 Luciana F. Brito1,2†
Luciana F. Brito1,2† Marina Gil López1,2†
Marina Gil López1,2† Lucas Straube1,2
Lucas Straube1,2 Luciane M. P. Passaglia3
Luciane M. P. Passaglia3 Volker F. Wendisch1,2*
Volker F. Wendisch1,2*- 1Genetics of Prokaryotes, Faculty of Biology, Bielefeld University, Bielefeld, Germany
- 2Center for Biotechnology, Bielefeld University, Bielefeld, Germany
- 3Department of Genetics, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Due to the importance of phosphorus (P) in agriculture, crop inoculation with phosphate-solubilizing bacteria is a relevant subject of study. Paenibacillus sonchi genomovar Riograndensis SBR5 is a promising candidate for crop inoculation, as it can fix nitrogen and excrete ammonium at a remarkably high rate. However, its trait of phosphate solubilization (PS) has not yet been studied in detail. Here, differential gene expression and functional analyses were performed to characterize PS in this bacterium. SBR5 was cultivated with two distinct P sources: NaH2PO4 as soluble phosphate source (SPi) and hydroxyapatite as insoluble phosphate source (IPi). Total RNA of SBR5 cultivated in those two conditions was isolated and sequenced, and bacterial growth and product formation were monitored. In the IPi medium, the expression of 68 genes was upregulated, whereas 100 genes were downregulated. Among those, genes involved in carbon metabolism, including those coding for subunits of 2-oxoglutarate dehydrogenase, were identified. Quantitation of organic acids showed that the production of tricarboxylic acid cycle-derived organic acids was reduced in IPi condition, whereas acetate and gluconate were overproduced. Increased concentrations of proline, trehalose, and glycine betaine revealed active osmoprotection during growth in IPi. The cultivation with hydroxyapatite also caused the reduction in the motility of SBR5 cells as a response to Pi depletion at the beginning of its growth. SBR5 was able to solubilize hydroxyapatite, which suggests that this organism is a promising phosphate-solubilizing bacterium. Our findings are the initial step in the elucidation of the PS process in P. sonchi SBR5 and will be a valuable groundwork for further studies of this organism as a plant growth-promoting rhizobacterium.
Introduction
Phosphorus (P) is a macronutrient required for plant growth. Plant roots can absorb P in the form of orthophosphates, either H2PO4– or HPO42–, but the concentration of these ions in the soil is in the micromolar range (Shin et al., 2004; Ai et al., 2009). This is due to the complex dynamics of P in the soil; this nutrient has a unique characteristic, which is its high fixation in soil. Mineral P can be, for example, found associated with the surface of iron or aluminum oxides in the soil, making it poorly soluble and therefore unavailable for plant nutrition (Shen et al., 2011). Those factors lead to the overuse of chemical P fertilizers and animal manure applied to agricultural land, which—although soil fertility and crop production are improved—caused severe environmental damage in the past decades, such as eutrophication of rivers and lakes, and the input of cadmium and radionuclides in the soil from contaminated fertilizers (Vance et al., 2003; Attallah et al., 2019; Li et al., 2020; Lee et al., 2020). Moreover, the input demand of P into croplands is expected to increase up to 86% by 2050 (Mogollón et al., 2018). It is also estimated an absolute shortage of P supply from industrial fertilizers in the future, which is aggravated by the depletion of the P present in agricultural soils caused by erosion (Alewell et al., 2020). Therefore, P is receiving more attention as a non-renewable resource (Cordell et al., 2009). Improvement of P acquisition and use by plants is critical for economic and environmental reasons. In this context, phosphate solubilization (PS) performed by organisms in the soil is an interesting target for study. PS ability is well characterized in mycorrhizal fungi (Delavaux et al., 2017) and phosphate-solubilizing bacteria (PSB), mostly associated with the plant rhizosphere (Rodríguez and Fraga, 1999). Endophytic and mycorrhizal fungal strains have a significant impact on plant P nutrition, being used by many agronomists as soil inoculants to enhance plant growth (Khan et al., 2010). Among rhizosphere fungi, the most common P-solubilizing genera are Aspergillus, Penicillium, and Trichoderma (Sharma et al., 2013; Wang et al., 2020). There is a substantial number of PSB genera widespread in the rhizosphere (Chen et al., 2006), of which some are well-established as plant growth-promoting rhizobacteria (PGPR), e.g., Pseudomonas (Parvaze et al., 2007), Bacillus (Chen et al., 2006), Enterobacter (Chung et al., 2005), and Azotobacter (Kumar et al., 2001). The use of PSB as inoculants leads to increased P uptake by the plant and improved crop yield (Nosratabad et al., 2017; Din et al., 2020; Magallon-Servin et al., 2020). Besides providing soluble P to plants, these organisms promote plant growth and development by other activities, such as nitrogen fixation and production of plant phytohormones (Zaidi et al., 2009).
Among the PS mechanisms utilized by PSB, the production of organic acids is well recognized and most common in rhizobacteria. The organic acids excreted by PSB have different modes of action that lead to PS: mineral ions bound to precipitated Pi can be chelated by these organic acids or P can be dissolved by decreasing the pH (Rashid et al., 2004). Among the genera considered as PSB, Paenibacillus has shown to be promising for inoculation purposes. Paenibacillus members that present PS activity have been isolated from the rhizosphere of various crops (Seldin et al., 1998; Tcherpakov et al., 1999; Beneduzi et al., 2010). In some Paenibacillus species such as Paenibacillus mucilaginosus (Hu et al., 2006), Paenibacillus elgii (Narayan et al., 2010), Paenibacillus kribbensis (Marra et al., 2012), Paenibacillus polymyxa, Paenibacillus macerans (Wang et al., 2012), and Paenibacillus xylanilyticus (Pandya et al., 2015), the PS ability has been confirmed. In a comparative genomic analysis, it was suggested that a wide group of Paenibacillus can perform PS by producing the organic acid gluconate, as several Paenibacillus species (i.a. P. polymyxa, Paenibacillus peoriae, and Paenibacillus sp.) showed high conservation of the genes coding for the enzymes glucose 1-dehydrogenase and gluconate dehydrogenase, which are involved in the synthesis of this compound (Xie et al., 2016).
Paenibacillus sonchi genomovar Riograndensis is a rod-shaped, Gram-positive, motile, nitrogen-fixing bacterium (Beneduzi et al., 2010; Sant’Anna et al., 2017). The strain SBR5 was isolated from the rhizosphere of wheat (Triticum aestivum) plants cultivated in Rio Grande do Sul, Brazil (Beneduzi et al., 2008). In addition to nitrogen fixation, SBR5 possesses further plant growth-promoting activities, such as the ability to produce indole-3- acetic acid and to excrete a high amount of ammonium (Fernandes et al., 2014; Ambrosini et al., 2018). The complete genome of P. sonchi SBR5 (Genbank accession LN831776) was sequenced and fully annotated and consists of one chromosome of 7,893,056 bp, containing 6,705 protein-coding genes, 87 tRNAs, and 27 rRNAs genes (Brito et al., 2015). RNAseq has been used to chart its global transcriptomic landscape (Brito et al., 2017a). Moreover, molecular tools for gene expression (Brito et al., 2017b) and CRISPR interference-based gene repression (Brito et al., 2020) were developed for this organism. Hence, a methodology is in place to study the poorly characterized PS by SBR5. To elucidate the PS process in this bacterium, we performed RNAseq and physiological analyses with two inorganic P sources: soluble and insoluble.
Materials and Methods
Strains and Growth Conditions
The PGPR bacterium P. sonchi genomovar Riograndensis SBR5 was obtained from the strain collection of the Department of Genetics at Universidade Federal do Rio Grande do Sul, Brazil. SBR5 and Escherichia coli DH5α cells were used as expression host and molecular cloning host, respectively. All bacterial strains utilized in the present study are listed in Supplementary Table 1. E. coli cells were routinely cultivated in Luria–Bertani medium, composed of 10 g L–1 tryptone, 5 g L–1 yeast extract, and 10 g L–1 NaCl, at 37°C and 120 rpm, and chloramphenicol (20 μg ml–1) and ampicillin (100 μg ml–1) were added to cultivation broth when necessary. Precultures of SBR5 transformants were routinely cultivated in Caso broth (DSMZ 220), containing 15 g L–1 peptone from casein, 5 g L–1 peptone from soymeal, and 5 g L–1 NaCl, and chloramphenicol (5 μg ml–1) and ampicillin (10 μg ml–1) were added to medium when necessary. Bacterial cells were grown in Caso broth agar plates, and a single colony was picked and transferred to 500 ml shaking flasks containing 50 ml of the same medium, and the flask was incubated overnight at 30°C and 120 rpm. One milliliter of the bacterial suspension was inoculated into 500 ml shaking flasks containing 50 ml of distinct broth media differing with regard to the P source: (1) Pikovskaya broth prepared as Pikovskaya (1948) with 5 g L–1 of Ca5(PO4)3(OH) (hydroxyapatite) as P source (IPi); (2) Pikovskaya broth with 3.2 g L–1 NaH2PO4 as P source (SPi). The pH of the media was adjusted to 7.0 before autoclaving, and the pH neutrality was maintained by the addition of 12 g L–1 of 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid. The cells grown in each P source were used as inoculum in fresh media of the same condition. For each condition tested, six biological replicates were used: three for total RNA isolation and three for further determination of growth characteristics. Bacterial cells of three replicate cultivations were harvested after 6 h of growth, in the middle of the exponential phase (Supplementary Figure 1). The harvesting procedure was done according to Irla et al. (2016); 20 ml of each sample was collected in 50-ml falcon tubes filled with shaved ice and centrifuged at 4°C for 10 min at 4,000 rpm. The resulting cell pellets were immediately frozen in liquid nitrogen after centrifugation and stored at −80°C until further use. Additionally, SBR5 was cultivated in CGXII minimal medium (Keilhauer et al., 1993) with some modifications. CGXII medium was composed of 20 g L–1 ammonium sulfate, 5 g L–1 urea, and 12 g L–1 2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid, with supplementation of a trace salts solution (Keilhauer et al., 1993), 9 g L–1 glucose, and 0.1 mg L–1 biotin. The phosphate content of the medium varied with gradually increasing concentrations of NaH2PO4 (0, 13, 26, 39, 52, and 65 mM) to obtain microbial growth kinetics. The precultures were grown in CGXII with either sufficient (70 mM) or depleted (20 mM) Pi. For growth analysis, SBR5 cells were cultivated in IPi, SPi, low phosphate (LPi), and high phosphate (HPi) media. LPi and HPi media were prepared according to Qi et al. (1997), with some modifications. Both media were composed of 4 g L–1 ammonium sulfate, 0.18 g L–1 sodium citrate, 0.5 g L–1 ferric chloride, 0.15 g L–1 manganese sulfate, 4.2 g L–1 magnesium sulfate, 6 g L–1 Trizma, 1.3 mg L–1 zinc chloride, and 12.3 mg L–1 (LPi) or 0.49 g L–1 (HPi) K2HPO4; both media were supplemented 9 g L–1 glucose, 0.5 g L–1 casamino acids, and 0.1 mg L–1 biotin. Two milliliters of cultivation broth were taken for measurements of optical density at 600 nm (OD600 nm) and pH (pH indicator sticks, Macherey-Nagel, Düren, Germany), and supernatants and cell pellets (n = 3) were collected by centrifugation (10 min at 4,000 rpm). The initial OD600 nm of main cultivations was approximately 0.05. During the cultivations, P. sonchi cells were routinely shaken at 120 rpm at a temperature of 30°C.
RNA Isolation and Preparation and Sequencing of Complementary DNA Libraries
To isolate the total RNA from SBR5 cells, bacterial cell pellets previously harvested and stored at −80°C were thawed in ice, and RNA was extracted individually for each cultivation condition. All the procedures regarding RNA isolation and RNA quality control were done according to Brito et al. (2017a). The three replicates of extracted RNA samples of each condition were pooled in equal parts, and the pool of total RNA was subsequently used for the preparation of complementary DNA (cDNA) libraries. For each condition, a whole transcriptome library was prepared. For removal of ribosomal RNA, Ribo-Zero Plus rRNA Depletion Kit (Illumina, San Diego, United States) was used according to manufacturer’s recommendations. The preparation of this library and the cDNA sequencing were carried out according to Mentz et al. (2013).
Bioinformatics Analysis
The sequence reads were mapped onto the reference genome of P. sonchi genomovar Riograndensis SBR5 (Brito et al., 2015). To prepare the reads for mapping, the tool Trimmotatic version 0.33 (Bolger et al., 2014) was used to trim the sequences to a minimal length of 35 bp. Trimmed reads were mapped to the reference genome of SBR5 through the software for short read alignment Bowtie (Langmead, 2010). The visualization of mapped reads and the differential gene expression analysis were performed using the software ReadXplorer (Hilker et al., 2014), in which the statistical method DEseq was used to analyze the resultant RNAseq data (Anders and Huber, 2010). The cutoff values for designating a gene as differentially expressed included a change in expression level (base mean) higher than 30 and an adjusted P-value equal to or less than 0.05. When differentially expressed genes coding for proteins of unknown function were detected, the gene sequences were submitted to BLASTx analysis to identify the protein family conservations (Altschup et al., 1990).
Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction
To validate the data obtained by RNAseq analysis, real-time quantitative reverse transcription-polymerase chain reaction (qRT-PCR) was performed utilizing a LightCycler system (Roche Diagnostics, Penzberg, Germany). The RNA samples utilized to compose the RNA pools for cDNA library preparation in this study were also utilized as templates for qRT-PCR. Differentially expressed genes selected for this assay and the characteristics of the primers utilized are listed in Supplementary Table 2. Furthermore, we selected the genes glpQ, phoH, phoU, pstB3, and pstS, which are differentially expressed in the condition of Pi starvation in bacteria (Baek and Lee, 2007; Myers et al., 2016; Devine, 2018), to be targeted in the qRT-PCR analysis. Therefore, to obtain messenger RNA from SBR5 in Pi starvation conditions, SBR5 cells were cultivated in HPi medium until they reached exponential phase and subsequently transferred to LPi medium or HPi control medium for RNA isolation at the times 15, 60 min, 6, and 24 h. For analysis of the isolated RNA, qRT-PCR was performed utilizing a CFX Connect Real-Time PCR Detection System (BioRad Laboratories, Hercules, United States). Therefore, each sample concentration was adjusted to 50 ng RNA μl–1, and 1 μl thereof was pipetted into a reaction mixture of the SensiFAST SYBR No-ROX Kit (Bioline, Luckenwalde, Germany), following manufacturer’s instructions. To evaluate relative gene expression, 16S rRNA was used as reference control (Sperb et al., 2016). The melting-curve data-based quantification cycle (Cq) values were used for further calculation. For the calculation of relative gene expression, the following equation was used: ΔΔCq = 2–(CqIpI – Cq16S) / 2–(CqSPi – Cq16S) (Livak and Schmittgen, 2001).
