Serological Surveillance of Rabies in Free-Range and Captive Common Vampire Bats Desmodus rotundus
 Jane Megid1*
Jane Megid1*  Julio Andre Benavides Tala1,2
Julio Andre Benavides Tala1,2  Laís Dário Belaz Silva1
Laís Dário Belaz Silva1  Fernando Favian Castro Castro1,3
Fernando Favian Castro Castro1,3  Bruna Letícia Devidé Ribeiro1 Camila Michele Appolinário1
Bruna Letícia Devidé Ribeiro1 Camila Michele Appolinário1  Iana Suly Santos Katz4 Karin Corrêa Scheffer4 Sandriana Ramos Silva4
Iana Suly Santos Katz4 Karin Corrêa Scheffer4 Sandriana Ramos Silva4  Adriana Ruckert Rosa4 Gisely Toledo Barone5 Luzia Fátima Alves Martorelli5† Marilene Fernandes de Almeida5†
Adriana Ruckert Rosa4 Gisely Toledo Barone5 Luzia Fátima Alves Martorelli5† Marilene Fernandes de Almeida5†- 1School of Veterinary Medicine and Animal Science, São Paulo State University (Unesp), Botucatu, Brazil
- 2Centro de Investigación para la sustestabilidad, Facultad de Ciencias de la vida, Universidad Andrés Bello, Santiago, Chile
- 3Animal Health Department, Universidad Antonio Narino Cauca Popayan, Popayán, Colômbia
- 4Diagnostics Sector, Immunology Laboratory Pasteur Institute of São Paulo, São Paulo, Brazil
- 5Centro de Controle de Zoonoses, Coordenadoria de vigilância em saúde, São Paulo, Brazil
The control of vampire bat rabies (VBR) in Brazil is based on the culling of Desmodus rotundus and the surveillance of outbreaks caused by D. rotundus in cattle and humans in addition to vaccination of susceptible livestock. The detection of anti-rabies antibodies in vampire bats indicates exposure to the rabies virus, and several studies have reported an increase of these antibodies following experimental infection. However, the dynamics of anti-rabies antibodies in natural populations of D. rotundus remains poorly understood. In this study, we took advantage of recent outbreaks of VBR among livestock in the Sao Paulo region of Brazil to test whether seroprevalence in D. rotundus reflects the incidence of rabies in nearby livestock populations. Sixty-four D. rotundus were captured during and after outbreaks from roost located in municipalities belonging to three regions with different incidences of rabies in herbivores. Sixteen seropositive bats were then kept in captivity for up to 120 days, and their antibodies and virus levels were quantified at different time points using the rapid fluorescent focus inhibition test (RFFIT). Antibody titers were associated with the occurrence of ongoing outbreak, with a higher proportion of bats showing titer >0.5 IU/ml in the region with a recent outbreak. However, low titers were still detected in bats from regions reporting the last outbreak of rabies at least 3 years prior to sampling. This study suggests that serological surveillance of rabies in vampire bats can be used as a tool to evaluate risk of outbreaks in at risk populations of cattle and human.
Introduction
Rabies is an infectious disease of viral etiology that causes acute encephalitis, with rapid and usually fatal evolution in all mammals (1). The etiological agent is the RNA virus belonging to the family Rhabdoviridae and genus Lyssavirus, and it is a zoonosis distributed worldwide that now affects mainly low-income countries (2). Rabies virus is transmitted by contact with infected saliva through bites or scratches. Dogs are the main reservoir of rabies and are responsible for most fatal cases in humans worldwide (2). However, in Latin America, with success obtained by the countries to control rabies mediated by dogs through the mass vaccination campaign (3, 4), bats have become the main reservoir of rabies over the last decade and are responsible for thousands of cases in livestock and for most of the remaining cases in humans (1, 4). Within bats, the common vampire bat Desmodus rotundus is the most important reservoir because of the high occurrence of spillover of rabies from this species of bat to other animals and humans, and vampire bat rabies (VBR) remains unpredictable and uncontrolled in several areas of the continent.
