 Epaminondas Doxakis
Epaminondas Doxakis Yuan Chao Xue
Yuan Chao Xue Anca F. Savulescu
Anca F. Savulescu- 1Center of Basic Research, Biomedical Research Foundation of the Academy of Athens, Athens, Greece
- 2Department of Pathology, University of Texas Medical Branch, Galveston, TX, United States
- 3Division of Chemical, Systems and Synthetic Biology, Institute for Infectious Disease and Molecular Medicine, Faculty of Health Sciences, University of Cape Town, Cape Town, South Africa
Introduction
The traditional view that RNA-binding proteins (RBPs) function at single, discrete checkpoints (e.g., splicing, translation) inadequately captures their regulatory complexity, particularly in neurons (Cajigas et al., 2012; Miura et al., 2013; Tushev et al., 2018). A given RBP can act sequentially across the entire transcript life cycle, from transcription and splicing through export, localization, translation, and decay, on long, isoform-diverse transcripts deployed across distinct neuronal compartments. Neuronal phenotypes frequently emerge from coupled regulatory steps rather than isolated checkpoints. A life cycle framework clarifies where an RBP acts on specific neuronal transcripts and how these actions propagate across subsequent regulatory steps.
Synaptic mRNAs carry characteristically long 3′ untranslated regions (3′UTRs) with roughly twice as many predicted miRNA sites per kilobase, expanding the regulatory surface for RBPs and miRNAs (Paschou et al., 2012). During neuronal differentiation, many genes actively lengthen both poly(A) tails and 3′UTRs, highlighting that 3′UTR extension is not a static feature but a regulated neuronal program (Miura et al., 2013; Kiltschewskij et al., 2023). This increased regulatory complexity necessitates systematic approaches that track RBP actions across checkpoints rather than attributing effects to single regulatory steps. Adopting a life cycle perspective prevents misattribution by identifying where along the regulatory pathway an RBP influences specific neuronal transcripts.
Scope and use of non-neuronal data
Although most foundational studies of RBP mechanisms have been conducted in non-neuronal systems, the core regulatory principles are largely conserved across cell types (Corley et al., 2020; Engel et al., 2020). Where neuronal-specific data exist, these are prioritized; non-neuronal findings are included when they illuminate fundamental mechanisms likely operative in neurons.
Mechanistic checkpoints across the transcript life cycle
Regulatory commitment begins in the nucleus
Several RBPs function at transcriptional checkpoints before any post-transcriptional decisions occur. For instance, heterogeneous nuclear ribonucleoprotein A1 (hnRNPA1) binds single-stranded DNA and G-quadruplex-forming promoter regions and engages the 7SK-HEXIM-P-TEFb axis to influence RNA polymerase II (RNAPII) pause-release and elongation (Zhang et al., 2006; Barrandon et al., 2007; Nishikawa et al., 2019). RNAPII elongation rate directly influences exon choice and 3′-end selection. Reduced elongation shifts poly(A) site usage toward proximal sites, whereas accelerated elongation favors distal sites in vivo (Geisberg et al., 2020; Yague-Sanz et al., 2020). In parallel, U1 small nuclear ribonucleoprotein (U1 snRNP)-mediated “telescripting” suppresses premature cleavage and polyadenylation across nascent RNAs, including long introns, thereby preserving full-length pre-mRNA and shaping alternative polyadenylation (APA) outcomes (Kaida et al., 2010; Berg et al., 2012; So et al., 2019).
Nuclear factors subsequently bias 3′-end choice in neurons. Cleavage factor Im complex 25 kDa subunit (CFIm25, NUDT21) promotes distal polyadenylation; reduced CFIm25 levels shift usage toward proximal sites, shorten 3′UTRs in the mouse hippocampus, and produce learning deficits alongside cortical hyperexcitability in vivo (Alcott et al., 2020). In Drosophila, embryonic lethal abnormal visual-like (ELAV/Hu) paralogs drive global neuronal 3′UTR extension (Hilgers et al., 2012; Oktaba et al., 2015). In mammals, neuronal ELAVLs (ELAV2-4, HuB/C/D) regulate alternative polyadenylation at defined loci such as ELAVL1 (HuR), where distal site usage during differentiation produces long 3′UTR isoforms with reduced translation and stability (Dai et al., 2012; Mansfield and Keene, 2012). Recent work identifies PQBP1 as a regulator of APA in neural progenitor cells. PQBP1 interacts with UGUA motifs and can impede recruitment of the CFIm complex, maintaining cell-specific poly(A) site profiles and balancing progenitor proliferation and differentiation (Liu et al., 2024).
These 3′UTR decisions have direct functional consequences. At the brain-derived neurotrophic factor (Bdnf) locus, short- and long-3′UTR isoforms follow different routes: the short isoform remains in the soma, whereas the long isoform localizes to dendrites and supports local functions in hippocampal neurons (An et al., 2008). The long 3′UTR imposes translational restraint at baseline but permits rapid activity-dependent translation, in contrast to the constitutively active short 3′UTR (Lau et al., 2010). Neurotrophins and distinct RBP assemblies further modulate dendritic targeting of these Bdnf isoforms (Vicario et al., 2015).
Export is not a neutral handoff: nuclear assembly decisions carry through to shape cytoplasmic fate
The transcription and export (TREX) complex assembles during splicing and 3′-end processing, licensing nuclear RNA export factor 1 (NXF1/TAP) to bind mature messenger ribonucleoproteins (mRNPs) and thereby coupling nuclear processing to export competence and downstream cytoplasmic availability (Viphakone et al., 2012; Puhringer et al., 2020). Co-transcriptional deposition of the exon junction complex at exon-exon junctions provides a platform that interfaces with NXF1 loading, linking splicing to export and downstream surveillance mechanisms such as nonsense-mediated decay and translation enhancement (Le Hir et al., 2000; Viphakone et al., 2019).
Export routes can switch under defined cellular stresses. During heat shock, HuR -mediated RNA export shifts to a chromosome region maintenance 1/exportin 1 (CRM1/XPO1)-dependent route via its shuttling ligands pp32/ANP32A and APRIL/ANP32B; under these conditions export becomes leptomycin B-sensitive, and CRM1 co-immunoprecipitates with HuR only following heat shock (Gallouzi et al., 2001). In addition, after nuclear export, 3′UTRs can undergo remodeling through endonucleolytic cleavage. For instance, cytoplasmic cleavage of the inositol monophosphatase 1 (IMPA1) 3′UTR generates a more translatable isoform, which is required for maintaining axon integrity (Andreassi et al., 2021).
