 Maura Rojas-Pirela1,2,3
Maura Rojas-Pirela1,2,3 Lisvaneth Medina1
Lisvaneth Medina1 Maria Verónica Rojas2
Maria Verónica Rojas2 Ana Isabel Liempi1
Ana Isabel Liempi1 Christian Castillo1,4Elizabeth Pérez-Pérez3Jesús Guerrero-Muñoz1
Christian Castillo1,4Elizabeth Pérez-Pérez3Jesús Guerrero-Muñoz1 Sebastian Araneda1,5
Sebastian Araneda1,5 Ulrike Kemmerling1*
Ulrike Kemmerling1*- 1Instituto de Ciencias Biomédicas, Facultad de Medicina, Universidad de Chile, Santiago, Chile
- 2Instituto de Biología, Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
- 3Facultad de Farmacia y Bioanálisis, Universidad de Los Andes, Mérida, Venezuela
- 4Núcleo de Investigación Aplicada en Ciencias Veterinarias y Agronómicas, Facultad de Medicina Veterinaria y Agronomía, Universidad de Las Américas, Santiago, Chile
- 5Facultad de Salud y Odontología, Universidad Diego Portales, Santiago, Chile
Apicomplexans are a group of pathogenic protists that cause various diseases in humans and animals that cause economic losses worldwide. These unicellular eukaryotes are characterized by having a complex life cycle and the ability to evade the immune system of their host organism. Infections caused by some of these parasites affect millions of pregnant women worldwide, leading to various adverse maternal and fetal/placental effects. Unfortunately, the exact pathogenesis of congenital apicomplexan diseases is far from being understood, including the mechanisms of how they cross the placental barrier. In this review, we highlight important aspects of the diseases caused by species of Plasmodium, Babesia, Toxoplasma, and Neospora, their infection during pregnancy, emphasizing the possible role played by the placenta in the host-pathogen interaction.
Introduction
The phylum Apicomplexa constitutes a broad group of parasitic protists that include more than 6000 species (Votýpka et al., 2017), distributed in a wide diversity of environments, including soil, freshwater, and marine habitats (del Campo et al., 2019). Many of these parasites have significant clinical and economic relevance since they cause important human and veterinary diseases worldwide. In this context, malaria (Plasmodium species), babesiosis (Babesia sp.), toxoplasmosis (Toxoplasma gondii), neosporosis (Neospora sp.), and cryptosporidiosis (Cryptosporidium parvum) are diseases caused by apicomplexan parasites (Chakraborty et al., 2017; Martínez-Ocampo, 2018).
All those parasites present complex life cycles, infecting one or more hosts and undergoing multiple morphological and metabolic differentiation (Martínez-Ocampo, 2018; Jeninga et al., 2019). Additionally, in the life cycles of most of the apicomplexan, an alternation between asexual and sexual reproduction can be observed. Thus, in the different life cycles, three different states can be observed: sporozoite (infective stage), merozoite (a result of asexual reproduction), and gametocyte (germ cells) (Votýpka et al., 2017).
Apicomplexan parasites are characterized by having a complex of apical secretory organelles composed of micronemes, rhoptries, and dense granules. These organelles vary between species and are involved in host cell attachment, motility, invasion, and intracellular parasitophorous vacuole formation (Votýpka et al., 2017). In addition, all apicomplexan examined so far (except Cryptosporidium and Gregarine) possess an apicoplast (Salomaki and Kolisko, 2019), a relict plastid involved in several essential metabolic pathways, such as fatty acid, isoprenoid, and heme biosynthesis, that are crucial for parasite replication and establishment of infection (Ralph et al., 2004; Chakraborty, 2016).
Apicomplexan presents different transmission routes (Votýpka et al., 2017). However, for some species such as Neospora, Babesia microti (B. microti), and Toxoplasma gondii (T. gondii), the infection from mother to fetus constitutes an important transmission route (Joseph et al., 2012; Hide, 2016; Khan et al., 2020). The incidence of congenital malaria, caused by Plasmodium, is relatively low in endemic regions. However, some reports have documented that the incidence can reach 37% (Ouédraogo et al., 2012; Olupot-Olupot et al., 2018). Regarding T. gondii, when primo-infection occurs during pregnancy, the congenital transmission rates are high (Hide, 2016), increasing from the first to the third trimester of pregnancy (McAuley, 2014). Importantly, these congenital infections can be detrimental to maternal and child health (Maldonado et al., 2017; Tegegne et al., 2019). The frequency of the transplacental transmission of these parasites is associated with different factors such as maternal and developing fetus immune systems and genetic background (Ortiz-Alegría et al., 2010; Ouédraogo et al., 2012; Gómez-Chávez et al., 2019), the passive transfer of maternal antibodies (Ortiz-Alegría et al., 2010; Dobbs and Dent, 2016) characteristics of the fetal environment (De Silva et al., 1982), and the functioning of the placenta as a physical barrier and immune organ (Robbins et al., 2012; Sharma and Shukla, 2017). Thus, the placenta is a crucial organ determining the probability of infection (Castillo et al., 2018).
The present review is focused on the congenital transmission of Plasmodium, Babesia, T. gondii, Neospora and the role of the placenta.
Diseases Caused by Apicomplexan Parasites: Social and Economic Impact
Diseases caused by apicomplexan parasites affect millions of people, mainly in low- and middle-income countries located in the tropical and subtropical regions (Votýpka et al., 2017). Moreover, some of those infections have been included in the group of Neglected Tropical Diseases (NTD) (PLoS Neglected Tropical Disease, 2020), implying a significant social and economic impact that can reach millions of dollars each year (Laha et al., 2015; Ozawa et al., 2019; Stelzer et al., 2019).
Malaria is caused by five members of the genus Plasmodium, where P. falciparum and P. vivax species represent the most significant threat (Geleta and Ketema, 2016). Malaria is an important public health problem; it was estimated in 2018 that 228 million people were infected worldwide, and 405,000 malaria deaths were reported (WHO, 2019). Children aged under 5 years and pregnant women constitute the most vulnerable population (Gabrielli et al., 2016; WHO, 2019). The attempts to control and eradicate malaria present an estimated economic impact of 2.7 billion dollars (WHO, 2019).
