 Chao-Tao Tang1†
Chao-Tao Tang1† Qian Liang1†Li Yang1†Xiao-Lu Lin2†Shan Wu1†Yong Chen1Xin-Tian Zhang1Yun-Jie Gao1Zhi-Zheng Ge1*
Qian Liang1†Li Yang1†Xiao-Lu Lin2†Shan Wu1†Yong Chen1Xin-Tian Zhang1Yun-Jie Gao1Zhi-Zheng Ge1*- 1Division of Gastroenterology and Hepatology, Key Laboratory of Gastroenterology and Hepatology, Ministry of Health, Shanghai Institute of Digestive Disease, Renji Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
- 2Department of Digestive Endoscopy, Provincial Clinic Medical College, Fujian Medical University, Fujian Provincial Hospital, Fuzhou, China
Background: Gastric cancer (GC), one of the most common cancers worldwide, is highly malignant and fatal. Ras-related protein in brain 31 (RAB31), a member of the RAB family of oncogenes, participates in the process of carcinogenesis and cancer development; however, its role in GC progression is unknown.
Methods: In our study, 90 pairs of tissue microarrays were used to measure the levels of RAB31 protein by immunochemistry, and 22 pairs of fresh tissue were used to measure the levels of RAB31 mRNA by quantitative PCR. We also investigated the effects of RAB31 on tumor growth both in vitro and in vivo.
Results: RAB31 was overexpressed in GC tissues, and its overexpression predicted poor survival in patients. In a nude mouse model, depletion of RAB31 inhibited tumor growth. In vitro, silencing of RAB31 suppressed cell viability, promoted cell cycle arrest, enhanced apoptosis, and affected the expression of cell cycle and apoptotic proteins; these effects were mediated by glioma-associated oncogene homolog 1 (GLI1). Co-immunoprecipitation and immunofluorescence assays confirmed that RAB31 interacted with GLI1. In addition, luciferase reporter assays and Western blotting showed that microRNA-30c-2-3p modulated the RAB31/GLI1 pathway by targeting the 3′-untranslated region of RAB31.
Conclusions: Collectively, these data show that RAB31 is regulated by microRNA-30c-2-3p, and functions as an oncogene in GC tumorigenesis and development by interacting with GLI1. Therefore, targeting the miR-30c-2-3p/RAB31/GLI1 axis may be a therapeutic intervention for gastric cancer.
Introduction
Gastric cancer (GC) is one of the most common cancers worldwide and has a high fatality rate (1). Helicobacter pylori, recognized as the main cause of this disease, causes damage to the gastric epithelium due to sustained inflammation and direct invasion (2). GC patients are prone to local recurrence and distant metastases, resulting in a low 5-year survival rate (3). Although there are many treatment strategies for GC, such as surgery, chemotherapy, and radiotherapy, their success rates have been low (3, 4). Therefore, there is an urgent need to develop more effective, targeted therapies for this disease.
Ras-related protein in brain 31 (RAB31) (also called Rab22b) is a member of the RAB family, which belongs to the Ras superfamily of small GTPases and is also characterized as the RAB5 subfamily (5). RAB31 was first isolated in 1996 from melanoma cells and was found to have significant homology with RAB22; hence, it was named RAB22b (6). As with other members of the RAB family, RAB31 is expressed throughout the cell and mainly functions to regulate vesicular intracellular membrane transport (7). Initially, RAB31 was found to be involved in the transport of many substances such as glucose transporter type 4, epidermal growth factor receptor, and mannose-6-phosphate receptors (8–10). Recently, it was also found to play a significant role in human cancers such as breast cancer, glioblastoma, and hepatocellular cancer (11, 12). In breast cancer, RAB31 was first identified as an independent prognostic factor, and was subsequently found to interact with the mucin 1 C-terminal subunit oncoprotein to promote cancer progression and subsequent tamoxifen resistance in patients (13–16). However, the function of RAB31 in GC is unknown. Therefore, we determined if it plays a similar role in GC progression.
MicroRNAs (miRNAs) are short single-stranded non-coding RNA molecules ~20–25 nucleotides in length that play a critical role in gene expression (17). Their key function is to directly bind the 3′ untranslated regions (3′-UTRs) of complementary messenger RNAs (mRNAs) to degrade or inhibit their translation (17). Increasing evidence has confirmed that miRNAs also bind to coding regions (18). In addition, many miRNAs such as miR-30a-5p (19) and miR-155 (20) have been confirmed as oncogenes or suppressors of tumor growth and metastasis. miRNAs play vital roles in various pathological processes such as cell proliferation, cell stemness, and the aggressiveness of GC cells (21, 22).
Glioma-associated oncogene homolog 1 (GLI1), a transcript of Hedgehog (Hh) signaling, strongly promotes GC development (23). In the etiology of GC, it has been confirmed that GLI1 is involved in H. pylori infection and promotes chronic information (24, 25). Furthermore, the mechanism through which GLI1 enhances GC development involves many oncogenes, such as NAPDH oxidase 4 (26), Mucin 5AC, Oligomeric Mucus/Gel-Forming (27), and Wnt proteins (28), leading to increased resistance to chemotherapy drugs (29, 30). Recently, it was shown that Hh signaling via Smoothened and GLI1 requires involvement of the RAB family (31). RAB23 interacts with GLI1 to promote the progression of tumors such as breast, prostate, and ovarian cancers (32–34).
In this study, we analyzed the level of RAB31 expression in GC tissue and the correlation with clinical information. Overexpression of RAB31 promoted GLI1 expression, subsequently inducing the activation of downstream oncogenes and enhancing GC cell proliferation and inhibiting apoptosis. In addition, miR-30c-2-3p regulated the expression of RAB31 by binding the 3′-UTR, and regulated GLI1 expression to reverse the effects of RAB31.
Materials and Methods
Patients and Tissue Samples
A total of 90 patients with an initial diagnosis of GC between December 2009 and June 2010 at Renji Hospital, affiliated with the Medical College, Shanghai Jiaotong University School (Shanghai, China) were included in this study. All patients provided signed informed consent and did not undergo preoperative treatment. The surgical tissues were fixed in formalin and embedded in paraffin. Detailed clinicopathological information such as age, gender, and tumor clinical stage is shown in Table 1. We also collected 22 pairs of fresh tumor tissues and para-tumor tissues from GC patients to measure the level of RAB31 mRNA. The process of extracting mRNA from tissues was very similar to that used for extracting cellular mRNA.
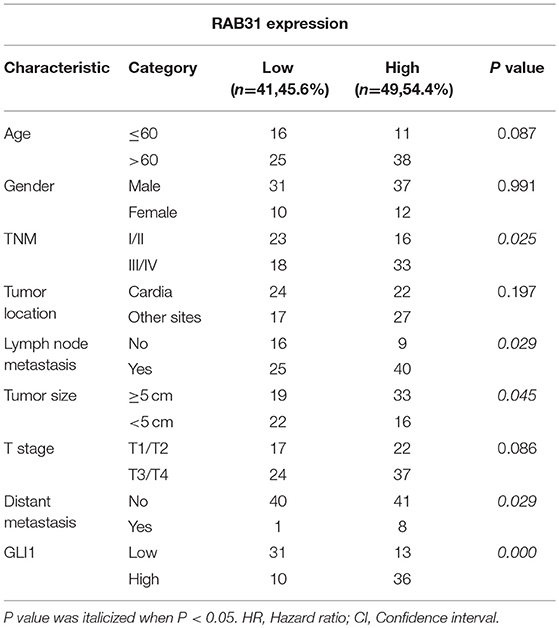
Table 1. Correlation of RAB31 expression with the clinicopathological characteristics of GC.
Bioinformatics Analysis
To explore the level of RAB31 expression in GC tissues, we used the Oncomine Cancer Microarray database (https://www.oncomine.org) and assessed the levels of RAB31 among different tissue specimens. Additionally, we investigated the correlation between RAB31 and GLI1 expression with the Gene Expression Profiling Interactive Analysis (GEPIA) tool and a gene set enrichment analysis (GSEA) which was performed as depicted in our previous study (35). We also used miRWalk 2.0, miRanda, and Targetscan to predict miRNAs that target RAB31. Then we performed Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis to identify the miRNAs involved in the Hh signaling pathway, and investigated the level of miRNA expression in GC tissues using the YM500v2 database (http://ngs.ym.edu.tw/ym500v2/index.php)(36). Finally, we created a survival curve of the predicted miRNAs using the The Cancer Genome Atlas (TCGA) database.
Cell Culture
All of the GC cell lines utilized were the same as those previously described (26). Cells were cultured in RPMI 1640 medium (Gibco, Gaithersburg, MD, USA) supplemented with 10% fetal bovine serum (FBS), and maintained at 37°C in an atmosphere of 5% CO2.
Transfection
First, cells were seeded in 6-well plates for 24 h. Then we transfected plasmids and small interfering RNA (siRNA) using the Genmute reagent (SignaGen Laboratories, Rockville, MD, USA) according to the manufacturer's instructions. The pCMV-RAB31 cDNA plasmid was constructed by OBIO Company (Shanghai, China); the corresponding control plasmid was pCDNA3.1(+). The RAB31 cDNA plasmid comprising 588 bp was constructed according to the NM sequence (NM_006868), and cloned into the pcDNA3.1(+) eukaryotic expression vector. The wild-type and mutant RAB31 3′UTR plasmids were constructed by NEWGEORGE (Shanghai, China), and the psiCHECK™−2 vector served as the empty vector control. The wild-type RAB31 3′UTR plasmid sequence was identified in the Pubmed database according to the RAB31 gene symbol, and was constructed according to the database. The sequences of the wild-type and mutant RAB31 3′UTR plasmids are provided in Data Sheet 1. Transfection of RAB31 miRNA mimics and inhibitor was performed using the Dharmocon reagent (Dharmocon, USA), and co-transfection of the miRNA inhibitor and RAB31 siRNA was done using Lipofectamine™ 3000 (Invitrogen™, Carlsbad, CA, USA). The siRNA sequences were designed by GenePharma Company (Shanghai, China) and are presented in Table S1.