Localization and translation are often co-regulated in neurons
Many neurite-bound transcripts carry RNA G-quadruplex motifs that recruit fragile X mental retardation protein (FMRP). FMRP promotes projection-side localization while stalling elongation, coupling transport to translational restraint with stimulus-triggered release (Darnell et al., 2011; Goering et al., 2020). Among cytoskeletal transcripts subject to FMRP-linked ribosome stalling in neurons is MAP1B (Darnell et al., 2011). Further, local translation and mitochondrial tethering of Pink1 mRNA by synaptojanin 2 binding protein (SYNJ2BP) and synaptojanin 2 (SYNJ2A) are required for activation of the PINK1/Parkin pathway in axons (Harbauer et al., 2022).
Localization codes operate within neuronal transcripts. The β-actin zipcode bound by zipcode-binding protein 1 (ZBP1/IGF2BP1) mediates dendritic transport and local translation, and the A2 response element recognized by hnRNPA2/B1 supports activity-regulated dendritic delivery (Eom et al., 2003; Shan et al., 2003; Patel et al., 2012; Leal et al., 2014). Coding regions can also direct targeting. In motor neuron axons, Cox7c mRNA co-transports with mitochondria through a mechanism that depends on the coding region, not the 3′UTR (Cohen et al., 2022). Mechanistically, RBPs couple cargo to motors via adaptors such as adenomatous polyposis coli (APC), which links select mRNAs to kinesin complexes for bidirectional transport in neurons (Baumann et al., 2022).
This cross-regulatory coordination appears in neuron-focused crosslinking and immunoprecipitation (CLIP) datasets, which show RBP occupancy spanning splicing enhancers and 3′UTRs, indicating interactions across multiple regulatory steps. In the brain, the RBP neuro-oncological ventral antigen (NOVA) binds splice-regulatory elements and 3′UTRs, primarily regulating neuronal splicing programs while also affecting alternative polyadenylation at select loci (Licatalosi et al., 2008). RNA-binding FOX protein (RBFOX) family binding maps define intronic sites that predict neuronal splicing programs; additionally, cytoplasmic RBFOX1 binds 3′UTRs in neurons and increases target mRNA stability (Weyn-Vanhentenryck et al., 2014; Lee et al., 2016).
Noncoding RNAs add regulatory complexity. Long non-coding RNA (lncRNA)-mRNA base pairing through inverted Alu elements can create Staufen-binding sites in trans, triggering Staufen-mediated decay (SMD) (Gong and Maquat, 2011). Moreover, circular RNAs (circRNAs) can escort RBPs to their targets, as demonstrated by circNSUN2 forming a ternary complex with IGF2BP2 and HMGA2 mRNA that stabilizes the message (Chen et al., 2019).
RNA modifications further modulate these relationships
In the hippocampus, m6A reader YTH N6-methyladenosine RBP F1 (YTHDF1) enhances translation of synaptic transcripts in an activity-dependent manner and is required for learning and long-term potentiation (Shi et al., 2018). In the nucleus, the m6A modification increases HNRNPC access by locally relaxing RNA structure, which alters splicing and RNA abundance, linking epitranscriptomic marks to RBP occupancy and downstream processing (Liu et al., 2015). Additional modifications provide export- and binding-sensitive regulatory levers. m5C deposition by NOP2/Sun RNA methyltransferase (NSUN2) and recognition by Aly/REF export factor (ALYREF) promote mRNA export (Yang et al., 2017). mRNA pseudouridylation represents another widespread modification that increases under serum starvation; TruB pseudouridine synthase family member 1 (TRUB1) serves as a major contributor to mRNA pseudouridine in mammalian cells (Carlile et al., 2014; Safra et al., 2017). When pseudouridine occurs at stop codons, it can reduce termination efficiency (Karijolich and Yu, 2011). These examples illustrate how chemical modifications and cellular conditions can reroute the same transcript through different regulatory outcomes. The modifications create context-dependent switches that alter RBP binding patterns and downstream transcript processing, demonstrating another layer of life cycle regulation beyond protein-RNA interactions alone.
A single-gene paradigm illustrates how regulatory steps combine
AU-rich element RNA binding protein 1 (AUF1/HNRNPD) binds proximal and distal elements in the SNCA 3′UTR and coordinates multiple regulatory steps in cellular systems: it influences pre-mRNA maturation, is necessary for efficient nuclear export, promotes deadenylation-linked decay of isoforms with shorter 3′UTRs, and reduces ribosome engagement, together lowering α-synuclein output (Kattan et al., 2023). Within this same transcript, Pumilio RNA-binding protein (PUM1) binds two conserved sites in the SNCA 3′UTR and preferentially suppresses the long 3′UTR isoform. PUM1 redistributes SNCA between soma and axons, normalizes α-synuclein levels in patient-derived neurons with SNCA locus triplication, and modulates microRNA responsiveness, indicating coordinated control across isoform choice, subcellular routing, and post-transcriptional sensitivity (Cabaj et al., 2025). This single-locus analysis demonstrates how multiple RBPs can coordinate transcript processing, export, stability, localization, and translation to control protein output through interconnected rather than independent mechanisms.
Stress conditions rewire RBP-mRNA partnerships and translation
The RBPs T-cell intracellular antigen 1 (TIA1) and TIA1-related protein TIAR (TIAL1) accumulate in the cytoplasm during mild heat shock and nucleate stress granules (SGs) when eukaryotic initiation factor 2 alpha (eIF2α) becomes phosphorylated (Kedersha et al., 1999). Additionally, TIA1 oxidation decreases SG assembly and increases sensitivity to apoptosis (Arimoto-Matsuzaki et al., 2016). The TIA1 proximity interactome reveals that TIA1 partners shift between basal conditions and sodium-arsenite stress, indicating that the protein machinery associated with specific RNAs adapts to cellular conditions (Gourdomichali et al., 2022). During osmotic shock, hnRNPA1 becomes hyperphosphorylated and accumulates in the cytoplasm (Allemand et al., 2005). Under sodium arsenite, heat shock, or hyperosmotic stress, hnRNPA1 also relocates to SGs (Guil et al., 2006). Depletion of nuclear hnRNPA1 alters alternative splicing, linking stress-driven relocalization of splicing regulators to splicing control (Allemand et al., 2005). Phase-separated condensates formed by RBPs represent distinct regulatory compartments that reorganize under stress (Brangwynne et al., 2009; Lin et al., 2024).
At the level of output, translation during stress becomes selectively altered for defined transcript subsets. When eIF2α is phosphorylated, mRNAs with upstream open reading frames (uORFs) in their 5′ leaders, such as activating transcription factor 4 (ATF4), undergo preferential translation via delayed reinitiation (Harding et al., 2000; Vattem and Wek, 2004). When cap-dependent initiation becomes attenuated, cap-independent translation via internal ribosome entry site (IRES)-like elements can sustain protein output; the SNCA 5′UTR supports such activity (Koukouraki and Doxakis, 2016). Through a distinct pathway, mechanistic target of rapamycin complex 1 (mTORC1) inhibition triggers La-related protein 1 (LARP1)-dependent repression of 5′-terminal oligopyrimidine (5′-TOP) mRNAs (Fonseca et al., 2015; Philippe et al., 2020).
