Standardized protocol for reproductive phenology monitoring of fucalean algae of the genus Cystoseira s.l. with potential for restoration
 Fabio Rindi1,2
Fabio Rindi1,2  Alba Vergés3
Alba Vergés3  Irene Zuchegna1 Silvia Bianchelli1,2
Irene Zuchegna1 Silvia Bianchelli1,2  Sònia de Caralt3
Sònia de Caralt3  Cristina Galobart4 Jorge Santamaría4 Francesco Martini1,4
Cristina Galobart4 Jorge Santamaría4 Francesco Martini1,4  Margalida Monserrat5,6
Margalida Monserrat5,6  Sotiris Orfanidis7 Cèlia Sitjà4
Sotiris Orfanidis7 Cèlia Sitjà4  Soultana Tsioli7
Soultana Tsioli7  Jana Verdura5
Jana Verdura5  Luisa Mangialajo5
Luisa Mangialajo5  Simonetta Fraschetti2,8
Simonetta Fraschetti2,8  Roberto Danovaro1,2
Roberto Danovaro1,2  Emma Cebrian4*
Emma Cebrian4*- 1Dipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, Ancona, Italy
- 2National Biodiversity Future Center (NBFC), Palermo, Italy
- 3Institut Ecologia Aquàtica, Universitat de Girona, Girona, Spain
- 4Department of Marine Ecology, Centre d’Estudis Avançats de Blanes (CEAB-CSIC), Blanes, Spain
- 5Université Côte d’Azur, CNRS, ECOSEAS, Nice, France
- 6Consorzio Interuniversitario per le Scienze del Mare, CoNISMa, Roma, Italy
- 7Fisheries Research Institute (ELGO-DIMITRA), Kavala, Greece
- 8Department of Biology, University of Naples Federico II, Naples, Italy
Active marine restoration is strongly encouraged to prevent the loss of the valuable habitats formed by Cystoseira sensu lato species, since they enhance biodiversity and preserve ecosystem functions and services. Current restoration interventions are mainly based on recruitment enhancement methods by deploying bags with fertile receptacles in situ or by outplanting juveniles grown ex situ under laboratory conditions. These methods allow the recovery of endangered species avoiding the depletion of the donor populations. In all cases, a priori knowledge of the reproductive phenology and recruitment periods of the species to be restored is essential, since the success of restoration techniques relies on collecting fertile branches and the obtention and survival of recruits. For their collection, identified donor populations characterized by dense Cystoseira s.l. cover should be studied. Specifically, monitoring the reproductive phenology of populations is crucial to detect the period of the year in which they develop mature reproductive structures and to understand how it might be linked to environmental conditions. Then, these general patterns on the reproductive phenology of Cystoseira s.l. species are essential to determine the most suitable time and conditions to plan for the most effective restoration action. Here, we provide a cost-effective and friendly protocol that can be easily and widely implemented for all Cystoseira s.l. species. We pose that this protocol provides a standardized and useful methodology to understand the environmental factors driving the optimal periods for sampling fertile branches across the Mediterranean Sea, and thus it can be an essential tool to plan future restoration actions.
Introduction
In the Mediterranean Sea, macroalgal forests formed by populations of Cystoseira sensu lato (genera Cystoseira C. Agardh, Ericaria Stackhouse and Gongolaria Boehmer) dominate shallow and mesophotic areas of rocky reefs. Unfortunately, declines and local extinctions of these fucalean populations have been reported throughout the Mediterranean (Bianchi et al., 2014; Thibaut et al., 2015; Mariani et al., 2019), mainly caused by deterioration of water quality, habitat destruction and overgrazing by herbivores (Thibaut et al., 2005; Mangialajo et al., 2008; Sala et al., 2012; Blanfuné et al., 2016; Orfanidis et al., 2021). The loss of Cystoseira s.l. forests is a concern, since they are very productive habitats and are home to a large number of associated species (Ballesteros, 1988). Moreover, these communities offer important ecosystem services, such as being important nursery habitats for fisheries species (Cheminée et al., 2013). Since Mediterranean coastal areas are among the most altered and degraded systems in the world (Lotze et al., 2011), the pressures that Cystoseira s.l. habitats are now facing are likely to increase, probably leading to more frequent population extinctions. Once the populations become extinct, their natural recovery is almost impossible, even after mitigation of the stressors responsible for their disappearance (Pinedo et al., 2013; Tamburello et al., 2022). In these circumstances, active restoration emerges as the only suitable tool to recover these ecosystems (Verdura et al., 2018; Orlando-Bonaca et al., 2022; Tamburello et al., 2022).
Early trials of restoration of Cystoseira s.l. in the Mediterranean were attempted in the early 2000s and consisted of transplants of adult thalli (Falace et al., 2006; Susini et al., 2007; Robvieux, 2013) or transfers of cobbles with settled juvenile specimens (Perkol-Finkel and Airoldi, 2010; Perkol-Finkel et al., 2012). Since most Cystoseira s.l. species are considered threatened or endangered by the Barcelona Convention (Appendix II; UNEP/MAP, 2013), any technique that requires harvesting whole individuals is considered undesirable, as it will cause a depletion of the donor populations and consequently is a potential threat (Cebrian et al., 2021). This has led to the exploration of less invasive restoration actions (see Gianni et al., 2013 and Cebrian et al., 2021 for reviews).
The high potential of these seaweeds to generate gametes and zygotes under optimal conditions makes the use of germlings obtained from fertile receptacles (usually present at the apices of the branches) a sustainable option for restoring Cystoseira s.l. forests without depleting natural populations (Falace et al., 2018; Verdura et al., 2018). Current restoration approaches are based on enhancement of recruitment obtained by stimulating sexual reproduction and consequent production of numerous germlings (which are then expected to grow into adult specimens and gradually form self-maintaining populations). The germlings can be produced in controlled conditions in the laboratory (ex situ methods; Falace et al., 2018; Verdura et al., 2018; De La Fuente et al., 2019; Clausing et al., 2022; Lardi et al., 2022) or directly in the field (in situ methods; Verdura et al., 2018; Medrano et al., 2020a). In both cases, the availability of fertile branches containing large numbers of mature gametes is a key requirement. Knowledge of the reproductive phenology of donor populations is crucial to detect the time of the year in which mature reproductive structures develop, understand how reproduction is influenced by environmental conditions, and finally determine the most suitable period and conditions to carry out effective restoration action (Cebrian et al., 2021; Smith et al., 2023). However, this knowledge is only partially available for some species (Sauvageau, 1912; Ercegović, 1952; Gómez-Garreta et al., 1982; Hoffmann et al., 1992; Pizzuto et al., 1995; Serio, 1995a; Serio, 1995b; Alongi et al., 1999; Pardi et al., 2000; Gómez Garreta et al., 2001; Marzocchi et al., 2003; Falace et al., 2005; Capdevila et al., 2015; Medrano et al., 2020b; Battelli and Catra, 2023; for a summary of the information available see Cebrian et al., 2021). Furthermore, phenological data published in individual studies typically have been obtained from a single site or geographical area; to date, there is not a single Cystoseira s.l. species for which phenological data are available at a pan-Mediterranean scale.
The great intraspecific year-to-year variability in the reproductive potential documented for some Cystoseira s.l. populations (Bevilacqua et al., 2019) leads to a high unpredictability of available mature gametes and highlights the need for standardized phenological protocols for different populations and species across the Mediterranean Sea.
