 Ming-Cheng Chiang
Ming-Cheng Chiang Edward Chern
Edward Chern- 1niChe Lab for Stem Cell and Regenerative Medicine, Department of Biochemical Science and Technology, National Taiwan University, Taipei, Taiwan
- 2Research Center for Developmental Biology and Regenerative Medicine, National Taiwan University, Taipei, Taiwan
Recently, increasing studies have emphasized the importance of commensal bacteria in humans, including microbiota in the oral cavity, gut, vagina, or skin. Ocular surface microbiota (OSM) is gaining great importance as new methodologies for bacteria DNA sequencing have been published. This review outlines the current understanding and investigation of OSM and introduces the new concept of the gut–eye axis. Moreover, we have collected current studies that focus on the relationship between ophthalmic infectious disease and alterations in the OSM or human gut microbiota. Finally, we discuss the current application of probiotics in ophthalmic infectious disease, its limitations to date, and futural directions.
Introduction
Microbiota is a collective term for polymicrobial communities, including bacteria, virus, fungi, and archaea. These may inhabit a particular site, shape the microenvironment, and play a role in metabolism, disease development, and immunomodulation in the human body (De Sordi et al., 2019). Microbiota in the gastrointestinal tract is well recognized to modulate homeostasis of the digestive system. Its clinical applications are numerous; however, the ocular surface has its own microbiota that is related to several ophthalmic diseases (Zegans and Van Gelder, 2014). Moreover, OSM can inhibit the growth of pathogens and is highly related to eye infections. OSM is composed of several microorganisms, ranging from bacteria to fungus. The variations in OSM can be caused by personal habits, systemic diseases, or antibiotic usage, leading to disturbances in the local immune function. Moreover, the gut–eye axis, which illustrates the relationship between gastrointestinal microbiota and the ocular microenvironment, has been proposed. Combined with better sequencing skill and furthering understanding of human microbiota, the application of probiotics to improve ocular or gastrointestinal microbiota and for ophthalmic infectious disease treatment is an important object of study.
Properties of OSM and OSM identification
Local immune system and microbiota
The ocular surface is a defensive frontline of the innate immune system. It is composed of the cornea and sclera, and their overlying tissue, such as the conjunctiva and tear film. It forms a physical barrier through the presence of epithelial intercellular tight junctions (Mantelli et al., 2013). The tear film contains several anti-infectious molecules, including lysozymes, lactoferrin, lipocalin, and β-defensins (McDermott, 2013). Resident conjunctiva lymphocytes, plasma cells, macrophages, and dendritic cells (DC) can be triggered to produce antibodies and upregulate the downstream immune response, eliminating non-self microorganisms by phagocytosis and intracellular degradation (Bauer et al., 2002; Petrillo et al., 2020). The ocular surface is armored with various antimicrobial strategies; however, it can tolerate some commensal colonies, such as OSM. Lipopolysaccharides (LPSs) are the major component of the outer membrane of Gram-negative bacteria (GNB), which serves as a barrier resistant to host-derived antimicrobial compounds (Erridge et al., 2002). LPSs are pathogen-associated molecular patterns (PAMPs). This means that they can be recognized as a molecular hallmark of invading microbes by the host (Erridge, 2010). In addition, lipoteichoic acid in Gram-positive bacteria (GPB), lipoproteins, and mycobacterial lipoglycans also meet the criteria of PAMPs (Ray et al., 2013). Pattern recognition receptors (PRRs) are surface proteins of antigen-presenting cells (APCs) capable of recognizing PAMPs (Gordon, 2002). Once PRRs recognize and link to the PAMP, a downstream signaling process is initiated and production of pro-inflammatory cytokines are upregulated as the PRR–PAMP pair is an invasive signal for the immune system (Mogensen, 2009). However, PRRs contain various subfamilies. Toll-like receptors (TLRs), most widely known among all, are closely related to the immune silence of the ocular surface (Takeuchi and Akira, 2010).
TLRs
TLRs are transmembrane receptors, which can be divided into two groups depending on their cellular localization and PAMP ligands: One group comprises TLR1, TLR2, TLR4, TLR5, TLR6, and TLR11, which are categorized together as they are located on cell surfaces and recognize microbial membrane components; the other group is composed of TLR3, TLR7, TLR8, and TLR9, which are located in intracellular vesicles and recognize microbial nucleic acids. Each TLR has its own specific characteristic and function. TLR2 senses lipoproteins by forming a heterodimer with either TLR1 or TLR6. TLR4 combined with myeloid differentiation factor 2 appears to recognize LPSs. Hayashi et al. (2001) found bacterial flagellin, a binding ligand of TLR5. TLR11 recognizes the profilin-like molecule derived from Toxoplasma gondii; however, the TLR11 functional protein does not exist in humans because a stop codon is contained in its open reading frame. TLR3 can identify the double-stranded RNA of some viruses, while TLR7 and TLR8 recognize single-stranded RNA. TLR9 can be activated by microbial DNA sequences containing unmethylated CpG dinucleotides. After binding with specific ligands and being activated, TLRs trigger downstream signal pathways and inflammatory gene transcription, contributing to antimicrobial response.
Immune silence
A complicated defensive system inhibits microbial growth on the ocular surface; however, some microorganisms are tolerable to the human immune system. Immune system homeostasis is maintained by the balance between regulatory and stimulative signals as antigen-loaded DCs migrate to lymph nodes and induce regulatory or immunogenic T cells (Horwitz et al., 2019). The type of T-cell expansion that is produced is determined by the presence of danger signals, such as PAMPs or endogenous signals derived from cell damage (Idzko et al., 2007). By contrast, an unclear mechanism imprints the tolerogenic profile on ocular surface DCs in basal conditions. Once DCs loaded with ocular surface-derived antigens contact circulating naive T cells, regulatory T cells are induced because tolerogenic profiles have been imprinted on DCs previously (Galletti et al., 2017). Several hypotheses have been published explaining the immune silence phenomenon on the ocular surface. TLR2 and TLR4 are expressed intracellularly in human corneal epithelial cells (Ueta et al., 2004). TLR5 is located at wing cells and the basal cell layer, instead of the outermost part of the cornea; therefore, bacteria cannot bind to TLR5 as long as the integrity of the epithelial barrier persists (Zhang et al., 2003). Thus, immune activity at the ocular surface is regulated by various mechanisms, leading to an immune-silent environment within which microbiota can survive.
Members of ocular microbiota
Culture-dependent method
The ocular surface is constantly exposed to the environment. Whether the stable presence of commensal bacteria exists remains under debate. In the past, OSM was harvested from tears or swabs of the ocular surface and cultured. Of all cultivable microorganisms from conjunctiva swabs, GPB, including Staphylococcus, Propionibacterium sp., diphtheroids, and Staphylococcus aureus, accounted for the majority. In addition, GNB, such as Pseudomonas sp., Enterobacter sp., and Escherichia coli, and fungus were found in healthy eyes, although less common. Overall, coagulase-negative staphylococci were the most common bacteria isolated from the conjunctiva, lids, or tears (Gordon, 2002). However, traditional culture methods for OSM identification are seldom used currently due to their limited detection of slow growing, fastidious, or uncultivable species.
Culture-independent method
Next-generation 16S rRNA sequencing has been applied to microorganism identification over time. rRNA is the most conserved and least variable gene in all cells, belonging to the same genus and species. 16S rRNA is a small ribosomal subunit containing 1,500 nucleotides, composed of hypervariable regions and strongly conserved regions. A primer is designed to bind conserved regions and amplify the variable parts between different genera and species. Thus, 16S rRNA sequencing is a tool to classify bacteria to the species level and recognize closely related bacterial species. As a result, this sequencing has enabled the classification of uncultivable bacteria and the tracing of phylogenetic relationships, leading to tremendous development in taxonomy.
Application to OSM
16S rRNA sequencing has been applied to OSM identification. Graham et al. identified the bacterial genera of conjunctiva samples from 91 subjects by culture and 16S rRNA sequencing. Much more genera could be identified by 16S rRNA sequencing than by the culture method. It revealed coagulase-negative Staphylococcus sp., Staphylococcus epidermidis, Bacillus sp., Rhodococcus sp., and uncultivable bacterium, including Corynebacterium sp., Klebsiella sp., and Erwinia sp. By contrast, only coagulase-negative Staphylococcus and Bacillus sp. were grown in culture (Graham et al., 2007). In addition, fungi were identified as a member of OSM. A total of two phyla, Basidiomycota and Ascomycota, and five genera, Malassezia, Rhodotorula, Davidiella, Aspergillus, and Alternaria, were found to account for >80% of fungal flora and were isolated from >80% of 45 tested samples (Wang et al., 2020). Viruses are found on the ocular surface. A study has demonstrated the presence of torque teno virus on the ocular surface in 65% of 107 healthy volunteers (Doan et al., 2016).
