 Ana Arnaiz
Ana Arnaiz Irene Rosa-Diaz
Irene Rosa-Diaz Maria C. Romero-Puertas
Maria C. Romero-Puertas Luisa M. Sandalio
Luisa M. Sandalio Isabel Diaz
Isabel Diaz- 1Centro de Biotecnologia y Genómica de Plantas, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Universidad Politécnica de Madrid, Madrid, Spain
- 2Department of Biochemistry and Molecular and Cellular Biology of Plants, Estación Experimental del Zaidín, CSIC, Granada, Spain
- 3Departamento de Biotecnología-Biología Vegetal, Escuela Técnica Superior de Ingeniería Agronómica, Alimentaria y de Biosistemas, Universidad Politécnica de Madrid, Madrid, Spain
Reactive nitrogen species (RNS), mainly nitric oxide (NO), are highly reactive molecules with a prominent role in plant response to numerous stresses including herbivores, although the information is still very limited. This perspective article compiles the current progress in determining the NO function, as either a signal molecule, a metabolic intermediate, or a toxic oxidative product, as well as the contribution of molecules associated with NO metabolic pathway in the generation of plant defenses against phytophagous arthropods, in particular to insects and acari.
Introduction
Plants are in constant struggle with a variety of biotic stresses in nature that limit their survival. Among them, phytophagous arthropods are one of the most devastating groups. These herbivores employ specialized feeding modes to obtain nutrients causing leaf defoliation, chlorosis, biomass destruction, growth delay, and even worse consequences under severe infestations leading to an important negative impact in crop yields. Plants have developed sophisticated protection strategies against herbivore combining constitutive and inducible defenses, as the result of their long coexistence during the last 100 million years (Santamaria et al., 2013, 2018a). While constitutive defenses are constantly present, inducible ones are just activated in response to a specific threat, being their nature and mechanism of action directly targeted to the precise feeder and dependent on the plant species and developmental stage. The induction of defenses starts when plasma membrane-specific receptors (pattern recognition receptors, PRRs) recognize conserved herbivore-associated molecular patterns (HAMPs), microbe-associated molecular patterns (MAMPs) derived from herbivore symbionts, or damage-associated molecular patterns (DAMPs) linked to the herbivore injury. The perception of these molecular patterns promotes downstream short-term responses, first at the membrane level (potential depolarization, Ca2+ influxes, etc.), followed by the generation of reactive oxygen and/or nitrogen species (ROS and RNS) as signaling molecules, the activation of kinase cascades, and the synthesis of hormones to finally regulate the expression of defense genes (Fürstenberg-Hägg et al., 2013; Santamaria et al., 2018a). These cues prompt a set of defense events known as pattern-triggered immunity (PTI), by activating signal transduction pathways to synthesize defense metabolites (Jones and Dangl, 2006; Zipfel, 2014; Santamaria et al., 2018a). Alternatively, plant intracellular receptors identify herbivore molecules, elicitors or effectors, that selectively can either trigger or compromise plant immunity altering the defense machinery. This additional response, termed effector-triggered immunity (ETI), is considered an amplified reaction of the PTI (Tsuda and Katagiri, 2010). Early responses take place within minutes to hours after herbivore detection to then induce late-term responses whose products include defensive molecules with toxic, anti-nutritive, deterrent, or repellent properties and volatiles to attract natural enemies of the phytophagous pest (Santamaria et al., 2018a; Stahl et al., 2018; Erb and Reymond, 2019). The whole process is under the regulation of a complex hormonal crosstalk between jasmonic acid (JA), salicylic acid (SA), and ethylene (ET), besides other phytohormones. The known antagonistic relation between SA and JA allows a fine-tune regulation of the defense process (Erb et al., 2012; Schmiesing et al., 2016). Generally, JA-depending pathway is activated by chewing insects, whereas SA regulates responses induced by sucking-feeders (Bari and Jones, 2009; Pieterse et al., 2012), and a balance between JA/SA modulates defenses against sucking mites (Wei et al., 2014; Zhurov et al., 2014; Santamaria et al., 2020a).
Despite all the information available about the plant defense against arthropods, our knowledge on oxidative and particularly on nitrosative signaling is poorly understood. Levels of ROS and RNS, mainly hydrogen peroxide (H2O2) and nitric oxide (NO), increase during insect and acari infestation, and the redox status balance in the cell determines their function since moderate ROS/RNS concentrations differentially sense defense signaling, but an excess of oxidative stress produces chemical oxidation and induces programmed cell death (Foyer and Noctor, 2005; Bittner et al., 2017; Santamaria et al., 2017, 2018b).
No Metabolism
Nitric oxide is clearly recognized as an intra- and intercellular signaling molecule involved in the regulation of a huge range of plant processes ranging from development to resistance and defense responses to biotic and abiotic stresses (Sanchez-Vicente et al., 2019). Two pathways coexist in plants to produce NO, reductive and oxidative ones, involving nitrite and arginine as substrates, respectively (Leon and Costa-Broseta, 2020; Figure 1). Within reductive pathways, NO production arises by both enzymatic and non-enzymatic reactions and is usually dependent on oxygen and NO2– concentrations. Nitrate reductase (NR), a multifunctional cytoplasmic enzyme, whose main function is nitrate assimilation to produce NO2– in a NADPH-dependent way (Campbell, 2001), also shows nitrite reductase (NR) activity, although this represents only 1% of its reductase ability under normal conditions (Yamasaki and Sakihama, 2000; Rockel et al., 2002; Astier et al., 2019). NO production through the action of NR has been demonstrated using different approaches. The mitochondrial electron transport chain (mETC) under anaerobic/hypoxic conditions and the xanthine dehydrogenase–oxidase under anaerobic conditions or phosphate deficiency may also produce NO (Wang et al., 2010; Gupta et al., 2011; Cantu-Medellin and Kelley, 2013). On the other hand, under specific environmental conditions, such as low pH and high concentrations of NO3–, non-enzymatic reduction into NO takes place (Wendehenne et al., 2001; Bethke et al., 2004; Stöhr and Stremlau, 2006; Fancy et al., 2017).