Babesiosis is a globally distributed disease with malaria-like symptoms that affects elderly and immunocompromised patients (Kim et al., 2016), caused by erythrocytic parasites of the genus Babesia. Many of the 100 species of Babesia constitute a significant threat to humans, domestic animals, and livestock (Young et al., 2019), being in humans, B. microti, B. divergens, B. duncani, and B. venatorum the causal agents (Sun et al., 2014; Young et al., 2019). The mortality rate of this disease is approximately 5%; however, in case of infection through blood transfusion, the mortality rate increases to 19% (Katz and Dodd, 2019). In addition to human health effects, some Babesia species cause a significant loss to the cattle industry due to death, a loss of beef production of infected animals, and death (Mosqueda et al., 2012).
Toxoplasmosis is a zoonotic disease that affects approximately one-third of the world population (Ngô et al., 2017; Stelzer et al., 2019). The infection in immunocompetent individuals is generally asymptomatic; however, severe symptoms are observed in newborns (i.e., mental retardation, ocular disease) when the primoinfection occurs in the pregnant mother. Additionally, infection during pregnancy can cause abortion (Tegegne et al., 2016). Toxoplasmosis can be fatal in immunosuppressed patients since the reactivation of latent infection can lead to the development of encephalitis and, in some cases, reactivation of malignancies (Maciel et al., 2000; Wang Z.D. et al., 2017). In the case of animals, it is considered one of the leading causes of reproductive losses in small ruminants worldwide, which also play an important role in transmitting the parasite to humans. In some countries, such as Australia, the losses attributed to this disease can be up to 70 million dollars (Stelzer et al., 2019).
Neosporosis is a devastating worldwide disease responsible for abortions, neonatal mortality, and central nervous system diseases in animals. It is mainly caused by the species Neospora caninum (N. caninum) and Neospora hughesi (N. hughesi) and is transmitted by horizontal and vertical routes (Dubey, 2003; Donahoe et al., 2015). N. caninum affects dogs and cattle but occasionally infects horses, sheep, and deer, while N. hughesi only infects horses causing Equine Protozoal Myeloencephalitis (Dubey, 2003; Wobeser et al., 2009).
Neosporosis is associated with sporadic abortions (between 10 and 12.5%) in cattle herds with a frequent congenital transmission. However, the percentage of abortions increases (30–57%) in herds when the parasite is acquired by the pregnant mother (Dubey et al., 2007). N. caninum infections have a significant global economic impact; for instance, in New Zealand, the losses attributed to this disease exceed US$ 35.7 million (Reichel et al., 2013).
Host–Apicomplexan Interactions at the Mammalian Placenta
As mentioned above, apicomplexan parasites can be transmitted congenitally. Within this group Plasmodium spp. and T. gondii are the most documented (Carlier et al., 2012). The placenta is a transitory organ that acts as the interface between the mother and fetus, which mediates nutrition and gas exchange between the fetus and the mother, ensuring growth and normal embryo-fetal development and supporting the maternal changes associated with pregnancy (Griffiths and Campbell, 2015; Castillo et al., 2018). The chorioallantoic is the main placenta in mammals during middle to late gestation and develops from the endometrium of the uterus and the trophoblast of the embryo. According to the extent of trophoblast invasion into the uterus, placentation is classified into hemochorial (highly invasive), endotheliochorial (moderate invasive) or epitheliochorial (low invasive) (Liempi et al., 2020). In the case of the human placenta, it is classified as discoidal, villous, and hemochorial, made up of a fetal portion that (originates from the Chorion frondosum) and a maternal portion or decidua basalis (originates from the endometrium) (Castillo et al., 2018).
The functional units in the human placenta are the chorionic villi, formed by the trophoblast a lining epithelium formed by two types of cell populations, undifferentiated cytotrophoblasts (CT) and fully differentiated syncythiotrophoblasts (ST), and the villous stroma (VS) (Gude et al., 2004; Castillo et al., 2018). The trophoblast is connected to and separated from the villous stroma (VS), the fetal connective tissue by a basal lamina, a specialized structure of extracellular matrix (ECM) (Benirschke et al., 2012). Therefore, trophoblast, basal laminae, and VS, the latter containing fetal capillaries, form the placental barrier that pathogens must cross to infect the fetus during transplacental transmission (Kemmerling et al., 2019). However, the placenta can protect the developing fetus from some kind of pathogens (Zeldovich et al., 2013). That is the case of the kinetoplastid Trypanosoma cruzi (T. cruzi), where only a few parasite antigens and DNA can be identified in human chorionic villi (Duaso et al., 2010). Additionally, the presence of T. cruzi activates local placental defense mechanisms such as the epithelial turnover of the BLT and secretion of pro-inflammatory cytokines through Toll-like receptors (TLR) activation and NFkB signaling (Liempi et al., 2016, 2019; Castillo et al., 2017a, b).
The ovine and bovine placenta are of cotyledonary shape, villous and epitheliochorial. The fetal component is formed by the fusion of the avascular chorion and the vascular allantois. The placental barrier is composed by six tissue layers: maternal capillary endothelium, maternal endometrial connective tissue, maternal endometrial epithelium, trophoblast, chorionic connective tissue, and fetal endothelium (Benirschke et al., 2012; Liempi et al., 2020).
The type of placental barrier has been associated with the probability of transmission of pathogens. Thus, it has been proposed that in the hemochorial placenta, where the trophoblast is in direct contact with maternal blood, the placental infection and, therefore, the transmission to the fetus is facilitated. Moreover, considering that the complexity of the placental barriers increases from hemochorial (human) to epitheliochorial (ovine), it could be assumed that the parasites cross the hemochorial barrier more easily. However, the same hemochorial barrier favors the transfer of maternal antibodies to the fetus, and in less invasive placentas, a greater variety of pathogens is observed (Capellini et al., 2015; Liempi et al., 2020).
Unfortunately, most pathogens, including apicomplexan, can surpass the placental barrier and infect the fetus. As we will see in the following sections, these parasites use very similar adhesion mechanisms to the placenta, based on the modulation of the expression of adhesion molecules on the surface of placental cells. Additionally, the presence of these pathogens induces alterations in the immune response and a change in the Th1/Th2 balance, favoring the activation of defense mechanisms based on cellular immunity, which harms placental function and fetal growth.