In order to ensure comparability of future phenological studies, here we propose a cost-effective and user-friendly protocol for monitoring the reproductive phenology applicable to all Cystoseira s.l. species. The aim of the proposed protocol is to guide restoration actions by detecting the optimal periods for sampling of fertile branches across the Mediterranean Sea.
Materials and equipment
Material: reproductive structures in Cystoseira s.l.
Implementation of the protocol requires familiarity with the reproductive structures of Cystoseira s.l. and capacity to recognize their stages of maturation. All members of the order Fucales (to which Cystoseira s.l. belong) have a diplontic monophasic life history, with only one diploid generation and gametic meiosis (Gómez Garreta et al., 2001; Heesch et al., 2021). The reproductive structures are swellings called receptacles, which may be formed either in apical or intercalary portions of the branches, depending on the species (Figure 1). Details of the morphology of receptacles of Mediterranean Cystoseira s.l., including pictures useful to recognize them, are available in Valiante (1883), Gómez Garreta et al. (2001) and Cormaci et al. (2012).

Figure 1 Gongolaria barbata (A) A thallus bearing mature receptacles; (B) detail of mature receptacles.
Cavities known as conceptacles are sunken into the surfaces of receptacles, with only an open pore (ostiole) visible from the receptacle surface (Graham et al., 2016). Each conceptacle contains oogonia (female gametangia), antheridia (male gametangia) and some sterile branched hairs called paraphyses. All species of Cystoseira s.l. are monoecious; the conceptacles are hermaphroditic and contain both oogonia and antheridia (Figure 2A). In some species, however, unisexual thalli are known to occur: male thalli containing only antheridial conceptacles exist in Gongolaria barbata (Stackhouse) Kuntze (Sauvageau, 1912; Martini and Rindi, pers. obs.; Figure 2B), whereas female thalli containing only oogonial conceptacles have been reported for Ericaria amentacea (C. Agardh) Molinari & Guiry, Ericaria mediterranea (Sauvageau) Molinari & Guiry, Ericaria selaginoides (Linnaeus) Molinari & Guiry and Gongolaria montagnei (J. Agardh) Kuntze (Sauvageau, 1912). For some species, different conceptacles within the same receptacle have been documented (e.g., for Cystoseira compressa (Esper) Gerloff & Nizamuddin, Sauvageau (1912) recorded male, female and hermaphroditic conceptacles in a same receptacle).
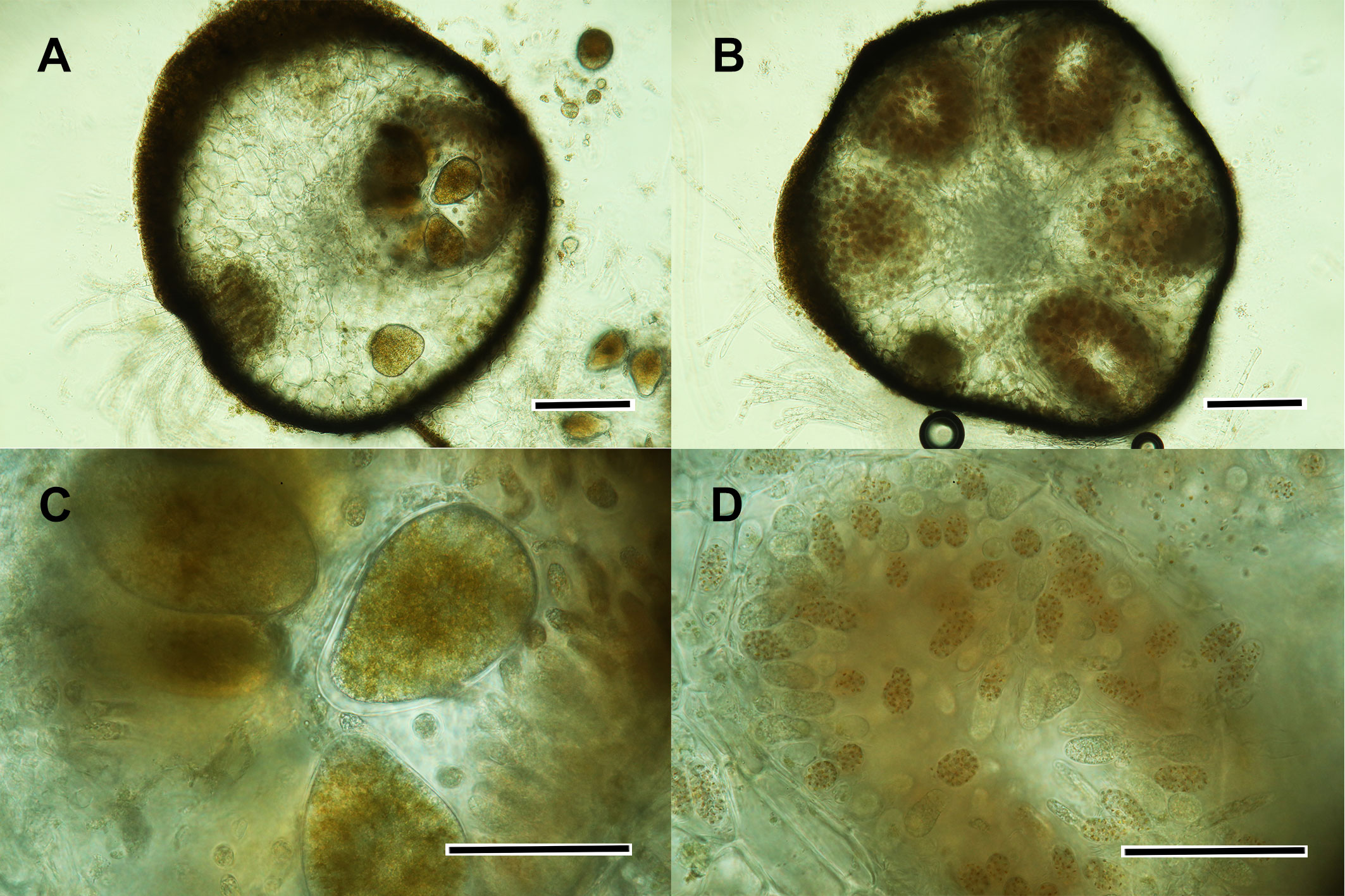
Figure 2 Gongolaria barbata (A) cross section of a hermaphroditic receptacle; (B) cross section of an antheridial conceptacle; (C) detail of some oogonia; (D) detail of an antheridial conceptacle. Scale bars: (A, B) 200 μm; (C, D) 50 μm.
When mature, each conceptacle contains several oogonia (5-6 to about 12, the number varying among species). Typically, the oogonia observed in a conceptacle are in different stages of maturation. The oogonia are produced in the part of the conceptacle opposite to the ostiole, and a ring of antheridial branches occurs in the part near the ostiole (Guern, 1962). The oogonia are easily recognized in cross sections as dark masses with spherical, ovoid or elongated shape (Figure 2C). Each oogonium produces a single egg cell, spherical (90-130 μm in diameter) or ovoid (110-130 x 140-180 μm) in shape. The antheridia are ovoid to club-shaped; they are borne on hair-like antheridial branches and contain 64 antherozoids (male gametes). In some species (e.g., Gongolaria barbata) each antherozoid encloses a stigma formed by carotenoid pigments, in which case the colored dots of the antherozoids give the antheridial layer a distinctive brownish orange color (Figure 2D). In other species (e.g., Cystoseira compressa) the stigma is lacking and the antheridia form a greyish layer, in which case they may be difficult to distinguish from paraphyses (Guern, 1962).
