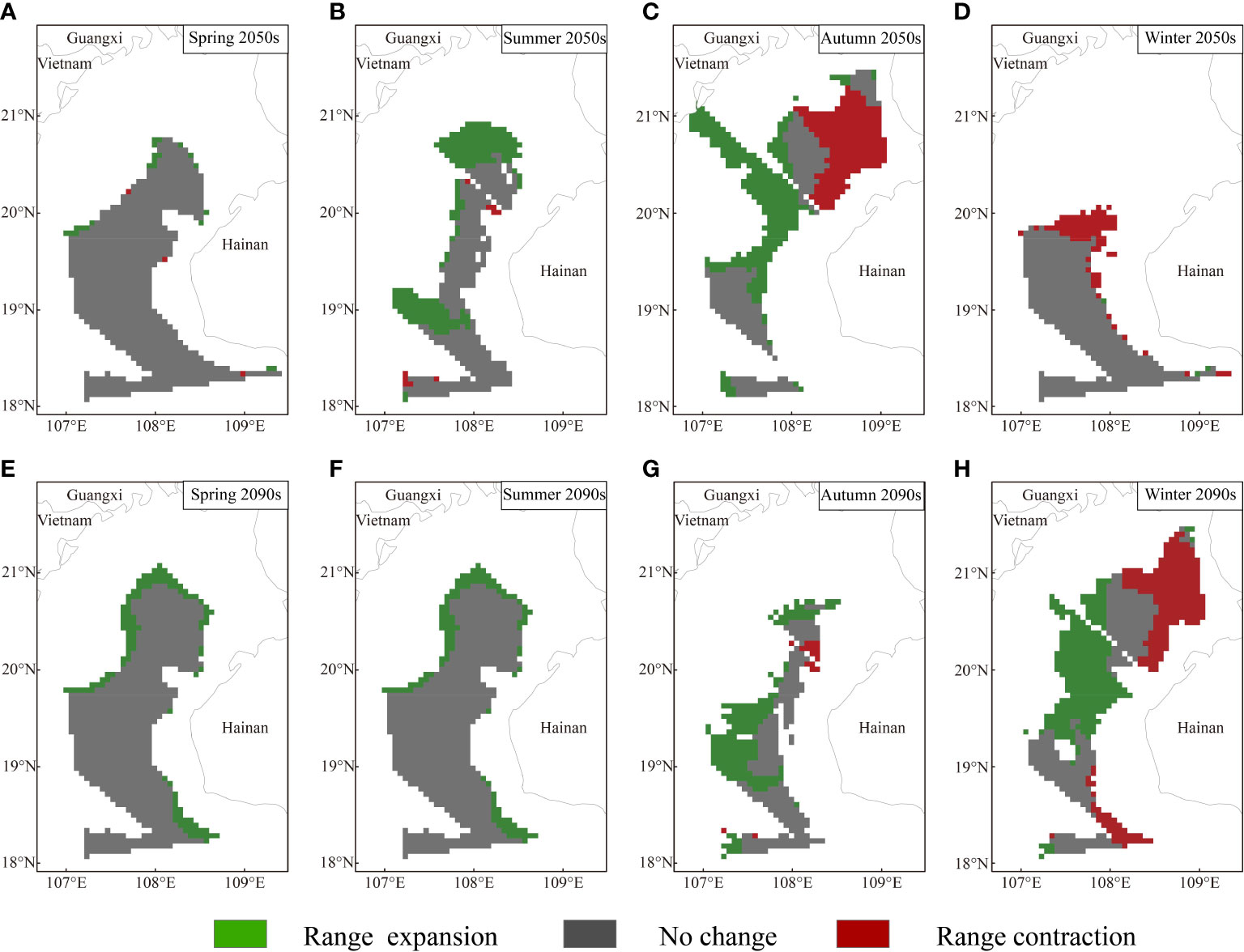
Figure 5 Projected range expansion or contraction of Trichiurus japonicus in the Beibu Gulf under RCP126. Area of geographic distribution changes among the species distribution models based on binary predictions for the current and future (i.e., in the 2050s and 2090s) climate conditions: (A) spring 2050s; (B) summer 2050s; (C) autumn 2050s; (D) winter 2050s; (E) spring 2090s; (F) summer 2090s; (G) autumn 2090s; (H) winter 2090s.
Under RCP126, the area of total potential distribution increased in all seasons except in winter, with the range-expansion predictions exceeding the range-contraction predictions (Figure 5). The short-term (2050s) scene shows changes of future potential suitable habitat in all seasons, wherein the potentially suitable habitats in spring show slight expansion in the margins of the northern parts of the current suitable habitat area. Range-expansion areas in summer could be divided as follows: a general trend of northward expansion of suitable habitat in the area near Guangxi, with suitable habitat mostly around 19°N, but expanding to the northwest. In autumn, northwestern areas of Hainan are characterized by a loss of suitable habitat areas, while gains in suitable area are observed in central parts of the study area. Areas that recognized as potentially suitable habitats in winter are located in southwestern parts of the study area, and the predicted decrease in suitable areas were located mainly in the northern parts of the current suitable areas. Compared with the findings in the 2050s, the proportion of range-expansion area decreases and the range-contraction area increases in the 2090s.
Under RCP370, especially in the 2090s, the projected risk increases for the distribution range of T. japonicus in the Beibu Gulf (Figure 6). In the 2050s, the overall suitable distribution area increases across seasons except for in autumn. However, the suitable habitat areas reflecting total range expansion are smaller in spring and winter, comprising 3.005% range-expansion area and 6.454% range-contraction area when compared with the total study area. In the 2090s, the potential distribution area based on all suitable habitat areas decreases in all seasons except winter. Moreover, among seasons, the suitable habitat area suffers the largest and most dramatic decline in summer, and the new expansion areas that were seen in the 2050s disappear. The expansion area of suitable distribution in the 2090s accounts for only 1.682% of the total study area.