Congenital Malaria
Malaria represents a high risk to the pregnant woman, fetus, and newborn. In some endemic areas, the prevalence of exposure to infection during pregnancy is 35%. Globally, malaria causes each year over 10,000 maternal and 75.000–200,000 neonatal deaths (Dombrowski et al., 2018; WHO, 2019). Congenital malaria is characterized by the sequestration of erythrocytes infected with Plasmodium parasites and the infiltration of immune cells within the intervillous space (IVS) of the placenta (Figure 1A), and the acquisition of a dark color of the basal plate, the fetal membranes (FM), fetal capillaries (FC), and monocytes (Mo) in the IVS, due to the deposition of the hemozoin malarial pigment (Parekh et al., 2010; Sharma and Shukla, 2017). The presence of parasites causes a pro-inflammatory environment in the placenta, leading to structural and functional alterations (Figure 1B). Placental damage, in turn, alters the nutrient exchange system between mother and fetus, leading to pregnancy-related complications including abortion, stillbirth, intrauterine growth retardation (IUGR), and low birth weight (LBW) (Briand et al., 2016; Sharma and Shukla, 2017). It has been proposed that due to hormonal and immunological changes in pregnancy (McLean et al., 2015), together with the fact that the parasite is localized inside red blood cells, it can more easily evade the maternal immune response (Sharma and Shukla, 2017). Interestingly, first-time pregnant women are more susceptible to malaria infection than women conceived a second or third time. This resistance to malaria infection in multigravida women is attributed to the development of placental parasite-specific immunity (Soulard et al., 2011).
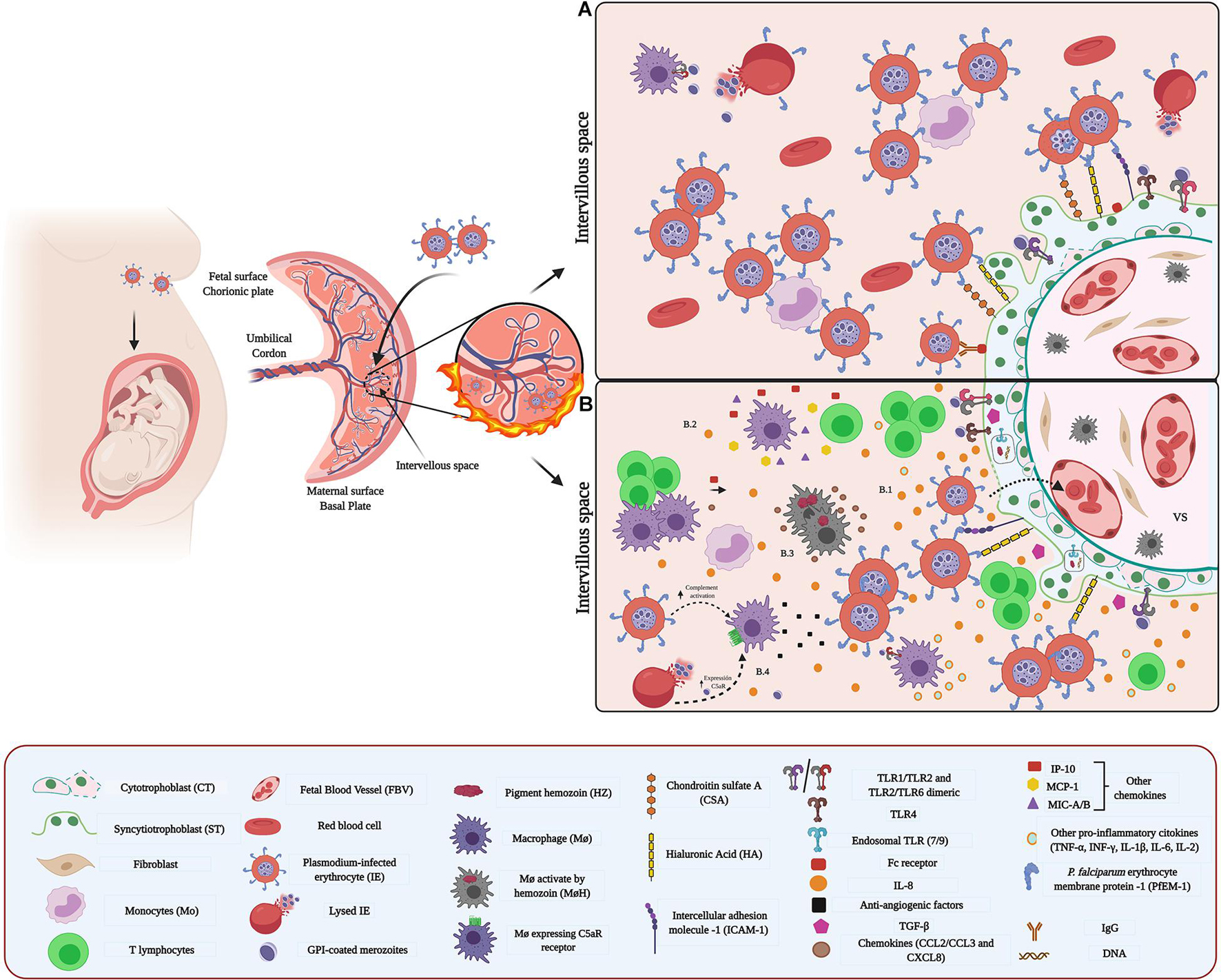
Figure 1. Host–pathogen interaction between Plasmodium and placenta: (A) Adhesion and sequestration of parasite-IEs. IEs express parasite-derived proteins on their surface. For example, the PfEMP1 protein family is highly expressed in placental IEs and acts as a surface antigen and ligand for their adhesion and sequestration. PfEMP1 selectively binds to specific placental receptors such as CSA. Other receptor molecules such as HI and ICAM-1, expressed in ST, participate as key molecules in the adhesion process. (B) Microenvironment and inflammatory response (B.1,B.2) Inflammation in the IVS is attributed to chemokines and cytokines secreted by maternal Mo, T cells, and ST. (B.3) Parasite’s hemozoin induces activation of MΦ, and subsequent release of chemokines. (B.4) Schizonts activate C5 and rupture releasing parasite components containing GPI that induce expression of C5aR and activate MΦ.
Any of the Plasmodium species can cause congenital malaria; however, depending on the geographic region, infection by a specific species may predominate. Thus, in endemic regions of the Indian and African subcontinent, most congenital malaria cases are due to P. falciparum and P. vivax (Bhatia et al., 2016), while in European countries, P. malariae and P. vivax are predominate (Vottier et al., 2008). In South America, P. vivax accounts for the majority of malaria cases (Recht et al., 2017). In addition, P. ovale is a rare causative agent of congenital malaria (Jenkins, 1957; Penazzato et al., 2007), is limited to specific areas of tropical Africa and islands in the Western Pacific (Collins and Jeffery, 2005).