Molecular Cloning, Plasmid DNA Construction, and Preparation of Recombinant Strains
Molecular cloning was performed as described previously (Sambrook, 2001). Chemically competent cells of E. coli DH5α were prepared for cloning (Hanahan and Harbor, 1983). All plasmids obtained in this study and all oligonucleotide sequences utilized in PCR are listed in Supplementary Table 1. Genomic DNA of P. sonchi SBR5 was isolated using NucleoSpin Microbial DNA kit (Machery-Nagel, Düren, Germany). The NucleoSpin Gel and PCR Clean-up kit (Machery-Nagel) was used for PCR cleanup, and plasmids were isolated using the GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific, Waltham, United States). Plasmid backbones were digested using restriction enzyme BamHI (Thermo Fisher Scientific). Double-stranded DNA inserts were amplified using Phusion DNA polymerase (New England Biolabs, Ipswich, England) and the overlapping regions joined by Gibson assembly (Gibson, 2014). For colony PCR, Taq polymerase (New England Biolabs) was used according to the manufacturer’s recommendations. The sequence correctness of resultant plasmids was checked by sequencing in the Center for Biotechnology in Bielefeld, Germany. Plasmid DNA was used to transform P. sonchi SBR5 cells as in Brito et al. (2017b).
Quantification of Free Inorganic Orthophosphates in Cell Pellets and Supernatants
PS efficiency of SBR5 cultivated in the different conditions (IPi, SPi, LPi, and HPi) was determined through the quantification of orthophosphates present in the supernatants or cell lysates. The cells were lysed by ultrasonication for 6 min at on and off cycles of 0.5 s and amplitude 55% (Ultraschalldesintegrator Sonoplus GM 200, Sonotrode M72, Bandelin electronic GmbH & Co KG, Berlin, Germany) and centrifuged for 30 min at 4°C and 13,200 rpm. This assay was performed through the molybdenum-blue method, as described by Murphy and Riley (1958).
Metabolite Analyses
For quantification of glucose and organic acids in the supernatants and cell lysates of SBR5, a high-performance liquid chromatography system (HPLC, 1200 series, Agilent Technologies Deutschland GmbH, Böblingen, Germany) was used as in Zahoor et al. (2014). Quantification was done by calibration with external standards. By this method, gluconate, oxoglutarate, acetate, citrate, succinate, oxalate, and malate of cell supernatants at 20 h, at the end of RNAseq cultivation (Supplementary Figure 1), were determined. Moreover, oxoglutarate, trehalose, and glycine betaine were also measured in supernatants and cell lysates, along with the growth of SBR5 in IPi and SPi. Furthermore, 2-oxoglutarate dehydrogenase (2-OGDH) activity of SBR5 crude extracts in IPi or SPi was measured according to Nguyen et al. (2015). To obtain the concentration of amino acids present in cell supernatants of SBR5 at the end of its growth in RNAseq cultivation (Supplementary Figure 1), HPLC samples were derivatized with ortho-phthaldialdehyde and separated on a system consisting of a pre-column (LiChrospher 100 RP8 EC-5μ, 40 × 4.6 mm, CS-Chromatographie Service GmbH, Langerwehe, Germany) and the main column (LiChrospher 100 RP8 EC-5μ, 125 × 4.6 mm, CS-Chromatographie Service GmbH, Langerwehe, Germany) and detected by fluorescence (FLD G1321A, 1200 series, Agilent Technologies) (Jorge et al., 2017b). Asparagine was used as an internal standard, and external standards of glutamate, valine, arginine, and ornithine were prepared for calibration. For proline measurement, samples were derivatized with 9-fluorenylmethyl chloroformate as in Jensen et al. (2015). Furthermore, trehalose, glycine betaine, oxoglutarate, proline, and glutamate were measured in supernatants and cell lysates of SBR5 along with its growth in SPi or IPi.
Determination of Osmolality Levels
Osmolality measurements of IPi and SPi samples (supernatant and lysates from cell pellets) were done by freezing point depression using an Osmomat auto (Gonotec, Berlin, Germany) according to the manufacturer’s instructions. Supernatant samples and cell lysates were directly used for the measurement. The total osmolality of each sample was determined by comparative measurements of the freezing points of pure water and the test solutions using 50 μl of the sample.
Flow Cytometry
Recombinant P. sonchi SBR5 cells were cultivated in SPi or IPi supplemented with chloramphenicol or ampicillin. Cells were harvested in the mid-exponential phase of growth and washed three times in NaCl 0.9%. Next, the OD600 nm of the samples was normalized to 0.5. To quantify the fluorescence intensities in the cell suspensions, they were submitted to flow cytometry (Beckman Coulter, Brea, United States) and the data analyzed in the Beckman Coulter Kaluza Flow Analysis Software. The settings for the emission signal and filters within the flow cytometer for detection of GfpUV and Crimson fluorescence were 550/525 bandpass FL9 filter and 710/660 bandpass FL6 filter, respectively.
Statistical Analysis
Student’s t-test was used to assess statistical differences in mean orthophosphate concentrations over time in the supernatants of SBR5 cultivated in IPi condition. Results were considered statistically significant when p-values were <0.01 or <0.001.
Results
Growth and Gene Expression of P. sonchi SBR5 With Different Pi Concentrations
Growth of P. sonchi SBR5 in CGXII supplemented with increasing Pi concentrations (0-, 13-, 26-, 39-, 52-, and 65-mM NaH2PO4) was compared, with precultures grown with sufficient (70 mM) or depleted (20 mM) Pi. Only sub-millimolar Pi concentrations reduced growth when precultures were grown with sufficient Pi (data not shown), which indicated that an inner cell P storage, such as polyphosphate granule (Racki et al., 2017), may exist. Therefore, precultures were grown in depleted Pi. A Monod constant for NaH2PO4 of approximately 10 mM was determined, as this concentration supported a half-maximal growth rate (Supplementary Figure 2). The final biomass (ΔOD) of SBR5 cells was negatively affected by Pi depletion when the supplied NaH2PO4 concentration was inferior to 26 mM (Supplementary Figure 2).
To analyze a Pi down shock, precultures grown in HPi were transferred to HPi or LPi media, containing 5 or 0.13 mM of Pi, respectively. LPi condition is expected to be similar to IPi condition with respect to the soluble Pi concentration in the culture broth. During growth in LPi and HPi conditions, pH values of supernatant remained neutral (data not shown), and growth in LPi condition was only slightly lower than in HPi (Figure 1A). The Pi concentration in the supernatants decreased 30% during growth in LPi condition (Figure 1B), whereas the intracellular Pi concentration dropped drastically during growth under both conditions (Figure 1C).

Figure 1. Growth (A) and orthophosphate concentrations in the supernatants (B) and cell lysates (C) of P. sonchi SBR5 cultivated in high phosphate medium (HPi, closed circles) or low phosphate medium (LPi, open circles). CDW, cell dry weight. Data given as means and standard deviations of biological triplicates.
To verify if Pi inducible genes known from other bacteria (glpQ, phoH, phoU, pstB3, and pstS) (Baek and Lee, 2007; Myers et al., 2016; Devine, 2018) show increased RNA levels in SBR5 upon transfer from HPi to LPi culture broth, P. sonchi SBR5 cells cultivated in HPi medium were transferred to HPi or LPi, and RNA samples were taken after 15, 60 min, 6, or 24 h for qRT-PCR analysis. All the targeted genes showed increased RNA levels upon transfer to the LPi medium (Figure 2). The genes pstB3 and pstS, coding for components of the high-affinity Pi transport system, and the gene phoU, encoding a phosphate-specific transport system accessory protein, were highly expressed 15, 60 min, and 6 h after the transfer to LPi condition but downregulated at 24 h. The gene phoH, coding for a Pi starvation protein, was expressed only at 15 min after the transfer to LPi condition, whereas the glycerophosphodiester phosphodiesterase gene qlpQ showed increased ΔCq levels at 6 and 24 h after the transfer to LPi condition. Pi induction of pstS was strongest (approximately hundredfold after the transfer from HPi to LPi condition; Figure 2), indicating pstS as a good indicator of Pi starvation.
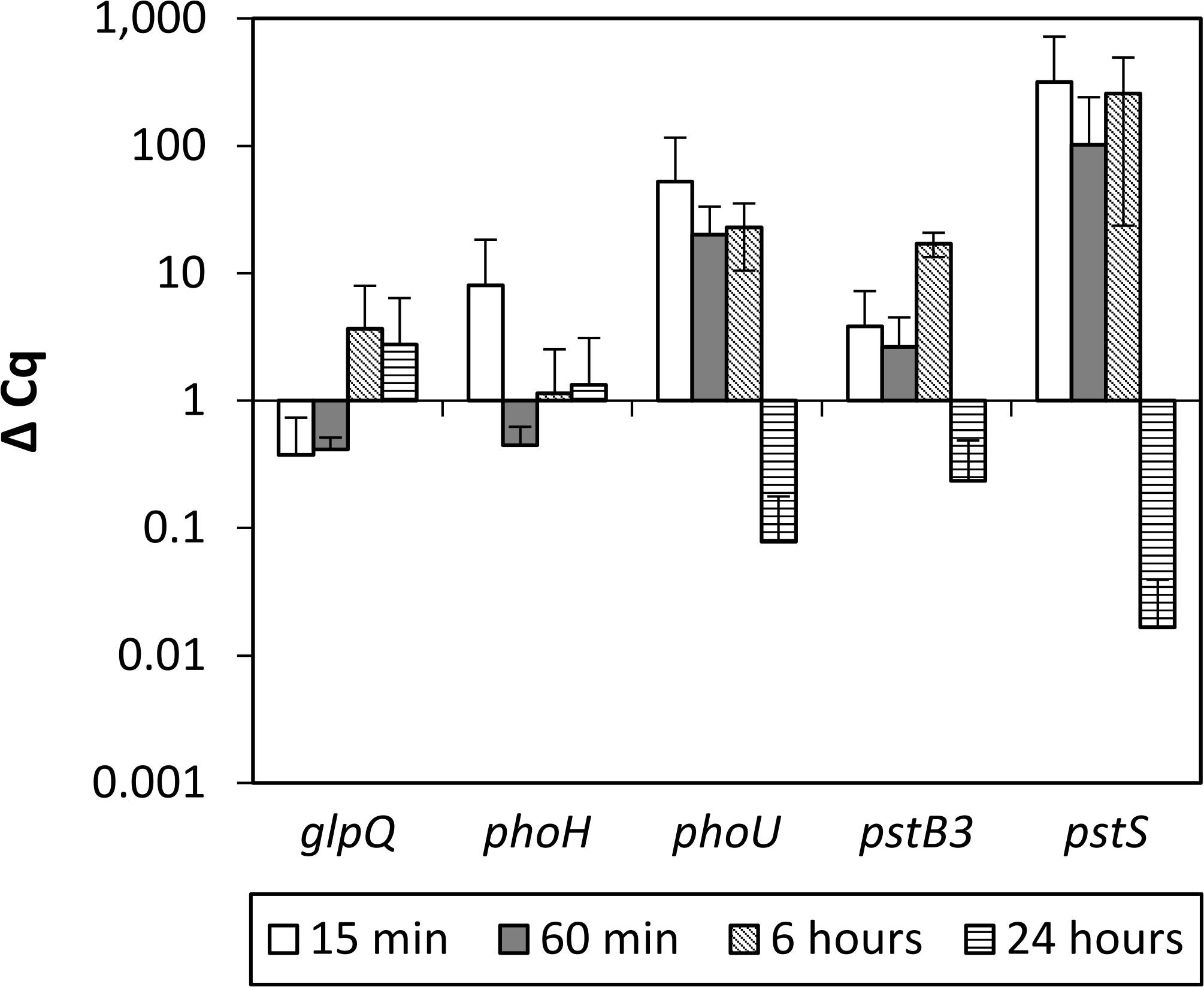
Figure 2. qRT-PCR analysis of P. sonchi SBR5 in low phosphate shock, grown in high phosphate medium (HPi) and transferred to low phosphate medium (LPi). ΔCq values for the indicated genes (glpQ, phoH, phoU, pstB3, and pstS), and the indicated RNA harvesting times after transferring are given as means and standard deviations of biological triplicates.
Growth of P. sonchi SBR5 in Soluble (SPi) and Insoluble (IPi) Containing P Media
Growth of P. sonchi SBR5 in either 5 g L–1 hydroxyapatite (IPi) or 30 mM NaH2PO4 (SPi) was compared with precultures grown with depleted Pi. The maximum growth rate under both conditions was comparable (0.18 h–1, data not shown), but SBR5 grew to a higher biomass concentration (OD600 nm) under the IPi condition (Figure 3A). During RNAseq cultivation, the medium pH dropped, which was more prominent in IPi medium (pH dropped from 7 ± 0 to 4 ± 0) in comparison with SPi medium (from 7 ± 0 to 6 ± 0) (Supplementary Figure 1). SBR5 cultures cultivated in the IPi and SPi conditions represented in Figure 3 presented the same effect in pH (data not shown).
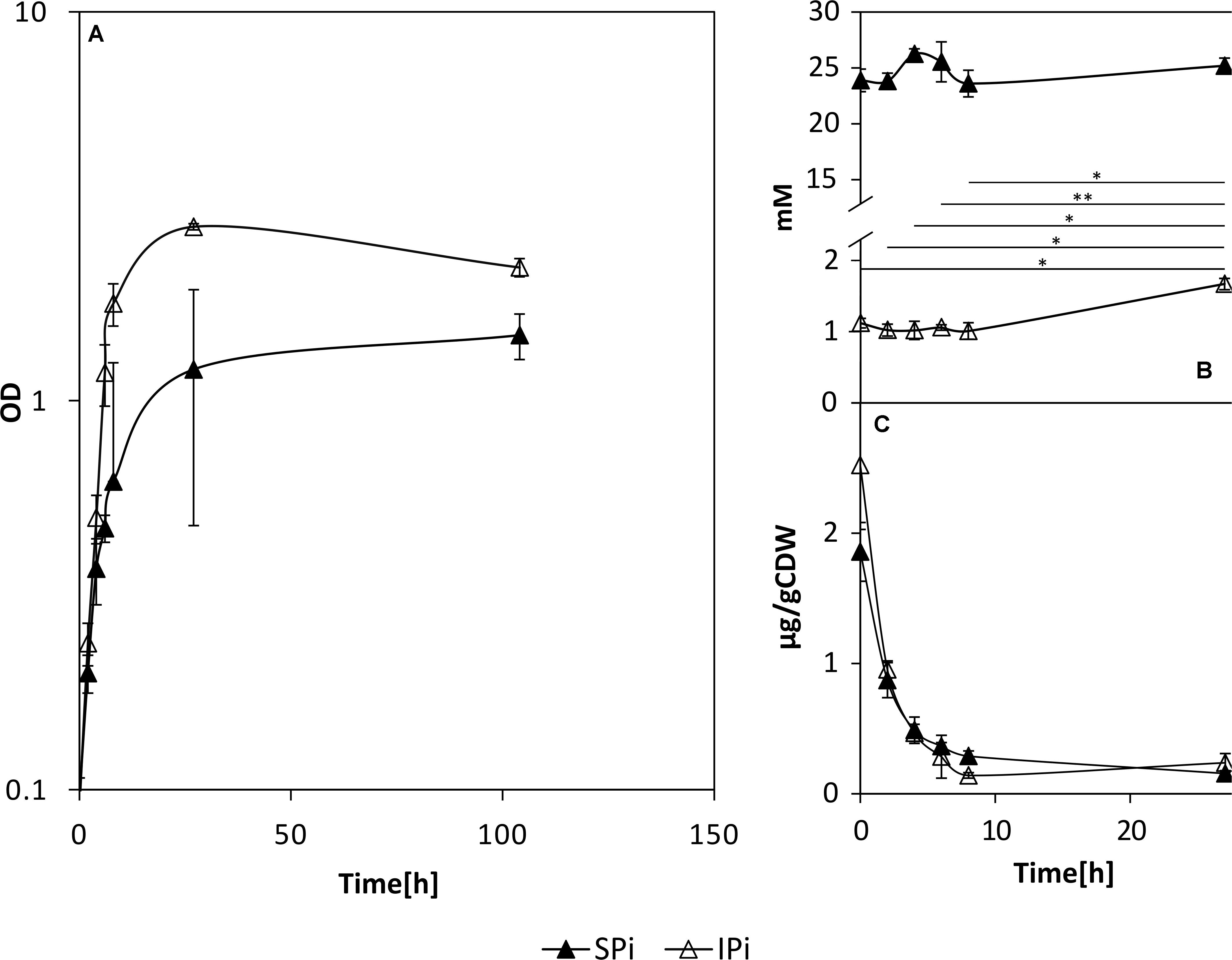
Figure 3. Growth (A) and orthophosphate concentrations in the supernatants (B) and cell lysates (C) of P. sonchi SBR5 cultivated with IPi or SPi. Pikovskaya broth (Pikovskaya, 1948) was used for growth with IPi, and Pikovskaya broth with phosphate source replaced by NaH2PO4 for growth with SPi. Data are given as means and standard deviations of biological triplicates. Student’s paired t-test was performed to examine the significance level and error probability (*p < 0.01; **p < 0.001). CDW, cell dry weight.
