 Tian-Bo Ding
Tian-Bo Ding Jie Li1
Jie Li1 Er-Hu Chen
Er-Hu Chen Jin-Zhi Niu
Jin-Zhi Niu- 1Key Laboratory of Integrated Crop Pest Management of Shandong Province, College of Plant Health and Medicine, Qingdao Agricultural University, Qingdao, China
- 2Key Laboratory of Entomology and Pest Control Engineering, College of Plant Protection, Southwest University, Chongqing, China
Tomato yellow leaf curl virus (TYLCV) and Tomato chlorosis virus (ToCV) are two of the most devastating cultivated tomato viruses, causing significant crop losses worldwide. As the vector of both TYLCV and ToCV, the whitefly Bemisia tabaci Mediterranean (MED) is mainly responsible for the rapid spread and mixed infection of TYLCV and ToCV in China. However, little is known concerning B. tabaci MED's molecular response to TYLCV and ToCV infection or their co-infection. We determined the transcriptional responses of the whitefly MED to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection using Illumina sequencing. In all, 78, 221, and 60 differentially expressed genes (DEGs) were identified in TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected whiteflies, respectively, compared with non-viruliferous whiteflies. Differentially regulated genes were sorted according to their roles in detoxification, stress response, immune response, transport, primary metabolism, cell function, and total fitness in whiteflies after feeding on virus-infected tomato plants. Alterations in the transcription profiles of genes involved in transport and energy metabolism occurred between TYLCV&ToCV co-infection and single infection with TYLCV or ToCV; this may be associated with the adaptation of the insect vector upon co-infection of the two viruses. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses demonstrated that the single infection with TYLCV or ToCV and the TYLCV&ToCV co-infection could perturb metabolic processes and metabolic pathways. Taken together, our results provide basis for further exploration of the molecular mechanisms of the response to TYLCV, ToCV single infection, and TYLCV&ToCV co-infection in B. tabaci MED, which will add to our knowledge of the interactions between plant viruses and insect vectors.
Introduction
More than 75% of plant viruses are transmitted by insect vectors, most of which belong to hemipteran (suborder Homoptera) families, such as whiteflies, aphids, and planthoppers (Hogenhout et al., 2008). Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), commonly known as the sweetpotato whitefly, has caused tremendous damage to the tomato crops in greenhouses and fields worldwide (Valverde et al., 2004). B. tabaci is a cryptic species complex consisting of at least 39 cryptic species (Alemandri et al., 2015), among which the Middle East-Asia Minor (MEAM1) (formerly referred to as biotype B) and the Mediterranean (MED) (formerly referred to as biotype Q) species have become the most destructive alien species in many regions of the world (De Barro et al., 2011). The whitefly MED was first detected in China in 2003 (Chu et al., 2006), and has gradually displaced MEAM1 and become the dominant cryptic species of B. tabaci in China (Pan et al., 2011; Rao et al., 2011). The whitefly is an effective vector of numerous plant viruses, the majority of which are Begomoviruses. Several viruses belonging to other genera, including Crinivirus, Ipomovirus, Torradovirus, and Carlavirus, can also be transmitted by whiteflies (Navas-Castillo et al., 2011; Polston et al., 2014).
Tomato yellow leaf curl virus (TYLCV) (Geminiviridae), the type member of the genus Begomovirus, is one of the most devastating viruses of cultivated tomato worldwide; the virus is transmitted by B. tabaci in a persistent circulative manner (Czosnek, 2007; Hogenhout et al., 2008). In China, TYLCV was first detected in 2006 in Shanghai (Wu et al., 2006), and has rapidly spread to many provinces, including Zhejiang, Jiangsu, Shandong, Hebei, and Beijing (Ji et al., 2008; Mugiira et al., 2008; Sun et al., 2009; Zhang et al., 2010; Zhou et al., 2010; Pan et al., 2012). A previous study has confirmed that B. tabaci MEAM1 and MED contributed to the TYLCV epidemic across China, and that the introduction of MED accelerated the prevalence of TYLCV (Pan et al., 2012). Recently, another devastating tomato virus, Tomato chlorosis virus (Closteroviridae: Crinivirus), has reached an outbreak level in several areas of the Chinese mainland, including Beijing, Tianjin, Shandong, Henan, Jiangsu, Neimenggu, and Guangdong (Zhao et al., 2013; Liu et al., 2014; Gao et al., 2015; Hu et al., 2015; Wu et al., 2016; Zheng et al., 2016; Tang et al., 2017; Wei et al., 2018), resulting in significant crop losses. ToCV is uniquely transmitted in a semi-persistent mode by two distinct whitefly genera, Bemisia and Trialeurodes, viz. B. tabaci, T. abutilonea, and T. vaporariorum (Wisler et al., 1998; Wintermantel and Wisler, 2006). Furthermore, previous research suggests that the rapid spread of ToCV in China was associated with the transmission by MED (Dai et al., 2016; Shi et al., 2018). Notably, the co-infection of TYLCV and ToCV have been detected in several regions of China, including Shandong and Jiangsu Provinces (Zhao et al., 2014; Wu et al., 2016). Following the confirmation of the transmission of TYLCV and ToCV by B. tabaci MED in China, we speculated that the TYLCV&ToCV co-infection in the field was probably due to the spread of B. tabaci MED.
Interactions between plant viruses and insect vectors are important for both the dispersal of the plant viruses and the population dynamics of the insect (Stout et al., 2006). Plant viruses can have direct or indirect impacts on insect vectors. For example, two Begomoviruses [Tabacco curly shoot virus (TbCSV) and Tomato yellow leaf curl China virus (TYLCCNV)] infecting tobacco plants significantly increased the fecundity and longevity of their insect vector B. tabaci MEAM1 (Jiu et al., 2007). In addition, TYLCV infection also benefitted its vector B. tabaci MED by improving the growth, survival, and reproduction (Su et al., 2015). However, ToCV infection decreased the performance of B. tabaci MED on tomato plants as measured by declines in longevity and fecundity (Li et al., 2018).
Next-generation sequencing has been shown to be an efficient means of examining the interaction mechanisms between plant viruses and insects. Many transcriptome studies have been performed to analyze the responses to diverse viral infections in whiteflies. When B. tabaci MEAM1 was infected with TYLCCNV, the immune responses were activated, and detoxification activity and energy costs were simultaneously attenuated (Luan et al., 2011, 2013). Early studies also revealed that a number of genes involved in transport, binding, metabolism, signal transduction, receptors and lysosomes were differentially regulated when B. tabaci MEAM1 fed on TYLCV- and ToCV-infected tomato plants (Kaur et al., 2017; Hasegawa et al., 2018). However, previous viral transcriptome studies were mainly performed on B. tabaci MEAM1, and the focus was limited to the response to a single virus infection. Little is known concerning how B. tabaci MED responds to single TYLCV and ToCV infection. Moreover, the molecular response of B. tabaci to the co-infection with TYCLV and ToCV remains unknown.
In this study, we compared the transcriptional responses in B. tabaci MED after feeding on TYLCV- and ToCV-infected, TYLCV&ToCV co-infected and uninfected tomato plants during a 24-h acquisition access period (AAP), respectively. A number of genes involved in defense response, transport, primary metabolism, cell function, and fitness responded to viral infection. We also used the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases to further annotate the functions of the differentially expressed genes (DEGs). Additionally, we compared the similarities and differences between MED whiteflies infected by a single virus (TYLCV or ToCV) and whiteflies co-infected by TYLCV&ToCV. The results provide a comprehensive view of the molecular response to diverse forms of viral infection in the MED whitefly and yield new insights into the interactions between insect vectors and multiple viruses. To our knowledge, this is the first study to analyze the transcriptional changes in response to viral co-infection in whitefly vectors.
Materials and Methods
Insect Culture and Virus Source
A colony of B. tabaci MED originating in 2012 from Ji'nan, Shandong, China has been maintained on cotton plants (Gossypium hirsutum L. cv. Lu-Mian 21), a non-host for TYLCV and ToCV for 6 years. The TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected tomato plants (Solanum lycopersicum M) were collected from Qingdao, Shandong, China in 2014, and the viruses were maintained using tomato plants (S. lycopersicum M. cv. Zhongza 9) via whitefly transmission as described previously (Li et al., 2018). Both the whiteflies and the plants were cultured in separate climate chambers at 27°C, 60% RH, and a 16:8 (L: D) of photoperiod. The B. tabaci MED population was confirmed using the Vsp I-based mtCOI-RFLP method (Chu et al., 2012).
Virus Acquisition and Sample Collection
To obtain the viruliferous and non-viruliferous whiteflies, 2,400 female adults were collected and transferred onto TYLCV-infected, ToCV-infected, TYLCV&ToCV co-infected, and uninfected tomato plants for 24 h. Approximately 200 live whiteflies were collected from virus-infected or uninfected tomato plants following a 24-h AAP for each of the two biological replications. The whiteflies were frozen in liquid nitrogen and stored at −80°C. To confirm virus acquisition and determine the efficiency of acquisition for each virus, DNA was extracted from 15 individuals fed on TYLCV-infected and TYLCV&ToCV co-infected tomato plants using a TIANamp Micro DNA Kit (TIANGEN, China). Total RNA was extracted from 15 individuals fed on ToCV-infected and TYLCV&ToCV co-infected tomato plants using TRIzol Reagent (Thermo Fisher, USA). The first-strand cDNA was synthesized following the procedures for the PrimerScript RT Reagent Kit (Perfect real-time) (TaKaRa, Japan). PCR analyses for the detection of TYLCV and ToCV were conducted using Golden Star T6 Super PCR Mix (TsingKe, China) with the primers TYLCV-F/R (Li et al., 2012) and ToCV-F/R (Dovas et al., 2002), respectively (Supplementary Table S1). The individuals fed on the uninfected tomato plants served as the negative control.
cDNA Library Preparation and Sequencing
Total RNA was extracted separately from viruliferous and non-viruliferous whiteflies using TRIzol Reagent (Thermo Fisher, USA) according to the manufacturer's protocol. RNA quality and concentration were verified using 1% agarose gels, a NanoPhotometer® spectrophotometer (IMPLEN, USA), and the Qubit RNA Assay Kit in a Qubit® 2.0 Fluorometer (Life Technologies, USA). The integrity of total RNA was determined using the RNA Nano 6000 Assay Kit of the Bioanalyzer 2100 system (Agilent Technologies, USA).
The cDNA libraries were generated from 1.5 μg RNA of each sample using NEBNext® UltraTM RNA Library Prep Kit for Illumina (NEB, USA) following the manufacturer's instructions, and the index codes were added to attribute sequences to each sample. The quality of the libraries was evaluated using the Agilent Bioanalyzer 2100 system. The cDNA libraries were sequenced for 125/150 bp paired-end reads on an Illumina HiSeq Xten platform (Novogene Bioinformatics Technology Co. Ltd, China).
Transcriptome Assembly and Differential Expression Analysis
In order to obtain clean data, the raw reads were cleaned by removing any reads containing adapter, reads containing poly-N, and low quality reads. The genomes of B. tabaci MED (Xie et al., 2017) and MEAM1 (Chen et al., 2016) were used for alignment of the clean reads. Indices of the reference genomes were built using Bowtie v2.2.3 (Langmead and Salzberg, 2012), and the high-quality paired-ended clean reads were aligned to the reference genome using HISAT 2.0.4 (Kim et al., 2015). The reads numbers mapped to each gene were counted using HTSeq v0.6.1. The FPKM (expected number of Fragments Per Kilobase of transcript sequence per Million base pairs sequenced) of each gene was calculated based on the length of the gene and the reads count mapped to that gene. Differential expression analysis of viruliferous and non-viruliferous whiteflies was performed using the DESeq2 R package (1.0) (Love et al., 2014). The resulting P-values were adjusted using Benjamini and Hochberg's approach for controlling the false discovery rate (FDR) (Benjamini and Hochberg, 1995). Genes with an adjusted P-value (q-value) < 0.05 found by DESeq were assigned as differentially expressed. The identification of genes related to viruses infection was conducted based on the gene annotation and differential expression analysis.