Equipment
The protocol requires both field and laboratory work. For the fieldwork, standard equipment for snorkelling or SCUBA diving is required, depending on the depth at which the population to monitor occurs. For species growing in the infralittoral fringe or in littoral rockpools (e.g., Cystoseira compressa, Ericaria amentacea) wellington boots or waders may be sufficient if monitoring is performed in conditions of low tide. In this case, however, care should be taken not to trample or cause any other damage to Cystoseira s.l. thalli. Writing boards are necessary to annotate the fertility data collected by underwater observation. Sealable ziploc bags or plastic containers are necessary to store the receptacles collected for the maturity assessment until transfer to the laboratory.
Laboratory work requires stereoscopes and light microscopes. Sharp tools such as razor blades or scalpels are necessary to cut thin sections of the receptacles collected. Standard laboratory glassware (slides and coverslips) is required for the microscopic observation of the sections. If possible, it is highly recommended to have a digital camera connected to the light microscope used for the observation. This will allow to obtain high-quality pictures of the sections examined, documenting photographically their level of maturity.
Method
Based on a careful assessment, an appropriate donor population should be selected before to start a restoration action. The donor population should be extensive and dense, ideally with a more or less continuous cover extending for many tens to hundreds of square meters.
The monitoring of the population should be carried out at monthly intervals, at minimum over a whole annual cycle (12 months). However, we recommend that it should be continued as long as funding, workforce available and other logistical considerations allow. Two variables are quantified: fertility and maturity.
Fertility
Fertility is the reproductive variable that defines the amount of receptacles present. It is measured individually for a thallus as percentage of branches bearing receptacles over the total branches of the thallus. The branches to consider for this are the last order branches (Figure 3). Five categories of fertility are defined, based on the following scale:
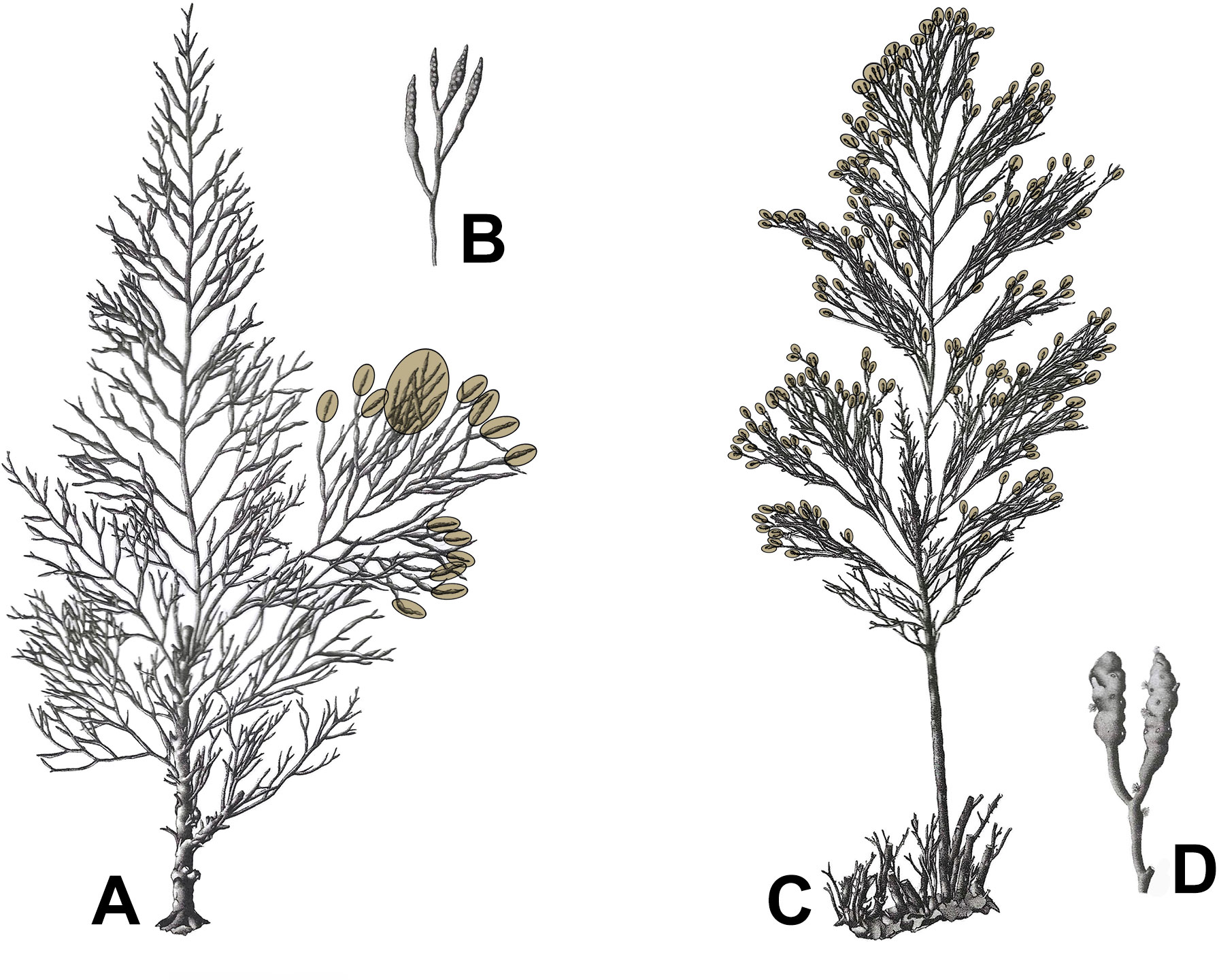
Figure 3 Fertility in Gongolaria barbata (A, B) and Ericaria crinita (C, D). Tips of branches delimited by brown ovals bear receptacles. In Gongolaria barbata fertility level is F1 (< 25% of the branches bearing receptacles); in Ericaria crinita it is F4 (> 75% of the branches bearing receptacles). (A) Thallus of Gongolaria barbata; (B) receptacles of Gongolaria barbata; (C) thallus of Ericaria crinita; (D) receptacles of Ericaria crinita. Adapted from Ercegović (1952).
NF: non fertile, a thallus devoid of receptacles;
F1: < 25% of the branches bears receptacles;
F2: 25 to 50% of the branches bears receptacles;
F3: 50 to 75% of the branches bears receptacles;
F4: > 75% of the branches bears receptacles.
Presence or absence of the receptacles is assessed visually in the field. A number of thalli between 50 and 100 should be checked for fertility, although a lower number can be used in particular cases (e.g., small populations or circumstances that allow to spend only a short time in the field); it should, however, never be lower than 20. The thalli to check must be selected randomly, ensuring that their distribution is spatially well spread within the population. If a Cystoseira s.l. forest extends over a large area, it is possible that different parts of the population may be subjected to somewhat different environmental conditions. In this case, production of receptacles and maturation of gametes might be shifted at different times in different parts of the population.
Maturity
Maturity is the reproductive variable that refers to the developmental stage of the gametangia. It is assessed in the laboratory by microscopic observation.
In the field, receptacles are collected from some of the thalli selected for the fertility check. The number of thalli from which the receptacles are collected will depend on the extent of the population and the total number of thalli present. To avoid a strong impact on the reproductive potential of the donor population, no more than 5% of the total receptacles in the population should be collected. In general, 5 to 10 thalli should be sampled. Adult and fully developed thalli, ideally bearing a large number of receptacles, are to be preferred. In the field they should not be clustered, but widely spread over the whole extent of the population. Two-three receptacles should be collected from each thallus.