Quantitative PCR
Tissue RNA extraction was performed as previously described (26). Briefly, all of the fresh tissues were homogenized using Trizol reagent (Takara, Tokyo, Japan), and extacted using chloroform and isopropyl alcohol. The concentrations of total RNA were measured using NanoDrop 2000 (Thermo Scientific, Wilmington, DE, USA). The complementary DNA was synthesized with reagents purchased from Takara according to the manufacturer's instructions. Quantitative PCR (qPCR) was performed using SYBR Green and other reagents (Takara), according to a previously described method (37). The primer sequences of RAB31 were as follows: forward 5′-ATCTTTGGGCTGGGTTTG-3′ and reverse 5′-ATGGGCTCATTAGTGGGTAG-3′. The primer sequences of 18S and GLI1 were the same as previously reported. The primer sequences of hsa-mir-30c-2-3p and U6 (HmiRQP0395 and HmiRQP9001, respectively) were constructed by GeneCopeia (GuangZhou, China).
Cell Proliferation and Colony Formation Assays
The protocols used for the cell proliferation and colony formation assays were in accordance with a previous study (26). Briefly, for the cell proliferation assay, cells (2 × 103) were incubated in 96-well plates overnight and treated the next day. The cell viability reagent was added to the plates in serum-free medium (1:10) and incubated for 2 h, after which the optical density at 450 nm was measured; this was repeated every 24 h for 96 h. For the colony formation assay, 800 cells were seeded in 6-well plates and cultured for 12 days. Then cell colonies were fixed in formaldehyde and stained with 1% crystal violet. Finally, the number of colonies was compared among RAB31-depletion group, control group and RAB31-overexpression group.
Cell Cycle and Apoptosis Assays
These assays were conducted as previously described (35). Cells were seeded in 6-well plates and cultured for 72 h. After the cells were collected, they were washed once and fixed in 70% ethyl alcohol for 24 h. Then the cell cycle was measured using flow cytometry after being stained with propidium iodide (PI) (BD Biosciences, Franklin Lakes, NJ, USA) for 30 min. For the apoptosis assay, the cells and supernatant were collected together and washed with 1X binding buffer. Apoptosis was measured after double staining with Annexin V FITC/PI (BD Biosciences) for 15 min. The results of the cell cycle and apoptosis were analyzed using FlowJo VX10 software.
Western Blotting
Western blotting was performed according to a previously described protocol (35). Briefly, cells in a 6-well plate were lysed on ice with RIPA buffer containing protease inhibitors, and the lysates were centrifuged at 12,000 rpm at 4°C for 10 min. The protein concentration was measured using the BCA Protein Assay Kit (23227; Thermo Fisher Scientific, Waltham, MA, USA). Then proteins were resolved on 10% SDS-PAGE gels and electrotransferred to the Pure Nitrocellulose Blotting Membrane (Pall Corporation, Port Washington, NY, USA). The membrane was blocked in 5% milk, and incubated overnight at 4°C with the following primary antibodies: RAB31 (1:400, 16182-1-AP; Proteintech, Rosemont, IL, USA), GLI1 (1:1,000, ab134906; Abcam, Cambridge, MA, USA), cyclin D1 (1:5,000, ab134175; Abcam), c-Myc (1:1,000, ab39688; Abcam), B-cell lymphoma 2 (Bcl-2) (1:1,000, ab32124, Abcam), and Bcl-2-associated X protein (BAX) (1:1,000, ab32503, Abcam). After washing, the membrane was incubated at room temperature for 1 h with the same secondary antibodies as those used in our previous study (1:3,000; WeiAo Pharmaceutical, Sichuan, China) (35). The protein signals were detected using the ChemiDocTM Imaging System (Bio-Rad, Hercules, CA, USA).
Immunohistochemistry
The GC specimens were first processed into tissue microarrays. Then the microarrays were dewaxed, followed by antigen retrieval and incubation overnight at 4°C with the following primary antibodies: RAB31 (1:50, 16182-1-AP; Proteintech) and GLI1 (1:100, ab134906; Abcam). Finally, the microarrays were washed and incubated with secondary antibody for 1 h, followed by staining with DAB reagent. The intensity of staining and the percentage of positively stained cells were analyzed according to a previously described method (37). The immunohistochemistry (IHC) results were scored as follows: high expression (scores 7–9), moderate expression (scores 4–7), and low expression (scores 0–3), and blindly and independently assessed by pathologists.
Luciferase Reporter Assay
A 522 bp fragment of RAB31 3′-UTR that contained a site for has-mir-30c-2-3p binding was designed and cloned into a luciferase reporter plasmid that included a wild-type and mutant (NEWGEORGE). According to the instructions of the Luc-PairTM Duo-Luciferase Assay Kit 2.0 (GeneCopoeia), cells were seeded into 96-well plates for 24 h. Then the cells were co-transfected with the 3′-UTR plasmid and has-mir-30c-2-3p mimics or the negative control (NC) using Lipofectamine™ 3000 (Invitrogen™). After 24 h, the fluorescence intensity was measured by the Luc-PairTM Duo-Luciferase Assay Kit 2.0.
Immunofluorescence Assay
GC cells were incubated in chamber slides for 24 h, fixed in 4% paraformaldehyde for 20 min, and permeabilized with 0.1% Triton X-100 for 1 h. Next, cells were incubated with RAB31 (1:100, H00011031-M03; Abnova, Taipei, Taiwan) or GLI1 (1:100, ab49314; Abcam) primary antibody overnight at 4°C, after which the slides were washed three times. Finally, cells were incubated with goat anti-rabbit or anti-mouse IgG fluorescent secondary antibody (Thermo Fisher Scientific), and the nuclei were stained with DAPI. The cells were examined by confocal fluorescence microscopy at wavelengths of 488 and 594 nm.
Co-immunoprecipitation
A co-immunoprecipitation (Co-IP) experiment was performed to determine if there was an interaction between RAB31 and GLI1 proteins. GC cells were seeded in 6-well plates overnight. Then cells were scraped in the presence of RIPA lysis buffer. A volume of 2 μL primary antibody, 50 μL Protein A-Agarose (sc-2001; Santa Cruz Biotechnology, Dallas, TX, USA), and 500 μL phosphate-buffered saline (PBS) was mixed together and incubated for 2 h. Next, the cellular proteins were incubated with RAB31 (16182-1-AP; Proteintech), GLI1 (ab134906; Abcam), or IgG (BL003A, Bio-Sharp, Shanghai, China) primary antibody overnight at 4°C on a rocking platform. The next day, the cells were centrifuged at 3,000 rpm at 4°C and washed twice with PBS containing protein inhibitors, followed by Western blotting with the primary antibodies used for the Co-IP experiments.
Nude Mouse Subcutaneous Tumor Model
To determine the effects of RAB31 knockdown on tumor growth in vivo, we subcutaneously implanted MKN45 GC cells (5 × 106 in 0.15 mL PBS) into 4-week-old male BALB/c nude mice, as previously described (37). Once the xenograft tumors measured 3–4 mm, we randomly divided the mice into three groups: PBS, control, and RAB31 adenovirus groups. Then we treated the mice every second day and measured the tumor size for 15 days. Finally, after 20 days, we removed the tumors from the mice and measured the tumor weights.
Statistical Analysis
Statistical analysis was conducted as previously described (26). All of the values are shown as the mean ± standard deviation (SD) and were analyzed using SPSS and GraphPad software. The Kaplan–Meier survival curve of RAB31 was created to analyze the survival time of GC patients, and Cox regression analysis was performed to predict independent risk factors. A significant difference was identified using the two-tailed Student's t-test. P < 0.05 were considered statistically significant.
Results
Overexpression of RAB31 in GC Patients Predicts Poor Survival
To evaluate the expression of RAB31 in GC tissues, we measured RAB31 expression using the Oncomine database. As shown in Figures 1A–D, the level of RAB31 expression was noticeably higher in tumor tissues than in non-cancer tissues. To confirm the high level of RAB31 mRNA expression in tumor tissues, we collected 22 pairs of fresh tissues from GC patients and performed qPCR. RAB31 mRNA expression was markedly higher in the tumor tissues than in the normal tissues, and RAB31 mRNA could be regarded as a predictor based on an area under the curve of 0.8409 (Figures 1E,F). In addition, using the GEPIA tool, we found that expression of RAB31 varied in different clinicopathological stages and was enhanced in advanced GC, which was determined from the violin plot (Figure 1G). To investigate the level of RAB31 protein expression, we performed IHC experiments using tissue microarrays, and found that RAB31 protein expression was also higher in GC tissues compared to normal tissues (Figure 1H). Based on the IHC results, we divided patients into two groups (low RAB31 and high RAB31 expression), and evaluated specific clinical features between the two groups. As shown in Table 1, features such as tumor-node-metastasis (TNM), lymph node metastasis, and tumor size were significantly correlated with RAB31 expression. Moreover, we created a Kaplan–Meier survival curve and found that patients with high RAB31 expression had poorer survival (Figure 1I), consistent with the database results (Figure 1J). In a regression analysis model of survival, we found that patients with RAB31 overexpression had a poorer prognosis regardless of the results of the univariable or multiunivariable analysis (Table 2). TNM stage, lymph node metastasis, and distant metastasis were also prognostic predictors (Table 2).
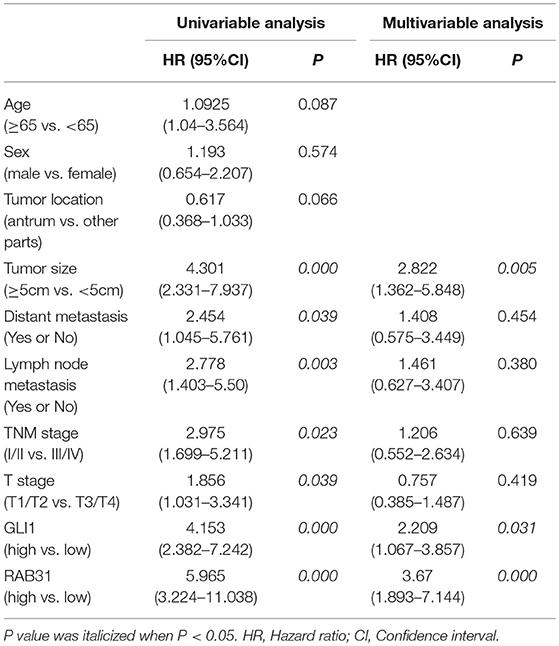
Table 2. Univariate and multivariate analyses of prognostic factors for survival among patients with GC.