GO and KEGG Pathway Analysis
GO enrichment analysis of the differential expression of genes across the samples was carried out using the GOseq R package (Young et al., 2010). Additionally, the statistical enrichment of the differential expression genes in KEGG pathways was implemented by the KOBAS software.
RT-qPCR Validation
To validate the DEG analysis results, the expression profiles of 11 genes were measured by RT-qPCR with SDHA as the internal control gene (Li et al., 2013). Primers for RT-qPCR were designed using the Primer 3.0 software (http://bioinfo.ut.ee/primer3-0.4.0/) and are listed in Supplementary Table S1. All of the whitefly samples for the RT-qPCR validation were prepared according to the method described above. Each treatment contained three biological replications. The same total RNA extraction and cDNA synthase methods were used as described above for the detection of ToCV in whiteflies. RT-qPCR assays were performed in 20 μL using the SYBR Premix Ex TaqTM II (Perfect Real Time) (TaKaRa, Dalian, China) according to the manufacturer's instructions. The reactions were conducted using a qTower 2.2 real-time PCR Thermal Cycler (Analytikjena, Germany) under the following conditions: 95°C for 2 min; 40 cycles of 95°C for 15 s and 60°C for 30 s; melting curve generation (60°C to 95°C). The relative expression ratios were calculated using the 2−ΔΔCT method (Livak and Schmittgen, 2001).
Results
Virus Infection Status of Whiteflies Selected for RNA-Seq
In order to make sure the basis of comparison between virus-infected and non-viruliferous colonies of B. tabaci MED, the infection rates were estimated for the whiteflies feeding on virus-infected tomato plants for 24 h. Both the infection rates of TYLCV and ToCV reached 100% in the whiteflies after a 24-h AAP feeding on TYLCV and ToCV singly infected tomato plants (Supplementary Figure S1). However, the lower infection rates of TYLCV (93.33%) and ToCV (80%) were observed in the whiteflies feeding on TYLCV&ToCV co-infection plants (Supplementary Figure S1).
Overview of Illumina Sequencing and Transcriptome Assembly
To determine the transcriptome profiles of B. tabaci MED in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection, we performed RNA-seq analysis of female adults that had fed on the respective tomato plants for 24 h Eight cDNA libraries of viruliferous and non-viruliferous whiteflies were sequenced, generating 30,475,776 and 43,101,948 raw reads, respectively (Table 1). After cleaning and quality checks, 29,813,838 to 41,978,676 clean reads were obtained and mapped to the whitefly (B. tabaci MED) reference genome, for the mapping rates of 80.85 to 82.36% (Table 1). Additionally, Pearson's correlation analysis indicated that the two replicated libraries of each treatment were highly correlated (Pearson's r = 0.958–0.968) (Supplementary Figure S2).
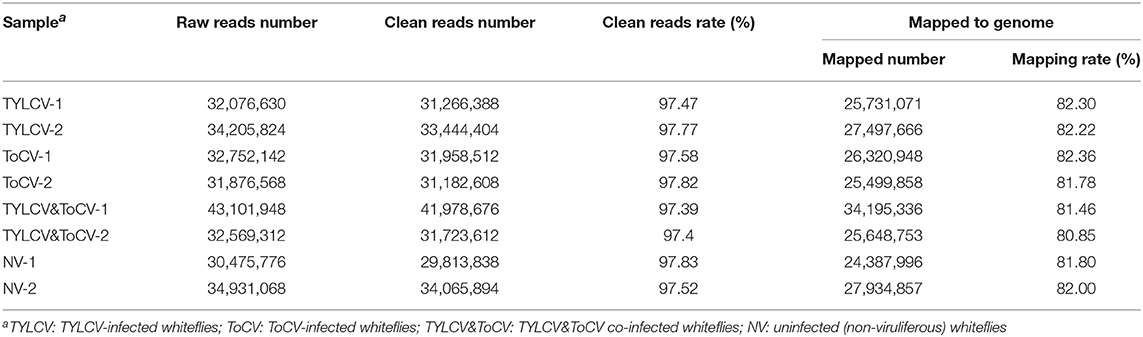
Table 1. Summary statistics of RNA-Seq libraries from Bemisia tabaci MED fed for 24 h on virus-infected or uninfected tomato plants.
Global Patterns of Gene Expression in Response to Different Patterns of Viral Infection
A total of 359 genes were differentially expressed in whiteflies feeding on TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected tomato plants compared to whiteflies that fed on uninfected tomato plants (Figure 1A). In TYLCV and ToCV infected whiteflies, 78 (43 upregulated, 35 downregulated) and 221 (88 upregulated, 133 downregulated) DEGs were detected, while only 60 (38 upregulated, 22 downregulated) genes were differentially expressed due to TYLCV&ToCV co-infection (Figure 1A).
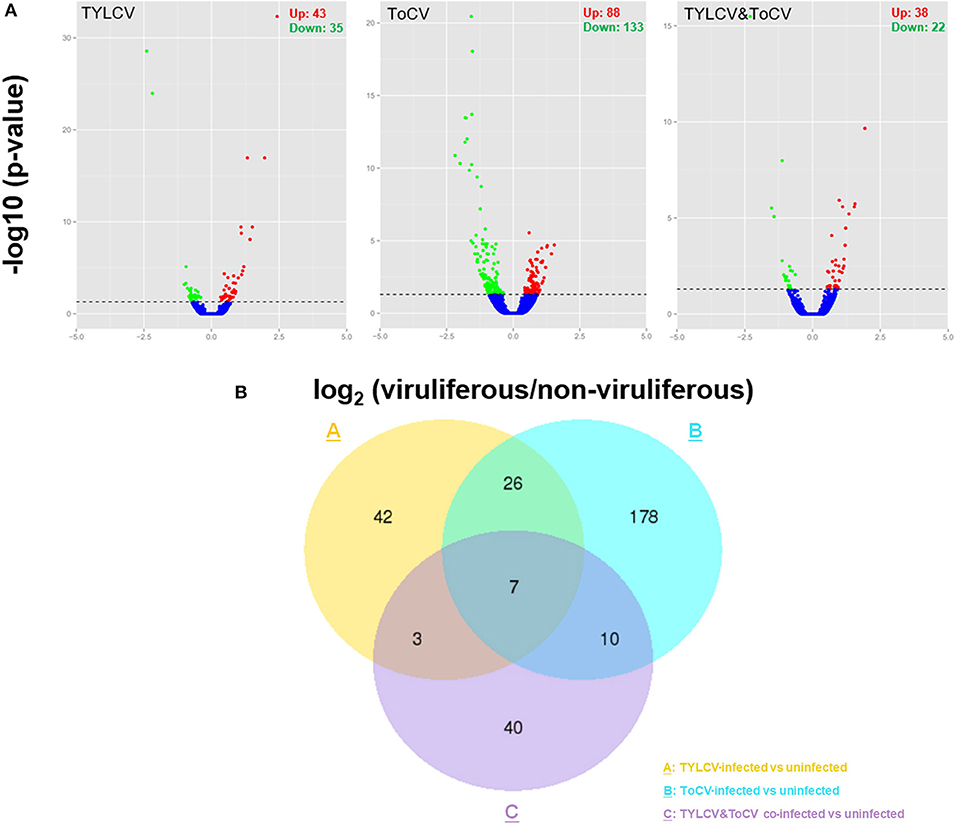
Figure 1. Differentially expressed genes (DEGs) in Bemisia tabaci MED in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection. (A) Volcano plots of differentially expressed genes in whiteflies infected with TYLCV, ToCV, and TYLCV&ToCV compared to non-viruliferous whiteflies. Dots above the horizontal dotted line indicate the DEGs with an adjusted P-value < 0.05. Red dots denote significantly upregulated genes, while green dots denote significantly downregulated genes. (B) Venn diagram depicting unique and common DEGs in whiteflies in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection.
Although most of the DEGs in response to different patterns of viral infection were diverse, there were seven common genes differentially expressed in whiteflies feeding on TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected tomato plants (Figure 1B). Totals of 42.31% and 14.93% of the virus-responsive genes were shared between TYLCV- and ToCV-infected whiteflies. Compared to non-viruliferous whiteflies, only ten DEGs were in common between TYLCV-infected and TYLCV&ToCV co-infected whiteflies. However, greater proportions of common genes altered by ToCV (7.69%) and TYLCV&ToCV (28.33%) were identified (Figure 1B).
Differentially Regulated Genes Associated With Detoxification
Compared to non-viruliferous whiteflies, three differentially regulated cytochrome P450 (P450) genes involved in detoxification were identified in TYLCV-infected whiteflies. One P450 gene was significantly upregulated, while the other two P450 genes were significantly downregulated (Table 2). We found a total of 12 DEGs encoding detoxification enzymes in ToCV-infected whiteflies, including five P450s (one upregulated, four downregulated), a downregulated carboxylesterase (CarE), four downregulated UDP-glucuronosyltransferases (UGTs), and two upregulated ATP-binding cassette transporters (ABCs) (Table 2). Only one P450 gene, BTA015105.1, was identified and downregulated in response to TYLCV&ToCV co-infection (Table 2); the same gene was also detected in TYLCV and ToCV singly infected whiteflies with the same expression pattern.
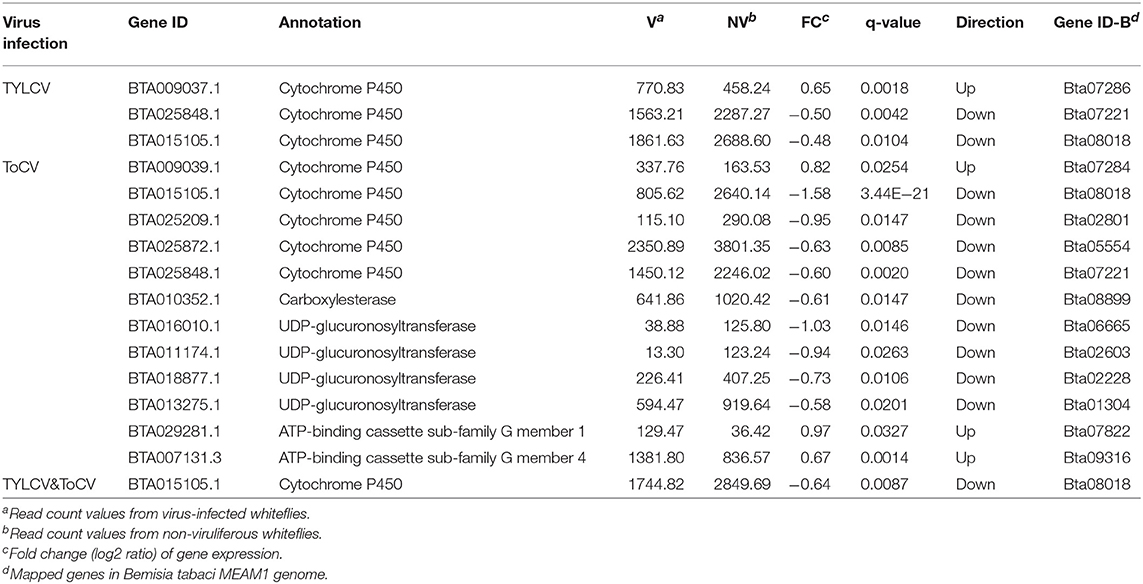
Table 2. Differentially regulated genes associated with detoxification.
Differentially Regulated Genes Associated With Stress and Immune Response
A majority of stress response related genes were upregulated in TYLCV-infected whiteflies; these included two alpha-crystallin B chain-like genes, a heat shock protein 70 (HSP70), and a heat shock protein 90 (HSP90) (Table 3). While the gene BTA014707.1, which encodes superoxide dismutase (SOD), was significantly downregulated (Table 3). In accordance with TYLCV-infected whiteflies, four of five stress responsive genes were upregulated in whiteflies feeding on ToCV-infected tomato plants, including an HSP70, a Gulia lazarillo, a peroxidase, and a glutathione peroxidase (Table 3). The co-infection with TYLCV and ToCV could also significantly depress the transcription level of the SOD gene BTA014707.1 (Table 3).