The P source in IPi was 5 g L–1 hydroxyapatite, which was not promptly available for the cells in the first hours of growth (Figure 3B). In the first hours of growth, the supernatants of cells cultivated in IPi contained approximately 1 mM of orthophosphates. This is approximately 20 times less than the concentration of Pi present in supernatants of cells cultivated in SPi, in which the Pi concentration was kept relatively constant (∼22 mM) along with the growth of SBR5. Nevertheless, the concentration of orthophosphates in the supernatants increased to approximately 2 mM at 24 h. The difference between the initial and final concentration of Pi in supernatants of IPi condition shows an increase of approximately 1 mM of orthophosphates in IPi medium during the growth of P. sonchi SBR5 (Figure 3B). Cell lysate orthophosphate concentrations under SPi and IPi conditions were drastically depleted in the first hours of growth (Figure 3C).
Differential Gene Expression Analysis and Validation of Expression Pattern by Real-Time Quantitative Reverse Transcription-Polymerase Chain Reaction
Gene expression analysis regarding the P metabolism and PS in bacteria has previously been performed through microarray (Bianco and Defez, 2010) and RNAseq technologies (Zeng et al., 2017). Here, we carried out RNASeq analysis to determine differential gene expression of P. sonchi SBR5 cultivated with a soluble P source (NaH2PO4, SPi medium) and an insoluble P source (hydroxyapatite, IPi medium). Sequencing of cDNA libraries generated from RNA obtained from those cultures resulted in 2,729,614 reads for the SPi sample and 2,773,600 reads for the IPi sample. Of the resultant reads, 2,596,992 and 2,611,860 reads of SPi and IPi libraries, respectively, were mapped onto the genome of SBR5 (Supplementary Table 3). Our DESeq analysis revealed higher expression of 68 genes in IPi (Table 1) and reduced expression of 100 genes in IPi as compared with SPi condition (Table 2), as represented in Supplementary Figure 3.
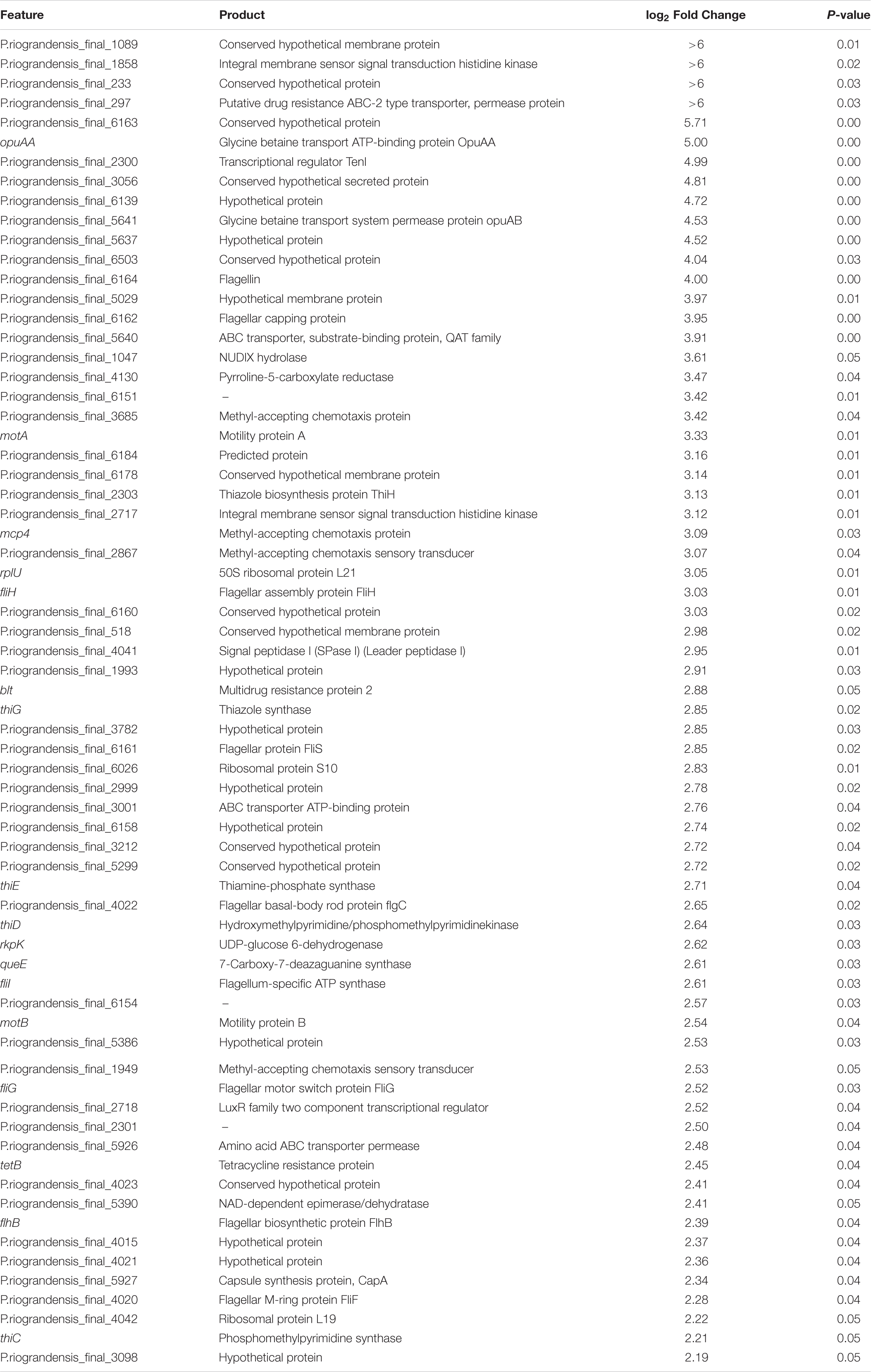
Table 1. List of P. sonchi SBR5 genes showing increased RNA levels during cultivation with IPi in comparison to SPi.

Table 2. List of P. sonchi SBR5 genes showing reduced RNA levels during cultivation with IPi in comparison to SPi.
The results obtained in RNAseq were confirmed by the analysis of gene expression patterns by qRT-PCR for 12 genes selected from the identified differentially expressed genes. The expression patterns of the 12 candidate genes detected by qRT-PCR were in accordance with the gene expression patterns obtained in the RNAseq analysis (Supplementary Figure 4), demonstrating the reliability of our RNAseq analysis.
Differential expression of genes coding for the two pho regulon components PhoP-PhoR was not observed in this work; however, the gene for the phosphate-binding protein PstS was downregulated (Table 2). Interestingly, the expression of two genes related to central carbon metabolism was reduced under IPi condition: the odhAB operon that codes for two subunits of the 2-OGDH (Table 2). Similarly, carbohydrate storage genes, i.e., treA coding for trehalose-6-phosphate hydrolase, treP coding for trehalose phospholyase, and glgA and glgC, coding for glucose-1-phosphate adenylyltransferase and glycogen synthase, respectively, showed reduced expression under IPi condition (Table 2). By contrast, most SBR5 genes involved in thiamine biosynthesis were upregulated in IPi condition: thiH, thiG, thiC, thiD, and thiE (Table 1). Moreover, the increased expression of the gene coding for capsule synthesis protein CapA (Table 1) may indicate a stress response (Candela and Fouet, 2006). The expression of an operon comprising genes coding for flagellar protein FliS (P.riograndensis_final_6161), flagellar capping protein (P.riograndensis_final_6162), a hypothetical protein similar to FlaG protein (P.riograndensis_final_6163), and a flagellin protein (P.riograndensis_final_6164) was upregulated under IPi condition (Table 1). In addition, eight genes related to flagellation and motility were also upregulated: motAB genes coding for motility proteins A and B, flagellar basal body rod protein gene flgC (P.riograndensis_final_4022), flagellar biosynthetic protein gene flhB, and genes fliFGHI coding for flagellar M-ring protein (P.riograndensis_final_4020), flagellar motor switch protein, flagellar assembly protein FliH, and flagellum-specific ATP synthase (Table 1). Furthermore, expression of an operon that includes two genes related to iron and siderophore metabolism (yusV-P.riograndensis_final_1900) was downregulated during cultivation in IPi medium (Table 2). Finally, one gene encoding for pyrroline-5-carboxylase reductase (proC) and an operon comprising genes related to the transport of glycine betaine (opuAA and opuAB-P.riograndensis_final_5641) were upregulated in SBR5 grown in IPi condition (Table 1).
Promoter Activity Assays Using Transcriptional Fusions to Promoterless Genes for Fluorescent Proteins and Flow Cytometry
The promoterless gfpUV reporter gene was fused with promoter sequences of genes upregulated (opuAA) or downregulated (pstS and odhA) under IPi condition. P. sonchi strains P2opuAA-gfpUV, P2pstS-gfpUV, P2odhA-gfpUV, and empty vector control pNW33Nkan were cultivated in SPi or IPi media and analyzed by flow cytometry means. SBR5(pNW33Nkan) strain presented GfpUV background signal in both cultivation conditions (Figure 4A). The GfpUV fluorescence intensities of SBR5(P2pstS-gfpUV) and SBR5(P2odhA-gfpUV) were lower in IPi as compared with SPi condition (Figures 4B,C). In contrast, the GfpUV fluorescence intensity of SBR5(P2opuAA-gfpUV) was higher in IPi as compared with SPi condition (Figure 4D). For further corroboration, a strain was constructed to simultaneously monitor the activity of opuAA gene promoter (fused to promoterless red-fluorescence reporter gene crimson) and pstS promoter (fused to promoterless gfpUV). Flow cytometry revealed opuAA promoter activity in cells cultivated in IPi, whereas pstS promoter activity was not detected (Figure 4E). By contrast, when cultivated in SPi, promoter activity was detected for pstS but not for opuAA (Figure 4F). Thus, reporter gene expression analysis confirmed that opuAA is induced under IPi condition, whereas pstS and odhA gene expression is lower under IPi condition compared with SPi condition.
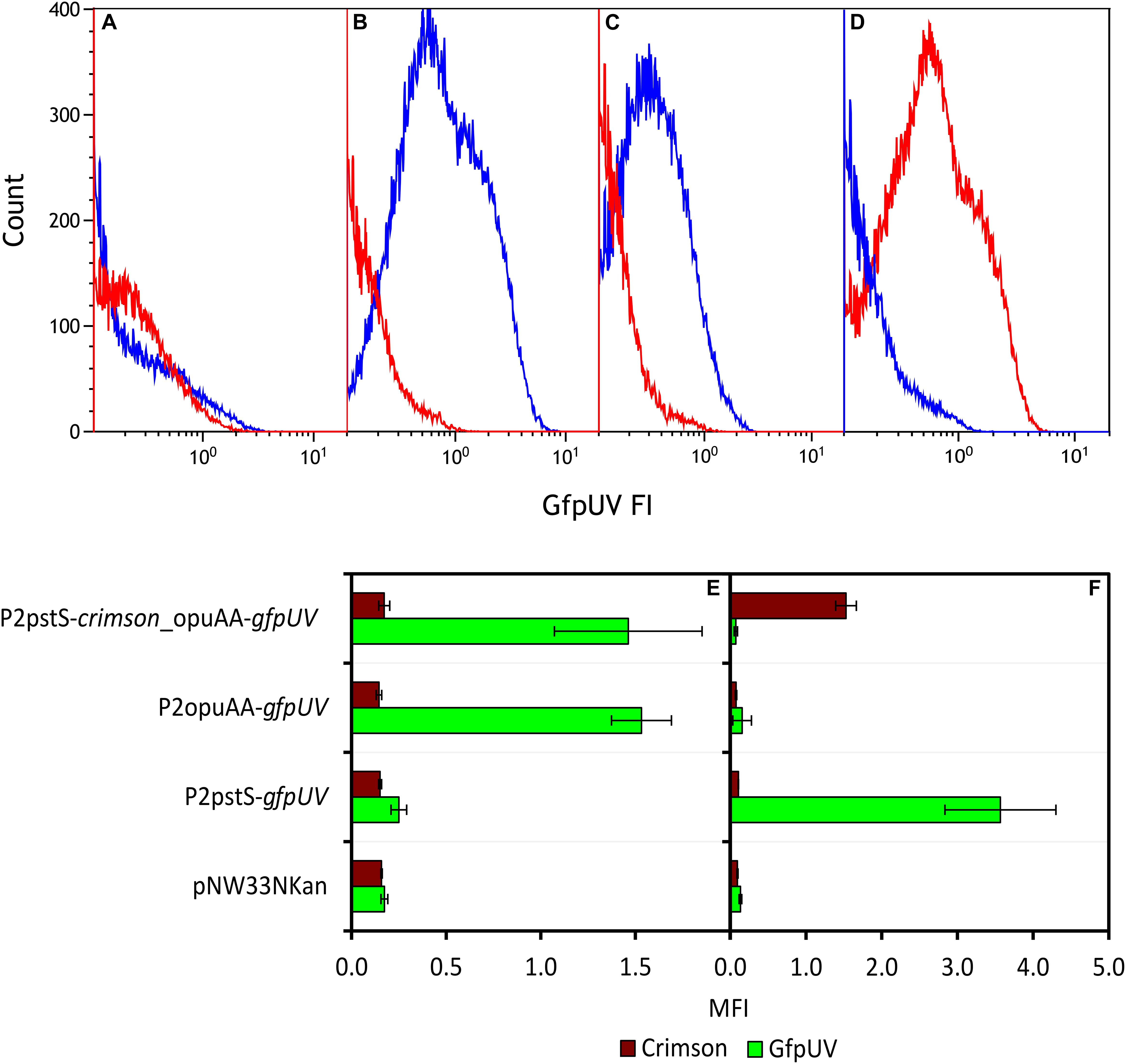
Figure 4. Expression analysis of various promoters in P. sonchi SBR5 grown in IPi or SPi. (A–D) Histograms show flow cytometry results of GfpUV fluorescence intensity (GfpUV FI) of SBR5 carrying different vectors (A: pNW33N; B: P2pstS-gfpUV; C: P2odhA-gfpUV; D: P2opuAA-gfpUV) cultivated in IPi (red) and SPi (blue). (E,F) Crimson and GfpUV mean fluorescence intensities (MFI) of SBR5 carrying vectors pNW33N, P2pstS-gfpUV, P2odhA-gfpUV, P2opuAA-gfpUV, and P2pstS-crimson_opuAA-gfpUV cultivated in IPi (E) or SPi (F). MFIs are given as means and standard deviations of triplicate cultivations measured by flow cytometry of 20,000 cells.