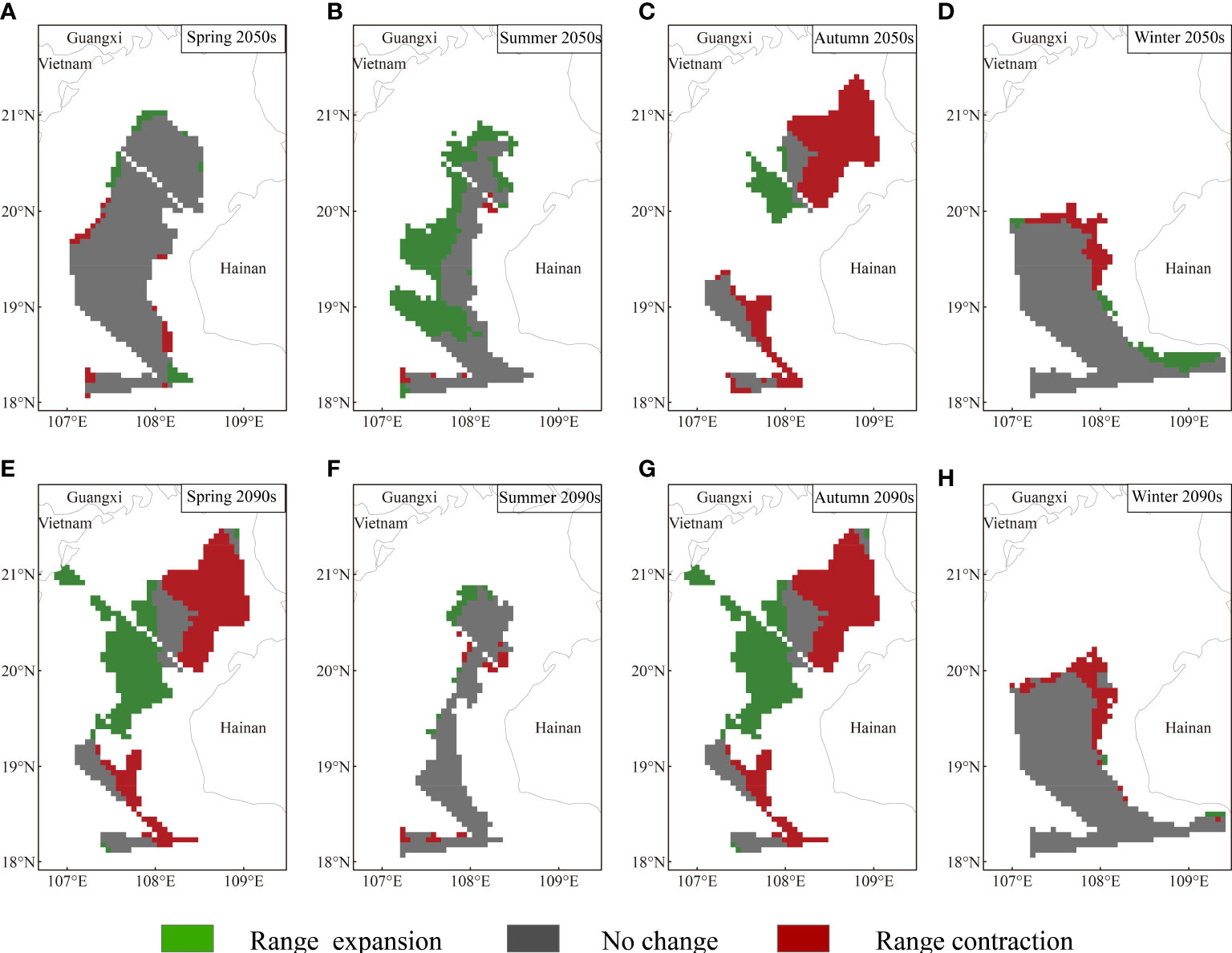
Figure 6 Projected range expansion or contraction for Trichiurus japonicus in the Beibu Gulf under RCP370. Area of geographic distribution changes among the species distribution models based on binary predictions for current and future (i.e., in the 2050s and 2090s) climate conditions: (A) spring 2050s; (B) summer 2050s; (C) autumn 2050s; (D) winter 2050s; (E) spring 2090s; (F) summer 2090s; (G) autumn 2090s; (H) winter 2090s.
Under RCP585, the trend in suitable habitat for T. japonicus varied among seasons. The area of suitable habitat shows an increasing trend but is smaller than the area of decrease in winter in the 2050s (Figure 7); the area of suitable habitat shows the opposite trend in other seasons, although with less variation in spring. The areas of suitable habitat near Vietnam increase substantially in summer, with the model-predicted area of suitable habitat expansion at 8,900.18 km2. Trends in potentially suitable habitat in autumn and winter were similar under the three scenarios. Compared with that in the 2050s, the suitability of the future potential suitable habitat of T. japonicus in the 2090s decreases in autumn and winter, but increases in spring and summer. Also in the 2090s, the range-expansion area increases and the range-contraction area decreases in summer and both areas increase in spring, but the area of range expansion is larger.
