Microbial Therapeutics Designed for Infant Health
 Claire Watkins
Claire Watkins Catherine Stanton
Catherine Stanton C. Anthony Ryan
C. Anthony Ryan R. Paul Ross
R. Paul Ross- 1APC Microbiome Institute, University College Cork, Cork, Ireland
- 2Teagasc Food Research Centre, Fermoy, Ireland
- 3School of Microbiology, University College Cork, Cork, Ireland
- 4Department of Neonatology, Cork University Maternity Hospital, Cork, Ireland
- 5School of Science, Engineering and Food Science, University College Cork, Cork, Ireland
Acknowledgment of the gut microbiome as a vital asset to health has led to multiple studies attempting to elucidate its mechanisms of action. During the first year of life, many factors can cause fluctuation in the developing gut microbiome. Host genetics, maternal health status, mode of delivery, gestational age, feeding regime, and perinatal antibiotic usage, are known factors which can influence the development of the infant gut microbiome. Thus, the microbiome of vaginally born, exclusively breastfed infants at term, with no previous exposure to antibiotics, either directly or indirectly from the mother, is to be considered the “gold standard.” Moreover, the use of prebiotics as an aid for the development of a healthy gut microbiome is equally as important in maintaining gut homeostasis. Breastmilk, a natural prebiotic source, provides optimal active ingredients for the growth of beneficial microbial species. However, early life disorders such as necrotising enterocolitis, childhood obesity, and even autism have been associated with an altered/disturbed gut microbiome. Subsequently, microbial therapies have been introduced, in addition to suitable prebiotic ingredients, which when administered, may aid in the prevention of a microbial disturbance in the gastrointestinal tract. The aim of this mini-review is to highlight the beneficial effects of different probiotic and prebiotic treatments in early life, with particular emphasis on the different conditions which negatively impact microbial colonisation at birth.
Introduction
From birth through to the initial stages of weaning, intestinal microbial composition has a significant impact on infant gut health. Recent advances in culture-independent sequencing technologies has allowed for the identification of key microbial species involved in the initial colonization process, including those facultative anaerobes such as Streptococcus, Staphylococcus, and Enterobacter spp. (1, 2). Mode of delivery and feeding regime are two important factors which influence microbial colonization at birth (Figure 1). Host genetics may also impact development of the gut microbiome, with recent studies focusing on similar microbial patterns between monozygotic twin pairs and their fraternal siblings (3, 4). Indeed, the duration of breast feeding and introduction of formula feed can play a significant role in shaping the gut microbiome (5–7). Thus, it is imperative that we understand how the introduction of particular microbial species and prebiotic additives may restore balance and ameliorate the effects associated with gastrointestinal (GI) disorders.
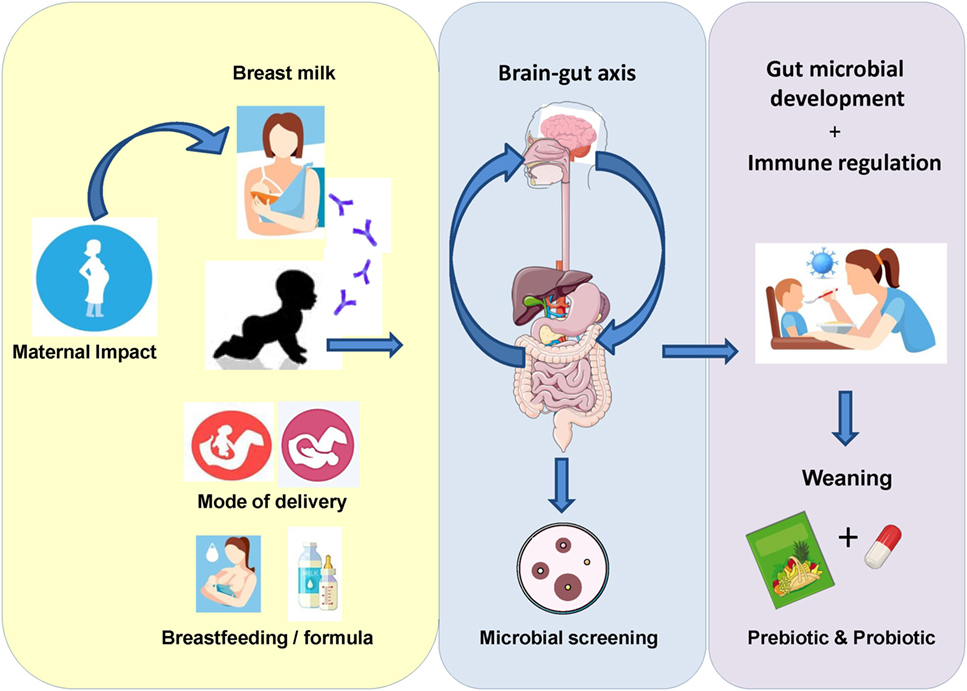
Figure 1. Initial exposure to the microbial environment surrounding the infant can have a significant impact on gut microbiota development. External factors, such as maternal health status, mode of delivery, gestational age, and feeding regime, can impact the colonization and flux of microorganisms during this critical period in life. Subsequently, multiple studies have begun to focus on how these factors can affect the gut microbiome in early life. Moreover, in order to improve the health status of the infant gut, current focus is on the effect of probiotics and prebiotics in terms of their potential multifaceted health benefits. The current mini-review outlines a number of studies where either pro- or pre-biotics were utilized as a microbial therapeutic to improve infant health.