High-Performance Liquid Chromatography Analyses: Carbon Metabolism and Osmoprotection
Because the excretion of organic acids by PSB is considered a crucial factor in PS (Rashid et al., 2004), the production of organic acids by SBR5 was monitored under IPi and SPi conditions. A different composition of organic acids was detected in supernatants of SBR5 when this organism was cultivated in IPi or SPi conditions (Figure 5A). The organic acids succinate, oxoglutarate, and citrate, which are components of the tricarboxylic acid (TCA) cycle, were present in the supernatants of SBR5 in SPi condition (Figure 5A). However, in the IPi condition, the concentration of succinate and oxoglutarate decreased 100 and 81%, respectively, in comparison with SPi condition, whereas the citrate concentration was similar to that in the SPi condition. Besides, the amount of acetate and gluconate was 85 and 48% higher, respectively, when SBR5 was cultivated in IPi as compared with SPi (Figure 5A). Arginine and ornithine could not be detected (Figure 5B). Moreover, the 2-OGDH-encoding genes odhAB were downregulated when SBR5 was cultivated in IPi condition (Table 2). This result was confirmed by enzymatic assay of 2-OGDH in crude extracts of SBR5, in which 2-OGDH was inactive in crude extracts of cells grown in IPi, whereas activity was observed in SPi condition (Figure 5C).
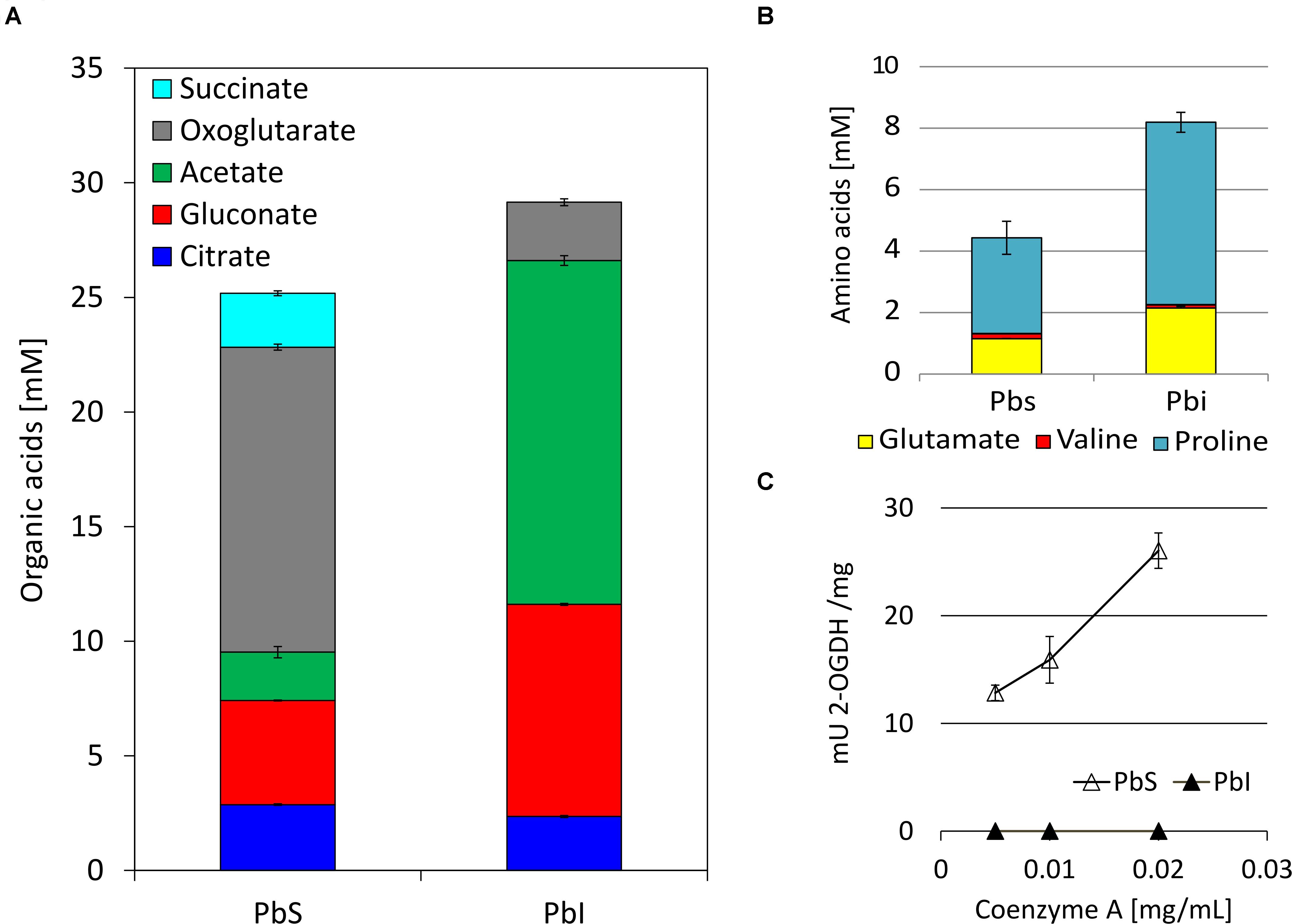
Figure 5. Accumulation of organic acids (A) and amino acids (B) in the culture medium and 2-oxoglutarate dehydrogenase (2-OGDH)-specific activities (mU 2-OGDH/mg) in crude extracts (C) of P. sonchi SBR5 cultivated with IPi medium in comparison with SPi. Concentrations of organic acids and amino acids were determined by HPLC analysis of the supernatants collected after 20 h of growth. Arginine and ornithine could not be detected. Activity of 2-OGDH was measured with different concentrations of CoA (milligrams per milliliter). Data are given as means and standard deviations of biological triplicates.
Furthermore, the levels of the compatible solutes trehalose and glycine betaine in supernatants and cell lysates of SBR5 cultivated in SPi or IPi were determined. This was done because, although the levels of osmolality obtained in the supernatants of SPi and IPi cultures were similar (Figure 6A), the internal osmolality of cells was threefold higher than that in SPi cultivation broth (Figure 6B). Approximately 0.01 mM of trehalose was present in SBR5 supernatants at 0 h under both Pi conditions, indicating that this component was present in the media composition (Figure 6C). However, trehalose concentration in the supernatant increased during growth, peaking at 4.9 and 7.8 mM in SPi and IPi, respectively (Figure 6C). SBR5 cells cultivated in IPi medium accumulated trehalose intracellularly from 2 to 8 h of growth, peaking at 0.1 mg g CDW–1 at 4 h (Figure 6D). No glycine betaine was detected in the supernatant of SPi cultivation broth, but 24.3 mM of glycine betaine production was observed at 2 h of cultivation in IPi broth (Figure 6E). The glycine betaine concentration in SBR5 supernatants from IPi cultivation dropped completely at 24 h, as this component accumulated intracellularly (Figures 6E,F). The presence of glycine betaine in SBR5 cell lysates was observed from 4 h of growth, being threefold higher at 8 h of growth when those cells were cultivated in IPi in comparison with SPi medium (Figure 6F). Glycine betaine and trehalose were produced and/or accumulated in IPi cultivation broth.
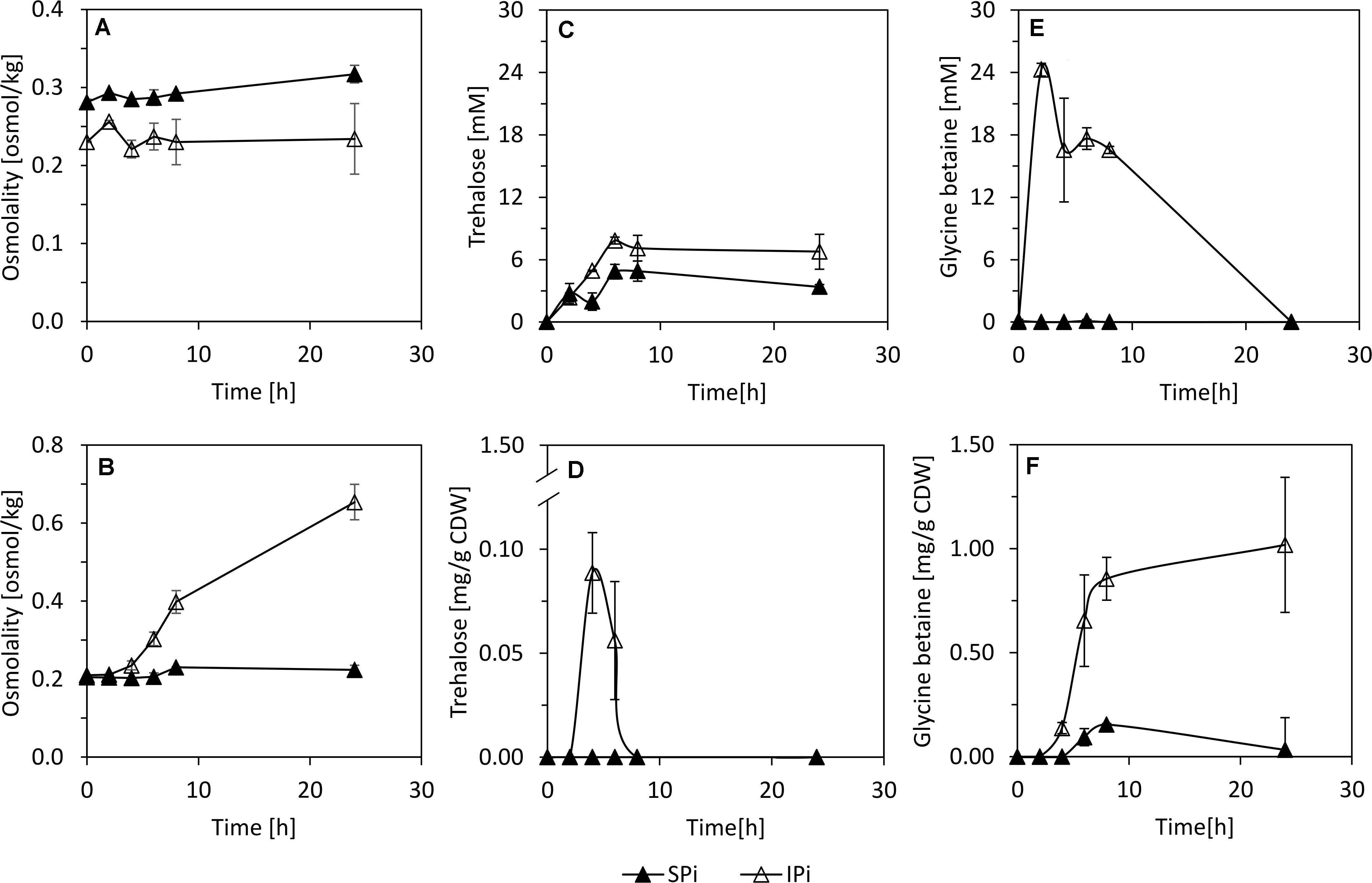
Figure 6. Supernatant (A,C,E) and intracellular (B,D,F) osmolarity, trehalose, and glycine betaine concentrations during growth of P. sonchi SBR5 with IPi and Spi. Pikovskaya broth (Pikovskaya, 1948) was used for growth with IPi, and Pikovskaya broth with phosphate source replaced by NaH2PO4 for growth with SPi. Data are given as means and standard deviations of biological triplicates. CDW, cell dry weight.
Glutamate and proline were secreted by SBR5 cells cultivated in IPi (Figures 5B, 7A). The concentration of proline in SBR5 supernatants at 24 h of bacterial growth was fivefold higher in IPi in comparison with SPi (Figures 7A,B). Moreover, the internal proline concentration reached ∼240 mg gCDW–1, which is twofold higher than that in SPi (Figures 7C,D). Although ∼9-mM oxoglutarate formation was observed in SPi supernatant (Figure 7B), no oxoglutarate was detected in the supernatant of IPi broth (Figure 7A).
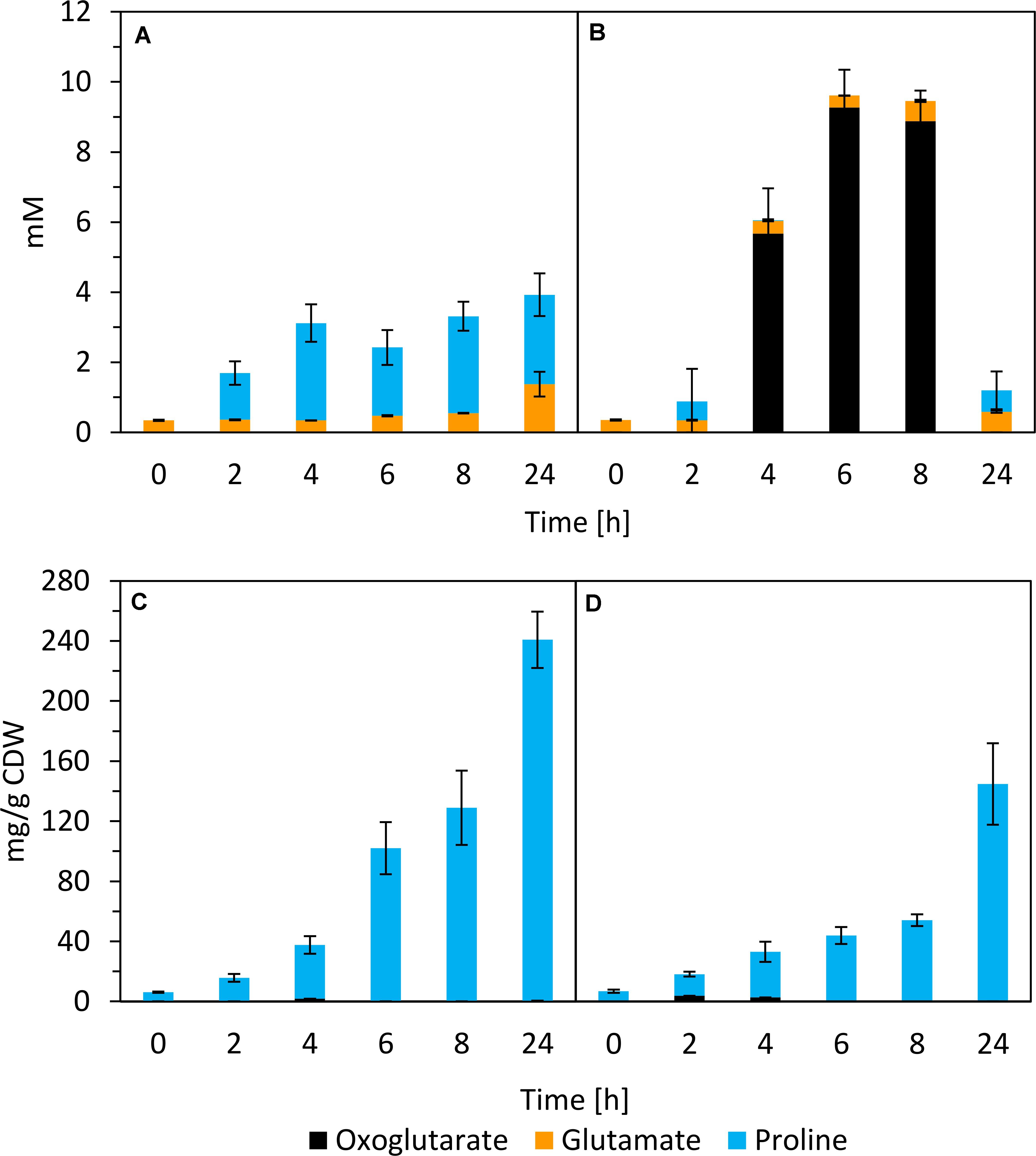
Figure 7. Quantification of oxoglutarate, glutamate, and proline in supernatants (A,B) and crude extracts (C,D) of P. sonchi SBR5 along with its growth in IPi (right) or SPi (left). Pikovskaya broth (Pikovskaya, 1948) was used for growth with IPi, and Pikovskaya broth with phosphate source replaced by NaH2PO4 for growth with SPi. Data are given as means and standard deviations of biological triplicates.