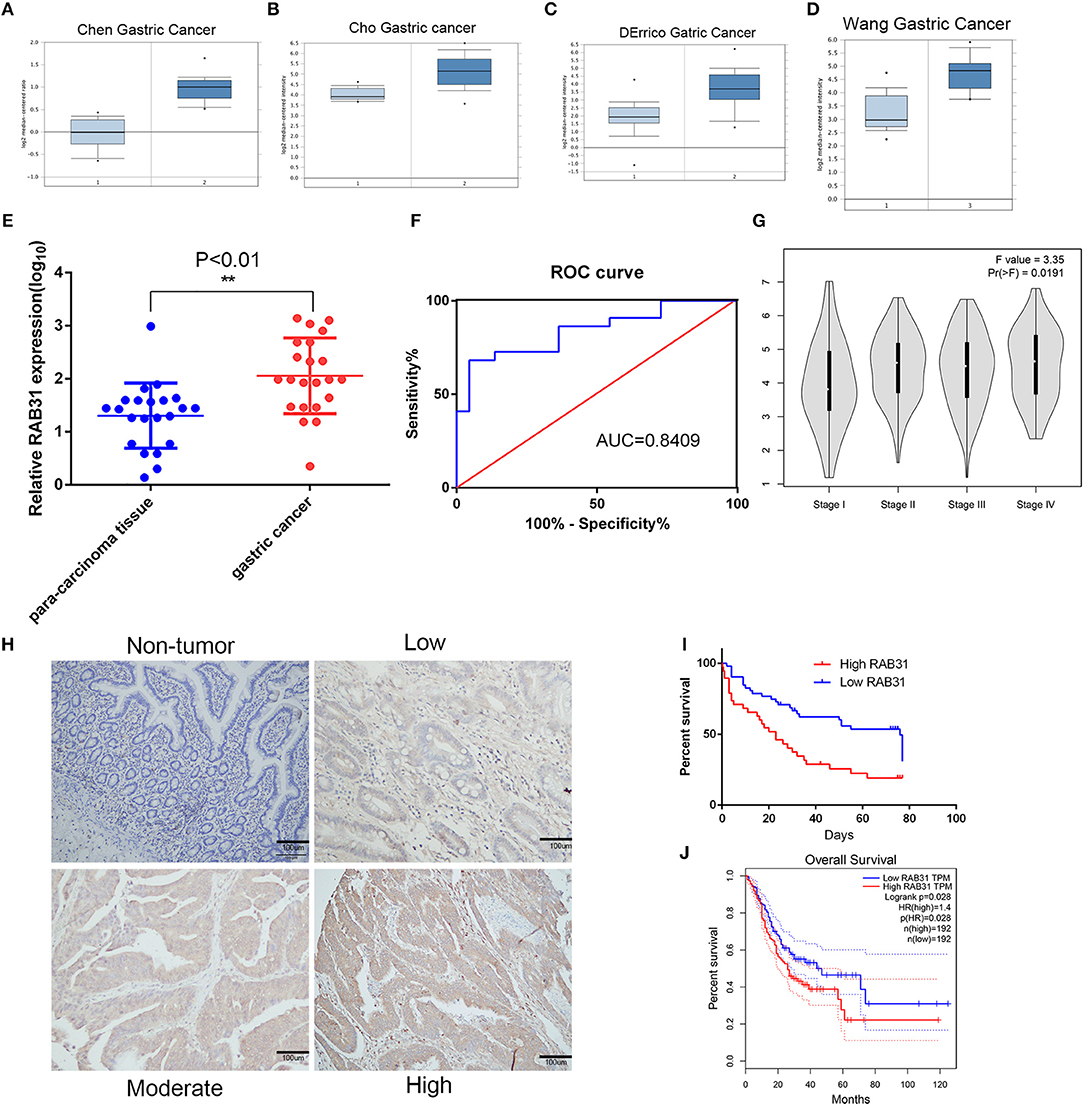
Figure 1. RAB31 is overexpressed in GC tissues and is positively correlated with poor prognosis. (A–D). Representative box plots of RAB31 expression from the Oncomine database (P < 0.001). (E) Expression level of RAB31 in cancer and para-carcinoma tissues of 22 patients as determined by qPCR (P < 0.01). (F) The receiver operating characteristic of specificity and sensitivity was created to assess potential predictive values. (G) The different expression levels of RAB31 in different stages of GC are shown by the violin plot, P < 0.05. (H) Representative IHC images of RAB31 staining in gastric tumor and non-tumor tissues (scale bar, 100 μm). (I) Overall survival analysis of GC patients with low versus high RAB31 expression. Survival rate was calculated by Kaplan–Meier survival analysis (n = 90; P < 0.001, log-rank test). (J) Survival curve was created using the GEPIA tool (P < 0.05).
RAB31 Promotes GC Cell Proliferation in vitro and in vivo and Inhibits Apoptosis
To determine the role of RAB31 in GC carcinogenesis, we measured RAB31 expression in GC cell lines. As shown in Figure 2A, RAB31 expression was highest in AGS and MKN45 cell lines. Then, we transfected siRAB31 into AGS and MKN45 cells and measured the effects of RAB31 knockdown by Western blotting (Figure 2B). As expected, RAB31 expression was reduced after transfection of siRAB31. Hence, we measured cell viability and colony formation using AGS and MKN45 cells following downregulation of RAB31 expression, and found that downregulation impaired cell growth and colony formation compared to control cells transfected with siNC (Figures 2C–E). Cell cycle and apoptosis assays showed that knockdown of RAB31 significantly halted the cell cycle in the G1 phase and promoted apoptosis (Figures 2F,G). Due to the decreased cell proliferation demonstrated by the in vitro assays, we examined the effects of RAB31 silencing on tumor growth in vivo using a tumor model in which AGS cells were injected subcutaneously into nude mice. As shown in Figures 3A,B, the size and weight of tumors with transfection of RAB31 siRNA adenovirus were smaller than those treated with scrambled adenovirus or PBS. Moreover, we created a tumor growth curve by measuring tumor size every other day, and found that RAB31 siRNA tumors grew more slowly than those in the control group (Figure 3C). Subsequently, we investigated the effects of RAB31 on tumor growth by IHC and found that Ki67 and RAB31 expression was lower in the RAB31 siRNA group than in the NC group (Figure 3D).
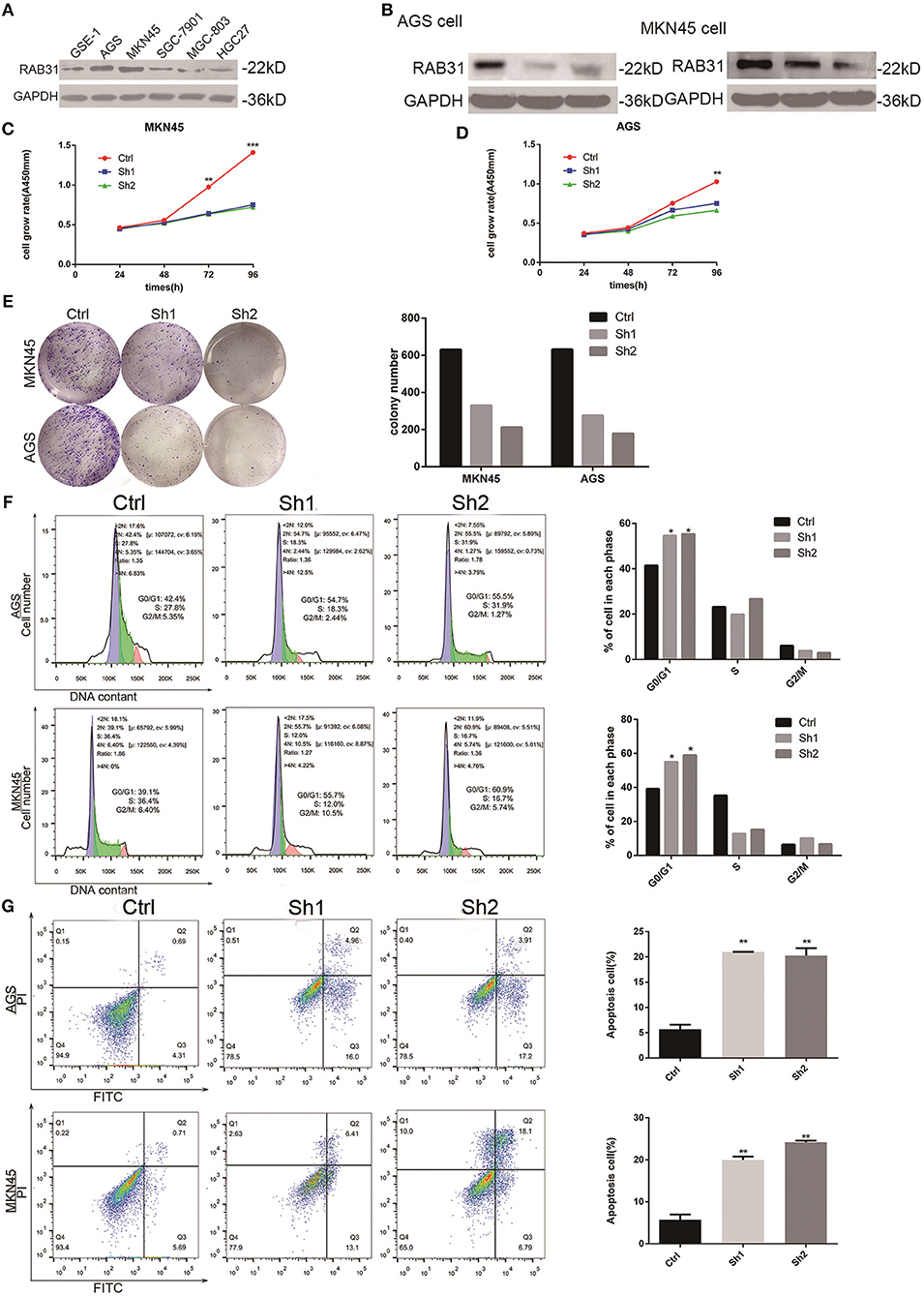
Figure 2. Depletion of RAB31 inhibits GC cell proliferation, blocks cell cycle progression, and promotes apoptosis in vitro. (A) Expression of RAB31 protein in six GC cell lines. (B) Western blot analysis of RAB31 in AGS and MKN45 cell lines after transfection of RAB31 siRNA; GAPDH was used as the control. (C,D) Viability of AGS and MKN45 cells after depleting RAB31 expression, as determined by the CCK8 assay. (E) Images from the colony formation assays in AGS and MKN45 cells are shown on the left; the average number of colonies formed in three experiments is shown by histogram on the right (P < 0.01). (F,G) Cell cycle and apoptosis were assessed by flow cytometry in AGS and MKN45 cell lines after knockdown of RAB31 expression; representative images are shown on the left and the results of triplicate assays are shown on the right. Results are shown as the mean ± SD (*P < 0.05, **P < 0.01, and ***P < 0.001).
