 Murray P. Cox1Yanan Guo1David J. Winter2Diya Sen3
Murray P. Cox1Yanan Guo1David J. Winter2Diya Sen3 Nicholas C. Cauldron4Jason Shiller3Ellie L. Bradley5Austen R. Ganley6
Nicholas C. Cauldron4Jason Shiller3Ellie L. Bradley5Austen R. Ganley6 Monica L. Gerth7
Monica L. Gerth7 Randy F. Lacey7Rebecca L. McDougal3Preeti Panda8Nari M. Williams9Niklaus J. Grunwald10
Randy F. Lacey7Rebecca L. McDougal3Preeti Panda8Nari M. Williams9Niklaus J. Grunwald10 Carl H. Mesarich5
Carl H. Mesarich5 Rosie E. Bradshaw1*
Rosie E. Bradshaw1*- 1Laboratory of Molecular Plant Pathology/Bioprotection Aotearoa, School of Natural Sciences, Massey University, Palmerston North, New Zealand
- 2Institute of Environmental Science and Research (ESR), Porirua, New Zealand
- 3Scion, Rotorua, New Zealand
- 4Department of Botany and Plant Pathology, Oregon State University, Corvallis, OR, United States
- 5Laboratory of Molecular Plant Pathology/Bioprotection Aotearoa, School of Agriculture and Environment, Massey University, Palmerston North, New Zealand
- 6School of Biological Sciences and Digital Life Institute, University of Auckland, Auckland, New Zealand
- 7Bioprotection Aotearoa, School of Biological Sciences, Victoria University of Wellington, Wellington, New Zealand
- 8Plant and Food Research, Lincoln, New Zealand
- 9Plant and Food Research, Hawkes Bay, New Zealand
- 10Horticultural Crops Disease and Pest Management Research Unit, USDA Agricultural Research Service, Corvallis, OR, United States
Phytophthora species are notorious plant pathogens, with some causing devastating tree diseases that threaten the survival of their host species. One such example is Phytophthora agathidicida, the causal agent of kauri dieback – a root and trunk rot disease that kills the ancient, iconic and culturally significant tree species, Agathis australis (New Zealand kauri). A deeper understanding of how Phytophthora pathogens infect their hosts and cause disease is critical for the development of effective treatments. Such an understanding can be gained by interrogating pathogen genomes for effector genes, which are involved in virulence or pathogenicity. Although genome sequencing has become more affordable, the complete assembly of Phytophthora genomes has been problematic, particularly for those with a high abundance of repetitive sequences. Therefore, effector genes located in repetitive regions could be truncated or missed in a fragmented genome assembly. Using a combination of long-read PacBio sequences, chromatin conformation capture (Hi-C) and Illumina short reads, we assembled the P. agathidicida genome into ten complete chromosomes, with a genome size of 57 Mb including 34% repeats. This is the first Phytophthora genome assembled to chromosome level and it reveals a high level of syntenic conservation with the complete genome of Peronospora effusa, the only other completely assembled genome sequence of an oomycete. All P. agathidicida chromosomes have clearly defined centromeres and contain candidate effector genes such as RXLRs and CRNs, but in different proportions, reflecting the presence of gene family clusters. Candidate effector genes are predominantly found in gene-poor, repeat-rich regions of the genome, and in some cases showed a high degree of duplication. Analysis of candidate RXLR effector genes that occur in multicopy gene families indicated half of them were not expressed in planta. Candidate CRN effector gene families showed evidence of transposon-mediated recombination leading to new combinations of protein domains, both within and between chromosomes. Further analysis of this complete genome assembly will help inform new methods of disease control against P. agathidicida and other Phytophthora species, ultimately helping decipher how Phytophthora pathogens have evolved to shape their effector repertoires and how they might adapt in the future.
Introduction
Phytophthora is an oomycete genus named for its reputation as a ‘plant destroyer’. Some species are notorious pathogens of food crops (Fones et al., 2020). For example, P. infestans has been known as a potato pathogen since the 19th century and P. sojae was identified as a soybean pathogen in the 1950s (Govers and Gijzen, 2006). Since 2000, however, the number of Phytophthora species identified as pathogens in forest ecosystems has increased around the world (Hansen, 2015). For example, P. ramorum was discovered as the causal agent of sudden oak death in the United States (Rizzo et al., 2002), P. kernoviae on beech in England (Brasier et al., 2005) and P. pluvialis on pines in New Zealand (Dick et al., 2014). More Phytophthora species are being discovered and identified as new disease outbreaks are investigated and new species identified (Hansen, 2015; Scott et al., 2019; Keriö et al., 2020).
Phytophthora agathidicida, which is in Phytophthora Clade 5, causes a dieback disease of New Zealand kauri trees known as kauri dieback (Weir et al., 2015). The pathogen invades and colonises the roots and vascular tissue, resulting in resin secretion (gummosis) from trunk lesions, crown decline and eventual death of the tree (Bradshaw et al., 2020). P. agathidicida is currently thought to have a narrow host range and limited geographic distribution (Bradshaw et al., 2020). While it is generally considered an introduced pathogen, its origin is unknown (Winkworth et al., 2021). The host of this disease, New Zealand kauri (Agathis australis (D.Don) Loudon), is part of the ancient Araucariaceae family, with trees capable of living for over 1,500 years (Ahmed and Ogden, 1987). Kauri is an iconic species with immense cultural significance for indigenous Māori people and is an ecologically vital foundation species for many of New Zealand’s native forests (Wyse et al., 2014; Black et al., 2018; Lambert et al., 2018).
An important focus of oomycete research has been the identification and analysis of proteinaceous effectors that are produced by the pathogens. These include intracellular effectors such as RXLR and CRN (crinkling and necrosis) proteins, and apoplastic effectors such as elicitins, NLPs (necrosis-like proteins) and some CAZymes (carbohydrate-active enzymes) that collectively facilitate pathogen invasion of their hosts (Amaro et al., 2017; Wang et al., 2019b; Rocafort et al., 2020; Chepsergon et al., 2021; Bradley et al., 2022; Wang et al., 2022; Wilson and McDowell, 2022). In some cases, plant hosts have evolved to recognise specific pathogen effectors by means of immune receptors, which then elicit plant defence responses. Pathogens, however, can evolve to avoid recognition through deletion, mutation or silencing of these effectors, provided that a significant associated fitness penalty is not incurred (Pais et al., 2018; Dong and Ma, 2021).
The availability of pathogen genome sequences has revolutionised many aspects of plant pathology. These sequences facilitate an unprecedented level of knowledge that can be exploited for many purposes such as developing diagnostic tools, screening for pathogen effectors to identify host resistance and making disease predictions based on patterns of pathogen dispersal and evolution (Hamelin and Roe, 2020; Keriö et al., 2020). Draft genome sequences are now available for many Phytophthora pathogens of forest trees (Tyler et al., 2006; Quinn et al., 2013; Feau et al., 2016; Grenville-Briggs et al., 2017; Vetukuri et al., 2018; McGowan et al., 2020; Thorpe et al., 2021), including an earlier draft genome of P. agathidicida (Studholme et al., 2016). However, this previously assembled P. agathidicida genome sequence, like those of many other Phytophthora species obtained using short-read Illumina sequencing, was highly fragmented with >3,000 scaffolds (Haas et al., 2009; Studholme et al., 2016; Fletcher and Michelmore, 2018; Guo et al., 2020a). The prevalence of repeated sequences makes Phytophthora genomes recalcitrant to assembly. Repetitive, transposon-rich regions of oomycete genomes are often enriched in effector genes, facilitating the adaptive evolution of those genes (Dong et al., 2015; Zhang et al., 2019). Thus, annotation of genome assemblies in which some repetitive regions are missing often excludes some effector genes. For instance, a high-quality genome assembly of P. cinnamomi led to the identification of candidate effector genes that were not present in an earlier, more fragmented, genome assembly (Engelbrecht et al., 2021).
The importance of high-quality genome assemblies for accurate and complete identification of candidate effector genes has led to considerable efforts to exploit long-read sequencing technologies to create better-assembled genome sequences (Fletcher and Michelmore, 2018). For example, scaffolds numbering in the thousands from short-read (principally Illumina) technologies have been reduced to hundreds for some species including P. capsici (Cui et al., 2019; Stajich et al., 2021), P. cinnamomi (Engelbrecht et al., 2021) and P. sojae (Fang et al., 2020), and less than 30 scaffolds for P. ramorum (Carleson et al., 2022). However, despite recent advances in long-read sequencing technologies and assembly techniques (Fletcher and Michelmore, 2018), it has proven difficult to achieve chromosome-level assemblies of Phytophthora genomes, principally due to their high repeat content, frequent heterozygosity and relatively large size.
Here we report the chromosome-level sequence assembly of the 57 Mb P. agathidicida genome and chromosome-level distributions of key classes of genes and structural genomic elements. This genome assembly will not only provide an important foundation for better understanding the molecular mechanisms underpinning virulence and pathogenicity by the kauri dieback pathogen, but will also act as a reference for other Phytophthora species.
Materials and methods
Phytophthora agathidicida isolate
The type strain of Phytophthora agathidicida B.S. Weir, Beever, Pennycook & Bellgard, isolate NZFS3770 (International Collection of Microorganisms [ICMP] 17,027), was initially collected in 2006 from Great Barrier Island (Aotea), New Zealand (WGS84 coordinates 175.417921E 36.225347S). The Ngāti Rehua Ngātiwai ki Aotea Trust has cultural authority over this isolate, which is hereafter referred to as P. agathidicida 3770.
DNA extraction and sequencing
For genomic DNA (gDNA) extraction, P. agathidicida 3770 mycelium was grown on cellophane membranes on V8 agar plates (Weir et al., 2015) at 22°C in the dark for 5 days before harvesting and freeze-drying. High molecular weight genomic DNA was extracted using a Blood and Cell Culture DNA Midi Kit (Qiagen, Hilden, Germany) and a 100G Genomic-tip 100/G (Qiagen). Genomic DNA quality and quantity were assessed on a 0.4% agarose gel and by using a Qubit Fluorometer (Life Technologies, Singapore). For whole-genome sequencing, a P. agathidicida genomic DNA library (subread length 20–30 kb) was prepared and sequenced on a PacBio Sequel II platform by Novogene (Hong Kong), via the Massey University Genome Service, yielding 7.7 million reads, average length 19.9 kb, N50 = 32.7 kb. The same genomic DNA sample was also sequenced on an Illumina Novaseq™ 6000 platform, yielding 34 million 150 bp paired-end reads.
Hi-C libraries and sequencing
For Hi-C analysis, P. agathidicida 3770 mycelium was grown in triplicate in V8 broth with shaking at 180 rpm for 4 days at 22°C in the dark. Mycelium was harvested from each replicate flask and divided such that half was used for the Hi-C library (pooled from three replicates) and half retained for RNA extraction (three separate replicates).
The Hi-C library was made using a Proximo Hi-C (Fungal) kit (Phase Genomics, Seattle, WA), following the manufacturer’s instructions. The quality and quantity of the Hi-C library were checked using a LabChip® GX Touch HT Nucleic Acid Analyser (Perkin Elmer, Melbourne, Australia), with the library then sequenced on an Illumina Novaseq™ 6000 platform, yielding 310 million 150 bp paired-end reads.
RNA extraction and sequencing
For Iso-Seq analysis, RNA was extracted from P. agathidicida 3770 grown under a diverse range of growth conditions in order to maximise the number and diversity of genes represented; namely (1) mycelia grown in clarified V8, potato dextrose (PD) and plich broth (Meijer et al., 2014) at 22°C for 3 days without shaking; (2) mycelium incubated for 1.5, 3, 6 and 24 h in the presence of kauri leaves (from seeds originally sourced from Waipoua forest, New Zealand; Ye et al., 2011); and (3) a mixture of zoospores and cysts (Lawrence et al., 2017). RNA was extracted from the combined samples by using an RNeasy Plant Mini Kit (Qiagen). A cDNA Iso-Seq library with 1–10 kb reads was made by Novogene by pooling a size-selected library with a non-size-selected library to increase the diversity and yield of fragments >4 kb. This pooled library was sequenced on a PacBio Sequel (Novogene), yielding 34 million subreads with an average read length of 2.36 kb.
To complement the Hi-C data, RNA was also extracted from the P. agathidicida 3770 mycelium grown for Hi-C analysis (see above) using an RNeasy Plant Mini Kit (Qiagen). Gene expression was calculated from the three biological replicates, sequenced to a depth of over 30 million 150 bp paired-end reads per replicate, on the Illumina Novaseq™ 6000 platform. Reads mapping uniquely to single gene model exons were identified with STAR 2.7.10a (Dobin et al., 2013), using the gene annotations described below, and gene expression levels were subsequently classified by percentiles.
We took advantage of the new genome assembly and annotation described in this study to re-map pre-existing RNA-seq reads from another isolate of P. agathidicida, NZFS3813 (Herewini et al., 2018), that had been grown on kauri leaves and roots (from seeds originally sourced from Waipoua forest, NZ) in a project developed with the cultural authority of Te Roroa. The experimental design, growth conditions and RNA extraction were described previously (Guo et al., 2020a). RNA library construction and Illumina sequencing were performed by the Beijing Genomics Institute (BGI), as part of the Scion ‘Healthy Trees Healthy Future’ Programme, to yield 150 bp paired-end reads, with approximately 150 million reads from each of three replicates for 6 and 24 h samples, and 75 million reads for 48 and 72 h samples. We also prepared in vitro RNA samples for Pa3813 to enable comparison with in planta gene expression. For this, P. agathidicida 3813 was grown in V8 broth at 22°C for 5 days without shaking. RNA was extracted using a Spectrum™ Plant Total RNA Kit (Sigma) and sequenced with 150 bp paired-end reads by Novogene using an Illumina NovaSeq™ 6000.
The sequencing reads from all experimental conditions were mapped to the P. agathidicida 3770 genome using HISAT2 v2.2.1 (Kim et al., 2015, 2019), with default settings except without repeat-masking and with the addition of the ‘--no-unal’ setting to suppress the inclusion of unaligned reads in the output alignment file that are expected to derive from the kauri host. The default mapping parameters used in the Hisat2 mapping tool do not preclude multiple read mapping in the case of duplicated genes. Read counts were calculated for each gene using ‘featureCounts’ (Liao et al., 2014) in the R package Rsubread v2.10.5 (Liao et al., 2019). Differential gene expression between the in vitro samples and each of the other experimental conditions was performed with edgeR v3.38.4 (Robinson et al., 2010) using the quasi-likelihood pipeline (Chen et al., 2016). Gene expression data were transformed and expressed as log2 fragments per kilobase of exon per million mapped fragments (FPKM).
Genome assembly and synteny analysis
Assemblies were produced from the PacBio data using three programs with different assembly algorithms, Canu v2.1 (Koren et al., 2017), Mecat2 v2020.02.28 (Xiao et al., 2017) and Flye v2.8.3 (Kolmogorov et al., 2019). The three algorithms complemented each other, with discontinuities present in the primary assembly from Canu often being well resolved in the others. The small number of non-assembled gaps could be resolved by identifying multiple reads with >10 kb of sequence matching each broken flank. As a conservative strategy, these gaps were closed by scaffolding the two contigs with a string of 10 Ns. The final assembly was polished with Pilon v1.23 (Walker et al., 2014) using 150 bp paired-end Illumina MiSeq reads produced as part of this project using the same gDNA sample as used for the PacBio libraries. Centromeres were identified by their characteristic cross-like patterns in the Hi-C contact map. Following standard practice, chromosomes were ordered by size, oriented with short arms first. The ribosomal DNA locus was manually annotated and, by convention, trimmed to two complete rDNA units together with the partial flanking units present at each terminus. Telomeres were trimmed to the last complete canonical repeat.