These stress-responsive mechanisms illustrate how cellular conditions can rapidly reconfigure RBP-transcript relationships and redirect the same transcripts through alternative regulatory pathways, thereby introducing temporal dynamics to the life cycle framework.
An experimental design that captures cross-regulatory coordination
A practical framework that tracks mRNA transcripts across regulatory steps (transcription, processing, export, localization, translation, decay) captures complex regulation more accurately than relying on single assays (Figure 1). Integrating information from different experimental approaches is essential to understanding the broader regulatory scheme controlled by RBPs in neurons. The experimental design outlined below can be repeated under defined stress conditions when appropriate, to separate primary from secondary effects and assign mechanisms to the correct regulatory step of the transcript life cycle (Figure 1).
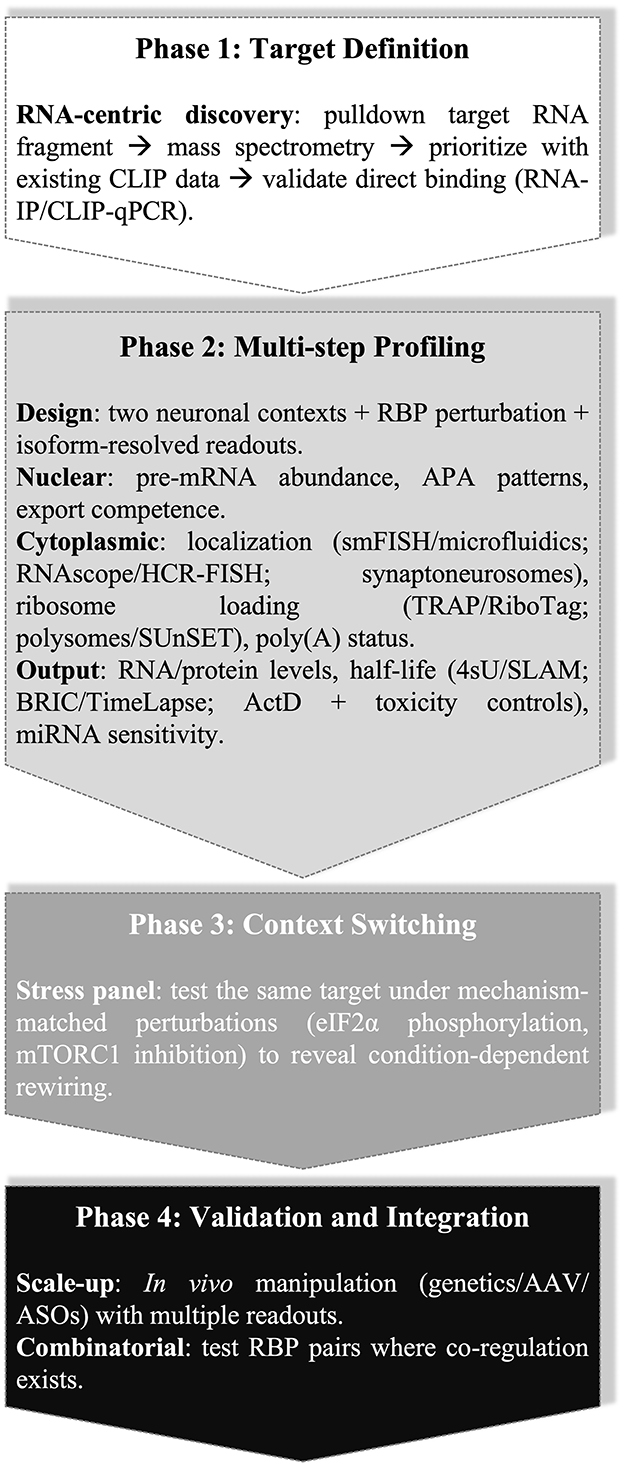
Figure 1. Four-phase, isoform-aware workflow for mapping RBP control across the transcript life cycle in neurons.
Begin with RNA-centric candidate discovery for a defined target transcript. Perform biochemical pulldown of a 3′UTR segment or exon, followed by mass spectrometry. Prioritize candidates using CLIP-class binding evidence where available, and confirm direct binding on endogenous RNA by RNA immunoprecipitation (RNA-IP) or CLIP-qPCR (Yoon and Gorospe, 2016; Kattan et al., 2023). Proceed to an isoform-aware experimental panel using two neuronal contexts that differ in baseline RBP expression or RNA metabolism. Following relevant RBP perturbation, quantify pre-mRNA (transcriptional input), mature isoforms (splicing and alternative polyadenylation outcomes), and nuclear-cytoplasmic distribution with fraction-purity controls (e.g., MALAT1 and/or RNU6 for nuclear; GAPDH mRNA for cytoplasm), using conventional fractionation with isoform-specific RT-qPCR or RNA-seq, and assess subcellular localization using single-molecule FISH (smFISH) (Raj et al., 2008), axon-soma microfluidics (Taylor et al., 2005), or synapse-targeted micro-local perfusion (microLP) (Taylor et al., 2010). When smFISH or microfluidics are unavailable, synaptoneurosome preparations provide synapse-enriched RNA for biochemical assays (Westmark et al., 2011). RNAscope (Wang et al., 2012) and hybridization chain reaction (HCR) FISH (Choi et al., 2014) are additional in situ options.
Profile ribosome engagement using FLAG-RPL22 IP (Kattan et al., 2023), translating ribosome affinity purification (TRAP) (EGFP-L10a) (Heiman et al., 2014), or ribosomal tagging (RiboTag) (HA-RPL22) in Cre-defined neurons (Sanz et al., 2009), followed by quantification of isoforms using distal 3′UTR primers. Classical sucrose-gradient polysome profiling with isoform-specific qPCR is an effective substitute (Chasse et al., 2017), and SUnSET provides a complementary non-genetic readout of protein synthesis (Schmidt et al., 2009).
To integrate transcript stability with translation effects, measure poly(A) tail lengths using the extended poly(A) test (ePAT), and analyze isoform-specific tail-length shifts alongside ribosome engagement, half-life, and protein output (Janicke et al., 2012; Subtelny et al., 2014; Lima et al., 2017). Test miRNA sensitivity, since 3′UTR switching can expose or occlude binding sites, and RBPs can remodel Ago proteins access through competition or structural changes (Bhattacharyya et al., 2006; Surgucheva et al., 2013; Kim et al., 2021; Cabaj et al., 2025).