Upon collection, the receptacles must be placed in sealable ziploc bags or plastic containers (receptacles from different thalli must be placed in separate ziploc bags, individually numbered) without seawater but keeping them moist. The bags are then transferred to the laboratory, where the receptacles are checked under a stereoscope and sectioned. This should be done within the same day of collection; if not possible, the bags can be stored in a cool and dark place until processing (e.g., in a fridge for a maximum of 24-48 hours). If a fast check is not possible, the material can be placed in a freezer and stored for a longer period. Whenever possible, however, this should be avoided, since defrosting will damage the reproductive structures (especially delicate ones, such as the antheridia), making the assessment more problematic.
In the laboratory, thin transversal sections of the receptacles should be cut using a razor blade or a scalpel. It is recommended to make sections of all two-three receptacles collected from each thallus. The sections should be cut in the median portions of the receptacles (Figure 4A). Since not all conceptacles within a receptacle may be exactly at the same stage of maturity, it is recommended to cut several sections of the same receptacle (at least three-four, Figure 4B).

Figure 4 Gongolaria barbata (A) a mature receptacle; dotted lines indicate the median parts of the receptacle, where sections should be cut; (B) the same receptacle after 5 sections have been cut. Scale bars: 1 cm.
In the sections, the level of maturity will be assessed by noting the amount (number) and size of the oogonia. In the specific case of Gongolaria barbata, for which male specimens have been recorded in some populations, it is also recommendable to note if the conceptacles examined are hermaphroditic or antheridial.
The following three levels of maturity are defined:
1. Initial: the oogonia are not yet formed or they are still small, immature and in small numbers (Figures 5A, B);
2. Medium: the oogonia are in phase of development, in part mature and in part immature (Figures 5C, D);
3. Mature: the oogonia are numerous, mostly large and mature (Figures 5E, F).
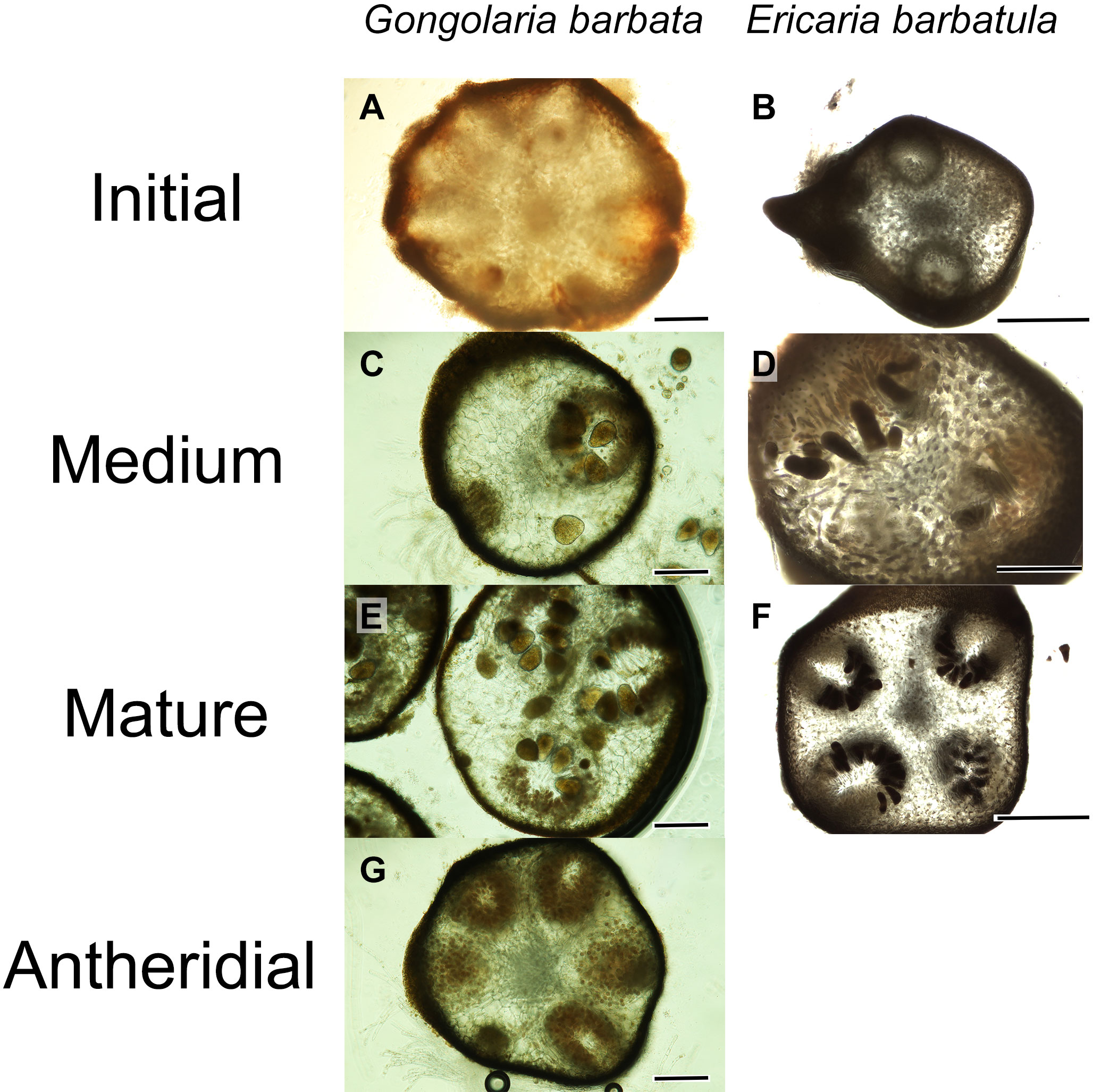
Figure 5 Cross sections illustrating maturity levels in receptacles of Gongolaria barbata and Ericaria barbatula (A) initial stage in G. barbata; (B) initial stage in E. barbatula; (C) medium stage in G. barbata; (D) medium stage in E. barbatula; (E) mature stage in G. barbata; (F) mature stage in E. barbatula; (G) antheridial conceptacle in G. barbata. Scale bars: (A–G): 200 μm; (B, F): 500 μm.
For Gongolaria barbata, attention should be paid not to confuse immature hermaphroditic conceptacles with antheridial conceptacles (Figure 5G). If male thalli occur in a population of Gongolaria barbata selected for monitoring, mature antheridial should be added as a fourth category to the three defined for the hermaphroditic conceptacles (Figure 5G).
The level of maturity is recorded for all receptacles collected. If different numbers of receptacles are collected in different monitoring dates (or from different sites), the maturity data will be best presented as percentages of receptacles belonging to the three categories defined above.
Results: a case study of monitoring
As example of application of the proposed protocol, we present here phenological data collected in a 13-months survey for a population of Gongolaria barbata of the central Adriatic shore (Piscinetta del Passetto, Ancona, Italy; 43°37’09” N, 13°31’54” E). The site is a shallow inlet 1-1.5 m deep, partially sheltered by a natural reef; its bedrock hosts a population of Gongolaria barbata consisting of a few hundreds of thalli. Vegetative and reproductive phenology were monitored from January 2022 to January 2023 at approximately monthly intervals (13 sampling dates: 15 January 2022; 16 February 2022; 14 March 2022; 27 April 2022; 12 May 2022; 14 June 2022; 20 July 2022; 22 August 2022; 3 October 2022; 26 October 2022; 17 November 2022; 8 December 2022; 6 January 2023). In each sampling date, fertility was assessed snorkelling by visual assessment of 20 randomly selected thalli. Two-three receptacles were randomly collected from 10 out of the 20 selected thalli for which fertility was assessed. The collected receptacles, placed in separate ziploc bags, were examined in the laboratory on the same day or the day after, and their level of maturity was determined by microscopic observation.