By feeding every night on livestock, VBR causes significant economic losses in Latin America, particularly to small-scale farmers (5). D. rotundus can also feed on the blood of human beings, resulting in occasional and unpredictable outbreaks of VBR in remote settings such as the Amazon region (6). The epidemiology of VBR circulation among D. rotundus that results in rabies cases among livestock is driven by several natural factors that are still poorly understood. Anthropogenic and ecological features favor the presence of bats (e.g., distribution of cattle herd and land occupation) (7), and factors favoring VBR among bats are also involved. However, understanding VBR circulation requires the capacity of measuring rabies exposure or infections within the bat population.
Most studies understanding rabies circulation among bats rely on official reports of livestock mortality, but this data is often biased by variable levels of under-reporting across the landscap (8, 9). Herbivores are accidental hosts of the rabies virus. Because they are a dead-end host, they only contribute as sentinels in the existence of the virus in the bat population (7). Despite its limitations, studies based on livestock mortality have shown that rabies can circulate in the landscape in different ways including wave-like spread into new areas, metapopulation dynamics, or endemically (9–11). However, the dynamics of the virus in endemic areas where the virus has been established for longer periods of time is less understood. Therefore, surveillance of rabies in bats, although logistically challenging, can bring unique insides into the circulation of the virus and our ability to predict and prevent future outbreaks in humans and livestock (9, 11).
The recognition of bats as reservoirs of the disease made epidemiological surveillance extend to these species. Rabies virus is rarely isolated from an infected bat because infected bats are often lower than 1% of the population (11). Because the presence of anti-rabies antibodies correlates with exposure to the virus, serological studies can contribute to understanding VBR circulation among bats. Several studies have shown the presence of antibodies to rabies in bats that do not die from the disease, as seen in other animals and also in humans (12, 13), implying that exposure to rabies in this species does not necessarily leads to mortality (14). For example, in the Botucatu region of Brazil, an endemic region of VBR, 45% of D. rotundus vampire bats had virus neutralizing antibodies (VNA) titers ranging from 0.10 to 0.20 IU/ml, 9.31% had between 0.20 and 0.3 IU/ml, and 11% had VNA levels >0.30 IU/ml (14). Similarly, D. rotundus bats from 16 colonies were tested in the endemic region of Paraiba in Brazil and all animals presented anti-rabies antibodies titers in some level, with titers higher than 0.5 IU/ml in 30.1% of bats (15). Delpietro et al. (16) reported the detection of 30% antibodies in bats after an outbreak of bovine rabies in Argentina. Before the outbreak, 2% of the bats had the antibodies, and during the outbreak, the percentage increased to 4%, suggesting that the presence of high proportion of seropositivity results from a high circulation of the VBR among bats at the time of an outbreak.
Here, we took advantage of recent outbreaks of VBR among livestock in the Sao Paulo region of Brazil to further test whether seroprevalence in D. rotundus reflects the incidence of rabies in bat populations. Field work was combined with captive experiments to follow seropositive animals and increase our understanding of within-host anti-rabies antibodies dynamics among D. rotundus.
Materials and Methods
Virus Neutralizing Antibodies (VNA) in D. rotundus Bats During and After Outbreaks
We captured and collected samples of 64 D. rotundus from December 2015 to June 2016 from colonies in areas with three different epidemiological scenarios (Figure 1) of outbreaks in livestock of nearby farms attacked by these bats (according to the Ministry of Agriculture, Livestock and Supply from Brazil). Bats were aleatorily captured using nets installed outside the roost. In the first location, 23 bats were caught from roosts located in the University farm of UNESP, Botucatu, São Paulo. This farm just had a rabies outbreak 1 month before sampling involving seven cattle and horses. A second collection of five bats was performed 30 days after in this same spot. The second site was Bofete, where eight bats were captured and the nearby farms had reported rabies among livestock in 2013 and 2016. After 30 days, 16 bats were captured in the same farms. The third site of capture was Anhembi where six bats were captured and rabies has been reported on livestock in 2015 for the last time. After 30 days, six more bats were captured in the same place.