Adhesion of Plasmodium to the Placenta
Plasmodium parasites invade red blood cells and induce the expression of parasite-derived proteins on the cell surface of the infected erythrocyte (IEs) (Chan et al., 2014). These parasite-derived proteins, known as variant surface antigens (VSAs), enable the IEs to adhere to the endothelium and subsequently infect different organs, including the placenta (Autino et al., 2012). In addition, VSAs bind to serum proteins forming red blood cell aggregates (rosetting), allowing the evasion of the host immune response, and establish chronic infection (Wahlgren et al., 2017). Among, VSAs P. falciparum erythrocyte membrane protein 1 (PfEMP1) is a major surface antigen involved in vascular adhesion and sequestration of IEs (Chan et al., 2014). Notably, fEMP1 proteins are encoded by a polymorphic var multigene family (approximately 60 copies per genome), characterized by expression through allelic exclusion, allowing switching between PfEMP1 proteins and subsequent modification of the antigenic and binding properties of IEs (Wahlgren et al., 2017).
PfEMP1 proteins mediate the adhesion of IEs to different receptors and surface molecules including integrins (Chesnokov et al., 2018), endothelial protein C receptor (EPCR) (Gillrie et al., 2015), CD36 (Hsieh et al., 2016), intercellular adhesion molecule-1 and 2 (ICAM-1/2) (Lennartz et al., 2019), proteoglycans, such as chondroitin sulphate A (CSA) (Pehrson et al., 2016), glycosaminoglycans (GAGs), such as hyaluronic acid (HA) (Beeson and Brown, 2004), P-selectin, E-Selectin (Metwally et al., 2017) and Platelet/endothelial cell adhesion molecule-1 (PECAM-1) (Berger et al., 2013). In the placenta, the PfEMP1 variant VAR2CSA is predominantly expressed in IEs (Tuikue Ndam et al., 2005) and is the main VSA responsible for parasite-binding tropism (Nunes and Scherf, 2007; Smith, 2014) in this organ (Benavente et al., 2018). Notably, a selective accumulation of mature asexual stages of P. falciparum- IEs occurs in the IVS and chorionic villi surface (Beeson et al., 2002; Muthusamy et al., 2004; Figure 1A). The accumulation of IEs is mediated by receptors and molecules on the ST surface, macrophages (MΦ) monocytes (Mo), the fibrinoid deposits, and blood vessels in the VS (Beeson et al., 1999; Matejevic et al., 2001; Sugiyama et al., 2001).
Chondroitin sulphate A is the principal placental IEs receptor (Fried et al., 2006; Chishti, 2015). However, other molecules such as HA, non-immune immunoglobulins (IgG/IgM), and ICAM-1, present on the ST, can act as adhesion receptors (Maubert et al., 1997; Sartelet et al., 2000; Flick et al., 2001; Beeson and Brown, 2004; Barfod et al., 2011; Figure 1A). CD38 is only localized in the cytoplasm of stromal cells, fibroblasts, and MΦ, where the presence of IEs might influence its expression without being directly implicated in the sequestration of these (Sartelet et al., 2000). On the other side, HA is associated mainly with the aggregation or clumping of IEs (Beeson and Brown, 2004). Importantly, the expression of the different receptors is influenced by P. falciparum-induced proinflammatory cytokines (Vásquez et al., 2013).
In addition, other factors determine the accumulation and the sequestration of IEs in the placenta, including the low placental blood flow associated with trophoblast conformational changes, for the formation of cytotrophoblastic prolongations, known as “Coan-Burton bridges” (Moraes et al., 2013), IEs cell deformability (Safeukui et al., 2018), and the presence of Mo (Sugiyama et al., 2001). The adhesion and invasion of IEs also depend on the type of Plasmodium species. Thus, P. vivax and P. falciparum can bind to placental CSA and HA. However, P. vivax cannot bind to CD36 and ICAM-1 (Chotivanich et al., 2012), ligands widely used by P. falciparum.
So far, the Plasmodium mechanisms of adhesion to the placenta are the best known among apicomplexan parasites. However, as we will see in the following sections, these mechanisms are preserved in related apicomplexan parasites such Babesia, Toxoplasma, and Neospora. In these organisms, the parasite proteins expressed in the infected cells, or the proteins present on the parasite’s surface play fundamental roles in the adhesion and infection processes in the placenta.
Immune Response Against Plasmodium
The development of severe malaria during pregnancy depends on parasite and host factors, including the immune response (Clark, 2019). During pregnancy, an increase of hormones, including cortisol, progesterone, estradiol, and testosterone, polarize the immune response toward a Th2-type one, characterized by an increase of regulatory T cells (Tregs) and anti-inflammatory cytokines (Robinson and Klein, 2012). The presence of the parasite alters this response causing infiltration of immune cells and a strong inflammatory response (Suguitan et al., 2003; Chêne et al., 2014).
Toll-like receptors, particularly TLRs-2 -4 -7-and -9, expressed on the trophoblast and cells of the VS, play a key role in the immune response against Plasmodium (Lucchi et al., 2011; Zhu et al., 2011; Barboza et al., 2014, 2017; Reis et al., 2020). TLR- 2 and -4 recognize glycosylphosphatidylinositol (GPI) anchors (Krishnegowda et al., 2005), while nucleic acids and hemozoin are recognized by TLR7 and TLR9 (Coban et al., 2005; Parroche et al., 2007; Figure 1B). The expression of TLR-4 and -9 is regulated by Plasmodium (Barboza et al., 2017) and polymorphisms of the TLRs have been associated with the susceptibility to infection (Hamann et al., 2010).
The binding of IEs and parasite antigens to TLR expressed on the trophoblast promotes the activation of MAPK pathways leading to the secretion of pro-inflammatory cytokines, including TNF-α, IFN-γ, TGF-β, and IL-8, and consequent activation of immune cell (Lucchi et al., 2008; Reis et al., 2020; Figure 1B). Concomitantly, the chemokines interferon γ–induced protein-10 (IP-10), monocyte chemoattractant protein-1 (MCP-1), and macrophage inflammatory protein (MIP)-1a/b expressed in maternal white blood cells (WBCs) contribute to the accumulation of Mo and MΦ in the IVS (Suguitan et al., 2003). Then, the presence of the immune cells and cyto/chemokines leads to a Th1-type immune response, where in addition to the cytokines mentioned above, IL1β, IL-2, IL-6, (Dobaño et al., 2018) and IL-8 (Fried et al., 1998) are secreted (Figure 1B). Thus, T cells proliferation is induced, and MΦ phagocytosis is enhanced, aiming to limit the parasite’s replication (Ozarslan et al., 2019).