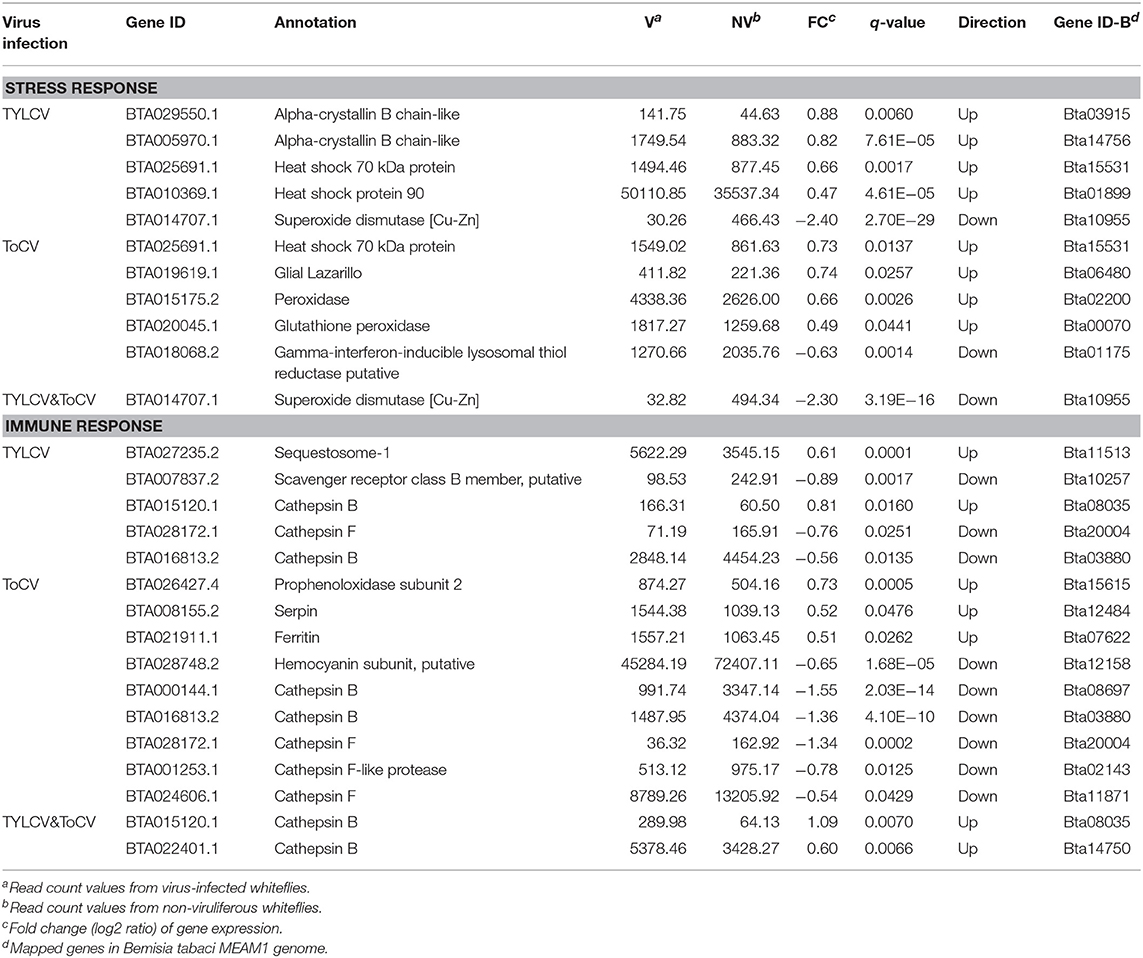
Table 3. Differentially regulated genes associated with stress and immune response.
Five DEGs related to the immune response were identified in TYLCV-infected whiteflies, including one upregulated sequestosome-1, one downregulated scavenger receptor, and three differentially expressed genes encoding the cathepsins (one upregulated and two downregulated) (Table 3). Additionally, nine immune related genes were differentially expressed in whiteflies when infected with ToCV (Table 3). The genes encoding prophenoloxidase subunit 2, serpin, and ferritin were upregulated, while one hemocyanin subunit gene was downregulated (Table 3). Five genes associated with the lysosome, including two cathepsin B genes and three cathepsin F genes, were all downregulated (Table 3). Only two cathepsin B genes associated with the lysosome were classified as being associated with the immune response in TYLCV&ToCV co-infected whiteflies, and both were upregulated (Table 3).
Differentially Regulated Genes Associated With Transport
Four genes associated with transport were all downregulated in TYLCV-infected whiteflies compared with non-viruliferous whiteflies (Table 4). A total of 18 DEGs implicated in transport were identified in ToCV-infected whiteflies, and the expression levels of 14 genes were decreased (Table 4). Among these downregulated genes, five genes (BTA02849.1, BTA029270.2, BTA019411.1, BTA024233.1, and BTA016669.3) were identified as glucose transporters (Table 4). However, among the DEGs in TYLCV&ToCV co-infected whiteflies, four genes related to transport were all upregulated (Table 4).
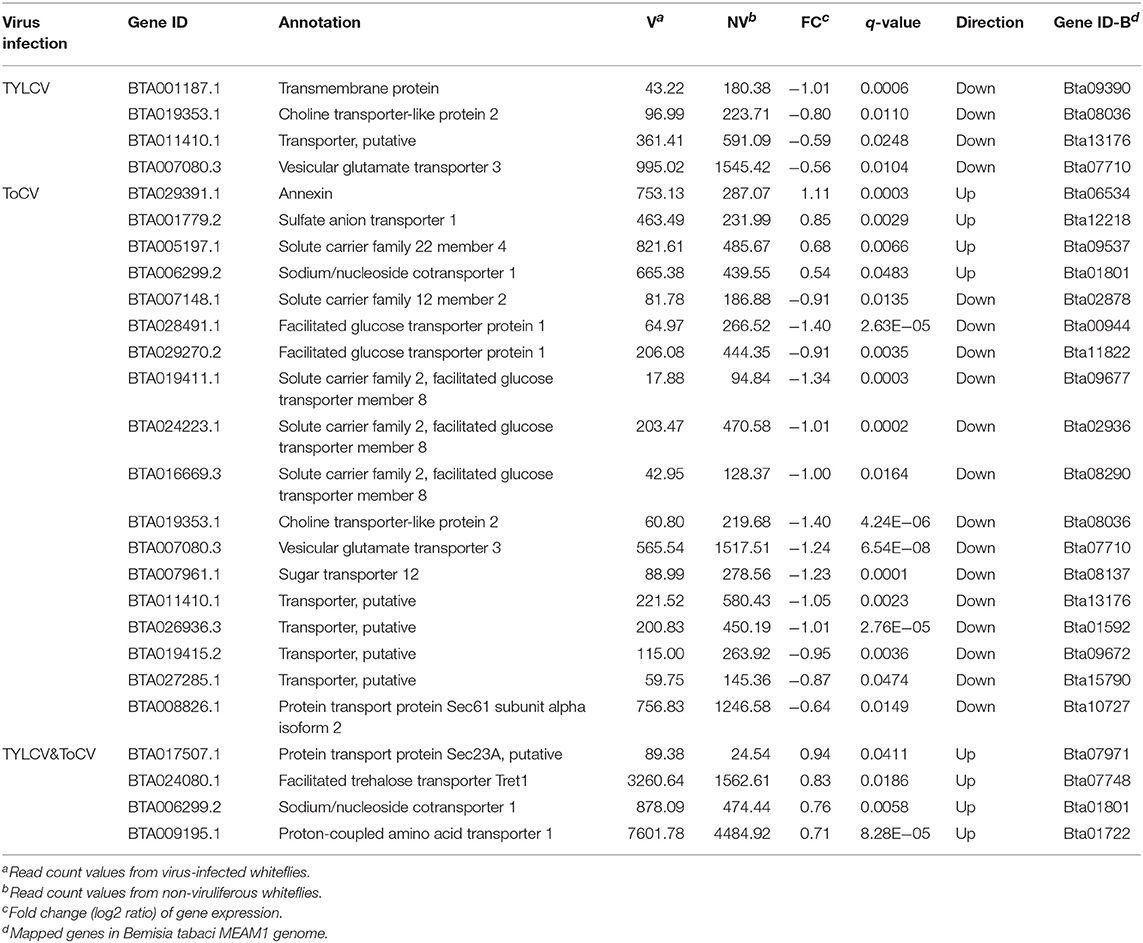
Table 4. Differentially regulated genes associated with transport.
Differentially Regulated Genes Associated With Energy Metabolism, Lipid Metabolism, and Protein Synthesis and Amino Acid Metabolism
After feeding on TYLCV-infected tomato plants, only one gene that was considered to be involved in energy metabolism, annotated as ATP synthase gamma chain, was significantly downregulated in whiteflies (Table 5). Six genes involved in carbohydrate metabolism and three genes involved in ATP metabolism were differentially expressed in ToCV-infected whiteflies (Table 5). Moreover, four genes (BTA020543.1, BTA021845.1, BTA020828.1, and BTA001223.1) associated with carbohydrate metabolism and two genes (BTA029804.1 and BTA020850.1) associated with ATP metabolism appeared among the DEGs in TYLCV&ToCV co-infected whiteflies (Table 5).
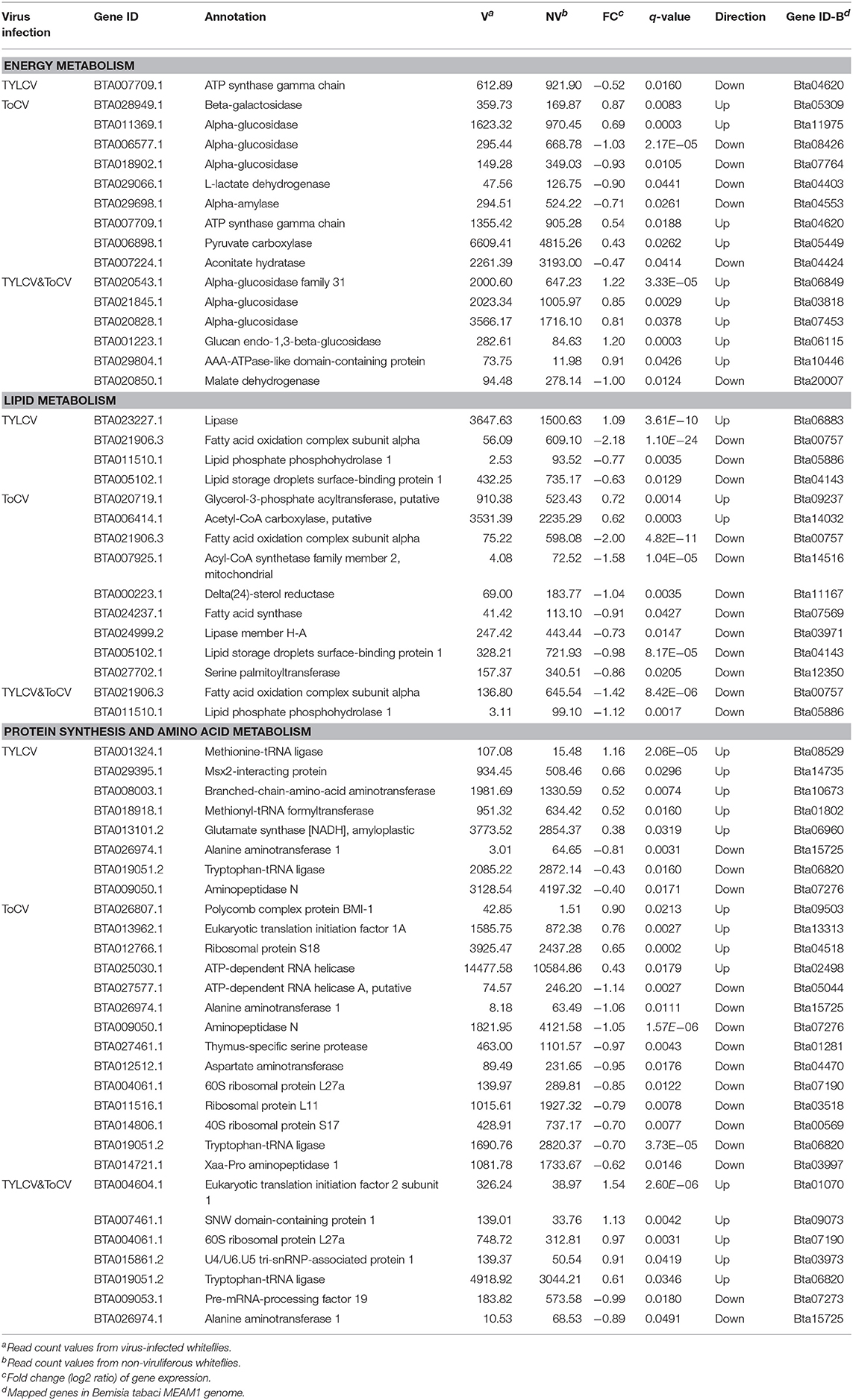
Table 5. Differentially regulated genes associated with energy metabolism, lipid metabolism, and protein synthesis and amino acid metabolism.