Results are shown in Figure 6. The highest levels of fertility were recorded in April and May 2022, when respectively 90% and 100% of the sampled thalli were in the fertility category 4 (> 75% of the branches bearing receptacles) (Figure 6A). In terms of maturity, receptacles occurred in the studied population throughout the whole study period except in July and August 2022. These months correspond to the annual highest sea surface temperatures recorded at the study site (26-28°C). At this time, the Gongolaria barbata specimens shed most of their branches, resulting in the reduced habitat devoid of receptacles typical of this species in summer months.
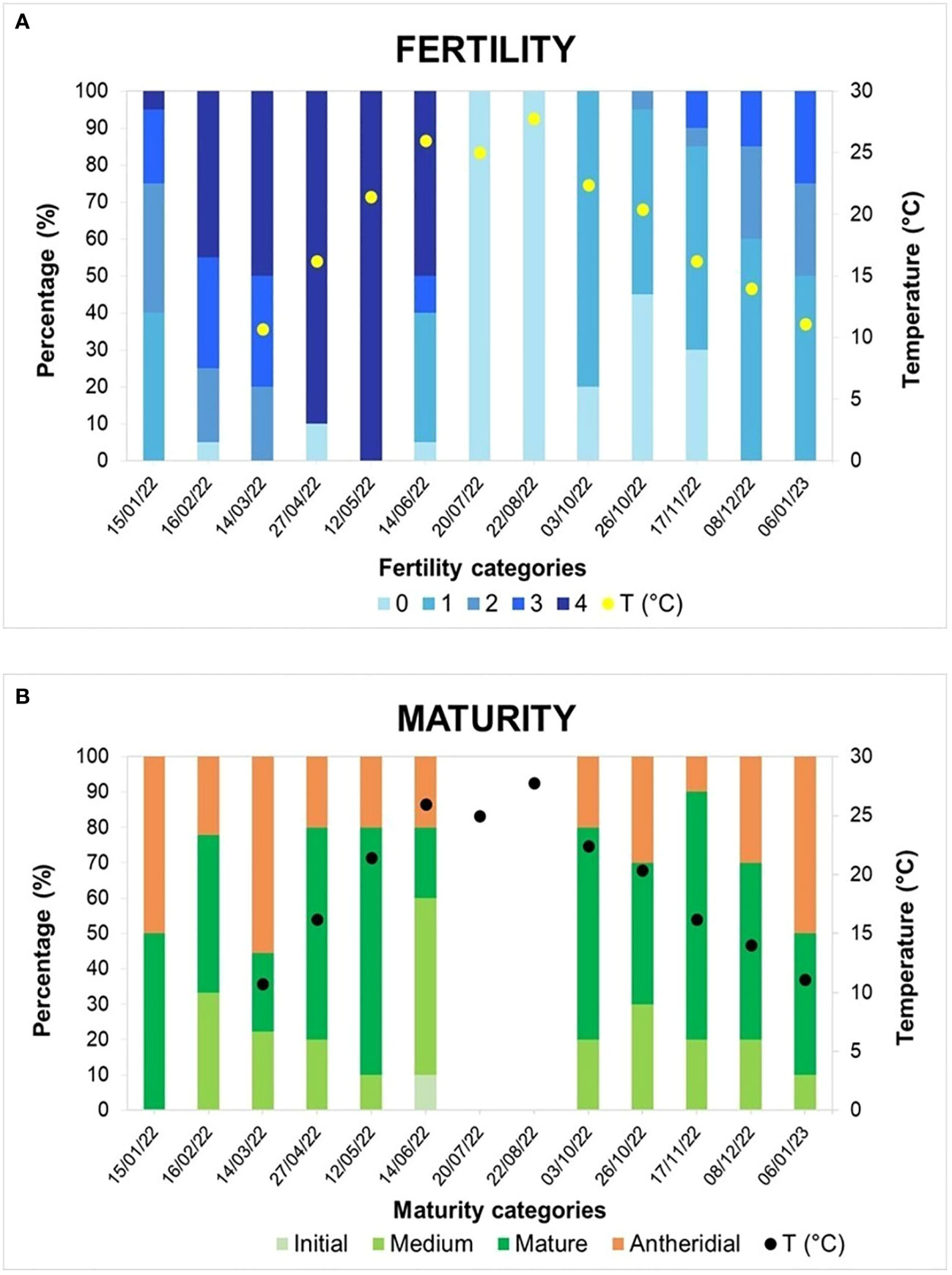
Figure 6 Reproductive phenology patterns detected in the monitored population of Gongolaria barbata from the Piscinetta del Passetto in the period January 2022-January 2023. Temperature values recorded in sampling dates from March 2022 to January 2023 are reported. (A) Fertility (B) maturity.
Microscopic examination of the receptacles showed that the studied population of Gongolaria barbata consisted of both hermaphroditic and male individuals (Figure 6B). Hermaphroditic thalli in medium or full stage of maturity were recorded throughout the study period except for July and August 2022 (when conceptacles were lacking). The peak of maturity was reached in May and November 2022, when fully mature hermaphroditic conceptacles were found in 70% of the sampled thalli. Male thalli bearing only antheridial conceptacles were recorded in all sampling dates in which receptacles were present; in some dates they represented approximately 50% of the sampled thalli (January 2022, March 2022 and January 2023). They were easily recognized as the mature antheridia formed a dense brownish orange layer on the internal surface of the conceptacles.
The reproductive phenology of Gongolaria barbata in the Piscinetta del Passetto agrees with information reported for this species in the literature. In Mediterranean populations of this species, gamete maturation and release take place mostly in spring and early summer (Sauvageau, 1912; Hamel, 1931; Ercegović, 1952; Marzocchi et al., 2003; Cebrian et al., 2021). In the case of the Piscinetta population, however, fertility and full maturity were extended to a much wider temporal span, including autumn and winter months. A remarkable feature of the studied population was the abundance of male thalli. To our knowledge, this phenomenon had previously been reported only by Sauvageau (1912) for populations from Banyuls-sur-Mer, France.
Discussion
Phenological monitoring of Cystoseira s.l. is crucial to detect in what period of the year populations develop reproductive structures. These data allow assessing fertility and maturity patterns for Cystoseira s.l. species/populations and provide indications about the environmental factors influencing the onset of gametogenesis. The protocol presented here is a user-friendly, easily reproducible tool responding to this need. It is primarily designed for marine scientists based in the Mediterranean area and involved in conservation and restoration of brown algal forests. However, with some preliminary training it can also be used by people who are not professional marine botanists, such as undergraduate students, staff of environmental protection agencies or citizen scientists. Although designed for restoration purposes, this protocol will also be useful for studies with different focus (e.g., reproductive responses of these seaweeds to future climatic changes) (see, for example, Monserrat et al., 2022). In this sense, the proposed protocol could be also applied to other fucoid species (such are Sargassum sp. or Fucus sp. genera among others), which also have an important role as habitat formers of underwater marine macroalgae forests (e.g., Coleman and Wernberg, 2017) and suffer regional declines (Mangialajo et al., 2008). In fact, those genera have been target in several phenological studies, however, although most of them have focused on the % of the plants being reproductive (Frequency of reproductive thalli)(Coleman and Brawley, 2005; El Atouani et al., 2021), fertility and maturity of conceptacles is usually disregarded. In this sense, we show that similar values in the percentage of fertile individuals in spring-early summer can mask very different values of fertility and maturity, leading to different reproductive potential among months. In terms of preliminary knowledge, a prerequisite is familiarity with the habit of receptacles and capacity to recognize them in the field. For this, useful descriptions and iconography can be found in references available from the web (Valiante, 1883: https://www.algaebase.org/search/bibliography/detail/?biblio_id=48364; Cormaci et al., 2012: https://www.algaebase.org/search/bibliography/detail/?biblio_id=50107).