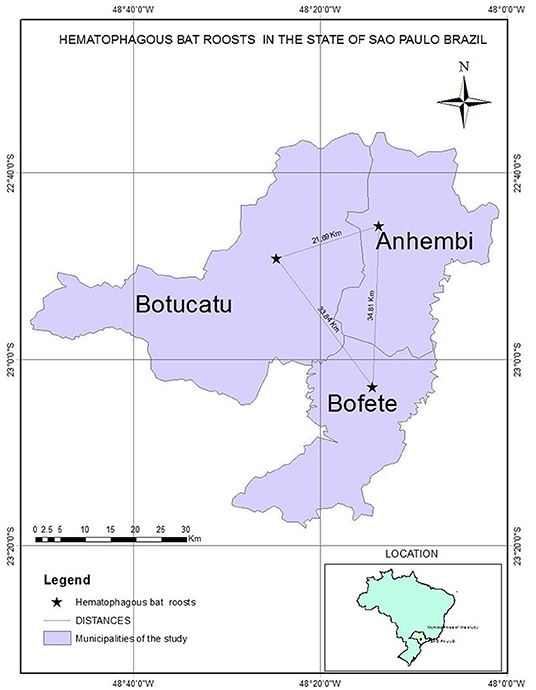
Figure 1. Map showing the three municipalities with different epidemiological scenarios.
Serum Sampling and Rabies Neutralizing Antibody Testing
From the second capture, six bats from Anhembi, four bats from Botucatu, and six from Bofete were aleatorily separated and kept in captivity to study their rabies antibodies for 4 months. The animals were quarantined under observation in isolated cages in a controlled environment in the animal facility of the infectious diseases sector—School of Veterinary Medicine and Animal Science-UNESP/Botucatu (approved by the Ethical Committee of Animals Uses in Veterinary Medicine and Animal Production of São Paulo State University “Julio de Mesquita Filho,” number 85/2015, and SISBIO (Biodiversity Authorization and Information System) license number 51231). Animals were fed with defibrinated bovine blood negative for anti-rabies antibodies using automatic drinking bottles, in addition to daily cleaning of the cages. In all bats captured in nature and not kept in captivity, blood samples were followed by euthanasia using halothane anesthetic induction and deepening of the anesthetic plane with 0.1 ml intraperitoneal ketamine. Bats kept in captivity were subjected to monthly blood collection with anesthetic induction. Blood was collected from the saphenous vein using needles 25 G ½ (0.45 × 13 mm) and capillaries (microhematocrit) with prior asepsis with 70% alcohol. Sera were stored at −20°C until antirabies neutralizing antibodies titer evaluation by the rapid fluorescent focus inhibition test (RFFIT) (17). For the RFFIT (18), a constant dose of a previously titrated (calibrated to give 80% fluorescent focus infected cells) cell culture-adapted RABV challenge virus (CVS) was incubated with 3-fold dilutions of the sera. After incubation of the serum-virus mixtures, a suspension of a clone of BHK-21 (BSR) cells was added. After 20 h of incubation, the cell monolayer was acetone-fixed and labeled with anti-rabies virus antibody conjugated to FITC (19). The optimal challenge dose (the dilution giving 80% infected cells for each virus production) was calculated. Titers of sera were calculated by comparison with a reference serum calibrated to the WHO reference serum. A minimal threshold of 0.5 IU/ml was considered for protection.
Animals that died during observation period (7) and those euthanized on the day of capture (or at the end of the 4 month period) were subjected to the same protocols of cardiac puncture for collection of blood used for RFFIT, and brain was submitted to qRT-PCR for rabies diagnosis as previously described (20), resulting all negatives to rabies.
Given the low number of samples from each type of colony, we used a Pearson's chi-squared test using the R function prop test to compare the proportion of seropositive individuals between day 0 and day 30 of sampling and between sampling locations.