We also identified and analyzed four genes associated with lipid metabolism in TYLCV-infected whiteflies, all of which were downregulated except lipase (Table 5). A majority of the lipid metabolism genes were downregulated in ToCV-infected whiteflies; the transcription levels of only two genes encoding glycerol-3-phosphate acyltransferase and acetyl-CoA carboxylase increased (Table 5). Similar with TYLCV-infection and ToCV-infection, two lipid metabolism related genes (fatty acid oxidation complex subunit alpha and lipid phosphate phosphohydrolase 1) were significantly downregulated in TYLCV&ToCV co-infected whiteflies (Table 5).
For protein synthesis and amino acid metabolism, five genes were upregulated and three genes were downregulated in TYLCV-infected whiteflies, while 14 genes (four upregulated and ten downregulated) were responsive to ToCV infection in B. tabaci MED (Table 5). A total of seven genes associated with protein synthesis and amino acid metabolism were identified in TYLCV&ToCV co-infected whiteflies (Table 5). The two genes encoding tryptophan-tRNA ligase and alanine aminotransferase, which were found in the two single virus-infected whiteflies, were also detected in TYLCV&ToCV co-infected whiteflies (Table 5), suggesting common functions in response to viral infection.
Differentially Regulated Genes Associated With Cell Function and Other Functions
Only one gene involved in cell function, which is encoding Alpha-tubulin N-acetyltransferase, was upregulated in response to TYLCV infection (Table 6). Compared to TYLCV infection, more differentially regulated genes (three upregulated, six downregulated) associated with cell function were identified in ToCV-infected whiteflies (Table 6). In addition, two genes involved in cell functions (one upregulated, one downregulated) were altered in TYLCV&ToCV co-infected whiteflies (Table 4), while the gene BTA030040.1 encoding condensing-2 complex subunit D3 was also differentially expressed in ToCV-infected whiteflies.
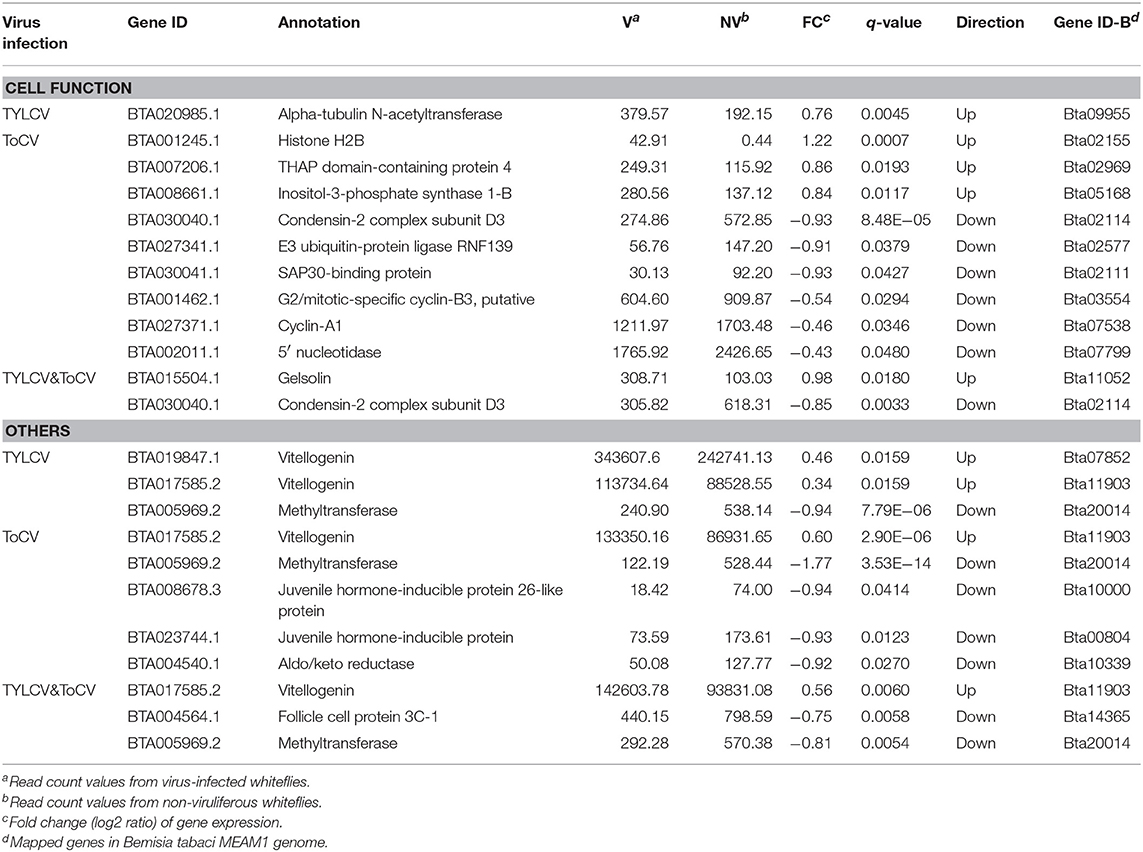
Table 6. Differentially regulated genes associated with cell function and other functions.
In response to TYLCV infection, ToCV infection and TYLCV&ToCV co-infection, the vitellogenin genes were all upregulated in whiteflies (Table 6). We also detected that two genes encoding juvenile hormone-inducible proteins (Table 6), exhibited the downregulated transcription profiles with ToCV infection. The follicle cell protein, which is also associated with reproduction, was identified in TYLCV&ToCV co-infected whiteflies (Table 6). Moreover, our analysis showed both single infection with TYLCV or ToCV and TYLCV & ToCV co-infection could reduce the transcription level of methyltransferase (Table 6).
Differentially Regulated Genes Associated With Unknown Protein
Among the TYLCV- and ToCV-responsive DEGs, seven and ten genes' functions were still unknown, while the comment gene BTA023229.1 showed opposite regulation between these two viruses infection (Supplementary Table S2). Seven unknown proteins were differentially expressed in TYLCV&ToCV co-infected whiteflies, with three upregulated and four downregulated, while the gene BTA027611.2 was similarly downregulated in ToCV-infected whiteflies (Supplementary Table S2).
GO and KEGG Enrichment Analysis of DEGs
GO assignment was performed to classify the functions of the DEGs in response to viral infection. There were 60, 175, and 46 genes among DEGs in response to TYLCV infection, ToCV infection and TYLCV&ToCV co-infection, respectively. These were categorized under biological process, cellular component, and molecular function categories, respectively (Figure 2; Supplementary Table S3). Under the biological process category, metabolic process, and single-organism metabolic process represented the most abundant subcategories in TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected whiteflies. The third most represented GO terms in TYLCV-infected and TYLCV&ToCV co-infected whiteflies were both protein metabolic process, while organonitrogen compound metabolic process was present as the third largest group in ToCV-infected whiteflies. Fewer DEGs were enriched in the cellular component category, and cytoplasm was the largest subcategory among the three types of viruliferous whiteflies. In TYLCV- and ToCV-infected whiteflies, catalytic activity, hydrolase activity, and anion binding were the most abundant molecular function categories. For whiteflies exposed to TYLCV&ToCV, the DEGs were significantly enriched in catalytic activity, hydrolase activity, and peptidase activity under the molecular function category.
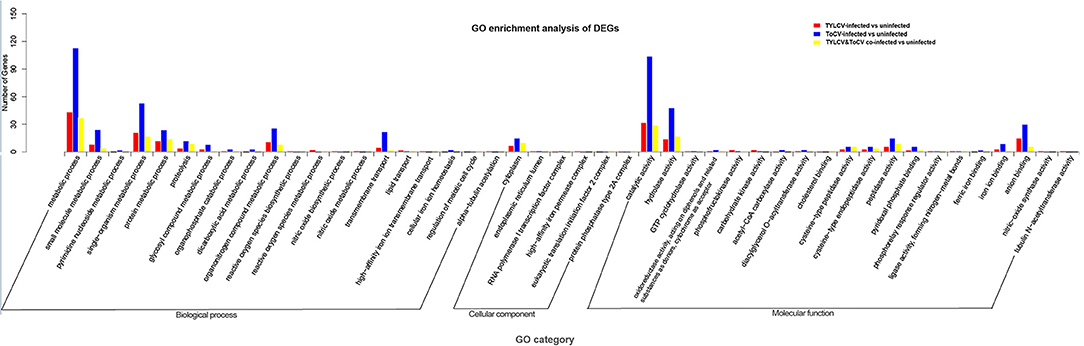
Figure 2. GO enrichment analysis of DEGs in Bemisia tabaci MED in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection. Bars represent the number of DEGs classified into 46 GO terms under three main categories (biological process, cellular component, and molecular function).
Moreover, we found that 57, 184, and 44 genes among TYLCV, ToCV, and TYLCV&ToCV responsive genes, respectively, were mapped to the KEGG database and classified into 11 categories (Figure 3; Supplementary Table S4). The terms of the four most represented pathways in whiteflies fed on TYLCV were: metabolic pathways (10), protein processing in the endoplasmic reticulum (4), lysosome (3), and biosynthesis of amino acids (3). In ToCV-infected whiteflies, the highest number of genes also belonged to metabolic pathways (37), followed by ribosome (8), lysosome (7), and fatty acid metabolism (7). Compared to non-viruliferous whiteflies, the four terms of pathways most represented in TYLCV&ToCV co-infected whiteflies were metabolic pathways (7), lysosome (4), protein processing in the endoplasmic reticulum (4), and spliceosome (3).
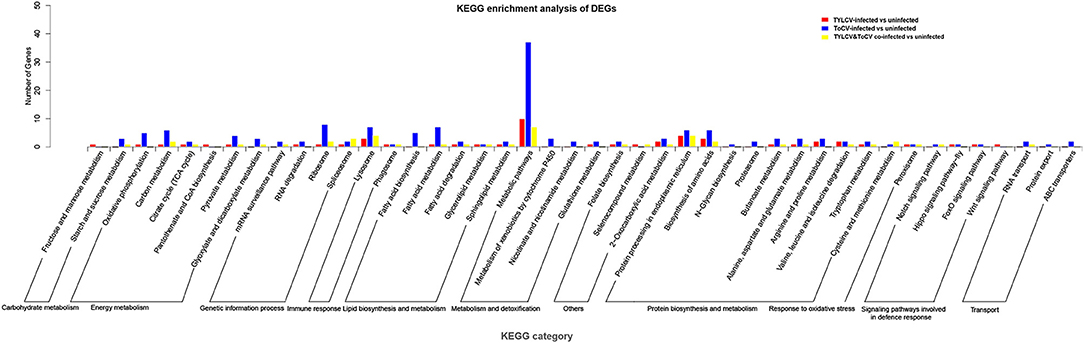
Figure 3. KEGG pathways enrichment analysis of DEGs in Bemisia tabaci MED in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection. Bars represent the number of DEGs classified into 44 KEGG terms under 11 main categories (metabolism and detoxification, response to oxidative stress, transport, signaling pathways involved in defense response, immune response, genetic information process, carbohydrate metabolism, protein biosynthesis and metabolism, lipid biosynthesis and metabolism, energy metabolism, and others).