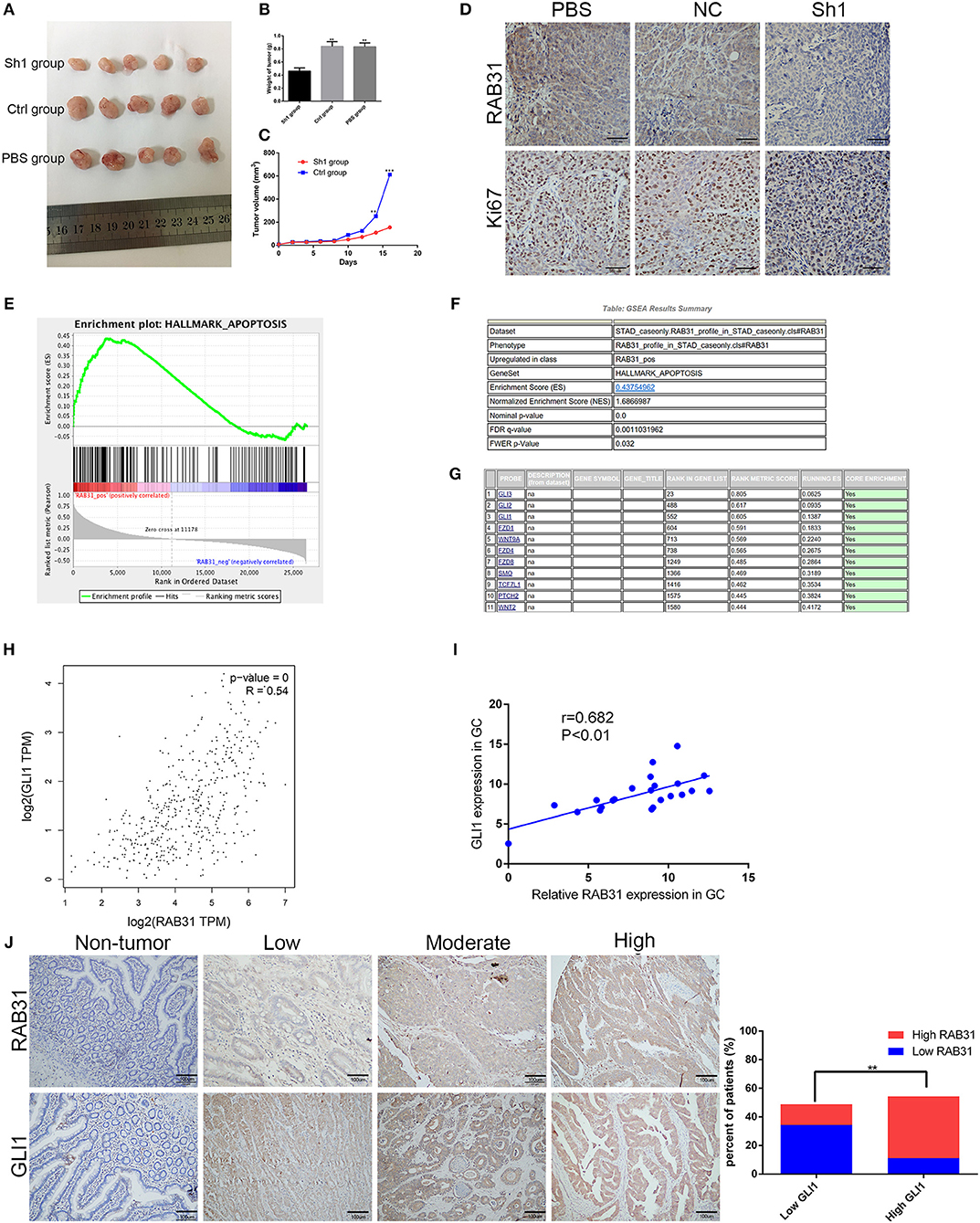
Figure 3. Depletion of RAB31 suppressed tumor growth in vivo and biological data predicted RAB31 function via interaction with GLI1. (A) Xenograft tumors were excised from the mice. (B,C) Measurements of xenograft tumor volumes and weights. (D) IHC staining of RAB31 and Ki-67 in tumor specimens from mouse tumor tissues (Scale bar, 100 μm). (E) GSEA analysis of the TCGA dataset showed the association between RAB31 expression and apoptosis signaling pathways. The enrichment score (ES, green line) equals the degree to which the gene set is overrepresented. (G) Genes involved in apoptosis are listed according to rank metric score. (H) The relationship between RAB31 and GLI1 was determined using the GEPIA tool. (I) Fresh tissues from 22 pairs of patients were analyzed for a correlation between RAB31 and GLI1 mRNA levels by qPCR. (J) Association of RAB31 expression with GLI1 expression in 90 primary human GC specimens; the representative images are shown on the left (scale bar, 100 μm). The columns show the significant results on the right (**P < 0.01, ***P < 0.001).
RAB31 Promotes GC Cell Proliferation and Inhibits Apoptosis via Targeting GLI1
The GLI1 pathway has been well documented as a downstream signaling pathway of RAB subfamily members (31, 38). Therefore, we performed gene set enrichment analysis (GSEA) to determine the function of RAB31 in GC, and found that GLI1 was involved in RAB31-mediated inhibition of apoptosis (Figures 3E–G). Then we tested the correlation between RAB31 and GLI1 expression by GEPIA, and found that RAB31 expression was positively associated with GLI1 expression (Figure 3H). Next, we performed qPCR using 22 pairs of fresh GC tissues and found that RAB31 expression was highly correlated with GLI1 expression (Figure 3I), consistent with the database results. To further analyze the relationship between RAB31 and GLI1, IHC of 90 tissue microarrays was performed to confirm the correlation between RAB31 and GLI1 expression. A marked association between the expression of RAB31 and GLI1 protein was observed in GC tissue (Figure 3J). Hence, we attempted to determine whether GLI1 mediates the function of RAB31 in GC cell growth and apoptosis. To this end, we created a cell model that overexpressed RAB31 by transducing RAB31 plasmid (Figure 4A). Subsequently, we performed several cell function experiments in RAB31-overexpressing cells with simultaneous GLI1 knockdown. According to our cell viability and colony formation assays, overexpression of RAB31 enhanced cell proliferation and knockdown of GLI1 expression blocked the effects induced by overexpression (Figures 4B–D). Similar results were obtained from the cell cycle and apoptosis assays, as overexpression of RAB31 promoted cell cycle progression and inhibited apoptosis, which was impaired by silencing of GLI1 expression (Figures 4E,F). To further demonstrate the molecular mechanisms of GLI1 regulation of RAB31, Western blotting was performed to determine if GLI1 protein levels were regulated by RAB31. The expression of GLI1 was reduced by silencing of RAB31, and expression of the cell cycle-related proteins cyclin D1 and c-Myc was also decreased, as well as that of the anti-apoptotic protein Bcl-2 and the pro-apoptosis protein BAX (Figures 5A,B). The enhanced expression of cyclin D1, c-Myc, and Bcl-2 proteins, and the decreased expression of BAX were due to RAB31 overexpression, which was impaired by knockdown of GLI1 expression (Figures 5C,D). We confirmed that RAB31 interacts with GLI1 by Co-IP and reverse Co-IP assays (Figures 5E,F). To verify the co-localization of RAB31 and GLI1, we performed immunofluorescence assays. As shown in Figure 5G, there was strong fluorescence intensity in the nucleus and moderate intensity in the cytoplasm and cytomembrane, illustrating that RAB31 protein was co-localized with GLI1. We also measured the immunofluorescence intensity of GLI1 and RAB31 proteins after overexpressing RAB31, and found that the intensity of GLI1 and RAB31 and that of the merged picture also increased. These results showed that the co-localization and correlation between RAB31 and GLI1 was stronger inmunofluorescence intensity (Figures 5H,I). Taken together, these data demonstrate that GLI1 mediates the effects of abnormal RAB31 expression on GC cell proliferation and apoptosis.