For decay measurements, avoid treatments that disrupt endogenous recognition and execution steps. In mammalian cells, PAN2-PAN3 initiates poly(A) shortening while CCR4-NOT cooperates with poly(A)-binding protein to complete deadenylation (Uchida et al., 2004; Yi et al., 2018). Shortened tails permit decapping by DCP2 assembled on the EDC4 scaffold with DCP1, and decapped RNA undergoes 5′ to 3′ degradation by 5′-3′ exoribonuclease 1 (XRN1) (Chang et al., 2014; Brothers et al., 2023). Estimate isoform-specific half-lives using 4sU pulse-chase coupled to thiol(SH)-linked alkylation for the metabolic sequencing of RNA (SLAM-seq) or SLAM-qPCR, which preserves translation during measurement (Herzog et al., 2017). Alternative approaches include bromouridine immunoprecipitation chase sequencing (BRIC-seq) or BRIC-qPCR with 5-bromouridine labeling (Tani et al., 2012) and TimeLapse-seq for chemical recoding of 4sU (Schofield et al., 2018). Where chemistry or platforms are constrained, conventional actinomycin D chase with qPCR provides decay estimates, but interpretation requires toxicity controls such rRNA processing markers and p53-responsive transcripts (Bensaude, 2011). Avoid global translation inhibitors (e.g., cycloheximide) when assaying translation-dependent decay, since they suppress SMD and nonsense-mediated decay (NMD); instead, infer decay from labeling methods or brief, toxicity-controlled transcriptional blocks (Gong and Maquat, 2011).
When stress-dependence appears likely, repeat the isoform-aware panel under defined, mechanism-matched perturbations: heat shock to probe CRM1-dependent nuclear export switches (Gallouzi et al., 2001); sodium arsenite for eIF2α phosphorylation and stress-granule formation (Kedersha et al., 1999; Gourdomichali et al., 2022); mTORC1 inhibition to test 5′-TOP control via LARP1 (Fonseca et al., 2015; Philippe et al., 2020); cap-dependent attenuation to assay IRES-mediated translation, such as the SNCA 5′UTR (Koukouraki and Doxakis, 2016); or ER stress (e.g., thapsigargin) to test uORF-dependent translation (ATF4) (Harding et al., 2000; Vattem and Wek, 2004).
Two extensions add mechanistic weight once single-factor effects are mapped. First, test in vivo by altering RBP levels with conditional genetics, focal adenoviral-associated viruses (AAV) delivery, or intracerebroventricular antisense oligonucleotides, and measure the same pre-mRNA, isoform, localization, ribosome engagement, and protein endpoints in targeted brain regions or cell types. Second, when two RBPs independently regulate the same target, consider pairwise perturbations guided by co-occupancy evidence and mechanistic precedent, as shown for neuronal ELAVLs and AUF1 on APP (Fragkouli et al., 2017).
Future technological integration
Single-nucleus RNA-seq and spatial transcriptomics add cell-type and spatial context to the isoform-aware panel, enabling tissue-level maps of neuronal programs and RNA localization, and placing RBP effects in their native cellular niches (Chen et al., 2015; Hu et al., 2017; Rodriques et al., 2019; Booeshaghi et al., 2021). Patient-derived iPSC neurons with mutant transactive response DNA binding protein 43 kDa (TDP-43) or fused in sarcoma (FUS) show disease-relevant phenotypes (Bilican et al., 2012; Devlin et al., 2015; Higelin et al., 2016). In these human neurons, genome-scale CRISPRi screens can identify modifier genes and compensatory pathways (Tian et al., 2019). Long-read/native RNA sequencing resolves full-length isoforms on single molecules and can report RNA modifications, exposing RBP-sensitive isoform choices and marks that alter RBP binding (Garalde et al., 2018; Workman et al., 2019; Begik et al., 2021).
Concluding remarks
Neuronal RNA regulation operates through coupled, sequential steps where early processing choices constrain later outcomes and where effects at one checkpoint propagate through subsequent stages.
Early decisions made in the nucleus set the stage for what follows. U1-guided protection and 3′-end choice shape isoforms; TREX and the exon junction complex link splicing and 3′-end formation to export competence; export routes can switch during stress; and post-export remodeling, including 3′UTR cleavage, can change local availability. Together, these events carry forward to influence cytoplasmic fate.
In the cytoplasm, localization and translation are often coupled. Transport factors can hold ribosomes in check during transit and release them with activity. Decay frequently depends on translation status. The SNCA example shows how more than one RBP can coordinate processing, export, stability, localization, ribosome engagement, and protein output at a single locus. Chemical modifications and noncoding RNAs add context-dependent layers that tune these relationships.
Two considerations are central. CLIP occupancy alone does not prove function, and effects are commonly isoform-specific and cell-type-specific. The practical answer is to pair defined perturbations with isoform-aware, subcellularly resolved, ribosome-informed readouts in at least two neuronal backgrounds, and to repeat the panel under relevant stress conditions. This approach moves work from isolated events to integrated lifecycle control, encouraging experiments that follow the same RNA across regulatory stages within defined cellular contexts.
Author contributions
ED: Conceptualization, Writing – original draft, Writing – review & editing, Visualization. YX: Writing – review & editing. AFS: Writing – review & editing, Visualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alcott, C. E., Yalamanchili, H. K., Ji, P., van der Heijden, M. E., Saltzman, A., Elrod, N., et al. (2020). Partial loss of CFIm25 causes learning deficits and aberrant neuronal alternative polyadenylation. Elife 9:e50895. doi: 10.7554/eLife.50895