Overproduction of cytokine IL-10, an immunoregulating cytokine, is associated with response to placental malaria infections (Suguitan et al., 2003; Chêne et al., 2014) attenuating the detrimental effects on the placental barrier exerted by the exacerbated inflammatory response (Okamgba et al., 2018). Alternatively, the increase in IL-10 could modulate lymphoid dendritic cell (DC) subpopulations shifting the Th1/Th2 balance toward a TH1 response and consequently promote infection (Diallo et al., 2008).
Mo-secreted IL-1β in response to IEs alters the amino acid transport system in the trophoblast impairing their transfer to the fetus and contributes to PM-associated FGR’s pathogenesis. Moreover, the accumulation of Mo in the malarial placenta promotes the decrease of insulin-like growth factor-1, one of the most influential factors in fetal growth and life (Umbers et al., 2011).
Different MΦ populations are related to functional modification of the placenta and maternal-fetal tolerance (Ordi et al., 1998; Gaw et al., 2019). Thus, maternal-intervillous monocytes (MIM) are associated with massive chronic villositis and placental damage (Ordi et al., 1998). Likewise, a decrease in the anti-inflammatory M2-type Hofbauer cells (HBC), located in the VS, is associated with LBW in placental malaria (Gaw et al., 2019).
Plasmodium also activates the complement cascade (C) and subsequently modulates the release of the anti-angiogenic factors (Conroy et al., 2011; Chau et al., 2017; Figure 1B). Together, an excessive mononuclear cells activation, such as MΦ, and an increase of soluble vascular endothelial growth factor (VEGF) receptor-1 (sFlt-1), which reduce the bioavailability of the pro-angiogenic VEGF and placental growth factor (PGF), impair normal angiogenesis (Azimi-Nezhad, 2014; Chau et al., 2017) and trigger functional placental insufficiency, and ultimately, FGR (Conroy et al., 2011).
The presence of Hemozoin (Hz) is another determining factor in the placental malaria immune environment. Hz can stimulate the trophoblast, MΦ, and DCs to secrete chemokines and cytokines [CCL2, CCL3, and CXCL8 and tumor necrosis factor (TNF)], contributing to the recruitment of peripheral blood mononuclear cells, especially Mo (Lucchi et al., 2011) and inflammatory response (Moormann et al., 1999; Abrams et al., 2003; Figure 1B). In addition, phagocytosis of this pigment increases the expression of matrix metalloproteinase (MMPs), particularly MMP-9, in the trophoblast (Clark, 2019) inducing cleavage and liberation of the extracellular domain of Syndecan-1; therefore allowing the adherence of IEs and promote their accumulation in the placenta (Clark, 2019).
Congenital Babesiosis
Congenital babesiosis, caused by B. microti and Babesia sp., is considered a rare disease; to date, a total of 10 cases have been reported in the United States (Esernio-Jenssen et al., 1987; New et al., 1997; Sethi et al., 2009; Aderinboye and Syed, 2010; Cornett et al., 2012; Joseph et al., 2012; Yager et al., 2014; Surra and Jesus, 2015; Saetre et al., 2018). Although transplacental babesiosis has been reported, so far, no histological studies have been conducted in Babesia-infected placentas. However, it has been proposed that B. microti sporozoites can invade red blood cells and cross the placental barrier and infect the fetus (Figure 2A). On the other side, the presence of the parasite in amniotic fluid has been demonstrated (Cornett et al., 2012). During gestational babesiosis, clinical manifestations similar to the HELLP syndrome (hemolysis, elevated liver enzymes, and low platelets) may develop, causing significant maternal and neonatal morbidity and mortality (Khangura et al., 2019). In infants, in addition to presenting hemolytic anemia, thrombocytopenia, and fever, hepatosplenomegaly is one of the most common symptoms (Saetre et al., 2018).
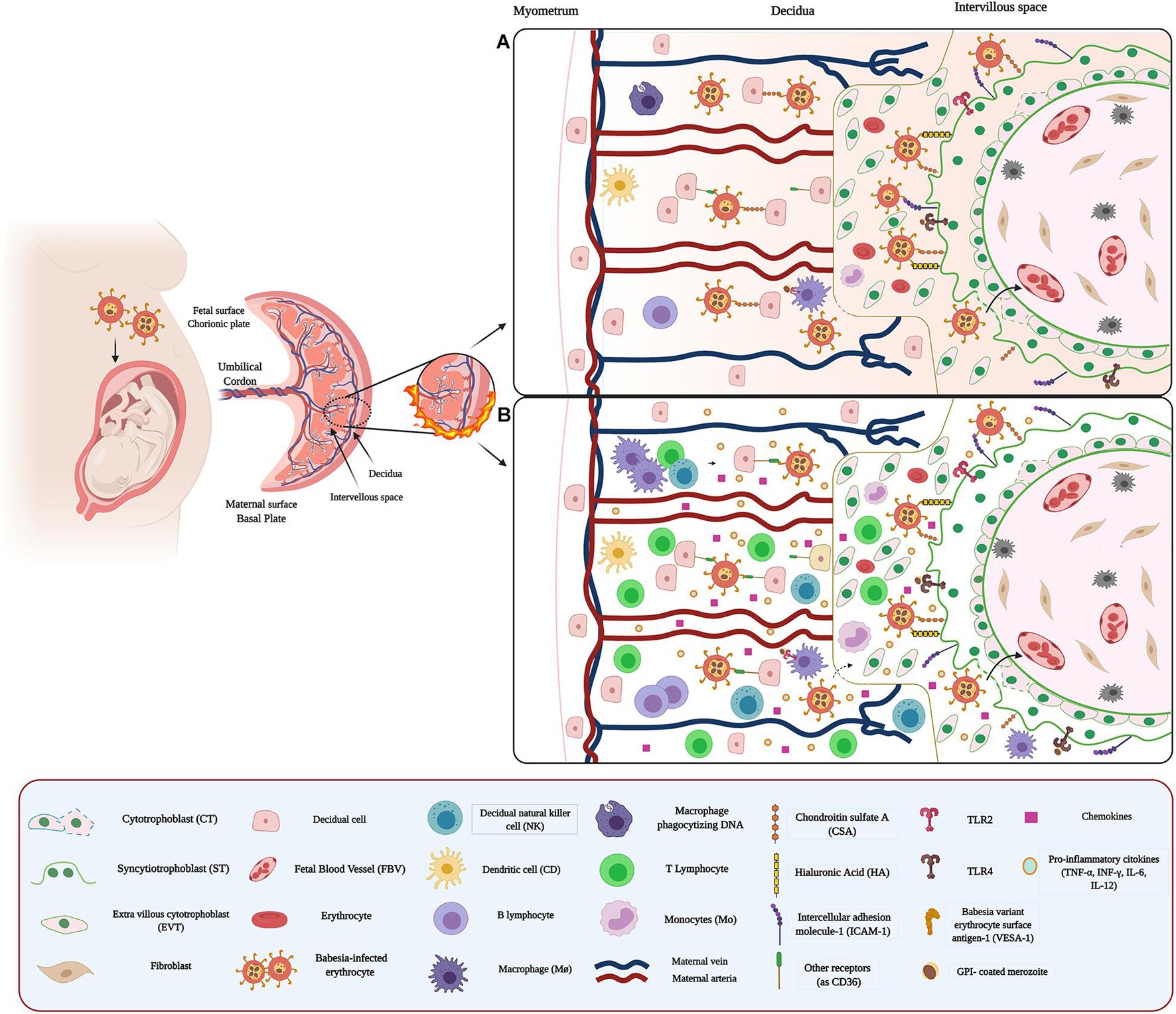
Figure 2. Host–pathogen interaction between Babesia and placenta: (A) Adhesion and sequestration of Babesia-IEs Infection of erythrocytes by the parasite induces expression of parasite-derived proteins on the surface of the IEs. Babesia-infected IEs highly express the VESA1 family and possibly act as a surface antigen and ligand for adhesion and sequestration of IEs in the placenta. VESA1 selectively binds to specific placental receptors, such as CSA, HA, and ICAM-1, expressed by ST, participate as key molecules in this adhesion process. In addition, some receptors such as CD36 could also facilitate the adhesion of IEs in other regions of the placenta, such as the decidua. (B) Microenvironment and inflammatory response against Babesia. Inflammation in the IVS and decidua could be attributed to chemokines and cytokines secreted by maternal MΦ, Mo, B cell, and T cells, as well as ST.