For the assessment of fertility, estimation of the percentage of branches bearing receptacles may represent a partial source of uncertainty, since it may be influenced by observer-related subjectivity (and, even for the same observer, it may be subject to variation depending on the environmental conditions, i.e., sampling carried out in warm or cold water). For this reason, fertility is quantified in terms of five semi-quantitative categories, instead of exact percentage. This substantially reduces the risk of errors and ensures comparability of measurements taken by different observers.
Assessment of maturity is based on abundance and size of oogonia, for which presence and level of maturity are easily assessed by microscopical observation of cross sections (Figures 2, 5). For Gongolaria barbata, as previously mentioned, care should also be taken not to mistake antheridial conceptacles with immature hermaphroditic conceptacles. If accidentally only male receptacles are collected and used for a restoration action, fertilization will not take place and the operation will result into a complete failure.
We strongly recommend that for each monitoring date some important environmental data should be measured and reported, in particular seawater temperature and photoperiod. This information will allow to infer possible correlations between key abiotic factors and the onset of reproduction in the population monitored. Many experimental studies and field observations have shown that light, temperature or their combination play a key role in triggering the onset of fertility in temperate seaweeds (Liu et al., 2017 and references therein). A series of papers, predominantly addressing fertility in red and brown algae under laboratory conditions, highlighted the importance of photoperiod (daylength) and temperature as the most important environmental factors controlling reproduction in most seaweeds (Liu et al., 2017). For Mediterranean Cystoseira s.l., unfortunately, studies of this type are still very limited. Orfanidis (1991) showed that Gongolaria barbata (as former Cystoseira barbata) from the northern Aegean Sea was able to survive between -1°C and 30°C, growing optimally between 10°C and 20°C; he was, however, not able to induce fertility in the specimens used for his experiments. Recently, Papadimitriou et al. (2022), based on laboratory experiments, demonstrated that in Ericaria barbatula (Kützing) Molinari & Guiry formation of receptacles and fertilization took place only in short day conditions. This was the first study documenting a photoperiodic effect in a Mediterranean Cystoseira s.l. species. The presence of a few receptacles throughout the cultivating period under SD conditions is consistent with mature receptacles being found in the branches of a few specimens on September 27, 2015, which were collected from Cape Vrasidas, the same site as the material used in the present study. Throughout the entire year, fertile fronds seem not to be rare, since they have also been found in Ericaria abies-marina, which is morphologically similar to Ericaria barbatula, on the island of Gran Canaria (Valdazo et al., 2020).
Temperature may play an especially important role in restoration projects, as it may deeply affect both gametogenesis and survival of juvenile specimens. In the last decades, marine heatwaves (MHW) have become increasingly frequent in many Mediterranean regions. These phenomena strongly affect a multitude of benthic organisms, causing mass mortalities (Garrabou et al., 2022). If a MHW or some other thermal anomaly takes place at the time of gametogenesis of a Cystoseira s.l. population, reproduction and recruitment will be compromised. Bearing in mind that intensity and frequency of MHW are expected to increase, their consequences on the vegetative and reproductive phenology of Cystoseira s.l. could potentially impact the functioning of marine Mediterranean shallow rocky bottoms. MHW will also have disruptive effects if they take place shortly after a reproductive period, when numerous juvenile individuals are present. Germlings and juvenile thalli are typically the most sensitive stages in the life history of these seaweeds and are impacted by environmental disturbances much more heavily than adult specimens (de Caralt et al., 2020; Verdura et al., 2021).
Temperature can be measured using a portable temperature sensor or, if possible, a logger fixed in situ providing measurements at short time intervals (such as HOBO data loggers). Although not strictly necessary, data for additional variables such as light irradiance, salinity, nutrients and sediment load are also desirable. These will better define the environmental conditions in which gametangia reach maturation. We stress, however, that data of environmental variables collected in previous years should not be used as an absolute reference to guide restoration actions. Year-to-year variation in reproductive potential attributed to thermal anomalies have been documented: as an example, Bevilacqua et al. (2019) reported that in 2019 a population of Gongolaria barbata of the Gulf of Trieste became reproductive three months earlier than expected; the same population did not reproduce at all in the two subsequent years. A successful restoration action requires accurate knowledge of the reproductive phenology of the donor population shortly before the action is planned. We recommend that the protocol presented here should become a tool of widespread use for this purpose.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
EC, FR, AV, SC, SF, SB, SO, RD, JV, and LM participated in the study conception and design, all authors participated in data collection, FR, AV, and EC wrote the first draft of the manuscript, finalized by all the co-authors. All authors contributed to the article and approved the submitted version.
Funding
Financial support has been provided by the EASME–EMFF (Sustainable Blue Economy) Project AFRIMED (http://afrimed-project.eu/, grant agreement N. 789059), by Biodiversa+, the European Biodiversity Partnership under the 2021-2022 BiodivProtect joint call for research proposals, co-funded by the European Commission (GA N°101052342) and with the funding organisations, Agencia Estatal de Investigación AEI / 10.13039/501100011033/ NextGenerationEU/PRTR (PCI2022-135070-2, PCI2022-135052-2) and the Spanish Ministry of Science and Innovation (FoRestA Grant/Award No. PID2020- 112985GB-I00).
SB, RD, and FR acknowledge support from the National Biodiversity Future Center of Italy, funded by the European Community under the National Recovery and Resilience Plan (NRRP) for Italy.
Acknowledgments
Giulia Bellanti is acknowledged for assistance in the preparation of the figures.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer MC declared a past collaboration with the authors MMB to the handling editor.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alongi G., Catra M., Cormaci M. (1999). Fenologia morfologica e riproduttiva di Cystoseira elegans Sauvageau (Cystoseiraceae, Phaeophyta). Boll. Acc. Gioenia. Sci. Nat. 31, 189–200.
Ballesteros E. (1988). Estructura y dinámica de la comunidad de Cystoseira mediterranea Sauvageau en el Mediterráneo noroccidental. Investig. Pesq. 52, 313–334.