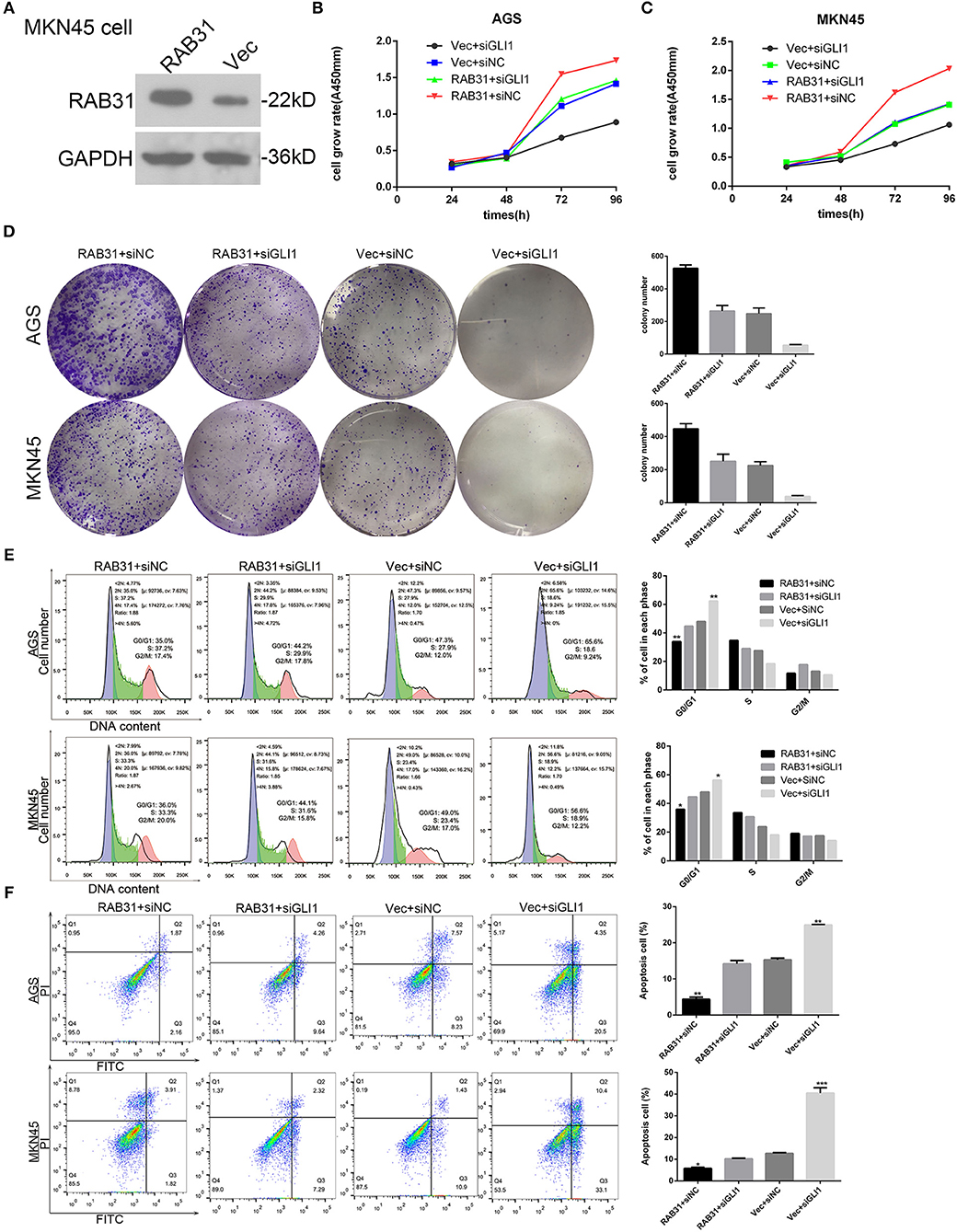
Figure 4. RAB31 promotes GC progression through activation of the GLI1 pathway in vivo. (A) Western blotting was performed to evaluate the overexpression of RAB31 after transfection of plasmids into MKN45 cells. (B–F) AGS and MKN45 cells with enhanced RAB31 expression and vector control were transfected simultaneously with GLI1 siRNA and siNC, respectively. (B,C) The viability of AGS and MKN45 cells was determined by the CCK8 assay. (D) Representative images of colony formation assays are shown on the left; the average colony formation from three experiments is shown by the histogram on the right (P < 0.01). (E,F) Cell cycle and apoptosis were assessed by flow cytometry. Representative images are on the left and the results of triplicate assays are shown on the right. Results are shown as the mean ± SD (*P < 0.05, **P < 0.01, and ***P < 0.001).
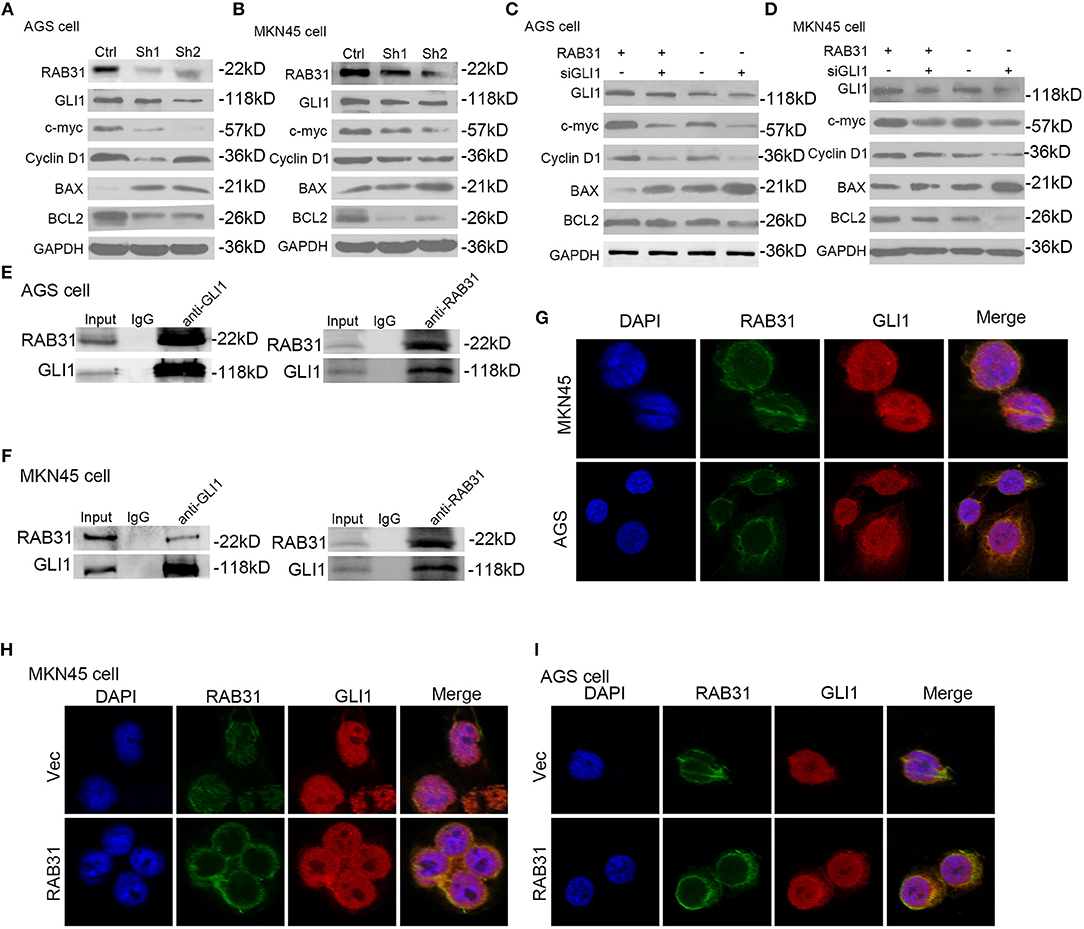
Figure 5. RAB31 mediated its downstream proteins via GLI1. (A,B) Expression of GLI1, cyclin D1, c-myc, BAX, and Bcl-2 were detected by Western blotting in AGS and MKN45 cells with RAB31 expression silenced. (C,D) Western blotting was conducted to detect the expression of GLI1, cyclin D1, c-myc, BAX, and Bcl-2 in AGS and MKN45 cells with simultaneous RAB31 plasmid and GLI1 siRNA co-transfection. (E,F) Co-IP assay was utilized to investigate the interaction between RAB31 and GLI1 expression. The assay was conducted using a GLI1 antibody to pulldown the RAB31 protein or a RAB31 antibody to pulldown the GLI1 protein. (G) Co-localization of the GLI1 and RAB31 proteins was evaluated by immunofluorescence. (H,I) Immunofluorescence was conducted to measure the fluorescence intensity of RAB31 and GLI1 in AGS and MKN45 cells transfected with the RAB31 plasmid and vector controls.
Low miR-30c-2-3p Expression in GC Tissues Predicts Poor Survival and Regulates Cell Proliferation
To evaluate the regulation of RAB31 by miRNA, we used miRWalk 2.0, miRanda, and Targetscan tools to predict the miRNAs that may bind the 3′-UTR of RAB31 (Data Sheet 2). Then we performed KEGG pathway analysis of the identified miRNAs to detect the miRNAs involved in the GLI1 pathway; these miRNAs were ranked by P-value (Data Sheet 2). As shown in Figure 6A, miR-30c-2-3p was highly involved in the RAB31/GLI1 pathway. Additionally, we tested the level of miR-30c-2-3p expression in GC tissues using the YM500v2 database (Data Sheet 3) (36). The heat map and bar graph of miR-30c-2-3p expression between tumor tissues para-carcinoma tissues showed that miR-30c-2-3p expression was low in GC; therefore, miR-30c-2-3p is a potential tumor suppressor gene (Figures 6B,C). Likewise, Kaplan–Meier analysis, which was performed using The Cancer Genome Atlas (TCGA) database and was based on the average expression of miR-30c-2-3p, showed that GC patients with low miR-30c-2-3p expression had poorer survival (Figure 6D). In addition, we confirmed the role of miR-30c-2-3p in GC cell growth and apoptosis in vitro. As shown in Figure 6E, we constructed a cell model with low miR-30c-2-3p expression or miR-30c-2-3p overexpression by transfecting an miR-30c-2-3p inhibitor or mimic. Downregulation of miR-30c-2-3p in AGS and MKN45 cells by transfecting an miR-30c-2-3p inhibitor enhanced cell growth and colony formation, which were blocked by transfection of miR-30c-2-3p mimics (Figures 6F–H). In addition, flow cytometry showed that miR-30c-2-3p mimics blocked the cell cycle transition between the G0/G1 and S1 phase in AGS and MKN45 cells, and promoted apoptosis, compared with the miR-30c-2-3p inhibitor group (Figures 6I,J).