Validation of DEGs by RT-qPCR
To validate the data from the DGE analyses, RT-qPCR was conducted on 11 randomly selected differentially expressed genes (Figure 4). We analyzed the transcription profiles of nine annotated genes (BTA017585.2, vitellogenin; BTA021906.3, fatty acid oxidation complex subunit alpha; BTA001187.1, transmembrane protein; BTA009039.1, P450; BTA000144.1, cathepsin B; BTA028491.1, facilitated glucose transporter protein 1; BTA005969.2, methyltransferase; BTA009053.1, pre-mRNA-processing factor 19; BTA011510.1, lipid phosphate phosphohydrolase 1), and two genes encoding unknown proteins (BTA000384.1 and BTA004369.1). The results showed that 10 of the selected genes exhibited concordant expression patterns for both RT-qPCR and DGE (Figure 4). However, the expression trend of one gene (BTA000144.1) was inconsistent between RT-qPCR and DGE (Figure 4), which might have been due to the sensitivity of biases existing between the two methods. Nevertheless, the high consistency between the transcription profiles obtained by RT-qPCR and DGE confirmed the reliability of our DGE results.
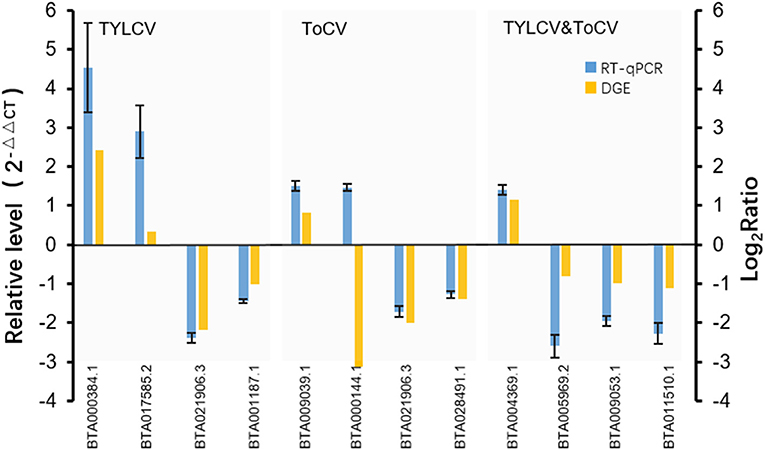
Figure 4. Verification of DEGs by RT-qPCR. Expression levels of 11 selected genes were measured by RT-qPCR using the 2−ΔΔCT method. The 11 selected genes contained nine annotated genes (BTA017585.2, vitellogenin; BTA021906.3, fatty acid oxidation complex subunit alpha; BTA001187.1, transmembrane protein; BTA009039.1, P450; BTA000144.1, cathepsin B; BTA028491.1, facilitated glucose transporter protein 1; BTA005969.2, methyltransferase; BTA009053.1, pre-mRNA-processing factor 19; BTA011510.1, lipid phosphate phosphohydrolase 1), and two genes encoding unknown proteins (BTA000384.1, BTA004369.1). The X axis represents the gene ID numbers of the 11 selected genes; the left Y axis represents the relative gene expression levels measured by RT-qPCR, while the right Y axis represents the log2Ratio of the genes resulting from DGE.
Discussion
Several transcriptome studies on whiteflies responding to TYLCV or ToCV infection have been conducted, and these have provided a large amount of valuable information on the interaction among tomato viruses, whiteflies, and plants (Kaur et al., 2017; Geng et al., 2018; Hasegawa et al., 2018). However, these studies mainly focused on infection by a single virus and were conducted on B. tabaci MEAM1. Thus, in the present study, we performed transcriptome analyses of B. tabaci MED after the whiteflies had fed on TYLCV-infected, ToCV-infected, TYLCV&ToCV co-infected, and uninfected tomatoes for 24 h to compare the gene expression pattern differences between whiteflies infected by a single virus and by both viruses. A total of 265,433,932 clean reads were obtained from eight cDNA libraries, with an average mapping rate of 81.85% to the whitefly (B. tabaci MED) reference genome (Xie et al., 2017). Comparative transcriptome analyses identified 359 DEGs between viruliferous and non-viruliferous whiteflies. The number of DEGs in response to ToCV infection (221 genes) was greater than that in response to TYLCV infection (78 genes). This was in accordance with previous studies with MEAM1, wherein more genes were differentially expressed on acquisition for 24 h with ToCV than TYLCV (Kaur et al., 2017; Hasegawa et al., 2018). Only 60 DEGs were detected in whiteflies feeding on TYLCV&ToCV co-infected tomato plants compared to whiteflies feeding on uninfected tomatoes. The number of TYLCV&ToCV-responsive DEGs was clearly less than those of TYLCV or ToCV, and this may be related to the complex interactions between the two viruses in the insect vectors or in the plants.
Detoxification enzymes of insects, including P450s, CarEs, UGTs, and ABCs, are important in the metabolism of xenobiotics, such as plant allelochemicals and insecticides (Scott et al., 1998; Ferry et al., 2004; Despres et al., 2007; Dermauw and Van Leeuwen, 2014). More genes associated with detoxification were regulated in ToCV-infected whiteflies than in TYLCV-infected and TYLCV&ToCV co-infected whiteflies. P450s were the main detoxification enzymes among the DEGs in virus-infected whiteflies, especially in TYLCV-infected and TYLCV&ToCV co-infected whiteflies. Furthermore, most of P450 genes were downregulated in response to the virus infection (2 of 3 genes for TYLCV infection, 4 of 5 genes for ToCV infection, 1 of 1 gene for TYLCV&ToCV co-infection). Four downregulated UGT genes were detected in ToCV-infected whiteflies, indicating that the glucuronidation of plant toxins might be inhibited by ToCV infection. Moreover, we found that the majority of detoxification enzyme genes were downregulated in the virus-infected whiteflies, which was consistent with a DGE analysis of B. tabaci MEAM1 in response to TYLCCNV infection (Luan et al., 2013). Whitefly susceptibility to insecticides might therefore be altered by virus infection. ABC transporters are mainly involved in the transport of xenobiotics/plant allelochemicals in arthropods (Dermauw and Van Leeuwen, 2014). The ABC gene ABCB FT/P-gp of Drosophila melanogaster is involved in colchicine transport (Wu et al., 1991; Tapadia and Lakhotia, 2005). Additionally, an ABC transporter with similar functions in nicotine excretion was identified in the tobacco hornworm Manduca sexta (Murray et al., 1994; Gaertner et al., 1998; Govind et al., 2010). It is noteworthy that two ABC genes identified in ToCV-infected whiteflies were significantly upregulated. These results suggest that improving the transport efficiency might be a whitefly strategy for coping with the plant toxins induced by viral infection.
The alpha-crystallin B chain-like proteins belonging to the small heat shock proteins (sHSP) as well as the heat shock proteins (HSP70 and HSP90) were significantly upregulated in TYLCV-infected whiteflies. The sHSPs are involved in the destabilizing effects of stressful conditions on cellular integrity (Jong et al., 1993), while HSPs are involved in preventing aggregation of denatured proteins in response to several stress factors, including oxidative, osmotic, and temperature stresses (Lindquist, 1986; Johnston et al., 1998; Sorensen et al., 2003). In addition to the HSP70 gene, we also identified the Glilial Lazarillo (GLaz) gene, a homolog of apolipoprotein D, exhibiting a positive response to the ToCV infection. This protein has been shown to play a protective role in stress situations in D. melanogaster (Sanchez et al., 2006). In contrast with the regulation of detoxification enzyme genes, most of the genes associated with stress response showed increased transcription profiles in response to either TYLCV or ToCV infection.
The innate immune system is the major weapon used by insects to fight against foreign invaders such as pathogens (Hoffmann, 1995; Irving et al., 2001). When infected by plant viruses, the cellular and humoral immune response systems are activated in insects (Luan et al., 2011; Xu et al., 2012; Shrestha et al., 2017). In the present study, higher numbers of immune response-related genes were activated in ToCV-infected whiteflies than in TYLCV-infected or TYLCV&ToCV co-infected whiteflies. Furthermore, a number of cathepsin genes were differentially expressed in whiteflies after feeding on virus-infected tomato plants. The cathepsins have been implicated in virus transmission, apoptosis, and signaling (Kubo et al., 2012; Sim et al., 2012; Saikhedkar et al., 2015). Similar to our findings, Hasegawa et al. (2018) and Kaur et al. (2017) have identified many differentially regulated genes belonging to the cathepsin B and F families in B. tabaci MEAM1 after feeding on TYLCV- and ToCV-infected tomatoes. Only two upregulated cathepsin B genes were found in TYLCV&ToCV co-infected whiteflies, suggesting the possible involvement of immune responses and transmission of these two viruses. Previous studies have shown that the high expression of immune genes in whiteflies with symbionts could lead to a fitness cost (Ghosh et al., 2018). Then, it can be speculated that the inducement of immune response genes may attenuate the performance of TYLCV&ToCV co-infected whiteflies on host plants. Additionally, the altered gene encoding a class B scavenger receptor, a type of surface receptor that is considered to be a regulator of phagocytosis (Franc et al., 1999; Geng et al., 2018), was found in TYLCV-infected whiteflies. Autophagy-related genes have previously been shown to be important in resistance to Begomovirus infection in whiteflies (Luan et al., 2011; Wang et al., 2016). We also identified an upregulated gene encoding sequestosome-1, which is classified as an autophagosome cargo protein, in response to TYLCV infection in whiteflies. After feeding on ToCV-infected tomato plants, the transcription level of the gene encoding the hemocyanin subunit was significantly decreased, in contrast to the transcription profile of the same gene in TYLCV-infected B. tabaci MEAM1 (Hasegawa et al., 2018). Hemocyanins are involved in antiviral functions in arthropods (Dolashka and Voelter, 2013), and these results suggest that the immune response modes of hemocyanins may differ according to diverse viral infections.
Our analysis identified a downregulated choline transporter-like protein gene associated with TYLCV infection. In humans, the choline transporter is a cell membrane transporter that carries choline to cholinergic neurons for acetylcholine synthesis (Okuda and Haga, 2000). The transcription level of the choline transporter-like protein was significantly decreased in ToCV-infected whiteflies, suggesting that both TYLCV and ToCV inhibit the normal transmission of neuron signals in whiteflies. Several members of the annexin group have essential roles in vesicular trafficking, adhesion, apoptosis, and viral infection (Iseki et al., 2009; Patel et al., 2011; Ma et al., 2012). For example, annexin A1 (ANXA1) was confirmed to play a detrimental role in influenza infection and positively regulated virus titers in viral infection experiments using mice (Arora, 2014). A similar role was found for salmon annexin 1 during the infection of infectious pancreatic necrosis virus (IPNV) (Hwang et al., 2007). Thus, the overexpression of the annexin gene in ToCV-infected whiteflies may contribute to the survival and transmission of the virus in vectors. The glucose transporter proteins are related to the interaction with viruses (Huang et al., 2015), and a number of facilitated glucose transporter genes were differentially regulated in ToCV-infected whiteflies B. tabaci MEAM1 (Kaur et al., 2017). In our study, a total of five facilitated glucose transporter genes were detected in ToCV-infected whiteflies, which suggests that these genes may participate in the interaction with ToCV in vivo and may be associated with virus transmission. We noticed that all four transport related genes were upregulated in TYLCV&ToCV co-infected whiteflies, while most of those genes in TYLCV- and ToCV-infected whiteflies were downregulated. The transporter proteins may exhibit positive actions, such as improving the transferring efficiency of trehalose by the upregulated facilitated trehalose transporter Tret 1 gene, in order to defend against the viral co-infection.