Thiamine Biosynthesis Confirmation by Flow Cytometry
Our RNAseq analysis revealed the upregulation of many thiamine biosynthesis-related genes in SBR5 cultivated in IPi condition (Table 1). Therefore, we have performed a thiamine biosynthesis detection using a thiamine pyrophosphate (TPP)-based biosensor. The strain SBR5(P2pyk_RthiC-gfpUV) carries the constitutive promoter of the pyruvate kinase-encoding gene pyk of SBR5 with its native 5′ untranslated region (5′UTR) replaced by the 5′UTR of the P. riograndensis_final_150, thiC gene coding for phosphomethylpyrimidine synthase (Brito et al., 2017a). The 5′UTR of thiC gene contains a TPP riboswitch previously characterized, which “switches off” the expression of gfpUV reporter gene in the presence of thiamine (Brito et al., 2017a). The strain SBR5 carrying the plasmid P2pyk-gfpUV (Brito et al., 2017b), with original pyk 5′UTR and the empty vector pNW33N were used as control. All recombinant strains were cultivated in modified IPi (IPim) or SPi (SPim), both media containing 0.1 mg/L of biotin but lacking yeast extract. Cells were harvested after 6 h, and GfpUV mean fluorescence intensity (MFI) was measured by flow cytometry. As expected, the MFI of empty vector control strain presented GfpUV background signal (Figure 8) in both media. Recombinant SBR5 with native pyk promoter driving the expression of gfpUV showed elevated GfpUV MFI in both media in comparison with the empty vector control. However, in the presence of thiC TPP riboswitch, cells cultivated in IPim “switched off” gfpUV expression, being GfpUV MFI levels similar to the empty vector control (Figure 8), indicating thiamine biosynthesis in this condition.
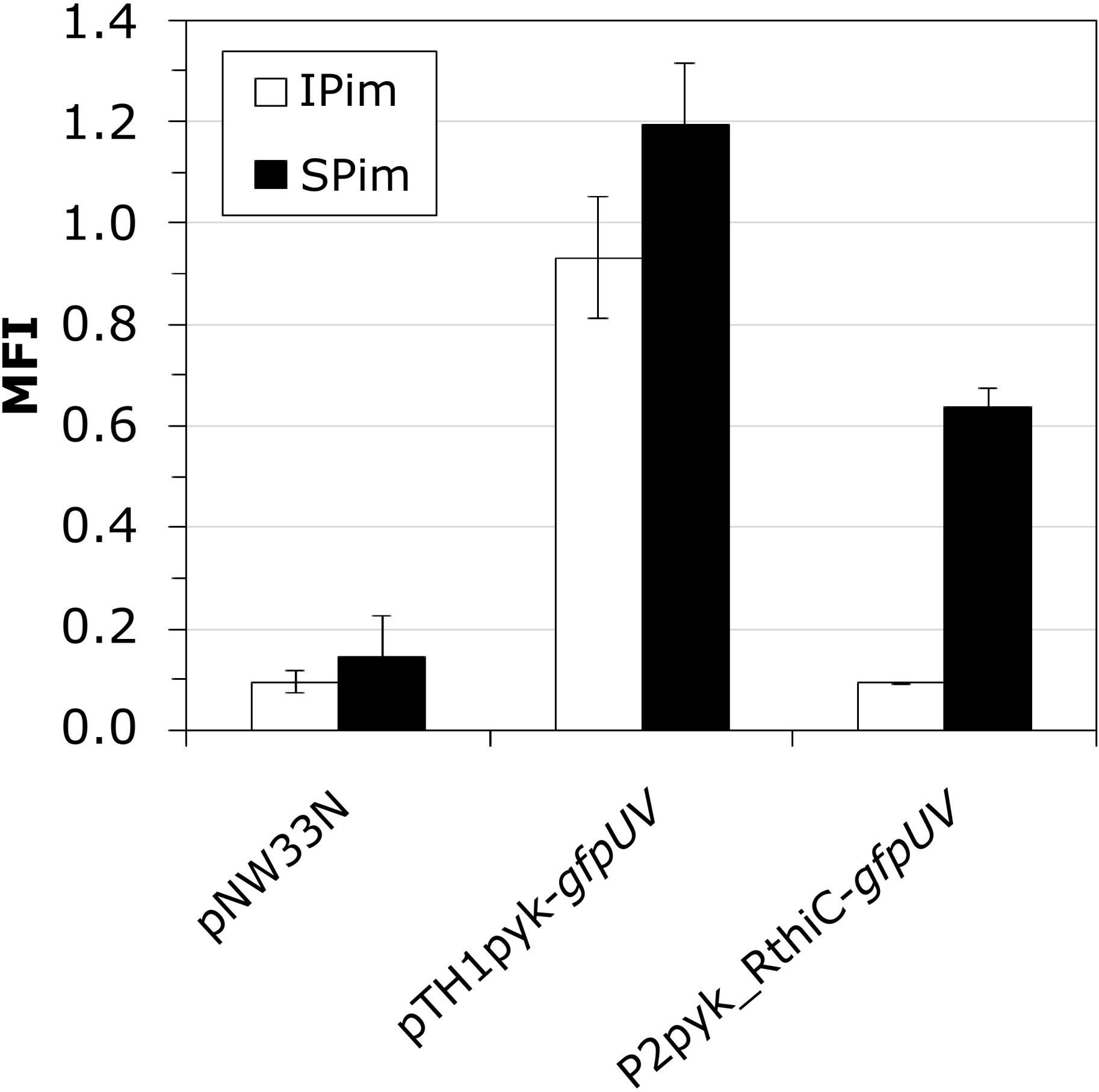
Figure 8. Monitoring intracellular thiamin concentrations by a thiamin-responsive riboswitch fused to GfpUV. P. sonchi SBR5 cells carried plasmids either with gfpUV gene under control of pyk promoter and 5′UTR of the endogenous thiC gene (pTH1pyk-gfpUV), pyk promoter carrying its native 5′UTR (p2pyk_RthiC-gfpUV), or the empty vector plasmid pNW33N. P. sonchi cells were cultivated in modified IPi or with SPi. MFI (mean fluorescence intensities) are given as means and standard deviations of triplicate cultivations measured by flow cytometry of 20,000 cells. Modified IPi and SPi media were prepared without yeast extract but with addition of biotin (100 μg/ml).
Discussion
Here, the activity of P. sonchi genomovar Riograndensis SBR5 under IPi condition was characterized. Functional analyses were performed based on the currently available genomic and genome-wide transcriptomic databases (Brito et al., 2015, 2017a) and on the gene expression analysis performed in the present study. We aimed to relate some genes of P. sonchi SBR5, which were differentially expressed with two distinct P sources (SPi and IPi) to the physiological analysis of PS processes. Our findings revealed that IPi condition (with hydroxyapatite as P source) changed carbon metabolism and vitamin biosynthesis of SBR5 and expressed flagellation genes and activated osmoprotection.
It is a general phenotype of PSB that the PS activity is induced by low levels of exogenous soluble phosphate and inhibited by its high levels (Zeng et al., 2017). The phosphate in SPi condition was not completely utilized (Figure 3B) but supported the growth of P. sonchi SBR5 (Figure 3A), indicating that SPi provided a sufficient supply of P for SBR5. However, soluble phosphate was not promptly available for SBR5 at the beginning of growth in IPi medium, containing hydroxyapatite (Figure 3B), lacking the necessary Pi supply for the cells. Altogether, that could explain the increase of ∼1 mM orthophosphate concentration after 24 h in the IPi medium (Figure 3B). This value is similar to values observed for other Paenibacillus species: ∼1 mM by P. polymyxa and ∼1.2 mM by P. mucilaginosus from CaHPO4 and phosphorite after 3 and 5 days of incubation, respectively (Hu et al., 2006; Wang et al., 2012). However, the increase in orthophosphate concentration observed in IPi condition represents approximately 3.4 % of hydroxyapatite solubilization rate, which is dramatically inferior to the solubilization rates (up to 100%) reported in fungi (Mendes et al., 2013). It is known that the PS activity of PSB can be repressed by soluble phosphate by a feedback inhibition (Zeng et al., 2017), which is a severe limitation to the extensive application of PSB. However, the molecular mechanism of soluble phosphate regulation on PS activity of PSB remains to be elucidated.
In the past decades, enzymatic processes have been characterized to be responsible for the solubilization of organic phosphate substrates in bacteria. PS promoted by acid phosphatase was observed in Pseudomonas sp. (Jorquera et al., 2008), Burkholderia cepacia (Rodriguez et al., 2000), Enterobacter aerogenes, Enterobacter cloacae, Citrobacter freundi, Proteus mirabalis, and Serratia marcenscens (Thaller et al., 1995). Moreover, organic Pi solubilization through phytase activity was observed in Bacillus subtilis, Pseudomonas putida, and Pseudomonas mendocina (Richardson and Hadobas, 1997), and phosphotase activity was detected in Klebsiella aerogenes (Ohtake et al., 1996) and P. fluorescens (Lidbury et al., 2016). However, although P. sonchi SBR5 possesses some of those PS-related enzymes, upregulation of their respective genes was not observed in the differential gene expression analysis under PS conditions (Tables 1, 2). This could be because the phosphate sources utilized in SPi (NaH2PO4) and IPi (hydroxyapatite) conditions were not from an organic origin. However, an inspection of the complete genome sequence of SBR5 (Brito et al., 2015) allowed the detection of some candidate genes coding for enzymes that promote organic PS and enzymes related to P metabolism, e.g., an alkaline phosphatase (P.riograndensis_final_731) and a phytase (P.riograndensis_final_2549). The Pst phosphate-specific transport system is a major phosphate transport system characterized in B. subtilis (Qi et al., 1997). The pst operon of B. subtilis is composed of pstS, pstC, pstA, pstB1, and pstB2 genes. PstS is a binding protein, PstC and PstA are two integral inner membrane proteins, and PstB1 and PstB2 are ATP-binding proteins (Qi et al., 1997; Atalla and Schumann, 2003). Those genes were found in the genome of P. sonchi SBR5 (P.riograndensis_final_3645-3648) and were expressed at low levels in our previous RNAseq study, in which no phosphate depletion treatment was applied (Brito et al., 2017a). Here, the gene pstS was downregulated in IPi condition (Table 2) as confirmed by gfpUV expression analysis, in which pstS promoter-driven expression reduced GfpUV fluorescence in comparison with SPi condition (Figure 4). Our orthophosphate quantification of IPi supernatants revealed P concentrations ∼10 times smaller than the Monod constant depicted in Supplementary Figure 2. This indicates that P is not promptly available for SBR5 cells in the first hours of growth (Figure 3B). Although pst P transport genes are stimulated upon P starvation, the underlying regulatory mechanism remains to be studied. Moreover, cultivation of SBR5 in low and high phosphate conditions (LPi and HPi media), together with phosphate quantification and qRT-PCR, revealed that phosphate starvation response of SBR5 could be mediated by a regulatory system. Some bacteria present a global regulatory mechanism of Pi management in a situation of Pi depletion that involves a two-component regulatory system and is named pho regulon (Santos-Beneit, 2015). The pho regulon is controlled by the two-component PhoP-PhoR signal transduction system (Shi and Hulett, 1999). Homologous genes are present in the genome sequence of P. sonchi SBR5 (P.riograndensis_final_2307-2308). Here, qRT-PCR showed that genes involved in the regulation of the pho regulon genes phoU and phoH (Santos-Beneit, 2015) were highly expressed in LPi condition (Figure 2). Phosphate depletion led to the expression of σB-mediated general stress response genes in B. subtilis (Prágai and Harwood, 2002). B. subtilis responds to phosphate starvation stress by regulating genes encoding the phosphate starvation Pho proteins (Prágai et al., 2004). Genes classically associated with their regulation were targeted by qRT-PCR in the present study, being highly expressed by SBR5 cultivated in LPi condition. Among them are pstS and pstB3 and the glycerophosphodiester phosphodiesterase gene glpQ (Figure 2), in which the product hydrolyzes deacylated phospholipids to alcohol and glycerol-3-phosphate, which is subsequently utilized in the cell (Jorge et al., 2017a). Finally, the fact that the concentration of orthophosphate in LPi medium dropped approximately 30% at 24 h indicates consumption of the low P available in Pi depletion condition (Figure 1B), which is in accordance with the high expression of high-affinity Pi uptake system pst genes (Figure 2).