Allemand, E., Guil, S., Myers, M. P., Moscat, J., Caceres, J. F., and Krainer, A. R. (2005). Regulation of heterogenous nuclear ribonucleoprotein A1 transport by phosphorylation in cells stressed by osmotic shock. Proc. Natl. Acad. Sci. U. S. A. 102, 3605–3610. doi: 10.1073/pnas.0409889102
An, J. J., Gharami, K., Liao, G. Y., Woo, N. H., Lau, A. G., Vanevski, F., et al. (2008). Distinct role of long 3' UTR BDNF mRNA in spine morphology and synaptic plasticity in hippocampal neurons. Cell 134, 175–187. doi: 10.1016/j.cell.2008.05.045
Andreassi, C., Luisier, R., Crerar, H., Darsinou, M., Blokzijl-Franke, S., Lenn, T., et al. (2021). Cytoplasmic cleavage of IMPA1 3' UTR is necessary for maintaining axon integrity. Cell Rep. 34:108778. doi: 10.1016/j.celrep.2021.108778
Arimoto-Matsuzaki, K., Saito, H., and Takekawa, M. (2016). TIA1 oxidation inhibits stress granule assembly and sensitizes cells to stress-induced apoptosis. Nat. Commun. 7:10252. doi: 10.1038/ncomms10252
Barrandon, C., Bonnet, F., Nguyen, V. T., Labas, V., and Bensaude, O. (2007). The transcription-dependent dissociation of P-TEFb-HEXIM1-7SK RNA relies upon formation of hnRNP-7SK RNA complexes. Mol. Cell. Biol. 27, 6996–7006. doi: 10.1128/MCB.00975-07
Baumann, S. J., Grawenhoff, J., Rodrigues, E. C., Speroni, S., Gili, M., Komissarov, A., et al. (2022). APC couples neuronal mRNAs to multiple kinesins, EB1, and shrinking microtubule ends for bidirectional mRNA motility. Proc. Natl. Acad. Sci. U. S. A. 119:e2211536119. doi: 10.1073/pnas.2211536119
Begik, O., Lucas, M. C., Pryszcz, L. P., Ramirez, J. M., Medina, R., Milenkovic, I., et al. (2021). Quantitative profiling of pseudouridylation dynamics in native RNAs with nanopore sequencing. Nat. Biotechnol. 39, 1278–1291. doi: 10.1038/s41587-021-00915-6
Bensaude, O. (2011). Inhibiting eukaryotic transcription: which compound to choose? How to evaluate its activity? Transcription 2, 103–108. doi: 10.4161/trns.2.3.16172
Berg, M. G., Singh, L. N., Younis, I., Liu, Q., Pinto, A. M., Kaida, D., et al. (2012). U1 snRNP determines mRNA length and regulates isoform expression. Cell 150, 53–64. doi: 10.1016/j.cell.2012.05.029
Bhattacharyya, S. N., Habermacher, R., Martine, U., Closs, E. I., and Filipowicz, W. (2006). Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell 125, 1111–1124. doi: 10.1016/j.cell.2006.04.031
Bilican, B., Serio, A., Barmada, S. J., Nishimura, A. L., Sullivan, G. J., Carrasco, M., et al. (2012). Mutant induced pluripotent stem cell lines recapitulate aspects of TDP-43 proteinopathies and reveal cell-specific vulnerability. Proc. Natl. Acad. Sci. U. S. A. 109, 5803–5808. doi: 10.1073/pnas.1202922109
Booeshaghi, A. S., Yao, Z., van Velthoven, C., Smith, K., Tasic, B., Zeng, H., et al. (2021). Isoform cell-type specificity in the mouse primary motor cortex. Nature 598, 195–199. doi: 10.1038/s41586-021-03969-3
Brangwynne, C. P., Eckmann, C. R., Courson, D. S., Rybarska, A., Hoege, C., Gharakhani, J., et al. (2009). Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 324, 1729–1732. doi: 10.1126/science.1172046
Brothers, W. R., Ali, F., Kajjo, S., and Fabian, M. R. (2023). The EDC4-XRN1 interaction controls P-body dynamics to link mRNA decapping with decay. EMBO J. 42:e113933. doi: 10.15252/embj.2023113933
Cabaj, M., Mazzara, P. G., Gaertner, Z. A., Wang, R., Pauers, M. M., Randolph, L. K., et al. (2025). Alpha-synuclein abundance and localization are regulated by the RNA-binding protein PUMILIO1. Cell Rep. 44:116145. doi: 10.1016/j.celrep.2025.116145
Cajigas, I. J., Tushev, G., Will, T. J., tom Dieck, S., Fuerst, N., and Schuman, E. M. (2012). The local transcriptome in the synaptic neuropil revealed by deep sequencing and high-resolution imaging. Neuron 74, 453–466. doi: 10.1016/j.neuron.2012.02.036
Carlile, T. M., Rojas-Duran, M. F., Zinshteyn, B., Shin, H., Bartoli, K. M., and Gilbert, W. V. (2014). Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 515, 143–146. doi: 10.1038/nature13802
Chang, C. T., Bercovich, N., Loh, B., Jonas, S., and Izaurralde, E. (2014). The activation of the decapping enzyme DCP2 by DCP1 occurs on the EDC4 scaffold and involves a conserved loop in DCP1. Nucleic Acids Res. 42, 5217–5233. doi: 10.1093/nar/gku129
Chasse, H., Boulben, S., Costache, V., Cormier, P., and Morales, J. (2017). Analysis of translation using polysome profiling. Nucleic Acids Res. 45:e15. doi: 10.1093/nar/gkw907
Chen, K. H., Boettiger, A. N., Moffitt, J. R., Wang, S., and Zhuang, X. (2015). RNA imaging: spatially resolved, highly multiplexed RNA profiling in single cells. Science 348:aaa6090. doi: 10.1126/science.aaa6090
Chen, R. X., Chen, X., Xia, L. P., Zhang, J. X., Pan, Z. Z., Ma, X. D., et al. (2019). N(6)-methyladenosine modification of circNSUN2 facilitates cytoplasmic export and stabilizes HMGA2 to promote colorectal liver metastasis. Nat. Commun. 10:4695. doi: 10.1038/s41467-019-12651-2
Choi, H. M. T., Beck, V. A., and Pierce, N. A. (2014). Next-generation in situ hybridization chain reaction: higher gain, lower cost, greater durability. ACS Nano 8, 4284–4294. doi: 10.1021/nn405717p
Cohen, B., Altman, T., Golani-Armon, A., Savulescu, A. F., Ibraheem, A., Mhlanga, M. M., et al. (2022). Co-transport of the nuclear-encoded Cox7c mRNA with mitochondria along axons occurs through a coding-region-dependent mechanism. J. Cell Sci. 135:jcs259436. doi: 10.1242/jcs.259436
Corley, M., Burns, M. C., and Yeo, G. W. (2020). How RNA-binding proteins interact with RNA: molecules and mechanisms. Mol. Cell 78, 9–29. doi: 10.1016/j.molcel.2020.03.011
Dai, W., Zhang, G., and Makeyev, E. V. (2012). RNA-binding protein HuR autoregulates its expression by promoting alternative polyadenylation site usage. Nucleic Acids Res. 40, 787–800. doi: 10.1093/nar/gkr783