Most of the differentially regulated genes associated with energy metabolism encode glucosidases containing alpha-glucosidases and beta-glucosidases. Glucosidases are mainly involved in the hydrolysis of carbohydrates; they also play important roles in normal cellular function and pathogen defense (Bourne and Henrissat, 2001). Two of three alpha-glucosidase genes were downregulated in ToCV-infected whiteflies. Expression of beta-glucosidase genes in Frankliniella occidentalis was also depressed by TSWV infection (Zhang et al., 2013). Here, all four glucosidase genes were all upregulated in response to TYLCV&ToCV co-infection, indicating that the activation of glucosidase genes may be involved in the immune defense to co-infection with the two viruses and the complex interactions between whiteflies and viruses.
Lipid metabolism of insect vectors can be disturbed by plant viruses; this has been demonstrated in several studies and is considered to be a hallmark of cellular changes associated with viral infection (Luan et al., 2011; Xu et al., 2012; Zhang et al., 2013). Most genes in this study associated with lipid metabolism were downregulated in TYLCV-infected (3 of 4 genes) and ToCV-infected (7 of 9 genes) whiteflies, while both of the two lipid metabolism-related genes identified in TYLCV&ToCV co-infected whiteflies were downregulated. This result indicates that both single infection with TYLCV or ToCV and co-infection with TYLCV&ToCV can significantly inhibit the lipid metabolism of B. tabaci MED. Luan et al. (2011) also found that most genes involved in lipid metabolism were downregulated in B. tabaci MEAM1 when feeding on TYLCCNV-infected plants. However, other studies have shown contrasting results in TSWV-infected thrips and demonstrated that lipid metabolism in F. occidentalis and F. fusca was active in response to TSWV infection (Zhang et al., 2013; Shrestha et al., 2017).
Higher numbers of DEGs involved in protein synthesis and amino acid metabolism were identified in ToCV-infected whiteflies than in TYLCV-infected and TYLCV&ToCV co-infected whiteflies, and the majority of those genes (10 of 14 genes) were downregulated in response to ToCV infection. This suggests that protein synthesis and amino acid metabolism in whiteflies can be inhibited by ToCV infection, which is consistent with the results of previous studies on Sogatella furcifera and Campoletis sonorensis (Shelby and Webb, 1997; Xu et al., 2012). Two genes encoding eukaryotic translation initiation factors (eIF), which are involved in the initiation phase of eukaryotic translation, were upregulated in ToCV-infected and TYLCV&ToCV co-infected whiteflies. Wang et al. (2014) revealed that eIF4B of the host could inhibit influenza A virus (IAV) replication by upregulating the expression level of a key protein (interferon-induced transmembrane protein 3, IFITM3) that protects the host from virus infection. Thus, it can be inferred that the upregulation of eIF genes in viruliferous whiteflies may be a strategy in response to ToCV infection and TYLCV&ToCV co-infection. Interestingly, the two genes encoding 60S ribosomal protein L27a and tryptophan-tRNA ligase were downregulated in ToCV-infected whiteflies, but they exhibited opposite regulation profiles in TYLCV&ToCV co-infected whiteflies. The shift in gene expression may result from the alteration of insect and host plant physiologies caused by the co-infection with the two viruses.
Virus infection can lead to cellular DNA damage and the host cells will activate repair mechanisms (Huang et al., 2011). In ToCV-infected whiteflies, the histone H2B gene, which is involved in repair of DNA and regulation of transcription (Ronnigen et al., 2015), was significantly upregulated. This suggests that the histone H2B gene may help attenuate the cell damage wrought by ToCV infection. Additionally, viruses are able to alter the host cell cycle to achieve the replication and expression of their genomes, a phenomenon that has been demonstrated in geminivirus infection (Emmett et al., 2005; Ascencio-lbanez et al., 2008; Geng et al., 2018). Although ToCV cannot replicate in insect vectors, two cyclin genes (G2/motic-specific cyclin-B3 and cyclin-A1), which function in controlling the progression of cells through the cell cycle (Galderisi et al., 2003), were downregulated in ToCV-infected whiteflies. This result demonstrates that ToCV infection also disturbs the normal cell cycle in whiteflies. TYLCV&ToCV co-infection induced the expression of the gelsolin gene, an important actin regulator that is associated with the inhibition of apoptosis (Koya et al., 2000). These alterations of the cell functions caused by viruses may be an adaption or a defense strategy of insect vectors in response to viral infection.
Plant viruses can have direct and indirect effects that influence the fecundity, longevity, and survival rate of the vectors (Belliure et al., 2005; Jiu et al., 2007; Li et al., 2018). Consistent with this, our DGE analysis found several reproduction-related genes and development-related genes differentially regulated in the virus-infected whiteflies. Two vitellogenin genes were upregulated in TYLCV-infected whiteflies, supporting the result that feeding on TYLCV-infected tomato plants can increase the fecundity of B. tabaci MED (Su et al., 2015). Previous study showed that whiteflies MED had a shorter development time on ToCV-infected tomato plants than on healthy tomato plants (Shi et al., 2018). We also found that two juvenile hormone-inducible protein genes were both downregulated in ToCV-infected whiteflies.
GO enrichment analysis showed that the DEGs in TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected whiteflies were all highly enriched in three functional subcategories: the metabolic process, catalytic activity, and single-organism metabolic process. Previous reports demonstrate that plant viruses can change the defense response and nutrition status of the host plants (Shi et al., 2013; Mauck et al., 2014), which might alter the expression patterns of genes involved in metabolic processes and catalytic activity. We also found several differentially regulated genes enriched in the cytoplasm, indicating that those virus-responsive genes were mainly distributed in the cytoplasm among the cellular component. Additionally, there were several DEGs classified as iron ion binding in whiteflies infected with TYLCV (three genes), ToCV (nine genes), and TYLCV&ToCV (one gene) compared with non-viruliferous whiteflies. Iron-binding proteins are vital in iron transport and sequestering iron, but an overabundance can lead to oxidative stress (Strickler-Dinglasan et al., 2006). It is also known that the major iron-binding proteins in insects play important roles in iron transport (Bartfield and Law, 1990) and immunity (Yoshiga et al., 1997, 1999). Therefore, the DEGs belonging to the iron ion binding group may participate in the defense responses to viral infection and oxidative stress.
In accordance with the results of the GO assignment, the KEGG pathway analysis showed that most DEGs in TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected whiteflies were enriched in metabolic pathways. A majority of the genes involved in metabolic pathways were downregulated both in the TYLCV-infected (8 of 10 genes) and TYLCV&ToCV (5 of 7 genes) co-infected whiteflies. A previous study showed that 72.18% of genes enriched in primary metabolism were downregulated in TSWV-infected F. occidentalis (Zhang et al., 2013), which is similar to our results. However, there were 14 upregulated genes enriched in metabolic pathways in ToCV-infected whiteflies. Kaur et al. (2017) also confirmed the upregulation of metabolic pathways in ToCV-infected B. tabaci MEAM1. The lysosome pathway was among the most represented categories in TYLCV-infected, ToCV-infected, and TYLCV&ToCV co-infected whiteflies compared with non-viruliferous whiteflies, indicating an immune response in whiteflies during the early stages of viral infection. As expected, a number of DEGs were also enriched in the lysosome pathway in ToCV-infected B. tabaci MEAM1 compared with non-viruliferous whiteflies (Kaur et al., 2017), and several lysosome genes were also found to be differentially regulated in TYLCV-infected B. tabaci MEAM1 (Hasegawa et al., 2018). Notably, all four genes from the lysosome pathway in TYLCV&ToCV co-infected whiteflies were upregulated, while only one gene was upregulated in both TYLCV- and ToCV-infected whiteflies. We can speculate that co-infection with two viruses may induce a stronger antiviral response than single infection with either TYLCV or ToCV. Additionally, four and three genes upregulated in TYLCV infected and TYLCV&ToCV co-infected whiteflies, respectively, were significantly enriched in the pathway of protein processing in the endoplasmic reticulum, indicating that protein synthesis might be enhanced by TYLCV infection and TYLCV&ToCV co-infection.
Conclusion
We conducted a transcriptome analysis on the whitefly B. tabaci MED in response to TYLCV infection, ToCV infection, and TYLCV&ToCV co-infection. B. tabaci MED genes responsive to viral infection were identified, including genes associated with defense response, transport, primary metabolism, cell function, and total fitness. Shifts in the expression of genes involved in transport and energy metabolism occurred between TYLCV&ToCV co-infection and single infection with TYLCV or ToCV, indicating different responses to diverse patterns of viral infection. GO and KEGG pathway enrichment analyses revealed that the metabolic process and metabolic pathways were significantly disturbed by single infection with TYLCV or ToCV as well as TYLCV&ToCV co-infection. These data increase our understanding of the whitefly-virus interaction and provide new insights into the molecular mechanisms involved in response to co-infection with different viruses. The findings may be useful for discovery of novel molecular targets that could block the spread of whitefly-transmitted viruses and help control insect vector whiteflies.
Data Availability
The RNA-seq reads have been submitted to the SRA at NCBI under the accession PRJNA490883.
Author Contributions
T-BD and DC conceived the study. T-BD conducted the experiments. T-BD, JL, E-HC, and J-ZN analyzed the data. T-BD drafted the manuscript. E-HC and J-ZN revised and enhanced the manuscript. All authors read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of P. R. China (31501707), High-level Talents Funds of Qingdao Agricultural University (6631115033), the Taishan Mountain Scholar Constructive Engineering Foundation of Shandong, and the Agricultural Science and Technology Innovation Project of Shandong Academy of Agricultural Sciences (CXGC2016B11). We also would like to thank LetPub (www.letpub.com) for providing linguistic assistance during the preparation of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2019.00302/full#supplementary-material
References
Alemandri, V., Vaghi Medina, C. G., Dumon, A. D., Arguello Caro, E. B., Mattio, M. F., Garcia Medina, S., et al. (2015). Three members of the Bemisia tabaci (Hemiptera: Aleyrodidae) cryptic species complex occur sympatrically in Argentine horticultural crops. J. Econ. Entomol. 108, 405–413. doi: 10.1093/jee/tov017
Arora, S. (2014). Functional characterization of annexin A1 in toll like receptor 7 signaling and influenza virus infection. Doctor's thesis, National University of Singapore, Singapore.
Ascencio-lbanez, J. T., Sozzani, R., Lee, T. J., Chu, T. M., Wolfinger, R. D., Cella, R., et al. (2008). Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 148, 436–454. doi: 10.1104/pp.108.121038
Bartfield, N. S., and Law, J. H. (1990). Isolation and molecular cloning of transferrin from the tobacco hornworm, Manduca sexta. J. Biol. Chem. 265, 21684–21691.
Belliure, B., Janssen, A., Maris, P. D., and Sabelis, M. (2005). Herbivore arthropods benefit from vectoring plant virus. Ecol. Lett. 8, 70–79. doi: 10.1111/j.1461-0248.2004.00699.x
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
Bourne, Y., and Henrissat, B. (2001). Glycoside hydrolases and glycosyltransferases: families and functional modules. Curr. Opin. Struc. Biol. 11, 593–600. doi: 10.1016/S0959-440X(00)00253-0
Chen, W. B., Hasegawa, D. K., Kaur, N., Kliot, A., Valle Pinheiro, P., Luan, J. B., et al. (2016). The draft genome of whitefly Bemisia tabaci MEAM1, a global crop pest, provides novel insights into virus transmission, host adaptation, and insecticide resistance. BMC Biol. 14:110. doi: 10.1186/s12915-016-0321-y
Chu, D., Hu, X., Gao, X., Zhao, H., Nichols, R. L., and Li, X. (2012). Use of mitochondrial cytochrome oxidase I polymerase chain reaction-restriction fragment length polymorphism for identifying subclades of Bemisia tabaci Mediterranean group. J. Econ. Entomol. 105, 242–251. doi: 10.1603/EC11039
Chu, D., Zhang, Y. J., Brown, J. K., Cong, B., Xu, B. Y., Wu, Q. J., et al. (2006). The introduction of the exotic Q biotype of Bemisia tabaci Mediterranean group. J. Econ. Entomol. 105, 242–251.