Under IPi condition, several genes involved in thiamine biosynthesis were upregulated (Figure 9). Thiamine is indispensable for the activity of the carbohydrate and branched-chain amino acid metabolic enzymes (Du et al., 2011). TPP is the active form of this vitamin and functions as a cofactor of several important enzymes in carbohydrate and amino acid metabolism (Melnick et al., 2004). Biosynthesis of thiamine has been studied in several groups of PGPR, including Azotobacter, Pseudomonas, and Azospirillum (Palacios et al., 2014). As biotin, thiamine is part of the vitamin B group, which is suggested to improve the plant root colonization by PGPR (Palacios et al., 2014). Thiamine also acts as a cofactor of indolepyruvate decarboxylase, a synthesizing enzyme of principal plant hormone in PGPR indole-3-acetic acid (Schütz et al., 2003). In E. coli, thiamine precursors hydroxymethyl-pyrimidine diphosphate and hydroxyethyl-thiazole phosphate are utilized to synthetize TPP (Begley et al., 1999). Thiamine biosynthesis protein ThiC converts aminoimidazole ribotide to hydroxymethyl-pyrimidine phosphate, which is subsequently phosphorylated by the bifunctional hydroxymethyl-pyrimidine phosphate P kinase ThiD to yield hydroxymethyl-pyrimidine pyrophosphate. On the other hand, the thiazole moiety of thiamine is derived from tyrosine, cysteine, and 1-deoxy-D-xylulose-5-phosphate. In a yet unresolved chain of reactions featuring thiF, thiS, thiG, thiH, and thiI gene products, hydroxyethyl-thiazole phosphate is formed. Hydroxymethyl-pyrimidine pyrophosphate and hydroxyethyl-thiazole phosphate are joined by one enzymatic step mediated by the ThiE protein (Figure 9) (Lünse et al., 2014). In the present study, among the mentioned thiamine biosynthesis genes, only thiF, thiS, and thiI were not detected in RNAseq analysis (Table 1). Moreover, although the production of thiamine by SBR5 was not measured, a TPP riboswitch was used to monitor intracellular TPP. As shown by Brito et al. (2017a), the 5′UTR of the phosphomethylpyrimidine synthase-encoding gene (P.riograndensis_final_150) contains the sequence of a TPP riboswitch that “switches off” gene expression in the presence of 5 μM thiamine. Here, gfpUV reporter gene expression was “switched off” in IPim condition, indicating thiamine biosynthesis by P. sonchi (Figure 8). These findings suggest that thiamine might be important for SBR5 to cope with low phosphate concentrations during growth in IPi condition.
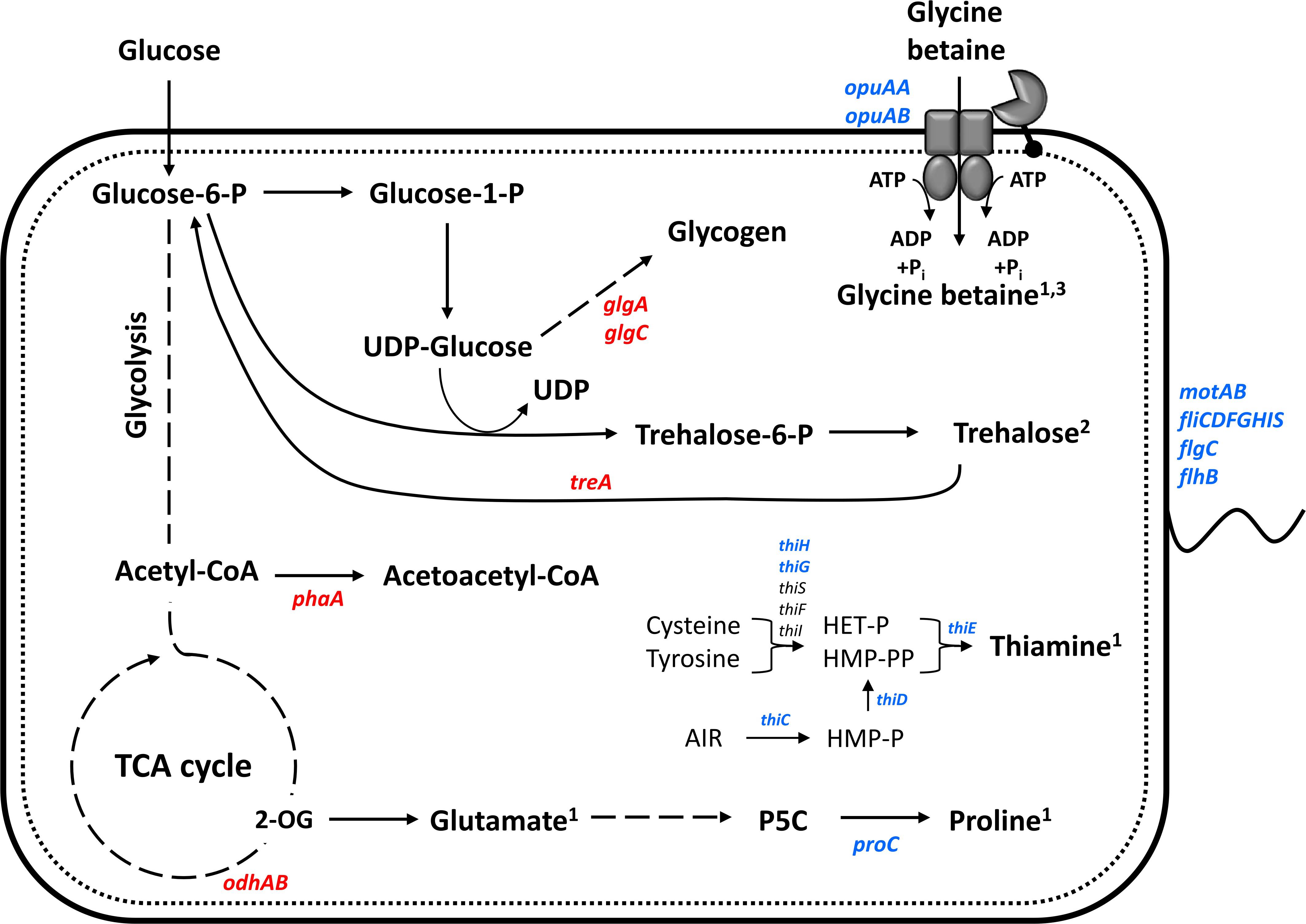
Figure 9. Schematic representation of physiological and gene expression differences between P. riograndensis SBR5 grown with IPi and SPi. Genes showing higher RNA levels during growth with IPi as compared with SPi are depicted in blue, whereas those with reduced RNA levels in red. RNAseq together with the physiological analyses of the present study indicates that proline is produced and glycine betaine produced and accumulated, and trehalose is accumulated inside the cell for osmoprotection. Thiamine biosynthesis is activated in insoluble phosphate condition. Continuous lines: one-step processes; dashed lines: processes with more than one-step. 1Produced component; 2putatively accumulated component; 3imported component.
SBR5 acidifies the IPi medium, whereas it maintains the medium pH near neutral under SPi condition (Supplementary Figure 1), which was expected because acidification typically is the main PS mechanism of PSB (Chen et al., 2006; Marra et al., 2015). The decrease of pH as a mechanism to perform PS has been reported in fungi (Whitelaw, 2000), and it is mostly related to the production of organic acids, as in Aspergillus and Penicillium (Li et al., 2016). Organic acid release is classically related to PS in bacteria (Illmer and Schinner, 1992). Production of oxalate and malate, which were previously related to PS in Pseudomonas (Vyas and Gulati, 2009), was not observed in SBR5. In the PGPR Burkholderia multivorans, the expression of genes related to carbon metabolism, including genes encoding for sugar ABC transporters and 2-OGDH, was upregulated when P was depleted in the medium (Zeng et al., 2017). The switch of carbon metabolism pathways is closely related to the depletion of available soluble phosphate (Thomas et al., 2012). Here, downregulation of 2-OGDH genes odhA and odhB was observed during the growth of SBR5 in IPi condition (Table 2), which was confirmed by reporter gene fusion analysis (Figure 4C). 2-OGDH is a key enzyme that catalyzes the step in TCA cycle in which oxoglutarate is converted to succinyl-CoA (Nguyen et al., 2015). Thereby, it is suggested that the metabolic flux toward the TCA cycle is reduced when SBR5 is cultivated in a medium with IPi, which leads to a general decrease of accumulation of TCA metabolites, e.g., oxoglutarate and succinate (Figures 5A, 7A). Low production of oxoglutarate in contrast to high production of gluconate was also observed in soil bacterial isolates that perform PS (Mardad et al., 2013). Succinate is a product of a step in the TCA cycle that follows the reaction catalyzed by 2-OGDH, and it is also one of the major organic acids present in root exudates of plants in the rhizosphere. Repression of glucose utilization by succinate is termed as succinate-mediated catabolite repression (Bringhurst and Gage, 2002). Divya et al. (2011) showed that succinate and malate individually and as mixtures repressed gluconate production and PS in Pseudomonas aeruginosa. It is also a component that represses PS phenotype in Klebsiella pneumoniae (Rajput et al., 2013, 2015). Based on that and our findings, the reduction of the metabolic flux toward the TCA cycle might be one strategy utilized by SBR5 to perform PS.
In RNAseq cultivation, the total concentration of analyzed organic acids at the end of SBR5 growth in IPi condition was 15% higher than in SPi (Figure 5A). This is in accordance with our observation regarding the acidification of the growth medium in the IPi condition (Supplementary Figure 1). Gluconate and acetate, known as active agents in PS, were highly produced in IPi condition (Figure 5A). Gluconate is produced by the majority of the PSB, being often the most quantitatively produced organic acid for PS ends (Yi et al., 2008; Vyas and Gulati, 2009; Shahid and Hameed, 2012; Marra et al., 2015). The production of acetate by PSB isolates to perform PS has also been reported (Mardad et al., 2013). The gene encoding the enzyme glucose dehydrogenase is present in the genome of P. sonchi SBR5 (P.riograndensis_final_6601). Glucose dehydrogenase oxidizes glucose to convert it to gluconate (Werra et al., 2009). There is a high conservation of this gene in Paenibacillus species (Xie et al., 2016). Although gluconate is accumulated in IPi condition, P.riograndensis_final_6601 was slightly transcribed in landscape transcriptome of SBR5 (Brito et al., 2017a) and not detected in our RNAseq analysis. Phosphotransacetylase and acetate kinase are enzymes involved in the reversible interconversion of acetyl-CoA to acetate (Wendisch et al., 1997). The genes P.riograndensis_final_3937 and P.riograndensis_final_4369 (coding for phosphotransacetylase and acetate kinase, respectively) were not differentially expressed, but their transcript abundances were considered intermediate to high in landscape transcriptome analysis (Brito et al., 2017a). In Sinorhizobium meliloti, the genes encoding phosphotransacetylase and acetate kinase are induced by phosphate deficiency and controlled by the pho regulon (Summers et al., 1999).
The expression of flagellin protein-encoding gene (P.riograndensis_final_6164), FliS (P.riograndensis_final_6161), flagellar capping protein (P.riograndensis_final_6162), hypothetical protein similar to FlaG (P.riograndensis_final_6163), motAB genes, FlgC-encoding gene (P.riograndensis_final_4022), flagellar biosynthetic protein FlhB-encoding gene, and genes fliFGHI were upregulated under IPi condition (Table 1). Flagella biosynthesis is energy-intensive, and flagella genes are typically expressed from strong promoters. Besides, flagellin proteins are translated from near-consensus RBS (Guttenplan and Kearns, 2013), which is also true for SBR5, where the RBS sequence 5′GGGGAGG assigned to the flagellin gene P.riograndensis_final_6164 is similar to the consensus RBS sequence 5′aGGaGg (Brito et al., 2017a). However, no difference in swimming of SBR5 was detected between SPi and IPi soft agar media (0.2% agar, data not shown). Nevertheless, the acidification of media causes the cells to change the flagella morphology immediately in E. coli. When the bacteria were grown at pH 6 or 8, shorter and thinner forms of flagella were produced (Chang et al., 2013). Hence, this indicates that the upregulation of several flagellation-related genes in IPi condition might be due to the change of pH that occurs in this condition.
Another effect observed in our work was the increase in cell osmolality caused by the IPi medium, which led to the change of compatible solutes dynamics in SBR5 cells to induce osmoadaptation (Figure 6). The osmolality of the rhizosphere is likely to exceed that of bulk soil water as a result of the exclusion of salts such as NaCl from root tissue, the exudation of low molecular weight organic acids, and the secretion of mucilage by roots and bacteria (Miller and Wood, 1996). As the activities of roots and rhizobacteria modify the osmolality of the rhizosphere, osmoadaptive mechanisms will likely influence both the survival of rhizosphere bacteria and their interactions with plant roots. The study of osmoadaptation by rhizosphere bacteria has been limited to a small number of genera. In most cases, these bacteria have been examined because of interest in their interactions with plants rather than their degree of salt tolerance (Miller and Wood, 1996). Soil variables that influence the osmotic properties of the rhizosphere include its clay content and texture, its water content, and the nature and concentration of solutes in the soil water (Gregory, 1988). The osmolality of typical soil water has been estimated as less than 50 mOsm kg–1 (Gregory, 1988). Here, the cultivation of SBR5 in IPi condition led to an osmolality as high as 600 mOsm kg–1 (Figure 6B). This could be due to the increase of organic acid excretion in this condition, which was 15% higher than in SPi (Figure 5A). The increase in osmolality caused SBR5 cells to use osmoregulatory mechanisms such as production, uptake, and accumulation of compatible solutes (Figure 9). Proline and glycine betaine are among the principal compatible solutes accumulated as an osmotic response in bacteria (Sleator and Hill, 2002). The accumulation of the osmoprotectant glycine betaine from exogenous sources provides a high degree of osmotic tolerance to B. subtilis (Kappes et al., 1996). Proline was secreted by SBR5 to higher concentrations under IPi as compared with SPi condition (Figures 5B, 7A). Accordingly, there was upregulation of the gene P.riograndensis_final_4130 (proC) encoding pyrroline-5-carboxylate reductase (Table 1), which is the enzyme catalyzing the last step in the proline production pathway in Corynebacterium glutamicum (Jensen et al., 2015). Another evidence is the downregulation of the 2-OGDH complex genes odhAB, which could have caused a change in the metabolic flux from the TCA cycle intermediate 2-oxoglutarate to proline (Figure 9). The metabolic pull of oxoglutarate toward glutamate and proline biosynthesis may explain the reduced oxoglutarate concentration and the increased glutamate and proline concentrations in supernatants of IPi cultures in comparison with SPi (Figures 7A,B). Three lines of evidence showed intracellular accumulation of glycine betaine under IPi condition: opuAA and opuAB RNA levels were increased (Table 1), GfpUV reporter gene expression driven by opuAA promoter increased (Figures 4D,E), as did the intracellular glycine betaine concentration (Figure 6F). Lastly, trehalose was accumulated in SBR5 cells in IPi condition (Figure 6D). Accordingly, the reduced expression of treA, glgA, and glgC genes was observed. Trehalose is degraded by trehalase TreA, whereas GlgC and GlgA synthesize glycogen, a pathway competing for UDP-glucose with trehalose synthesis (Purvis et al., 2005). Thus, the changed medium and inner cell osmolality under IPi condition were counteracted by the secretion of proline and glutamate and the accumulation of glycine betaine and trehalose.
Taken together, our differential gene expression study revealed an extraordinarily complex transcriptional response of P. sonchi SBR5 to two distinct P sources, soluble and insoluble. On the one hand, P. sonchi SBR5 changes its carbon metabolism characteristics in the presence of insoluble phosphate by the reduction of the metabolic flux toward the TCA cycle. On the other hand, a drastic change in cell osmolality was detected, triggering osmoprotection mechanisms (Figure 9). We showed that the production of organic acids might be the most important strategy utilized by SBR5 to perform PS. The findings of our study will help us understand the mechanisms of insoluble phosphate-derived gene regulation and physiological activities by P. sonchi SBR5, providing the first step in the elucidation of the PS process in this organism, which could further improve the scope of its application as a crop inoculant.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/, GSE154303.