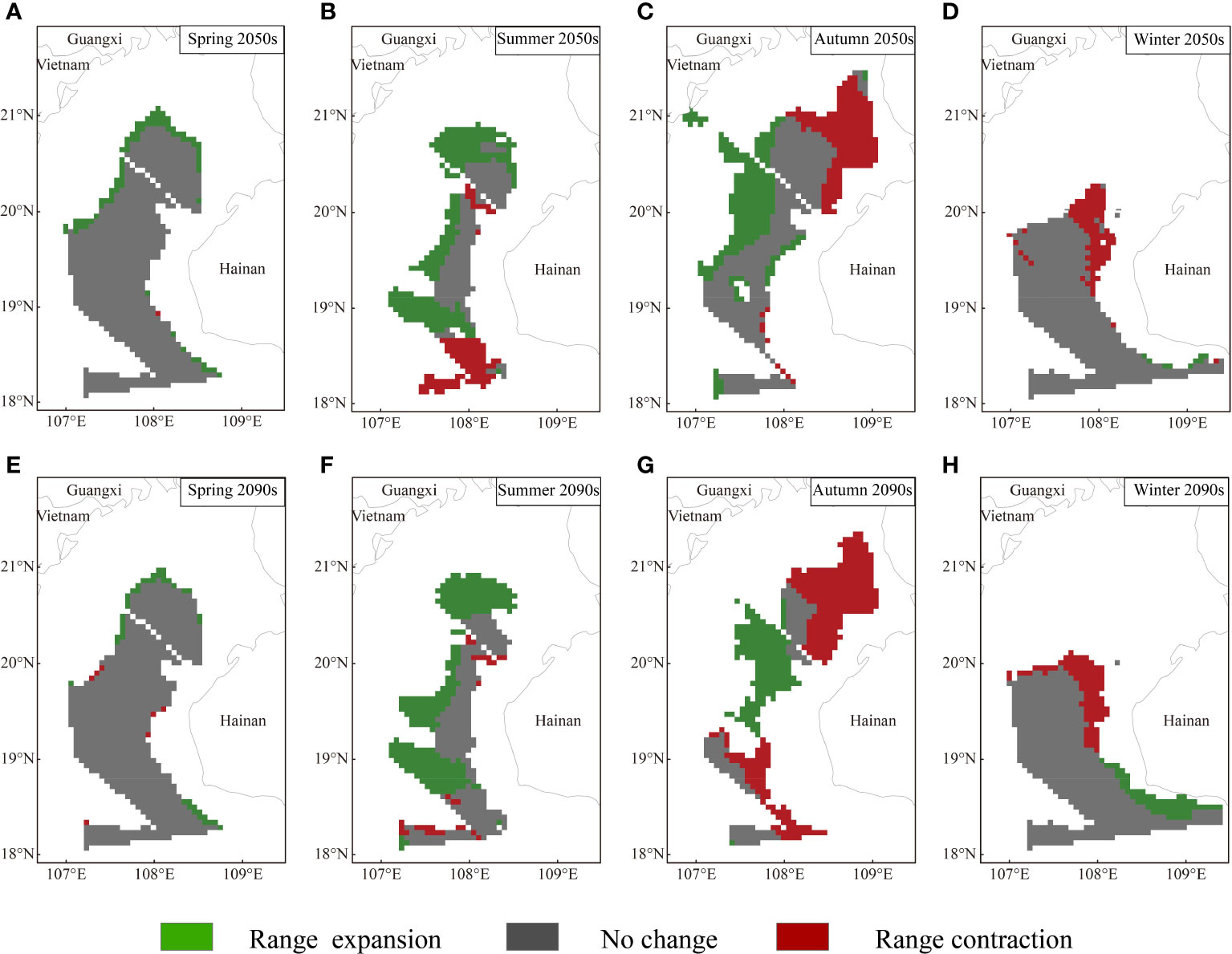
Figure 7 Projected range expansion or contraction for Trichiurus japonicus in the Beibu Gulf under RCP585. Area of geographic distribution changes among binary species distribution models based on binary predictions for current and future (i.e., in the 2050s and 2090s) climate conditions: (A) spring 2050s; (B) summer 2050s; (C) autumn 2050s; (D) winter 2050s; (E) spring 2090s; (F) summer 2090s; (G) autumn 2090s; (H) winter 2090s.
3.5 Core distributional shifts
The core distribution of T. japonicus in the Beibu Gulf showed pronounced seasonal variation, and the wintering population shifts toward the northeast (Figure 8). Under three climate-change scenarios, the core displacement was generated more toward central parts of the Beibu Gulf in spring, summer and autumn, but toward southern parts in winter (Figure 9D). The species’ core distribution in spring in both the 2050s and 2090s under the three climate-change scenarios, except for RCP126–2050 spring, shifts northeastward (Figure 9A). In spring in the 2050s under RCP126, the centroids of the areas shifted as far as 11.060 km away from the current distribution core, as the distribution cores in summer, autumn, and winter moved toward the northwest, north, and south, respectively. By contrast, we examined the centroid distances in different seasons, and found the migration distances of the centroids were shortest in summer and longest in summer (Figure 9B). The maximum migration distance of the centroids of areas was 40.152 km, under RCP126_2050 autumn (Figure 9C).