Darnell, J. C., Van Driesche, S. J., Zhang, C., Hung, K. Y. S., Mele, A., Fraser, C. E., et al. (2011). FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell 146, 247–261. doi: 10.1016/j.cell.2011.06.013
Devlin, A. C., Burr, K., Borooah, S., Foster, J. D., Cleary, E. M., Geti, I., et al. (2015). Human iPSC-derived motoneurons harbouring TARDBP or C9ORF72 ALS mutations are dysfunctional despite maintaining viability. Nat. Commun. 6:5999. doi: 10.1038/ncomms6999
Engel, K. L., Arora, A., Goering, R., Lo, H. G., and Taliaferro, J. M. (2020). Mechanisms and consequences of subcellular RNA localization across diverse cell types. Traffic 21, 404–418. doi: 10.1111/tra.12730
Eom, T., Antar, L. N., Singer, R. H., and Bassell, G. J. (2003). Localization of a beta-actin messenger ribonucleoprotein complex with zipcode-binding protein modulates the density of dendritic filopodia and filopodial synapses. J. Neurosci. 23, 10433–10444. doi: 10.1523/JNEUROSCI.23-32-10433.2003
Fonseca, B. D., Zakaria, C., Jia, J. J., Graber, T. E., Svitkin, Y., Tahmasebi, S., et al. (2015). La-related Protein 1 (LARP1) represses terminal oligopyrimidine (TOP) mRNA translation downstream of mTOR complex 1 (mTORC1). J. Biol. Chem. 290, 15996–16020. doi: 10.1074/jbc.M114.621730
Fragkouli, A., Koukouraki, P., Vlachos, I. S., Paraskevopoulou, M. D., Hatzigeorgiou, A. G., and Doxakis, E. (2017). Neuronal ELAVL proteins utilize AUF-1 as a co-partner to induce neuron-specific alternative splicing of APP. Sci. Rep. 7:44507. doi: 10.1038/srep44507
Gallouzi, I. E., Brennan, C. M., and Steitz, J. A. (2001). Protein ligands mediate the CRM1-dependent export of HuR in response to heat shock. RNA 7, 1348–1361. doi: 10.1017/S1355838201016089
Garalde, D. R., Snell, E. A., Jachimowicz, D., Sipos, B., Lloyd, J. H., Bruce, M., et al. (2018). Highly parallel direct RNA sequencing on an array of nanopores. Nat. Methods 15, 201–206. doi: 10.1038/nmeth.4577
Geisberg, J. V., Moqtaderi, Z., and Struhl, K. (2020). The transcriptional elongation rate regulates alternative polyadenylation in yeast. Elife 9:e59810. doi: 10.7554/eLife.59810
Goering, R., Hudish, L. I., Guzman, B. B., Raj, N., Bassell, G. J., Russ, H. A., et al. (2020). FMRP promotes RNA localization to neuronal projections through interactions between its RGG domain and G-quadruplex RNA sequences. Elife 9:e52621. doi: 10.7554/eLife.52621
Gong, C., and Maquat, L. E. (2011). lncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3' UTRs via Alu elements. Nature 470, 284–288. doi: 10.1038/nature09701
Gourdomichali, O., Zonke, K., Kattan, F. G., Makridakis, M., Kontostathi, G., Vlahou, A., et al. (2022). In situ peroxidase labeling followed by mass-spectrometry reveals TIA1 interactome. Biology 11:287. doi: 10.3390/biology11020287
Guil, S., Long, J. C., and Caceres, J. F. (2006). hnRNP A1 relocalization to the stress granules reflects a role in the stress response. Mol. Cell. Biol. 26, 5744–5758. doi: 10.1128/MCB.00224-06
Harbauer, A. B., Hees, J. T., Wanderoy, S., Segura, I., Gibbs, W., Cheng, Y., et al. (2022). Neuronal mitochondria transport Pink1 mRNA via synaptojanin 2 to support local mitophagy. Neuron 110, 1516–1531.e9. doi: 10.1016/j.neuron.2022.01.035
Harding, H. P., Novoa, I., Zhang, Y., Zeng, H., Wek, R., Schapira, M., et al. (2000). Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 6, 1099–1108. doi: 10.1016/S1097-2765(00)00108-8
Heiman, M., Kulicke, R., Fenster, R. J., Greengard, P., and Heintz, N. (2014). Cell type-specific mRNA purification by translating ribosome affinity purification (TRAP). Nat. Protoc. 9, 1282–1291. doi: 10.1038/nprot.2014.085
Herzog, V. A., Reichholf, B., Neumann, T., Rescheneder, P., Bhat, P., Burkard, T. R., et al. (2017). Thiol-linked alkylation of RNA to assess expression dynamics. Nat. Methods 14, 1198–1204. doi: 10.1038/nmeth.4435
Higelin, J., Demestre, M., Putz, S., Delling, J. P., Jacob, C., Lutz, A. K., et al. (2016). FUS mislocalization and vulnerability to DNA damage in ALS patients derived hiPSCs and aging motoneurons. Front. Cell. Neurosci. 10:290. doi: 10.3389/fncel.2016.00290
Hilgers, V., Lemke, S. B., and Levine, M. (2012). ELAV mediates 3' UTR extension in the Drosophila nervous system. Genes Dev. 26, 2259–2264. doi: 10.1101/gad.199653.112
Hu, P., Fabyanic, E., Kwon, D. Y., Tang, S., Zhou, Z., and Wu, H. (2017). Dissecting cell-type composition and activity-dependent transcriptional state in mammalian brains by massively parallel single-nucleus RNA-Seq. Mol. Cell 68, 1006–1015.e1007. doi: 10.1016/j.molcel.2017.11.017
Janicke, A., Vancuylenberg, J., Boag, P. R., Traven, A., and Beilharz, T. H. (2012). ePAT: a simple method to tag adenylated RNA to measure poly(A)-tail length and other 3' RACE applications. RNA 18, 1289–1295. doi: 10.1261/rna.031898.111
Kaida, D., Berg, M. G., Younis, I., Kasim, M., Singh, L. N., Wan, L., et al. (2010). U1 snRNP protects pre-mRNAs from premature cleavage and polyadenylation. Nature 468, 664–668. doi: 10.1038/nature09479
Karijolich, J., and Yu, Y. T. (2011). Converting nonsense codons into sense codons by targeted pseudouridylation. Nature 474, 395–398. doi: 10.1038/nature10165
Kattan, F. G., Koukouraki, P., Anagnostopoulos, A. K., Tsangaris, G. T., and Doxakis, E. (2023). RNA binding protein AUF1/HNRNPD regulates nuclear export, stability and translation of SNCA transcripts. Open Biol. 13:230158. doi: 10.1098/rsob.230158
Kedersha, N. L., Gupta, M., Li, W., Miller, I., and Anderson, P. (1999). RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha to the assembly of mammalian stress granules. J. Cell Biol. 147, 1431–1442. doi: 10.1083/jcb.147.7.1431
Kiltschewskij, D. J., Harrison, P. F., Fitzsimmons, C., Beilharz, T. H., and Cairns, M. J. (2023). Extension of mRNA poly(A) tails and 3'UTRs during neuronal differentiation exhibits variable association with post-transcriptional dynamics. Nucleic Acids Res. 51, 8181–8198. doi: 10.1093/nar/gkad499