Large-scale synteny comparisons of the P. agathidicida 3770 genome relative to other oomycete species was determined by comparison to the 70-contig draft genome sequence of P. sojae isolate P6497 (NCBI accession number GCA_009848525.1; Fang et al., 2020), and to the genome of Peronospora effusa isolate UA202013 (GCA_021491655.1; the only other completely assembled genome sequence of an oomycete species; Fletcher et al., 2021). These comparisons were performed with D-GENIES v1.3.1 (Cabanettes and Klopp, 2018), using a cross-mapping algorithm that employs minimap2 v2.24-r1122 (Li, 2018).
Repetitive elements analysis
We produced a de novo annotation of repeats in the P. agathidicida 3770 genome using the RepeatModeler v2.0.1 pipeline (Flynn et al., 2020). This pipeline uses RepeatScout to discover multicopy regions of the genome and RECON (Bao and Eddy, 2002) to discover known transposable element (TE) motifs. We used RepeatMasker v4.0.6 (Chen, 2004) to locate specific elements of each identified repetitive element family within the genome. Having already identified centromere locations with the Hi-C 3D data, we used BLASTn v2.5.0 to search the genome for elements with homology to the CoLT retrotransposons that occur in P. sojae centromeric regions (Fang et al., 2020). Genomic locations with more than 10 copies of a CoLT retrotransposon element confirmed the positions of centromeres in the P. agathidicda genome.
Gene prediction and functional annotation
Genes encoding tRNAs were identified using tRNAscan-SE v2.0 (Lowe and Eddy, 1997) and non-protein-coding RNAs by searching v14.5 of the Rfam database (Kalvari et al., 2021) using the cmscan option from Infernal v1.1.2 (Nawrocki and Eddy, 2013) for all other RNA classes. We identified putative mRNA transcripts from our IsoSeq data using v3.0 of the PacBio IsoSeq pipeline1.
Protein-coding genes were identified from the masked P. agathidicida 3770 genome using Augustus v3.3.3 (Stanke et al., 2006). We generated training data for Augustus gene-calling using our IsoSeq transcripts following a protocol originally designed for expressed sequence tags (Hoff and Stanke, 2019), then used minimap2 v2.17 to align transcripts to the reference genome and the PASA v2.4.1 pipeline to curate the resulting alignments. Proteins from the current version of the P. sojae P6497 genome (Tyler et al., 2006) were also used as evidence in our gene calling pipeline.
To assess the completeness of the P. agathidicida 3770 genome assembly, BUSCO v4.14 (Benchmarking Universal Single-Copy Orthologs; Seppey et al., 2019) was used in genome mode with the stramenopile lineage and Eukaryota gene sets from OrthoDB (Zdobnov et al., 2021), using default parameter values. The BUSCO values were compared with those of other oomycete genome assemblies produced from long-read sequences, including those of Pe. effusa, (Fletcher et al., 2021) P. ramorum (Carleson et al., 2022) and P. sojae (Fang et al., 2020). The accuracy of our automated gene-calling was assessed by comparing the resulting gene models to hand-curated models produced from other studies focusing on RXLR effectors (Guo et al., 2020a) and carbohydrate metabolism (Bradley, 2022).
Functional domains in predicted proteins were identified using InterProScan v5.53. Signal peptides were predicted using SignalP v3.0 (Bendtsen et al., 2004) and transmembrane domain prediction was done using TMHMM v2.0 (Krogh et al., 2001). SignalP v3.0 was chosen as it was previously deemed to be the most sensitive version for the prediction of signal peptides in oomycete effector proteins (Sperschneider et al., 2015).
A set of six gene classes were selected for further investigation based on their known roles in virulence in other Phytophthora species. Candidate CRNs and RXLRs were annotated in both the predicted proteins and all 6-frame translations of the genomes from getorf (−find 1 -minsize 210) using EMBOSS v6.6.0.0. ORFs in intergenic spaces between predicted genes that encode candidate effectors were supplemented into the annotations using a custom Python script2.
Putative RXLR effector proteins were identified using previously described approaches (McGowan and Fitzpatrick, 2017; Guo et al., 2020a). Briefly, sequences were tested using three prediction methods (Bhattacharjee et al., 2006; Whisson et al., 2007; Win et al., 2007) implemented in a Python script (Cock et al., 2013). The intersection of candidates returned by all methods were considered RXLRs.
Sequences were putatively called as CRNs if they were identified from either a regular expression or a machine-learning approach. The regular expression approach implemented in effectR v1.0.2 (Tabima and Grünwald, 2019) was used to search for any sequence containing the LFLAK motif within or immediately following the first 90 amino acids. For the machine learning approach, an HMM profile was created from all 315 CRNs from P. infestans (Haas et al., 2009) aligned with MAFFT v7.480 (--legacygappenalty --genafpair --maxiterate 1000; Katoh and Standley, 2013) and hmmsearch v3.1b2 was used to identify sequences with a positive bit score. Lastly, any candidate not containing a degenerate LFLAK motif (L[FY]LA[RK]) within or immediately following the first 90 amino acids was removed (McGowan and Fitzpatrick, 2017; Thorpe et al., 2021).
Proteins were considered CAZymes if they were predicted as such by at least two of the three tools used by dbCAN2 (released 21 Dec 2021; HMMER, DIAMOND, and eCAMI; Zhang et al., 2018). Candidate elicitins and NLPs were identified on the basis of their pfam domains: PF00964 and PF05630, respectively (Derevnina et al., 2016). G-protein-coupled receptors (GPCRs) were predicted using the GPCR-Pen pipeline (Begum et al., 2020).
Intergenic distance and 3D clustering analysis of gene classes
Intergenic 5′ and 3′ distances to the nearest gene (‘flanking intergenic regions’ or FIRs) were calculated for all genes, to determine the background distribution, as well as for the six gene classes of interest (CAZymes, CRNs, elicitins, GPCRs, NLPs and RXLRs). Custom code3 was used to plot the 5′ and 3′ distances against the background distribution. Monte Carlo probabilities that the median distance of each subset of genes is greater than that expected for the same number of randomly chosen genes was calculated via bootstrapping with replacement and corrected for multiple hypothesis testing using the false discovery rate approach. Individual genes were overlaid with the percentile of their expression level in culture.
Clustering of genes in the 3D space of the nucleus can be ascertained from Hi-C data. 3D interactions were calculated for the same six gene classes as above. Custom code4 calculated the mean number of pairwise Hi-C contacts between all genes in each class, and determined using Monte Carlo statistics whether their 3D interactions differed significantly from equivalently sized sets of genes chosen randomly across the genome. The Hi-C contact matrix had a window resolution of 10 kb. Probability values were corrected for multiple testing using the False Discovery Rate (FDR) implementation in the p.adjust function of R v4.2.0 (R Core Team, 2022).
Single nucleotide variants
Illumina genome data were available for 13 additional New Zealand isolates of P. agathidicida as NCBI BioProjects PRJNA290658, PRJNA486676 and PRJNA864064 (Studholme et al., 2016; Guo et al., 2020a). These Illumina reads were aligned to the P. agathidicida 3770 reference genome using BWA-mem v0.7.17-r1188 (Li, 2013). Duplicate reads were marked by Picard Tools MarkDuplicates v2.21.8.5 Variants were called using GATK v4.1.8.1 (Van der Auwera and O'Connor, 2020). First, all samples were separately called with HaplotypeCaller (Poplin et al., 2018) (setting the -ERC parameter of GVCF to produce per sample genotype/gVCF files). Next, gVCF files were merged with GenomicsDBImport and finally joint-genotyped with GenotypeGVCF. Variants were filtered to retain single nucleotide polymorphisms (SNPs) using GATK SelectVariants. Next, VCFTools was used for retaining only biallelic sites (--min-alleles 2 --max-alleles 2) and hard filters were applied for missing data (max-missing = 0.8), minimum depth (minDP = 10) and minimum quality (minQ = 30). The final vcf file containing filtered variants from all the genomes was used for downstream analyses.
To estimate mean SNPs per exon, all exons were extracted from the P. agathidicida 3770 GFF genome annotation file into a BED file. Locations of SNPs from the joint-called VCF file containing SNPs from all isolates were also extracted into a BED file. SNPs overlapping exons were counted by using bedtools intersect with the two BED files followed by the -c option to count the number of overlaps. The results were summarised as means using the summarise function in the dplyr package (Wickham et al., 2015) in R v3.6.2.
Ploidy and heterozygosity estimation and copy number variation analysis
For estimation of ploidy and heterozygosity, K-mer analysis was performed with P. agathidicida 3770 Illumina reads using the KMC algorithm (Deorowicz et al., 2013), then genome profiling was carried out with Genomescope v2.0 (Ranallo-Benavidez et al., 2020) using a K-mer size of 21.
Copy number variation (CNV) was analysed using CNVPytor (Suvakov et al., 2021), with the P. agathidicida 3770 assembly file used as the reference, followed by GC correction. BAM and VCF files for all the P. agathidicida isolate samples (outlined in Section “Single nucleotide variants”) were processed to extract read depth and allele frequency information using three different bin sizes (1, 5, and 10 kb) following protocols outlined in the manual.6 CNVs were called using the CNVpytor mean-shift method for all three bin sizes, and the results were compared manually. The final file contained copy number calls from the 5 kb bin size, with the column ‘CNV_level’ in the output file normalized to a copy number of 2. CNVs were visualised in KaryoploteR v3.15 (Gel and Serra, 2017) with the kpPlotRegions function and labelled as deletions (copy number < 2) or duplications (copy number > 2). Code for recreating the karyoploteR output is available online (https://github.com/diyasen2021/copynumber.R).
Results
A chromosome-level genome assembly of Phytophthora agathidicida 3770
The genome assembly
The previous genome assembly of P. agathidicida 3770 (Studholme et al., 2016; BioProject PRJNA290659) had 3,689 scaffolds and an estimated genome size of 37,328,500 bp. Here, we assembled the genome sequence into ten complete chromosomes with a genome size of 56,996,300 bp and a read depth of >100x (Figure 1A). To produce this chromosome-level genome assembly, PacBio reads were assembled with a combination of complementary algorithms to yield four whole chromosomes (telomere to telomere) and 14 chromosome fragments with one or no telomeres. Hi-C (chromatin conformation capture) reads and inspection of multiple long reads containing sequences from the ends of these fragments were then used to assemble the 14 chromosome fragments into six more chromosomes. Finally, Illumina reads were used to polish the sequence.
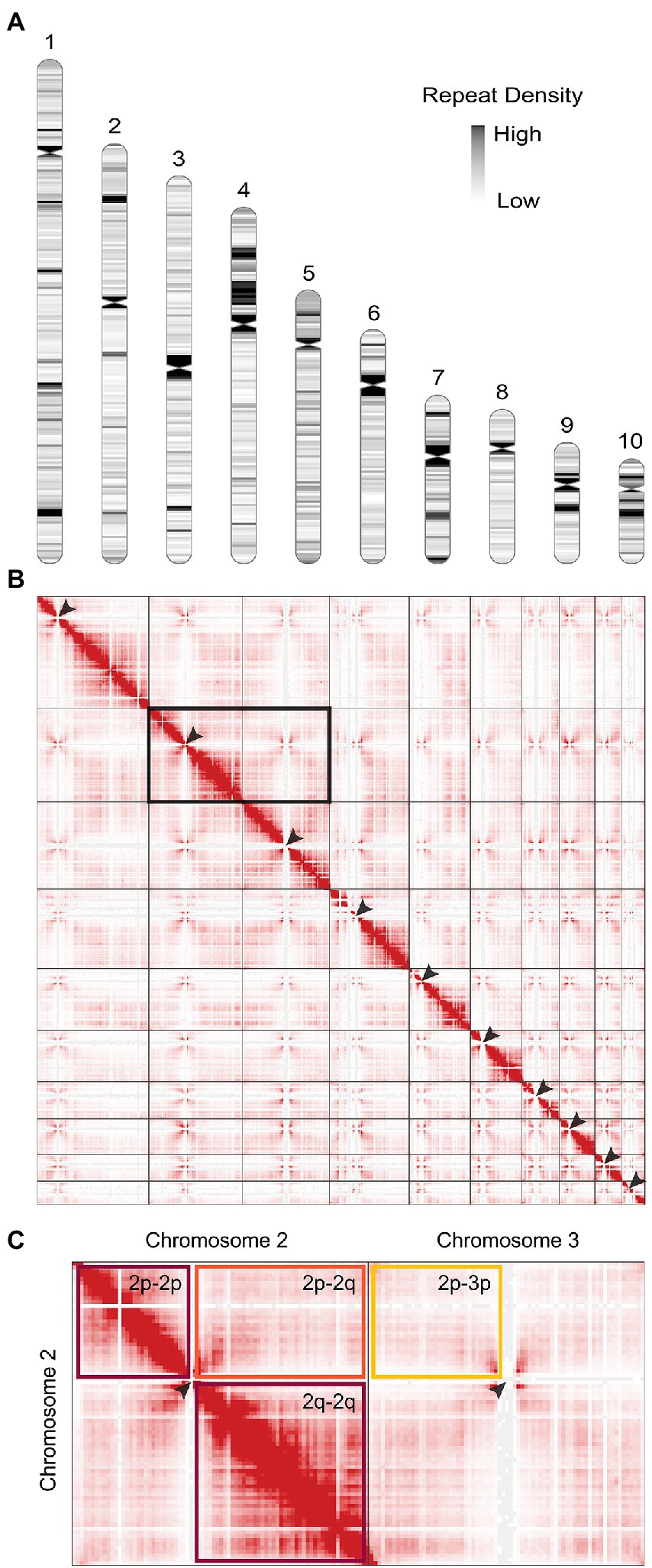
Figure 1. The genome of Phytophthora agathidicida. (A) Karyotype of the P. agathidicida 3770 genome, indicating centromere locations (pinched-in regions) and density of repeat elements across the ten complete chromosomes, although the full rDNA array is not depicted in chromosome 10. (B) Contact map of 3D DNA interactions within the P. agathidicida nucleus (red stippling). Arrows mark Hi-C patterns characteristic of centromere locations. (C) Enlarged example region of the DNA contact map shown in B. DNA interactions are frequent within the arms of each chromosome (maroon boxes), whilst interactions between chromosome arms are much reduced (orange box) and similar to interaction levels between different chromosomes (yellow box).
BUSCO analysis of the P. agathidicida genome assembly was 100% (complete and single copy genes) based on the Stramenopile lineage gene sets, indicating high accuracy of the assembly. This was a higher score than those for other oomycete genome assemblies based on long-read sequences, including those for Pe. effusa, P. ramorum and P. sojae (Supplementary Figure S1). Similarly, using the Eukaryota gene set, the complete and single copy BUSCO score for the P. agathidicida genome was higher than those for Pe. effusa and P. sojae, but similar to those of P. ramorum (Supplementary Figure S1). The number of predicted gene models increased from 17,691 (fragmented assembly from short-read sequencing data) to 17,963 (chromosome-level assembly). A k-mer analysis with Genomescope (Supplementary Figure S2) suggests that P. agathidicida 3770 is diploid, with an estimated low heterozygosity level of 0.02%.
Positions of the centromeres on the ten chromosomes were indicated by the frequency of Hi-C interactions (Figures 1B,C). Only chromosome 3 is metacentric, 2 and 4 are submetacentric, whilst the others are acrocentric. The ten chromosomes vary in length from 2.2 Mb to 10.5 Mb with 52.9–53.6% GC content (Table 1). Chromosome 10 showed the highest repeat density (51.4%) and lowest gene density (Table 1), and chromosome 4 showed large blocks of repetitive sequences (Figure 1A) despite having a percentage repeats value similar to the genome mean. The ribosomal RNA gene array (rDNA) is at a single location on chromosome 10 and is estimated to contain 245 copies of the rDNA units (Supplementary Data S1), although the final reference assembly only includes two full repeats plus the two partial flanking units as outlined in the methods. Including the full 245 rDNA copies, chromosome 10 is actually ~4.7 Mb in length and the whole genome ~59.5 Mb. The mitochondrial sequence assembly was published previously (Winkworth et al., 2021; BioProject PRJNA738084).