Battelli C., Catra M. (2023). Morphological and reproductive phenology of Cystoseira foeniculacea f. tenuiramosa (Phaeophyceae, Fucales) from the lagoon of Strunjan (Gulf of Trieste, northern Adriatic). Acta Adr. 64, 33–44. doi: 10.32582/aa.64.1.9
Bevilacqua S., Savonitto G., Lipizer M., Mancuso P., Ciriaco S., Srijemsi M., et al. (2019). Climatic anomalies may create a long-lasting ecological phase shift by altering the reproduction of a foundation species. Ecology 100, e02838. doi: 10.1002/ecy.2838
Bianchi C. N., Corsini-Foka M., Morri C., Zenetos A. (2014). Thirty years after: dramatic change in the coastal marine ecosystems of Kos Island (Greece), 1981-2013. Med. Mar. Sci. 15 (3), 482–497. doi: 10.12681/mms.678
Blanfuné A., Boudouresque C. F., Verlaque M., Thibaut T. (2016). The fate of Cystoseira crinita, a forest-forming fucale (Phaeophyceae. Stramenopiles), in France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 181, 196–208. doi: 10.1016/j.ecss.2016.08.049
Capdevila P., Linares C., Aspillaga E., Navarro L., Kersting D., Hereu B. (2015). Recruitment patterns in the Mediterranean deep-water alga Cystoseira zosteroides. Mar. Biol. 162, 1165–1174. doi: 10.1007/s00227-015-2658-0
Cebrian E., Tamburello L., Verdura J., Guarnieri G., Medrano A., Linares C., et al. (2021). A roadmap for the restoration of Mediterranean macroalgal forests. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.709219
Cheminée A., Sala E., Pastor J., Bodilis P., Thiriet P., Mangialajo L., et al. (2013). Nursery value of Cystoseira forests for Mediterranean rocky reef fishes. J. Exp. Mar. Bio. Ecol. 442, 70–79. doi: 10.1016/j.jembe.2013.02.003
Clausing R., de la Fuente G., Falace A., Chiantore M. (2022). Accounting for environmental stress in restoration of intertidal foundation species. J. Appl. Ecol. 60, 305–318. doi: 10.1111/1365-2664.14334
Coleman M. A., Brawley S. H. (2005). Variability in temperature and historical patterns in reproduction in the Fucus distichus complex (Heterokontophyta; Phaeophyceae): implications for speciation and the collection of herbarium specimens. J. Phycol 41, 1110–1119. doi: 10.1111/j.1529-8817.2005.00147.x
Coleman M. A., Wernberg T. (2017). Forgotten underwater forests: the key role of fucoids on Australian temperate reefs. Ecol. Evol. 7, 8406–8418. doi: 10.1002/ece3.3279
Cormaci M., Furnari G., Catra M., Alongi G., Giaccone G. (2012). Flora marina bentonica del Mediterraneo: Phaeophyceae. Boll. Acc. Gioenia Sci. Nat. 45, 1–508.
de Caralt S., Verdura J., Vergés A., Ballesteros E., Cebrian E. (2020). Differential effects of pollution on adult and recruits of a canopy-forming alga: implications for population viability under low pollutant levels. Sci. Rep. 10, 17825. doi: 10.1038/s41598-020-73990-5
De La Fuente G., Chiantore M., Asnaghi V., Kaleb S., Falace A. (2019). First ex situ outplanting of the habitat- forming seaweed Cystoseira amentacea var. stricta from a restoration perspective. PeerJ 7, e7290. doi: 10.7717/peerj.7290
El Atouani S., Belattmania Z., Kaidi S., Engelen A. H., Serrão E. A., Chaouti A., et al. (2021). Spatiotemporal patterns of phenology of the alien Phaeophyceae Sargassum muticum on the Atlantic coast of Morocco. Sci. Mar. 85, 103–111. doi: 10.3989/scimar.05118.010
Ercegović A. (1952). Jadranske cistozire. Njihova morfologija, ekologija i razvitak (Institut za Oceanografiju i Ribarstvo). Split, 212 pp.
Falace A., Kaleb S., de la Fuente G., Asnaghi V., Chiantore M. (2018). Ex situ cultivation protocol for Cystoseira amentacea var. stricta (Fucales, Phaeophyceae) from a restoration perspective. PloS One 13, e0193011. doi: 10.1371/journal.pone.0193011
Falace A., Zanelli E., Bressan G. (2005). Morphological and reproductive phenology of Cystoseira compressa (Esper) Gerloff & Nizamuddin (Fucales, Fucophyceae) in the gulf of Trieste (North Adriatic Sea). Annales Ser. Hist. Nat. 15 (1), 71–78.
Falace A., Zanelli E., Bressan G. (2006). Algal transplantation as a potential tool for artificial reef management and environmental mitigation. Bull.Mar. Sci. 78, 161–166.
Garrabou J., Gómez-Gras D., Medrano A., Cerrano C., Ponti M., Schlegel R., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Ch. Biol. 28 (19), 5708–5725. doi: 10.1111/gcb.16301
Gómez Garreta A., Barcelo i Martí M. C., Ribera Siguán M. A., Rull Lluch J. (2001). “Cystoseira C. Agardh,” in Flora Phycologica Iberica Vol. 1 Fucales, vol. 1 . Ed. Gómez Garreta A. (Murcia: Universidad de Murcia), 99–166.
Gómez-Garreta A., Ribera Siguán M. A., Seoane Camba J. A. (1982). Estudio fenológico de varias especies del género Cystoseira en Mallorca. Collect. Bot. 13 (2), 841–855.
Graham L., Graham J. M., Wilcox L. W., Cook M. E. (2016). Algae. 3rd ed. (Upper Saddle River). 689 pp.
Guern M. (1962). Embryologie de quelques espèces du genre Cystoseira Agardh 1821 (Fucales). Vie Milieu 1962, 649–680.
Gianni F., Bartolini F., Airoldi L., Ballesteros E., Francour P., Guidetti P., et al. (2013). Conservation and restoration of marine forests in the Mediterranean Sea and the potential role of Marine Protected Areas. Adv. Oceanogr. Limnol 4, 83–101. doi: 10.1080/19475721.2013.845604
Heesch S., Serrano-Serrano M., Barrera-Redondo J., Luthringer R., Peters A. F., Destombe C., et al. (2021). Evolutiomn of life cycles and reproductive traits: insights from the brown algae. J. Evol. Biol. 34, 992–1009. doi: 10.1111/jeb.13880
Hoffmann L., Renard R., Demoulin V. (1992). Phenology, growth and biomass of Cystoseira balearica in Calvi (Corsica). Mar. Ecol. Progr. Ser. 80 (2), 249–254. doi: 10.3354/meps080249
Lardi I. L., Varkitzi I., Tsiamis K., Orfanidis S., Koutsoubas D., Falace A., et al. (2022). Early development of Gongolaria montagnei (Fucales, Phaeophyta) germlings under laboratory conditions, with a view to enhancing restoration potential in the Eastern Mediterranean. Bot. Mar. 65 (4), 279–287. doi: 10.1515/bot-2021-0105
Liu X., Bogaert K., Engelen A. H., Leliaert F., Roleda M. Y., De Clerck O. (2017). Seaweed reproductive biology: environmental and genetic controls. Bot. Mar. 60 (2), 89–108. doi: 10.1515/bot-2016-0091
Lotze H. K., Coll M., Dunne J. A. (2011). Historical changes in marine resources, food web structure and ecosystem functioning in the Adriatic Sea, Mediterranean. Ecosystems 14, 198–222. doi: 10.1007/s10021-010-9404-8
Mangialajo L., Chiantore M., Cattaneo-Vietti R. (2008). Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 358, 63–74. doi: 10.3354/meps07400
Mariani S., Cefalì M. E., Chappuis E., Terradas M., Pinedo S., Torras X., et al. (2019). Past and present of Fucales from shallow and sheltered shores in Catalonia. Reg. Stud. Mar. Sci. 32, 100824. doi: 10.1016/j.rsma.2019.100824. 1-18.
Marzocchi M., Curiel D., Dri C., Scattolin M. (2003). Fenologia morfologica e riproduttiva di Cystoseira barbata (Stackhouse) C. Agardh var. barbata (Fucales, Fucophyceae) nella laguna di Venezia (Nord adriatico). Lav. Soc Ven. Sc. Nat. 28, 21–23.