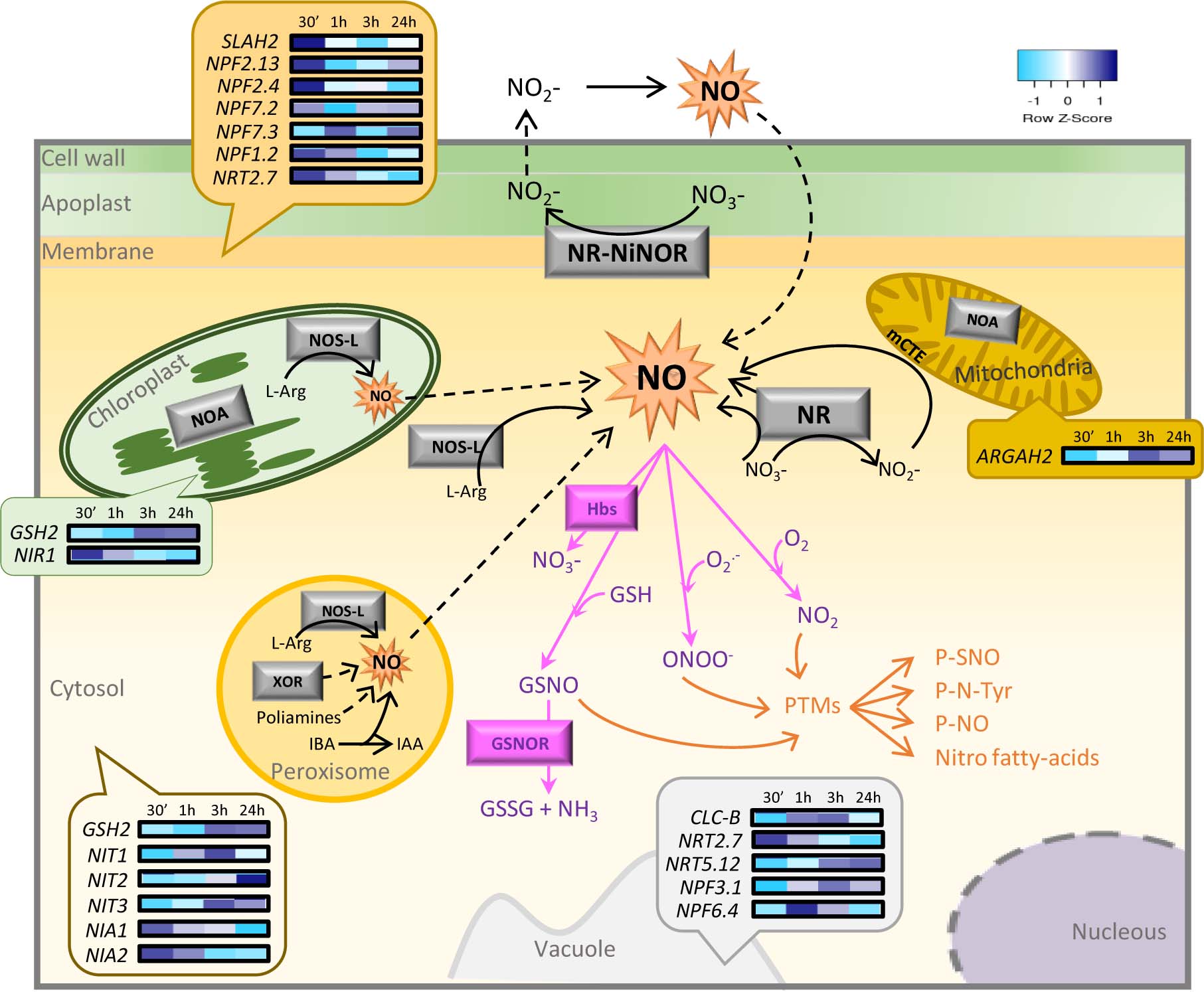
Figure 1. Schematic overview of NO sources and pathways in a plant cell and a heatmap of NO-associated genes expressed in the subcellular locations of A. thaliana after spider mite feeding. The diagram shows the main sources and pathways of NO (black arrows) including both oxidative and reductive pathways, the main scavengers (pink arrows) including superoxide ion, GSH, and hemoglobins, and the main NO mechanisms of action (orange arrows). Discontinued lines represent the mechanisms not experimentally demonstrated. A heatmap showing transcriptomic data of NO-associated genes from A. thaliana at different infestation times (30 min, 1, 3, and 24 h) with T. urticae is comprised within bubbles, positioned over the subcellular compartment where genes are expressed according to SUBA predictions, with a score ≥0.5. IAA, indole-3-acetic acid; IBA, indole-3-butyric acid; GSH, glutathione; GSNO, S-nitrosoglutathione; GSNOR, S-nitrosoglutathione reductase; Hbs, hemoglobins; L-Arg, L-arginine; mETC, mitochondrial electron transport chain; NR, nitrate reductase; NO, nitric oxide; NOS-L, nitric oxide synthase-like; NOA, NO-associated protein; P-NO, nitrosylated protein; P-N-Tyr, nitrated protein; P-SNO, S-nitrosylated protein; PTMs, post-translational modifications; XOR, xanthine oxidoreductase.
The oxidative pathway involves the activity of specialized enzymes as the nitric oxide synthases (NOSs), which oxidize L-arginine to form L-citrulline and NO, and they are well characterized in mammals (Alderton et al., 2001). However, controversial results about this activity have been shown in plants. Bioinformatics approaches have shown no NOS gene/protein in higher plants (Jeandroz et al., 2016; Hancock, 2019), excluding some algae (Foresi et al., 2015), and the typical mammalian NO–cGMP signaling pathway has been also questioned (widely reviewed in Astier et al., 2019). Nevertheless, NOS-like activity has been extensively described in plants by the use of NOS inhibitors and even by heterologous expression of mammalian NOS (Zeidler et al., 2004; Ali et al., 2007; Astier et al., 2018), and the denomination “NOS-like” is adopted for this activity.