Results
Virus Neutralizing Antibodies (VNA) During and After the Outbreak
The highest rabies antibodies titers were obtained from bats captured in Botucatu (0.45 ± 0.05), which had the most recent outbreaks. In this area, 13 animals out of 23 had protective antibody titers (0.64 ± 0.05). Average titer concentrations in the bats captured 30 days after (0.09 ± 0.01) decreased significantly (p = 0.0075), and none of those five bats had protective titers. Bats from Anhembi and Bofete had a similar average concentration of titers (0.11 ± 0.01 and 0.10 ± 0.01; p = 0.51), respectively, that were lower than in Botucatu (0.45 ± 0.05) at the first capture (p < 0.0001). However, bats from Anhembi maintained the same level of titer 30 days later (0.11 ± 0.03), whereas Bofete presented a titer >10 IU/ml, which increased the overall average concentration (0.70 ± 0.63) (Figure 2). Considering that this bat could interfere with the biological interpretation of our results, we run another statistics test excluding its titer (>10 IU/ml) to the analysis. The statistical analysis showed a significant difference of the VNA titers from bats captured in Bofete in both moments (p = 0.0129). Overall, the proportion of seropositive animals, on day 0, was higher in Botucatu compared to either Anhembi and Bofete (Pearson's test, p < 0.01), and there was no significant difference in the proportion of seropositive individuals between day 0 and 30 in neither of three locations (Pearson's test, p > 0.05). All captured bats were apparently in a good sanitary condition, and no clinical signs were observed. Details on specific VNA are given in Table 1, Figure 2.
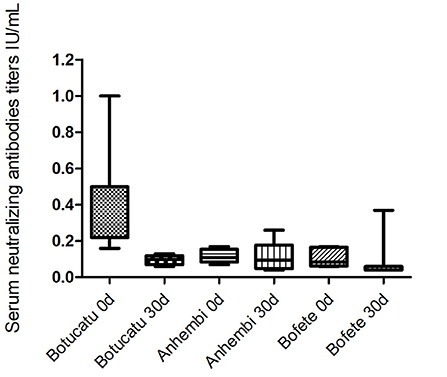
Figure 2. Geometric mean titers (GMT) of virus-neutralizing antibodies in the bats from the studied municipalities. Municipalities with different scenarios evaluated at 0 and 30 days.
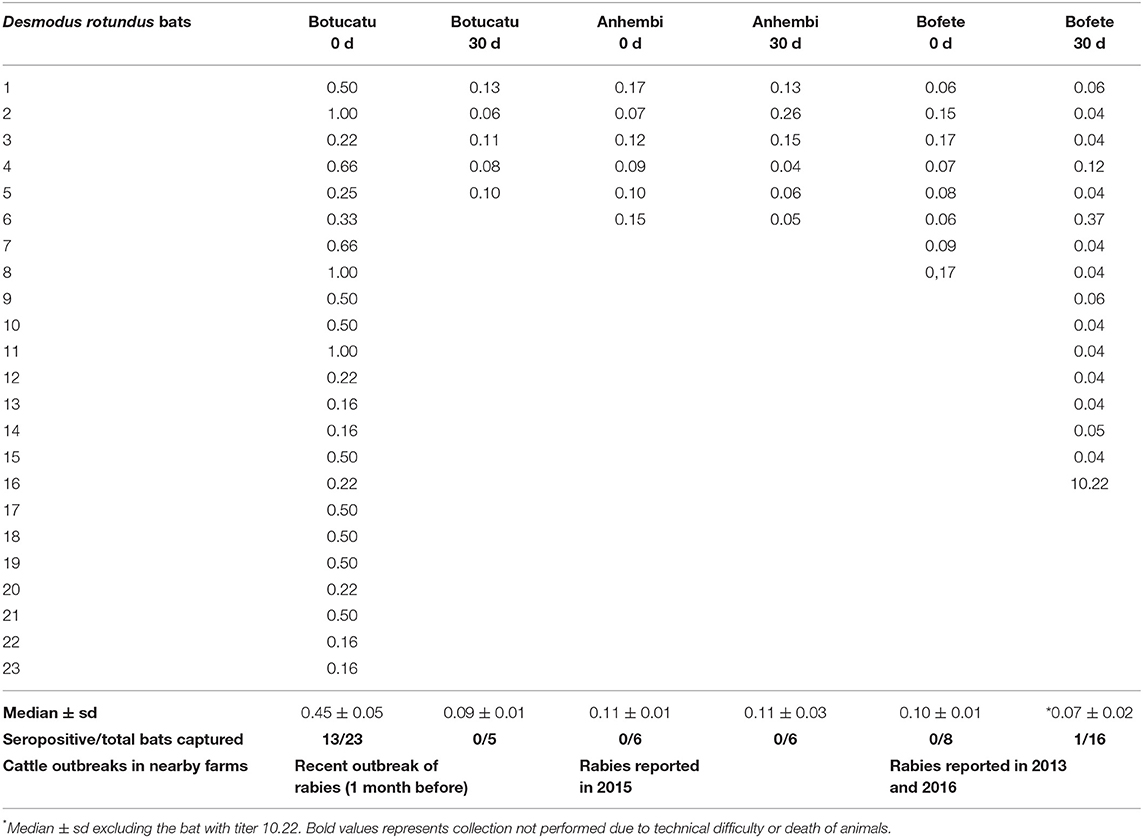
Table 1. Antirabies neutralizing antibodies titers in Desmodus rotundus bats captured, from December 2015 to june 2016, in geographic areas with different epidemiological situation for rabies among livestock.