In microbial ecology, the term “microbiome” is an assembly of the genomes of microbial symbionts that live inside and on humans, and “core microbiome” is defined as the set of genes present in a given habitat in all or the vast majority of humans (Turnbaugh et al., 2007). However, there is a lack of consensus about whether core microbiomes exist on the ocular surface. Dong et al. utilized 16S rRNA sequencing to survey OSM of both eyes in four subjects. A total of 12 genera—Pseudomonas, Propionibacterium, Bradyrhizobium, Corynebacterium, Acinetobacter, Brevundimonas, Staphylococcus, Aquabacterium, Sphingomonas, Streptococcus, Streptophyta, and Methylobacterium—are considered the putative core microbiomes as they were ubiquitous among all examined subjects (Dong et al., 2011). Ozkan et al. sampled the conjunctiva of 43 healthy subjects at three time points: baseline, 1 month, and 3 months. A total of 183 operational taxonomic units (OTUs) were collected after contamination filtering. Each subject carried 33 OTUs on average across the three time points. OTUs associated with Corynebacterium, Streptococcus, Anaerococcus, Sphingomonas, and Acinetobacter were found in one or more individuals throughout the whole study; however, no OTU was noted in all individuals at all time points or in all individuals in any one time point, indicating that the ocular surface may not carry consistent OTUs (Ozkan et al., 2017). This contrasts with the signature core microbiome of OTUs that are found on the human skin, in the gut, or in the oral cavity (Zaura et al., 2009; Zhu et al., 2010; Byrd et al., 2018). As a result, whether “core” residential microbiomes exist on the ocular surface remains unclear.
Influence parameters of OSM and current limitations in OSM identification
Inherent factors
A study of 106 infants in which conjunctival swabs were taken immediately after birth showed that infants delivered vaginally carried more bacterial species and colony-forming units than infants delivered via cesarean delivery (Isenberg et al., 1988). The Shannon index is used to reflect the number of species or types in a database. The higher the number, the higher the species diversity. This was believed to be lower in the elderly (60–70 years old) than in younger adults (25–35 years old) (Suzuki et al., 2020). However, the opposite outcome has been published by Wen et al. as they found a higher Shannon index in an elderly group (Wen et al., 2017). These outcomes may be led by different sequencing technologies, for example, 16S rRNA sequencing and metagenomic shotgun sequencing. Previous studies have shown mixed opinions on the relationship between gender and OSM (Ozkan et al., 2017; Cavuoto et al., 2018), with no consensus reached to date.
Ozkan et al. studied the bacterial biogeography in the human eyes and divided it into four areas: ocular surface, fornix conjunctiva tissue, lid margin, and skin 1 cm above the eyelid margin. Pseudomonas was often found on the fornix conjunctiva and lid margin, while Acinetobacter and Aeribacillus tended to be noted on the ocular surface. At the phylum level, Proteobacteria made up 90% of the OSM of the fornix conjunctival tissue, while Bacteroidetes appeared irregularly across all sites. Actinobacteria was more frequently found on the periocular skin than in other groups (Ozkan et al., 2019) (Figure 1). Detailed information at the genus level is shown in Table 1. Cavuoto et al. (2019) found that Firmicutes accounted for the majority of microbiota on the lid margin. This result differs from that of Ozkan et al. (2019); however, both found that Proteobacteria was predominant on the ocular surface, followed by Firmicutes and that Firmicutes had a slim lead over Proteobacteria on the periocular skin (Figure 2). A depth-stratified microbiota was identified by using samples collected from the same individual with different swab pressures. A greater relative abundance of Staphylococcus and Corynebacterium was noted with soft pressure swabs; the presence of Proteobacteria increased in swabs applied with firm pressure (Dong et al., 2011). Moreover, differences in OSM can exist between each eye in a same person (Zilliox et al., 2020).
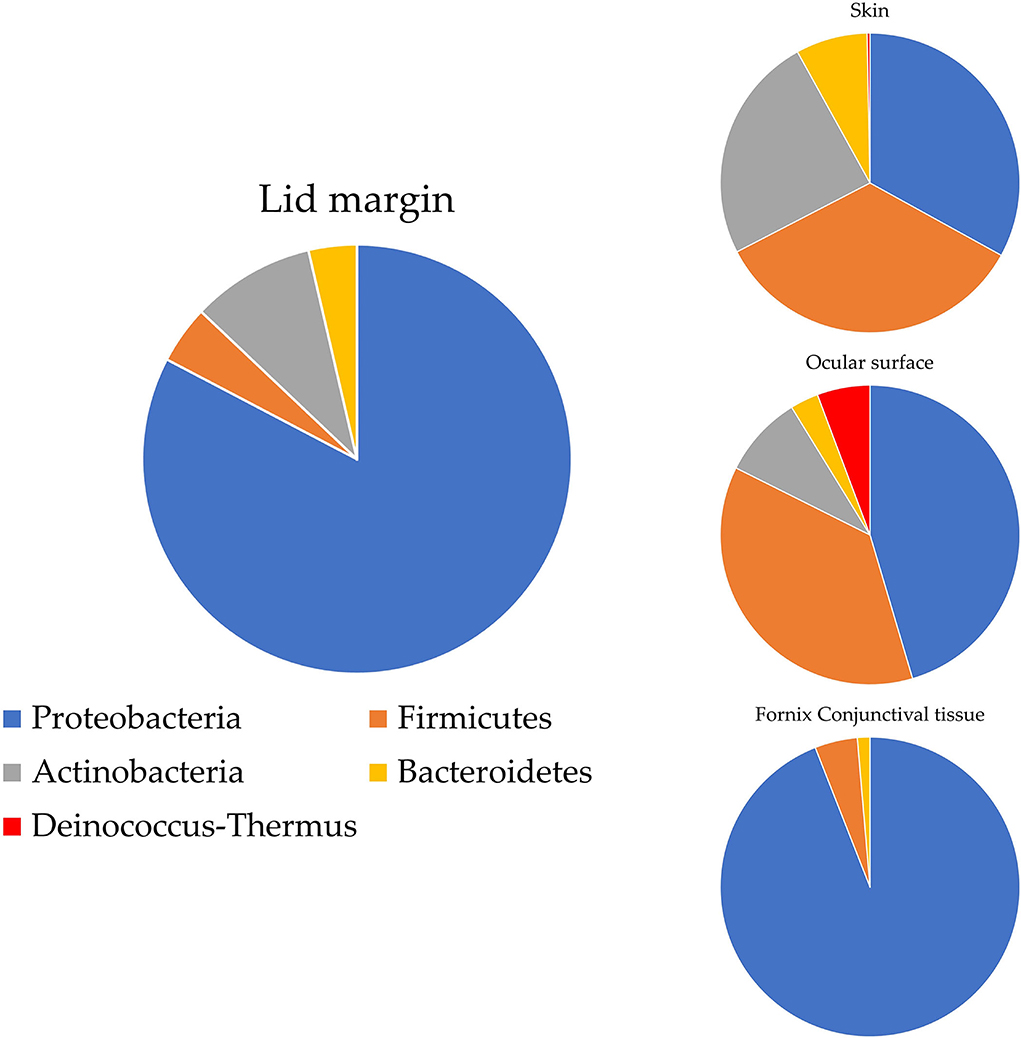
Figure 1. OSM distribution on ocular surface by Ozkan et al. (2019). Proteobacteria account for most of the ocular flora in most areas, while the periocular skin flora is predominated by Firmicutes and colonized with Actinobacteria more frequently than other areas.
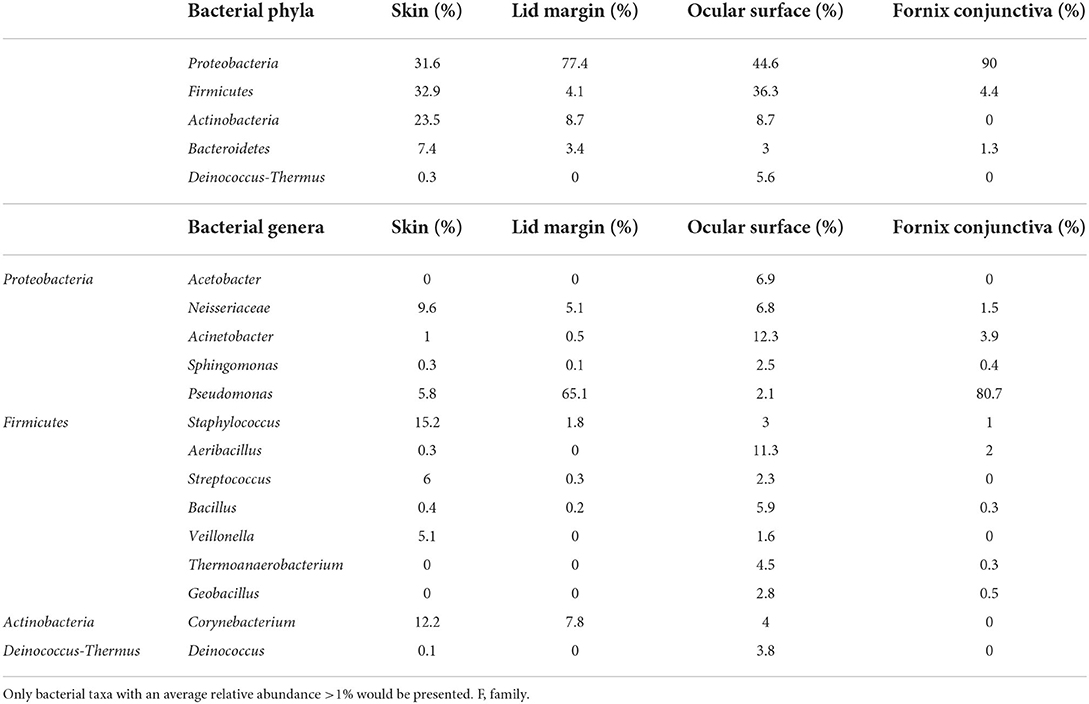
Table 1. Members of OSM on ocular surface by Ozkan et al. (2019).