Once synthesized, NO is highly reactive, and there are three main types of molecules that react with NO: ROS, glutathione (GSH), and metals (Romero-Puertas and Sandalio, 2016). NO rapidly reacts when present, with the radical superoxide (O2–) generating peroxynitrite (ONOO–), which is one of the most potent oxidant molecules in the cell leading to lipid peroxidation, protein nitration (Ischiropoulos and al-Mehdi, 1995; Radi, 2004), oxygenated forms of cysteine (Cys) residues (sulfenic, sulfinic, and sulfonic acids), and S-glutathionylation (Martínez-Ruiz et al., 2013). ONOO– has been shown to be produced under different stress conditions in plants (Romero-Puertas et al., 2007; Arasimowicz-Jelonek and Floryszak-Wieczorek, 2019). NO can also react with lipid peroxyl radical (LOO⋅) to produce nitro-fatty acids that are related to plant development and plant response to abiotic stress (Rubbo, 2013; Mata-Perez et al., 2017). Besides, the reaction of NO with GSH produces nitrosoglutathione (GSNO), which is considered an endogenous NO reservoir (Noctor et al., 2012) and acts as an S-nitrosylating agent. GSNO is metabolized by GSNO reductase (GSNOR) to transform GSNO into glutathione disulfide (GSSG) and ammonia. Thus, GSNOR controls intracellular levels of GSNO and NO and, therefore, plant responses under different conditions (Liu et al., 2001; Yun et al., 2016). On the other hand, globins are proteins able to metabolize NO producing NO3– (Perazzolli et al., 2004; Becana et al., 2020), and consequently, these proteins can control NO levels by detoxification or through post-translational modification (PTM) reactions (Perazzolli et al., 2006; Figure 1).
No Mechanism of Action: Crosstalk With ROS and H2S
Nitric oxide reactivity leads to its main mechanism of action being PTM of proteins, which are carried out by a series of RNS produced by the reaction of NO with other free radicals as described before. PTMs best studied in plants are: (i) S-nitrosylation/S-nitrosation, referred to the formation of a nitrosothiol group in cysteines, with more than thousand targets described in plants, although a small number have been characterized (Sanchez-Vicente et al., 2019; Sandalio et al., 2019); (ii) nitration, being mainly studied the addition of a nitro group to Tyr side chain, with more than hundred targets described and only a dozen characterized (Rubbo and Radi, 2008; Sanchez-Vicente et al., 2019), and (iii) nitrosylation of transition metals, with the formation of complex bonds to heme groups (Martinez-Ruiz and Lamas, 2009), scarcely studied in plants. NO-dependent PTMs result in the induction of different physiological responses and/or signaling processes as alteration of gene expression, metabolic changes, and phytohormone signaling. Furthermore, NO may regulate other signaling pathways, such as phosphorylation, oxidation, and ubiquitinylation (Cui et al., 2018; Leon and Costa-Broseta, 2020; Lindermayr et al., 2020). Therefore, the ability to regulate virtually all processes in the plant makes NO a do it all molecule (Delledonne, 2005).
Post-translational modification regulation of proteins is quite complex, however, due to the synergistic and antagonistic interplays between the different PTMs (Sandalio et al., 2019). Overlapping of different PTMs on the same protein is very often and follows common pattern in different species, which demonstrate the importance of multilevel PTM regulation in cell metabolism (Duan and Walther, 2015). NO crosstalk with other signaling molecules, such as the well-known ROS and the lesser-known sulfide (H2S), leads to an interplay between redox-dependent PTMs being targets the sulfur-containing amino acids, such as cysteine. Thus, the first step in Cys oxidation is S-nitrosylation while the main ROS involved in signaling, H2O2, leads Cys to the following steps, its reversible oxidation to sulfenic acid (–SOH; sulfenylation) and sulfinic acid (–SO2H; sulfinylation). Excessive ROS accumulation gives rise to the irreversible sulfonic acid (–SO3H; sulfonylation) derivative (Young et al., 2019). S-nitrosylation, sulfenylation, sulfinylation, and intra- and intermolecular disulfide bond formations are rapid and reversible mechanisms to regulate protein function, stability, and location of proteins (Sandalio et al., 2019; Young et al., 2019). Due to their transient nature, these sulfur modifications, which can be reversibly reduced by thioredoxin and glutaredoxin pathways, are regarded as redox switches, giving rise to rapid finely tuned regulation of metabolic pathways and signaling processes (Sandalio et al., 2019; Young et al., 2019). H2S, involved in regulating various processes essential for plant survival, has been demonstrated recently to be a signaling molecule in the same degree of NO and H2O2 in plant systems (Gotor et al., 2019; Hancock, 2019). The mechanism of action of H2S is related with its high affinity for metals from metalloproteins, but it also can oxidize Cys thiol groups to persulfide groups (R-S-SH) promoting covalent PTMs termed persulfidation, which could play a protective role for thiols against oxidative damage (Gotor et al., 2019). Interestingly, RNS and ROS levels are regulated by the interplay between ROS-, H2 S-, and NO-dependent PTMs. Curiously, S-nitrosylation prevents ROS-dependent oxidative damage to several proteins involved in the Calvin–Benson cycle, probably by inducing conformational changes in specific proteins (Tanou et al., 2012). Crosstalk between NO and H2S has been reported in acclimation processes in citrus plants (Molassiotis et al., 2016). On the other hand, antagonistic interplay between protein Tyr nitration and phosphorylation competing for the same Tyr sites has been reported, interfering with different cellular processes, such as cell signaling via MAP kinase cascades (Arasimowicz-Jelonek and Floryszak-Wieczorek, 2019). Although several proteins have been shown as targets of NO-dependent PTMs under different stress conditions, in particular, plant–herbivore interaction is a field that needs to be better explored.