Babesiosis has also been documented in other mammals such as canines, felines, cattle, and rodents (Schoeman, 2009; Tołkacz et al., 2017). In these animals, babesiosis is caused by several species of babesia, which are relatively host-specific. However, all Babesia species can be transferred to other hosts through infected blood (Cox, 1982). Interestingly, in rodents, the placental transmission of B. microti is higher than in other mammals, including humans (Tołkacz et al., 2017; Tufts and Diuk-Wasser, 2018).
Adhesion of Babesia to the Placenta
The pathogenesis of Babesia is strikingly similar to that of malaria (Figure 2A). Thus, alterations in the adhesive properties of IEs due to antigenic modifications of their membrane (Cooke et al., 2005; Hutchings et al., 2007), caused by the expression of a variant of the parasite-derived erythrocyte surface antigen protein (VESA), can be observed (Jackson et al., 2014). VESAs are proteins encoded by various families of ves1α and ves1β genes and secreted by the parasite (Allred et al., 2000; Allred, 2019). In addition, VESAs appear to play essential roles in pathogenicity (Pedroni et al., 2013), immune evasion (Jackson et al., 2014), persistence, and survival of the parasites (Allred, 2019). VESAs, variant erythrocyte surface antigenic- 1 (VESA1) protein, is one of the most studied in B. bovis. This protein acts as an endothelial cell ligand and mediator of antigenic variation in B. bovis-IEs (O’Connor and Allred, 2000; Wang et al., 2012). The host cell adhesion receptors for VESA have not yet been identified (Gallego-Lopez et al., 2019). However, structural similarities between fEMP1 and VESA1 suggest that the latter could bind to equivalent host cell receptors of PfEMP1 such as CD36 (Hutchings et al., 2007), ICAM-1, P-selectin, CSA, and CD31 (Gallego-Lopez et al., 2019; Figure 2A). Furthermore, studies performed with B. bigemina and B. ridhaini have demonstrated the adhesion IEs to thrombospondin (PST) (Parrodi et al., 1990) present in endothelial cells (Baruch et al., 1996).
Importantly, the severity of babesiosis is highly dependent on the babesia species (Cooke et al., 2005). For example, babesiosis caused by Babesia microti, B. bovis, B. Lengua, and B. canis is associated with the development of severe syndromes such as brain babesiosis and multi-organ failure due to the sequestration of IEs in the organ’s microvasculature (Wright et al., 1979; Aikawa et al., 1992; Bosman et al., 2013; Sondgeroth et al., 2013; Ripoll et al., 2018; Schetters, 2019).
Immune Response Against Babesia
There are few reports related to the placental immune response against Babesia. However, it has been suggested that pro-inflammatory cytokine activation and erythrocyte adhesion in babesiosis are similar to malaria (Krause et al., 2007; Figure 2B). Some studies have even documented that prior infection with B. microti protects against fatal malaria in mice and primates through a mechanism of cross-protection (van Duivenvoorde et al., 2010; Efstratiou et al., 2020).
Toll-like receptors also recognize Babesia through the presence of its surface of GPI-linked molecules. The main TLRs studied regarding Babesia species are TLR-3 and -4, which can signal downstream through MyD88 independent pathways (Skariah et al., 2017). Similarly, TLR2 could play a role in this inflammatory response, mediating MΦ activation in the placenta. Previous studies have shown that lipid extracts from B. bovis stimulate TLR-2-mediated MΦ activation (Gimenez et al., 2010; Figure 2B) and consequently induces the secretion of pro-inflammatory cytokines, including TNF-α, IL-1, IL-12, and the immunomodulating IL-8 and nitric oxide (NO) (Shoda et al., 2000; Terkawi et al., 2015). In addition, T lymphocytes and natural killer (NK) cells, responsible for INF-γ production, and B lymphocytes are essential elements in the immune response against this parasite (Krause et al., 2007; Vannier and Krause, 2012; Yi et al., 2018; Figure 2B). Notably, the DNA of this parasite also has immunomodulatory effects on both B cells and MΦ, through the induction of IL-12, TNF-α, and NO production (Shoda et al., 2001).