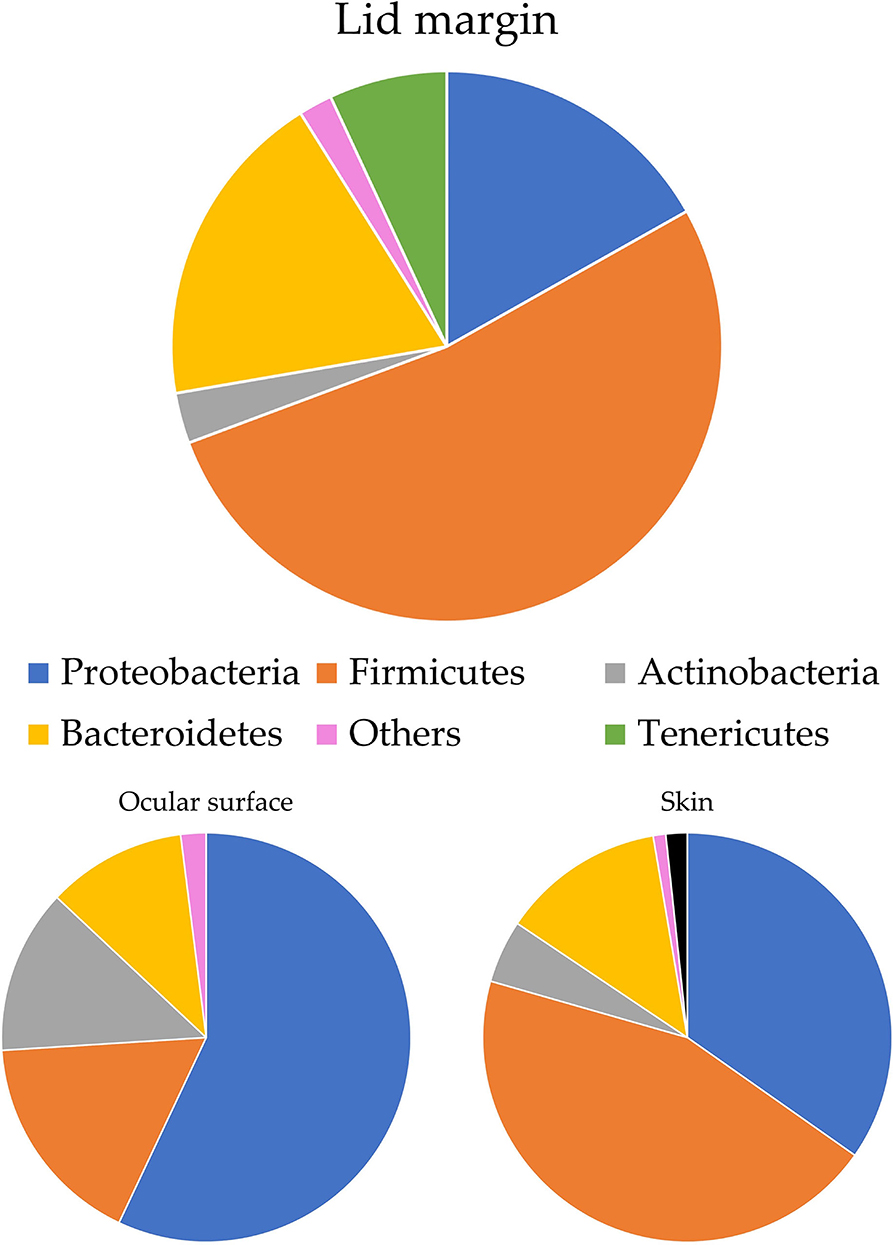
Figure 2. OSM distribution on ocular surface by Cavuoto et al. (2019). Firmicutes are the most abundant flora of the lid margin, while Ozkan et al. (2019) have reported that Proteobacteria is the most prevalent here. The two studies agree that Proteobacteria is the most common phyla of the ocular surface and that periocular skin is dominated by Firmicutes.
Acquired factors
A previous study pointed out that the OSM of newborns delivered vaginally carried more bacterial species and colony-forming units (Isenberg et al., 1988). However, conjunctival cultures collected 2 days after birth showed a more frequently positive outcome than those collected at birth, regardless of the delivery type, suggesting further shaping and modulation of OSM by diet and environment (Lee et al., 1989).
Antibiotics have been used for a long time to treat bacterial infections. However, antibiotic usage, either locally or systemically, can impact OSM. Dave et al. delivered azithromycin and fluoroquinolone intravitreally and then observed a significant change in ocular flora. Staphylococcus epidermidis and Staphylococcus aureus accounted for 54.5 and 18.2% of cultured isolates at baseline, respectively, which changed to 90.9% (P < 0.05) and 4.5% (P < 0.05) after azithromycin treatment. In the fluoroquinolone group, Staphylococcus epidermidis increased from 45.7 to 63.4% (P < 0.05) (Dave et al., 2013). Patients who underwent topical levofloxacin (LVFX) treatment for 1 month were recruited by Ono et al. to investigate the impact of antibiotics on OSM. The outcome showed substantially reduced bacterial diversity after LVFX usage (P < 0.01) (Ono et al., 2017). Moxifloxacin, a common topical-use antibiotic, is found to reduce culture positivity rates on the ocular surface (Celebi and Onerci Celebi, 2021). In addition, antibiotics do affect the drug resistance of microbiota as topical LVFX and moxifloxacin have been found to increase the mean MIC of ocular flora (Yin et al., 2013; Ono et al., 2017). Moreover, preoperative conjunctival isolates of coagulase-negative Staphylococcus without previous antibiotic treatment had better susceptibility to vancomycin and LVFX (Ta et al., 2003).
Contact lenses are widely used for vision correction, enhancing corneal healing, and drug delivery. Orthokeratology lens (OKL) wearers have a lower abundance of Bacillus, Tatumella, and Lactobacillus, while the abundance of Delftia decreases in soft contact lens wearers when compared with subjects who do not wear contact lenses (Zhang et al., 2017). Interestingly, lens wearers appear to have ocular flora more like skin flora with higher abundances of Methylobacterium, Lactobacillus, Acinetobacter, and Pseudomonas (Shin et al., 2016).
Eye drops are a major drug delivery route for ophthalmic disease. A significantly lower ocular surface-positive culture rate in the treatment group with glaucoma eye drop usage was observed than in the control group without any treatment (Honda et al., 2011). Preservatives commonly used in eye drops have aroused attention. Travoprost and latanoprost are eye drops for glaucoma, but they both use a different preservative agent: the former is SofZia-preserved, and the latter is benzalkonium chloride-preserved. Bacterial samples were collected from conjunctiva sacs of patients who have been treated with travoprost or latanoprost for more than 1 year. The analysis showed that S. epidermidis present in the latanoprost group was significantly more resistant to LVFX, gatifloxacin, moxifloxacin, and tobramycin (Ohtani et al., 2017).
Trauma is an influential factor of OSM. The abundance of Pseudomonas fluorescens and Pseudomonas aeruginosa is significantly elevated in patients following a traumatic corneal ulcer when compared with healthy subjects (Kang et al., 2020). Dry eye disease (DED) occurs when tears cannot provide adequate lubrication and nourishment to the ocular surface. It is classified into two subtypes: aqueous tear-deficient dry eye (ADDE) and hyper-evaporative DED (Craig et al., 2017). Compared with the normal healthy population, there is significantly lower diversity and relative abundance of OSM in patients with ADDE (Andersson et al., 2021). Another study has compared the difference in OSM between DED and non-DED subjects, revealing depleted levels of Pseudomonas plecoglossicida, Pseudomonadaceae, Gammaproteobacteria, and Proteobacteria in DED eyes (Li et al., 2019). Further reports have indicated that lifestyle and systemic disease may alter OSM. For example, OSM in chronic alcoholism shows a significantly higher presence of Staphylococcus aureus (Gunduz et al., 2016). In patients with diabetes mellitus, Proteobacteria makes up a large proportion of OSM, whereas Firmicutes is the most common phylum of the OSM in healthy populations (Ham et al., 2018).
Acquired factors
Conjunctiva is a low-biomass tissue. Even a little DNA contamination can produce false-positive signals, especially after PCR amplification, so a rigorous contaminant-filtering step is needed to eliminate the “noise” from the environment overwhelming the target signal (Glassing et al., 2016). Moreover, it is hard to distinguish the bacteria that are resident on the ocular surface or accidentally brought there by finger, contact lens, or wind. Within the microbiota community, complicated interactions, including competition and cooperation, happen simultaneously and reach a state of dynamic equilibrium; therefore, the OSM profile may differ at every sampling time point (Wintermute and Silver, 2010; Kang et al., 2021). The sampling site also contributes to this variation as the distribution of OSM is inconsistent (Ozkan et al., 2019). In addition, it is crucial to distinguish whether the origin of the recovered samples is from viable or nonviable organisms. A harvested sample may be simply microbes perishing in the antimicrobial microenvironment, instead of a community of colonizing organisms (Zegans and Van Gelder, 2014). Also, the baseline state may be complicated by diet, age, and environment, as previously mentioned. Further research is needed to enhance previous findings by providing a much more detailed examination.