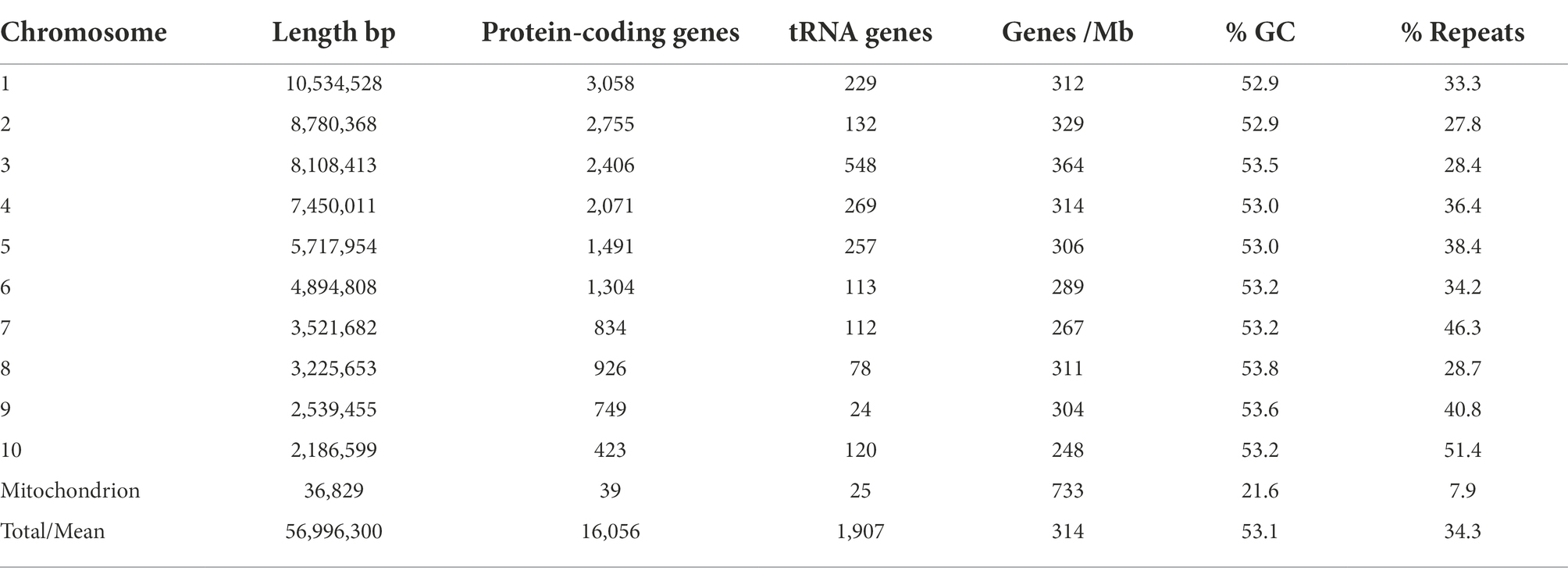
Table 1. Phytophthora agathidicida genome features by chromosome.
The assembled genome shows synteny to other assembled oomycete genomes
Comparative synteny analysis indicates substantial conservation of genome content between P. agathidicida, P. sojae and Peronospora effusa, with relatively little genome content unique to the three distinct species (Figure 2). Most of the conserved regions fall within long genomic blocks. Within the two completely assembled genome sequences, P. agathidicida chromosomes 7, 8 and 10 have essentially identical synteny with Pe. effusa across their entire length, despite being phylogenetically distant. This is suggestive of deep conservation of high-level genome organization, even though the DNA sequences themselves are divergent, as illustrated by low sequence identity (yellow and orange in the Figure 2 comparison plots). The comparative analysis also identifies species-specific genome rearrangements, such as an approximately 2 Mb inversion on P. agathidicida chromosome 3. Because the alternative non-inverted genomic structure is present in both P. sojae and Pe. effusa, the most parsimonious explanation is that this inversion occurred within the P. agathidicida lineage, with the ancestral state retained by the other two species. Given this high level of syntenic conservation, the chromosome-level genome of P. agathidicida may help guide potential scaffolding of more fragmented genome assemblies, as in this case for P. sojae.

Figure 2. Comparative genomics of the Phytophthora agathidicida genome. Synteny was determined between the ten chromosomes of P. agathidicida 3770 and the complete genome of Peronospora effusa UA202013 (top panel) and the fragmentary genome assembly of P. sojae P6497 (bottom panel). Matching colors indicate percent sequence identity.
Conserved centromere sequences and many unclassified repeats are present
Over a third of the P. agathidicida 3770 genome was identified as being comprised of repeat elements (Table 1). A de novo annotation of repeats in the genome (Supplementary Table S1; Supplementary Figure S3) showed that the most common repeats are Long Terminal Repeat (LTR) retrotransposons, in particular LTR/Copia (CoLT) retrotransposons (~7% of the genome). The locations of particularly long blocks of CoLT elements coincide with centromeric regions identified by Hi-C analysis (Figure 3); the same type of retroelement was also found in centromeres of P. sojae (Fang et al., 2020). In the P. agathidicida genome, there are many more classes of DNA repeat elements (21) than other characterised repeat classes such as LTRs and LINEs (12), so whilst the contribution of each type of DNA element is small (at most 2% of the genome), they are cumulatively substantial (Supplementary Figure S3). There was low sequence diversity within some classes of repeats, suggesting recent transposon activity in P. agathidicida (Supplementary Figure S4). However, a large proportion of the repeats could not be classified, suggesting that there might be many repeats unique to the Phytophthora genus.
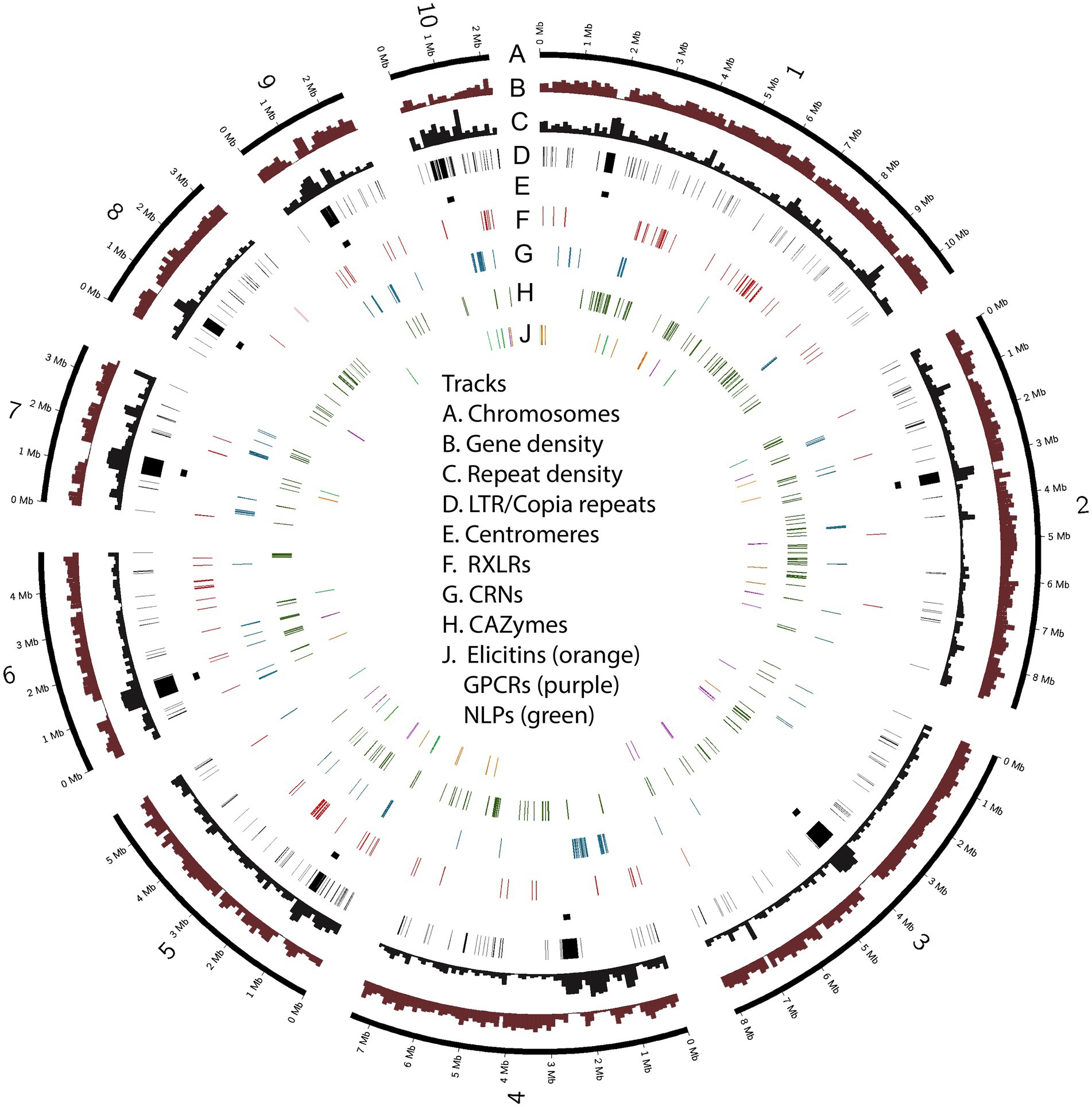
Figure 3. Chromosomes of Phytophthora agathidicida 3770. The circos plot illustrates the ten assembled chromosomes (numbered 1–10 on track A), along with densities of protein-coding genes and repetitive sequence (B, C); the positions of LTR/Copia (CoLT) repeats associated with centromeres (D, E) and genes in the six selected classes encoding predicted RXLR effectors, crinkling and necrosis proteins (CRNs), carbohydrate-active enzymes (CAZymes), elicitins, G-protein-coupled receptors (GPCRs) and necrosis-like proteins (NLPs) (F–J).
Gene distributions
Some key gene groups show a non-uniform distribution across the genome
Candidate genes from six classes that have been characterised as virulence factors in other Phytophthora species (Hua et al., 2013; Ma et al., 2015; Amaro et al., 2017; Chepsergon et al., 2020; Pirc et al., 2022) were selected for further investigation. These are genes encoding candidate cytoplasmic RXLR and CRN effectors, apoplastic effectors including elicitins, NLPs, CAZymes, and GPCRs involved in signaling (Supplementary Table S2). Compared to the previous fragmented genome assembly in which 78 candidate RXLR genes were identified, 147 RXLR genes were called in the new genome assembly, even though these RXLR candidates were identified using intersection of the same three prediction methods (Bhattacharjee et al., 2006; Whisson et al., 2007; Win et al., 2007). However, in the new genome assembly, ORFs as well as annotated gene models were searched for RXLRs, thus this modified method plus the more complete genome assembly account for the increased RXLR gene count.
The number of RXLR gene candidates (147) was similar to those found in some other Phytophthora species, such as P. chlamydospora (132; McGowan et al., 2020) and P. cactorum (135; Yang et al., 2018), but less than in many other species including P. ramorum (400; Carleson et al., 2022) and P. sojae (350; Tyler et al., 2006), although we had taken a conservative approach to RXLR identification by excluding RXLR-like genes from our numbers. In contrast, the P. agathidicida genome contained more CRN gene candidates (145) than most sequenced Phytophthora genomes except P. infestans (196; Haas et al., 2009). For example, in two well-assembled P. ramorum genomes, only 45 and 47 CRN gene candidates were identified using the same methodology as we used for P. agathidicida (Carleson et al., 2022).
Some of the gene classes showed an asymmetric distribution across chromosomes (Table 2; Figure 3). For example, 60% of the candidate RXLR genes and 64% of the candidate NLP genes were found on chromosomes 1 or 5, whilst chromosomes 3 and 8 had no NLP genes and very few candidate RXLR, CRN and elicitin genes. For chromosome 10, despite its smaller size (2.2 Mb), lower gene density and higher repeat density compared to the other chromosomes (Table 1), there were representatives from all the studied gene groups and also a higher proportion of SNPs among a set of New Zealand P. agathidicida isolates, particularly in exons (Table 2).
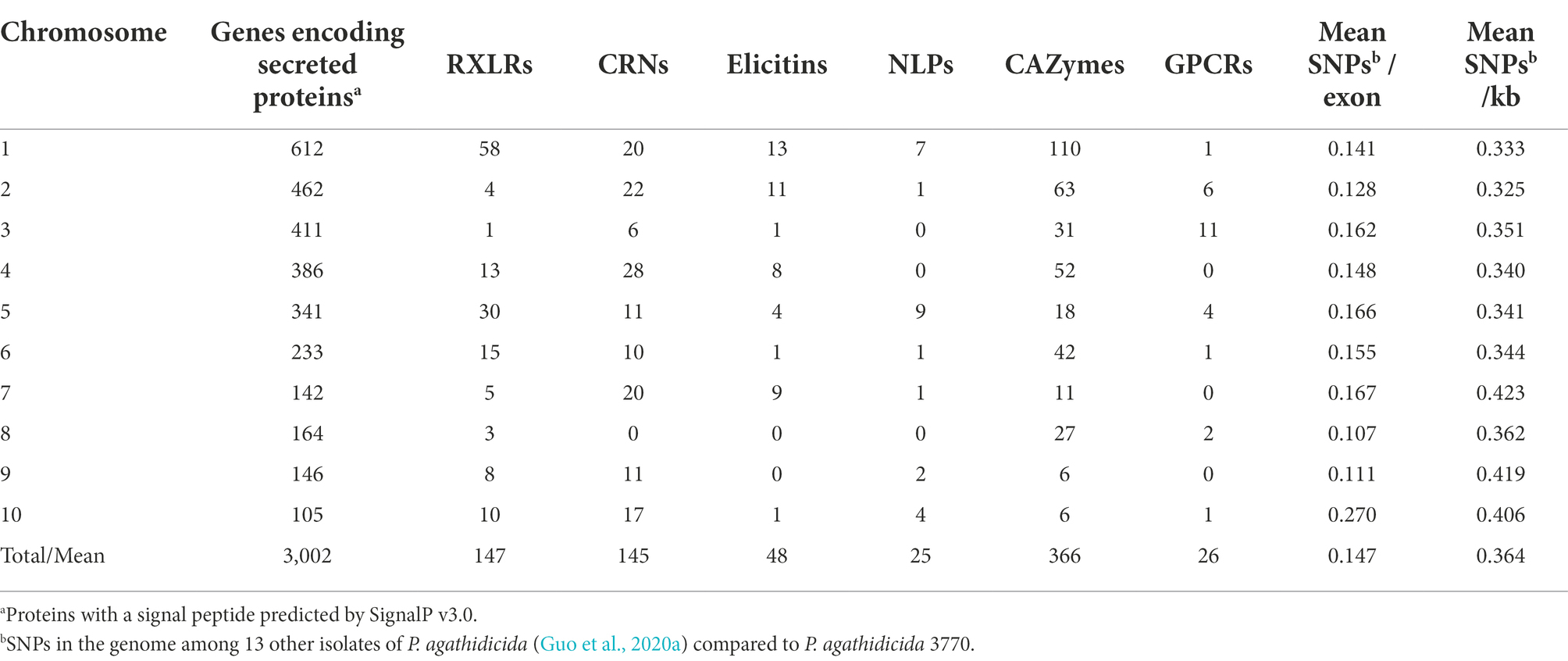
Table 2. Phytophthora agathidicida candidate gene groups and single nucleotide polymorphisms (SNPs) by chromosome.