GI Microbial Development at Birth
Many studies have begun to focus on the development of the infant gut microbiome over time (8–10). A study by our group found that in full-term (FT) cesarean delivered infants, an increased fecal abundance of Firmicutes and lower abundance of Actinobacteria was evident after the first week of life; however, the gut microbiota of preterm (PT) infants displayed a significantly greater abundance of Proteobacteria compared to the FT infant group. Interestingly, the gut microbiota profile of FT cesarean delivered infants resembled that of the vaginally delivered infants at 8 weeks of life (1). In terms of gestational age at birth, the PT infant gut has previously been characterized by delayed microbial colonization, with reduced levels of anaerobic taxa such as Bifidobacterium and Bacteroides (11, 12). Indeed, gut microbiota development in PT infants has been found to correlate with the infant’s postconceptual age (13). Moreover, Stewart et al. (14) described the impact of delivery mode on the PT gut microbiome and found no significant change in microbial diversity during the first 100 days of life.
Maternal–Infant Transmission
The acquisition of microbial strains may occur through multiple different pathways. For example, the administration of the probiotic Lactobacillus rhamnosus GG in a small subset of pregnant woman (between 30 and 36 weeks gestational age) found that maternal–infant transmission was successful and identified the strain in the feces of the infant cohort 6 months after birth (15). Indeed, maternal–infant transmission of mothers’ lactobacilli predominantly occurs when the infant is delivered vaginally (16, 17). It is understood that the vaginal tract harbors these lactobacilli to reduce the pH of the intestinal milieu and prevent the growth of potentially pathogenic microorganisms in the infant gut.
Interestingly, a number of studies have begun to examine microbial communities present within breastmilk. Nine genera have previously been identified as part of the “core” breastmilk microbiome, including Streptococcus, Staphylococcus, Serratia, Pseudomonas, Corynebacteria, Ralstonia, Propionibacterium, Sphingomonas, and Bradyrhizobiaceae (18). Several other studies have also identified the horizontal transfer of Lactobacillus, Staphylococcus, Enterococcus, and Bifidobacterium spp., from breastmilk to the infant gut (19–21). In a more recent study, Murphy et al. (22) reported the presence of 12 dominant genera in the breastmilk of lactating mothers. Results from this study described a number of frequently shared taxa, including Bifidobacterium, Lactobacillus, Staphylococcus, and Enterococcus, common in both breastmilk and infant feces during the first 3 months of life. Moreover, culture-dependent analysis identified Bifidobacterium breve and Lactobacillus plantarum present in both breastmilk and infant feces. Indeed, similar studies have also identified genomic patterns of Bifidobacterium and Lactobacillus spp. present in breastmilk and corresponding infant feces (23–25).
Recent studies have also begun to focus on microbial colonization which may occur in utero. Isolation of microorganisms from the umbilical cord blood of cesarean delivered infants (26), as well as the detection of bacteria in “first-pass” meconium (27, 28), suggest that the fetus may be colonized at a low abundance prior to exiting the womb. Moreover, research focused on the placental microbiome has found significant correlations between the placental and oral microbial communities (29). However, supporting evidence for the existence of a distinct placental microbiome is currently lacking (30). Thus, microbial colonization of the infant gut may be strongly influenced by the maternal microbiome, originating from several different niches, including the vaginal tract, breastmilk, and possibly the placenta.
Throughout the remainder of this mini-review, a number of studies will be discussed regarding prebiotic and probiotic treatments to prevent and/or treat conditions linked to GI health in early life.
Prebiotics
Human Milk Oligosaccharides (HMOs)
The current definition of prebiotics defined by Gibson et al. (31) describes “selectively fermented ingredients that result in specific changes, in the composition and/or activity of the GI microbiota, thus conferring benefit(s) upon host health.”
Human breastmilk is a natural prebiotic source which contains essential nutrients and growth factors required for development of a healthy gut microbiome. Selective proliferation of healthy intestinal bacteria is thought to be just one of the multiple benefits of exclusive breast feeding, in addition to the nutrient supply of HMOs and glycoconjugates it provides (32). As HMOs are not digested by the infant themselves, they reach the colon intact and act as an essential substrate for the growth of beneficial Bifidobacterium and Bacteroides spp. Breastfed infants have also been found to harbor gut microbial taxa with genes involved in the phosphotransferase system for carbohydrate uptake, in addition to harboring an increased abundance of microbial species commonly used as probiotics, such as L. johnsonii/L. gasseri, L. paracasei/L. casei, and B. longum (33). Moreover, Hill et al. (1) found that prolonged breast feeding (>4 months) had a significant effect on the microbial composition of cesarean delivered FT infants at 24 weeks of life, in comparison to vaginally delivered infants, suggesting that breastmilk may prove to be even more beneficial in caesarian delivered infants.
Prebiotics and Weaning
It is well known that the infant gut microbiome does not fully develop until an infant reaches 2–3 years of age. Therefore, it is important that we recognize the changes occurring in the infant gut during this transition from early infant feeding to solid foods. Indeed, the World Health Organisation (34) states that the appropriate age for complementary feeding is “6 to 23 months of age”; however, this can change in exceptionally difficult circumstances [e.g., very low birth weight (VLBW) infants]. The following studies investigate the role of diet and the introduction of galacto- and fructo-oligosaccharides (GOS and FOS) for improving gut microbiota development in early life.
In terms of diet, a recent study investigated the impact of different foods on the gut microbiota profile of a Danish infant cohort. Results from this study found strong correlations between microbial taxa present and the dietary intake of foods high in protein and fiber; specifically meats, cheeses, and Danish rye bread (35). Interestingly, breastmilk/early infant feeding was correlated with the presence of Bifidobacteriaceae, Enterococcaceae, and Lactobacillaceae, whereas Lachnospiriaceae abundance was positively correlated with protein intake and negatively correlated with Bifidobacteriaceae. Moreover, Pasteurellaceae abundance was positively correlated with fiber and health conscious food choices (high in vegetable fats, fruits or fish, but low in sugar). Findings from this study suggest that the transition from breast feeding to “family-like” foods rich in fiber and protein significantly affects development of the infant gut microbiome (35). Digestion of these foods provides a variety of fermentable substrates necessary for the growth of colonic bacteria and thus further investigation into the by-products of predigested foods may provide valuable information to positively modulate the infant gut throughout weaning.