Author Contributions
LB, MGL, and LS carried out the experimental procedures. LB analyzed the data of the present study. LB prepared a draft of the manuscript. LB, LP, and VW finalized the manuscript. VW coordinated the study. All the authors read and approved the manuscript.
Funding
LB acknowledges support as fellow of the Ciência sem Fronteiras program of Brazil (Science Without Borders program). We acknowledge the financial support of the German Research Foundation (DFG) and the Open Access Publication Fund of Bielefeld University for the article processing charge.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The content of this manuscript has been published in part as part of the thesis of LB, 2017. We thank Aaron Weihrauch for help with cloning experiments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.588605/full#supplementary-material
References
Ai, P., Sun, S., Zhao, J., Fan, X., Xin, W., Guo, Q., et al. (2009). Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J. 57, 798–809. doi: 10.1111/j.1365-313x.2008.03726.x
Alewell, C., Ringeval, B., Ballabio, C., Robinson, D. A., Panagos, P., and Borrelli, P. (2020). Global phosphorus shortage will be aggravated by soil erosion. Nat. Commun. 11:4546.
Altschup, S. F., Gish, W., Pennsylvania, T., and Park, U. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410.
Ambrosini, A., Sant’Anna, F. H., Heinzmann, J., de Carvalho Fernandes, G., Bach, E., and Passaglia, L. M. P. (2018). Paenibacillus helianthi sp. nov., a nitrogen fixing species isolated from the rhizosphere of Helianthus annuus L. Antonie van Leeuwenhoek 111, 2463–2471. doi: 10.1007/s10482-018-1135-4
Anders, S., and Huber, W. (2010). Differential expression analysis for sequence count data. Genome Biol. 11:R106.
Atalla, A., and Schumann, W. (2003). The pst operon of Bacillus subtilis is specifically induced by alkali stress. J. Bacteriol. 185, 5019–5022. doi: 10.1128/jb.185.16.5019-5022.2003
Attallah, M. F., Metwally, S. S., Moussa, S. I., and Soliman, M. A. (2019). Environmental impact assessment of phosphate fertilizers and phosphogypsum waste: elemental and radiological effects. Microchem. J. 146, 789–797. doi: 10.1016/j.microc.2019.02.001
Baek, J. H., and Lee, S. Y. (2007). Transcriptome analysis of phosphate starvation response in Escherichia coli. J. Microbiol. Biotechnol. 17, 244–252.
Begley, T. P., Downs, D. M., Ealick, S. E., McLafferty, F. W., Van Loon, A. P., Taylor, S., et al. (1999). Thiamin biosynthesis in prokaryotes. Arch. Microbiol. 171, 293–300.
Beneduzi, A., Costa, P. B., Parma, M., Melo, I. S., Bodanese-Zanettini, M. H., and Passaglia, L. M. (2010). Paenibacillus riograndensis sp. nov., a nitrogen-fixing species isolated from the rhizosphere of Triticum aestivum. Int. J. Syst. Evol. Microbiol. 60, 128–133. doi: 10.1099/ijs.0.011973-0
Beneduzi, A., Peres, D., da Costa, P. B., Bodanese Zanettini, M. H., and Passaglia, L. M. (2008). Genetic and phenotypic diversity of plant-growth-promoting bacilli isolated from wheat fields in southern Brazil. Res. Microbiol. 159, 244–250. doi: 10.1016/j.resmic.2008.03.003
Bianco, C., and Defez, R. (2010). Improvement of phosphate solubilization and Medicago plant yield by an indole-3-acetic acid-overproducing strain of Sinorhizobium meliloti. Appl. Environ. Microbiol. 76, 4626–4632. doi: 10.1128/aem.02756-09
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Bringhurst, R. M., and Gage, D. J. (2002). Control of inducer accumulation plays a key role in succinate-mediated catabolite repression in Sinorhizobium meliloti. J. Bacteriol. 184, 5385–5392. doi: 10.1128/jb.184.19.5385-5392.2002
Brito, L. F., Bach, E., Kalinowski, J., Ruckert, C., Wibberg, D., Passaglia, L. M., et al. (2015). Complete genome sequence of Paenibacillus riograndensis SBR5T, a Gram-positive diazotrophic rhizobacterium. J. Biotechnol. 207, 30–31. doi: 10.1016/j.jbiotec.2015.04.025
Brito, L. F., Irla, M., Kalinowski, J., and Wendisch, V. F. (2017a). Detailed transcriptome analysis of the plant growth promoting Paenibacillus riograndensis SBR5 by using RNA-seq technology. BMC Genomics 18:846. doi: 10.1186/s12864-017-4235-z
Brito, L. F., Irla, M., Walter, T., and Wendisch, V. F. (2017b). Magnesium aminoclay-based transformation of Paenibacillus riograndensis and Paenibacillus polymyxa and development of tools for gene expression. Appl. Microbiol. Biotechnol. 101, 735–747. doi: 10.1007/s00253-016-7999-1
Brito, L. F., Schültenkamper, K., Passaglia, L. M. P., and Wendisch, V. F. (2020). CRISPR interference-based gene repression in the plant growth promoter Paenibacillus sonchi genomovar Riograndensis SBR5. Appl. Microbiol. Biotechnol. 104, 5095–5106. doi: 10.1007/s00253-020-10571-6
Candela, T., and Fouet, A. (2006). Poly-gamma-glutamate in bacteria. Mol. Microbiol. 60, 1091–1098. doi: 10.1111/j.1365-2958.2006.05179.x
Chang, K. C., Cheng, S. J., Chen, Y. C., Huang, H. R., and Liou, J. W. (2013). Nanoscopic analysis on pH induced morphological changes of flagella in Escherichia coli. J. Microbiol. Immunol. 46, 405–412. doi: 10.1016/j.jmii.2012.08.004
Chen, Y. P., Rekha, P. D., Arun, A. B., Shen, F. T., Lai, W., and Young, C. C. (2006). Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 34, 33–41. doi: 10.1016/j.apsoil.2005.12.002
Chung, H., Park, M., Madhaiyan, M., Seshadri, S., Song, J., Cho, H., et al. (2005). Isolation and characterization of phosphate solubilizing bacteria from the rhizosphere of crop plants of Korea. Soil Biol. Biochem. 37, 1970–1974. doi: 10.1016/j.soilbio.2005.02.025
Cordell, D., Drangert, J. O., and White, S. (2009). The story of phosphorus: global food security and food for thought. Glob. Environ. Chang. 19, 292–305. doi: 10.1016/j.gloenvcha.2008.10.009
Delavaux, C. S., Smith−Ramesh, L. M., and Kuebbing, S. E. (2017). Beyond nutrients: a meta-analysis of the diverse effects of arbuscular mycorrhizal fungi on plants and soils. Ecology 98, 2111–2119. doi: 10.1002/ecy.1892
Devine, K. M. (2018). Activation of the PhoPR-mediated response to phosphate limitation is regulated by wall teichoic acid metabolism in Bacillus subtilis. Front. Microbiol. 9:2678.
Din, A. R. J. M., Rosli, M. A., Azam, Z. M., Othman, N. Z., and Sarmidi, M. R. (2020). Paenibacillus polymyxa role involved in phosphate solubilization and growth promotion of Zea mays under abiotic stress condition. Proc. Natl. Acad. Sci. U.S.A. 90, 63–71. doi: 10.1007/s40011-019-01081-1
Divya, K. P., Prayag, M., Archana, G., and Kumar, G. N. (2011). Repression of mineral phosphate solubilizing phenotype in the presence of weak organic acids in plant growth promoting fluorescent Pseudomonas. Bioresour. Technol. 102, 3055–3061. doi: 10.1016/j.biortech.2010.10.041
Du, Q., Wang, H., and Xie, J. (2011). Thiamin (vitamin B1) biosynthesis and regulation : a rich source of anti- microbial drug targets. Int. J. Biol. Sci. 7, 41–52. doi: 10.7150/ijbs.7.41
Fernandes, G. C., Trarbach, L. J., de Campos, S. B., Beneduzi, A., and Passaglia, L. M. (2014). Alternative nitrogenase and pseudogenes: unique features of the Paenibacillus riograndensis nitrogen fixation system. Res. Microbiol. 165, 571–580. doi: 10.1016/j.resmic.2014.06.002
Gibson, D. G. (2014). Programming biological operating systems: genome design, assembly and activation. Nat. Methods 11, 521–526. doi: 10.1038/nmeth.2894
Gregory, P. J. (1988). “Growth and functioning of plant roots,” in Russell’s Soil Conditions and Plant Growth, eds E. J. Russel and A. Wild (Harlow: Longman Scientific & Technical), 113–167.
Guttenplan, S. B., and Kearns, D. B. (2013). Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 37, 849–871. doi: 10.1111/1574-6976.12018
Hanahan, D., and Harbor, C. S. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/s0022-2836(83)80284-8
Hilker, R., Stadermann, K. B., Doppmeier, D., Kalinowski, J., Stoye, J., Straube, J., et al. (2014). ReadXplorer – visualization and analysis of mapped sequences. Bioinformatics 30, 2247–2254. doi: 10.1093/bioinformatics/btu205
Hu, X., Chen, J., Guo, J., and Hangzhou, X. (2006). Two phosphate- and potassium-solubilizing bacteria isolated from Tianmu Mountain. World J. Microb. Biotechnol. 22, 983–990. doi: 10.1007/s11274-006-9144-2
Illmer, P., and Schinner, F. (1992). Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem. 24, 389–395. doi: 10.1016/0038-0717(92)90199-8
Irla, M., Heggeset, T. M. B., Nærdal, I., Paul, L., Haugen, T., Le, S. B., et al. (2016). Genome-based genetic tool development for Bacillus methanolicus: theta- and rolling circle-replicating plasmids for inducible gene expression and application to methanol-based cadaverine production. Front. Microbiol. 7:1481. doi: 10.3389/fmicb.2016.01481
Jensen, J. V. K., Eberhardt, D., and Wendisch, V. F. (2015). Modular pathway engineering of Corynebacterium glutamicum for production of the glutamate-derived compounds ornithine, proline, putrescine, citrulline, and arginine. J. Biotechnol. 214, 85–94. doi: 10.1016/j.jbiotec.2015.09.017
Jorge, A. M., Schneider, J., Unsleber, S., Gohring, N., Mayer, C., and Peschel, A. (2017a). Utilization of glycerophosphodiesters by Staphylococcus aureus. Mol. Microbiol. 103, 229–241. doi: 10.1111/mmi.13552
Jorge, J. M., Nguyen, A. Q., Perez-Garcia, F., Kind, S., and Wendisch, V. F. (2017b). Improved fermentative production of gamma-aminobutyric acid via the putrescine route: systems metabolic engineering for production from glucose, amino sugars, and xylose. Biotechnol. Bioeng. 114, 862–873. doi: 10.1002/bit.26211
Jorquera, M. A., Hernández, M. T., Rengel, Z., Marschner, P., De, M., and Mora, L. (2008). Isolation of culturable phosphobacteria with both phytate-mineralization and phosphate-solubilization activity from the rhizosphere of plants grown in a volcanic soil. Biol. Fert. Soils 44, 1025–1034. doi: 10.1007/s00374-008-0288-0
Kappes, R. M., Kempf, B., and Bremer, E. (1996). Three transport systems for the osmoprotectant glycine betaine operate in Bacillus subtilis: characterization of OpuD. J. Bacteriol. 178, 5071–5079. doi: 10.1128/jb.178.17.5071-5079.1996
Keilhauer, C., Eggeling, L., and Sahm, H. (1993). Isoleucine synthesis in corynebacterium glutamicum: molecular analysis of the ilvB-ilvN-ilvC Operon. J. Bacteriol. 175, 5595–5603. doi: 10.1128/jb.175.17.5595-5603.1993
Khan, M. S., Zaidi, A., Ahemad, M., Oves, M., and Wani, P. A. (2010). Plant growth promotion by phosphate solubilizing fungi – current perspective. Arch. Agron. Soil Sci. 56, 73–98. doi: 10.1080/03650340902806469
Kumar, V., Behl, R. K., and Narula, N. (2001). Establishment of phosphate-solubilizing strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat cultivars under green house conditions. Microbiol. Res. 156, 87–93. doi: 10.1078/0944-5013-00081
Langmead, B. (2010). Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinformatics 11:Unit11.7.
Lee, E. K., Zhang, X., Adler, P. R., Kleppel, G. S., and Romeiko, X. X. (2020). Spatially and temporally explicit life cycle global warming, eutrophication, and acidification impacts from corn production in the U.S. J. Cleaner Prod. 242:118465. doi: 10.1016/j.jclepro.2019.118465
Li, H., Yang, Z., Dai, M., Diao, X., Dai, S., Fang, T., et al. (2020). Input of Cd from agriculture phosphate fertilizer application in China during 2006–2016. Sci. Total Environ. 698:134149. doi: 10.1016/j.scitotenv.2019.134149
Li, Z., Bai, T., Dai, L., Wang, F., Tao, J., Meng, S., et al. (2016). A study of organic acid production in contrasts between two phosphate solubilizing fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 6:25313.
Lidbury, I. D., Murphy, A. R., Scanlan, D. J., Bending, G. D., Jones, A. M., Moore, J. D., et al. (2016). Comparative genomic, proteomic and exoproteomic analyses of three Pseudomonas strains reveals novel insights into the phosphorus scavenging capabilities of soil bacteria. Environ. Microbiol. 18, 3535–3549. doi: 10.1111/1462-2920.13390
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lünse, C. E., Scott, F. J., Suckling, C. J., and Mayer, G. (2014). Novel TPP-riboswitch activators bypass metabolic enzyme dependency. Front. Chem. 2:53. doi: 10.3389/fchem.2014.00053
Magallon-Servin, P., Antoun, H., Taktek, S., and de-Bashan, L. E. (2020). Designing a multi-species inoculant of phosphate rock-solubilizing bacteria compatible with arbuscular mycorrhizae for plant growth promotion in low-P soil amended with PR. Biol. Fertil. Soils 56, 521–536. doi: 10.1007/s00374-020-01452-1
Mardad, I., Serrano, A., and Soukri, A. (2013). Solubilization of inorganic phosphate and production of organic acids by bacteria isolated from a Moroccan mineral phosphate deposit. Afr. J. Microbiol. Res. 7, 626–635.