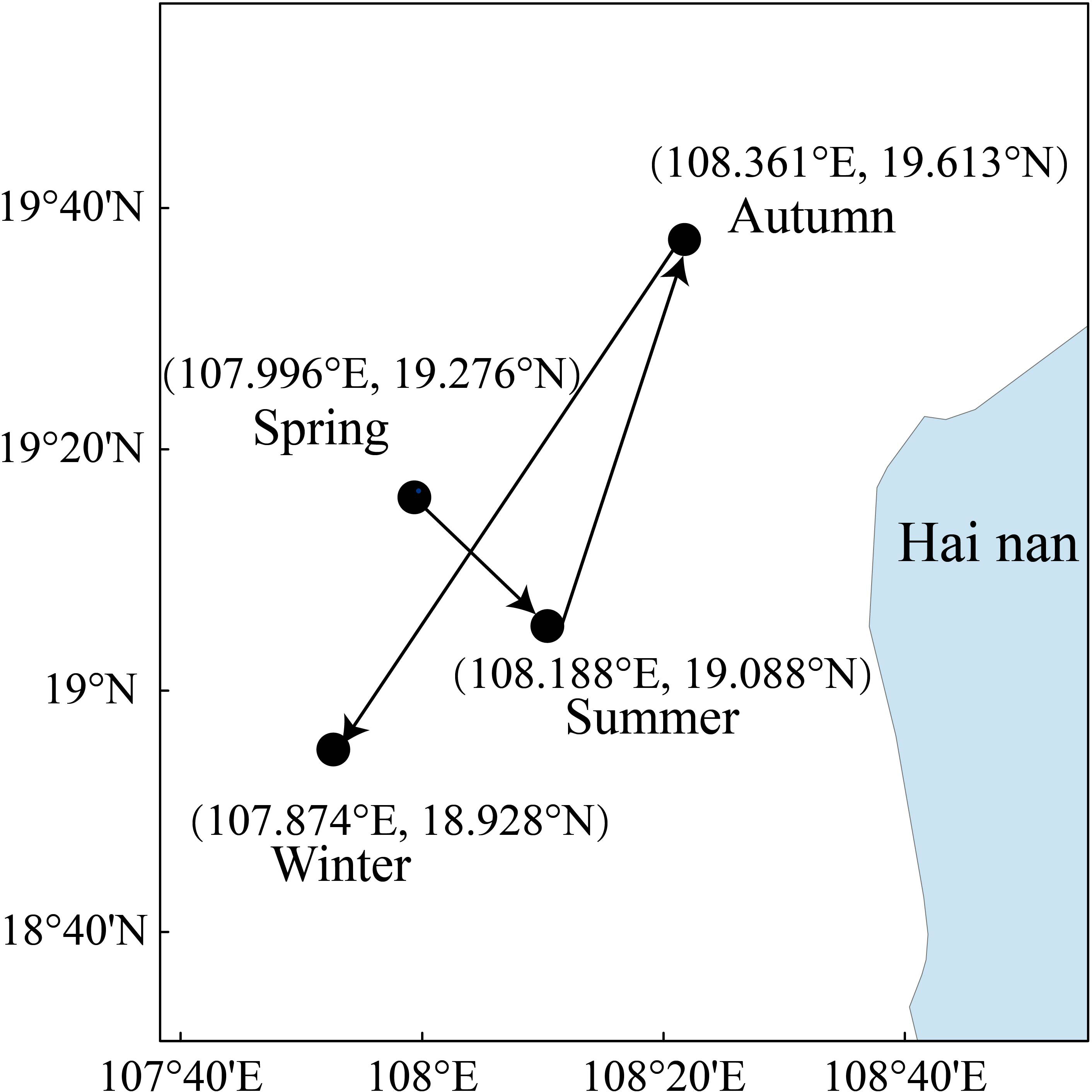
Figure 8 The current core seasonal distributions of Trichiurus japonicus in the Beibu Gulf, South China Sea.
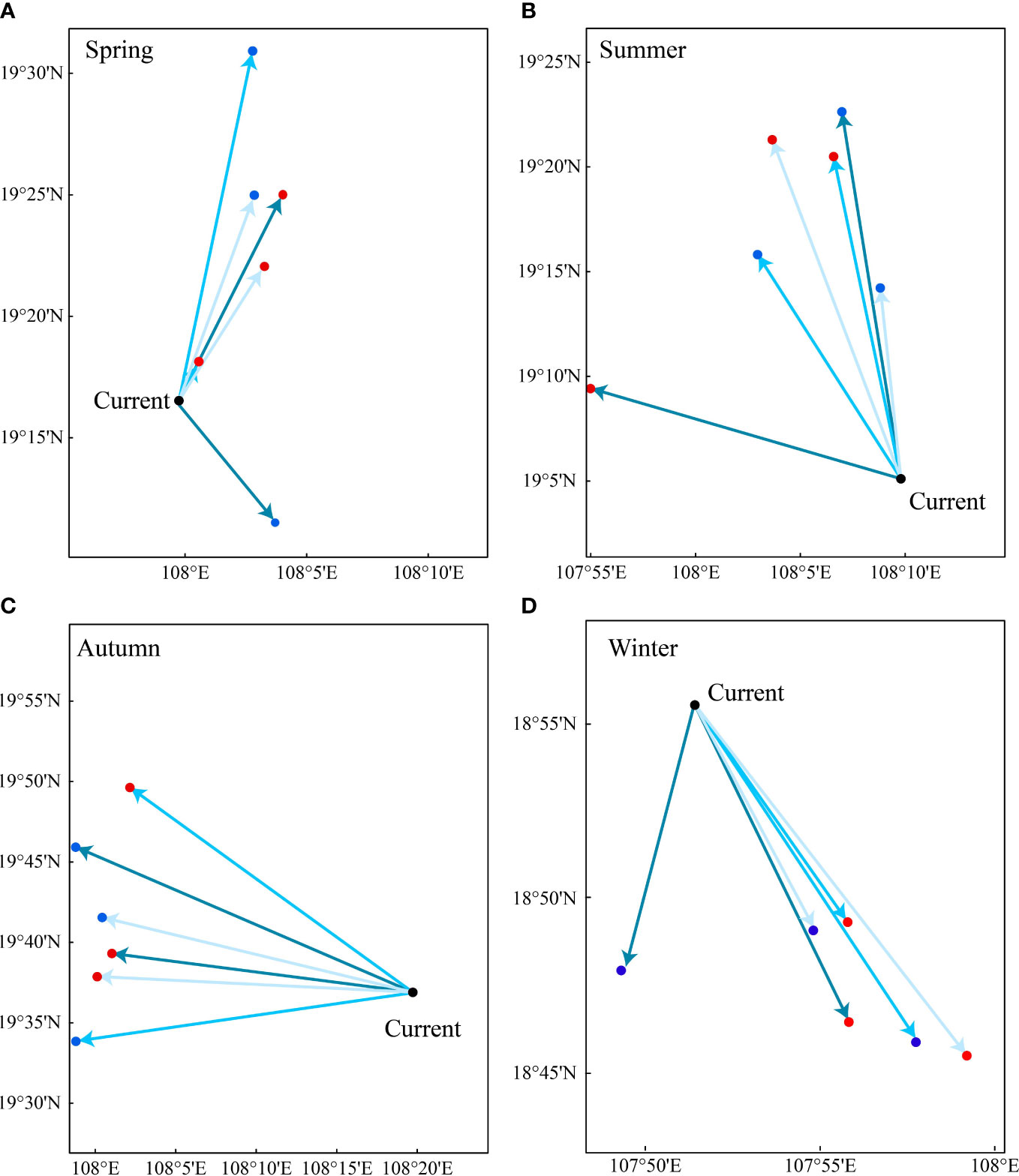
Figure 9 Current potential distribution pattern of Trichiurus japonicus in the Beibu Gulf, by season, and core distributional shifts of suitable habitat under future climate scenarios in the 2050s and the 2090s. Different colors represent different climate-change models: dark blue denotes SSP126, mid-blue SSP370, and light blue SSP585. The black dot represents the suitable habitat distribution center under the current climate scenario, blue dots represent that in the 2050s, and red dots that in the 2090s; red arrows indicate shifting routes of the suitable habitat distribution core. (A) spring; (B) summer; (C) autumn; (D) winter.