With respect to prebiotic supplementation of infant formulae, recent studies have investigated the use of GOS and FOS to reduce pH and produce a similar short-chain fatty acid (SCFA) profile to that of exclusively breastfed infants (36). Indeed, where infant formulae have been supplemented with GOS/FOS, a higher abundance of B. longum was found in the infant gut (37, 38). In addition, Haarman and Knol (38) found that infants consuming a standard formula (without prebiotic supplement) possessed a higher abundance of Bifidobacterium catenulatum and Bifidobacterium adolescentis, resembling a more adult-like microbiota. Alternatively, prebiotic inulin-type fructans and FOS can be found readily available in foods such as cereals, chicory, and bananas, which are recommended for infants during weaning. These previously mentioned studies, and others (Table 1), provide evidence for the beneficial use of prebiotics, GOS, and FOS, to help maintain a well-balanced microbial progression from infancy to early adulthood.
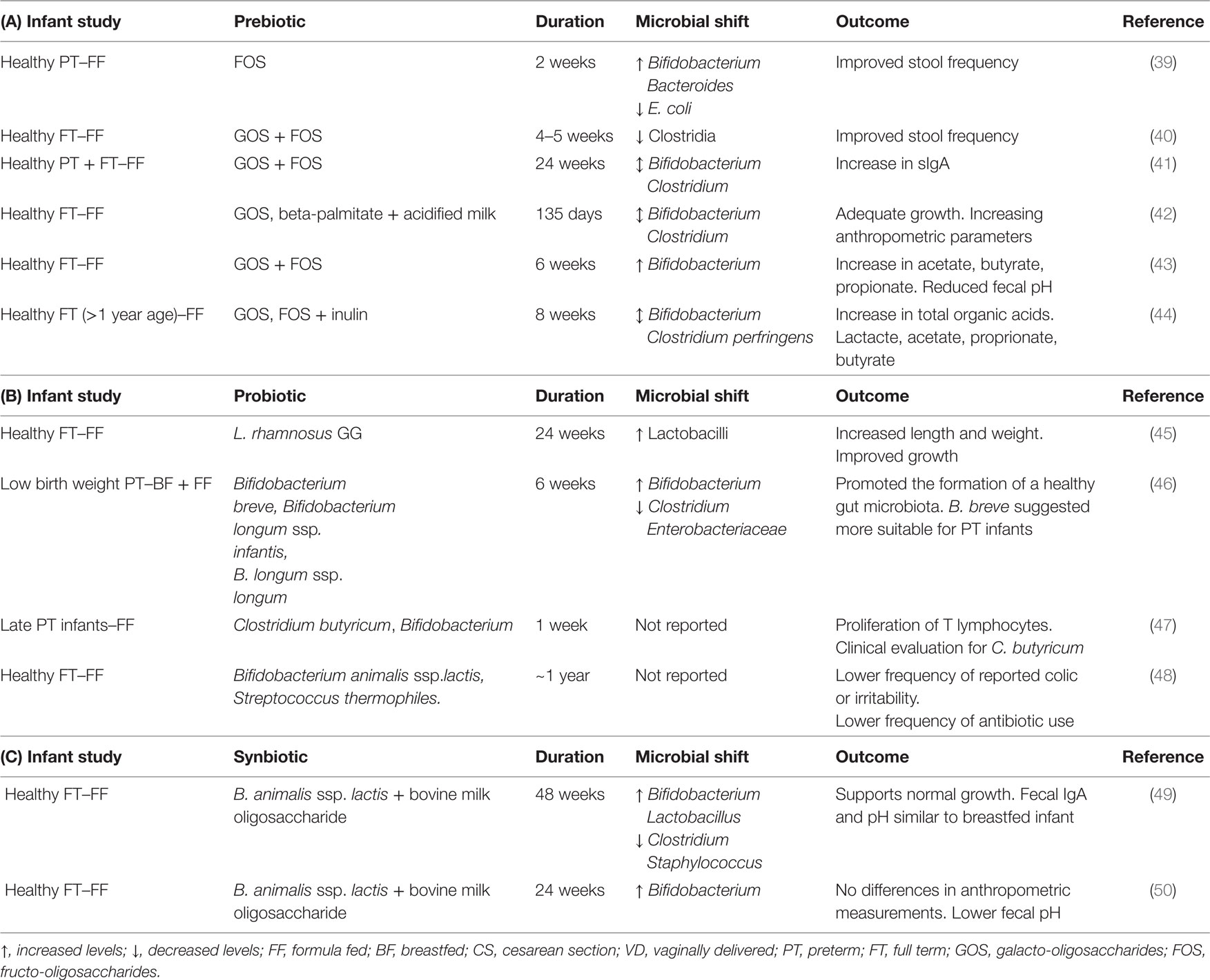
Table 1. (A) Prebiotics effective in altering the intestinal microbiota in human infant studies, (B) probiotic strains effective in altering the intestinal microbiota in a number of human infant studies, and (C) synbiotics effective in altering the intestinal microbiota in a number of human infant studies.
Although synbiotics, a combination of both a probiotic and a prebiotic (51), were not discussed in this review, the beneficial effects of bovine milk oligosaccharides and Bifidobacterium spp. on the infant gut have been noted in two human interventions (Table 1).