Relationship between OSM, immunity, and ophthalmic infectious disease
Crosstalk between OSM and innate immunity
OSM is reported to boost the local innate immunity, link it to adaptive immunity, and maintain a harmonious relationship between them, pathogenic microbials, and the host.
Corynebacterium mastitidis is a commensal organism residing on the ocular surface of humans and mice. In a mouse model, it triggers γδ T cells within the ocular mucosa to produce IL-17. IL-17 then activates downstream synthesis of antimicrobial substance into tears to strengthen protection from invasive Candida albicans and Pseudomonas aeruginosa, expounding the indispensable role of microbiota–T-cell interaction in ocular surface innate immunity (St Leger et al., 2017). A lack of bacterial colonization may weaken the corneal barrier as significantly greater corneal barrier disruption and more goblet cell loss were found within the lacrimal gland of the germ-free (GF) mice than in the conventional C57BL/6J mice (Wang et al., 2018).
Secretory IgA (SIgA), the predominant antibody in tears and the ocular surface, is a first-line defensive system in the ocular surface (Knop et al., 2008). In GF mice, protein and heavy-chain transcription of SIgA are significantly decreased on the ocular surface when compared with conventional mice. Similarly, both wild-type mice treated with ophthalmic gentamicin gel and GF mice show reduced gene expression of the downstream effectors of IL-17 (St Leger et al., 2017). Complement component 3 (C3) and C9 are important innate immune effectors, which decrease in eyewashes of GF mice compared with specific pathogen-free Swiss Webster mice (SPF SW mice) (Kugadas et al., 2016). These findings strongly suggest that OSM participates in ocular surface innate immunity to a great extent as either temporal interference in OSM via antibiotics or artificial depletion of OSM reduces the integrity of the ocular mucosal barrier and innate immunity.
Crosstalk between OSM and adaptive immunity
Interlukin-1 beta (IL-1ß) is a member of the interlukin-1 family. It connects innate immunity and adaptive immunity as it shows to increase nucleoprotein-specific CD4+ and CD8+ T-cell immunity (Van Den Eeckhout et al., 2021). It may serve as a possible driving adjuvant in Th-17 cell-mediated immune response (Wüthrich et al., 2013), and it is necessary for antigen-specific T-cell activation (Nambu et al., 2006). Kugadas et al. found significantly lower expression of corneal IL-1ß in P.aeruginosa PAO1-infected tissues from mice treated with local gentamycin. This illustrates that the existence of OSM could promote the release of IL-1ß during ocular infection (Kugadas et al., 2016).
Sjogren syndrome (SS) is an autoimmune disease attacking the exocrine glands. It leads to severe dryness of the mucosal surfaces. CD4+ T cells are involved in SS; overactive CD4+ T cells create glandular injury via the activation of B cells (Singh and Cohen, 2012; Brito-Zerón et al., 2016). Zaheer et al. compared CD25 knockout (CD25KO) mice, which spontaneously developed SS-like inflammation, in GF and conventional conditions. GF CD25KO mice had a higher total lymphocyte infiltration score, greater expression of IFN-γ and IL-12, and a higher frequency of autoreactive CD4+ T cells than conventional CD25KO mice. However, fecal transplantation from C57BL/6J mice reversed the phenotype of dry eyes and lowered the generation of pathogenetic CD4+ T cells in GF CD25KO mice. This study highlights that a lack of microbiota may accelerate the production of CD4+ T cells with greater pathogenicity and the immunomodulatory properties of microbiota. Furthermore, it implies that the microbiota in the gut may affect the ocular condition of its host (Zaheer et al., 2018).
In summary, OSM is involved in the release of antimicrobial molecular and signals, SIgA production, the regulation and promotion of the immune system, and the connection between native immunity and adaptive immunity.
Ophthalmic infectious disease and OSM
The knowledge of the characteristics of OSM in innate immunity and adaptive immunity has led to studies addressing the relationship between altered OSM and ophthalmic infectious disease.
Blepharitis is general inflammation along the edges of the eyelids (Lindsley et al., 2012). Increased relative abundances of Staphylococcus, Streptophyta, Corynebacterium, and Enhydrobacter were noted in the eyelash and tear samples from subjects with blepharitis, suggesting blepharitis may be caused by exposure to pollens, dusts, and soil particles (Lee et al., 2012). Previous reports indicated that an elevated proportion of skin microflora on the ocular surface may lead to blepharitis. This corresponds to Lee et al. (2012) as they found a significantly higher Staphylococcus proportion in subjects with blepharitis (Groden et al., 1991; Kulaçplu, 2001).
Dysbiosis in the ocular fungal microbiota is related to fungal keratitis (FK). Prashanthi et al. collected conjunctival swabs (SWs) from a healthy control (HC) group and additional corneal scrapings (CRs) from participants with FK. A significant difference in the Shannon index, Simpson index, and number of observed OTUs were noted between HC-SW and FK-CR groups. Ascomycota (mean abundance, 35.66%) and Basidiomycota (mean abundance, 37.05%) were the two most dominant phyla in the HC-SW. Ascomycota was the most dominant phyla in the FK-CR and FK-SW; however, the abundance was clearly increased (mean abundance was 88.36% and 69.99% in FK-CR and FK-SW, respectively). Basidiomycota was significantly lower in both FK-CR and FK-SW groups; it accounted for only ≤10.2%. Interaction networks were established based on pair-wise correlations. “Hub genera” refers to a genus interacting with more than 10 genera in the network, either positively or negatively. In HC-SW, Setosphaeria was the largest hub genera, which negatively interacted with 20 other genera, symbolizing its inhibitive role in the growth of Pseudozyma, Peniophora, Trichomonascus, and Talaromyces (Prashanthi et al., 2019).
Trachoma, caused by Chlamydia trachomatis, is the most common infectious cause of blindness. It is characterized by conjunctival inflammation, scarring, trichiasis, or entropion (Taylor et al., 2014). Zhou et al. recruited 210 residents in Gambia, 105 with healthy conjunctivae and 105 with clinical signs of trachoma; decreased diversity and increased abundance of Corynebacterium and Streptococcus were noted in the participants with signs of trachoma (Zhou et al., 2014).
Aforementioned studies explained the close connection between ophthalmic infectious disease and OSM; however, the interest lies in whether alterations or improvements in OSM can serve as a treatment strategy in ophthalmic infectious disease. Kugadas et al. indicated that GF mice are more susceptible to Pseudomonas aeruginosa-induced keratitis. This may be caused by commensal microbiota deficiency. Coagulase-negative Staphylococcus sp., the predominant bacterium in OSM, was topically applied to the ocular surface of GF mice. After 2 weeks, the mono-colonized GF mice showed restoration of resistance to P. aeruginosa and the same expression level of IL-1ß conjunctival transcription as Swiss Webster mice. The restoration of resistance toward pathogens and OSM homeostasis via topical application of commensal bacteria is an inspiring finding (Kugadas et al., 2016). In humans, topical or systemic administration of specific commensal bacteria to treat ophthalmic infectious disease has gathered great importance.
The gut–eye axis and potential of probiotics in treating ophthalmic infectious disease
Human gut microbiota (HGM) comprises more than 100 trillion microorganisms and can weigh up to 2 kg in an adult, which has coevolved in eclectic physiological responses, including digestion, metabolism of xenobiotics, antimicrobial peptide production, and immunomodulation by upregulation or downregulation of host gene expression (Fu et al., 2017). Up to 116 genes are significantly differentially expressed in GF mice and conventional mice (Nichols and Davenport, 2021). Components of HGM differ from person to person and can be influenced by diet, lifestyle, and medication treatment. Currently, microbiota colonized out of eyes, particularly in the gastrointestinal tract, are found to be related to several ophthalmic diseases, and such linkage is the so-called gut–eye axis (Clemente et al., 2012; Turner, 2018; Napolitano et al., 2021).
Astafurov et al. compared the oral cavity microbiota of a healthy population and that of patients with glaucoma via mouthwash specimens. Significantly higher bacterial loads are found in glaucoma cases, and linear discriminant analysis of DNA has confirmed differences in oral cavity microbiota of the two groups (Astafurov et al., 2014). HGM also participates in the modulation of age-related macular degeneration (AMD). Serotonin (5-HT) is a protective substance against retinal damage, and its synthesis is mainly regulated by gut spore-forming bacteria (Yano Jessica et al., 2015; Zhang and Davies, 2016). Corroborating outcomes are noted in another study, indicating that modifying HGM with a low-glycemia diet can delay the onset and progress of AMD features in animal models (Rowan et al., 2017).
Ophthalmic infectious disease is related to HGM. An increased abundance of the pathogenic fungi, Aspergillus and Malassezia, is noted in fecal samples collected from patients with bacterial keratitis (BK). The Shannon and Simpson indexes of HGM are significantly different between the BK and HC groups (Jayasudha et al., 2018). SPF SW mice are more susceptible to ocular P. aeruginosa infection and have increased corneal pathology with depletion of gut microbiota (Kugadas et al., 2016). In addition, a decline in SIgA expression in the tears has been noted in conventional SW mice upon oral antibiotic treatment (Kugadas and Gadjeva, 2016). These studies are indicative of the fact that the interference in gut microbiota may lead to decreased ocular immunity. It should not be neglected that the discrepancy between the HGM of the normal healthy population and that of patients with infectious ophthalmic disease was never limited in taxonomic abundance; functional differences in HGM between HC and FK patients were reported. A total of three KEGG pathways are significantly enriched in the FK group. Another six KEGG pathways are enriched in the HC group, including the biosynthesis of ansamycin and neomycin (Kalyana Chakravarthy et al., 2018).