A characteristic often noted in other filamentous plant pathogens, including Phytophthora species, is a compartmentalized genome in which genes involved in virulence or pathogenicity tend to be located in gene-sparse genomic regions enriched with repeats or transposable elements (Dong et al., 2015). As a consequence, effector genes tend to have longer intergenic distances to their neighboring genes when compared to other types of genes. To test whether P. agathidicida 3770 has this kind of compartmentalized genome, 5′ and 3′ distances to the nearest gene were calculated for genes in the six classes of interest: CAZymes, CRNs, elicitins, GPCRs, NLPs and RXLRs (Figure 4). The median 5′ and 3′ distance to the nearest gene is 512 bp for all genes identified in P. agathidicida 3770. Four gene classes show significantly larger median distances than this background rate: CRNs (11,835 bp), RXLRs (2,549 bp), NLPs (1,724 bp) and CAZymes (713 bp; all p < 1 × 10−4). Distances do not differ significantly for elicitins (643 bp, p = 0.74) or GPCRs (495 bp, p = 0.45).
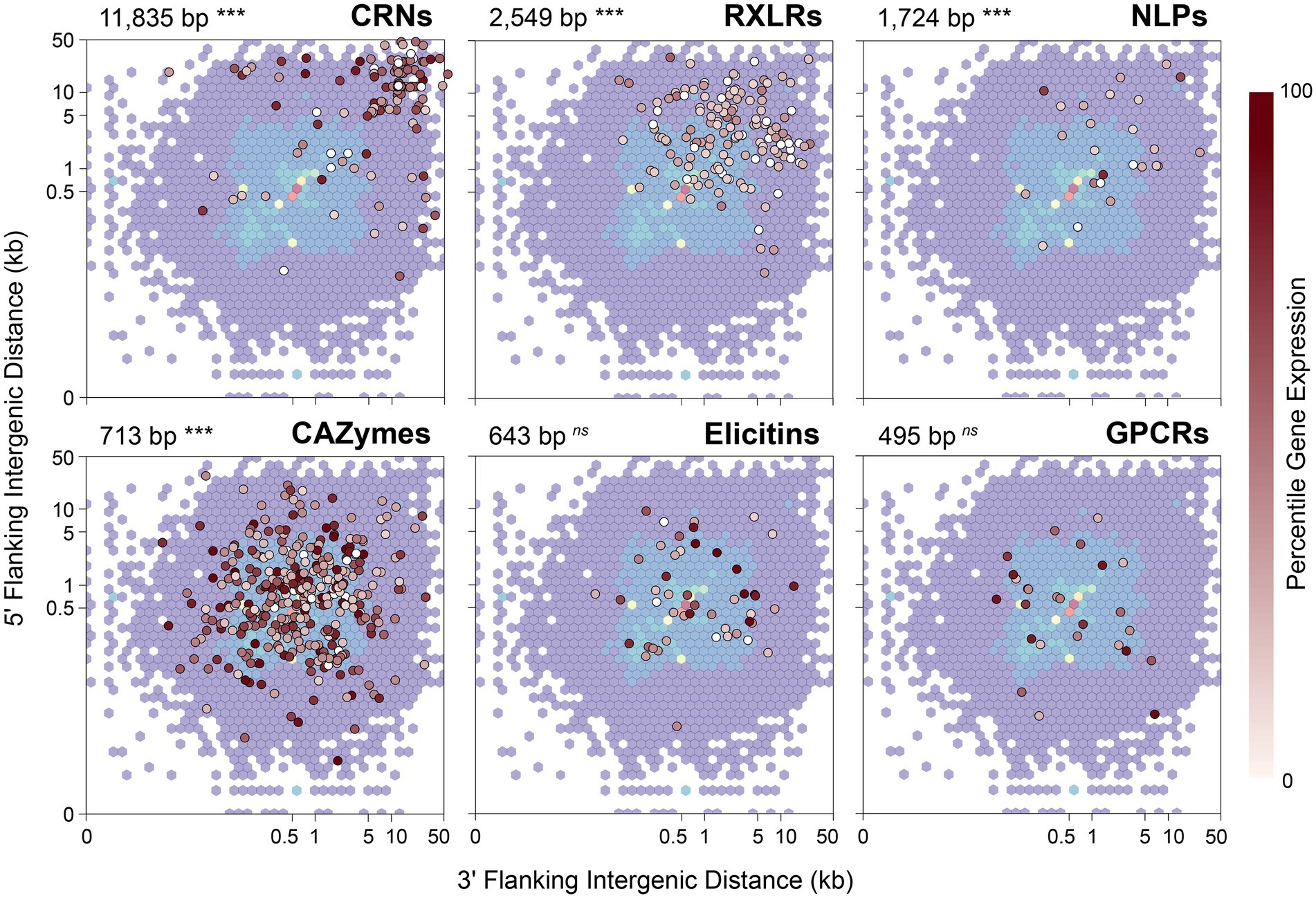
Figure 4. 5′ and 3′ distances to the nearest gene in the Phytophthora agathidicida genome. 5′ and 3′ distances for all genes identified in the P. agathidicida 3770 genome are shown by the background density distribution, which is overlaid in each panel with intergenic distances for genes in six gene classes: RXLRs, CRNs, NLPs, CAZymes, elicitins and GPCRs. Points for individual genes are colored by their percentile expression level in culture, with no expression indicated in white, low expression by lighter reds and high expression by darker reds. The median intergenic distance and its statistical significance are also shown for each gene class. ***p < 0.001; ns, not significant.
We considered that distances to flanking genes might influence gene expression. RNA extracted from the same P. agathidicida samples as used for Hi-C analysis was analysed by RNA sequencing and genes then classified by expression level percentile. However, no association was observed visually between intergenic distances and percentile levels of gene expression (Figure 4).
3D Genome clustering within gene classes reflects 2D arrangements on chromosomes
All six gene classes (CAZymes, CRNs, elicitins, GPCRs, NLPs and RXLRs) have 3D interaction profiles that differ significantly from gene sets chosen randomly from across the genome. CAZymes, elicitins, GPCRs, NLPs and RXLRs have larger numbers of 3D contacts (3- to 34-fold increase over a random gene set, all adjusted p < 0.001), indicating that on average these genes cluster more than average in the nucleus. Conversely, CRN genes exhibit fewer contacts (25-fold decrease) and therefore are less clustered in 3D space than average (adj p = 0.0006). As noted earlier, CRNs have markedly elevated 5′ and 3′ distances to the nearest gene along the chromosome (11,835 bp vs. 512 bp for all genes), reflecting their typical locations in repeat-rich and gene-poor regions of the genome. This physical isolation of CRN genes is probably associated with their greater isolation in 3D space.
Closer examination of genes in these six classes identified that many are physically adjacent along the genome, likely indicating that they arose through gene duplication, possibly followed by neofunctionalization. Genes that sit adjacent on a chromosome are by definition also close in 3D space, so we considered that the observed 3D clustering might simply be an artifact of the chromosomal co-location of many of these genes. To control for this feature, we excluded all genes that lay within 10 kb of any other gene in the same gene class and re-ran the analysis. After controlling for gene adjacency, the 3D clustering pattern disappears for elicitins, NLPs and RXLRs (all adj p > 0.05), but still holds, although more weakly, for GPCRs (6-fold increase in contacts over expectations, adj p = 0.036) and CAZymes (0.6-fold increase in contacts, adj p = 0.0002). The reduced 3D interaction of CRN genes is still strongly supported (21-fold decrease in contacts, adj p = 0.0006).
A complete genome offers the potential for deeper insights into gene expression and adaptation
As well as providing a chromosome-level perspective of the genes and other genome features, a complete genome assembly based on a high coverage with very long read sequences provides greater confidence in gene organisation in regions where there are tandem repeats. We present here several examples to illustrate the possibilities for further analysis of a fully assembled genome.
Visualising genetic diversity in copy number variants across the genome
In previous work, the genetic diversity of P. agathidicida among a collection of isolates from various locations in New Zealand was shown to be very low (~0.1% sequence diversity), indicating a near-clonal population (Guo et al., 2020a). However, an assessment of copy number variation (CNV), based on re-mapping of Illumina reads from the different isolates (including Pa3770) onto the complete P. agathidicida 3770 genome assembly (Supplementary Table S3), suggested there is some structural diversity between the isolate genomes, with partial aneuploidy potentially providing increased or decreased gene copy numbers in those regions (Figure 5). While there appear to be some CNVs that are common to many of the isolates, others stand out as being unique; for example, the higher copy number of chromosome 10 of P. agathidicida 3770. This highlights the value of a complete genome assembly for discerning structural genome diversity.

Figure 5. Copy number variants (CNVs) across the genomes of 14 isolates of P. agathidicida based on short read sequencing data. The rectangles at the bottom represent the ten chromosomes of P. agathidicida 3770. Tracks above depict copy number variants, where blue rectangles indicate duplications and red rectangles indicate deletions. P. agathidicida isolate numbers are displayed on the right.
Gaining a genome-wide view of gene expression
A complete genome assembly also offers the opportunity to re-map previously obtained gene expression sequence reads to provide a genome-wide perspective. Unpublished RNA-seq data of P. agathidicida isolate 3813 (BioProject PRJNA864064), grown in kauri leaves and roots over a time course of infection, were mapped to the new P. agathidicida 3770 gene models. Isolate P. agathidicida 3813 is very similar in sequence to P. agathidicida 3770; between these two isolates, only 10,435 SNPs were identified across the whole genome, with just 2,140 of these falling in gene-coding regions, providing confidence in the accuracy of mapping P. agathidicida 3813 reads onto the P. agathidicida 3770 genome. Levels of gene expression were evaluated for genes in the six gene classes mentioned above. Figure 6 shows the positions of these genes across the genome and their maximum expression values (at any time point) in planta. From this genome-wide perspective, the positions of hotspots of gene expression can be determined for several of the gene groups. For example, there are clusters of highly expressed CAZyme and RXLR genes on chromosome 1, elicitin genes on chromosome 7 and a region of high CRN gene expression on chromosome 4. Supplementary Figure S5 shows a similar plot except showing positions of genes that are significantly up-regulated in planta compared to in culture.
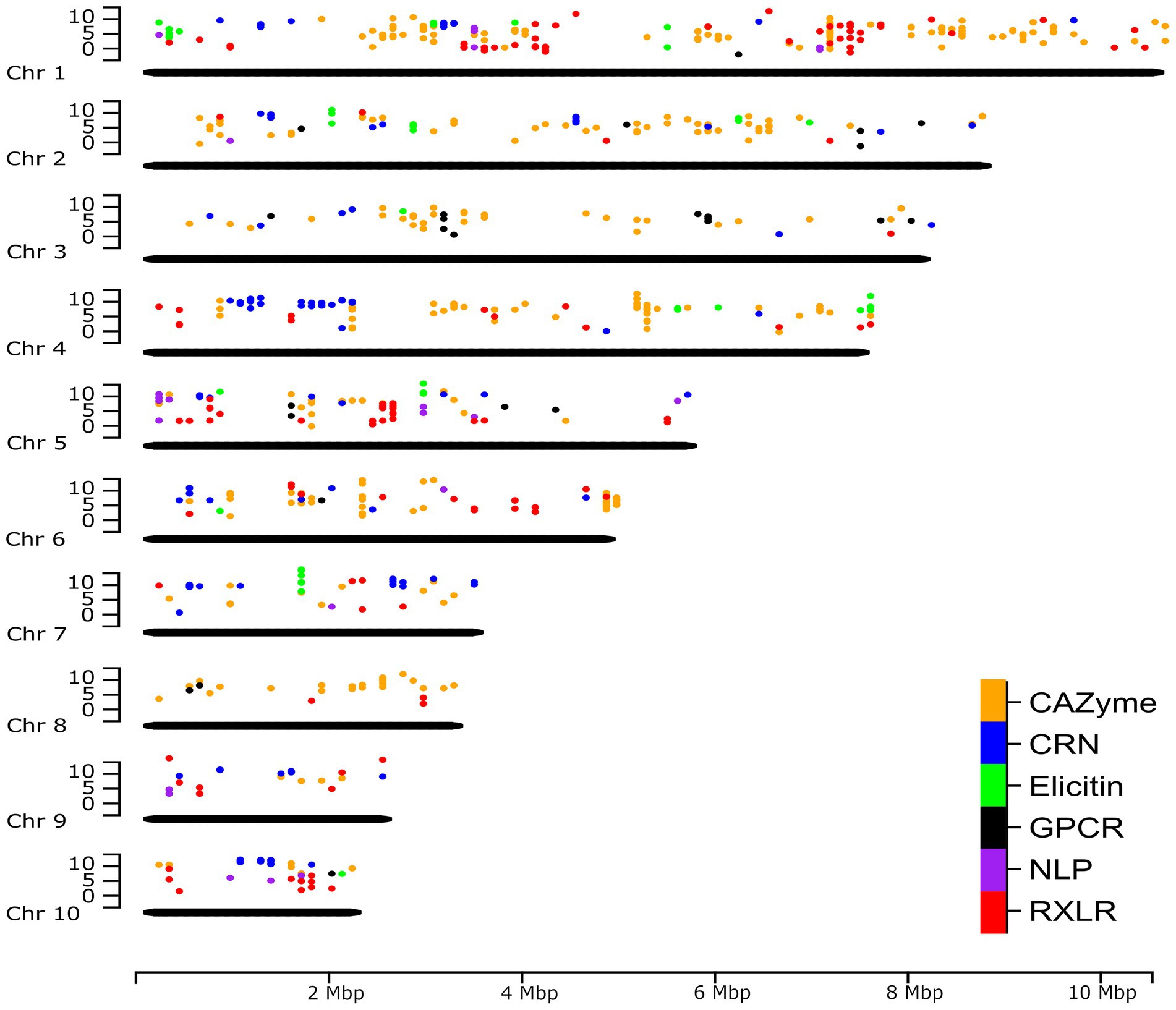
Figure 6. Genome-wide expression of putative effectors and other key gene classes. The positions of genes in six key gene classes in the P. agathidicida 3770 genome and the maximum expression value (log2 fragments per kilobase of exon per million mapped fragments [FPKM]) of those genes across a time course of expression on kauri roots at 6, 24, 48 and 72 h post-inoculation. Chr, chromosome.
Examining association of transposable elements with multi-copy CRN genes
In P. infestans, many of the large number of CRN genes are in repeat-rich regions and it was suggested that expansion of CRN gene families was facilitated by non-allelic homologous recombination involving repeats (Haas et al., 2009). A completely assembled genome can provide a clear picture of all or most of the repetitive elements and their locations with respect to protein-coding genes, facilitating studies of repeat-driven adaptation.
Although P. agathidicida has a relatively large number of CRN genes, many of them are grouped in gene clusters of various sizes (Figure 3; Supplementary Table S2). The clustered genes occur most often in repeated head-to-tail orientation, suggestive of tandem gene duplication events, sometimes with one or more other types of genes separating them. In an interesting example, 13 adjacent CRN genes in a cluster on chromosome 2 are separated from each other by repeating sets of repeats including DNA/Maverick and LTR/Gypsy transposons (Figure 7). Maverick repeats are in the same class of DNA transposons as helitrons, which were reported as being closely associated with a similar CRN gene cluster in P. infestans (Haas et al., 2009). CRNs are modular proteins with a recombination hotspot between the N and C termini at a conserved HVLVXXP motif (Haas et al., 2009; Amaro et al., 2017). The 13 clustered P. agathidicida CRN genes fall into three sub-groups based on differences in their predicted N- and C-terminal domain sequences (Figure 7; Supplementary Figure S6). These shared repeats and protein domain swaps suggest the cluster has originated via recombination and duplication events, a potential sequence of which is shown in Supplementary Figure S7. The nine identical genes (CRN31-39) labelled ‘AG’ in Figure 7 had maximum in planta expression values of only 40–65 FPKM (fragments per kilobase of exon per million mapped fragments), whilst the ‘AF’ (CRN27-28) and ‘BF’ (CRN29-30) types all showed higher expression values (between 196–273 FPKM; Supplementary Table S4).