4 Discussion
4.1 Seasonal variation in the current potential distribution of T. japonicus and associated environmental drivers
T. japonicus was distributed chiefly in central and southern parts of the Beibu Gulf, which agrees with the findings of Hou et al. (2021). The Beibu Gulf is characterized by complex oceanographic processes, including seasonal mixing, upwelling, and circulation. We analyzed the environmental variables in the gulf to forecast the habitat characteristics available to T. japonicus. In spring, the variation between SST and SBT was significant, which reflects the phenomenon of water-column stratification, such that SST or SBT is the most-important environmental factor (Sun et al., 2020). The most-significant contributor was bottom water temperature, which was the top predictor explaining T. japonicus distribution in summer and autumn. The southwest monsoon prevails in the gulf in summer, when high temperatures induced by solar heat flux and this monsoon limit the spawning activity of T. japonicus (Hou et al., 2021). Thus, we propose that the significant reduction in the suitable habitat area of the species is associated with the main season that T. japonicus feeds in the gulf (summer). The highest contribution to the model in summer was bottom water temperature, which in the Beibu Gulf is primarily associated with a middle-to-bottom cold water mass. This cold water mass reaches maximum intensity in summer, when the region surrounding it becomes rich in nutrients, and the bottom temperature significantly decreases (Gao et al., 2014). Decapterus maruadsi is the predominant bait of Trichiurus spp. And in autumn, the spatial distribution of suitable habitat largely matched the areas with the distribution of D. maruadsi (Chen et al., 2019). Maximum mixed layer depths occur in winter, which strengthens vertical convection mixing and provides nutrients needed to sustain plankton growth. This is beneficial for the feeding and overwintering of T. japonicus. Water temperature also had a high contribution to the models of T. japonicus geographic distribution in winter. The higher the temperature, the higher the probability of its distribution. In winter, the gulf temperatures are lower in the north and higher in the south, with a tongue-shape mass of warm water entering the gulf from the southwest; therefore, the temperature distribution in the southwestern areas is better suited as T. japonicus habitat (Huang et al., 2008; Ya et al., 2015).
4.2 Potential distributional shifts under future climate change in relation to environmental drivers
This study used T. japonicus as a case species and roughly estimated its distribution trends under possible pathways concerning future conditions of the mixed layer depth and salinity, based on MaxEnt niche modelling. Regarding climate-induced changes in the distribution of marine species, the FAO (2012) states that tropical zones are vulnerable to reduction in suitable habitat areas. Compared with that in the 2050s, under all three SSP scenarios, there is a higher risk of suitable habitats for T. japonicus in the Beibu Gulf in the 2090s. Under the three climate-change scenarios, the disadvantage to the distribution of T. japonicus is greatest in winter, both in the short-term and long-term. Potential suitable distributions for this species show a minor range extension in RCP 370_2050 winter, but in the rest of the scenes and years its distribution contracts toward the southern part of the gulf.
Mixed layer depth and salinity were shown to be major environmental stress factors affecting the distribution of T. japonicus. Thus, with changes in only these two factors, the maximum impact of suitable habitat distribution was observed during winter when compared with the findings for the other three seasons. Under RCP370_2090, areas having a suitable mixed layer depth range for T. japonicus showed no obvious change, while areas with a suitable salinity range showed contraction in winter in the northern part of the Beibu Gulf. Under the RC370 climate-change scenario, the projected salinity range in the study area was 31.437-33.721‰ in the 2050s and 31.100-33.518‰ in the 2090s. Hence, the salinity threshold may remain relatively constant in the Beibu Gulf but with a gradual decline in the optimum salinity range for T. japonicus over time. Cheng et al. (2020) studied the average salinity at a series of Argo floats in four ocean regions during 1960-2017; the results revealed a long-term trend of decreasing absolute salinity in the Pacific Ocean. The physiological responses of fish to environmental factors will prompt their movement to more suitable areas (Rodriguez-Burgos et al., 2022). Thus, we advise that salinity is the major environmental factor that will affect contraction of suitable habitat for T. japonicus in the long term.
In our results, the mixed layer depth is deeper in the 2050s and 2090s than in the current scenario. Under the same change trend, there is a higher risk to suitable habitat for T. japonicus in the 2090s. One explanation for this phenomenon could is that the contribution rates of the different environmental factors to the model will be affected by time. The present results align with previous studies (Mamun et al., 2018). For example, under RCP370, the gap between the relative contribution rate of the mixed layer depth (21.8%) and the salinity (17.4%) in the 2090s is smaller than the relative contribution rate of the mixed layer depth (31.7%) and the salinity (23.0%) in the 2050s (Figure 10).