Babesia, like Plasmodium, increases in the expression of MCP-1 and MIP-1a chemokines, contributing to the accumulation of Mo and MΦ in the IVS (Skariah et al., 2017; Zhao et al., 2020). Also, other chemokines such as CCL4 (MIP-1β) and CCL5 (RANTEs) have been associated with an inflammatory process in response to this parasite (Skariah et al., 2017).
Congenital Toxoplasmosis
Congenital toxoplasmosis is one of the leading causes of infant morbidity and mortality (Melamed et al., 2010). Although it is a disease with global distribution, its prevalence is markedly variable from one region to another (Melamed et al., 2010; Torgerson and Mastroiacovo, 2013), with a global estimated incidence of 190100 annual cases (Torgerson and Mastroiacovo, 2013). Congenital toxoplasmosis occurs predominantly after primary infection in pregnant women (Pappas et al., 2009). However, congenital transmission has been reported in pregnant women infected before pregnancy (Pons et al., 1995; Vogel et al., 1996), chronically infected women, in whom the T. gondii infection was reactivated due to immunosuppression by HIV or by drug treatment for underlying diseases (Marty et al., 1994; Bachmeyer et al., 2006), and in women previously infected with one serotype who developed a new infection with a second serotype acquired during pregnancy (Fortier et al., 1991; Valdès et al., 2011).
During congenital transmission, the parasite crosses the placenta directly or with the help of infected immune cells, Mo, and DCs, using them as “Trojan horses” (Figure 3A; Robert-Gangneux et al., 2011; Arranz-Solís et al., 2021), inducing phenotypic and functional alterations in infected cells, including overexpression of adhesion molecules and receptors, hypermotility, and downregulation of cytokine expression (Lachenmaier et al., 2011; Masocha and Kristensson, 2012; Sanecka and Frickel, 2012; Arranz-Solís et al., 2021). The parasite induces an inflammatory response (Figure 3B) that compromises mother and child health and the success of pregnancy (Robert-Gangneux et al., 2011). In addition, T. gondii causes severe neuro-ocular alterations such as chorioretinitis and the worst-case fetal death (Melamed et al., 2010). The severity of congenital Toxoplasmosis is inversely related to the gestational age at the time of primary maternal infection, implying that maternal infections in the first trimester of pregnancy lead to more serious clinical manifestations (McAuley, 2014). Besides, the clinical outcome of congenital Toxoplasmosis also is related to the T. gondii genotypes (Gilbert et al., 2008; McAuley, 2014), being the type I and II isolates related to more severe congenital Toxoplasmosis (Fuentes et al., 2001; Hutson et al., 2015; Rico-Torres et al., 2016; Lahmar et al., 2020; Arranz-Solís et al., 2021).
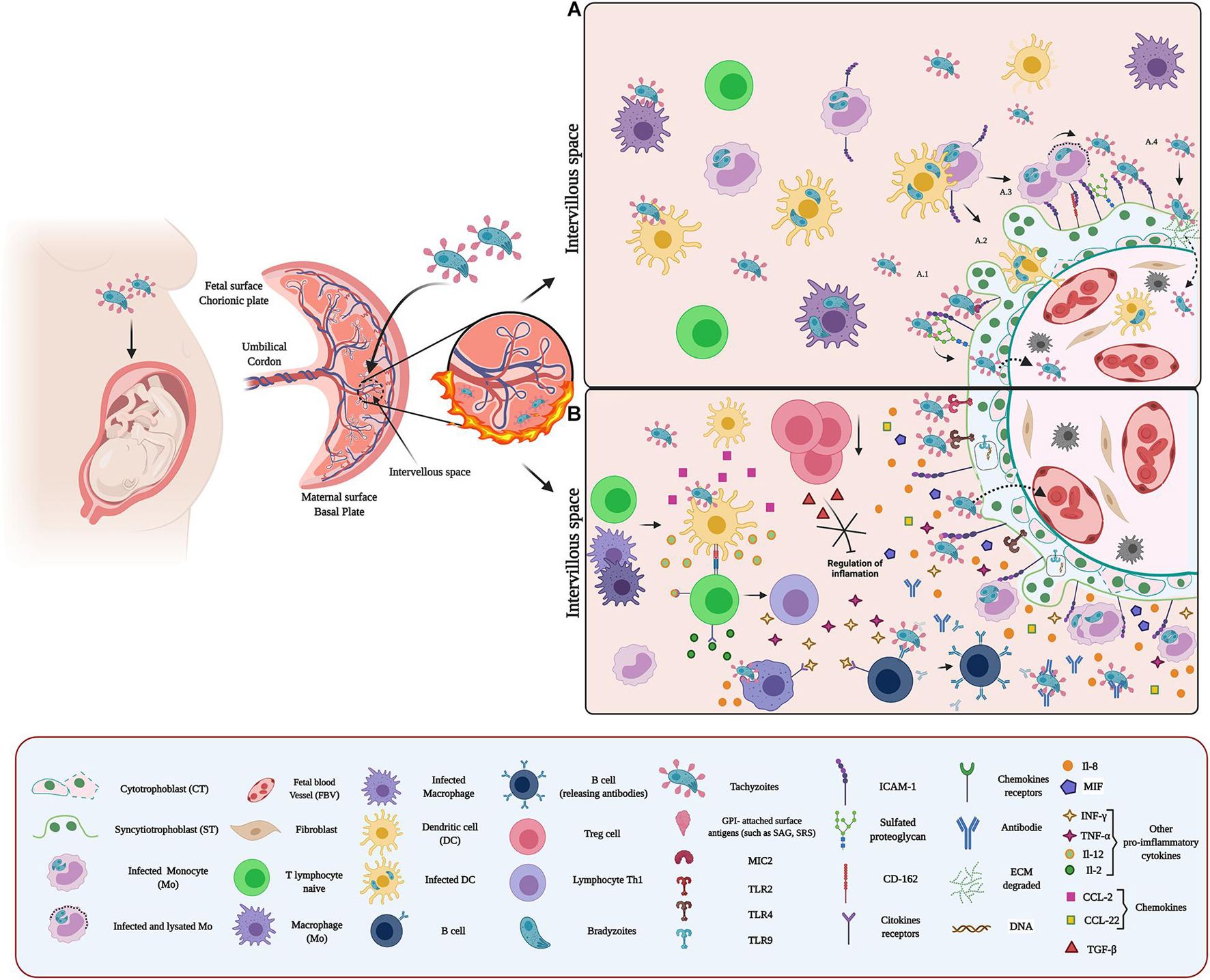
Figure 3. Host–pathogen interaction between Toxoplasma and placenta: (A) Adhesion of T. gondii in the placenta. Toxoplasma adhesion to the placenta can occur through proteins belonging to the SAG, SRS, and MIC families. These proteins expressed on the surface of the parasite act as surface antigens and ligands for molecules present on the surface of placental cells. These surface antigens could bind to GAGs, glycoproteins, adhesion molecules, and some extracellular matrix components. Once attached to the placenta, T. gondii crosses the placenta by different mechanisms: (i) without altering the integrity of the barrier (A.1); (ii) infecting immune cells (such as CDs) and using them as Trojan horses (A.2); (iii) exploiting the natural routes of leukocytes extravasation and coordinating its exit with the arrival at the target organ, thus minimizing attack by the immune system during the vehicle transition to target cells (A.3); and (iv) and promoting EMC degradation, through modulation of MMP expression and secretion (A.4). (B) Microenvironment and inflammatory response against T. gondii. Recognition of T. gondii by TLR (TLR2/4/9) induces IL-8 expression and secretion. Additionally, infection by this parasite promotes the expression of MIF and CCL-2. Recognition of the parasite by immune cells (such as CDs and MΦ) activates an inflammatory response characterized by cytokine and chemokine production such as IL-12, CCL-2, and CCL-22, which activate naive T cells. Subsequently, these cells differentiate to Th1 and produce cytokines (such as IL-2, INF-γ, and TNF-α) that promote an exacerbated pro-inflammatory environment, which stimulates the recruitment and activation of other immune cells at the site of infection. Additionally, infection by T. gondii promotes a decrease in Tregs, contributing to the exacerbated inflammatory reaction.