Marra, L. M., Oliveira, S. M. D., Ademar, P., and Ferreira, A. (2012). Biological nitrogen fixation and phosphate solubilization by bacteria isolated from tropical soils. Plant Soil 357, 289–307. doi: 10.1007/s11104-012-1157-z
Marra, L. M., Oliveira-longatti, S. M. D., Soares, C. R. F. S., Lima, J. M. D., Olivares, F. L., and Moreira, F. M. S. (2015). Initial pH of medium affects organic acids production but do not affect phosphate solubilization. Braz. J. Microbiol. 46, 367–375. doi: 10.1590/s1517-838246246220131102
Melnick, J., Lis, E., Park, J. H., Kinsland, C., Mori, H., Baba, T., et al. (2004). Identification of the two missing bacterial genes involved in thiamine salvage: thiamine pyrophosphokinase and thiamine kinase. J. Bacteriol. 186, 3660–3662. doi: 10.1128/jb.186.11.3660-3662.2004
Mendes, G. O., Freitas, A. L. M., Pereira, O. L., Silva, I. R., Vassilev, N. B., and Costa, M. D. (2013). Mechanisms of phosphate solubilization by fungal isolates when exposed to different P sources. Ann. Microbiol. 64, 239–249. doi: 10.1007/s13213-013-0656-3
Mentz, A., Neshat, A., Pfeifer-sancar, K., Pühler, A., Rückert, C., and Kalinowski, J. (2013). Comprehensive discovery and characterization of small RNAs in Corynebacterium glutamicum ATCC 13032. BMC Genomics 14:714. doi: 10.1186/1471-2164-14-714
Miller, K. J., and Wood, J. M. (1996). Osmoadaptation by rhizosphere bacteria. Ann. Rev. Microbiol. 50, 101–136. doi: 10.1146/annurev.micro.50.1.101
Mogollón, J. M., Beusen, A. H. W., van Grinsven, H. J. M., Westhoek, H., and Bouwman, A. F. (2018). Future agricultural phosphorus demand according to the shared socioeconomic pathways. Glob. Environ. Change 50, 149–163. doi: 10.1016/j.gloenvcha.2018.03.007
Murphy, J., and Riley, J. P. (1958). A single-solution method for the determination of soluble phosphate in sea water. J. Mar. Biol. Assoc. 37, 9–14. doi: 10.1017/s0025315400014776
Myers, C. L., Li, F. K., Koo, B. M., El-Halfawy, O. M., French, S., Gross, C. A., et al. (2016). Identification of two phosphate starvation-induced wall teichoic acid hydrolases provides first insights into the degradative pathway of a key bacterial cell wall component. J. Biol. Chem. 291, 26066–26082. doi: 10.1074/jbc.m116.760447
Narayan, S., Swarnalee, D., and Anil, D. (2010). Plant growth-promoting chitinolytic Paenibacillus elgii responds positively to tobacco root exudates. J. Plant Growth Regul. 29, 409–418. doi: 10.1007/s00344-010-9152-1
Nguyen, A. Q. D., Schneider, J., Reddy, G. K., and Wendisch, V. F. (2015). Fermentative production of the diamine putrescine: system metabolic engineering of Corynebacterium glutamicum. Metabolites 5, 211–231. doi: 10.3390/metabo5020211
Nosratabad, A. R. F., Etesami, H., and Shariati, S. (2017). Integrated use of organic fertilizer and bacterial inoculant improves phosphorus use efficiency in wheat (Triticum aestivum L.) fertilized with triple superphosphate. Rhizosphere 3, 109–111. doi: 10.1016/j.rhisph.2017.03.001
Ohtake, H., Wu, H., Imazu, K., Anbe, Y., Kato, J., and Kuroda, A. (1996). Bacterial phosphonate degradation, phosphite oxidation and polyphosphate accumulation. Resour. Conserv. Recy. 18, 125–134. doi: 10.1016/s0921-3449(96)01173-1
Palacios, O. A., Bashan, Y., and Luz, E. (2014). Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria — an overview. Biol. Fert. Soils 50, 415–432. doi: 10.1007/s00374-013-0894-3
Pandya, M., Rajput, M., and Rajkumar, S. (2015). Exploring plant growth promoting potential of non rhizobial root nodules endophytes of Vigna radiata. Microbiology 84, 80–89. doi: 10.1134/s0026261715010105
Parvaze, A. W., Mohammad Saghir, K., and Almas, Z. (2007). Synergistic effects of the inoculation with nitrogen-fixing and phosphate-solubilizing rhizobacteria on the performance of field-grown chickpea. J. Plant Nutr. Soil Sci. 170, 283–287. doi: 10.1002/jpln.200620602
Pikovskaya, R. I. (1948). Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiologiya 17, 362–370.
Prágai, Z., Allenby, N. E. E., O’Connor, N., Dubrac, S., Rapoport, G., Msadek, T., et al. (2004). Transcriptional regulation of the phoPR operon in Bacillus subtilis. J. Bacteriol. 186, 1182–1190. doi: 10.1128/jb.186.4.1182-1190.2004
Prágai, Z., and Harwood, C. R. (2002). Regulatory interactions between the Pho and sigma(B)-dependent general stress regulons of Bacillus subtilis. Microbiology 148, 1593–1602. doi: 10.1099/00221287-148-5-1593
Purvis, J. E., Yomano, L. P., and Ingram, L. O. (2005). Enhanced trehalose production improves growth of Escherichia coli under osmotic stress. Appl. Environ. Microbiol. 71, 3761–3769. doi: 10.1128/aem.71.7.3761-3769.2005
Qi, Y., Kobayashi, Y., and Hulett, F. M. (1997). The pst operon of Bacillus subtilis has a phosphate-regulated promoter and is involved in phosphate transport but not in regulation of the Pho regulon. J. Bacteriol. 179, 2534–2539. doi: 10.1128/jb.179.8.2534-2539.1997
Racki, L. R., Tocheva, E. I., Dieterle, M. G., Sullivan, M. C., Jensen, G. J., and Newman, D. K. (2017). Polyphosphate granule biogenesis is temporally and functionally tied to cell cycle exit during starvation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 114, E2440–E2449.
Rajput, M. S., Iyer, B., Pandya, M., and Jog, R. (2015). Derepression of mineral phosphate solubilization phenotype by insertional inactivation of iclR in Klebsiella pneumoniae. PLoS One 10:e0138235. doi: 10.1371/journal.pone.0138235
Rajput, M. S., Naresh Kumar, G., and Rajkumar, S. (2013). Repression of oxalic acid-mediated mineral phosphate solubilization in rhizospheric isolates of Klebsiella pneumoniae by succinate. Arch. Microbiol. 195, 81–88. doi: 10.1007/s00203-012-0850-x
Rashid, M., Khalil, S., Ayub, N., Alam, S., and Latif, F. (2004). Organic acids production and phosphate solubilization by phosphate solubilizing microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci. 7, 187–196. doi: 10.3923/pjbs.2004.187.196
Richardson, A. E., and Hadobas, P. A. (1997). Soil isolates of Pseudomonas spp. that utilize inositol phosphates. Can. J. Microbiol. 43, 509–516. doi: 10.1139/m97-073
Rodríguez, H., and Fraga, R. (1999). Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 17, 319–339. doi: 10.1016/s0734-9750(99)00014-2
Rodriguez, H., Rossolini, G. M., Gonzalez, T., Li, J., and Glick, B. R. (2000). Isolation of a gene from Burkholderia cepacia IS-16 encoding a protein that facilitates phosphatase activity. Curr. Microbiol. 40, 362–366. doi: 10.1007/s002840010071
Sambrook, J. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 2100.
Sant’Anna, F. H., Ambrosini, A., de Souza, R., de Carvalho Fernandes, G., Bach, E., Balsanelli, E., et al. (2017). Reclassification of Paenibacillus riograndensis as a genomovar of Paenibacillus sonchi: genome-based metrics improve bacterial taxonomic classification. Front. Microbiol. 8:1849. doi: 10.3389/fmicb.2017.01849
Santos-Beneit, F. (2015). The Pho regulon: a huge regulatory network in bacteria. Front. Microbiol. 6:402. doi: 10.3389/fmicb.2015.00402
Schütz, A., Golbik, R., Tittmann, K., Svergun, D. I., Koch, M. H., Hubner, G., et al. (2003). Studies on structure-function relationships of indolepyruvate decarboxylase from Enterobacter cloacae, a key enzyme of the indole acetic acid pathway. Eur. J. Biochem. 270, 2322–2331. doi: 10.1046/j.1432-1033.2003.03602.x
Seldin, L., Rosado, A. S., William, D., Cruz, D. A., Nobrega, A., Elsas, J., et al. (1998). Comparison of Paenibacillus azotofixans strains isolated from rhizoplane, rhizosphere, and non-root-associated soil from maize planted in two different Brazilian soils. Appl. Environ. Microbiol. 64, 3860–3868. doi: 10.1128/aem.64.10.3860-3868.1998
Shahid, M., and Hameed, S. (2012). Root colonization and growth promotion of sunflower (Helianthus annuus L.) by phosphate solubilizing Enterobacter sp. World J. Microbiol. Biotechnol. 28, 2749–2758. doi: 10.1007/s11274-012-1086-2
Sharma, S. B., Sayyed, R. Z., Trivedi, M. H., and Gobi, T. A. (2013). Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2:587.
Shen, J., Yuan, L., Zhang, J., Li, H., Bai, Z., and Chen, X. (2011). Phosphorus dynamics : from soil to plant. Plant Physiol. 156, 997–1005. doi: 10.1104/pp.111.175232
Shi, L., and Hulett, F. M. (1999). The cytoplasmic kinase domain of PhoR is sufficient for the low phosphate-inducible expression of Pho regulon genes in Bacillus subtilis. Mol. Microbiol. 31, 211–222. doi: 10.1046/j.1365-2958.1999.01163.x
Shin, H., Shin, H. S., Dewbre, G. R., and Harrison, M. J. (2004). Phosphate transport in Arabidopsis: Pht1;1 and Pht1;4 play a major role in phosphate acquisition from both low- and high-phosphate environments. Plant J. 39, 629–642.
Sleator, R. D., and Hill, C. (2002). Bacterial osmoadaptation: the role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 26, 49–71. doi: 10.1111/j.1574-6976.2002.tb00598.x
Sperb, E. R., Tadra-Sfeir, M. Z., Sperotto, R. A., Fernandes, G. C., Pedrosa, F. O., de Souza, E. M., et al. (2016). Iron deficiency resistance mechanisms enlightened by gene expression analysis in Paenibacillus riograndensis SBR5. Res. Microbiol. 167, 501–509. doi: 10.1016/j.resmic.2016.04.007
Summers, M. L., Denton, M. C., and Dermott, T. R. M. C. (1999). Genes coding for phosphotransacetylase and acetate kinase in Sinorhizobium meliloti are in an operon that is inducible by phosphate stress and controlled by PhoB. J. Bacteriol. 181, 2217–2224. doi: 10.1128/jb.181.7.2217-2224.1999
Tcherpakov, M., Ben-jacob, E., and Gutnick, D. L. (1999). Paenibacillus Dendritiformis Sp. Nov., proposal for a new pattern-forming species and its localization within a phylogenetic cluster. Int. J. Syst. Evol. Microbiol. 49, 239–246. doi: 10.1099/00207713-49-1-239
Thaller, M. C., Berlutti, F., Schippa, S., Iori, P., Passariello, C., and Rossolini, G. M. (1995). Heterogeneous patterns of acid phosphatases containing low-molecular-mass polypeptides in members of the family Enterobacteriaceae. Int. J. Syst. Bacteriol. 45, 255–261. doi: 10.1099/00207713-45-2-255
Thomas, L., Hodgson, D. A., Wentzel, A., Nieselt, K., Ellingsen, T. E., Moore, J., et al. (2012). Metabolic switches and adaptations deduced from the proteomes of Streptomyces coelicolor wild type and phop mutant grown in batch culture. Mol. Cell. Proteomics 11, M111.013797.
Vance, C. P., Uhde-stone, C., and Allan, D. L. (2003). Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 157, 423–447. doi: 10.1046/j.1469-8137.2003.00695.x
Vyas, P., and Gulati, A. (2009). Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 9:174. doi: 10.1186/1471-2180-9-174
Wang, J., Zhao, Y., and Maqbool, F. (2020). Capability of Penicillium oxalicum y2 to release phosphate from different insoluble phosphorus sources and soil. Folia Microbiol. doi: 10.1007/s12223-020-00822-4 [Epub ahead of print].
Wang, Y., Shi, Y., Li, B., Shan, C., Ibrahim, M., Jabeen, A., et al. (2012). Phosphate solubilization of Paenibacillus polymyxa and Paenibacillus macerans from mycorrhizal and non-mycorrhizal cucumber plants. Afr. J. Microbiol. Res. 6, 4567–4573.
Wendisch, V. F., Spies, M., Reinscheid, D. J., Schnicke, S., Sahm, H., and Eikmanns, B. J. (1997). Regulation of acetate metabolism in Corynebacterium glutamicum: transcriptional control of the isocitrate lyase and malate synthase genes. Arch. Microbiol. 168, 262–269. doi: 10.1007/s002030050497
Werra, P. D., Pe, M., Keel, C., and Maurhofer, M. (2009). Role of gluconic acid production in the regulation of biocontrol traits of Pseudomonas fluorescens CHA0. Appl. Environ. Microbiol. 75, 4162–4174. doi: 10.1128/aem.00295-09
Whitelaw, M. A. (2000). Growth promotion of plants inoculated with phosphate-solubilizing fungi. Adv. Agron. 69, 99–151. doi: 10.1016/s0065-2113(08)60948-7
Xie, J., Shi, H., Du, Z., Wang, T., Liu, X., and Chen, S. (2016). Comparative genomic and functional analysis reveal conservation of plant growth promoting traits in Paenibacillus polymyxa and its closely related species. Sci. Rep. 6:21329.
Yi, Y., Huang, W., and Ge, Y. (2008). Exopolysaccharide: a novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microbiol. Biotechnol. 24, 1059–1065. doi: 10.1007/s11274-007-9575-4
Zahoor, A., Otten, A., and Wendisch, V. F. (2014). Metabolic engineering of Corynebacterium glutamicum for glycolate production. J. Biotechnol. 192, 366–375. doi: 10.1016/j.jbiotec.2013.12.020
Zaidi, A., Khan, M., Ahemad, M., and Oves, M. (2009). Plant growth promotion by phosphate solubilizing bacteria. Acta Microbiol. Immunol. Hungar. 56, 263–284. doi: 10.1556/amicr.56.2009.3.6
Keywords: Paenibacillus sonchi, phosphate solubilization, PGPR – plant growth-promoting rhizobacteria, organic acids, osmoprotection, RNAseq
Citation: Brito LF, López MG, Straube L, Passaglia LMP and Wendisch VF (2020) Inorganic Phosphate Solubilization by Rhizosphere Bacterium Paenibacillus sonchi: Gene Expression and Physiological Functions. Front. Microbiol. 11:588605. doi: 10.3389/fmicb.2020.588605
Received: 29 July 2020; Accepted: 16 November 2020;
Published: 14 December 2020.
Edited by:
Hauke Smidt, Wageningen University & Research, NetherlandsReviewed by:
Sergio de los Santos Villalobos, Instituto Tecnológico de Sonora (ITSON), MexicoManoj Kumar Solanki, Agricultural Research Organization (ARO), Israel
Gilberto de Oliveira Mendes, Federal University of Uberlândia, Brazil
Copyright © 2020 Brito, López, Straube, Passaglia and Wendisch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Volker F. Wendisch, volker.wendisch@uni-bielefeld.de
†Present address: Luciana F. Brito, Department of Biotechnology and Food Science, Norwegian University of Science and Technology, Trondheim, Norway; Marina Gil López, Department of Biotechnology and Food Science, Norwegian University of Science and Technology, Trondheim, Norway