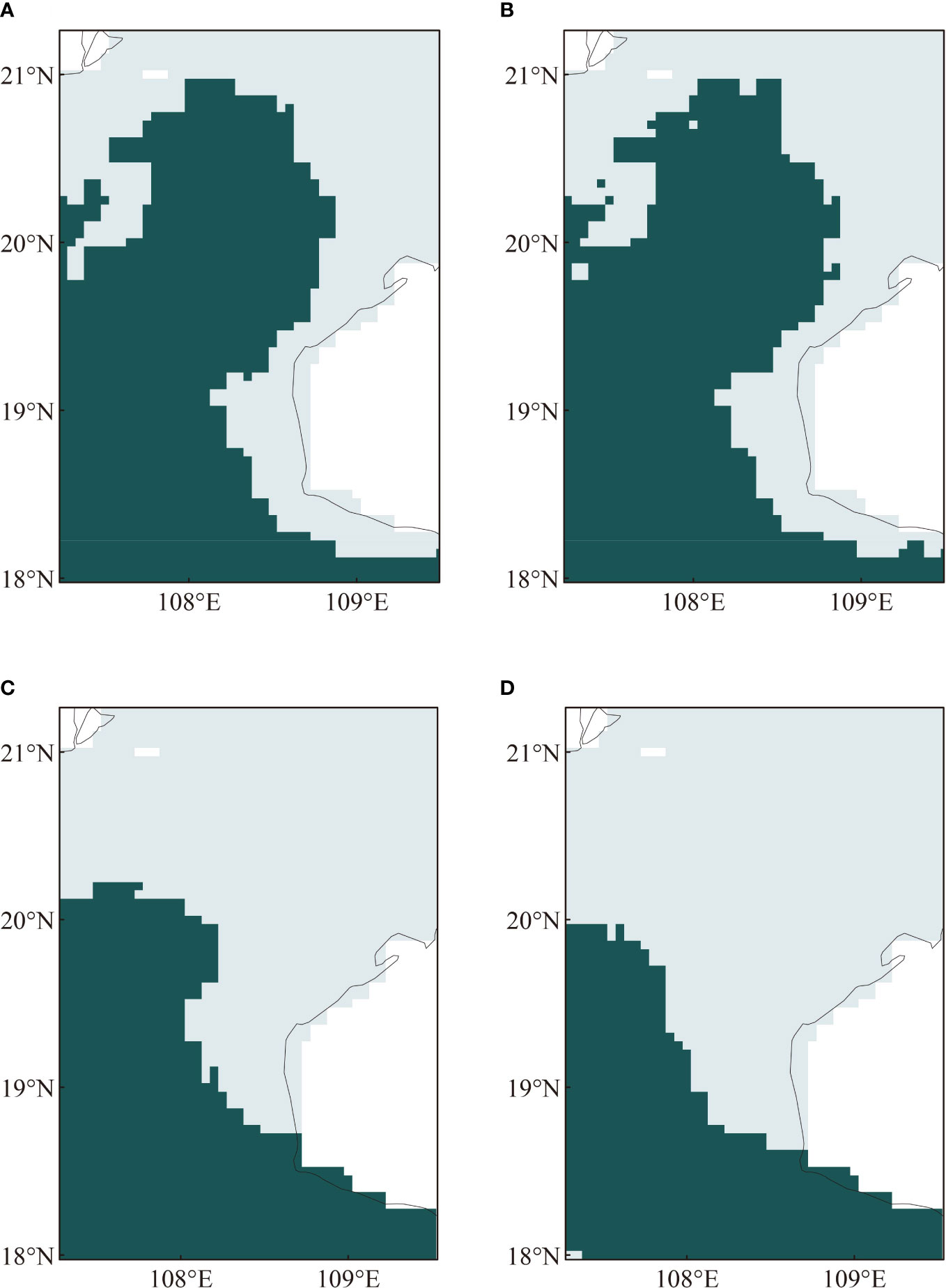
Figure 10 Distribution of the most suitable salinity and mixed layer depth combination (darker color) for Trichiurus japonicus in the Beibu Gulf, as projected for the future (in the 2050s and 2090s), under SSP370: (A) mixed layer depth, 2050s; (B) mixed layer depth, 2090s; (C) salinity, 2050s; (D) salinity, 2090s.
Different seasons of T. japonicus change in the status of winners and losers in the face of future climate change. Such seasonal changes may be particularly relevant for understanding how future climate will impact marine life as many key biological processes. According to the results of this study, it might result in reducing population repopulation efforts for the following spring if the species in winter encounters conditions predicted in RCP126 or RCP585 for 2050s and especially 2100s. It will also have negative effects on the functioning of such tropical ecosystems and the livelihoods of local fisherfolk. So we suggest that protection measures should be adopted as early as possible for the conservation of the fishing grounds of T. japonicus in the Beibu gulf in order to mitigate the effects of climate change on the fisheries and the ecosystem.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by the South China Sea Fisheries Research Institute, Chinese Academy of Fishery Science.
Author contributions
PX: conceptualization, Methodology, Formal analysis, Visualization, Writing - original draft. YX and MS: Investigation, Writing-review and editing. XZ and PJ: Formal analysis, Visualization. ZC: Methodology, Funding acquisition, Supervision, Validation, Writing-review and editing. JF: Conceptualization, Methodology, Supervision, Writing-review and editing. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by Key research and development project of Guangdong Province(2020B1111030001), Major Projects of Basic and Applied Basic Research Programs in Guangdong Province(2019B030302004), Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2020TD05). Central Public-Interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS (2021SD01), Financial Fund of the Ministry of Agricultureand Rural Affairs, P. R. of China (NFZX2021), National Key Research and Development Program of China (2018YFD0900906).