Probiotic Intervention
Health Benefits of Probiotics
Probiotics, described as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (52–55), have been investigated as potential prophylactics and/or treatments to re-establish gut homeostasis (Table 1). Moreover, the metabolism of indigestible oligosaccharides and plant polysaccharides by probiotic microorganisms, such as Bifidobacterium spp., contributes to the production of microbial bioactive molecules, such as SCFAs (56).
Subsequently, probiotic treatment is being extensively studied in different conditions associated with a disturbance in the gut. Necrotizing enterocolitis (NEC), childhood obesity and autism will be discussed next to highlight the link between a microbial disturbance in the gut and the beneficial use of probiotic prophylaxis in early life. The following conditions have been chosen due to their significant prevalence in current literature.
Necrotizing Enterocolitis
Necrotizing enterocolitis, where portions of the bowel undergo necrosis, is the second most common cause of mortality in PT infants. Subsequent studies focused on improving health outcomes have found an increased abundance of Proteobacteria prior to and throughout the condition, including potentially pathogenic organisms such as Salmonella and Escherichia coli (57, 58). In a longitudinal study examining gut microbiota development in PT twins, a twin pair discordant for NEC was discovered. Results from this study found clear changes attributable to antibiotic exposure and NEC development, with reduced microbial diversity and an increase in Escherichia spp. preceding NEC (59).
Probiotic prophylaxis has thus been investigated to examine whether this form of treatment could improve the quality of life in PT infants. In a comprehensive review by AlFaleh and Anabrees (60), it was found that enteral administration of probiotics reduced incidence of severe NEC, and NEC-related mortality, with the majority of infants being administered a combination of Lactobacillus, Bifidobacterium, and Streptococcus spp. via breastmilk. Interestingly, the administration of bovine lactoferrin, in combination with L. rhamnosus GG, was found effective in reducing incidences of NEC in VLBW infants (61). In addition, the routine use of Lactobacillus reuteri DSM 17938 (BioGaia®) was found to be highly successful in reducing rates of NEC in infants at highest risk (birth weights ≤ 1,000 g) (62).
Metabolic Syndrome (MS) and Obesity in Childhood
Metabolic syndrome, described by WHO in 1998, relates to any case of insulin resistance found in the presence of at least two of the following risk factors; hypertension, obesity, high triglyceride levels, or reduced high-density lipoprotein cholesterol levels. To examine the effectiveness of probiotics in preliminary animal trials against MS, L. paracasei, L. rhamnosus, and Bifidobacterium animalis subsp. lactis were found to improve glucose–insulin levels and hepatic steatosis in a high-fat diet (HFD)-induced murine model (63). However, in a systemic review on the use of early probiotic intervention in human clinical trials, inadequate evidence was found to support the use of the probiotic L. rhamnosus GG or L. paracasei F19, when administered to both mothers and infants, in the prevention of childhood MS (64).
To tackle the prevalence of childhood obesity, scientists have begun to unravel the link between diet, the gut microbiome and consequent energy intake and adiposity. Turnbaugh et al. (65) examined the hypothesis that particular communities of microorganisms could be involved directly with obesity in an obese mouse model. Results from the study found an increased capacity to harvest energy from diet, with an increased ratio of Firmicutes to Bacteroidetes (65). Further metagenomic analysis revealed that the microbiome of mice fed a high-fat/high-sugar (Western) diet, was significantly enriched in Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways involved in the fermentation of simple sugars (66). Thus, with the aim of reducing HFD-induced weight gain in humans, animal studies are examining the antiobesity effects of different probiotics (67, 68). In-depth analysis of these animal studies may provide opportunities for the introduction of probiotics to help reduce weight gain in early life.
Autism
The cause for autism spectrum disorder (ASD), a syndrome characterized by a deficit in social and communicative interactions, is yet unclear; however, recent studies have revealed a link between symptomatic cognitive dysfunctions and GI distress through a connection in the central nervous system, coining the term “brain–gut axis.” There is now evidence that probiotics alleviate GI distress in various murine models which mimic the symptomatic traits of ASD (69, 70). Studies have found that a member of the Bacteroides spp., Bacteroides fragilis, acts as a natural anti-inflammatory, capable of inhibiting inflammatory responses in a chemically induced murine model of experimental colitis (71, 72). Moreover, a maternal immune activation (MIA) model, which challenges the immune system and promotes inflammatory factors in pregnant dams, induces key features of ASD and thus serves as an appropriate murine model in testing B. fragilis as a potential therapeutic (70). Hsiao et al. (70) demonstrated the ability of B. fragilis to correct the levels of a MIA-induced serum metabolite which was found at significantly higher concentrations in MIA-offspring. Overall, B.fragilis improved gut permeability, as well as correcting ASD-related behavioral abnormalities. This suggests that a microbe-mediated therapy, such as B. fragilis, may alleviate various behavioral disorders during childhood.
Conclusion
Throughout this mini-review, we have discussed the introduction of microbial therapeutics, in addition to prebiotic supplementation, to highlight the health benefits for their use in relieving GI distress in early life.
With regards to infant formulae, prebiotic supplementation with a mixture of GOS/FOS can help mimic the composition of breastmilk and promote the development of Bifidobacterium in the infant gut, in particular B. longum.