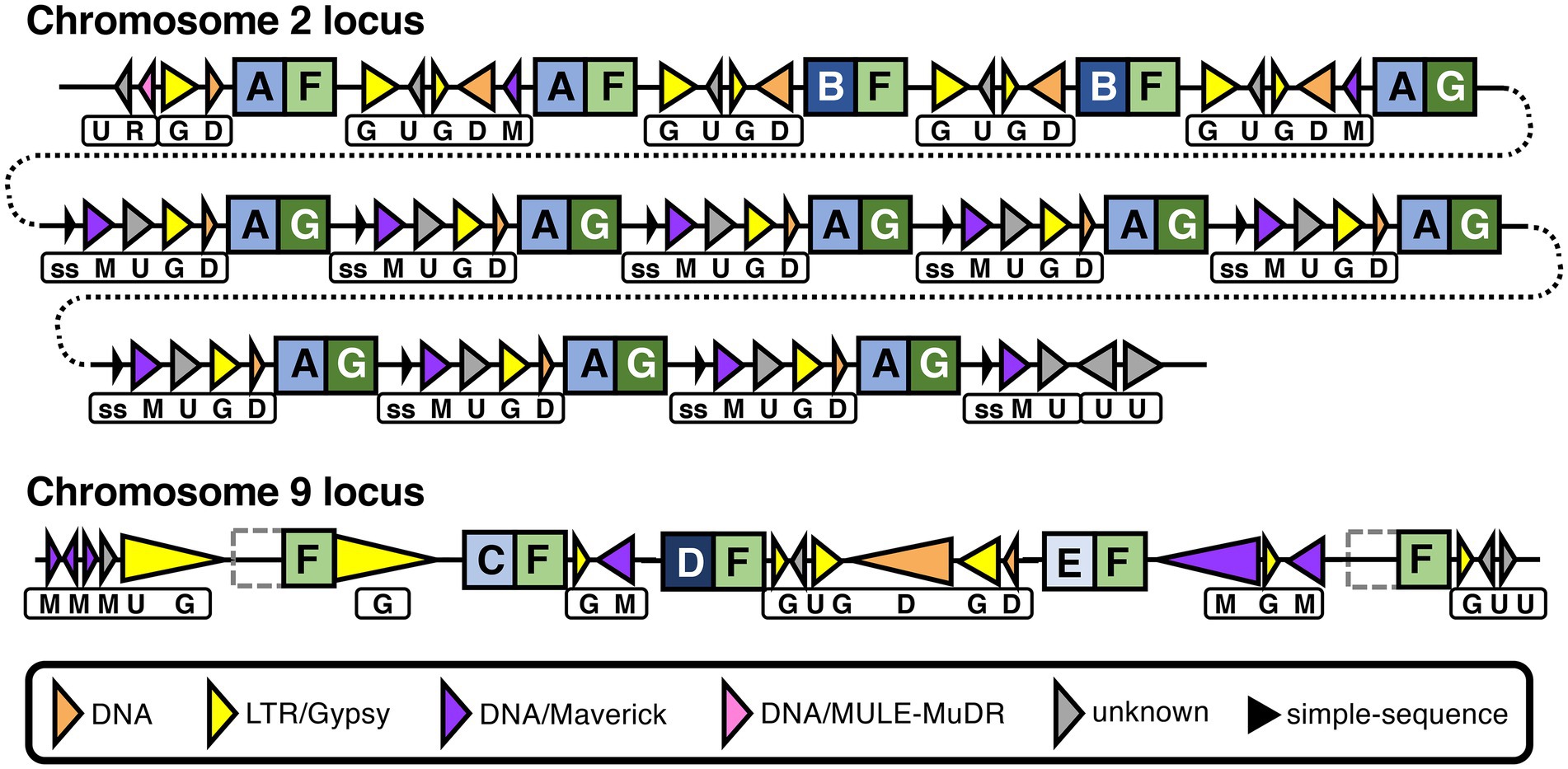
Figure 7. Recombination and repeats in two CRN gene clusters in the Phytophthora agathidicida genome. Genomic organization of CRN gene repeat clusters on chromosomes 2 and 9. CRN genes are indicated by square boxes with letters inside. N and C termini of predicted CRN proteins are indicated by blue and green boxes, respectively, with different shading and letters indicating different variants. Repeats are indicated by triangles, with transposons depicted as colored triangles and simple-sequence (ss) repeats as smaller, black triangles, as shown in the key. Triangle direction refers to the strand each repeat is encoded on. The different combinations of repeats between CRN genes are also indicated beneath by boxes with letters. The chromosome 2 cluster is shown over multiple lines (joined by dotted lines) for ease of visualization. In the chromosome 9 cluster, the left- and right-most CRN genes both encode proteins that lack the N-terminal domain, as indicated by grey dotted boxes. Diagram is not to scale, although large differences in repeat length are indicated schematically by small differences in triangle length.
On chromosome 9, a set of three adjacent CRN genes (CRN119-121) are all different from one another in the predicted N-terminal region (Figure 7; Supplementary Table S4; Supplementary Figure S6) but were predicted to have identical C termini apart from one amino acid. This is again suggestive of recombination at the same CRN gene hotspot. Moreover, these genes are interspersed with similar repeat elements as found between the 13 CRN genes on chromosome 2, and they also share an identical C-terminal amino acid sequence (F) with those from some of the chromosome 2 CRNs. Indeed, a total of 31 CRN genes in the genome encode proteins that have a C-terminus with at least 99.8% amino acid identity to what we have denoted as type F, along with another 12 genes that were not called as CRNs but appear to be associated with CRN gene clusters (Supplementary Table S5). Two of these 12 additional genes were found flanking the three-CRN gene cluster on chromosome 9; the upstream gene (CRN-like1) encodes a protein that lacks the complete N-terminal region and 100 amino acids of the C-terminus, while the downstream gene (CRN-like2) encodes a protein that lacks almost all of the N-terminal region (Supplementary Table S4). All five CRN and CRN-like genes on chromosome 9 shown in Figure 7 had maximum in planta expression values between 224 and 292 FPKM (Supplementary Table S4). Together these results suggest both intra- and inter-chromosomal recombination and duplication events have occurred in the P. agathidicida genome and that in some cases gene expression is affected.
Seeking evidence for adaptation in multi-copy RXLR genes
Of the 147 candidate RXLR genes in the P. agathidicida 3770 genome, 30 occur in one of 11 families with identical or near-identical genes and are located on chromosomes 1, 5, 6 or 9 (Table 1; Supplementary Table S6). Most of these gene families consist of two or more identical genes and many have other genes, sometimes a different RXLR gene, between them (Table 1). The largest family, on chromosome 1, is a set of nine tandemly repeated genes (RXLR29-1 to 29–9) which are identical in sequence, suggesting recent duplication events. In the previous genome annotation, only one of these nine RXLR29 genes was annotated (Guo et al., 2020a), highlighting the value of a fully-assembled genome for identifying families containing identical genes. Between each of the RXLR29 genes is a repeated sequence of an unknown repeat class (Supplementary Table S1). RXLR29 was previously shown to trigger plant cell death when delivered to Nicotiana tabacum by Agrobacterium tumefaciens-mediated transient transformation, suggesting it is recognised by immune receptors in the plant (Guo et al., 2020a). Curiously, based on the gene expression studies outlined in Section “Gaining a genome-wide view of gene expression,” none of these nine genes are expressed in planta or in culture.
Besides RXLR29, genes in six of the 11 other RXLR gene families also appear to be silenced, including gene families RXLR4, 11, 36, 46, 120 and 144, located on chromosomes 1, 5, 6 and 9 (Table 3; Supplementary Table S6). In contrast, the RXLR106 family, which consists of two identical genes, appears to be among the most highly expressed of the RXLR genes; this is the only identical RXLR gene pair that is located on two separate chromosomes (1 and 9), although, based on RNA-seq read mapping, it is not possible to tell whether only one or both of these identical genes are actually expressed. However, control of individual gene expression is also important, as seen on chromosome 9 where one of the most highly expressed singleton RXLR gene candidates (RXLR53) is located less than 3 kb from a CRN gene (CRN128) which is expressed at very low levels (Supplementary Table S2). A complete genome assembly offers great potential to study gene regulation on a genome level and the roles of certain classes of genome repeat elements and epigenetic factors provide a rich area for discovery.
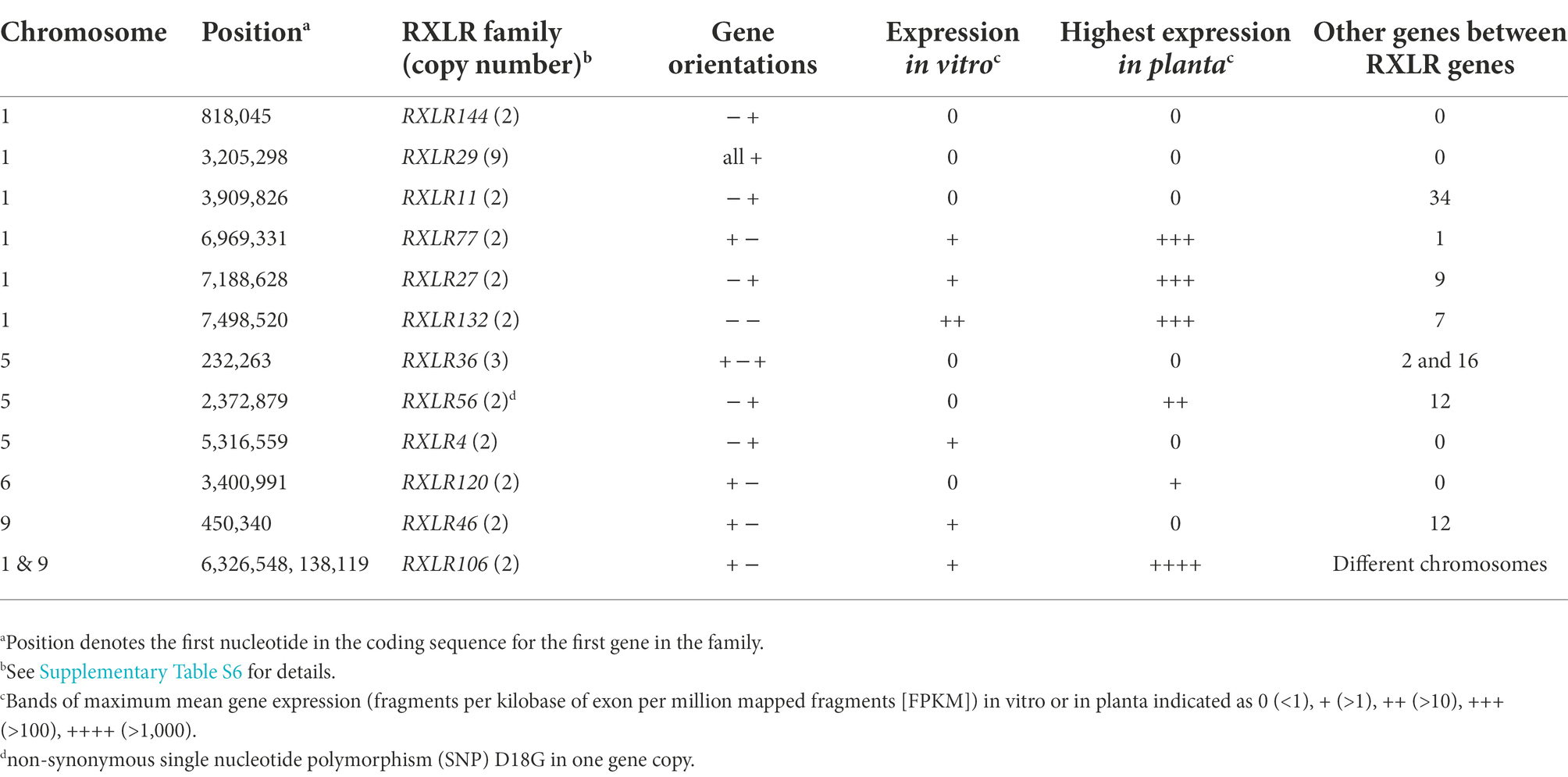
Table 3. Candidate RXLR gene families in the Phytophthora agathidicida genome.
Discussion
In this study, we combined data from long-read PacBio sequencing and Hi-C chromatin conformation capture to assemble the genome of the kauri dieback pathogen Phytophthora agathidicida to chromosome level. The assembly of the genome into ten chromosomes is a considerable improvement over the previous assembly of 3,689 scaffolds, which was derived using short-read Illumina sequences (Studholme et al., 2016). The newly generated genome has an estimated size of 57 Mb and appears to be accurate based on the high (100%) BUSCO score obtained using the genome assembly and the Stramenopile lineage gene set. The genome size is thus 20 Mb larger than that predicted in the previous assembly based on short read sequencing (Studholme et al., 2016). Because short-read assemblies often miss or collapse long repetitive regions, it seems likely that much of this extra 20 Mb comprises repeat sequences such as transposable elements. However, some members of gene families with identical or near-identical adjacent genes were also missed in the earlier assembly, such as the RXLR29 gene family.
Long-read technologies have been used for sequencing other Phytophthora genomes, including P. cactorum (Yang et al., 2018), P. capsici (Stajich et al., 2021), P. cinnamomi (Engelbrecht et al., 2021) and P. ramorum (Carleson et al., 2022), although none of these genomes were assembled to chromosome level and none of these authors reported using Hi-C data. In our work, whilst the Hi-C data enabled us to join a few sub-chromosomal scaffolds together, it was the availability of extremely high coverage long reads overlapping the junctions that primarily enabled us to complete the assembly. The Hi-C data did, however, confirm the overall assembly as well as indicate the centromere positions. The validity of the centromere positions was supported by the presence of LTR/Copia (CoLT) retrotransposon elements which have been associated with centromeres in other Phytophthora species (Fang et al., 2020).
Factors that likely influenced our ability to assemble the P. agathidicida genome to chromosome level include its relatively small genome size (57 Mb), low repeat content (34%), and very low heterozygosity (0.02%). The 57 Mb genome size of P. agathidicida is similar to that of P. ramorum (Carleson et al., 2022) but much smaller than those of P. capsici (94 Mb; Stajich et al., 2021), P. cinnamomi (107 Mb; Engelbrecht et al., 2021), P. sojae (95 Mb; Tyler et al., 2006) and P. infestans (240 Mb; Haas et al., 2009). In general, these differences in genome size reflect the repeat content (Engelbrecht et al., 2021) and the higher the repeat content the more challenging it is to assemble a complete genome. The heterozygosity level of 0.02% is considerably lower than those estimated for other Phytophthora species for which the same method was used, for example P. cinnamomi (1.36%; Engelbrecht et al., 2021), P. infestans (0.695%; Fletcher and Michelmore, 2018), P. chlamydospora (0.68%), P. gonapodyides (1.88%) and P. pseudosyringae (0.15%; McGowan et al., 2020).
Although chromosome-level assemblies of Phytophthora species are currently rare, repeat-rich genomes of fungal pathogens have been fully assembled using long-read and Hi-C sequence data. For example, the 67 Mb genome sequence of the fungal tomato pathogen Cladosporium fulvum (now called Fulvia fulva) was assembled to 14 chromosomes using these technologies (Zaccaron et al., 2022), representing a considerable advance over the 4,865 scaffolds of the earlier assembly, albeit from a different isolate of this species (de Wit et al., 2012). However, the 1 Gb genome sequence of the fungal myrtle rust pathogen Austropuccinia psidii, of which 91% is repetitive sequence, was assembled into 66 scaffolds rather than to chromosome-level (Tobias et al., 2021), highlighting the ongoing limitations to complete assembly imposed by repeats.