Acknowledgments
We thank Cynthia Kulongowski with Liwen Bianji (Edanz) (www.liwenbianji.cn) edited the language of a draft of this manuscript. We are grateful to the crew members of Zhongyuke 301 and Beiyu 60011 for their help with the data collection.In addition, we sincerely thank the reviewers for their valuable comments on an early draft of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1079590/full#supplementary-material
References
Angeles-Gonzalez L. E., Martinez-Meyer E., Yanez-Arenas C., Velazquez-Abunader I., Lopez-Rocha J. A., Torrejon-Magallanes J., et al. (2021). Climate change effect on octopus maya (Voss and solís-ramírez 1966) suitability and distribution in the yucatan peninsula, gulf of mexico: A correlative and mechanistic approach. Estuar. Coast. Shelf. S. 260, 107502. doi: 10.1016/j.ecss.2021.107502
Brown J. L. (2014). Sdm toolbox: A python-based gis toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 5, 694–700. doi: 10.1111/2041-210X.12200
Cai Y. C., Huang Z. R., Li J. J., Xu Y. W. (2020). Stock distribution of a new record species nemipterus mesoprion in offshore northern south China sea. South Chin. Fish. Sci. 16, 1–11. doi: 10.12131/20200064
Cheng L. J., Gruber T. N., Abraham J. P., Fasullo J. T., Li G. C., Mann M. E., et al. (2020). Improved estimates of changes in upper ocean salinity and the hydrological cycle. J. Clim. 33, 10357–10381. doi: 10.1175/JCLI-D-20-0366.1
Chen Z. Z., Zhang K., Cai Y. C., Sun M. S., Xu Y. W., Zhang J., et al. (2019). Fishery resources and environments in the northern south China sea (Beijing: China ocean press).
Cobos M. E., Peterson A. T., Barve N., Osorio-Olvera L. (2019). Kuenm: An r package for detailed development of ecological niche models using maxent. Peer J. 7, e6281. doi: 10.7717/peerj.6281
Cong M., Xu Y. Y., Tang L., Yang W. J., Jian M. F. (2020). Predicting the dynamic distribution of sphagnum bogs in China under climate change since the last interglacial period. PloS One 15, e0230969. doi: 10.1371/journal.pone.0230969
Davis T. R., Cadiou G., Champion C., Coleman M. A. (2020). Environmental drivers and indicators of change in habitat and fish assemblages within a climate change hotspot. Reg. Stud. Mar. Sci. 36, 101295. doi: 10.1016/j.rsma.2020.101295
Fishbase (2022) Trichiurus lepturus. Available at: https://www.fishbase.de/FieldGuide/FieldGuideSummary.php?genusname=Trichiurus&speciesname=lepturus&c_code=608 (Accessed August 16, 2022).
Food and Agriculture Organization (FAO (2012). State of the world’s fisheries and aquaculture 2012 (Rome: FAO).
Gao J. S., Shi M. C., Chen B., Zhao D. (2014). Responses of the circulation and water mass in the beibu gulf to the seasonal forcing regimes. Haiyang Xuebao 33, 1–11. doi: 10.1007/s13131-014-0506-6
Harte J., Newman E. A. (2014). Maximum information entropy: a foundation for ecological theory. Trends Ecol. Evol. 29, 384–389. doi: 10.1016/j.tree.2014.04.009
Hou G., Xu Y. W., Chen Z. Z., Zhang K., Huang W. S., Wang J. R., et al. (2021). Identification of eggs and spawning zones of hairtail fishes trichiurus (pisces: Trichiuridae) in northern south China Sea, using DNA barcoding. Front. Environ. Sci. Eng. 9. doi: 10.3389/fenvs.2021.703029
Huang Y. C., Li Y., Shao H., Li Y. H. (2008). Seasonal variations of sea surface temperature chlorophyll a and turbidity in beibu gulf, modis imagery study. J. Xiamen Univ. (Nat. Sci.), 48, 856–863.
Hu W., Du J., Su S., Tan H., Yang W., Ding L., et al. (2022). Effects of climate change in the seas of China: Predicted changes in the distribution of fish species and diversity. Ecol. Indic. 134, 108489.
Intergovernmental Panel on Climate Change (IPCC (2007). “Contribution of working group II to the fourth assessment report of the intergovernmental panel on climate change,” (Cambridge: Cambridge University Press).
Jiang Y. E., Sun D. R., Lin Z. J., Fang Z. Q. (2015). A new record of bothid fish, tosarhombus octoculatus,in the south China Sea. Yingyong Haiyang Xuebao 34, 397–401.
Lin S. Y., Zhao L., Feng J. (2022). Predicted changes in the distribution of antarctic krill in the cosmonaut sea under future climate change scenarios. Ecol. Indic. 142, 109234. doi: 10.1016/j.ecolind.2022.109234
Liu S. H., Liu Y., Alabia I. D., Tian Y. J., Ye Z. J., Yu H. Q., et al. (2020). Impact of climate change on wintering ground of japanese anchovy (engraulis japonicus) using marine geospatial statistics. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00604
Ma S. Y., Cheng J. H., Li J. C., Liu Y., Wan R., Tian Y. J. (2019). Interannual to decadal variability in the catches of small pelagic fishes from china seas and its responses to climatic regime shifts. Deep Sea Res. Part II Top. Stud. Oceanogr. 159, 112–129. doi: 10.1016/j.dsr2.2018.10.005
Mamun M., Kim S., An K. G. (2018). Distribution pattern prediction of an invasive alien species largemouth bass using a maximum entropy model (maxent) in the korean peninsula. J. Asia-Pacific Biodiversity J. Asia -Pac Biodivers. 11, 516–524. doi: 10.1016/j.japb.2018.09.007
Ministry of Agricultrue and Rural Affairs of the People’s Republic of China (MARA (2021). China Fishery statistical yearbook 2020 (Beijing: China Agriculture Press).
Ministry of Natural Resources of the Peoples’s Republic of China (MNR (2021) 2020 Sea level bulletin of China. Available at: http://gi.mnr.gov.cn/202104/P020210426570276410847.pdf (Accessed August 13, 2022).