In terms of the clinical use of probiotics, it is crucial that we develop standardized treatments which take into account the age group of a specific human cohort, in addition to health status of the group in question. In this respect, the appropriate dose of a probiotic must be determined. Moreover, it is vital that we re-evaluate the safety of alternative probiotics, coined “next-generation probiotics” (NGPs). A preliminary evaluation on the safe use of a Bacteroides xylanisolvens isolate has recently been reported (73), in addition to the beneficial effects of Faecalibacterium prausnitzii (74, 75), and bacterial strains belonging to the Eggerthellaceae family, which produce metabolites with anti-inflammatory and cardioprotective properties (76). However, guidelines outlined by the European Food Safety Authority, and the Food and Agriculture Organization of the United Nations, have made it difficult to introduce these bacteria as food supplements. In terms of economic potential, further research is required to upscale these NGPs for food and/or pharmaceutical industries. The industrial challenges may be overcome through high throughput selection of bacterial strains with the capacity to grow well in selective media and tolerate the presence of oxygen. In other words, the technological robustness of the strain in question must be tested, in addition to the suitable anaerobic media and encapsulation methods required to retain probiotic viability under good manufacturing practices. Moreover, in silico screening of bacterial genomes will ensure the safety of these strains through the detection of antibiotic resistance and virulence genes. Alternatively, live biotherapeutic products (LBPs) may create an opportunity to introduce NGPs to the market (77). An LBP has recently been described as “a biological product that: (1) contains live organisms, such as bacteria; (2) is applicable to the prevention, treatment, or cure of a disease or condition of human beings; and (3) is not a vaccine” (78). In addition, O’Toole et al. (77) described future testing of LBPs as biological medicinal products, thereby providing new opportunities to introduce NGPs as well characterized drugs to the market.
Overall, we conclude that additional studies are necessary to investigate the influence of prebiotics and probiotics in early life. It is important that we consider the mixed microbial communities present within foods and select those which will survive and adapt readily in an industrial environment (79, 80). More importantly, we suggest if new probiotics and prebiotics are to be considered health beneficial in the European market, the necessity for comprehensive, randomized controlled trials is vital. The current approach requires further strategy to provide consumers with valid information toward the use of probiotic and prebiotic supplementation in early childhood.
Author Contributions
RPR, CS, and CAR conceived the manuscript. CW drafted the manuscript. All authors reviewed the final version of the manuscript and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This research was supported by the Science Foundation Ireland (SFI)-funded APC Microbiome Institute; by Department of Agriculture Food and Marine (DAFM)-funded INFANTMET (Ref. No. 10FDairy) and ToddlerFood (Ref. No. 14F821) projects.
Abbreviations
GI, gastrointestinal; FT, full term; PT, preterm; VLBW, very low birth weight; GOS, galacto-oligosaccharides; FOS, fructo-oligosaccharides; SCFAs, short chain fatty acids; NEC, necrotizing enterocolitis; MS, metabolic syndrome; HFD, high-fat diet; ASD, autism spectrum disorder; MIA, maternal immune activation; NGPs, next generation probiotics; LBPs, live biotherapeutic products.
References
1. Hill CJ, Lynch DB, Murphy K, Ulaszewska M, Jeffery IB, O’Shea CA, et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET cohort. Microbiome (2017) 5:21. doi:10.1186/s40168-016-0213-y
2. Koenig JE, Spor A, Scalfone N, Fricker AD, Stombaugh J, Knight R, et al. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci U S A (2011) 108(1):4578–85. doi:10.1073/pnas.1000081107
3. Lim MY, Yoon HS, Rho M, Sung J, Song YM, Lee K, et al. Analysis of the association between host genetics, smoking, and sputum microbiota in healthy humans. Sci Rep (2016) 6:23745. doi:10.1038/srep23745
4. Murphy K, O’ Shea CA, Ryan CA, Dempsey EM, O’ Toole PW, Stanton C, et al. The gut microbiota composition in dichorionic triplet sets suggests a role for host genetic factors. PLoS One (2015) 10(4):e0122561. doi:10.1371/journal.pone.0122561
5. David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature (2014) 505(7484):559–63. doi:10.1038/nature12820
6. O’Sullivan A, Farver M, Smilowitz JT. The influence of early infant-feeding practices on the intestinal microbiome and body composition in infants. Nutr Metab Insights (2015) 8(1):1–9. doi:10.4137/NMI.S29530
7. Mueller NT, Bakacs E, Combellick J, Grigoryan Z, Dominguez-Bello MG. The infant microbiome development: mom matters. Trends Mol Med (2015) 21(2):109–17. doi:10.1016/j.molmed.2014.12.002