No in Plant–Herbivore Interactions
Some publications have described the rapid accumulation and participation of NO as a common feature to insect-infested plants (Table 1). Different arthropods including hemipteran (Smith and Boyko, 2007; Moloi and van der Westhuizen, 2009; Liu et al., 2011; Mai et al., 2014; Wozniak et al., 2017; Li et al., 2019; Xu et al., 2020) and lepidopteran species (Arimura et al., 2008; Bricchi et al., 2010) cause a rapid and transient increase of NO levels in insect-damaged tissues. However, its physiological significance remains to be established. NO has not been linked to Vm depolarization as H2O2 has, but it has been related to Ca2+ homeostasis and cGMP signaling (Wu and Baldwin, 2009; Misra et al., 2011). Thus, it could exert its biological function through the mobilization of secondary messengers or by the modulation of protein kinase activity. NO interacts with ROS and phytohormones (Mur et al., 2013) and, in consequence, may indirectly act as regulator of the gene expression. In addition, the PTM of proteins mediated by NO, described above, may have potential regulatory effects in plant defense against herbivores as it does toward plant pathogens (Mur et al., 2006; Martinez-Medina et al., 2019).

Table 1. Participation of NO and NO-related enzymes in the plant defenses against phytophagous insects.
In seedling leaves of pea (Pisum sativum), Mai et al. (2014) described the convergence of NO and H2O2 accumulation with the induction of JA, ET, and SA, hormones that sequentially appeared within the first 24–96 h after the aphid Acyrthosiphon pisum feeding. The simultaneous generation of hormones and free radicals at the same time points suggested a synergistic defense action in pea plants to aphid infestation. Moreover, the application of exogenous NO donors (NO, GSNO, and SNP, sodium nitroprusside) to pea plants infested with A. pisum revealed the induction of defense reactions leading to a deterrent result on the pea aphid feeding and the reduction in its population growth (Wozniak et al., 2017). A side effect of SNP treatment is the release of cyanide, a potent respiratory poison with a deterrent effect on phytophagous arthropods who try to elude it or detoxify (Pentzold et al., 2014; Keisham et al., 2019). Campbell and Vallano (2018) analyzed the effects of atmospheric NO2 leaf uptake on tobacco (Nicotiana tabacum) metabolism and its impact in the tobacco responses to the lepidopteran Manduca sexta. Results showed that the foliar assimilation of NO2 increased the nitrogen-derived defensive metabolites, particularly of some alkaloids, and diminished insect feeding and growth. To avoid this defense mechanism, herbivore modified somehow the plant capacity to absorb the reactive nitrogen, prompting a decrease in foliar nitrogen uptake and limiting the concentration of metabolites in leaves. Moreover, accumulating evidences indicate that an interactive fashion of phytohormones and NO regulates guard cell ABA-signaling and stomatal closure, which restricts the foliar uptake of NO2 (Sun et al., 2019). In turn, only few available reports have demonstrated the function of enzymes and other molecules associated with NO metabolic pathway in the generation of plant defenses to pests. Li et al. (2019) showed that the NO production in rice (Oryza sativa) plants was associated with their responses to Nilaparvata lugens infestation, in both susceptible and resistant cultivars. The rice planthopper feeding induced the activity of the NOS-like enzyme only in the susceptible cultivar, whereas no significant alterations of the NR enzymatic activity were observed, in none of the two rice-infested cultivars. These results suggested the active role of NOS in rice defense mediated by NO. Likewise, Wünsche et al. (2011a) examined the function of the GSNOR enzyme in the plant–herbivore interaction by knocking-down GSNOR in Nicotiana attenuata plants. A decrease in JA and ET levels in the silenced plants was observed concomitant to an elevated susceptibility to M. sexta attack. Accordingly, the GSNOR-silenced tobacco plants showed a significant reduction of the trypsin proteinase inhibitor activity and in the diterpene glycosides content, both considered secondary defensive metabolites dependent on the JA derivatives. Wünsche et al. (2011b) also proved that the N. attenuata NO-associated protein 1 (NOA1) was required for the accumulation of JA and JA-Ile and the generation of defenses against M. sexta. NOA1-silenced tobacco plants compromised the production of most of the carbon-based defensive compounds while the synthesis of nitrogen-rich defense metabolites was not altered. These results were probably due to the role of NOA1 in plant chloroplast functions and in the allocation of carbon resources within phenylpropanoid pathway (Wünsche et al., 2011b). Very recently, Xu et al. (2020) have demonstrated that the hemipteran Bemisia tabaci infestation activated NO signaling in tobacco, leading to suppression of JA-dependent defenses and improving nymph performance. Additionally, they have confirmed the NOA1 involvement in the JA-mediated responses to B. tabaci.
The mechanism by which NO mediates the enhancement of plant defenses against pests is still poorly studied, but a recent publication by Li et al. (2019) has linked a mitogen-activated protein kinase, OsMAPK20-5, to NO production in N. lugens-infested rice plants. The OsMAPK20-5 gene expression was up-regulated by female adult feeding, which presumably could be a response to oviposition. Surprisingly, the levels of NO and ET increased after insect feeding in the OsMAPK20-5-silenced plants and consequently improved rice resistance to brown planthopper and oviposited eggs. According to the authors, OsMAPK20-5 could enable rice plants to control excessive hyperaccumulation of NO and ET and thereby to prevent autotoxicity. Importantly, in field trials, MAPK20-5-silenced rice lines displayed a wide protection not only to the N. lugens but also to the white-backed planthopper Sogatella furcifera. Therefore, NO could mediate defense responses in plants against pests acting as a signal molecule, a metabolic intermediate, or a toxic oxidative product.
Since no information on the NO’s role in the interplay between plant and phytophagous acari was available, we did a search of NO-related genes in the RNA sequencing of Arabidopsis thaliana in response to the spider mite Tetranychus urticae after 30 min, 1, 3, and 24 h of feeding (Supplementary Material; Santamaria et al., 2020b). Nineteen NO-associated genes, mainly encoding nitrate transporters, NRs, and nitrilases, were differentially expressed at different time points of infestation. Nitrate transporters showed different expression patterns based on their subcellular in silico location. Generally, those transporters located at the cytoplasmic membrane were rapidly induced by mite infestation, followed by the ones located at the vacuole. NIA1 and NIA2 genes that encode RNS were highly up-regulated at 30 min after mite feeding but were repressed at 24 h. Glutathione synthetase 2 (GSH2) gene putatively located at the chloroplast and cytosol and arginine amidohydrolase 2 (ARGAH2) gene product located at the mitochondria presented the opposite expression pattern being induced at longer infestation time (Figure 1). These differential expression profiles are according to the consecutive steps of plant defense to mite attack since after mite perception, signaling is first activated at the cell membrane level and then transmitted through the cytosol to the rest of the organelles to finally induce the expression of defensive genes. In addition, the identified genes were classified into five different over-expressed categories based on their Gene Ontology (GO) biological function, all of them related to RNS metabolic processes (Supplementary Table 1). These data suggested their functional significance during T. urticae infestation. Further studies are needed to clarify the NO and NO metabolic pathways in the plant defenses against acari feeders.