The P. agathidicida genome assembly provides a unique insight into how a Phytophthora genome is constructed. The high level of synteny with the chromosome-level assembly of the oomycete Peronospora effusa (Fletcher et al., 2021) and the 70-contig assembly of P. sojae (Fang et al., 2020) suggests conservation of high-level genome organisation, despite low sequence identity, and may assist in assembly of other oomycete genome sequences. Comparisons among the ten chromosomes of P. agathidicida revealed only minor variations in gene density, GC content and percentage of repeats, although some classes of genes, among the six classes we studied, were more prominent on some chromosomes than others. Based on our analyses, this asymmetric distribution of genes across the chromosomes was most likely due to physical clustering of genes in gene families. We found multicopy gene families encoding RXLRs and CRNs in P. agathidicida that most likely occurred due to gene duplication, in keeping with effector gene expansion observed in other Phytophthora species such as P. infestans (Haas et al., 2009), P. sojae (Tyler et al., 2006) and P. cactorum (Yang et al., 2018). This phenomenon can explain the irregular distribution of some of the gene classes on P. agathidicida chromosomes. For example, chromosome 10 has a relatively large number of CRN genes (17) for its small size but most of these occurred in two clusters of five and 11 genes. Chromosome 10 does, however, have some special features compared to the other chromosomes: lower gene density, higher repeat content (51%), increased copy number suggestive of partial aneuploidy and a higher proportion of SNPs among a set of New Zealand P. agathidicida isolates, particularly in protein-coding regions (Table 2). However, as chromosome 10 was present in all the other P. agathidicida isolates tested and because it contains the rDNA locus, it does not appear to be a dispensable or accessory chromosome. Such dispensable or accessory chromosomes are often found in fungi. This includes Fulvia fulva, where only five of 24 isolates examined were found to contain the smallest chromosome, chromosome 14 (Zaccaron et al., 2022).
The chromosome-level assembly of the P. agathidicida genome also provides high confidence for identifying gene locations relative to other features, including where there are multicopy genes in gene families and repetitive elements such as transposons. It is known that Phytophthora effector genes, such as RXLRs, are often found in gene-poor, repeat-rich regions of the genome, where the proximity of the repeats provides opportunities for rapid adaptation and evolution by processes such as gene duplication and recombination (Haas et al., 2009). An often-used method to assess the gene density surrounding certain gene classes is to assess the intergenic distance to the next gene. As found in P. cinnamomi (Engelbrecht et al., 2021), candidate CRN, RXLR and NLP genes of P. agathidicida were significantly enriched within gene-poor regions (Figure 4). The CAZyme genes showed a significant but relatively small median distance from other genes, perhaps influenced by the broad diversity of CAZyme classes we considered. Although we found evidence to support a compartmentalized genome in P. agathidicida, further work is required to determine if rates of evolutionary change differ between the two types of compartments, as was found in P. infestans (Raffaele et al., 2010).
Given the expansion and diversification of effector genes that occur in many Phytophthora pathogens (Haas et al., 2009; McGowan and Fitzpatrick, 2017), a deeper understanding of how these pathogens adapt to invade and subdue their hosts will require analysis of evolutionary processes occurring in multi-gene families. Where these gene families are comprised of tandem repeats of near-identical genes, high-quality genome assemblies based on long-read sequencing technology provide a clearer picture compared to fragmented assemblies from which some genes may be missing. As examples, we looked at regions in the P. agathidicida genome containing tandemly repeated CRN and RXLR gene family candidates.
P. agathidicida appears to have a relatively large number of CRN genes (145) which will form the basis for future studies. CRN proteins are of great interest in that they are highly conserved, being found in all plant-pathogenic oomycetes studied so far (Amaro et al., 2017). CRNs tend to have a conserved N-terminal domain with two conserved motifs (LXLFLAK and HVLVVVP) but a variable C-terminal functional domain which denotes their classification (Zhang et al., 2016). Most CRNs are expressed both in culture and in planta (Haas et al., 2009; Amaro et al., 2017). In the case of P. capsici in tomato, there were two patterns of gene expression with (a) upregulation in both early and late stages of infection and (b) a gradual increase from early to late stages (Stam et al., 2013). Despite their name, not all CRNs induce cell death in plants and some can suppress cell death (Liu et al., 2011; Amaro et al., 2017).
In the P. agathidicida genome, transposable DNA elements (such as DNA/Maverick), were associated with clusters of CRN genes, along with LTR/Gypsy and other repetitive elements. Some types of CRN genes were likewise associated with DNA elements (helitrons) in P. infestans (Haas et al., 2009). Helitrons duplicate by rolling circle replication and can mediate genome evolution by promoting duplication and recombination (Kapitonov and Jurka, 2007), leading to the suggestion that CRN evolution in P. infestans is mediated by helitrons (Haas et al., 2009). A gypsy retrotransposon was also found in the C-terminal domain of a CRN-coding gene, PITG_23144, in P. infestans (Haas et al., 2009). Inspection of two clusters of CRN genes in the P. agathidicida genome provided evidence for recombination, including domain swaps, at the ‘hotspot’ junction of the N- and C-terminal domains. One of the CRN gene clusters contained several tandem identical repeats, indicative of recent duplication events, whilst the other cluster was flanked by CRN-like genes that lacked most or all of the N-terminal domain. In addition to recombination/duplication, our results also suggest recent transposon activity in P. agathidicida based on low divergence of transposon sequences (Supplementary Figure S4), as also shown in P. infestans (Haas et al., 2009). A more than three-fold difference in gene expression was seen between different types of recombinant CRN paralogs in the larger P. agathidicida cluster we investigated, suggesting a possible functional consequence of transposon-mediated recombination. The evolution of CRN genes and the roles of transposable and other repetitive elements in this process is a fascinating area for further study. Expansion of CRN genes appears to be associated with species that include necrotrophy in their life cycle (Stam et al., 2013). Due to the hemibiotrophic nature of most Phytophthora species (Boevink et al., 2020), the CRN genes may be a focal point for adaptation in these pathogens, including P. agathidicida.
Almost twice as many RXLR genes (147) were identified in the new P. agathidicida genome assembly compared to the previous fragmented assembly (78). RXLRs are the best studied among all the effectors of Phytophthora species (Anderson et al., 2015; Deb et al., 2018; Wang et al., 2019a; Chepsergon et al., 2021) and are often the largest class of intracellular effectors in Phytophthora pathogens (Boevink et al., 2020; Midgley et al., 2022). RXLR proteins function in many different regions of the cell and have many different roles (Boevink et al., 2020; Midgley et al., 2022). Among these, the ability of some RXLRs to induce or suppress plant cell death are well known (Deb et al., 2018). For example, in P. infestans the RXLR effector Avr3A elicits cell death in potato when recognised as an avirulence factor (Armstrong et al., 2005). In our earlier work on P. agathidicida, we identified eight RXLR candidates that elicited cell death in the non-host Nicotiana spp. and one (PaRXLR40) that suppressed RXLR-induced cell death (Guo et al., 2020a). One of those cell-death eliciting RXLR candidates, RXLR29, was encoded by a gene that formed part of a cluster of nine identical genes when we examined the new chromosome-level genome assembly. Thus the increased RXLR numbers in the new P. agathidicida genome assembly are partly accounted for by multicopy gene families that were undetected in the earlier fragmented genome.
The nine gene copies in the P. agathidicida cell-death eliciting RXLR29 cluster are separated by repeats of an unknown class that could potentially have facilitated gene duplication. However, whilst gene duplication and divergence are known as drivers of adaptation (Goss et al., 2013), none of the nine RXLR29 copies were expressed during infection of kauri. Similarly six of the 11 other RXLR gene clusters in the P. agathidicida genome were silenced or expressed at very low levels (Supplementary Table S6). Silencing of effector gene expression is a well-established adaptation mechanism whereby pathogens can avoid recognition by a plant host, which could otherwise provoke a defence response, often including localised cell death (Qutob et al., 2013; Wang et al., 2019b, 2022). In contrast, the nine identical and four near-identical CRN genes on chromosome 2 (Figure 7) were clearly expressed (Supplementary Table S4), supporting the reliability of our RNA-seq mapping method to determine expression in identical multi-copy genes.
In agricultural systems, the identification of effectors that elicit host defence responses has enabled the screening and breeding of resistant cultivars (Vleeshouwers and Oliver, 2014; Chepsergon et al., 2021). Effectors of gymnosperm pathogens, both fungal and oomycete, have been shown to elicit defence responses in model angiosperm hosts, suggesting similar mechanisms in angiosperm and gymnosperm plant-pathogen interactions (Chen et al., 2015; Guo et al., 2020a,b; Hunziker et al., 2021; Tarallo et al., 2022). Gymnosperm trees such as conifers have diverse sets of immune receptors, some of which respond to drought as well as to pathogens (Van Ghelder et al., 2019). A recent study showed that the defence responses of disease-tolerant Pinus contorta included increased expression of genes in PAMP- and effector-triggered immunity pathways as well as of certain immune receptors in response to infection with the fungal pathogen Dothistroma septosporum (Lu et al., 2021). Thus, the study of effectors in oomycete forest pathogens has the potential to identify resistance traits in gymnosperm trees such as kauri. However, pathogens can quickly adapt by silencing, mutating or deleting their genes. In Phytophthora species, insights into adaptation of virulence traits have been gained by studying chromatin-level modification of gene expression (Wang et al., 2020b), diversifying selection (Zhang et al., 2019) and loss of heterozygosity (Lamour et al., 2012; Dale et al., 2019). A focus on core effectors that elicit defence responses offers the promise of more durable resistance; these are effectors that are highly conserved, highly expressed in planta and essential for virulence or pathogen growth such that loss of function would be detrimental to the pathogen (Chepsergon et al., 2021).
In some cases RXLRs that elicit cell death at a later stage of infection, such as P. capsici PcAvh1 (Chen et al., 2019), might facilitate the transition from biotrophy to necrotrophy. In contrast, P. sojae PsAvh52 suppresses cell death in early stages of infection in soybean and is required for full virulence (Li et al., 2018). Thus some RXLRs play a key role in orchestrating the pathogen lifestyle during both biotrophic and necrotrophic stages. The study of RXLRs with different roles such as these in P. agathidicida is likely to reveal important virulence factors that could be targets for disease control.
In the case of trees such as kauri that can live for over a thousand years, it would be naive to think that a single mechanism will achieve durable resistance. Disease resistance or tolerance needs to encompass a diversity of mechanisms and function within a broader ecological setting that includes the effects of soil, companion plants and, importantly, potential protective effects of the microorganisms that naturally live in, on and around the tree (i.e., the host holobiont) (Pautasso et al., 2015; Padamsee et al., 2016; Lewis et al., 2019; Terhonen et al., 2019; Wang et al., 2019b; Byers et al., 2020; Wang et al., 2020a). However, by increasing our knowledge of the molecular interactions between Phytophthora pathogens and their hosts, in a broad ecological setting, we stand a better chance of protecting those plant hosts against the devastating effects of disease. The complete genome sequence of P. agathidicida provides a valuable stepping stone towards a better understanding of these plant destroyers.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, PRJNA734652; https://www.ncbi.nlm.nih.gov/, PRJNA864064.
Author contributions
RB, MC, RM, NG, NW, and CM conceived and guided the study. YG, DW, DS, NC, EB, AG, MG, RL, PP, JS, RB, and MC designed and performed experiments and analyzed data. RB and MC led manuscript writing. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the New Zealand Ministry of Business, Innovation and Employment (Ngā Rākau Taketake – Myrtle Rust and Kauri Dieback Research, grant number C09X1817, and Healthy Trees Healthy Future Programme, grant number C04X1305) and the New Zealand Tertiary Education Commission (Bioprotection Aotearoa, grant number 39504).
Acknowledgments
Kauri germplasm was provided via Scion with permission from Taoho Patuawa on behalf of the Te Roroa Iwi Trust. This project was completed with cultural authority from Te Roroa, the mana whenua from the Waipoua region, and from the Ngāti Rehua Ngātiwai ki Aotea Trust from Aotea/Great Barrier Island, from where samples were obtained for this work. Elena Hilario (Plant and Food Research, Auckland NZ) is thanked for advice on chromatin conformation capture (Hi-C) library preparation.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1038444/full#supplementary-material
Footnotes
1. ^https://github.com/PacificBiosciences/IsoSeq
2. ^https://grunwaldlab.github.io/annot_toolbox/complement_gff/
3. ^https://mpcox.github.io/2D_FIR_distribution
4. ^https://mpcox.github.io/3D_contact_distribution
References
Ahmed, M., and Ogden, J. (1987). Population dynamics of the emergent conifer Agathis australis (D. Don) Lindl.(kauri) in New Zealand I. Population structures and tree growth rates in mature stands. N. Z. J. Bot. 25, 217–229. doi: 10.1080/0028825X.1987.10410068
Amaro, T., Thilliez, G. J. A., Motion, G. B., and Huitema, E. (2017). A perspective on CRN proteins in the genomics age: evolution, classification, delivery and function revisited. Front. Plant Sci. 8:99. doi: 10.3389/fpls.2017.00099
Anderson, R. G., Deb, D., Fedkenheuer, K., and McDowell, J. M. (2015). Recent progress in RXLR effector research. Mol. Plant-Microbe Interact. 28, 1063–1072. doi: 10.1094/MPMI-01-15-0022-CR
Armstrong, M. R., Whisson, S. C., Pritchard, L., Bos, J. I., Venter, E., Avrova, A. O., et al. (2005). An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proc. Natl. Acad. Sci. U. S. A. 102, 7766–7771. doi: 10.1073/pnas.0500113102
Bao, Z., and Eddy, S. R. (2002). Automated de novo identification of repeat sequence families in sequenced genomes. Genome Res. 12, 1269–1276. doi: 10.1101/gr.88502
Begum, K., Mohl, J. E., Ayivor, F., Perez, E. E., and Leung, M. Y. (2020). GPCR-PEnDB: a database of protein sequences and derived features to facilitate prediction and classification of G protein-coupled receptors. Database 2020:baaa087. doi: 10.1093/database/baaa087
Bendtsen, J. D., Nielsen, H., von Heijne, G., and Brunak, S. (2004). Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340, 783–795. doi: 10.1016/j.jmb.2004.05.028
Bhattacharjee, S., Hiller, N. L., Liolios, K., Win, J., Kanneganti, T.-D., Young, C., et al. (2006). The malarial host-targeting signal is conserved in the Irish potato famine pathogen. PLoS Pathog. 2:e50. doi: 10.1371/journal.ppat.0020050
Black, A., Waipara, N. W., and Gerth, M. (2018). Correspondence: save Maori people’s sacred tree species. Nature 561:177. doi: 10.1038/d41586-018-06629-1
Boevink, P. C., Birch, P. R., Turnbull, D., and Whisson, S. C. (2020). Devastating intimacy: the cell biology of plant–Phytophthora interactions. New Phytol. 228, 445–458. doi: 10.1111/nph.16650
Bradley, E.L. (2022). Identification and functional characterisation of glycoside hydrolases from the kauri dieback pathogen, Phytophthora agathidicida. PhD, Massey University, New Zealand.