Virus Neutralizing Antibodies (VNA) Profile of Bats Kept in Captivity
During the observation period, seven bats died and resulted negative by rabies virus qRT-PCR test, so it was possible to have the serological profile from nine bats during the 120 days, one bat from Botucatu, and four bats from Bofete and Anhembi, respectively. The serological profile of bats kept in captivity showed a constant decrease in the titer levels in all animals (Table 2, Figure 3) up to days 120, where all titers were lower than 0.10 (p < 0.0001). The statistical analysis of the VNA titers from day 0 compared to 60 days showed a significant difference (p = 0.0009), and the same result was obtained when comparing to 90 days (p = 0.0017) and 120 days (p = 0.0004). Statistical significance was also evidenced comparing 30–60 days (p = 0.0017), 90 days (p = 0.0090), and 120 days (p = 0.0004), demonstrating a decrease in the levels of VNA in bats in captivity. Although two bats showed an increase in the VNA at 30 days, it is not possible to make any conclusion due to the low number of animals studied.
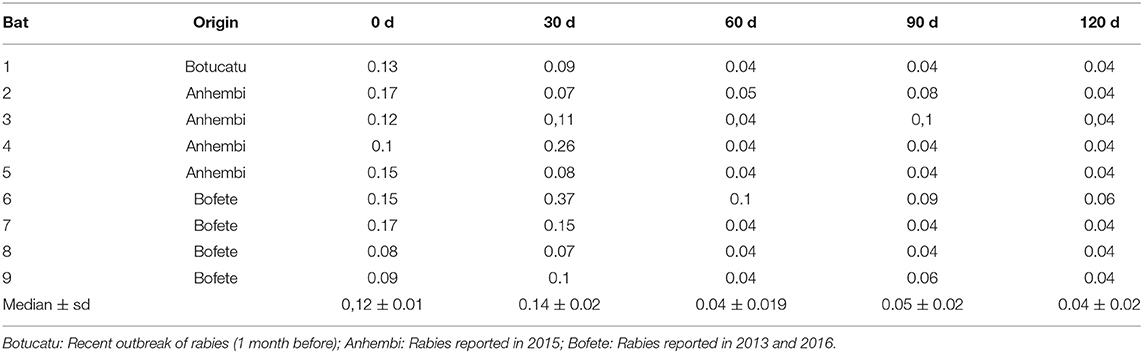
Table 2. Antirabies antibodies titers from bats captured in regions with different epidemiological situation.
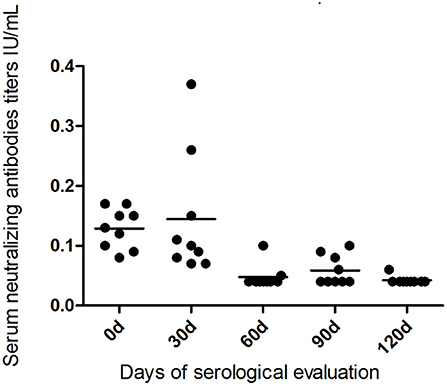
Figure 3. Serological profile of virus-neutralizing antibodies in the bats kept in captivity for 120 days from the studied municipalities.