Kim, S., Kim, S., Chang, H. R., Kim, D., Park, J., Son, N., et al. (2021). The regulatory impact of RNA-binding proteins on microRNA targeting. Nat. Commun. 12:5057. doi: 10.1038/s41467-021-25078-5
Koukouraki, P., and Doxakis, E. (2016). Constitutive translation of human alpha-synuclein is mediated by the 5'-untranslated region. Open Biol. 6:160022. doi: 10.1098/rsob.160022
Lau, A. G., Irier, H. A., Gu, J., Tian, D., Ku, L., Liu, G., et al. (2010). Distinct 3'UTRs differentially regulate activity-dependent translation of brain-derived neurotrophic factor (BDNF). Proc. Natl. Acad. Sci. U. S. A. 107, 15945–15950. doi: 10.1073/pnas.1002929107
Le Hir, H., Izaurralde, E., Maquat, L. E., and Moore, M. J. (2000). The spliceosome deposits multiple proteins 20-24 nucleotides upstream of mRNA exon-exon junctions. EMBO J. 19, 6860–6869. doi: 10.1093/emboj/19.24.6860
Leal, G., Afonso, P. M., and Duarte, C. B. (2014). Neuronal activity induces synaptic delivery of hnRNP A2/B1 by a BDNF-dependent mechanism in cultured hippocampal neurons. PLoS ONE 9:e108175. doi: 10.1371/journal.pone.0108175
Lee, J. A., Damianov, A., Lin, C. H., Fontes, M., Parikshak, N. N., Anderson, E. S., et al. (2016). Cytoplasmic Rbfox1 regulates the expression of synaptic and autism-related genes. Neuron 89, 113–128. doi: 10.1016/j.neuron.2015.11.025
Licatalosi, D. D., Mele, A., Fak, J. J., Ule, J., Kayikci, M., Chi, S. W., et al. (2008). HITS-CLIP yields genome-wide insights into brain alternative RNA processing. Nature 456, 464–469. doi: 10.1038/nature07488
Lima, S. A., Chipman, L. B., Nicholson, A. L., Chen, Y. H., Yee, B. A., Yeo, G. W., et al. (2017). Short poly(A) tails are a conserved feature of highly expressed genes. Nat. Struct. Mol. Biol. 24, 1057–1063. doi: 10.1038/nsmb.3499
Lin, Y., Zheng, J., Mai, Z., Lin, P., Lu, Y., Cui, L., et al. (2024). Unveiling the veil of RNA binding protein phase separation in cancer biology and therapy. Cancer Lett. 601:217160. doi: 10.1016/j.canlet.2024.217160
Liu, N., Dai, Q., Zheng, G., He, C., Parisien, M., and Pan, T. (2015). N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature 518, 560–564. doi: 10.1038/nature14234
Liu, X., Xie, H., Liu, W., Zuo, J., Li, S., Tian, Y., et al. (2024). Dynamic regulation of alternative polyadenylation by PQBP1 during neurogenesis. Cell Rep. 43:114525. doi: 10.1016/j.celrep.2024.114525
Mansfield, K. D., and Keene, J. D. (2012). Neuron-specific ELAV/Hu proteins suppress HuR mRNA during neuronal differentiation by alternative polyadenylation. Nucleic Acids Res. 40, 2734–2746. doi: 10.1093/nar/gkr1114
Miura, P., Shenker, S., Andreu-Agullo, C., Westholm, J. O., and Lai, E. C. (2013). Widespread and extensive lengthening of 3' UTRs in the mammalian brain. Genome Res. 23, 812–825. doi: 10.1101/gr.146886.112
Nishikawa, T., Kuwano, Y., Takahara, Y., Nishida, K., and Rokutan, K. (2019). HnRNPA1 interacts with G-quadruplex in the TRA2B promoter and stimulates its transcription in human colon cancer cells. Sci. Rep. 9:10276. doi: 10.1038/s41598-019-46659-x
Oktaba, K., Zhang, W., Lotz, T. S., Jun, D. J., Lemke, S. B., Ng, S. P., et al. (2015). ELAV links paused Pol II to alternative polyadenylation in the Drosophila nervous system. Mol. Cell 57, 341–348. doi: 10.1016/j.molcel.2014.11.024
Paschou, M., Paraskevopoulou, M. D., Vlachos, I. S., Koukouraki, P., Hatzigeorgiou, A. G., and Doxakis, E. (2012). miRNA regulons associated with synaptic function. PLoS One 7:e46189. doi: 10.1371/journal.pone.0046189
Patel, V. L., Mitra, S., Harris, R., Buxbaum, A. R., Lionnet, T., Brenowitz, M., et al. (2012). Spatial arrangement of an RNA zipcode identifies mRNAs under post-transcriptional control. Genes Dev. 26, 43–53. doi: 10.1101/gad.177428.111
Philippe, L., van den Elzen, A. M. G., Watson, M. J., and Thoreen, C. C. (2020). Global analysis of LARP1 translation targets reveals tunable and dynamic features of 5' TOP motifs. Proc. Natl. Acad. Sci. U. S. A. 117, 5319–5328. doi: 10.1073/pnas.1912864117
Puhringer, T., Hohmann, U., Fin, L., Pacheco-Fiallos, B., Schellhaas, U., Brennecke, J., et al. (2020). Structure of the human core transcription-export complex reveals a hub for multivalent interactions. Elife 9:e61503. doi: 10.7554/eLife.61503
Raj, A., van den Bogaard, P., Rifkin, S. A., van Oudenaarden, A., and Tyagi, S. (2008). Imaging individual mRNA molecules using multiple singly labeled probes. Nat. Methods 5, 877–879. doi: 10.1038/nmeth.1253
Rodriques, S. G., Stickels, R. R., Goeva, A., Martin, C. A., Murray, E., Vanderburg, C. R., et al. (2019). Slide-seq: a scalable technology for measuring genome-wide expression at high spatial resolution. Science 363, 1463–1467. doi: 10.1126/science.aaw1219
Safra, M., Nir, R., Farouq, D., Vainberg Slutskin, I., and Schwartz, S. (2017). TRUB1 is the predominant pseudouridine synthase acting on mammalian mRNA via a predictable and conserved code. Genome Res. 27, 393–406. doi: 10.1101/gr.207613.116
Sanz, E., Yang, L., Su, T., Morris, D. R., McKnight, G. S., and Amieux, P. S. (2009). Cell-type-specific isolation of ribosome-associated mRNA from complex tissues. Proc. Natl. Acad. Sci. U. S. A. 106, 13939–13944. doi: 10.1073/pnas.0907143106
Schmidt, E. K., Clavarino, G., Ceppi, M., and Pierre, P. (2009). SUnSET, a nonradioactive method to monitor protein synthesis. Nat. Methods 6, 275–277. doi: 10.1038/nmeth.1314
Schofield, J. A., Duffy, E. E., Kiefer, L., Sullivan, M. C., and Simon, M. D. (2018). TimeLapse-seq: adding a temporal dimension to RNA sequencing through nucleoside recoding. Nat. Methods 15, 221–225. doi: 10.1038/nmeth.4582
Shan, J., Munro, T. P., Barbarese, E., Carson, J. H., and Smith, R. (2003). A molecular mechanism for mRNA trafficking in neuronal dendrites. J. Neurosci. 23, 8859–8866. doi: 10.1523/JNEUROSCI.23-26-08859.2003