Bradley, E. L., Ökmen, B., Doehlemann, G., Henrissat, B., Bradshaw, R. E., and Mesarich, C. H. (2022). Secreted glycoside hydrolase (GH) proteins as effectors and invasion patterns of plant-associated fungi and oomycetes. Front. Plant Sci. 13:853106. doi: 10.3389/fpls.2022.853106
Bradshaw, R., Bellgard, S., Black, A., Burns, B., Gerth, M., McDougal, R., et al. (2020). Phytophthora agathidicida: research progress, cultural perspectives and knowledge gaps in the control and management of kauri dieback in New Zealand. Plant Pathol. 69, 3–16. doi: 10.1111/ppa.13104
Brasier, C. M., Beales, P. A., Denman, S., and Joan, R. (2005). Phytophthora kernoviae sp. nov., an invasive pathogen causing bleeding stem lesions on forest trees and foliar necrosis of ornamentals in the UK. Mycol. Res. 109, 853–859. doi: 10.1017/S0953756205003357
Byers, A.-K., Condron, L., Donavan, T., O'Callaghan, M., Patuawa, T., Waipara, N., et al. (2020). Soil microbial diversity in adjacent forest systems–contrasting native, old growth kauri (Agathis australis) forest with exotic pine (Pinus radiata) plantation forest. FEMS Microbiol. Ecol. 96:fiaa047. doi: 10.1093/femsec/fiaa047
Cabanettes, F., and Klopp, C. (2018). D-GENIES: dot plot large genomes in an interactive, efficient and simple way. PeerJ 6:e4958. doi: 10.7717/peerj.4958
Carleson, N. C., Press, C. M., and Grünwald, N. J. (2022). High-quality, phased genomes of Phytophthora ramorum clonal lineages NA1 and EU1. Mol. Plant-Microbe Interact. 35, 360–363. doi: 10.1094/MPMI-11-21-0264-A
Chen, N. (2004). Using repeat masker to identify repetitive elements in genomic sequences. Curr. Protoc. Bioinformat. 25, 4–10. doi: 10.1002/0471250953.bi0410s25
Chen, Y., Lun, A. T., and Smyth, G. K. (2016). From reads to genes to pathways: differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 5. doi: 10.12688/f1000research.8987.2
Chen, H., Quintana, J., Kovalchuk, A., Ubhayasekera, W., and Asiegbu, F. O. (2015). A cerato-platanin-like protein HaCPL2 from Heterobasidion annosum sensu stricto induces cell death in Nicotiana tabacum and Pinus sylvestris. Fungal Genet. Biol. 84, 41–51. doi: 10.1016/j.fgb.2015.09.007
Chen, X. R., Zhang, Y., Li, H. Y., Zhang, Z. H., Sheng, G. L., Li, Y. P., et al. (2019). The RXLR effector PcAvh1 is required for full virulence of Phytophthora capsici. Mol. Plant-Microbe Interact. 32, 986–1000. doi: 10.1094/MPMI-09-18-0251-R
Chepsergon, J., Motaung, T. E., Bellieny-Rabelo, D., and Moleleki, L. N. (2020). Organize, don’t agonize: strategic success of Phytophthora species. Microorganisms 8:917. doi: 10.3390/microorganisms8060917
Chepsergon, J., Motaung, T. E., and Moleleki, L. N. (2021). “Core” RxLR effectors in phytopathogenic oomycetes: a promising way to breeding for durable resistance in plants? Virulence 12, 1921–1935. doi: 10.1080/21505594.2021.1948277
Cock, P. J., Grüning, B. A., Paszkiewicz, K., and Pritchard, L. (2013). Galaxy tools and workflows for sequence analysis with applications in molecular plant pathology. PeerJ 1:e167. doi: 10.7717/peerj.167
Cui, C., Herlihy, J. H., Bombarely, A., McDowell, J. M., and Haak, D. C. (2019). Draft assembly of Phytophthora capsici from long-read sequencing uncovers complexity. Mol. Plant-Microbe Interact. 32, 1559–1563. doi: 10.1094/MPMI-11-21-0264-A
Dale, A. L., Feau, N., Everhart, S. E., Dhillon, B., Wong, B., Sheppard, J., et al. (2019). Mitotic recombination and rapid genome evolution in the invasive forest pathogen Phytophthora ramorum. mBbio 10, e02452–e02418. doi: 10.1128/mBio.02452-18
de Wit, P. J. G. M., van der Burgt, A., Okmen, B., Stergiopoulos, I., Abd-Elsalam, K. A., Aerts, A. L., et al. (2012). The genomes of the fungal plant pathogens Cladosporium fulvum and Dothistroma septosporum reveal adaptation to different hosts and lifestyles but also signatures of common ancestry. PLoS Genet. 8:e1003088. doi: 10.1371/journal.pgen.1003088
Deb, D., Anderson, R. G., How-Yew-Kin, T., Tyler, B. M., and McDowell, J. M. (2018). Conserved RxLR effectors from oomycetes Hyaloperonospora arabidopsidis and Phytophthora sojae suppress PAMP- and effector-triggered immunity in diverse plants. Mol. Plant Microbe Interact. 31, 374–385. doi: 10.1094/MPMI-07-17-0169-FI
Deorowicz, S., Debudaj-Grabysz, A., and Grabowski, S. (2013). Disk-based k-mer counting on a PC. BMC Bioinformat. 14, 1–12. doi: 10.1186/1471-2105-14-160
Derevnina, L., Dagdas, Y. F., De la Concepcion, J. C., Bialas, A., Kellner, R., Petre, B., et al. (2016). Nine things to know about elicitins. New Phytol. 212, 888–895. doi: 10.1111/nph.14137
Dick, M. A., Williams, N. M., Bader, M. K.-F., Gardner, J. F., and Bulman, L. S. (2014). Pathogenicity of Phytophthora pluvialis to Pinus radiata and its relation with red needle cast disease in New Zealand. N. Z. J. For. Sci. 44:6. doi: 10.1186/s40490-014-0006-7
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2013). STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21. doi: 10.1093/bioinformatics/bts635
Dong, S., and Ma, W. (2021). How to win a tug-of-war: the adaptive evolution of Phytophthora effectors. Curr. Opin. Plant Biol. 62:102027. doi: 10.1016/j.pbi.2021.102027
Dong, S., Raffaele, S., and Kamoun, S. (2015). The two-speed genomes of filamentous pathogens: waltz with plants. Curr. Opin. Genet. Dev. 35, 57–65. doi: 10.1016/j.gde.2015.09.001
Engelbrecht, J., Duong, T. A., Prabhu, S. A., Seedat, M., and van den Berg, N. (2021). Genome of the destructive oomycete Phytophthora cinnamomi provides insights into its pathogenicity and adaptive potential. BMC Genomics 22, 302–315. doi: 10.1186/s12864-021-07552-y
Fang, Y., Coelho, M. A., Shu, H., Schotanus, K., Thimmappa, B. C., Yadav, V., et al. (2020). Long transposon-rich centromeres in an oomycete reveal divergence of centromere features in Stramenopila-Alveolata-Rhizaria lineages. PLoS Genet. 16:e1008646. doi: 10.1371/journal.pgen.1008646
Feau, N., Taylor, G., Dale, A. L., Dhillon, B., Bilodeau, G. J., Birol, I., et al. (2016). Genome sequences of six Phytophthora species threatening forest ecosystems. Genomics Data 10, 85–88. doi: 10.1016/j.gdata.2016.09.013
Fletcher, K., and Michelmore, R. (2018). "From short reads to chromosome-scale genome assemblies," in Plant Pathogenic Fungi and Oomycetes. Humana Press, New York, NY, 151–197
Fletcher, K., Shin, O.-H., Clark, K. J., Feng, C., Putman, A. I., Correll, J. C., et al. (2021). Ancestral chromosomes for the Peronosporaceae inferred from a telomere-to-telomere genome assembly of Peronospora effusa. Mol. Plant Microbe Interact. 35, 450–463. doi: 10.1094/MPMI-09-21-0227-R
Flynn, J. M., Hubley, R., Goubert, C., Rosen, J., Clark, A. G., Feschotte, C., et al. (2020). RepeatModeler2 for automated genomic discovery of transposable element families. Proc. Natl. Acad. Sci. 117, 9451–9457. doi: 10.1073/pnas.1921046117
Fones, H. N., Bebber, D. P., Chaloner, T. M., Kay, W. T., Steinberg, G., and Gurr, S. J. (2020). Threats to global food security from emerging fungal and oomycete crop pathogens. Nat. Food 1, 332–342. doi: 10.1038/s43016-020-0075-0
Gel, B., and Serra, E. (2017). KaryoploteR: an R/Bioconductor package to plot customizable genomes displaying arbitrary data. Bioinformatics 33, 3088–3090. doi: 10.1093/bioinformatics/btx346
Goss, E. M., Press, C. M., and Gruenwald, N. J. (2013). Evolution of RXLR-class effectors in the oomycete plant pathogen Phytophthora ramorum. PLoS One 8:e79347. doi: 10.1371/journal.pone.0079347
Govers, F., and Gijzen, M. (2006). Phytophthora genomics: the plant destroyers' genome decoded. Mol. Plant-Microbe Interact. 19, 1295–1301. doi: 10.1094/MPMI-19-1295
Grenville-Briggs, L. J., Kushwaha, S. K., Cleary, M. R., Witzell, J., Savenkov, E. I., Whisson, S. C., et al. (2017). Draft genome of the oomycete pathogen Phytophthora cactorum strain LV007 isolated from European beech (Fagus sylvatica). Genomics Data 12, 155–156. doi: 10.1016/j.gdata.2017.05.010
Guo, Y., Dupont, P.-Y., Mesarich, C. H., Yang, B., McDougal, R. L., Panda, P., et al. (2020a). Functional analysis of RXLR effectors from the New Zealand kauri dieback pathogen Phytophthora agathidicida. Mol. Plant Pathol. 21, 1131–1148. doi: 10.1111/mpp.12967
Guo, Y., Hunziker, L., Mesarich, C. H., Chettri, P., Dupont, P.-Y., Ganley, R. J., et al. (2020b). DsEcp2-1 is a polymorphic effector that restricts growth of Dothistroma septosporum in pine. Fungal Genet. Biol. 135:103300. doi: 10.1016/j.fgb.2019.103300
Haas, B. J., Kamoun, S., Zody, M. C., Jiang, R. H., Handsaker, R. E., Cano, L. M., et al. (2009). Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 461, 393–398. doi: 10.1038/nature08358
Hamelin, R. C., and Roe, A. D. (2020). Genomic biosurveillance of forest invasive alien enemies: a story written in code. Evol. Appl. 13, 95–115. doi: 10.1111/eva.12853
Hansen, E. M. (2015). Phytophthora species emerging as pathogens of forest trees. Curr. Forest. Rep. 1, 16–24. doi: 10.1007/s40725-015-0007-7
Herewini, E. M., Scott, P. M., Williams, N. M., and Bradshaw, R. E. (2018). In vitro assays of Phytophthora agathidicida on kauri leaves suggest variability in pathogen virulence and host response. N. Z. Plant Protect. 71, 285–288. doi: 10.30843/nzpp.2018.71.127
Hoff, K. J., and Stanke, M. (2019). Predicting genes in single genomes with AUGUSTUS. Curr. Protoc. Bioinformatics 65:e57. doi: 10.1002/cpbi.57
Hua, C., Meijer, H. J., de Keijzer, J., Zhao, W., Wang, Y., and Govers, F. (2013). GK4, a G-protein-coupled receptor with a phosphatidylinositol phosphate kinase domain in Phytophthora infestans, is involved in sporangia development and virulence. Mol. Microbiol. 88, 352–370. doi: 10.1111/mmi.12190
Hunziker, L., Tarallo, M., Gough, K., Guo, M., Hargreaves, C., Loo, T. S., et al. (2021). Apoplastic effector candidates of a foliar forest pathogen trigger cell death in host and non-host plants. Sci. Rep. 11, 19958–19912. doi: 10.1038/s41598-021-99415-5
Kalvari, I., Nawrocki, E. P., Ontiveros-Palacios, N., Argasinska, J., Lamkiewicz, K., Marz, M., et al. (2021). Rfam 14: expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 49, D192–D200. doi: 10.1093/nar/gkaa1047
Kapitonov, V. V., and Jurka, J. (2007). Helitrons on a roll: eukaryotic rolling-circle transposons. Trends in Genetics 23, 521–529. doi: 10.1016/j.tig.2007.08.004
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Keriö, S., Daniels, H. A., Gomez-Gallego, M., Tabima, J. F., Lenz, R. R., Sondreli, K. L., et al. (2020). From genomes to forest management – tackling invasive Phytophthora species in the era of genomics. Can. J. Plant Pathol. 42, 1–29. doi: 10.1080/07060661.2019.1626910
Kim, D., Langmead, B., and Salzberg, S. L. (2015). HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360. doi: 10.1038/nmeth.3317
Kim, D., Paggi, J. M., Park, C., Bennett, C., and Salzberg, S. L. (2019). Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 37, 907–915. doi: 10.1038/s41587-019-0201-4
Kolmogorov, M., Yuan, J., Lin, Y., and Pevzner, P. A. (2019). Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 37, 540–546. doi: 10.1038/s41587-019-0072-8
Koren, S., Walenz, B. P., Berlin, K., Miller, J. R., Bergman, N. H., and Phillippy, A. M. (2017). Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 27, 722–736. doi: 10.1101/gr.215087.116
Krogh, A., Larsson, B., Von Heijne, G., and Sonnhammer, E. L. (2001). Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580. doi: 10.1006/jmbi.2000.4315
Lambert, S., Waipara, N., Black, A., Mark-Shadbolt, M., and Wood, W. (2018). "Indigenous biosecurity: Māori responses to kauri dieback and myrtle rust in Aotearoa New Zealand," in The Human Dimensions of Forest and Tree Health. Urquhart, J., Marzano, M., and Potter, C. Palgrave Macmillan, Cham, 109–137
Lamour, K. H., Mudge, J., Gobena, D., Hurtado-Gonzales, O. P., Schmutz, J., Kuo, A., et al. (2012). Genome sequencing and mapping reveal loss of heterozygosity as a mechanism for rapid adaptation in the vegetable pathogen Phytophthora capsici. Mol. Plant-Microbe Interact. 25, 1350–1360. doi: 10.1094/MPMI-02-12-0028-R
Lawrence, S. A., Armstrong, C. B., Patrick, W. M., and Gerth, M. L. (2017). High-throughput chemical screening identifies compounds that inhibit different stages of the Phytophthora agathidicida and Phytophthora cinnamomi life cycles. Front. Microbiol. 8:1340. doi: 10.3389/fmicb.2017.01340
Lewis, K. S., Black, A., Condron, L. M., Waipara, N. W., Scott, P., Williams, N., et al. (2019). Land-use changes influence the sporulation and survival of Phytophthora agathidicida, a lethal pathogen of New Zealand kauri (Agathis australis). For. Pathol. 49:e12502. doi: 10.1111/efp.12502
Li, H. (2013). Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 1303, 3997. doi:doi: 10.48550/arXiv.1303.3997.