The overuse of antibiotics has led the emergence of multidrug-resistant (MDR) bacteria; multidrug-resistant Pseudomonas aeruginosa (MDR-PA) and methicillin-resistant Staphylococcus aureus (MRSA) have been reported in ophthalmic infectious disease (Vazirani et al., 2015; Bispo et al., 2020). Therefore, an alternative therapy to antibiotics is required to overcome this issue. Numerous studies provide further demonstration by emphasizing the relationship between alterations in HGM and ophthalmic health. This clearly supports the collaborative contribution between OSM and HGM in the homeostasis of eyes. Novel therapies based on these findings have been studied in recent years. “Probiotics” are live nonpathogenic microorganisms administered to improve microbial balance, particularly in the gastrointestinal tract, which beneficially affects the host (Schrezenmeir and Vrese, 2001; Williams, 2010). They are expected to serve as brand new therapies for ophthalmic infectious disease via improving the HGM and adjusting the microenvironment in human eyes, thus ameliorating ophthalmic infection. A study has screened over 2,000 strains to select the bacterial strains that met 12 criteria including having specific biochemical activity, withstanding to low-pH environments, and exerting a beneficial impact on the host. Finally, two commercial probiotic strains, Lactobacillus rhamnosus GG and Lactobacillus acidophilus LA-1, were picked up. Following this, several strains belonging to Lactobacillus and Bifidobacterium have been confirmed as probiotics (Prasad et al., 1998; Picard et al., 2005; Lebeer et al., 2008).
Antimicrobial effect of probiotics to pathogens in ophthalmic infection
Probiotics can affect pathogens, as shown by current studies in the field. These highlight the antimicrobial capacity of probiotics toward pathogens that cause ophthalmic infection.
Biofilms are clusters of microorganisms that stick on the surface of plants or animals, being encased in an outer polymer layer that can be produced by microorganisms and able to evade the immune response or resist antibiotics (Hall-Stoodley et al., 2004). Bacillus spp. can form biofilm on the ocular surface. This was the target of Akova et al. They tested eight species of probiotics belonging to Lactobacillus for their anti-biofilm capacity; five of them significantly lowered biofilm formation compared with the control group (Akova et al., 2021).
A total of 17 culture-positive (Staphylococcus aureus in four isolates and S. epidermidis in 13 isolates) swabs were collected from patients with bacterial conjunctivitis. Antibiotic susceptibility profiling was performed, and all Staphylococcus spp. were resistant to oxacillin, penicillin G, and cephalexin. In total, six strains of probiotics belonging to Lactobacillus and Bifidobacterium were prepared to evaluate their antimicrobial potential. Interestingly, all the probiotics in this study expressed promising inhibition on bacterial growth, even on species with antibiotic resistance (Mohamed et al., 2020). Gonococcal conjunctivitis in neonates is caused by Neisseria gonorrhoeae colonizing in the maternal birth canal, which may result in severe vision impairment or blindness (Mallika et al., 2008). Biogenic substance of L. rhamnosus strain L60 could suppress the growth of Neisseria gonorrhoeae (Ruíz et al., 2015). Another pathogen of neonatal conjunctivitis is herpes simplex virus 2 (HSV-2), which infects neonates during vaginal birth (Brown et al., 2007). In a mammalian cell model, Lactobacillus crispatus prevented HSV-2 entry into cells by trapping the viral particles and blocking HSV-2 receptors (Mousavi et al., 2018).
Parasite infection also threatens human vision. Toxocariasis, caused by Toxocara canis infection, is a common zoonotic disease that has ophthalmic involvement, known as ocular larva migrans (Aydenizöz-Özkayhan et al., 2008; Chen et al., 2018). Traditionally, benzimidazole and steroids are the only remedy (Magnaval et al., 2001). However, a lower Toxocara canis burden was found in mice inoculated with Enterococcus faecalis CECT 7121 before infection compared with mice without inoculation (Basualdo et al., 2007).
A possible mechanism for these reports includes nutrition competition, immune modulation, or production of inhibitory substances reducing pathogen activity. Taken together, probiotics may become a new therapy for ocular toxocariasis.
Current application of probiotics in ophthalmic disease and limitations
Probiotics may be a promising therapy in future; however, several challenges are to be faced. Probiotic administration occurs via oral intake, fecal transplantation, and topical use. The first two are based on the gut–eye axis; expecting alterations in HGM may have positive or curative effects on ophthalmic disease. The final route is a more intuitive concept that directly changes OSM and modifies the ocular microenvironment.
The inhibition of pathogens can be caused by bacteriocin, ribosomally synthesized peptides, or proteins with antimicrobial activity produced by probiotics (Gálvez et al., 2007). Lactobacillus strains suppress bacterial growth by lowering the pH of their incubation medium with acetic and lactic acids (Fayol-Messaoudi et al., 2005). Commensal bacteria can inhibit pathogen growth through competition for nutrition and space (Miller and Iovieno, 2009; Kang et al., 2021). However, neither bacteriocin nor organic acids have been reported to approach eyes via systemic circulation or other routes. A previous study has shown higher expression of ansamycin and neomycin biosynthesis in healthy subjects than in patients with FK (Kalyana Chakravarthy et al., 2018). Nevertheless, whether these pathways are the key point in the inhibition of pathogen growth is unclear. Competition between commensal bacteria and pathogens may take place in adjacent areas alone. This is unlikely to suppress the growth of ocular pathogens directly from a distant organ. The oral administration or fecal transplantation of probiotics as new therapies for ophthalmic infectious disease treatment seems uncertain, and more research is required to pursue this line of investigation. For topical administration, probiotic eye drops have been developed for use in vernal keratoconjunctivitis (Iovieno et al., 2008) but not for infectious diseases.
In the field of utilizing probiotics as a new therapy for eye infection, much remains to be done. First, it is essential to investigate the precise mechanism and interaction between different probiotics and pathogens. Next, the “antimicrobial spectrum” of each strain should be established, for making it possible to select the most suitable probiotic strain depending on different pathogens. A key area for future research will be the comparison of the efficacy, clinical outcome, and cost-to-performance ratio between probiotics and existing antibiotics, and antiviral agents. Second, the positive effect of probiotics may only be asserted with a high number of viable cells reaching the ocular surface. Ameliorated manufacturing processes and storage ways are necessary as many probiotic bacteria may die during the manufacture and storage of products (Kailasapathy and Chin, 2000; Evivie et al., 2017). Third, the ocular surface contains a wide array of defense mechanisms against microorganisms. It is not clear whether probiotic bacteria can survive persistently. In addition, the ocular surface is exposed to the environment; therefore, numerous internal and external factors, including personal habits, contact lens wearing, eye rubbing, artificial tear usage, and systemic diseases, can alter its microenvironment and OSM state and then influence probiotic bacteria survival (Petrillo et al., 2020). Therefore, it is difficult to estimate the outcome of topically administered probiotic treatment for ophthalmic infectious disease. Last, although probiotics exert satisfying antimicrobial activities on different pathogens, the results are rarely confirmed in human clinical trials. More studies are urgently needed to confirm the efficacy and safety in human beings.
Conclusions and futural directions
With new sequencing technology, investigations of OSM are no longer limited to culture-dependent methods. The frequent fluctuation of ocular surface microenvironments and highly varied OSM per person mean that the existence of a core microbiota lacks empirical support. However, OSM modulates the immune system, inhibits microbial growth, and plays a crucial role in maintaining homeostasis. Different patterns in OSM and HGM have been reported in different ophthalmic infectious diseases; however, whether the alteration of microbiota is a cause or an outcome of infection is yet to be elucidated. Furthermore, some of current studies, despite being encouraging, did not utilize a filter when investigating OSM. Therefore, amelioration in methodology with a more rigorous or standardized filter is needed to exclude potential contaminations and make the studies more comparable. Futural studies should be alerted to. While research on these novel therapies is still at an early stage, probiotic application in ophthalmic infectious disease is a promising line of inquiry.
Author contributions
M-CC and EC contributed to the design and implementation of the research and the writing of the manuscript. All authors contributed equally and approved the final version of the manuscript.