Shi, H., Zhang, X., Weng, Y. L., Lu, Z., Liu, Y., Lu, Z., et al. (2018). m(6)A facilitates hippocampus-dependent learning and memory through YTHDF1. Nature 563, 249–253. doi: 10.1038/s41586-018-0666-1
So, B. R., Di, C., Cai, Z., Venters, C. C., Guo, J., Oh, J. M., et al. (2019). A complex of U1 snRNP with cleavage and polyadenylation factors controls telescripting, regulating mRNA transcription in human cells. Mol. Cell 76, 590–599.e4. doi: 10.1016/j.molcel.2019.08.007
Subtelny, A. O., Eichhorn, S. W., Chen, G. R., Sive, H., and Bartel, D. P. (2014). Poly(A)-tail profiling reveals an embryonic switch in translational control. Nature 508, 66–71. doi: 10.1038/nature13007
Surgucheva, I., Gunewardena, S., Rao, H. S., and Surguchov, A. (2013). Cell-specific post-transcriptional regulation of gamma-synuclein gene by micro-RNAs. PLoS ONE 8:e73786. doi: 10.1371/journal.pone.0073786
Tani, H., Mizutani, R., Salam, K. A., Tano, K., Ijiri, K., Wakamatsu, A., et al. (2012). Genome-wide determination of RNA stability reveals hundreds of short-lived noncoding transcripts in mammals. Genome Res. 22, 947–956. doi: 10.1101/gr.130559.111
Taylor, A. M., Blurton-Jones, M., Rhee, S. W., Cribbs, D. H., Cotman, C. W., and Jeon, N. L. (2005). A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2, 599–605. doi: 10.1038/nmeth777
Taylor, A. M., Dieterich, D. C., Ito, H. T., Kim, S. A., and Schuman, E. M. (2010). Microfluidic local perfusion chambers for the visualization and manipulation of synapses. Neuron 66, 57–68. doi: 10.1016/j.neuron.2010.03.022
Tian, R., Gachechiladze, M. A., Ludwig, C. H., Laurie, M. T., Hong, J. Y., Nathaniel, D., et al. (2019). CRISPR interference-based platform for multimodal genetic screens in human iPSC-derived neurons. Neuron 104, 239–255.e212. doi: 10.1016/j.neuron.2019.07.014
Tushev, G., Glock, C., Heumuller, M., Biever, A., Jovanovic, M., and Schuman, E. M. (2018). Alternative 3' UTRs Modify the Localization, Regulatory Potential, Stability, and Plasticity of mRNAs in Neuronal Compartments. Neuron 98, 495–511.e6. doi: 10.1016/j.neuron.2018.03.030
Uchida, N., Hoshino, S., and Katada, T. (2004). Identification of a human cytoplasmic poly(A) nuclease complex stimulated by poly(A)-binding protein. J. Biol. Chem. 279, 1383–1391. doi: 10.1074/jbc.M309125200
Vattem, K. M., and Wek, R. C. (2004). Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc. Natl. Acad. Sci. U. S. A. 101, 11269–11274. doi: 10.1073/pnas.0400541101
Vicario, A., Colliva, A., Ratti, A., Davidovic, L., Baj, G., Gricman, L., et al. (2015). Dendritic targeting of short and long 3' UTR BDNF mRNA is regulated by BDNF or NT-3 and distinct sets of RNA-binding proteins. Front. Mol. Neurosci. 8:62. doi: 10.3389/fnmol.2015.00062
Viphakone, N., Hautbergue, G. M., Walsh, M., Chang, C. T., Holland, A., Folco, E. G., et al. (2012). TREX exposes the RNA-binding domain of Nxf1 to enable mRNA export. Nat. Commun. 3:1006. doi: 10.1038/ncomms2005
Viphakone, N., Sudbery, I., Griffith, L., Heath, C. G., Sims, D., and Wilson, S. A. (2019). Co-transcriptional loading of RNA export factors shapes the human transcriptome. Mol. Cell 75, 310–323.e318. doi: 10.1016/j.molcel.2019.04.034
Wang, F., Flanagan, J., Su, N., Wang, L. C., Bui, S., Nielson, A., et al. (2012). RNAscope: a novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissues. J. Mol. Diagn. 14, 22–29. doi: 10.1016/j.jmoldx.2011.08.002
Westmark, P. R., Westmark, C. J., Jeevananthan, A., and Malter, J. S. (2011). Preparation of synaptoneurosomes from mouse cortex using a discontinuous percoll-sucrose density gradient. J. Vis. Exp. (55): e3196. doi: 10.3791/3196-v
Weyn-Vanhentenryck, S. M., Mele, A., Yan, Q., Sun, S., Farny, N., Zhang, Z., et al. (2014). HITS-CLIP and integrative modeling define the Rbfox splicing-regulatory network linked to brain development and autism. Cell Rep. 6, 1139–1152. doi: 10.1016/j.celrep.2014.02.005
Workman, R. E., Tang, A. D., Tang, P. S., Jain, M., Tyson, J. R., Razaghi, R., et al. (2019). Nanopore native RNA sequencing of a human poly(A) transcriptome. Nat. Methods 16, 1297–1305. doi: 10.1038/s41592-019-0617-2
Yague-Sanz, C., Vanrobaeys, Y., Fernandez, R., Duval, M., Larochelle, M., Beaudoin, J., et al. (2020). Nutrient-dependent control of RNA polymerase II elongation rate regulates specific gene expression programs by alternative polyadenylation. Genes Dev. 34, 883–897. doi: 10.1101/gad.337212.120
Yang, X., Yang, Y., Sun, B. F., Chen, Y. S., Xu, J. W., Lai, W. Y., et al. (2017). 5-methylcytosine promotes mRNA export - NSUN2 as the methyltransferase and ALYREF as an m(5)C reader. Cell Res. 27, 606–625. doi: 10.1038/cr.2017.55
Yi, H., Park, J., Ha, M., Lim, J., Chang, H., and Kim, V. N. (2018). PABP cooperates with the CCR4-NOT complex to promote mRNA deadenylation and block precocious decay. Mol. Cell 70, 1081–1088.e1085. doi: 10.1016/j.molcel.2018.05.009
Yoon, J. H., and Gorospe, M. (2016). “Cross-linking immunoprecipitation and qPCR (CLIP-qPCR) analysis to map interactions between long noncoding RNAs and RNA-binding proteins,” in Methods in Molecular Biology, Vol. 1402 (New York, NY: Humana Press) 11–17.
Keywords: RNA-binding proteins, transcript life cycle, alternative polyadenylation, RNA modifications, mRNA localization, local translation, stress
Citation: Doxakis E, Xue YC and Savulescu AF (2025) One RNA-binding protein, many decisions: integrating the transcript life cycle into neuronal regulation. Front. Mol. Neurosci. 18:1716825. doi: 10.3389/fnmol.2025.1716825
Received: 30 September 2025; Accepted: 29 October 2025;
Published: 12 November 2025.
Edited by:
Andrei Surguchov, University of Kansas Medical Center, United StatesReviewed by:
Mengting Niu, University of Electronic Science and Technology of China, ChinaCopyright © 2025 Doxakis, Xue and Savulescu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Epaminondas Doxakis, ZWRveGFraXNAYmlvYWNhZGVteS5ncg==