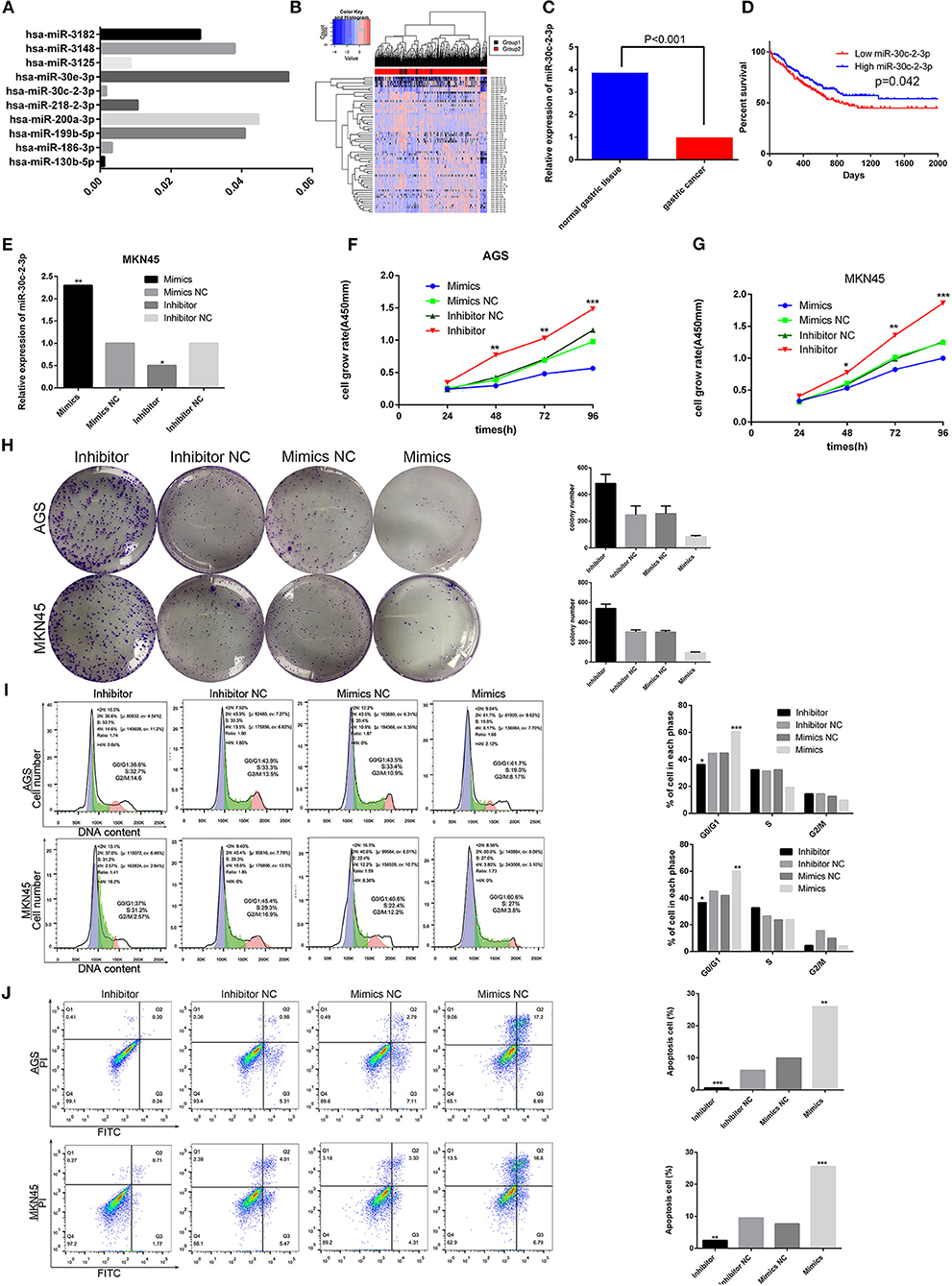
Figure 6. miR-30c-2-3p has low expression in GC and inhibits GC cell proliferation, blocks the cell cycle, and promotes apoptosis in vitro. (A) The results obtained on the Hh signaling pathway via analysis using Targetscan, miRWalk 2.0, miRDB databases, and KEGG pathway analysis are shown by the histogram, according to the P-value. (B,C) The heat plot showing miRNA expression in GC and the histogram showing miR-30c-2-3p expression were created using the YM500v2 tool. (D) Kaplan–Meier curve for overall survival was created using clinical information from the TCGA databank in GC patients with high and low miR-30c-2-3p expression levels (P < 0.05). (E) qPCR analysis of miR-30c-2-3p in MKN45 cells after transfection of mimics or the inhibitor. (F,G) Viability of AGS and MKN45 cells after upregulation or downregulation of miR-30-2-3p expression was determined by the CCK8 assay. (H) Significant images of colony formation assays using AGS and MKN45 cells are shown on the left, and the average colony formation from three experiments is shown by histogram on the right (P < 0.01). (I,J) Cell cycle and apoptosis were assessed by flow cytometry in AGS and MKN45 cell lines transfected with miR-30c-2-3p mimics and the inhibitor. Representative images are shown on the left and the results of triplicate assays are on the right. Results are shown as the mean ± SD (*P < 0.05, **P < 0.01, and ***P < 0.001).
MiR-30c-2-3p Is Involved in the RAB31/GLI1 Pathway Signaling by Targeting RAB31
To characterize the mechanisms of miR-30c-2-3p inhibition of cell proliferation, we performed a luciferase reporter assay to test whether miR-30c-2-3p binds the 3′-UTR of RAB31. We co-transfected wild-type or mutant plasmids of the 3′-UTR with miR-30c-2-3p mimics or a miR-30c-2-3p inhibitor into MKN45 cells and measured the fluorescence intensity. As expected, the miR-30c-2-3p mimics and miR-30c-2-3p inhibitor highly impaired or increased the fluorescence intensity, respectively (Figure 7A). Western blotting showed that expression of the RAB31 and GLI1 proteins and their downstream proteins was markedly mediated by miR-30c-2-3p (Figures 7B,C). To further demonstrate that miR-30c-2-3p functions as a suppressor gene by targeting RAB31, we co-transfected the miR-30c-2-3p inhibitor with RAB31 siRNA, and found that the enhanced cell proliferation induced by the miR-30c-2-3p inhibitor was impaired by RAB31 siRNA (Figures 7D,E). Consistent with the cell viability assay, the increased expression of RAB31 and GLI1 proteins and their downstream proteins induced by miR-30c-2-3p was impaired by RAB31 siRNA (Figures 7F, G). These data showed that miR-30c-2-3p regulated GLI1 and its downstream proteins by targeting RAB31.
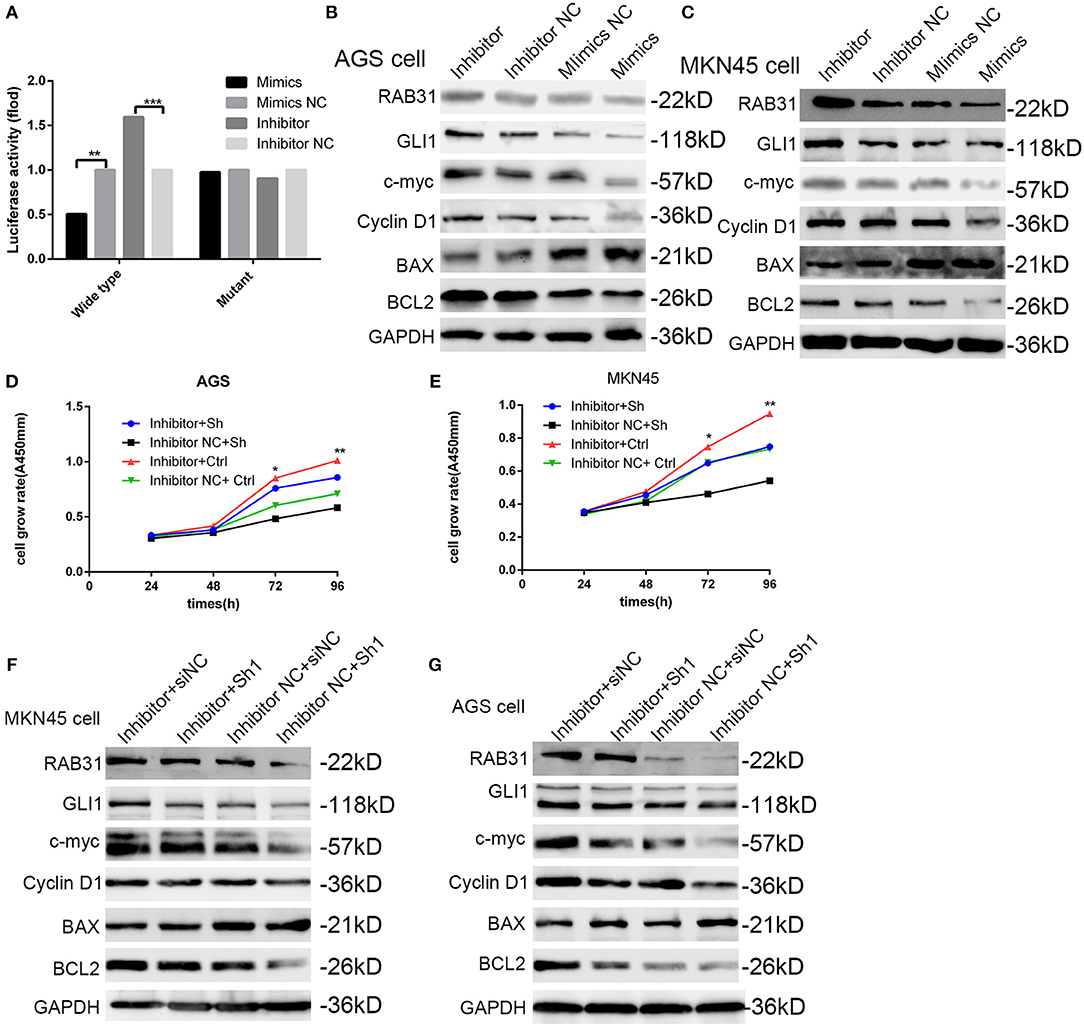
Figure 7. miR-30c-2-3p is involved in the RAB31/GLI1 pathway and regulates GC cell growth and apoptosis via binding to the 3′-UTR of RAB31. (A) Luciferase reporter assay was performed using MKN45 cells to determine if miR-30c-2-3p binds to the 3′-UTR of RAB31. (B,C) RAB31, GLI1, cyclin D1, BAX, and Bcl-2 expression was detected by Western blotting in AGS and MKN45 cells after transfection with miR-30c-2-3p mimics or the inhibitor; GAPDH was used as the control. (D,E) Cell viability was assessed in AGS and MKN45 cells transfected with the miR-30c-2-3p inhibitor and RAB31 siRNA. (F,G) Western blotting was conducted to investigate the levels of RAB31, GLI1, and its downstream proteins in AGS and MKN45 cells co-transfected with the miR-30c-2-3p inhibitor.
Discussion
The results of this study illustrated the roles of RAB31 in GC cell proliferation and apoptosis, and also showed that miR-30c-2-3p regulated RAB31, which subsequently mediated cell growth and apoptosis via the GLI1 signaling pathway. In addition, we found that the overexpression of RAB31 predicted poor survival times in GC patients.