In conclusion, the current information on how plant responses are regulated by NO and NO-related molecules constitutes still a set of unknown events to be explored, particularly, in the plant–acari interplay. An advanced understanding of the NO function in plant–herbivore interactions will be a strong tool to enhance crop performance and potentially lead to biotechnological approaches for pest control in agricultural systems.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
ID conceptualized the manuscript content. AA and IR-D performed the experiment. ID, AA, MR-P, and LS wrote different sections of the first draft. All authors contributed to the final version of the manuscript.
Funding
This work was financially supported by the Spanish Ministry of Science, Innovation and Universities (grants BIO2017-83472-R, RED2018-102407-T, RED2018-102397-T, and PGC2018-098372-B-100).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully appreciate comments and suggestions from Dr. M. E. Santamaria from the CBGP-UPM-INIA to enhance the manuscript interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.620086/full#supplementary-material
References
Alderton, W. K., Cooper, C. C., and Knowles, R. G. (2001). Nitric oxide synthases: structure, function and inhibition. Biochem. J. 357, 593–615. doi: 10.1042/0264-6021:3570593
Ali, R., Ma, W., Lemtiri-Chlieh, F., Tsaltas, D., Leng, Q., Von Bodman, S., et al. (2007). Death don’t have no mercy and neither does calcium: Arabidopsis CYCLIC NUCLEOTIDE GATED CHANNEL2 and innate immunity. Plant Cell 19, 1081–1095. doi: 10.1105/tpc.106.045096
Arasimowicz-Jelonek, M., and Floryszak-Wieczorek, J. (2019). A physiological perspective on targets of nitration in NO-based signalling networks in plants. J. Exp. Bot. 70, 4379–4389. doi: 10.1093/jxb/erz300
Arimura, G. I., Kopke, S., Kunert, M., Volpe, V., David, A., Brand, P., et al. (2008). Effects of feeding Spodoptera littoralis on lima bean leaves: IV. Diurnal and nocturnal damage differentially initiate plant volatile emission. Plant Physiol. 146, 965–973. doi: 10.1104/pp.107.111088
Astier, J., Gross, I., and Durner, J. (2018). Nitric oxide production in plants: an update. J. Exp. Bot. 69, 3401–3411. doi: 10.1093/jxb/erx420
Astier, J., Mounier, A., Santolini, J., Jeandroz, S., and Wendehenne, D. (2019). The evolution of nitric oxide signalling diverges between animal and green lineages. J. Exp. Bot. 70, 4355–4364. doi: 10.1093/jxb/erz088
Bari, R., and Jones, J. D. G. (2009). Role of plant hormones in plant defence responses. Plant Mol. Biol. 69, 473–488. doi: 10.1007/s11103-008-9435-0
Becana, M., Yruela, I., Sarath, G., Catalán, P., and Hargrove, M. S. (2020). Plant hemoglobins: a journey from unicellular green algae to vascular plants. New Phytol. 227, 1618–1635. doi: 10.1111/nph.16444
Bethke, P. C., Badger, M. R., and Jones, R. L. (2004). Apoplastic synthesis of nitric oxide by plant tissues. Plant Cell 16, 332–341. doi: 10.1105/tpc.017822
Bittner, N., Trauere-Kizilelma, U., and Hilker, M. (2017). Early plant defence against insect attack: involvement of reactive oxygen species in plant responses to insect egg deposition. Planta 245, 993–1007. doi: 10.1007/s00425-017-2654-3
Bricchi, I., Leitner, M., Foti, M., Mithofer, A., Boland, W., Maffei, M. E. (2010). Robotic mechanical wounding (MecWorm) versus herbivore-induce responses: early signalling and volatile emission in Lima bean (Phaseolus lunatus L.). Planta 232, 719–729. doi: 10.1007/s00425-010-1203-0
Campbell, S. A., and Vallano, D. M. (2018). Plant defences mediate interactions, herbivory and the direct foliar uptake of atmospheric reactive nitrogen. Nat. Comm. 9:4743.