Medrano A., Hereu B., Cleminson M., Pagès-Escolà M., Rovira G., Solà J., et al. (2020a). From marine deserts to algal beds: Treptacantha elegans revegetation to reverse stable degraded ecosystems inside and outside a No-Take marine reserve. Restor. Ecol. 28, 632–644. doi: 10.1111/rec.13123
Medrano A., Hereu B., Mariani S., Neiva J., Escol M. P. (2020b). Ecological traits, genetic diversity and regional distribution of the macroalga Treptacantha elegans along the Catalan coast (NW Mediterranean Sea). Sci. Rep. 10, 19219. doi: 10.1038/s41598-020-76066-6. 1-10.
Monserrat M., Comeau S., Verdura J., Alliouane S., Spennato G., Priouzeau F., et al. (2022). Climate change and species facilitation affect the recruitment of macroalgal marine forests. Sci. Rep. 12, 1–12. doi: 10.1038/s41598-022-22845-2
Orfanidis S. (1991). Temperature responses and distribution of macroalgae belonging to the warm-temperate Mediterranean-Atlantic distribution group. Bot. Mar. 34, 541–552. doi: 10.1515/botm.1991.34.6.541
Orfanidis S., Rindi F., Cebrian E., Fraschetti S., Nasto I., Taskin E., et al. (2021). Effects of natural and anthropogenic stressors on fucalean brown seaweeds across different spatial scales in the Mediterranean Sea. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.658417
Orlando-Bonaca M., Savonitto G., Asnaghi V., Trkov D., Pitacco V., Sisko M., et al. (2022). Where and how – new insight for brown algal forest restoration in the Adriatic. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.988584
Papadimitriou A., Nakou K., Papathanasiou V., Orfanidis S. (2022). Effects of photoperiod and temperature on ecophysiological responses of Ericaria barbatula (Phaeophyceae): a long-term study. Bot. Mar. 65 (4), 269–277. doi: 10.1515/bot-2022-0009
Pardi G., Piazzi L., Cinelli F. (2000). Demographic study of a Cystoseira humilis Kützing (Fucales: Cystoseiraceae) population in the western Mediterranean. Bot. Mar. 43, 81–86. doi: 10.1515/BOT.2000.007
Perkol-Finkel S., Airoldi L. (2010). Loss and recovery potential of marine habitats: an experimental study of factors maintaining resilience in subtidal algal forests at the Adriatic Sea. PloS One 5, e10791. doi: 10.1371/journal.pone.0010791
Perkol-Finkel S., Ferrario F., Nicotera V., Airoldi L. (2012). Conservation challenges in urban seascapes: promoting the growth of threatened species on coastal infrastructures. J. Appl. Ecol. 49, 1457–1466, doi: 10.1111/j.1365-2664.2012.02204.x.
Pinedo S., Zabala M., Ballesteros E. (2013). Long-term changes in sublittoral macroalgal assemblages related to water quality improvement. Bot. Mar. 56, 461–469. doi: 10.1515/bot-2013-0018
Pizzuto F., Presti C., Serio D. (1995). Struttura e periodismo di un popolamento a Cystoseira amentacea v. stricta Montagne (Fucales, Fucophyceae) del litorale catanese. Boll. Acc. Gioenia Sci. Nat. 28 (349), 23–43.
Robvieux P. (2013). Conservation des populations de Cystoseira en regions Provence-Alpes-Côte-d’Azur et Corse. [dissertation thesis] (Université Nice Sophia Antipolis).
Sala E., Ballesteros E., Dendrinos P., Di Franco A., Ferretti F, Foley, et al. (2012). The structure of mediterranean rocky reef ecosystems across environmental and human gradients, and conservation implications. PloS One 7 (2), e32742. doi: 10.1371/journal.pone.0032742
Sauvageau C. (1912). A propos des Cystoseira de Banyuls et Guéthary. Bull. Stn. Biol. d’Arcachon 14, 133–556.
Serio D. (1995a). Fenologia morfologica e riproduttiva di Cystoseira spinosa Sauvageau v. compressa (Ercegovic) Cormaci et al. (Fucales, Fucophyceae). Boll. Acc. Gioenia Sci. Nat. 28 (349), 5–22.
Serio D. (1995b). Fenologia morfologica e riproduttiva di Cystoseira zosteroides C. Agardh (Fucales, Fucophyceae). Boll. Acc. Gioenia Sci. Nat. 28 (349), 45–58.
Smith C. J., Verdura J., Papadopoulou N., Fraschetti S., Cebrian E., Fabbrizzi E, et al. (2013). A decision-support framework for the restoration of Cystoseira sensu lato forests. Front. Mar. Sci 10, 1159262. doi: 10.3389/fmars.2023.1159262
Susini M. L., Mangialajo L., Thibaut T., Meinesz A. (2007). Development of a transplantation technique of Cystoseira amentacea var. stricta and Cystoseira compressa. Hydrobiologia 580, 241–244. doi: 10.1007/s10750-006-0449-9
Tamburello L., Chiarore A., Fabbrizzi E., Colletti E., Franzitta G., Grech D., et al. (2022). Can we preserve and restore overlooked macroalgal forests? Sci. Tot. Env. 806, 150855. doi: 10.1016/j.scitotenv.2021.150855
Thibaut T., Blanfuné A., Boudouresque C. F., Verlaque M. (2015). Decline and local extinction of Fucales in the French Riviera: the harbinger of future extinctions? Mediterr. Mar. Sci. 16, 206–224. doi: 10.12681/mms.1032
Thibaut T., Pinedo S., Torras X., Ballesteros E. (2005). Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. Pollut. Bull. 50, 1472–1489. doi: 10.1016/j.marpolbul.2005.06.014
UNEP/MAP. (2013). United nations environment programme/mediterranean action plan [UNEP/MAP] (2013). Protocol concerning specially protected areas and biological diversity in the mediterranean. list of endangered species. Athina: UNEP/MAP.
Valdazo J., Viera-Rodriguez M. A., Tuya F. (2020). Seasonality in the canopy structure of the endngered brown macroalga Cystoseira abies-marina at Gran Canaria Island (Canary Islands, eastern Atlantic). Eur. J. Phycol. 55 (3), 253–265. doi: 10.1080/09670262.2019.1696989
Valiante R. (1883). Le Cystoseirae del Golfo di Napoli (Leipzing: Verlag von Wilhelm Egelmann). 30 pp.
Verdura J., Sales M., Ballesteros E., Cefalì M. E., Cebrian E. (2018). Restoration of a canopy-forming alga based on recruitment enhancement: methods and long-term success assessment. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01832
Keywords: macroalgae, phenology, Cystoseira, Mediterranean, conservation
Citation: Rindi F, Vergés A, Zuchegna I, Bianchelli S, de Caralt S, Galobart C, Santamaría J, Martini F, Monserrat M, Orfanidis S, Sitjà C, Tsioli S, Verdura J, Mangialajo L, Fraschetti S, Danovaro R and Cebrian E (2023) Standardized protocol for reproductive phenology monitoring of fucalean algae of the genus Cystoseira s.l. with potential for restoration. Front. Mar. Sci. 10:1250642. doi: 10.3389/fmars.2023.1250642
Received: 30 June 2023; Accepted: 25 September 2023;
Published: 19 October 2023.
Edited by:
Leonel Pereira, University of Coimbra, PortugalReviewed by:
Marcos Rubal, University of Minho, PortugalMariachiara Chiantore, University of Genoa, Italy
Copyright © 2023 Rindi, Vergés, Zuchegna, Bianchelli, de Caralt, Galobart, Santamaría, Martini, Monserrat, Orfanidis, Sitjà, Tsioli, Verdura, Mangialajo, Fraschetti, Danovaro and Cebrian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emma Cebrian, emma@ceab.csic.es