Montiel S. A. H., Coronado-Franco K. V., Selvaraj J. J. (2019). Predicted changes in the potential distribution of seerfish (scomberomorus sierra) under multiple climate change scenarios in the colombian pacific ocean. Ecol. Inform. 53, 100985. doi: 10.1016/j.ecoinf.2019.100985
Muscarella R., Galante P. J., Soley-Guardia M., Boria R. A., Kass J. M., Uriarte M., et al. (2014). ENMeval: An r package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods Ecol. Evol. 5, 1198–1205. doi: 10.1111/2041-210X.12261
Phillips S. J., Anderson R. P., Dudík M., Schapire R. E., Blair M. E. (2017). Opening the black box: An open-source release of maxent. Ecography 40, 887–893. doi: 10.1111/ecog.03049
Phillips S. J., Anderson R. P., Schapire R. E. (2006). Maximum entropymodeling of species geographic distributions. Ecol. Modell. 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Phillips S. J., Dudík M. (2008). Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 31, 161–175. doi: 10.1111/j.0906-7590.2008.5203.x
Rodriguez-Burgos A. M., Briceño-Zuluaga F. J., Ávila Jiménez J. L., Hearn A., Peñaherrera-Palma C., Espinoza E., et al. (2022). The impact of climate change on the distribution of sphyrna lewini in the tropical eastern pacific. Mar. Environ. Res. 180, 105696–105696. doi: 10.1016/j.marenvres.2022.105696
Shi D. F., Zhang K., Cai Y. C., Geng P., Xu Y. W., Sun M. S., et al. (2020). Population structure of trichiurus japonicus in northern south China Sea and parameters of its growth. South Chin. Fish. Sci. 16, 51–59. doi: 10.12131/20200055
Sill S. R., Dawson T. P. (2021). Climate change impacts on the ecological dynamics of two coral reef species, the humphead wrasse (cheilinus undulatus) and crown-of-thorns starfish (ancanthaster planci). Ecol. Inform. 65, 101399. doi: 10.1016/j.ecoinf.2021.101399
Sun P., Chen Q., Fu C., Zhu W., Li J., Zhang C., et al. (2020). Daily growth of young-of-the-year largehead hairtail (Trichiurus japonicus) in relation to environmental variables in the east China sea. J. Mar. Syst. 201, 103243. doi: 10.1016/j.jmarsys.2019.103243
Tian Y. J. (2020). Climate-induced nonlinearity in pelagic communities and non-stationary relationships with physical drivers in the kuroshio ecosystem. Fish. Fish. 22, 1–17. doi: 10.1111/faf.12502
Xiao F., Daniel S. P., Ye L., Pandey R., Papeş M. (2019). Collinearity in ecological niche modeling: Confusions and challenges. Ecol. Evol. 9, 10365–10376. doi: 10.1002/ece3.555
Ya H. Z., Gao J. S., Dong D. X. (2015). Analysis of variation characteristic and driving factor of sea surface temperature in beibu gulf. Guangxi Sci. 22, 260–265. doi: 10.13656/j.cnki.gxkx.20150708.004
Yan Y. R., Zhang W. K., Lu H. S., Wang X. F. (2012). Using stable isotopess to analyze feeding haibits and trophic position of hairtail from the beibu gulf, south China Sea. Oceanol. Limnol. Sin. 43, 192–200.
Zhang K., Geng P., Li J. J., Xu Y. W., Kalhoro M. A., Sun M. S., et al. (2022a). Influences of fisheries management measures on biological characteristics of threadfin bream (Nemipterus virgatus) in the beibu gulf, south China Sea. Haiyang Xuebao 41, 24–33.
Zhang Z. X., Mammola S., Xian W. W., Zhang H. (2020b). Modelling the potential impacts of climate change on the distribution of ichthyoplankton in the yangtze estuary, China. Divers. Distrib. 26, 126–137. doi: 10.1111/ddi.1300
Zhang Z. X., Mammola S., Zhang H. (2020a). Does weighting presence records improve the performance of species distribution models? a test using fish larval stages in the yangtze estuary. Sci. Total Environ. 741, 140393. doi: 10.1016/j.scitotenv.2020.140393
Zhang M., Wang X. H., Du F. Y., Sun D. R. (2022b). Assessment of trichiurus haumela stocks in the beibu gulf based on length-based bayesian biomass estimation method. Haiyang Xuebao 44, 11–21. doi: 10.12284/hyxb2022002
Zhao Y., Deng X. W., Xiang W., Chen L., Shuai O. Y. (2021). Predicting potential suitable habitats of chinese fir under current and future climatic scenarios based on maxent model. Ecol. Inform. 64, 101393. doi: 10.1016/j.ecoinf.2021.101393
Zhao Z. Y., Xiao N., Shen M., Li J. S. (2022). Comparison between optimized maxent and random forest modeling in predicting potential distribution: A case study with quasipaa boulengeri in China. Sci. Total Environ. 842, 156867. doi: 10.1016/j.scitotenv.2022.156867
Keywords: habitat suitability, maximum entropy model, potential distribution, seasonal change, climate-change biology
Citation: Xiong P, Xu Y, Sun M, Zhou X, Jiang P, Chen Z and Fan J (2023) The current and future seasonal geographic distribution of largehead hairtail Trichiurus japonicus in the Beibu Gulf, South China Sea. Front. Mar. Sci. 9:1079590. doi: 10.3389/fmars.2022.1079590
Received: 25 October 2022; Accepted: 29 November 2022;
Published: 12 January 2023.
Edited by:
Fan Zhang, Shanghai Ocean University, ChinaCopyright © 2023 Xiong, Xu, Sun, Zhou, Jiang, Chen and Fan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zuozhi Chen, chenzuozhi@scsfri.ac.cn; Jiangtao Fan, fjt@scsfri.ac.cn