Campbell, W. H. (2001). Structure and function of eukaryotic NAD(P)H:nitrate reductase. Cell. Mol. Life Sci. 58, 194–204. doi: 10.1007/PL00000847
Cantu-Medellin, N., and Kelley, E. E. (2013). Xanthine oxidoreductase-catalyzed reduction of nitrite to nitric oxide: insights regarding where, when and how. Nitric Oxide 1, 19–26. doi: 10.1016/j.niox.2013.02.081
Cui, B., Pan, Q., Clarke, D., Villarreal, M. O., Umbreen, S., Yuan, B., et al. (2018). S-nitrosylation of the zinc finger protein SRG1 regulates plant immunity. Nat. Commun. 9:4226. doi: 10.1038/s41467-018-06578-3
Delledonne, M. (2005). NO news is good news for plants. Curr. Opin. Plant Biol. 8, 390–396. doi: 10.1016/j.pbi.2005.05.002
Duan, G., and Walther, D. (2015). The roles of post-translational modifications in the context of protein interaction networks. PLoS Comput. Biol. 11:e1004049. doi: 10.1371/journal.pcbi.1004049
Erb, M., Meldau, S., and Howe, G. A. (2012). Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 17, 250–259. doi: 10.1016/j.tplants.2012.01.003
Erb, M., and Reymond, P. (2019). Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 29, 527–557. doi: 10.1146/annurev-arplant-050718-095910
Fancy, N. N., Bahlmann, A. K., and Loake, G. J. (2017). Nitric oxide function in plant abiotic stress. Plant Cell Environ. 40, 462–472. doi: 10.1111/pce.12707
Foresi, N., Mayta, M. L., Lodeyro, A. F., Scuffi, D., Correa-Aragunde, N., Garcia-Mata, C., et al. (2015). Expression of the tetrahydrofolate-dependent nitric oxide synthase from the green alga Ostreococcus tauri increases tolerance to abiotic stresses and influences stomatal development in Arabidopsis. Plant J. 82, 806–821. doi: 10.1111/tpj.12852
Foyer, C. H., and Noctor, G. (2005). Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 17, 1866–1875. doi: 10.1105/tpc.105.033589
Fürstenberg-Hägg, J., Zagrobelny, M., and Bak, S. (2013). Plant defense against insect herbivores. Int. J. Mol. Sci. 14, 10242–10297. doi: 10.4161/psb.21663
Gotor, C., Garcia, I., Aroca, A., Laureano-Marin, A. M., Arenas-Alfonseca, L., Jurado-Flores, A., et al. (2019). Signaling by hydrogen sulfide and cyanide through post-translational modification. J. Exp. Bot. 70, 4251–4265. doi: 10.1093/jxb/erz225
Gupta, K. J., Fernie, A. R., Kaiser, W. M., and van Dongen, J. T. (2011). On the origins of nitric oxide. Trends Plant Sci. 16, 160–168. doi: 10.1016/j.tplants.2010.11.007
Hancock, J. T. (2019). Hydrogen sulfide and environmental stresses. Environ. Exp. Bot. 161, 50–56. doi: 10.1016/j.envexpbot.2018.08.034
Ischiropoulos, H., and al-Mehdi, A. B. (1995). Peroxynitrite-mediated oxidative protein modifications. FEBS Lett. 364, 279–282. doi: 10.1016/0014-5793(95)00307-u
Jeandroz, S., Wipf, D., Stuehr, D. J., Lamattina, L., Melkonian, M., Tian, Z., et al. (2016). Occurrence, structure, and evolution of nitric oxide synthase-like proteins in the plant kingdom. Sci. Signal. 9:re2. doi: 10.1126/scisignal.aad4403
Jones, J., and Dangl, J. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Keisham, M., Jain, P., Singh, N., von Toerne, C., Bhatla, S. C., and Lindermay, C. (2019). Deciphering the nitric oxide, cyanide and iron-mediated actions of sodium nitroprusside in cotyledons of salt stressed sunflower seedlings. Nitric Oxide 88, 10–26. doi: 10.1016/j.niox.2019.03.008
Leon, J., and Costa-Broseta, A. (2020). Present knowledge and controversies, deficiencies, and misconceptions on nitric oxide synthesis, sensing, and signaling in plants. Plant Cell Environ. 43, 1–15. doi: 10.1111/pce.13617
Li, J., Liu, X., Wang, Q., Huangfu, J., Schuman, M. C., and Lou, Y. (2019). A group D MAPK protects plants from autotoxicity by suppressing herbivore-induced defense signaling. Plant Physiol. 179, 1386–1401. doi: 10.1104/pp.18.01411
Lindermayr, C., Rudolf, E. E., Durner, J., and Groth, M. (2020). Interactions between metabolism and chromatin in plant models. Mol. Metab. 41:101069. doi: 10.1016/j.molmet.2020.01.015
Liu, L., Hausladen, A., Zeng, M., Que, L., Heitman, J., and Stamler, J. S. (2001). A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature 410, 490–494. doi: 10.1038/35068596
Liu, Y., He, J., Jiang, L., Wu, H., Xiao, Y., Liu, Y., et al. (2011). Nitric oxide production is associated with response to brown planthopper infestation in rice. J. Plant Physiol. 168, 739–745. doi: 10.1016/j.jplph.2010.09.018
Mai, V. C., Drzewiecka, K., Jelen, H., Narozna, D., Rucinska-Sobkowiak, R., Kesy, J., et al. (2014). Differential induction of Pisum sativum defense signaling molecules in response to pea aphid infestation. Plant Sci. 22, 1–12. doi: 10.1016/j.plantsci.2014.01.011
Martinez-Medina, A., Pescador, L., Terron-Camero, L. C., Pozo, M. J., and Romero-Puertas, M. C. (2019). Nitric oxide in plant-fungal interactions. J. Exp. Bot. 29, 4489–4503. doi: 10.1093/jxb/erz289
Martínez-Ruiz, A., Araujo, I. M., Izquierdo-Alvarez, A., Hernansanz-Agustín, P., Lamas, S., and Serrador, J. (2013). Specificity in S-nitrosylation: a short-range mechanism for NO signaling? Antioxid. Redox Signal. 19, 1220–1235. doi: 10.1089/ars.2012.5066
Martinez-Ruiz, A., and Lamas, S. (2009). Two decades of new concepts in nitric oxide signaling: from the discovery of a gas messenger to the mediation of nonenzymatic posttranslational modifications. IUBMB Life 61, 91–98. doi: 10.1002/iub.144
Mata-Perez, C., Sanchez-Calvo, B., Padilla, M. N., Begara-Morales, J. C., Valderrama, R., Corpas, F. J., et al. (2017). Nitro-fatty acids in plant signaling: new key mediators of nitric oxide metabolism. Redox Biol. 11, 554–561. doi: 10.1016/j.redox.2017.01.002
Misra, A. N., Misra, M., and Singh, R. (2011). Nitric oxide: a ubiquitous signaling molecule with diverse roles in plants. Afr. J. Plant Sci. 5, 57–74. doi: 10.5897/AJPS.9000178
Molassiotis, A., Job, D., Ziogas, V., and Tanou, G. (2016). Citrus plants: a model system for unlocking the secrets of NO and ROS-inspired priming against salinity and drought. Front. Plant Sci. 7:229. doi: 10.3389/fpls.2016.00229
Moloi, M. J., and van der Westhuizen, A. J. (2009). Involvement of nitric oxide during the Russian wheat aphid resistance. S. Afr. J. Bot. 75:412.