GC is one of the most malignant cancers worldwide, and is responsible for 8–15% of all cancer-related deaths (39). The rapid growth and early metastasis of tumor cells, which are dependent on nutrient transport, are the main causes of GC malignancy and the poor prognosis of GC patients (40). RAB GTPases are critical regulators of membrane trafficking, and mediate many biological processes of which dysregulation may lead to disease (41–44). Recently, they were identified as being important stimuli for tumorigenesis (45–47). For example, RAB1 serves as a significant mediator of endoplasmic reticulum-to-Golgi transport, which regulates cellular function and is linked to different cellular signaling pathways such as nutrient signaling, Notch signaling, and autophagy (45). Similarly, RAB31 has been shown to promote cell proliferation and inhibit apoptosis in some cancers,such as breast, hepatocellular, and ovarian cancers (11, 14, 48). Here, we uncovered a novel mechanism by which RAB31 promotes GC progression. We found that the expression of RAB31 was upregulated in GC tissues through qPCR analysis of fresh tissues and IHC of tissue microarrays (Figures 1E, H). We also found that the overexpression of RAB31 was significantly associated with specific clinicopathological characteristics and a shorter survival time, which strongly suggests that RAB31 may be a novel prognostic biomarker for GC. In addition, in vitro and in vivo assays demonstrated that the depletion of RAB31 attenuated cell growth and promoted apoptosis.
Previously, we confirmed that GLI1 is a significant mediator of GC progression, in accordance with the conclusions of other studies (26, 49, 50). Furthermore, RAB23, similar to RAB31, has been confirmed as a powerful regulator of tumorigenesis via the interaction with GLI1 signaling (38, 51, 52). Therefore, we performed GSEA using the TCGA databank and found that RAB31 regulates apoptosis by targeting GLI1. We also performed Co-IP assays and immunofluorescence experiments and found that RAB31 interacted and co-localized with GLI1 (Figures 5E, F). The rescue experiments showed that the depletion of GLI1 attenuated the effects on cell proliferation, the cell cycle, and apoptosis induced by the overexpression of RAB31, suggesting that GLI1 regulates the oncogenic functions of RAB31. Furthermore, through analysis of the miRNA database, we found that the RAB31/GLI1 axis was regulated by miR-30c-2-3p. Previous studies showed that miR-30c-2-3p functions as a tumor suppressor gene by targeting cellular signaling factors in various cancers such as breast cancer and renal cell carcinoma (53, 54). In GC, it was shown that miR-30c-2-3p exhibits low expression and is linked to clinical features such as TNM stage (55). Similarly, we found that patients with low miR-30c-2-3p expression were predicted to have poor survival rates (Figures 6C, D). Moreover, in the in vitro assays, the upregulation or downregulation of miR-30c-2-3p expression inhibited cell growth or promoted cell growth, respectively. Similar to other miRNAs that degrade or inhibit specific mRNAs by binding the 3′-UTR of targeted genes (56), we demonstrated that miR-30c-2-3p functioned through binding the 3′-UTR of RAB31 and subsequently mediated GLI1 expression. Interestingly, the inhibition of miR-30c-2-3p rescued the growth inhibition of GC cells induced by silencing of RAB31 (Figures 7D, G). Thus, the results of our study provide evidence to support the novel regulatory axis of miR-30c-2-3p/RAB31/GLI1 in the development of GC.
Conclusions
In conclusion, the results of our study provide robust evidence to support the function of RAB31 in the regulation of GC cell proliferation and apoptosis. Moreover, we uncovered a novel mechanism whereby downregulation of RAB31 suppresses the expression of GLI1 and inhibits the expression of cyclin D1, c-Myc, BAX, and Bcl-2 proteins, leading to GC cell growth retardation, cell cycle arrest, and apoptosis. We also demonstrated that miR-30c-2-3p is involved in the RAB31/GLI1 axis by targeting the 3′-UTR of RAB31 and regulating cell proliferation. Therefore, our data demonstrates the great significance of miR-30c-2-3p/RAB31/GLI1 signaling in the development of GC, which may serve as a potential target for GC therapy.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article and its additional files.
Ethics Approval and Consent to Participate
The research was approved by the Ethics Committee of Renji Hospital, School of Medicine, Shanghai Jiao Tong University.
Author Contributions
C-TT, QL, and Z-ZG conceived and designed the study. C-TT and LY performed the experiments and drafted the manuscript. SW and X-LL helped to analyze the data. Y-JG, X-TZ, and YC helped to collect the information on patients. Z-ZG supervised the experiments and revised the manuscript. All of the authors reviewed the manuscript and approved the final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 81670505), the Shanghai Municipal Education Commission: Gaofeng Clinical Medicine grant support (No. DLY201501), and the Three-year action plan for Shin Kang of Shanghai (No. 16CR3113B).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2018.00554/full#supplementary-material
Abbreviations
GC, Gastric cancer; GLI1, GLI Family Zinc Finger 1; RAB31, Member RAS Oncogene Family 31; IHC, immunohistochemistry; GSEA, gene set enrichment analysis; GEPIA, Gene Expression Profiling Interactive Analysis.
References
1. Parkin DM. Global cancer statistics in the year 2000. Lancet Oncol. (2001) 2:533–43. doi: 10.1016/S1470-2045(01)00486-7
2. Yuasa Y. Control of gut differentiation and intestinal-type gastric carcinogenesis. Nat Rev Cancer (2003) 3:592–600. doi: 10.1038/nrc1141
3. Lordick F, Siewert JR. Recent advances in multimodal treatment for gastric cancer: a review. Gastric Cancer (2005) 8:78–85. doi: 10.1007/s10120-005-0321-z
4. Wohrer SS, Raderer M, Hejna M. Palliative chemotherapy for advanced gastric cancer. Ann Oncol. (2004) 15:1585–95. doi: 10.1093/annonc/mdh422
5. Diekmann Y, Seixas E, Gouw M, Tavares-Cadete F, Seabra MC, Pereira-Leal JB. Thousands of rab GTPases for the cell biologist. PLoS Comput Biol. (2011) 7:e1002217. doi: 10.1371/journal.pcbi.1002217
6. Chen D, Guo J, Miki T, Tachibana M, Gahl WA. Molecular cloning of two novel rab genes from human melanocytes. Gene (1996) 174:129–34. doi: 10.1016/0378-1119(96)00509-4
7. Rodriguez-Gabin AG, Cammer M, Almazan G, Charron M, Larocca JN. Role of rRAB22b, an oligodendrocyte protein, in regulation of transport of vesicles from trans Golgi to endocytic compartments. J Neurosci Res. (2001) 66:1149–60. doi: 10.1002/jnr.1253
8. Hou JC, Pessin JE. Ins (endocytosis) and outs (exocytosis) of GLUT4 trafficking. Curr Opin Cell Biol. (2007) 19:466–73. doi: 10.1016/j.ceb.2007.04.018
9. Ng EL, Ng JJ, Liang F, Tang BL. Rab22B is expressed in the CNS astroglia lineage and plays a role in epidermal growth factor receptor trafficking in A431 cells. J Cell Physiol. (2009) 221:716–28. doi: 10.1002/jcp.21911
10. Rodriguez-Gabin AG, Yin X, Si Q, Larocca JN. Transport of mannose-6-phosphate receptors from the trans-Golgi network to endosomes requires Rab31. Exp Cell Res. (2009) 315:2215–30. doi: 10.1016/j.yexcr.2009.03.020
11. Chua CE, Tang BL. The role of the small GTPase Rab31 in cancer. J Cell Mol Med. (2015) 19:1–10. doi: 10.1111/jcmm.12403
12. Sui Y, Zheng X, Zhao D. Rab31 promoted hepatocellular carcinoma (HCC) progression via inhibition of cell apoptosis induced by PI3K/AKT/Bcl-2/BAX pathway. Tumour Biol. (2015) 36:8661–70. doi: 10.1007/s13277-015-3626-5
13. Kotzsch M, Sieuwerts AM, Grosser M, Meye A, Fuessel S, Meijer-van Gelder ME, et al. Urokinase receptor splice variant uPAR-del4/5-associated gene expression in breast cancer: identification of rab31 as an independent prognostic factor. Breast Cancer Res Treat (2008) 111:229–40. doi: 10.1007/s10549-007-9782-6
14. Grismayer B, Solch S, Seubert B, Kirchner T, Schafer S, Baretton G, et al. Rab31 expression levels modulate tumor-relevant characteristics of breast cancer cells. Mol Cancer (2012) 11:62. doi: 10.1186/1476-4598-11-62
15. Jin C, Rajabi H, Pitroda S, Li A, Kharbanda A, Weichselbaum R, et al. Cooperative interaction between the MUC1-C oncoprotein and the Rab31 GTPase in estrogen receptor-positive breast cancer cells. PLoS ONE (2012) 7:e39432. doi: 10.1371/journal.pone.0039432
16. Kharbanda A, Rajabi H, Jin C, Raina D, Kufe D. Oncogenic MUC1-C promotes tamoxifen resistance in human breast cancer. Mol Cancer Res. (2013) 11:714–23. doi: 10.1158/1541-7786.MCR-12-0668
17. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell (2004) 116:281–97. doi: 10.1016/S0092-8674(04)00045-5