Czosnek, H. (2007). Tomato Yellow Leaf curl virus Disease: Management, Molecular Biology, Breeding for Resistance. Dordrecht: Springer.
Dai, H. J., Liu, Y. G., Zhu, X. P., Liu, Y. J., and Zhao, J. (2016). Tomato chlorosis virus (ToCV) transmitted by Bemisia tabaci biotype Q of Shouguang in Shandong Province. J. Plant Protect. 43, 162–167. doi: 10.13802/j.cnki.zwbhxb.2016.01.024
De Barro, P., Liu, S. S., Boykin, L. M., and Dinsdale, A. B. (2011). Bemisia tabaci: a statement of species status. Annu. Rev. Entomol. 56, 1–19. doi: 10.1146/annurev-ento-112408-085504
Dermauw, W., and Van Leeuwen, T. (2014). The ABC gene family in arthropods: comparative genomics and role in insecticide transport and resistance. Insect. Biochem. Molec. 45, 89–110. doi: 10.1016/j.ibmb.2013.11.001
Despres, L., David, J. P., and Gallet, C. (2007). The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 22, 298–307. doi: 10.1016/j.tree.2007.02.010
Dolashka, P., and Voelter, W. (2013). Antiviral activity of hemocyanins. Invert. Surviv. J. 10, 120–127.
Dovas, C. I., Katis, N. I., and Avgelis, A. D. (2002). Multiplex detection of criniviruses associated with epidemics of a yellowing disease of tomato in Greece. Plant Dis. 86, 1345–1349. doi: 10.1094/PDIS.2002.86.12.1345
Emmett, S. R., Dove, B., Mahoney, L., Wurm, T., and Hiscox, J. A. (2005). The cell cycle and virus infection. Methods Mol. Biol. 296, 197–218. doi: 10.1385/1-59259-857-9:197
Ferry, N., Edwards, M. G., Gatehouse, J. A., and Gatehouse, A. M. R. (2004). Plant-insect interactions: molecular approaches to insect resistance. Curr. Opin. Biotech. 15, 155–161. doi: 10.1016/j.copbio.2004.01.008
Franc, N. C., Heitzier, P., and White, K. (1999). Requirement for croquemort in phagocytosis of apoptotic cells in Drosophila. Science 284, 1991–1994. doi: 10.1126/science.284.5422.1991
Gaertner, L. S., Murray, C. L., and Morris, C. E. (1998). Transepithelial transport of nicotine and vinblastine in isolated Malpighian tubules of the tobacco hornworm (Manduca sexta) suggests a P-glycoprotein-like mechanism. J. Exp. Biol. 201, 2637–2645.
Galderisi, U., Jori, F. P., and Giordano, A. (2003). Cell cycle regulation and neural differentiation. Oncogene 22, 5208–5219. doi: 10.1038/sj.onc.1206558
Gao, L. L., Sun, G. Z., Wang, Y., Gao, W., Zhang, C. X., Zhang, A. S., et al. (2015). Molecular detection and identification of Tomato chlorosis virus in Tianjin. Acta Agriculturae Boreali-Sinica 30, 211–215. doi: 10.7668/hbnxb.2015.03.036
Geng, L., Qian, L. X., Shao, R. X., Liu, Y. Q., Liu, S. S., and Wang, X. W. (2018). Transcriptome profiling of whitefly guts in response to Tomato yellow leaf curl virus infection. Virol. J. 15:14. doi: 10.1186/s12985-018-0926-6
Ghosh, S., Bouvaine, S., Richardson, S. C. W., Ghanim, M., and Maruthi, M. N. (2018). Fitness costs associated with infections of secondary endosymbionts in the cassava whitefly species Bemisia tabaci. J. Pest Sci. 91, 17–28. doi: 10.1007/s10340-017-0910-8
Govind, G., Mittapalli, O., Griebel, T., Allmann, S., Bocker, S., and Baldwin, I. T. (2010). Unbiased transcriptional comparisons of generalist and specialist herbivores feeding on progressively defenseless Nicotiana attenuata plants. PLoS ONE 5:e8735. doi: 10.1371/journal.pone.0008735
Hasegawa, D. K., Chen, W. B., Zheng, Y., Kaur, N., Wintermantel, W. M., and Simmons, A. M. (2018). Comparative transcriptome analysis reveals networks of genes activated in the whitefly, Bemisia tabaci when fed on tomato plants infected with Tomato yellow leaf curl virus. Virology 513, 52–64. doi: 10.1016/j.virol.2017.10.008
Hogenhout, S. A., Ammar, El-D, Desouky, A., Whitfield, A. E., and Redinbaugh, M. G. (2008). Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 46, 327–359. doi: 10.1146/annurev.phyto.022508.092135
Hu, J. A., Wan, X. J., Li, Z. J., Huang, W., Li, W. G., and Ying, F. Q. (2015). Molecular identification on Tomato chlorosis virus in Henan Province. China Vegetables 12, 25–28. doi: 10.3969/j.issn.1000-6346.2015.12.007
Huang, H. T., Chan, H. L., Shih, T. Y., and Chen, L. L. (2015). A study of the role of glucose transporter 1 (Glut1) in white spot syndrome virus (WSSV) infection. Fish Shellfish Immun. 46, 305–314. doi: 10.1016/j.fsi.2015.06.034
Huang, N., Wu, W., Yang, K., Passarelli, A. L., Rohrmann, G. F., and Glem, R. J. (2011). Baculovirus infection induce a DNA damage response that is required for efficient viral replication. J. Virol. 85, 12547–12556. doi: 10.1128/JVI.05766-11
Hwang, H. J., Chang, H. M., Han, G. K., Kim, J. Y., Lee, J. M., Park, J. W., et al. (2007). Identification and functional analysis of salmon annexin 1 induced by a virus infection in a fish cell line. J. Virol. 81:13816. doi: 10.1128/JVI.02822-06
Irving, P., Troxler, L., Heuer, T. S., Belvin, M., Kopczynski, C., Reichhart, J. M., et al. (2001). A genome-wide analysis of immune responses in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 98, 15119–15124. doi: 10.1073/pnas.261573998
Iseki, Y., Imoto, A., Okazaki, T., Harigae, H., and Takahashi, S. (2009). Identification of annexin 1 as a PU. 1 target gene in leukemia cells. Leukemia Res. 33, 1658–1663. doi: 10.1016/j.leukres.2009.04.010
Ji, Y. H., Xiong, R. Y., Cheng, Z. B., Zhou, T., Zhao, T. M., Yu, W. G., et al. (2008). Molecular diagnosis of Tomato yellow leaf curl disease in Jiangsu Province. Acta Horticulturae Sinica 35, 1815–1818. doi: 10.3724/SP.J.1005.2008.01083
Jiu, M., Zhou, X. P., Tong, L., Xu, J., Yang, X., Wan, F. H., et al. (2007). Vector-virus mutualism accelerates population increase of an invasive whitefly. PLoS ONE 2:e182. doi: 10.1371/journal.pone.0000182
Johnston, J. A., Ward, C. L., and Kopito, R. R. (1998). Aggresomes: a cellular response to misfolded proteins. J. Cell Biol. 143, 1883–1898. doi: 10.1083/jcb.143.7.1883
Jong, W. W. D., Leunissen, J. A., and Voorter, C. E. (1993). Evolution of the alpha-crystallin/small heat-shock protein family. Mol. Biol. Evol. 10:103.
Kaur, N., Chen, W., Zheng, Y., Hasegawa, D. K., Ling, K. S., Fei, Z., et al. (2017). Transcriptome analysis of the whitefly, Bemisia tabaci MEAM1 during feeding on tomato infected with the crinivirus, Tomato chlorosis virus, identifies a temporal shift in gene expression and differential regulation of novel orphan genes. BMC Genom. 18:370. doi: 10.1186/s12864-017-3751-1
Kim, D., Langmead, B., and Salzberg, S. L. (2015). HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360. doi: 10.1038/nmeth.3317
Koya, R. C., Fujita, H., Shimizu, S., Ohtsu, M., Takimoto, M., Tsujimoto, Y., et al. (2000). Gelsolin inhibits apoptosis by blocking mitochondrial membrane potential loss and cytochrome c release. J. Biol. Chem. 275, 15343–15349. doi: 10.1074/jbc.275.20.15343
Kubo, Y., Hayashi, H., Matsuyama, T., Sato, H., and Yamamoto, N. (2012). Retrovirus entry by endocytosis and cathepsin proteases. Adv. Virol. 2012:640894. doi: 10.1155/2012/640894
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Li, C. B., Cui, Y. L., Zhang, L. Y., and Li, C. Y. (2012). Molecular detection of Tomato yellow leaf curl virus (TYLCV). Hereditas 34, 366–370. doi: 10.3724/SP.J.1005.2012.00366
Li, J., Ding, T. B., Chi, H., and Chu, D. (2018). Effects of Tomao chlorosis virus on the performance of its key vector, Bemisia tabaci, in China. J. Appl. Entomol. 142, 296–304. doi: 10.1111/jen.12477
Li, R. M., Xie, W., Wang, S. L., Wu, Q. J., Yang, N. N., Yang, X., et al. (2013). Reference gene selection for qRT-PCR analysis in the sweetpotato whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 8:e53006. doi: 10.1371/journal.pone.0053006
Lindquist, S. L. (1986). The heat-shock response. Annu. Rev. Biochem. 55, 1151–1191. doi: 10.1146/annurev.bi.55.070186.005443
Liu, Y. G., Wei, J. P., Qiao, N., Li, M. Q., Liu, X. M., and Zhu, X. P. (2014). The occurrence and management of Tomato chlorosis virus in Shandong Province. China Vegetables 5, 67–69. doi: 10.3969/j.issn.1000-6346.2014.05.018
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real time quantitative PCR and the 2(−ΔΔCT) method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq 2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Luan, J. B., Li, J. M., Varela, N., Wang, Y. L., Li, F. F., Bao, Y. Y., et al. (2011). Global analysis of the transcriptional response of whitefly to Tomato yellow leaf curl china virus reveals the relationship of coevolved adaptations. J. Virol. 85, 3330–3340. doi: 10.1128/JVI.02507-10
Luan, J. B., Wang, Y. L., Wang, J., Wang, X. W., and Liu, S. S. (2013). Detoxification activity and energy cost is attenuated in whiteflies feeding on Tomato yellow leaf curl China virus-infected tobacco plants. Insect Mol. Biol. 22, 597–607. doi: 10.1111/imb.12048
Ma, H., Kien, F., Maniere, M., Zhang, Y., and Lararde, N. (2012). Human annexin A6 interacts with influenza A virus protein M2 and negatively modulates infection. J. Virol. 86, 1789–1801. doi: 10.1128/JVI.06003-11
Mauck, K. E., De Moraes, C. M., and Mescher, M. C. (2014). Biochemical and physiological mechanisms underlying effects of Cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant Cell Environ. 37, 1427–1439. doi: 10.1111/pce.12249
Mugiira, R. B., Liu, S. S., and Zhou, X. P. (2008). Tomato yellow leaf curl virus and Tomato leaf curl Taiwan virus invade south-ease coast of China. J. Phytopathol. 156, 217–221. doi: 10.1111/j.1439-0434.2007.01345.x
Murray, C. L., Quaglia, M., Arnason, J. T., and Morris, C. E. (1994). A putative nicotine pump at the metabolic blood-brain-barrier of the tobacco hornworm. J. Neurobiol. 25, 23–34. doi: 10.1002/neu.480250103
Navas-Castillo, J., Fiallo-Olive, E., and Sanchez-Campos, S. (2011). Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 49, 219–248. doi: 10.1146/annurev-phyto-072910-095235
Okuda, T., and Haga, T. (2000). Functional characterization of the human high-affinity choline transporter. FEBS Lett. 484, 92–97. doi: 10.1016/S0014-5793(00)02134-7