8. Ringel-Kulka T, Cheng J, Ringel Y, Salojärvi J, Carroll I, Palva A, et al. Intestinal microbiota in healthy U.S. young children and adults – a high throughput microarray analysis. PLoS One (2013) 8(5):e64315. doi:10.1371/journal.pone.0064315
9. Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, et al. Human gut microbiome viewed across age and geography. Nature (2012) 486(7402):222–7. doi:10.1038/nature11053
10. Kashtanova DA, Popenko AS, Tkacheva ON, Tyakht AB, Alexeev DG, Boytsov SA. Association between the gut microbiota and diet: fetal life, early childhood, and further life. Nutrition (2016) 32(6):620–7. doi:10.1016/j.nut.2015.12.037
11. Arboleya S, Binetti A, Salazar N, Fernández N, Solís G, Hernández-Barranco A, et al. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol Ecol (2012) 79(3):763–72. doi:10.1111/j.1574-6941.2011.01261.x
12. Forsgren M, Isolauri E, Salminen S, Rautava S. Late preterm birth has direct and indirect effects on infant gut microbiota development during the first six months of life. Acta Paediatr (2017) 106:1103–9. doi:10.1111/apa.13837
13. La Rosa PS, Warner BB, Zhou Y, Weinstock GM, Sodergren E, Hall-Moore CM, et al. Patterned progression of bacterial populations in the premature infant gut. Proc Natl Acad Sci U S A (2014) 111(34):12522–7. doi:10.1073/pnas.1409497111
14. Stewart CJ, Embleton ND, Clements E, Luna PN, Smith DP, Fofanova TY, et al. Cesarean or vaginal birth does not impact the longitudinal development of the gut microbiome in a cohort of exclusively preterm infants. Front Microbiol (2017) 8:1008. doi:10.3389/fmicb.2017.01008
15. Schultz M, Göttl C, Young RJ, Iwen P, Vanderhoof JA. Administration of oral probiotic bacteria to pregnant women causes temporary infantile colonisation. J Pediatr Gastroenterol Nutr (2004) 38(3):293–7. doi:10.1097/00005176-200403000-00012
16. MacIntyre DA, Chandiramani M, Lee YS, Kindinger L, Smith A, Angelopoulos N, et al. The vaginal microbiome during pregnancy and the postpartum period in a European population. Sci Rep (2015) 5:8988. doi:10.1038/srep08988
17. Ravel J, Gajer P, Abdo Z, Schneider GM, Koenig SS, McCulle SL, et al. Vaginal microbiome of reproductive-age women. Proc Natl Acad Sci U S A (2011) 108(1):4680–7. doi:10.1073/pnas.1002611107
18. Hunt KM, Foster JA, Forney LJ, Schutte UM, Beck DL, Abdo Z, et al. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS One (2011) 6:e21313. doi:10.1371/journal.pone.0021313
19. Martín R, Jiménez E, Heilig HG, Fernández L, Marín ML, Zoetendal EG, et al. Isolation of bifidobacteria from breast milk and assessment of the bifidobacterial population by PCR-denaturing gradient gel electrophoresis and quantitative real-time PCR. Appl Environ Microbiol (2009) 75:965–9. doi:10.1128/AEM.02063-08
20. Martín V, Maldonado A, Moles L, Rodríguez-Banos M, Del Campo R, Fernández L, et al. Sharing of bacterial strains between breast milk and infant faeces. J Hum Lact (2012) 28:36–44. doi:10.1177/0890334411424729
21. Fernández L, Langa S, Martín V, Maldonado A, Jiménez E, Martín R, et al. The human milk microbiota: origin and potential roles in health and disease. Pharmacol Res (2013) 69(1):1–10. doi:10.1016/j.phrs.2012.09.001
22. Murphy K, David C, O’Callaghan T, O’Shea CA, Dempsey EM, O’Toole PW, et al. The composition of human milk and infant faecal microbiota over the first three months of life: a pilot study. Sci Rep (2017) 7:40597. doi:10.1038/srep40597
23. Solís G, de Los Reyes-Gavilan CG, Fernández N, Margolles A, Gueimonde M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe (2010) 16(3):307–10. doi:10.1016/j.anaerobe.2010.02.004
24. Martín R, Jiménez E, Olivares M, Marín ML, Fernández L, Xaus J, et al. Lactobacillus salivarius CECT 5713, a potential probiotic strain isolated from infant feces and breast milk of a mother-child pair. Int J Food Microbiol (2006) 112(1):35–43. doi:10.1016/j.ijfoodmicro.2006.06.011
25. Makino H, Martin R, Ishikawa E, Gawad A, Kubota H, Sakai T, et al. Multilocus sequence typing of bifidobacterial strains from infant’s faeces and human milk: are bifidobacteria being sustainably shared during breastfeeding? Benef Microbes (2015) 6(4):563–72. doi:10.3920/BM2014.0082
26. Jiménez E, Fernández L, Marín ML, Martín R, Odriozola JM, Nueno-Palop C, et al. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr Microbiol (2005) 51(4):270–4. doi:10.1007/s00284-005-0020-3
27. Hansen R, Scott KP, Khan S, Martin JC, Berry SH, Stevenson M, et al. First-pass meconium samples from healthy term vaginally-delivered neonates: an analysis of the microbiota. PLoS One (2015) 10(7):e0133320. doi:10.1371/journal.pone.0133320
28. Ardissone AN, de la Cruz DM, Davis-Richardson AG, Rechcigl KT, Li N, Drew JC, et al. Meconium microbiome analysis identifies bacteria correlated with premature birth. PLoS One (2014) 9(3):e90784. doi:10.1371/journal.pone.0090784
29. Aagaard K, Ma J, Antony KM, Ganu R, Petrosino J, Versalovic J. The placenta harbors a unique microbiome. Sci Transl Med (2014) 6:237. doi:10.1126/scitranslmed.3008599
30. Lauder AP, Roche AM, Sherrill-Mix S, Bailey A, Laughlin AL, Bittinger K, et al. Comparison of placenta samples with contamination controls does not provide evidence for a distinct placenta microbiota. Microbiome (2016) 4(1):29. doi:10.1186/s40168-016-0172-3