Mur, L. A., Carver, T. L., and Prats, E. (2006). No way to live; the various roles of nitric oxide in plant-pathogen interactions. J. Exp. Bot. 57, 489–505. doi: 10.1093/jxb/erj052
Mur, L. A., Prats, E., Pierre, S., Hall, M. A., and Hebelstrup, K. H. (2013). Integrating nitric oxide into salicylic acid and jasmonic acid/ethylene plant defense pathways. Front. Plant Sci. 4:215. doi: 10.3389/fpls.2013.00215
Noctor, G., Mhamdi, A., Chaouch, S., Han, Y., Neukermans, J., Marquez-Garcia, B., et al. (2012). Glutathione in plants: an integrated overview. Plant Cell Environ. 35, 454–484.
Pentzold, S., Zagrobelny, M., Roelsgaard, P. S., Moller, B. L., and Bak, S. (2014). The multiple strategies of an insect herbivore to overcome plant cyanogenic glucoside defence. PLoS One 9:e91337. doi: 10.1371/journal.pone.0091337
Perazzolli, M., Dominici, P., Romero-Puertas, M. C., Zago, E., Zeier, J., Sonoda, M., et al. (2004). Nonsymbiotic hemoglobin AHb1 modulates nitric oxide bioactivity in Arabidopsis thaliana. Plant Cell 16, 2785–2794. doi: 10.1105/tpc.104.025379
Perazzolli, M., Romero-Puertas, M. C., and Delledonne, M. (2006). Modulation of nitric oxide bioactivity by plant haemoglobins. J. Exp. Bot. 57, 479–488. doi: 10.1093/jxb/erj051
Pieterse, C. M. J., Van der Does, D., Zamioudis, C., Leon-Reyes, A., and Van Wees, S. C. M. (2012). Hormonal modulation of plant immunity. Ann. Rev. Cell Dev. Biol. 28, 489–521. doi: 10.1146/annurev-cellbio-092910-154055
Radi, R. (2004). Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. U.S.A. 101, 4003–4008. doi: 10.1073/pnas.0307446101
Rockel, P., Strube, F., Rockel, A., Wildt, J., and Kaiser, W. M. (2002). Regulation of nitric oxide (NO) production by plant nitrate reductase in vivo and in vitro. J Exp. Bot. 53, 103–110. doi: 10.1093/jexbot/53.366.103
Romero-Puertas, M. C., Laxa, M., Mattè, A., Zaninotto, F., Finkemeier, I., Jones, A. M. E., et al. (2007). S-nitrosylation of peroxiredoxin II E promotes peroxynitrite-mediated tyrosine nitration. Plant Cell 19, 4120–4130. doi: 10.1105/tpc.107.055061
Romero-Puertas, M. C., and Sandalio, L. M. (2016). Nitric oxide level is self-regulating and also regulates its ROS partners. Front. Plant Sci. 7:137. doi: 10.3389/fpls.2016.00316
Rubbo, H. (2013). Nitro-fatty acids: novel anti-inflammatory lipid media-tors. Braz. J. Med. Biol. Res. 46, 728–734. doi: 10.1590/1414-431X20133202
Rubbo, H., and Radi, R. (2008). Protein and lipid nitration: role in redox signaling and injury. Biochim. Biophys. Acta 1780, 1318–1324. doi: 10.1016/j.bbagen.2008.03.007
Sanchez-Vicente, I., Fernandez-Espinosa, M. G., and Lorenzo, O. (2019). Nitric oxide molecular targets: reprogramming plant development upon stress. J. Exp. Bot. 70, 4441–4460. doi: 10.1093/jxb/erz339
Sandalio, L. M. S., Gotor, C., Romero, L. C., and Romero-Puertas, M. C. (2019). Multilevel regulation of peroxisomal proteome by post-translational modifications. Int. J. Mol. Sci. 20:4881. doi: 10.3390/ijms20194881
Santamaria, M. E., Arnaiz, A., Gonzalez-Melendi, P., Martinez, M., and Diaz, I. (2018a). Plant perception and short-term responses to phytophagous insects and mites. Int. J. Mol. Sci. 19:1356. doi: 10.3390/ijms19051356
Santamaria, M. E., Arnaiz, A., Velasco-Arroyo, B., Grbic, V., Diaz, I., and Martinez, M. (2018b). Arabidopsis response to the spider mite Tetranychus urticae depends on the regulation of reactive oxygen species homeostasis. Sci. Rep. 8:9432. doi: 10.1038/s41598-018-27904-1
Santamaria, M. E., Arnaiz, A., Rosa-Diaz, I., Gonzalez-Melendi, P., Romero-Hernandez, G., Ojeda-Martinez, D., et al. (2020a). Plant defences against Tetranychus urticae: mind the gaps. Plants 9:464. doi: 10.3390/plants9040464
Santamaria, M. E., Garcia, A., Arnaiz, A., Rosa-Diaz, I., Romero-Hernandez, G., Diaz, I., et al. (2020b). Comparative transcriptomics unravels hidden biological processes in a plant/arthropod herbivore interaction. J. Integ. Plant Biol (in press). doi: 10.1111/jipb.13026
Santamaria, M. E., Martinez, M., Arnaiz, A., Ortego, F., Grbic, V., and Diaz, I. (2017). MATI, a novel protein involved in the regulation of herbivore-associated signalling pathways. Front. Plant Sci. 8:975. doi: 10.3389/fpls.2017.00975
Santamaria, M. E., Martinez, M., Cambra, I., Grbic, V., and Diaz, I. (2013). Understanding plant defence responses against herbivore attacks: an essential first step towards the development of sustainable resistance against pests. Transgenic Res. 22, 697–708. doi: 10.3390/ijms19051356
Schmiesing, A., Emonet, A., Gouhier-Darimont, C., and Reymond, P. (2016). Arabidopsis MYC transcription factors are the target of hormonal salicylic acid/jasmonic acid cross talk in response to Pieris brassicae egg extract. Plant Physiol. 170, 2432–2443. doi: 10.1104/pp.16.00031
Smith, C. M., and Boyko, E. V. (2007). The molecular bases of plant resistance and defense responses to aphid feeding: current status. Entomol. Exp. Appl. 122, 1–16. doi: 10.1111/j.1570-7458.2006.00503.x