Li, H. (2018). Minimap2: pairwise alignment for nucleotide sequences. Bioinformatics 34, 3094–3100. doi: 10.1093/bioinformatics/bty191
Li, H. Y., Wang, H. N., Jing, M. F., Zhu, J. Y., Guo, B. D., Wang, Y., et al. (2018). A Phytophthora effector recruits a host cytoplasmic transacetylase into nuclear speckles to enhance plant susceptibility. elife 7:e40039. doi: 10.7554/eLife.40039
Liao, Y., Smyth, G. K., and Shi, W. (2014). FeatureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930. doi: 10.1093/bioinformatics/btt656
Liao, Y., Smyth, G. K., and Shi, W. (2019). The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 47:e47. doi: 10.1093/nar/gkz114
Liu, T., Ye, W., Ru, Y., Yang, X., Gu, B., Tao, K., et al. (2011). Two host cytoplasmic effectors are required for pathogenesis of Phytophthora sojae by suppression of host defenses. Plant Physiol. 155, 490–501. doi: 10.1104/pp.110.166470
Lowe, T. M., and Eddy, S. R. (1997). tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 25, 955–964. doi: 10.1093/nar/25.5.955
Lu, M., Feau, N., Vidakovic, D. O., Ukrainetz, N., Wong, B., Aitken, S. N., et al. (2021). Comparative gene expression analysis reveals mechanism of Pinus contorta response to the fungal pathogen Dothistroma septosporum. Mol. Plant-Microbe Interact. 34, 397–409. doi: 10.1094/MPMI-10-20-0282-R
Ma, Z., Song, T., Zhu, L., Ye, W., Wang, Y., Shao, Y., et al. (2015). A Phytophthora sojae glycoside hydrolase 12 protein is a major virulence factor during soybean infection and is recognized as a PAMP. Plant Cell 27, 2057–2072. doi: 10.1105/tpc.15.00390
McGowan, J., and Fitzpatrick, D. A. (2017). Genomic, network, and phylogenetic analysis of the oomycete effector arsenal. mSphere 2, e00408–e00417. doi: 10.1128/mSphere.00408-17
McGowan, J., O’Hanlon, R., Owens, R. A., and Fitzpatrick, D. A. (2020). Comparative genomic and proteomic analyses of three widespread Phytophthora species: Phytophthora chlamydospora, Phytophthora gonapodyides and Phytophthora pseudosyringae. Microorganisms 8:653. doi: 10.3390/microorganisms8050653
Meijer, H. J. G., Mancuso, F. M., Espadas, G., Seidl, M. F., Chiva, C., Govers, F., et al. (2014). Profiling the secretome and extracellular proteome of the potato late blight pathogen Phytophthora infestans. Mol. Cell. Proteomics 13, 2101–2113. doi: 10.1074/mcp.M113.035873
Midgley, K., van den Berg, N., and Swart, V. (2022). Unraveling plant cell death during Phytophthora infection. Microorganisms 10:1139. doi: 10.3390/microorganisms10061139
Nawrocki, E. P., and Eddy, S. R. (2013). Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 29, 2933–2935. doi: 10.1093/bioinformatics/btt509
Padamsee, M., Johansen, R. B., Stuckey, S. A., Williams, S. E., Hooker, J. E., Burns, B. R., et al. (2016). The arbuscular mycorrhizal fungi colonising roots and root nodules of New Zealand kauri Agathis australis. Fungal Biol. 120, 807–817. doi: 10.1016/j.funbio.2016.01.015
Pais, M., Yoshida, K., Giannakopoulou, A., Pel, M. A., Cano, L. M., Oliva, R. F., et al. (2018). Gene expression polymorphism underpins evasion of host immunity in an asexual lineage of the Irish potato famine pathogen. BMC Evol. Biol. 18, 93–11. doi: 10.1186/s12862-018-1201-6
Pautasso, M., Schlegel, M., and Holdenrieder, O. (2015). Forest health in a changing world. Microb. Ecol. 69, 826–842. doi: 10.1007/s00248-014-0545-8
Pirc, K., Clifton, L. A., Yilmaz, N., Saltalamacchia, A., Mally, M., Snoj, T., et al. (2022). An oomycete NLP cytolysin forms transient small pores in lipid membranes. Science. Advances 8:eabj9406. doi: 10.1126/sciadv.abj9406
Poplin, R., Ruano-Rubio, V., DePristo, M.A., Fennell, T.J., Carneiro, M.O., Van der Auwera, G.A., et al. (2018). Scaling accurate genetic variant discovery to tens of thousands of samples. BioRxiv, 201178. doi:doi: 10.1101/201178
Quinn, L., O'Neill, P. A., Harrison, J., Paskiewicz, K. H., McCracken, A. R., Cooke, L. R., et al. (2013). Genome-wide sequencing of Phytophthora lateralis reveals genetic variation among isolates from Lawson cypress (Chamaecyparis lawsoniana) in Northern Ireland. FEMS Microbiol. Lett. 344, 179–185. doi: 10.1111/1574-6968.12179
Qutob, D., Chapman, B. P., and Gijzen, M. (2013). Transgenerational gene silencing causes gain of virulence in a plant pathogen. Nat. Commun. 4:1349. doi: 10.1038/ncomms2354
R Core Team (2022). R: a language and environment for statistical computing. R Foundation for statistical computing, vienna, austria. https://www.R-project.org
Raffaele, S., Farrer, R. A., Cano, L. M., Studholme, D. J., MacLean, D., Thines, M., et al. (2010). Genome evolution following host jumps in the Irish potato famine pathogen lineage. Science 330, 1540–1543. doi: 10.1126/science.1193070
Ranallo-Benavidez, T. R., Jaron, K. S., and Schatz, M. C. (2020). GenomeScope 2.0 and Smudgeplot for reference-free profiling of polyploid genomes. Nature. Communications 11, 1–10. doi: 10.1038/s41467-020-14998-3
Rizzo, D., Garbelotto, M., Davidson, J., Slaughter, G., and Koike, S. (2002). Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 86, 205–214. doi: 10.1094/PDIS.2002.86.3.205
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rocafort, M., Fudal, I., and Mesarich, C. H. (2020). Apoplastic effector proteins of plant-associated fungi and oomycetes. Curr. Opin. Plant Biol. 56, 9–19. doi: 10.1016/j.pbi.2020.02.004
Scott, P., Taylor, P., Gardner, J., Puértolas, A., Panda, P., Addison, S., et al. (2019). Phytophthora aleatoria sp. nov., associated with root and collar damage on Pinus radiata from nurseries and plantations. Australas. Plant Pathol. 48, 313–321. doi: 10.1007/s13313-019-00631-5
Seppey, M., Manni, M., and Zdobnov, E. M. (2019). “BUSCO: assessing genome assembly and annotation completeness” in Gene Prediction (Cham: Springer), 227–245.
Sperschneider, J., Williams, A. H., Hane, J. K., Singh, K. B., and Taylor, J. M. (2015). Evaluation of secretion prediction highlights differing approaches needed for oomycete and fungal effectors. Front. Plant Sci. 6:1168. doi: 10.3389/fpls.2015.01168
Stajich, J. E., Vu, A. L., Judelson, H. S., Vogel, G. M., Gore, M. A., Carlson, M. O., et al. (2021). High-quality reference genome sequence for the oomycete vegetable pathogen Phytophthora capsici strain LT1534. Microbiol. Res. Announc. 10, e00295–e00221. doi: 10.1128/MRA.00295-21
Stam, R., Jupe, J., Howden, A. J., Morris, J. A., Boevink, P. C., Hedley, P. E., et al. (2013). Identification and characterisation CRN effectors in Phytophthora capsici shows modularity and functional diversity. PLoS One 8:e59517. doi: 10.1371/journal.pone.0059517
Stanke, M., Schöffmann, O., Morgenstern, B., and Waack, S. (2006). Gene prediction in eukaryotes with a generalized hidden Markov model that uses hints from external sources. BMC Bioinformat. 7, 1–11. doi: 10.1186/1471-2105-7-62
Studholme, D. J., McDougal, R. L., Sambles, C., Hansen, E., Hardy, G., Grant, M., et al. (2016). Genome sequences of six Phytophthora species associated with forests in New Zealand. Genomics Data 7, 54–56. doi: 10.1016/j.gdata.2015.11.015
Suvakov, M., Panda, A., Diesh, C., Holmes, I., and Abyzov, A. (2021). CNVpytor: a tool for copy number variation detection and analysis from read depth and allele imbalance in whole-genome sequencing. GigaScience 10:giab074. doi: 10.1093/gigascience/giab074
Tabima, J. F., and Grünwald, N. J. (2019). effectR: an expandable R package to predict candidate RxLR and CRN effectors in oomycetes using motif searches. Mol. Plant-Microbe Interact. 32, 1067–1076. doi: 10.1094/MPMI-10-18-0279-TA
Tarallo, M., McDougal, R. L., Chen, Z., Wang, Y., Bradshaw, R. E., and Mesarich, C. H. (2022). Characterization of two conserved cell death elicitor families from the Dothideomycete fungal pathogens Dothistroma septosporum and Fulvia fulva (syn. Cladosporium fulvum). Front. Microbiol. 13:964851. doi: 10.3389/fmicb.2022.964851
Terhonen, E., Blumenstein, K., Kovalchuk, A., and Asiegbu, F. O. (2019). Forest tree microbiomes and associated fungal endophytes: functional roles and impact on forest health. Forests 10:42. doi: 10.3390/f10010042
Thorpe, P., Vetukuri, R. R., Hedley, P. E., Morris, J., Whisson, M. A., Welsh, L. R., et al. (2021). Draft genome assemblies for tree pathogens Phytophthora pseudosyringae and Phytophthora boehmeriae. G3 11:jkab282. doi: 10.1093/g3journal/jkab282
Tobias, P. A., Schwessinger, B., Deng, C. H., Wu, C., Dong, C., Sperschneider, J., et al. (2021). Austropuccinia psidii, causing myrtle rust, has a gigabase-sized genome shaped by transposable elements. G3 11:jkaa015. doi: 10.1093/g3journal/jkaa015
Tyler, B. M., Tripathy, S., Zhang, X., Dehal, P., Jiang, R. H., Aerts, A., et al. (2006). Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science 313, 1261–1266. doi: 10.1126/science.1128796
Van der Auwera, G.A., and O'Connor, B.D. (2020). Genomics in the Cloud: Using Docker, GATK, and WDL in Terra. Sebastopol, CA: O'Reilly Media.
Van Ghelder, C., Parent, G. J., Rigault, P., Prunier, J., Giguère, I., Caron, S., et al. (2019). The large repertoire of conifer NLR resistance genes includes drought responsive and highly diversified RNLs. Sci. Rep. 9, 11614–11613. doi: 10.1038/s41598-019-47950-7
Vetukuri, R. R., Tripathy, S., Malar, C. M., Panda, A., Kushwaha, S. K., Chawade, A., et al. (2018). Draft genome sequence for the tree pathogen Phytophthora plurivora. Genome Biol. Evol. 10, 2432–2442. doi: 10.1093/gbe/evy162
Vleeshouwers, V. G. A. A., and Oliver, R. P. (2014). Effectors as tools in disease resistance breeding against biotrophic, hemibiotrophic, and necrotrophic plant pathogens. Mol. Plant-Microbe Interact. 27, 196–206. doi: 10.1094/MPMI-10-13-0313-IA
Walker, B. J., Abeel, T., Shea, T., Priest, M., Abouelliel, A., Sakthikumar, S., et al. (2014). Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS One 9:e112963. doi: 10.1371/journal.pone.0112963
Wang, L., Chen, H., Li, J., Shu, H., Zhang, X., Wang, Y., et al. (2020b). Effector gene silencing mediated by histone methylation underpins host adaptation in an oomycete plant pathogen. Nucleic Acids Res. 48, 1790–1799. doi: 10.1093/nar/gkz1160
Wang, S., McLellan, H., Bukharova, T., He, Q., Murphy, F., Shi, J., et al. (2019a). Phytophthora infestans RXLR effectors act in concert at diverse subcellular locations to enhance host colonization. J. Exp. Bot. 70, 343–356. doi: 10.1093/jxb/ery360
Wang, Y., Pruitt, R. N., Nürnberger, T., and Wang, Y. (2022). Evasion of plant immunity by microbial pathogens. Nat. Rev. Microbiol. 56, 21–40. doi: 10.1146/annurev-phyto-080516-035303
Wang, J., Shen, D., Ge, C., Du, Y., Lin, L., Liu, J., et al. (2020a). Filamentous Phytophthora pathogens deploy effectors to interfere with bacterial growth and motility. Front. Microbiol. 11:581511. doi: 10.3389/fmicb.2020.581511
Wang, Y., Tyler, B. M., and Wang, Y. (2019b). Defense and counterdefense during plant-pathogenic oomycete infection. Annu. Rev. Microbiol. 73, 667–696. doi: 10.1146/annurev-micro-020518-120022
Weir, B. S., Paderes, E. P., Anand, N., Uchida, J. Y., Pennycook, S. R., Bellgard, S. E., et al. (2015). A taxonomic revision of Phytophthora Clade 5 including two new species, Phytophthora agathidicida and P. cocois. Phytotaxa 205, 21–38. doi: 10.11646/PHYTOTAXA.205.1.2
Whisson, S. C., Boevink, P. C., Moleleki, L., Avrova, A. O., Morales, J. G., Gilroy, E. M., et al. (2007). A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 450, 115–118. doi: 10.1038/nature06203
Wickham, H., François, R., Henry, L., and Müller, K. (2015). “dplyr: a grammar of data manipulation,” in R package version 0.4. R Foundation for statistical computing, Vienna, Austria.
Wilson, R. A., and McDowell, J. M. (2022). Recent advances in understanding of fungal and oomycete effectors. Curr. Opin. Plant Biol. 68:102228. doi: 10.1016/j.pbi.2022.102228
Win, J., Morgan, W., Bos, J., Krasileva, K. V., Cano, L. M., Chaparro-Garcia, A., et al. (2007). Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell Online 19, 2349–2369. doi: 10.1105/tpc.107.051037
Winkworth, R. C., Bellgard, S. E., McLenachan, P. A., and Lockhart, P. J. (2021). The mitogenome of Phytophthora agathidicida: evidence for a not so recent arrival of the “kauri killing” Phytophthora in New Zealand. PLoS One 16:e0250422. doi: 10.1371/journal.pone.0250422
Wyse, S. V., Burns, B. R., and Wright, S. D. (2014). Distinctive vegetation communities are associated with the long-lived conifer Agathis australis (New Zealand kauri, Araucariaceae) in New Zealand rainforests. Austral Ecol. 39, 388–400. doi: 10.1111/aec.12089
Xiao, C.-L., Chen, Y., Xie, S.-Q., Chen, K.-N., Wang, Y., Han, Y., et al. (2017). MECAT: fast mapping, error correction, and de novo assembly for single-molecule sequencing reads. Nat. Methods 14, 1072–1074. doi: 10.1038/nmeth.4432
Yang, M., Duan, S. C., Mei, X. Y., Huang, H. C., Chen, W., Liu, Y. X., et al. (2018). The Phytophthora cactorum genome provides insights into the adaptation to host defense compounds and fungicides. Sci. Rep. 8:6534. doi: 10.1038/s41598-018-24939-2
Ye, W., Wang, X., Tao, K., Lu, Y., Dai, T., Dong, S., et al. (2011). Digital gene expression profiling of the Phytophthora sojae transcriptome. Mol. Plant-Microbe Interact. 24, 1530–1539. doi: 10.1094/MPMI-05-11-0106
Zaccaron, A. Z., Chen, L.-H., Samaras, A., and Stergiopoulos, I. (2022). A chromosome-scale genome assembly of the tomato pathogen Cladosporium fulvum reveals a compartmentalized genome architecture and the presence of a dispensable chromosome. Microb. Genom. 8:000819. doi: 10.1099/mgen.0.000819
Zdobnov, E. M., Kuznetsov, D., Tegenfeldt, F., Manni, M., Berkeley, M., and Kriventseva, E. V. (2021). OrthoDB in 2020: evolutionary and functional annotations of orthologs. Nucleic Acids Res. 49, D389–D393. doi: 10.1093/nar/gkaa1009
Zhang, D., Burroughs, A. M., Vidal, N. D., Iyer, L. M., and Aravind, L. (2016). Transposons to toxins: the provenance, architecture and diversification of a widespread class of eukaryotic effectors. Nucleic Acids Res. 44, 3513–3533. doi: 10.1093/nar/gkw221
Zhang, X., Liu, B., Zou, F., Shen, D., Yin, Z., Wang, R., et al. (2019). Whole genome re-sequencing reveals natural variation and adaptive evolution of Phytophthora sojae. Front. Microbiol. 10:2792. doi: 10.3389/fmicb.2019.02792
Keywords: Phytophthora, oomycete, kauri dieback, forest disease, chromosome-level genome assembly, chromatin conformation capture, effectors
Citation: Cox MP, Guo Y, Winter DJ, Sen D, Cauldron NC, Shiller J, Bradley EL, Ganley AR, Gerth ML, Lacey RF, McDougal RL, Panda P, Williams NM, Grunwald NJ, Mesarich CH and Bradshaw RE (2022) Chromosome-level assembly of the Phytophthora agathidicida genome reveals adaptation in effector gene families. Front. Microbiol. 13:1038444. doi: 10.3389/fmicb.2022.1038444
Edited by:
James Hane, Curtin University, AustraliaReviewed by:
David J. Studholme, University of Exeter, United KingdomKusch Stefan, RWTH Aachen University, Germany
Copyright © 2022 Cox, Guo, Winter, Sen, Cauldron, Shiller, Bradley, Ganley, Gerth, Lacey, McDougal, Panda, Williams, Grunwald, Mesarich and Bradshaw. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosie E. Bradshaw, r.e.bradshaw@massey.ac.nz