Discussion
Serological studies of VBR comparing populations of bats with different scenarios of rabies outbreaks in cattle are rare but crucial to understand within-bat dynamics of rabies. We found that vampire bats from the region of Botucatu, which had experienced a recent outbreak of rabies in cattle 30 days prior to sampling, showed a high proportion of seropositive individuals compared to two other sampling sites where the latest outbreak in cattle was reported at least 3 years prior to sampling. In Botucatu, 30 days after the outbreak, half of the bats had antibodies considered to be protective against rabies, which decreased in proportion 60 days after the outbreak in the region. Seropositive animals kept in captivity showed a variable decreased in titer after 4 months, suggesting a rapid loss of antibodies to non-protective levels. Our study suggests that rabies serology in bats can be an accurate indicator to identify if rabies had circulated in bats sampled.
The high proportion of antibodies considered as protective for rabies among bats sampled in Botucatu suggests the presence and circulation of the virus (8, 11) and corroborates cases reported among livestock in nearby farms. Although the second collection of bats 30 days later was much smaller (N = 5), there was a decrease on the proportion of seropositive animals. This decrease suggests that not all bats in a colony are exposed to the virus and that rabies does not necessarily persist within a colony after an outbreak (16).
Although outbreaks from cattle in Bofete and Anhembi were not reported for at least 3 years, antibodies were detected among bats in the first capture in both colonies, but not high enough to be considered protective (<0.5 IU/ml). This could reflect recent exposure to low levels of the virus that does not result in disease symptoms including exposure shortly after birth, subclinical or asymptomatic infections, sublethal infection, carrier state, or latent virus activated by stress (11, 21–23). In fact, Blackwood et al. (11) suggested that the probability of developing a lethal infection upon exposure to rabies is low in vampire bats (~10%), which enables viral persistence in the slowly reproducing bat colonies. This is also observed in other bats species (14, 24).
Low levels of antibodies within a bat colony without recent exposure could also result from long-term persistence of antibodies if rabies exposure happened several years in that colony (8, 25). However, the duration of rabies antibodies after exposure without re-exposure remains poorly understood for vampire bats. In this study, anti-rabies antibodies decreased to low levels 60 days after capture of seropositive individuals but fluctuations and sudden increases were also observed in some individuals. Antibodies fluctuation has been reported in free bats but explained by re-exposure to the virus, which was not possible in our laboratory settings (26). This result suggests possible rabies virus persistence in the bat tissue with periodic antigenic stimulation. This calls for future studies on the mechanisms behind anti-rabies antibody production and on the immunological and protective consequences of having low levels of antibodies in healthy bats.
The increase of anti-rabies antibodies titers on the second capture observed in Bofete suggests that rabies circulation can be detected in the bat population despite cases not been reported among livestock. This highlights the possibility of using bat serology surveillance to predict risk in that population and therefore affects livestock in the region too. Therefore, cost-effective analysis and more understanding on the serological behavior of bat population are needed to evaluate whether rabies serological surveillance among bats can be added as a tool to improve our understanding and prevention of rabies risk in both livestock and humans.
Overall, our study shows that anti-rabies antibodies among vampire bats could reflect the current risk of rabies outbreaks among nearby cattle. Furthermore, we observed a decrease in antibody titers 2 month after animals had high titers, although titers fluctuated in time and between individual bats. Future studies are required to assess whether the observed titer levels >0.5 IU/ml are indeed protective against rabies on bats and whether low-titers can also reflect rabies circulation among bats and subsequent risk to livestock and humans.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by CEUA FMVZ.