Pan, H. P., Chu, D., Ge, D. Q., Wang, S. L., Wu, Q. J., Xie, W., et al. (2011). Further spread of and domination by Bemisia tabaci (Hemiptera: Aleyrodidae) biotype Q on field crops in China. J. Econ. Entomol. 104, 978–985. doi: 10.1603/EC11009
Pan, H. P., Chu, D., Yan, W. Q., Su, Q., Liu, B. M., Wang, S. L., et al. (2012). Rapid spread of Tomato yellow leaf curl virus in China is aided differentially by two invasive whiteflies. PLoS ONE 7:e34817. doi: 10.1371/journal.pone.0034817
Patel, D. M., Ahmad, S. F., Weiss, D. G., Gerke, V., and Kuznetsov, S. A. (2011). Annexin Al is a new functional linker between actin filaments and phagosomes during phagocytosis. J. Cell Sci. 124(Pt 4), 578–588. doi: 10.1242/jcs.076208
Polston, J. E., De Barro, P., and Boykin, L. M. (2014). Transmission specificities of plant viruses with the newly identified species of the Bemisia tabaci species comples. Pest Manag. Sci. 70, 1547–1552. doi: 10.1002/ps.3738
Rao, Q., Luo, C., Zhang, H., Guo, X., and Devine, G. J. (2011). Distribution and dynamics of Bemisia tabaci invasive biotypes in central China. B. Entomol. Res. 101, 81–88. doi: 10.1017/S0007485310000428
Ronnigen, T., Shah, A., Oldenburg, A. R., Vekterud, K., Delbarre, E., Moskaug, J. O., et al. (2015). Prepatterning of differentiation-driven nuclear lamin A/C-associated chromatin domains by GlcNAcylated histone H2B. Genome Res. 25, 1825–1835. doi: 10.1101/gr.193748.115
Saikhedkar, N., Summanwar, A., Joshi, R., and Giri, A. (2015). Cathepsins of lepidopteran insects: aspects and prospects. Insect Biochem. Molec. 64, 51–59. doi: 10.1016/j.ibmb.2015.07.005
Sanchez, D., Lopez-Arias, B., Torroja, L., Canal, I., Wang, X. H., Bastiani, M. J., et al. (2006). Loss of Glial Lazarillo, a homolog of apolipoprotein D, reduces lifespan and stress resistance in Drosophila. Curr. Biol. 16, 680–686. doi: 10.1016/j.cub.2006.03.024
Scott, J. G., Liu, N., and Wen, Z. (1998). Insect cytochromes P450: diversity, insecticide resistance and tolerance to plant toxins. Comp. Biochem. and Phys. C 121, 147–155. doi: 10.1016/S0742-8413(98)10035-X
Shelby, K. S., and Webb, B. A. (1997). Polydnavirus infection inhibits translation of specific growth-associated host proteins. Insect Biochem. Molec. 27, 263–270. doi: 10.1016/S0965-1748(96)00095-1
Shi, X. B., Pan, H. P., Xie, W., Wu, Q. J., Wang, S. L., Liu, Y., et al. (2013). Plant virus differentially alters the plant's defense response to its closely related vectors. PLoS ONE 8:e83520. doi: 10.1371/journal.pone.0083520
Shi, X. B., Tang, X., Zhang, X., Zhang, D. Y., Li, F., Yan, F., et al. (2018). Transmission efficiency, preference and behavior of Bemisia tabaci MEAM1 and MED under the influence of Tomato chlorosis virus. Front. Plant Sci. 8:2271. doi: 10.3389/fpls.2017.02271
Shrestha, A., Champagne, D. E., Culbreath, A. K., Rotenberg, D., Whitfield, A. E., and Srinivasan, R. (2017). Transcriptome changes associated with Tomato spotted wilt virus infection in various life stages of its thrips vector, Fankliniella fusca (Hinds). J. Gen. Virol. 98, 2156–2170. doi: 10.1099/jgv.0.000874
Sim, S., Ramirez, J. L., and Dimopoulos, G. (2012). Dengue virus infection of the Aedes aegypti salivary gland and chemosensory apparatus induces genes that modulated infection and blood-feeding behavior. PLoS Pathog. 8:e1002631. doi: 10.1371/journal.ppat.1002631
Sorensen, J. G., Kristensen, T. N., and Loeschcke, V. (2003). The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 6, 1025–1037. doi: 10.1046/j.1461-0248.2003.00528.x
Stout, M. J., Thaler, J. S., and Thomma, B. P. H. J. (2006). Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Annu. Rev. Entomol. 51, 663–689. doi: 10.1146/annurev.ento.51.110104.151117
Strickler-Dinglasan, P. M., Guz, N., Attardo, G., and Aksoy, S. (2006). Molecular Characterization of iron binding proteins from Glossina morsitans morsitans (Diptera: Glossinidae). Insect Biochem. Molec. 36, 921–933. doi: 10.1016/j.ibmb.2006.09.003
Su, Q., Preisser, E. L., Zhou, X. M., Xie, W., Liu, B. M., Wang, S. L., et al. (2015). Manipulation of host quality and defense by a plant virus improves performance of whitefly vectors. J. Econ. Entomol. 108, 11–19. doi: 10.1093/jee/tou012
Sun, Z. W., Yang, J. X., Zhang, M. Z., and Li, X. D. (2009). Occurrence and control of Tomato yellow leaf curl virus disease in Shandong Province. China Vegetables 21, 5–6.
Tang, Y. F., He, Z. F., She, X. M., and Lan, G. B. (2017). Molecular identification of Tomato chlorosis virus infecting tomato in Guangdong Province. Plant Prot. 43, 133–137. doi: 10.3969/j.issn.0529-1542.2017.02.022
Tapadia, M. G., and Lakhotia, S. C. (2005). Expression of mdr49 and mdr65 multidrug resistance genes in larval tissues of Drosophila melanogaster under normal and stress conditions. Cell Stress Chaperone. 10, 7–11. doi: 10.1379/CSC-67R.1
Valverde, R. A., Sim, J., and Lotrakul, P. (2004). Whitefly transmission of sweet potato viruses. Virus Res. 100, 123–128. doi: 10.1016/j.virusres.2003.12.020
Wang, L. L., Wang, X. R., Wei, X. M., Huang, H., Wu, J. X., Chen, X. X., et al. (2016). The autophagy pathway participates in resistance to tomato yellow leaf curl virus infection in whitefies. Autophagy 12, 1560–1574. doi: 10.1080/15548627.2016.1192749
Wang, S., Chi, X. J., Wei, H. T., Chen, Y. H., Chen, Z. L., Huang, S. L., et al. (2014). Influenza A virus-induced degradation of eukaryotic translation initiation factor 4B contributes to viral replication by suppressing IFITM3 protein expression. J. Virol. 88, 8375–8385. doi: 10.1128/JVI.00126-14
Wei, K. K., Li, J., Ding, T. B., and Chu, D. (2018). Research progress on distribution, identification method of Tomato chlorosis virus (ToCV) and whitefly transmission characteristics. China Vegetables 1, 19–24.
Wintermantel, W. M., and Wisler, G. C. (2006). Vector specificity, host range, and genetic diversity of Tomato chlorosis virus. Plant Dis. 90, 814–819. doi: 10.1094/PD-90-0814
Wisler, G. C., Li, R. H., Liu, H. Y., Lowry, D., and Duffus, J. E. (1998). Tomato chlorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopathology 88, 402–409. doi: 10.1094/PHYTO.1998.88.5.402
Wu, C. T., Budding, M., Griffin, M. S., and Croop, J. M. (1991). Isolation and characterization of Drosophila multidrug resistance gene homologs. Mol. Cell. Biol. 11, 3940–3948. doi: 10.1128/MCB.11.8.3940
Wu, J. B., Dai, F. M., and Zhou, X. P. (2006). First report of Tomato yellow leaf curl virus in China. Plant Dis. 90:1359. doi: 10.1094/PD-90-1359C
Wu, S. H., Li, T. F., Zhao, W. H., Cheng, Z. B., Guo, Q. Y., Zhao, T. M., et al. (2016). Molecular identification on mixed infection of Tomato yellow leaf curl virus and Tomato chlorosis virus on tomato in Jiangsu Province. Acta Horticulturae Sinica 43, 89–99.
Xie, W., Chen, C. H., Yang, Z. Z., Guo, L. T., Yang, X., Wang, D., et al. (2017). Genome sequencing of the sweetpotato whitefly Bemisia tabaci MED/Q. GigaScience 6, 1–7. doi: 10.1093/gigascience/gix018
Xu, Y., Zhou, W. W., Zhou, Y. J., Wu, J. X., and Zhou, X. P. (2012). Transcriptome and comparative gene expression analysis of Sogatella furcifera (Horvath) in response to Southern rice black-streaked dwarf virus. PLoS ONE 7:e36238. doi: 10.1371/journal.pone.0036238
Yoshiga, T., Georgieva, T., Dunkov, B. C., Harizanova, N., Ralchev, K. L., and Law, J. H. (1999). Drosophila melanogaster transferrin. Eur. J. Biochem. 260, 414–420. doi: 10.1046/j.1432-1327.1999.00173.x
Yoshiga, T., Hernandez, V. P., Fallon, A. M., and Law, J. H. (1997). Mosquito transferrin, an acute-phase protein that is up-regulated upon infection. Proc. Natl. Acad. Sci. U.S.A. 94, 12337–12342. doi: 10.1073/pnas.94.23.12337
Young, M. D., Wakefield, M. J., Smyth, G. K., and Oshlack, A. (2010). Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 11:R14. doi: 10.1186/gb-2010-11-2-r14
Zhang, A. H., Zhang, S. M., Liu, S., Di, D. P., and Miao, H. Q. (2010). Occurrence and distribution of Tomato yellow leaf curl disease in Hebei Province. Plant Prot. 36, 127–129. doi: 10.3969/j.issn.0529-1542.2010.04.027
Zhang, Z. J., Zhang, P. J., Li, W. D., Zhang, J. M., Huang, F., Yang, J., et al. (2013). De novo transcriptome sequencing in Frankliniella occidentalis to identify genes involved in plant virus transmission and insecticide resistance. Genomics 101, 296–305. doi: 10.1016/j.ygeno.2013.02.005
Zhao, L. M., Li, G., Liu, Y. G., Guo, J. J., Wei, J. P., and Zhu, X. P. (2014). Molecular identification on mixed infections of Tomato chlorosis virus and Tomato yellow leaf curl virus. China Vegetables 12, 15–20. doi: 10.3969/j.issn.1000-6346.2014.12.006
Zhao, R., Wang, R., Wang, N., Fan, Z. F., Zhou, T., Shi, Y. C., et al. (2013). First report of Tomato chlorosis virus in China. Plant Dis. 97:1123. doi: 10.1094/PDIS-12-12-1163-PDN
Zheng, H. X., Xia, J. X., Zhou, X. M., and Zhang, Y. J. (2016). Be on alert of rapid diffusion of Tomato chlorosis virus transmitted by whitefly in China. China Vegetables 4, 22–26. doi: 10.3969/j.issn.1000-6346.2016.04.009
Keywords: Bemisia tabaci, Tomato yellow leaf curl virus, Tomato chlorosis virus, co-infection, transcriptome
Citation: Ding T-B, Li J, Chen E-H, Niu J-Z and Chu D (2019) Transcriptome Profiling of the Whitefly Bemisia tabaci MED in Response to Single Infection of Tomato yellow leaf curl virus, Tomato chlorosis virus, and Their Co-infection. Front. Physiol. 10:302. doi: 10.3389/fphys.2019.00302
Received: 19 October 2018; Accepted: 07 March 2019;
Published: 03 April 2019.
Edited by:
Patrizia Falabella, University of Basilicata, ItalyReviewed by:
Jing Zhao, Weifang University of Science and Technology, ChinaMurad Ghanim, Agricultural Research Organization (ARO), Israel
Copyright © 2019 Ding, Li, Chen, Niu and Chu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dong Chu, Y2hpbmFjaHVkb25nQHNpbmEuY29t