31. Gibson GR, Scott KP, Rastall RA, Tuohy KM, Hotchkiss A, Dubert-Ferrandon A, et al. Dietary prebiotics: current status and new definition. Food Sci Technol Bull Funct Foods (2010) 7:1–19. doi:10.1616/1476-2137.15880
32. Chatterton DE, Nguyen DN, Bering SB, Sangild PT. Anti-inflammatory mechanisms of bioactive milk proteins in the intestine of newborns. Int J Biochem Cell Biol (2013) 45(8):1730–47. doi:10.1016/j.biocel.2013.04.028
33. Bäckhed FJ, Roswall Y, Peng Q, Feng H, Jia P, Kovatcheva-Datchary Y, et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe (2015) 17(5):690–703. doi:10.1016/j.chom.2015.04.004
34. World Health Organisation. Infant and Young Child Feeding: Model Chapter for Textbooks for Medical Students and Allied Health Professionals. (2009). Available from: https://www.ncbi.nlm.nih.gov/books/NBK148965/
35. Laursen MF, Andersen LB, Michaelsen KF, Mølgaard C, Trolle E, Bahl MI, et al. Infant gut microbiota development is driven by transition to family foods independent of maternal obesity. mSphere (2016) 10(1):e69–15. doi:10.1128/mSphere.00069-15
36. Fanaro S, Boehm G, Garssen J, Knol J, Mosca F, Stahl B, et al. Galacto-oligosaccharides and long-chain fructo-oligosaccharides as prebiotics in infant formulas: a review. Acta Paediatr (2005) 94(449):22–6. doi:10.1080/08035320510043538
37. Barrett E, Deshpandey AK, Ryan CA, Dempsey EM, Murphy B, O’Sullivan L, et al. The neonatal gut harbours distinct bifidobacterial strains. Arch Dis Child Fetal Neonatal Ed (2015) 100(5):F405–10. doi:10.1136/archdischild-2014-306110
38. Haarman M, Knol J. Quantitative real-time PCR assays to identify and quantify fecal Bifidobacterium species in infants receiving a prebiotic infant formula. Appl Environ Microbiol (2005) 71(5):2318–24. doi:10.1128/AEM.71.5.2318-2324.2005
39. Kapiki A, Costalos C, Oikonomidou C, Triantafyllidou A, Loukatou E, Pertrohilou V. The effect of a fructo-oligosaccharide supplemented formula on gut flora of preterm infants. Early Hum Dev (2007) 83(5):335–9. doi:10.1016/j.earlhumdev.2006.07.003
40. Costalos C, Kapiki A, Apostolou M, Papathoma E. The effect of a prebiotic supplemented formula on growth and stool microbiology of term infants. Early Hum Dev (2008) 84(1):45–9. doi:10.1016/j.earlhumdev.2007.03.001
41. Scholtens PA, Alliet P, Raes M, Alles MS, Kroes H, Boehm G, et al. Fecal secretory immunoglobulin A is increased in healthy infants who receive a formula with short-chain galacto-oligosaccharides and long-chain fructo-oligosaccharides. J Nutr (2008) 138(6):1141–7.
42. Civardi E, Garofoli F, Longo S, Mongini ME, Grenci B, Mazzucchelli I, et al. Safety, growth, and support to healthy gut microbiota by an infant formula enriched with functional compounds. Clin Nutr (2017) 36(1):238–45. doi:10.1016/j.clnu.2015.11.006
43. Holscher HD, Faust KL, Czerkies LA, Litov R, Ziegler EE, Lessin H, et al. Effects of prebiotic-containing infant formula on gastrointestinal tolerance and fecal microbiota in a randomized controlled trial. JPEN J Parenter Enteral Nutr (2012) 36(1):95S–105S. doi:10.1177/0148607111430087
44. Chen YL, Liao FH, Lin SH, Chien YW. A prebiotic formula improves the gastrointestinal bacterial flora in toddlers. Gastroenterol Res Pract (2016) 2016:3504282. doi:10.1155/2016/3504282
45. Vendt N, Grünberg H, Tuure T, Malminiemi O, Wuolijoki E, Tillmann V, et al. Growth during the first 6 months of life in infants using formula enriched with Lactobacillus rhamnosus GG: double blind, randomized trial. J Hum Nutr Diet (2006) 19(1):51–8. doi:10.1111/j.1365-277X.2006.00660
46. Ishizeki S, Sugita M, Takata M, Yaeshima T. Effect of administration of bifidobacteria on intestinal microbiota in low-birth-weight infants and transition of administered bifidobacteria: a comparison between one-species and three-species administration. Anaerobe (2013) 23:38–44. doi:10.1016/j.anaerobe.2013.08.002
47. Zhang SF, Tang ZS, Tong L, Tao XX, Suo QF, Xu XM. Effects of Clostridium butyricum and Bifidobacterium on BTLA expression on CD4+ T cells and lymphocyte differentiation in late preterm infants. Microb Pathog (2016) 100:112–8. doi:10.1016/j.micpath.2016.09.008
48. Saavedra JM, Abi-Hanna A, Moore N, Yolken RH. Long-term consumption of infant formulas containing live probiotic bacteria: tolerance and safety. Am J Clin Nutr (2004) 79(2):261–7.
49. Radke M, Picaud JC, Loui A, Cambonie G, Faas D, Lafeber HN, et al. Starter formula enriched in prebiotics and probiotics ensures normal growth of infants and promotes gut health: a randomized clinical trial. Pediatr Res (2017) 81(4):622–31. doi:10.1038/pr.2016.270
50. Cooper P, Bolton KD, Velaphi S, de Groot N, Emady-Azar S, Pecquet S, et al. Early benefits of a starter formula enriched in prebiotics and probiotics on the gut microbiota of healthy infants born to HIV+ mothers: a randomized double-blind controlled trial. Clin Med Insights Pediatr (2016) 10:119–30. doi:10.4137/CMPed.S40134
51. Schrezenmeir J, de Vrese M. Probiotics, prebiotics, and synbiotics – approaching a definition. Am J Clin Nutr (2001) 73(2):361S–4S.