Stahl, E., Hilfiker, O., and Reymond, P. (2018). Plant-arthropod interactions: who is the winner? Plant J. 93, 703–729. doi: 10.1111/tpj.13773
Stöhr, C., and Stremlau, S. (2006). Formation and possible roles of nitric oxide in plant roots. J. Exp. Bot. 57, 463–470. doi: 10.1093/jxb/erj058
Sun, L. R., Yue, C. M., and Hao, F. S. (2019). Uptake on roles of nitric oxide in regulating stomatal closure. Plant Signal Behav. 14:e1649569. doi: 10.1080/15592324.2019.1649569
Tanou, G., Filippou, P., Belghazi, M., Job, D., Diamantidis, G., Fotopoulos, V., et al. (2012). Oxidative and nitrosative-based signaling and associated post-translational modifications orchestrate the acclimation of citrus plants to salinity stress. Plant J. 72, 585–599. doi: 10.1111/j.1365-313X.2012.05100.x
Tsuda, K., and Katagiri, F. (2010). Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 13, 459–465. doi: 10.1016/j.pbi.2010.04.006
Wang, B. L., Shi, L., Li, Y. X., and Zhang, W. H. (2010). Boron toxicity is alleviated by hydrogen sulfide in cucumber (Cucumis sativus L.) seedlings. Planta 231, 1301–1309. doi: 10.1007/s00425-010-1134-9
Wei, J., van Loon, J. J. A., Gols, R., Menzel, T. R., Li, N., Kang, L., et al. (2014). Reciprocal crosstalk between jasmonate and salicylate defence-signalling pathways modulates plant volatile emission and herbivore host-selection behaviour. J. Exp. Bot. 65, 3289–3298. doi: 10.1093/jxb/eru181
Wendehenne, D., Pugin, A., Klessig, D. F., and Durner, J. (2001). Nitric oxide: comparative synthesis and signaling in animal and plant cells. Trends Plant Sci. 6, 177–183. doi: 10.1016/s1360-1385(01)01893-3
Wozniak, A., Formela, M., Bilman, P., Grzeskiewicz, K., Bednarski, W., Marczak, L., et al. (2017). The dynamics of the defense strategy of pea induced by exogenous nitric oxide in response to aphid infestation. Int. J. Mol. Sci. 18:329. doi: 10.3390/ijms18020329
Wu, J., and Baldwin, I. T. (2009). Herbivory-induced signalling in plants: perception and action. Plant, Cell Environ. 32, 1161–1174. doi: 10.1111/j.1365-3040.2009.01943.x
Wünsche, H., Baldwin, I. T., and Wu, J. (2011a). S-Nitrosoglutathione reductase (GSNOR) mediates the biosynthesis of jasmonic acid and ethylene induced by feeding of the insect herbivore Manduca sexta and is important for jasmonate-elicited responses in Nicotiana attenuata. J. Exp. Bot. 62, 4605–4616. doi: 10.1093/jxb/err171
Wünsche, H., Baldwin, I. T., and Wu, J. (2011b). Silencing NOA1 elevates herbivory-induced jasmonic acid accumulation and compromises most of the carbon-based defense metabolites in Nicotiana attenuata. J. Integr. Plant Biol. 53, 619–631. doi: 10.1111/j.1744-7909.2011.01040.x
Xu, Y., Qu, C., Sun, X., Jia, Z., Xue, M., Zhao, H., et al. (2020). Nitric oxide boots Bemisia tabaci performance through the suppression of jasmonic acid signalling pathway in tobacco plants. Front. Physiol. 11:847. doi: 10.3389/fphys.2020.00847
Yamasaki, H., and Sakihama, Y. (2000). Simultaneous production of nitric oxide and peroxynitrite by plant nitrate reductase: in vitro evidence for the NR-dependent formation of active oxygen species. FEBS Lett. 468, 89–92. doi: 10.1016/S0014-5793(00)01203-5
Young, D., Pedre, B., Ezerinxa, D., De Smet, B., Lewandowska, A., Tossounian, M., et al. (2019). Protein promiscuity in H2O2. Antioxid. Redox Sign. 30, 1285–1324. doi: 10.1089/ars.2017.7013
Yun, B. W., Skelly, M. J., Yin, M., Yu, M., Mun, B. G., Lee, S. U., et al. (2016). Nitric oxide and S-nitrosoglutathione function additively during plant immunity. New Phytol. 211, 516–526. doi: 10.1111/nph.13903
Zeidler, D., Zähringer, U., Gerber, I., Dubery, I., Hartung, T., Bors, W., et al. (2004). Innate immunity in Arabidopsis thaliana: lipopolysaccharides activate nitric oxide synthase (NOS) and induce defense genes. Proc. Natl. Acad. Sci. U.S.A. 101, 15811–15816. doi: 10.1073/pnas.0404536101
Zhurov, V., Navarro, M., Bruinsma, K. A., Arbona, V., Santamaria, M. E., Cazaux, M., et al. (2014). Reciprocal responses in the interaction between Arabidopsis and the cell-content-feeding chelicerate herbivore spider mite. Plant Physiol. 164, 384–399. doi: 10.1104/pp.113.321555
Keywords: nitric oxide, phytophagous arthropods, plant defenses, reactive nitrogen species, signaling molecules
Citation: Arnaiz A, Rosa-Diaz I, Romero-Puertas MC, Sandalio LM and Diaz I (2021) Nitric Oxide, an Essential Intermediate in the Plant–Herbivore Interaction. Front. Plant Sci. 11:620086. doi: 10.3389/fpls.2020.620086
Received: 21 October 2020; Accepted: 24 November 2020;
Published: 08 January 2021.
Edited by:
Andrea Chini, National Center for Biotechnology (CNB), SpainReviewed by:
Ran Li, Zhejiang University, ChinaByung-Wook Yun, Kyungpook National University, South Korea
Copyright © 2021 Arnaiz, Rosa-Diaz, Romero-Puertas, Sandalio and Diaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isabel Diaz, i.diaz@upm.es
†These authors have contributed equally to this work