Author Contributions
FC, LB, BR, CA, and JM Bats capture, blood collection and work execution. JB, JM, and LB Foreground research, writing and editing. IK, KS, SS, AR, GB, and LA Serology. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2021.681423/full#supplementary-material
References
1. World Health Organization. WHO Expert Consultation on Rabies: Third Report (Geneva). Vol. 1012 (2018).
2. Hampson K, Coudeville L, Lembo T, Sambo M, Kieffer A, Attlan M, et al. Estimating the global burden of endemic canine rabies. PLoS Negl Trop Dis. (2015) 9:1–20. doi: 10.1371/journal.pntd.0003786
3. Schneider MC, Almeida GA, Souza LM, Morares NB, Diaz RC. Controle da raiva no Brasil de 1980 a 1990. Rev Saúde Pública. (1996) 30:196–203. doi: 10.1590/S0034-89101996000200012
4. Wada MY, Rocha SM, Maia-Elkhoury ANS. Situação da Raiva no Brasil, 2000 a 2009. Epidemiol Serv Saúde Brasília. (2011) 20:509–18. doi: 10.5123/S1679-49742011000400010
5. Benavides JA, Rojas Paniagua E, Hampson K, Valderrama W, Streicker DG. Quantifying the burden of vampire bat rabies in Peruvian livestock. PLoS Negl Trop Dis. (207) 11:e0006105. doi: 10.1371/journal.pntd.0006105
6. Grenhall AM. Etología y ecología de Los murciélagos vampiros. Reunión de consulta sobre la atención a personas expuestas a rabia transmitida por vampiros. Washington, DC: Organización Panamericana de la Salud/Organización Mundial de la Salud (1991).
7. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Análise de indicadores epidemiológicos da raiva dos herbívoros no Brasil. Brasília: Ministério da Agricultura, Pecuária e Abastecimento (2013).
8. de Thoisy B, Bourhy H, Delaval M, Pontier D, Dacheux L, Darcissac E, et al. Bioecological drivers of rabies virus circulation in a neotropical bat community. PLoS Negl Trop Dis. (2016) 10:e0004378. doi: 10.1371/journal.pntd.0004378
9. Benavides JA, Valderrama W, Streicker DG. Spatial expansions travelling waves of rabies in vampire bats. Proc R Soc B Biol Sci. (2016) 283:20160328. doi: 10.1098/rspb.2016.0328
10. Streicker DG, Winternitz JC, Satterfield DA, Condori-Condori RE, Broos A, Tello C, et al. Host–pathogen evolutionary signatures reveal dynamics and future invasions of vampire bat rabies. Proc Natl Acad Sci USA. (2016) 113:10926–31. doi: 10.1073/pnas.1606587113
11. Blackwood JC, Streicker DG, Altizer S, Rohani P. Resolving the roles of immunity, pathogenesis, and immigration for rabies persistence in vampire bats. Proc Natl Acad Sci USA. (2013) 110:20837–42. doi: 10.1073/pnas.1308817110
12. Benavides JA, Velasco-Villa A, Godino LC, Satheshkumar PS, Nino R, Rojas-Paniagua E, et al. Abortive vampire bat rabies infections in Peruvian peridomestic livestock. PLOS Negl Trop Dis. (2020) 14:e0008194. doi: 10.1371/journal.pntd.0008194
13. Gilbert AT, Petersen BW, Recuenco S, Niezgoda M, Gómez J, Laguna-Torres VA, et al. Evidence os rabies virus exposure among humans in the peruvian amazon. Am J Trop Med Hyg. (2012) 87:206–15. doi: 10.4269/ajtmh.2012.11-0689
14. Langoni H, Souza LC, Zetun CB, Silva TCC, Hoffmann JL, Silva RC. Serological survey for rabies in serum samples from vampire bats (Desmodus rotundus) in Botucatu region, SP. Brazil. J Venomous Anim Toxins Trop Dis. (2008) 14:651–8. doi: 10.1590/S1678-91992008000400008
15. Ferrari JJF. Morcego hematófago comum Desmodus rotundus no Vale do Paraíba, Estado de São Paulo: abrigos diurnos, agrupamentos, lesões corporais e sorologia antirrábica [monographie on the Internet]. Universidade de São Paulo, São Paulo. Tese de Doutorado em Saúde Publica (2015). Available online at: http://www.teses.usp.br/teses/disponiveis/6/6132/tde-03082015-113130/pt-br.php (accessed August 07, 2021).