18. Hausser J, Syed AP, Bilen B, Zavolan M. Analysis of CDS-located miRNA target sites suggests that they can effectively inhibit translation. Genome Res. (2013) 23:604–15. doi: 10.1101/gr.139758.112
19. Ye YY, Mei JW, Xiang SS, Li HF, Ma Q, Song XL, et al. MicroRNA-30a-5p inhibits gallbladder cancer cell proliferation, migration and metastasis by targeting E2F7. Cell Death Dis. (2018) 9:410. doi: 10.1038/s41419-018-0444-x
20. De Gregorio R, Pulcrano S, De Sanctis C, Volpicelli F, Guatteo E, von Oerthel L, et al. miR-34b/c regulates Wnt1 and enhances mesencephalic dopaminergic neuron differentiation. Stem Cell Rep. (2018) 10:1237–50. doi: 10.1016/j.stemcr.2018.02.006
21. Yu L, Wu D, Gao H, Balic JJ, Tsykin A, Han TS, et al. Clinical utility of a STAT3-regulated miRNA-200 family signature with prognostic potential in early gastric cancer. Clin Cancer Res. (2018) 24:1459–72. doi: 10.1158/1078-0432.CCR-17-2485
22. Huang T, Zhou Y, Zhang J, Wong CC, Li W, Kwan JSH, et al. SRGAP1, a crucial target of miR-340 and miR-124, functions as a potential oncogene in gastric tumorigenesis. Oncogene (2018) 37:1159–74. doi: 10.1038/s41388-017-0029-7
23. Xie K, Abbruzzese JL. Developmental biology informs cancer: the emerging role of the hedgehog signaling pathway in upper gastrointestinal cancers. Cancer Cell (2003) 4:245–7. doi: 10.1016/S1535-6108(03)00246-0
24. Wessler S, Krisch LM, Elmer DP, Aberger F. From inflammation to gastric cancer - the importance of Hedgehog/GLI signaling in Helicobacter pylori-induced chronic inflammatory and neoplastic diseases. BMC (2017) 15:15. doi: 10.1186/s12964-017-0171-4
25. Merchant JL, Ding L. Hedgehog signaling links chronic inflammation to gastric cancer precursor lesions. Cell Commun Signal. (2017) 3:201–10. doi: 10.1016/j.jcmgh.2017.01.004
26. Tang CT, Lin XL, Wu S, Liang Q, Yang L, Gao YJ, et al. NOX4-driven ROS formation regulates proliferation and apoptosis of gastric cancer cells through the GLI1 pathway. Cell Signal (2018) 46:52–63. doi: 10.1016/j.cellsig.2018.02.007
27. Kageyama-Yahara N, Yamamichi N, Takahashi Y, Nakayama C, Shiogama K, Inada K, et al. Gli regulates MUC5AC transcription in human gastrointestinal cells. PLoS ONE (2014) 9:e106106. doi: 10.1371/journal.pone.0106106
28. Kim JH, Shin HS, Lee SH, Lee I, Lee YS, Park JC, et al. Contrasting activity of Hedgehog and Wnt pathways according to gastric cancer cell differentiation: relevance of crosstalk mechanisms. Cancer Sci. (2010) 101:328–35. doi: 10.1111/j.1349-7006.2009.01395.x
29. Yu B, Gu D, Zhang X, Liu B, Xie J. The role of GLI2-ABCG2 signaling axis for 5Fu resistance in gastric cancer. J Genet Genomics (2017) 44:375–83. doi: 10.1016/j.jgg.2017.04.008
30. Hu Q, Hou YC, Huang J, Fang JY, Xiong H. Itraconazole induces apoptosis and cell cycle arrest via inhibiting Hedgehog signaling in gastric cancer cells. J Exp Clin Cancer Res. (2017) 36:50. doi: 10.1186/s13046-017-0526-0
31. Oro AE. The primary cilia, a “Rab-id” transit system for hedgehog signaling. Curr Opin Cell Biol. (2007) 19:691–6. doi: 10.1016/j.ceb.2007.10.008
32. Zhang W, Yu F, Wang Y, Zhang Y, Meng L, Chi Y. Rab23 promotes the cisplatin resistance of ovarian cancer via the Shh-Gli-ABCG2 signaling pathway. Oncol Lett. (2018) 15:5155–60. doi: 10.3892/ol.2018.7949
33. Liu Y, Zeng C, Bao N, Zhao J, Hu Y, Li C, et al. Effect of Rab23 on the proliferation and apoptosis in breast cancer. Oncol Rep. (2015) 34:1835–44. doi: 10.3892/or.2015.4152
34. Chang J, Xu W, Liu G, Du X, Li X. Downregulation of Rab23 in prostate cancer inhibits tumor growth in vitro and in vivo. Oncol Res. (2017) 25:241–8. doi: 10.3727/096504016X14742891049118
35. Lin XL, Yang L, Fu SW, Lin WF, Gao YJ, Chen HY, et al. Overexpression of NOX4 predicts poor prognosis and promotes tumor progression in human colorectal cancer. Oncotarget (2017) 8:33586–600. doi: 10.18632/oncotarget.16829
36. Cheng WC, Chung IF, Tsai CF, Huang TS, Chen CY, Wang SC, et al. YM500v2: a small RNA sequencing (smRNA-seq) database for human cancer miRNome research. Nucleic Acids Res. (2015) 43:D862–7. doi: 10.1093/nar/gku1156
37. Yan TT, Ren LL, Shen CQ, Wang ZH, Yu YN, Liang Q, et al. miR-508 defines the stem-like/mesenchymal subtype in colorectal cancer. Cancer Res. (2018) 78:1751–65. doi: 10.1158/0008-5472.CAN-17-2101
38. Chi S, Xie G, Liu H, Chen K, Zhang X, Li C, et al. Rab23 negatively regulates Gli1 transcriptional factor in a Su(Fu)-dependent manner. Cell Signal (2012) 24:1222–8. doi: 10.1016/j.cellsig.2012.02.004
39. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. (2015) 65:87–108. doi: 10.3322/caac.21262
40. Matsuoka T, Yashiro M. Rho/ROCK signaling in motility and metastasis of gastric cancer. World J Gastroenterol. (2014) 20:13756–66. doi: 10.3748/wjg.v20.i38.13756
41. Kiral FR, Kohrs FE, Jin EJ, Hiesinger PR. Rab GTPases and membrane trafficking in neurodegeneration. Curr Biol. (2018) 28:R471–86. doi: 10.1016/j.cub.2018.02.010
42. Jalagadugula G, Goldfinger LE, Mao G, Lambert MP, Rao AK. Defective RAB1B-related megakaryocytic ER-to-Golgi transport in RUNX1 haplodeficiency: impact on von Willebrand factor. Blood Adv. (2018) 2:797–806. doi: 10.1182/bloodadvances.2017014274
43. Fremont S, Echard A. Membrane Traffic in the late steps of cytokinesis. Curr Biol. (2018) 28:R458–70. doi: 10.1016/j.cub.2018.01.019
44. Actis Dato V, Grosso RA, Sanchez MC, Fader CM, Chiabrando GA. Insulin-induced exocytosis regulates the cell surface level of low density lipoprotein-related protein-1 in Muller Glial cells. Biochem J. (2018) 475:1669–85. doi: 10.1042/BCJ20170891
45. Yang XZ, Li XX, Zhang YJ, Rodriguez-Rodriguez L, Xiang MQ, Wang HY, et al. Rab1 in cell signaling, cancer and other diseases. Oncogene (2016) 35:5699–704. doi: 10.1038/onc.2016.81
46. Wang S, Hu C, Wu F, He S. Rab25 GTPase: functional roles in cancer. Traffic (2017) 8:64591–9. doi: 10.18632/oncotarget.19571
47. Shaughnessy R, Echard A. Rab35 GTPase and cancer: linking membrane trafficking to tumorigenesis. Traffic (2018) 19:247–52. doi: 10.1111/tra.12546
48. Pan Y, Zhang Y, Chen L, Liu Y, Feng Y, Yan J. The critical role of Rab31 in cell proliferation and apoptosis in cancer progression. Mol Neurobiol. (2016) 53:4431–7. doi: 10.1007/s12035-015-9378-9
49. Zhao Y, Huang J, Zhang L, Qu Y, Li J, Yu B, et al. MiR-133b is frequently decreased in gastric cancer and its overexpression reduces the metastatic potential of gastric cancer cells. BMC Cancer (2014) 14:34. doi: 10.1186/1471-2407-14-34
50. Ohta M, Tateishi K, Kanai F, Watabe H, Kondo S, Guleng B, et al. p53-Independent negative regulation of p21/cyclin-dependent kinase-interacting protein 1 by the sonic hedgehog-glioma-associated oncogene 1 pathway in gastric carcinoma cells. Cancer Res. (2005) 65:10822–9. doi: 10.1158/0008-5472.CAN-05-0777
51. Yang L, Clinton JM, Blackburn ML, Zhang Q, Zou J, Zielinska-Kwiatkowska A, et al. Rab23 regulates differentiation of ATDC5 chondroprogenitor cells. J Biol Chem. (2008) 283:10649–57. doi: 10.1074/jbc.M706795200
52. Sun HJ, Liu YJ, Li N, Sun ZY, Zhao HW, Wang C, et al. Sublocalization of Rab23, a mediator of Sonic hedgehog signaling pathway, in hepatocellular carcinoma cell lines. Mol Med Rep. (2012) 6:1276–80. doi: 10.3892/mmr.2012.1094
53. Shukla K, Sharma AK, Ward A, Will R, Hielscher T, Balwierz A, et al. MicroRNA-30c-2–3p negatively regulates NF-kappaB signaling and cell cycle progression through downregulation of TRADD and CCNE1 in breast cancer. Mol Oncol. (2015) 9:1106–19. doi: 10.1016/j.molonc.2015.01.008
54. Mathew LK, Lee SS, Skuli N, Rao S, Keith B, Nathanson KL, et al. Restricted expression of miR-30c-2–3p and miR-30a-3p in clear cell renal cell carcinomas enhances HIF2alpha activity. Cancer Discov. (2014) 4:53–60. doi: 10.1158/2159-8290.CD-13-0291
55. Li CY, Liang GY, Yao WZ, Sui J, Shen X, Zhang YQ, et al. Identification and functional characterization of microRNAs reveal a potential role in gastric cancer progression. Clin Transl Oncol. (2017) 19:162–72. doi: 10.1007/s12094-016-1516-y
Keywords: RAB31, GLI1, gastric cancer, miR-30c-2-3p, prognosis, proliferation
Citation: Tang C-T, Liang Q, Yang L, Lin X-L, Wu S, Chen Y, Zhang X-T, Gao Y-J and Ge Z-Z (2018) RAB31 Targeted by MiR-30c-2-3p Regulates the GLI1 Signaling Pathway, Affecting Gastric Cancer Cell Proliferation and Apoptosis. Front. Oncol. 8:554. doi: 10.3389/fonc.2018.00554
Received: 16 August 2018; Accepted: 08 November 2018;
Published: 26 November 2018.
Edited by:
Pashtoon Murtaza Kasi, Mayo Clinic, United StatesReviewed by:
Toru Furukawa, School of Medicine, Tohoku University, JapanQingfeng Zhu, Johns Hopkins Medicine, United States
Copyright © 2018 Tang, Liang, Yang, Lin, Wu, Chen, Zhang, Gao and Ge. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhi-Zheng Ge, zhizhengge@aliyun.com
†These authors have contributed equally to this work