Funding
This study was supported by grants from the Ministry of Science and Technology (MOST), Taiwan (MOST 109-2320-B- 002-051-MY2).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akova, B., Kivanç, S. A., and Kivanç, M. (2021). Antibiofilm effect of probiotic lactic acid bacteria against Bacillus spp obtained from the ocular surface. Eur. Rev. Med. Pharmacol. Sci. 25, 7799–7805. doi: 10.26355/eurrev_202112_27626
Andersson, J., Vogt, J. K., Dalgaard, M. D., Pedersen, O., Holmgaard, K., Heegaard, S., et al. (2021). Ocular surface microbiota in patients with aqueous tear-deficient dry eye. Ocul. Surf. 19, 210–217. doi: 10.1016/j.jtos.2020.09.003
Astafurov, K., Elhawy, E., Ren, L., Dong, C. Q., Igboin, C., Hyman, L., et al. (2014). Oral microbiome link to neurodegeneration in glaucoma. PLoS ONE. 9, e104416. doi: 10.1371/journal.pone.0104416
Aydenizöz-Özkayhan, M., Yagci, B. B., and Erat, S. (2008). The investigation of Toxocara canis eggs in coats of different dog breeds as a potential transmission route in human toxocariasis. Vet. Parasitol. 152, 94–100. doi: 10.1016/j.vetpar.2007.12.002
Basualdo, J., Sparo, M., Chiodo, P., Ciarmela, M., and Minvielle, M. (2007). Oral treatment with a potential probiotic (Enterococcus faecalis CECT 7121) appears to reduce the parasite burden of mice infected with Toxocara canis. Ann. Trop. Med. Parasitol. 101, 559–562. doi: 10.1179/136485907X193824
Bauer, D., Schmitz, A., Van Rooijen, N., Steuhl, K. P., and Heiligenhaus, A. (2002). Conjunctival macrophage-mediated influence of the local and systemic immune response after corneal herpes simplex virus-1 infection. Immunology. 107, 118–128. doi: 10.1046/j.1365-2567.2002.01477.x
Bispo, P. J. M., Ung, L., Chodosh, J., and Gilmore, M. S. (2020). Hospital-Associated Multidrug-Resistant MRSA Lineages Are Trophic to the Ocular Surface and Cause Severe Microbial Keratitis. Front. Public Health. 8. doi: 10.3389/fpubh.2020.00204
Brito-Zerón, P., Baldini, C., Bootsma, H., Bowman, S. J., Jonsson, R., Mariette, X., et al. (2016). Sjögren syndrome. Nat. Rev. Dis. Primers. 2, 16047. doi: 10.1038/nrdp.2016.47
Brown, E. L., Gardella, C., Malm, G., Prober, C. G., Forsgren, M., Krantz, E. M., et al. (2007). Effect of maternal herpes simplex virus (HSV) serostatus and HSV type on risk of neonatal herpes. Acta Obstet. Gynecol. Scand. 86, 523–529. doi: 10.1080/00016340601151949
Byrd, A. L., Belkaid, Y., and Segre, J. A. (2018). The human skin microbiome. Nat. Rev. Microbiol. 16, 143–155. doi: 10.1038/nrmicro.2017.157
Cavuoto, K. M., Banerjee, S., Miller, D., and Galor, A. (2018). Composition and comparison of the ocular surface microbiome in infants and older children. Transl. Vis. Sci. Technol. 7, 16. doi: 10.1167/tvst.7.6.16
Cavuoto, K. M., Galor, A., and Banerjee, S. (2019). Anatomic characterization of the ocular surface microbiome in children. Microorganisms. 7. doi: 10.3390/microorganisms7080259
Celebi, A. R. C., and Onerci Celebi, O. (2021). The effect of topical ocular moxifloxacin on conjunctival and nasal mucosal flora. Sci. Rep. 11, 13782. doi: 10.1038/s41598-021-93233-5
Chen, J., Liu, Q., Liu, G. H., Zheng, W. B., Hong, S. J., Sugiyama, H., et al. (2018). Toxocariasis: a silent threat with a progressive public health impact. Infect Dis. Poverty. 7, 59. doi: 10.1186/s40249-018-0437-0
Clemente, J. C., Ursell, L. K., Parfrey, L. W., and Knight, R. (2012). The impact of the gut microbiota on human health: an integrative view. Cell. 148, 1258–1270. doi: 10.1016/j.cell.2012.01.035
Craig, J. P., Nichols, K. K., Akpek, E. K., Caffery, B., Dua, H. S., Joo, C. K., et al. (2017). TFOS DEWS II definition and classification report. Ocul. Surf. 15, 276–283. doi: 10.1016/j.jtos.2017.05.008
Dave, S. B., Toma, H. S., and Kim, S. J. (2013). Changes in ocular flora in eyes exposed to ophthalmic antibiotics. Ophthalmology. 120, 937–941. doi: 10.1016/j.ophtha.2012.11.005
De Sordi, L., Lourenço, M., and Debarbieux, L. (2019). The battle within: interactions of bacteriophages and bacteria in the gastrointestinal tract. Cell Host Microbe. 25, 210–218. doi: 10.1016/j.chom.2019.01.018
Doan, T., Akileswaran, L., Andersen, D., Johnson, B., Ko, N., Shrestha, A., et al. (2016). Paucibacterial microbiome and resident DNA virome of the healthy conjunctiva. Invest. Ophthalmol. Vis. Sci. 57, 5116–5126. doi: 10.1167/iovs.16-19803
Dong, Q., Brulc, J. M., Iovieno, A., Bates, B., Garoutte, A., Miller, D., et al. (2011). Diversity of bacteria at healthy human conjunctiva. Invest. Ophthalmol. Vis. Sci. 52, 5408–5413. doi: 10.1167/iovs.10-6939
Erridge, C. (2010). Endogenous ligands of TLR2 and TLR4: agonists or assistants? J. Leukoc. Biol. 87, 989–999. doi: 10.1189/jlb.1209775
Erridge, C., Bennett-Guerrero, E., and Poxton, I. R. (2002). Structure and function of lipopolysaccharides. Microbes Infect. 4, 837–851. doi: 10.1016/S1286-4579(02)01604-0
Evivie, S. E., Huo, G. C., Igene, J. O., and Bian, X. (2017). Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutr. Res. 61, 1318034. doi: 10.1080/16546628.2017.1318034
Fayol-Messaoudi, D., Berger, C. N., Coconnier-Polter, M.-H., Moal, V., L-, L., et al. (2005). pH-, Lactic acid-, and non-lactic acid-dependent activities of probiotic Lactobacilli against Salmonella enterica Serovar TyphimuriumApplied and Environmental Microbiology. 71, 6008–6013. doi: 10.1128/AEM.71.10.6008-6013.2005
Fu, Z. D., Selwyn, F. P., Cui, J. Y., and Klaassen, C. D. (2017). RNA-seq profiling of intestinal expression of xenobiotic processing genes in germ-free mice. Drug Metab. Dispos. 45, 1225–1238. doi: 10.1124/dmd.117.077313
Galletti, J. G., Guzmán, M., and Giordano, M. N. (2017). Mucosal immune tolerance at the ocular surface in health and disease. Immunology. 150, 397–407. doi: 10.1111/imm.12716
Gálvez, A., Abriouel, H., López, R. L., and Omar, N. B. (2007). Bacteriocin-based strategies for food biopreservation. Int. J. Food Microbiol. 120, 51–70. doi: 10.1016/j.ijfoodmicro.2007.06.001
Glassing, A., Dowd, S. E., Galandiuk, S., Davis, B., and Chiodini, R. J. (2016). Inherent bacterial DNA contamination of extraction and sequencing reagents may affect interpretation of microbiota in low bacterial biomass samples. Gut Pathog. 8, 24. doi: 10.1186/s13099-016-0103-7
Gordon, S. (2002). Pattern recognition receptors: doubling up for the innate immune response. Cell. 111, 927–930. doi: 10.1016/S0092-8674(02)01201-1
Graham, J. E., Moore, J. E., Jiru, X., Moore, J. E., Goodall, E. A., Dooley, J. S., et al. (2007). Ocular pathogen or commensal: a PCR-based study of surface bacterial flora in normal and dry eyes. Invest. Ophthalmol. Vis. Sci. 48, 5616–5623. doi: 10.1167/iovs.07-0588
Groden, L. R., Murphy, B., Rodnite, J., and Genvert, G. I. (1991). Lid flora in blepharitis. Cornea. 10, 50–53. doi: 10.1097/00003226-199110010-00010
Gunduz, G., Gunduz, A., Polat, N., Cumurcu, B. E., and Yakupogullari, Y. (2016). The effect of chronic alcoholism on the conjunctival flora. Curr Eye Res. 41, 734–739. doi: 10.3109/02713683.2015.1056805
Hall-Stoodley, L., Costerton, J. W., and Stoodley, P. (2004). Bacterial biofilms: from the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108. doi: 10.1038/nrmicro821
Ham, B., Hwang, H. B., Jung, S. H., Chang, S., Kang, K. D., Kwon, M. J., et al. (2018). Distribution and diversity of ocular microbial communities in diabetic patients compared with healthy subjects. Curr. Eye Res. 43, 314–324. doi: 10.1080/02713683.2017.1406528
Hayashi, F., Smith, K. D., Ozinsky, A., Hawn, T. R., Yi, E. C., Goodlett, D. R., et al. (2001). The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature. 410, 1099–1103. doi: 10.1038/35074106