16. Del Pietro HA, Lord RD, Russo RG, Gury-Shomen F. Observations of Sylvatic Rabies in Northern Argentina during outbreaks of paralytic cattle rabies transmitted by vampire bats (Desmodus rotundus). J Wildlife Dis. (1992) 45:1168–73. doi: 10.7589/0090-3558-45.4.1169
17. Smith JS, Yager PA, Baer GM. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus neutralizing antibodies. In: Meslin FX. Kaplan MM, Koprowski H, editors. Laboratory Techniques in Rabies. 4th ed. Genebra: World Health Organization (1996). p. 181–92.
18. World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2017. Chapter 2.1.17. Rabies (infection with rabies vírus) (NB: Version adopted in May 2013) (Paris).
19. Almeida MF, Massad E, Aguiar EAC, Martorelli LFA, Joppert AMS. Neutralizing antirabies antibodies in urban terrestrial wildlife in Brazil. J Wildl Dis. (2001) 37:394–8. doi: 10.7589/0090-3558-37.2.394
20. Appolinário C, Allendorf SD, Vicente AF, Ribeiro BD, Fonseca CR, Antunes JM, et al. Fluorescent antibody test, quantitative polymerase chain reaction pattern and clinical aspects of rabies virus strains isolated from main reservoirs in Brazil. Braz J Infect Dis. (2015) 19:479–85. doi: 10.1016/j.bjid.2015.06.012
21. Turmelle AS, Jackson FR, Green D, McCracken GF, Rupprecht CE. Host immunity to repeates rabies vírus infection in big Brown bats. J Gen Virol. (2010) 91:2360–6. doi: 10.1099/vir.0.020073-0
22. Steece RS, Altenbach JS. Prevalence of rabies specific antibodies in the Mexican free-tailed bat (Tadarida brasiliensis mexicana) at Lava cave, New Mexico. J Wildl Dis. (1989) 25:490–6. doi: 10.7589/0090-3558-25.4.490
23. Hill RE, Beran GW, Clark WR. Demonstration of rabies virus-specific antibody in the sera of free-ranging Iowa raccoons (Procyon lotor). J Wildl Dis. (1992) 28:377–85. doi: 10.7589/0090-3558-28.3.377
24. Turmelle AS, Jackson FR, Green D, McCracken GF, Rupprecht CE. Ecology of rabies virus exposure in colonies of Brazilian Free-Tailed Bats (Tadarida brasiliensis) at natural and Man-Made Roosts in Texas. Vector-Borne Zoonotic Dis. (2010) 10:165–75. doi: 10.1089/vbz.2008.0163
25. Amengual B, Bourhy H, Lopez-Roig M, Serra-Cobo J. Temporal dynamics of european bat lyssavirus Type 1 and survival of myotis bats in natural colonies. PLoS ONE. (2007) 2:566. doi: 10.1371/journal.pone.0000566
Keywords: rabies, serology, RFFIT, vampire bats, antibodies, virus neutralizing antibodies
Citation: Megid J, Benavides Tala JA, Belaz Silva LD, Castro Castro FF, Ribeiro BLD, Appolinário CM, Katz ISS, Scheffer KC, Silva SR, Rosa AR, Barone GT, Alves Martorelli LF and de Almeida MF (2021) Serological Surveillance of Rabies in Free-Range and Captive Common Vampire Bats Desmodus rotundus. Front. Vet. Sci. 8:681423. doi: 10.3389/fvets.2021.681423
Received: 16 March 2021; Accepted: 28 July 2021;
Published: 29 September 2021.
Edited by:
Marco Antonio Natal Vigilato, Pan American Health Organization, United StatesReviewed by:
Felipe Rocha, Pan American Health Organization, BrazilFlorence Cliquet, Laboratoire de la rage et de la faune sauvage de Nancy, Agence Nationale de Sécurité Sanitaire de l'Alimentation, de l'Environnement et du Travail (ANSES), France
Semmannan Kalaiyarasu, ICAR-National Institute of High Security Animal Diseases (ICAR-NIHSAD), India
Copyright © 2021 Megid, Benavides Tala, Belaz Silva, Castro Castro, Ribeiro, Appolinário, Katz, Scheffer, Silva, Rosa, Barone, Alves Martorelli and de Almeida. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jane Megid, jane.megid@unesp.br
†Researcher Retired From the Centro de Controle de Zoonoses, São Paulo, Brazil