52. Morelli L, Capurso L. FAO/WHO guidelines on probiotics: 10 years later. J Clin Gastroenterol (2012) 46:S1–2. doi:10.1097/MCG.0b013e318269fdd5
53. Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol (2014) 11:506–14. doi:10.1038/nrgastro.2014.66
54. FAO/WHO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation. (Vol. 85). (2006). p. 0254–4725. Available from: http://www.fao.org/3/a-a0512e.pdf
55. FAO/WHO. Report on Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. (2001). Available from: ftp://ftp.fao.org/es/esn/food/probio_report_en.pdf
56. Vernocchi P, Del Chierico F, Putignani L. Gut microbiota profiling: metabolomics based approach to unravel compounds affecting human health. Front Microbiol (2016) 7:1144. doi:10.3389/fmicb.2016.01144
57. Wang Y, Hoenig JD, Malin KJ, Qamar S, Petrof EO, Sun J, et al. 16S rRNA gene-based analysis of fecal microbiota from preterm infants with and without necrotising enterocolitis. ISME J (2009) 3(8):944–54. doi:10.1038/ismej.2009.37
58. Björkström MV, Hall L, Söderlund S, Håkansson EG, Håkansson S, Domellöf M. Intestinal flora in very low-birth weight infants. Acta Paediatr (2009) 98:1762–7. doi:10.1111/j.1651-2227.2009.01471.x
59. Stewart CJ, Marrs EC, Nelson A, Lanyon C, Perry JD, Embleton ND, et al. Development of the preterm gut microbiome in twins at risk of necrotising enterocolitis and sepsis. PLoS One (2013) 8(8):e73465. doi:10.1371/journal.pone.0073465
60. AlFaleh K, Anabrees J. Probiotics for prevention of necrotising enterocolitis in preterm infants. Evid Based Child Health (2014) 9:584–671. doi:10.1002/ebch.1976
61. Manzoni P, Meyer M, Stolfi I, Rinaldi M, Cattani S, Pugni L, et al. Bovine lactoferrin supplementation for prevention of necrotising enterocolitis in very-low-birth-weight neonates: a randomized clinical trial. Early Hum Dev (2014) 90(1):S60–5. doi:10.1016/S0378-3782(14)70020-9
62. Hunter C, Dimaguila MAV, Gal P, Wimmer JE, Ransom JL, Carlos RQ, et al. Effect of routine probiotic, Lactobacillus reuteri DSM 17938, use on rates of necrotising enterocolitis in neonates with birthweight < 1000 grams: a sequential analysis. BMC Pediatr (2012) 12:142. doi:10.1186/1471-2431-12-142
63. Wang J, Tang H, Zhang C, Zhao Y, Derrien M, Rocher E, et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J (2015) 9(1):1–15. doi:10.1038/ismej.2014.99
64. Balasubramanian H, Patole S. Early probiotics to prevent childhood metabolic syndrome: a systematic review. World J Methodol (2015) 5(3):157–63. doi:10.5662/wjm.v5.i3.157
65. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature (2006) 444(7122):1027–131. doi:10.1038/nature05414
66. Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe (2008) 3(4):213–23. doi:10.1016/j.chom.2008.02.015
67. Park DY, Ahn YT, Park SH, Huh CS, Yoo SR, Yu R, et al. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS One (2013) 8(3):e59470. doi:10.1371/journal.pone.0059470
68. An HM, Park SY, Lee DK, Kim JR, Cha MK, Lee SW, et al. Antiobesity and lipid-lowering effects of Bifidobacterium spp. in high fat diet-induced obese rats. Lipids Health Dis (2011) 10:116. doi:10.1186/1476-511X-10-116
69. Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci (2012) 13:701–12. doi:10.1038/nrn3346
70. Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell (2013) 155(7):1451–63. doi:10.1016/j.cell.2013.11.024
71. Mazmanian SK, Round JL, Kasper DL. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature (2008) 453:620–5. doi:10.1038/nature07008
72. Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci U S A (2010) 107:12204–9. doi:10.1073/pnas.0909122107
73. Ulsemer P, Toutounian K, Schmidt J, Karsten U, Goletz S. Preliminary safety evaluation of a new Bacteroides xylanisolvens isolate. Appl Environ Microbiol (2012) 78(2):528–35. doi:10.1128/AEM.06641-11
74. Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermúdez-Humarán LG, Gratadoux JJ. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci U S A (2008) 105(43):16731–6. doi:10.1073/pnas.0804812105
75. Song H, Yoo Y, Hwang J, Na YC, Kim HS. Faecalibacterium prausnitzii subspecies-level dysbiosis in the human gut microbiome underlying atopic dermatitis. J Allergy Clin Immunol (2016) 137(3):852–60. doi:10.1016/j.jaci.2015.08.021
76. Selma MV, Beltrán D, Luna MC, Romo-Vaquero M, García-Villalba R, Mira A, et al. Isolation of human intestinal bacteria capable of producing the bioactive metabolite isourolithin a from ellagic acid. Front Microbiol (2017) 7(8):1521. doi:10.3389/fmicb.2017.01521
77. O’Toole PW, Marchesi JR, Hill C. Next-generation probiotics: the spectrum from probiotics to live biotherapeutics. Nat Microbiol (2017) 25(2):17057. doi:10.1038/nmicrobiol.2017.57
78. FDA. Early Clinical Trials with Live Biotherapeutic Products: Chemistry, Manufacturing, and Control Information; Guidance for Industry. (Vol. 81 FR). (2016). p. 43206–7. Available from: https://www.fda.gov/downloads/Biologi…/UCM292704.pdf
79. Walsh AM, Crispie F, Claesson MJ, Cotter PD. Translating omics to food microbiology. Annu Rev Food Sci Technol (2017) 8:113–34. doi:10.1146/annurev-food-030216-025729
Keywords: probiotics, prebiotics, gut microbiota, infant, health
Citation: Watkins C, Stanton C, Ryan CA and Ross RP (2017) Microbial Therapeutics Designed for Infant Health. Front. Nutr. 4:48. doi: 10.3389/fnut.2017.00048
Received: 06 April 2017; Accepted: 26 September 2017;
Published: 26 October 2017
Edited by:
Christophe Lacroix, ETH Zurich, SwitzerlandReviewed by:
Clara G. De Los Reyes-Gavilan, Consejo Superior de Investigaciones Científicas (CSIC), SpainAbelardo Margolles, Consejo Superior de Investigaciones Científicas (CSIC), Spain
Copyright: © 2017 Watkins, Stanton, Ryan and Ross. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: R. Paul Ross, p.ross@ucc.ie