Honda, R., Toshida, H., Suto, C., Fujimaki, T., Kimura, T., Ohta, T., et al. (2011). Effect of long-term treatment with eyedrops for glaucoma on conjunctival bacterial flora. Infect. Drug Resist. 4, 191–196. doi: 10.2147/IDR.S24250
Horwitz, D. A., Fahmy, T. M., Piccirillo, C. A., and La Cava, A. (2019). Rebalancing immune homeostasis to treat autoimmune diseases. Trends Immunol. 40, 888–908. doi: 10.1016/j.it.2019.08.003
Idzko, M., Hammad, H., van Nimwegen, M., Kool, M., Willart, M. A. M., Muskens, F., et al. (2007). Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritic cells. Nat. Med. 13, 913–919. doi: 10.1038/nm1617
Iovieno, A., Lambiase, A., Sacchetti, M., Stampachiacchiere, B., Micera, A., Bonini, S., et al. (2008). Preliminary evidence of the efficacy of probiotic eye-drop treatment in patients with vernal keratoconjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 246, 435–441. doi: 10.1007/s00417-007-0682-6
Isenberg, S. J., Apt, L., Yoshimori, R., Ward, Mc, Carty, J., and Alvarez, S. R. (1988). Source of the conjunctival bacterial flora at birth and implications for ophthalmia neonatorum prophylaxis. Am J Ophthalmol. 106, 458–462. doi: 10.1016/0002-9394(88)90883-5
Jayasudha, R., Chakravarthy, S. K., Prashanthi, G. S., Sharma, S., Garg, P., Murthy, S. I., et al. (2018). Alterations in gut bacterial and fungal microbiomes are associated with bacterial Keratitis, an inflammatory disease of the human eye. J. Biosci. 43, 835–856. doi: 10.1007/s12038-018-9798-6
Kailasapathy, K., and Chin, J. (2000). Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp. Immunol. Cell Biol. 78, 80–88. doi: 10.1046/j.1440-1711.2000.00886.x
Kalyana Chakravarthy, S., Jayasudha, R., Ranjith, K., Dutta, A., Pinna, N. K., Mande, S. S., et al. (2018). Alterations in the gut bacterial microbiome in fungal Keratitis patients. PLoS ONE. 13, e0199640. doi: 10.1371/journal.pone.0199640
Kang, Y., Lin, S., Ma, X., Che, Y., Chen, Y., Wan, T., et al. (2021). Strain heterogeneity, cooccurrence network, taxonomic composition and functional profile of the healthy ocular surface microbiome. Eye Vision. 8, 6. doi: 10.1186/s40662-021-00228-4
Kang, Y., Zhang, H., Hu, M., Ma, Y., Chen, P., Zhao, Z., et al. (2020). Alterations in the ocular surface microbiome in traumatic corneal ulcer patients. Invest. Ophthalmol. Vis. Sci. 61, 35. doi: 10.1167/iovs.61.6.35
Knop, E., Knop, N., and Claus, P. (2008). Local production of secretory IgA in the eye-associated lymphoid tissue (EALT) of the normal human ocular surface. Invest. Ophthalmol. Vis. Sci. 49, 2322–2329. doi: 10.1167/iovs.07-0691
Kugadas, A., Christiansen, S. H., Sankaranarayanan, S., Surana, N. K., Gauguet, S., Kunz, R., et al. (2016). Impact of microbiota on resistance to ocular pseudomonas aeruginosa-induced keratitis. PLoS Pathog. 12, e1005855. doi: 10.1371/journal.ppat.1005855
Kugadas, A., and Gadjeva, M. (2016). Impact of microbiome on ocular health. Ocul. Surf. 14, 342–349. doi: 10.1016/j.jtos.2016.04.004
Kulaçplu, D. N., Özbek, A., Uslu, H., Şahín, F., Güllülü, G., Koçer, I., et al. (2001). Comparative lid flora in anterior blepharitis. Turk. J. Med. Sci. 31, 359–63. Available online at: https://journals.tubitak.gov.tr/medical/vol31/iss4/12
Lebeer, S., Vanderleyden, J., and Keersmaecker, D. (2008). Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 72, 728–64. doi: 10.1128/MMBR.00017-08
Lee, P. W., Jun, A. K., and Cho, B. C. A. (1989). study of microbial flora of conjunctival sac in newborns. Korean J. Ophthalmol. 3, 38–41. doi: 10.3341/kjo.1989.3.1.38
Lee, S. H., Oh, D. H., Jung, J. Y., Kim, J. C., and Jeon, C. O. (2012). Comparative ocular microbial communities in humans with and without blepharitis. Investig. Ophthalmol. Vis. Sci. 53, 5585–5593. doi: 10.1167/iovs.12-9922
Li, Z., Gong, Y., Chen, S., Li, S., Zhang, Y., Zhong, H., et al. (2019). Comparative portrayal of ocular surface microbe with and without dry eye. J. Microbiol. 57, 1025–1032. doi: 10.1007/s12275-019-9127-2
Lindsley, K., Matsumura, S., Hatef, E., and Akpek, E. K. (2012). Interventions for chronic blepharitis. Cochrane Database Syst Rev. 2012, Cd005556. doi: 10.1002/14651858.CD005556.pub2
Magnaval, J. F., Glickman, L. T., Dorchies, P., and Morassin, B. (2001). Highlights of human toxocariasis. Korean J. Parasitol. 39, 1–11. doi: 10.3347/kjp.2001.39.1.1
Mallika, P., Asok, T., Faisal, H., Aziz, S., Tan, A., Intan, G., et al. (2008). Neonatal conjunctivitis - a review. Malays Fam. Physician. 3, 77–81.
Mantelli, F., Mauris, J., and Argüeso, P. (2013). The ocular surface epithelial barrier and other mechanisms of mucosal protection: from allergy to infectious diseases. Curr. Opin. Allergy Clin. Immunol. 13, 563–568. doi: 10.1097/ACI.0b013e3283645899
McDermott, A. M. (2013). Antimicrobial compounds in tears. Exp. Eye Res. 117, 53–61. doi: 10.1016/j.exer.2013.07.014
Miller, D., and Iovieno, A. (2009). The role of microbial flora on the ocular surface. Curr. Opin. Allergy Clin. Immunol. 9, 466–470. doi: 10.1097/ACI.0b013e3283303e1b
Mogensen, T. H. (2009). Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 22, 240–273. doi: 10.1128/CMR.00046-08
Mohamed, S., Elmohamady, M. N., Abdelrahman, S., Amer, M. M., and Abdelhamid, A. G. (2020). Antibacterial effects of antibiotics and cell-free preparations of probiotics against Staphylococcus aureus and Staphylococcus epidermidis associated with conjunctivitis. Saudi Pharmaceut. J. 28, 1558–1565. doi: 10.1016/j.jsps.2020.10.002
Mousavi, E., Makvandi, M., Teimoori, A., Ataei, A., Ghafari, S., Samarbaf-Zadeh, A., et al. (2018). Antiviral effects of Lactobacillus crispatus against HSV-2 in mammalian cell lines. Journal of the Chinese Medical Association. 81, 262–267. doi: 10.1016/j.jcma.2017.07.010
Nambu, A., Nakae, S., and Iwakura, Y. (2006). IL-1β, but not IL-1α, is required for antigen-specific T cell activation and the induction of local inflammation in the delayed-type hypersensitivity responses. Int. Immunol. 18, 701–712. doi: 10.1093/intimm/dxl007
Napolitano, P., Filippelli, M., Davinelli, S., Bartollino, S., dell'Omo, R., and Costagliola, C. (2021). Influence of gut microbiota on eye diseases: an overview. Ann Med. 53, 750–761. doi: 10.1080/07853890.2021.1925150
Nichols, R. G., and Davenport, E. R. (2021). The relationship between the gut microbiome and host gene expression: a review. Hum. Genet. 140, 747–760. doi: 10.1007/s00439-020-02237-0
Ohtani, S., Shimizu, K., Nejima, R., Kagaya, F., Aihara, M., Iwasaki, T., et al. (2017). Conjunctival bacteria flora of glaucoma patients during long-term administration of prostaglandin analog drops. Investig. Ophthalmol. Vis. Sci. 58, 3991–3996. doi: 10.1167/iovs.16-20853
Ono, T., Nejima, R., Iwasaki, T., Mori, Y., Noguchi, Y., Yagi, A., et al. (2017). Long-term effects of cataract surgery with topical levofloxacin on ocular bacterial flora. J. Cataract Refract. Surg. 43, 1129–1134. doi: 10.1016/j.jcrs.2017.06.037
Ozkan, J., Nielsen, S., Diez-Vives, C., Coroneo, M., Thomas, T., Willcox, M., et al. (2017). Temporal stability and composition of the ocular surface microbiome. Sci. Rep. 7, 9880. doi: 10.1038/s41598-017-10494-9
Ozkan, J., Willcox, M., Wemheuer, B., Wilcsek, G., Coroneo, M., Thomas, T., et al. (2019). Biogeography of the human ocular microbiota. Ocul. Surf. 17, 111–118. doi: 10.1016/j.jtos.2018.11.005
Petrillo, F., Pignataro, D., Lavano, M. A., Santella, B., Folliero, V., Zannella, C., et al. (2020). Current evidence on the ocular surface microbiota and related diseases. Microorganisms. 8. doi: 10.3390/microorganisms8071033
Picard, C., Fioramonti, J., Francois, A., Robinson, T., Neant, F., Matuchansky, C., et al. (2005). Review article: bifidobacteria as probiotic agents – physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 22, 495–512. doi: 10.1111/j.1365-2036.2005.02615.x
Prasad, J., Gill, H., Smart, J., and Gopal, P. K. (1998). Selection and characterisation of lactobacillus and bifidobacterium strains for use as probiotics. Int. Dairy J. 8, 993–1002. doi: 10.1016/S0958-6946(99)00024-2
Prashanthi, G. S., Jayasudha, R., Chakravarthy, S. K., Padakandla, S. R., SaiAbhilash, C. R., Sharma, S., et al. (2019). Alterations in the ocular surface fungal microbiome in fungal keratitis patients. Microorganisms. 7. doi: 10.3390/microorganisms7090309
Ray, A., Cot, M., Puzo, G., Gilleron, M., and Nigou, J. (2013). Bacterial cell wall macroamphiphiles: Pathogen-/microbe-associated molecular patterns detected by mammalian innate immune system. Biochimie. 95, 33–42. doi: 10.1016/j.biochi.2012.06.007
Rowan, S., Jiang, S., Korem, T., Szymanski, J., Chang, M.-L., et al. (2017). Involvement of a gut–retina axis in protection against dietary glycemia-induced age-related macular degeneration. Proc. Nat. Acad. Sci. 114, E4472–E81. doi: 10.1073/pnas.1702302114
Ruíz, F. O., Pascual, L., Giordano, W., and Barberis, L. (2015). Bacteriocins and other bioactive substances of probiotic lactobacilli as biological weapons against Neisseria gonorrhoeae. Pathog. Dis. 73. doi: 10.1093/femspd/ftv013
Schrezenmeir, J., and Vrese, M. (2001). Probiotics, prebiotics, and synbiotics–approaching a definition. Am. J. Clin Nutr. 73, 361s−4s. doi: 10.1093/ajcn/73.2.361s
Shin, H., Price, K., Albert, L., Dodick, J., Park, L., Dominguez-Bello, M. G., et al. (2016). Changes in the Eye Microbiota Associated with Contact Lens Wearing. MBio. 7, e00198. doi: 10.1128/mBio.00198-16
Singh, N., and Cohen, P. L. (2012). The T cell in Sjogren's syndrome: force majeure, not spectateur. J. Autoimmun. 39, 229–233. doi: 10.1016/j.jaut.2012.05.019
St Leger, A. J., Desai, J. V., Drummond, R. A., Kugadas, A., Almaghrabi, F., Silver, P., et al. (2017). An ocular commensal protects against corneal infection by driving an interleukin-17 response from mucosal γδ T cells. Immunity. 47, 148–58.e5. doi: 10.1016/j.immuni.2017.06.014
Suzuki, T., Sutani, T., Nakai, H., Shirahige, K., and Kinoshita, S. (2020). The microbiome of the meibum and ocular surface in healthy subjects. Invest. Ophthalmol. Vis. Sci. 61, 18. doi: 10.1167/iovs.61.2.18
Ta, C. N., Chang, R. T., Singh, K., Egbert, P. R., Shriver, E. M., Blumenkranz, M. S., et al. (2003). Antibiotic resistance patterns of ocular bacterial flora: a prospective study of patients undergoing anterior segment surgery. Ophthalmology. 110, 1946–1951. doi: 10.1016/S0161-6420(03)00735-8
Takeuchi, O., and Akira, S. (2010). Pattern recognition receptors and inflammation. Cell. 140, 805–820. doi: 10.1016/j.cell.2010.01.022
Taylor, H. R., Burton, M. J., Haddad, D., West, S., and Wright, H. (2014). Trachoma. Lancet. 384, 2142–2152. doi: 10.1016/S0140-6736(13)62182-0
Turnbaugh, P. J., Ley, R. E., Hamady, M., Fraser-Liggett, C. M., Knight, R., Gordon, J. I., et al. (2007). The human microbiome project. Nature. 449, 804–810. doi: 10.1038/nature06244
Turner, P. V. (2018). The role of the gut microbiota on animal model reproducibility. Animal Model Exp Med. 1, 109–115. doi: 10.1002/ame2.12022
Ueta, M., Nochi, T., Jang, M. H., Park, E. J., Igarashi, O., Hino, A., et al. (2004). Intracellularly expressed TLR2s and TLR4s contribution to an immunosilent environment at the ocular mucosal epithelium. J. Immunol. 173, 3337–3347. doi: 10.4049/jimmunol.173.5.3337
Van Den Eeckhout, B., Tavernier, J., and Gerlo, S. (2021). Interleukin-1 as innate mediator of T cell immunity. Front. Immunol. (2021), 11. doi: 10.3389/fimmu.2020.621931
Vazirani, J., Wurity, S., and Ali, M. H. (2015). Multidrug-resistant pseudomonas aeruginosa keratitis: risk factors, clinical characteristics, and outcomes. Ophthalmol. 122, 2110–2114. doi: 10.1016/j.ophtha.2015.06.007
Wang, C., Zaheer, M., Bian, F., Quach, D., Swennes, A. G., Britton, R. A., et al. (2018). Sjögren-like lacrimal keratoconjunctivitis in germ-free. Mice Int J. Mol Sci. 19 doi: 10.3390/ijms19020565
Wang, Y., Chen, H., Xia, T., and Huang, Y. (2020). Characterization of fungal microbiota on normal ocular surface of humans. Clin. Microbiol. Infect. 26, 123.e9–e13. doi: 10.1016/j.cmi.2019.05.011
Wen, X., Miao, L., Deng, Y., Bible, P. W., Hu, X., Zou, Y., et al. (2017). The influence of age and sex on ocular surface microbiota in healthy adults. Invest. Ophthalmol. Vis. Sci. 58, 6030–6037. doi: 10.1167/iovs.17-22957
Williams, N. T. (2010). Probiotics. Am. J. Health-System Pharm. 67, 449–458. doi: 10.2146/ajhp090168
Wintermute, E. H., and Silver, P. A. (2010). Dynamics in the mixed microbial concourse. Genes Dev. 24, 2603–2614. doi: 10.1101/gad.1985210
Wüthrich, M., LeBert, V., Galles, K., Hu-Li, J., Ben-Sasson, S. Z., Paul, W. E., et al. (2013). Interleukin 1 enhances vaccine-induced antifungal T-helper 17 cells and resistance against Blastomyces dermatitidis infection. J. Infect. Dis. 208, 1175–82. doi: 10.1093/infdis/jit283
Yano Jessica, M., Yu, K., Donaldson Gregory, P., Shastri Gauri, G., Ann, P., Ma, L., et al. (2015). Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 161, 264–276. doi: 10.1016/j.cell.2015.02.047
Yin, V. T., Weisbrod, D. J., Eng, K. T., Schwartz, C., Kohly, R., Mandelcorn, E., et al. (2013). Antibiotic resistance of ocular surface flora with repeated use of a topical antibiotic after intravitreal injection. JAMA Ophthalmol. 131, 456–461. doi: 10.1001/jamaophthalmol.2013.2379
Zaheer, M., Wang, C., Bian, F., Yu, Z., Hernandez, H., Souza, R. G., et al. (2018). Protective role of commensal bacteria in Sjögren Syndrome. J Autoimmun. 93, 45–56. doi: 10.1016/j.jaut.2018.06.004
Zaura, E., Keijser, B. J., Huse, S. M., and Crielaard, W. (2009). Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 9, 259. doi: 10.1186/1471-2180-9-259
Zegans, M. E., and Van Gelder, R. N. (2014). Considerations in understanding the ocular surface microbiome. Am. J. Ophthalmol. 158, 420–422. doi: 10.1016/j.ajo.2014.06.014
Zhang, H., Zhao, F., Hutchinson, D. S., Sun, W., Ajami, N. J., Lai, S., et al. (2017). Conjunctival microbiome changes associated with soft contact lens and orthokeratology lens wearing. Invest. Ophthalmol. Vis. Sci. 58, 128–136. doi: 10.1167/iovs.16-20231
Zhang, J., Xu, K., Ambati, B., and Fu-Shin, X. Y. (2003). Toll-like receptor 5-mediated corneal epithelial inflammatory responses to Pseudomonas aeruginosa flagellin. Investig. Ophthalmol. Vis. Sci. 44, 4247–4254. doi: 10.1167/iovs.03-0219
Zhang, L. S., and Davies, S. S. (2016). Microbial metabolism of dietary components to bioactive metabolites: opportunities for new therapeutic interventions. Genome Med. 8, 46. doi: 10.1186/s13073-016-0296-x
Zhou, Y., Holland, M. J., Makalo, P., Joof, H., Roberts, C. H., Mabey, D. C., et al. (2014). The conjunctival microbiome in health and trachomatous disease: a case control study. Genome Med. 6, 99. doi: 10.1186/s13073-014-0099-x
Zhu, B., Wang, X., and Human, L. i. L. (2010). gut microbiome: the second genome of human body. Protein Cell. 1, 718–725. doi: 10.1007/s13238-010-0093-z
Keywords: ocular surface microbiota, ophthalmic infectious disease, gut–eye axis, probiotic, microbiome
Citation: Chiang M-C and Chern E (2022) Ocular surface microbiota: Ophthalmic infectious disease and probiotics. Front. Microbiol. 13:952473. doi: 10.3389/fmicb.2022.952473
Received: 25 May 2022; Accepted: 01 August 2022;
Published: 19 August 2022.
Edited by:
Santi M. Mandal, Indian Institute of Technology Kharagpur, IndiaReviewed by:
Suman Saha, Priyamvada Birla Aravind Eye Hospital, IndiaJerome Ozkan, University of New South Wales, Australia
Copyright © 2022 Chiang and Chern. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Edward Chern, edchern@ntu.edu.tw