Toxoplasma gondii infects warm-blooded animals, including birds, cats, sheep, dogs, cows, goats, and pigs (Mondragon et al., 1998; Dubey et al., 2003; Stelzer et al., 2019); being isolates type II III the most frequent genotypes (Lachkhem et al., 2021). In some animals, the congenital transmission frequency is also high (Duncanson et al., 2001; Dos Santos et al., 2016) and is associated with fetal loss and economic losses (Stelzer et al., 2019). Interestingly, the potential risk of parasite transmission in animals through ingestion of contaminated milk has been suggested (Dubey et al., 2014; Ossani et al., 2017), increasing, therefore, the risk of zoonotic infection.
Adhesion of Toxoplasma gondii to the Placenta
Toxoplasma gondii cell invasion is a multi-step process that depends on the parasite’s motility, surface antigens, and the sequential secretion of proteins present in the apical secretory organelles (Carruthers and Boothroyd, 2007; Robert-Gangneux and Dardé, 2012).
The invasion process starts with low-affinity interaction between T. gondii surface molecules, including glycosylphosphatidylinositol (GPI)-attached surface antigens (SAG), SAG-related sequence proteins (SRS), and non-SAG-related surface antigens (SUSA) and the cell membrane of the host cell (Pollard et al., 2008; Blader and Saeij, 2009). The SAGs multigenic family comprises more than 150 genes distributed and tandemly arrayed throughout the genome (Lekutis et al., 2001; Jung et al., 2004). SAG and SRS1 proteins are predominantly expressed on the tachyzoite (rapidly dividing parasites) surface cells (Tomavo, 1996; Manger et al., 1998). Regarding the SUSA molecules, 31 genes have been identified, and the proteins are highly expressed in bradyzoites (slow dividing parasites) (Pollard et al., 2008). Significantly, all these surface antigens are also involved in immune modulation, virulence, and protection of the parasite to survive in the environment (Lekutis et al., 2001; Crawford et al., 2009), for what they are considered redundant systems of adhesion to the host cell (Manger et al., 1998). The host cell surface molecules include GAGs, proteoglycans, and glycosaminoglycan-like carbohydrates (Jacquet et al., 2001; He et al., 2002; Azzouz et al., 2013). In addition, proteins of apical secretory organelles also play a key role in parasite-cell adhesion. For example, rhoptry proteins (ROP 2, ROP 4) and dense granules protein (GRA2) are lectins that bind to sulfated GAGs and then participate in the moving junction complex (MJ complex) (Azzouz et al., 2013).
Additionally, T. gondii is recognized and adheres to ICAM-1 on the host cell surface. Thus, micronemal proteins (MICs), such as MIC2, binds to ICAM-1 and facilitate the transmigration of the parasite through the blood-brain barrier (Lachenmaier et al., 2011). Also, interactions with ECM laminin (Furtado et al., 1992) and sialylated oligosaccharides (Blumenschein et al., 2007) have been reported. In the placenta, T. gondii infects different cell types, including decidual cells, extravillous trophoblasts (EVT), and the trophoblast (Abbasi et al., 2003; Robbins et al., 2012). The parasite’s binding to these cells is mediated by sulfated proteoglycan and ICAM-1 (Abbasi et al., 2003; Barragan et al., 2005; Figure 3A.1). ICAM-1 is abundantly expressed at intercellular junctions and the surface of in human trophoblast cell line (BeWo) (Barragan et al., 2005). Also, T. gondii induces syncytial expression of this adhesion molecule (Juliano et al., 2006) increasing the adhesion of T. gondii-infected immune cells to the trophoblast (Pfaff et al., 2005). Thus, parasite infection of Mo and maternal leukocytes promotes their adhesion to trophoblasts through the upregulation of ICAM-1, contributing to placental and ultimately fetal infection (Xiao et al., 1997; Pfaff et al., 2005). Moreover, decidual DCs could serve as a vehicle for T. gondii to travel across the placenta and infect the fetus (Sanecka and Frickel, 2012; Arranz-Solís et al., 2021; Figure 3A.2). In addition, T. gondii can exploit natural routes for leukocyte extravasation to cross the placental barrier (Figure 3A.3). In vascular endothelium, CD162 is recognized by the vascular adhesion molecule-1 (VCAM-1) and E-selectin of infected cells mediating their attachment (Harker et al., 2013). Interestingly, CD162 has been identified in trophoblast cell lines (Multhaup et al., 2018).
The adhesion of infected immune cells to the endothelium serves as a signal to the parasite, allowing to schedule its leukocyte output at the target organ and minimizing the attack by the immune system (Baba et al., 2017). Thus T. gondii reaches the placenta inside maternal leukocytes and adheres to the trophoblast (Figure 3A.3). In the case of Mo, the presence T. gondii antigens, such as soluble tachyzoite antigen (STAg), induces the overexpression of macrophage migration inhibitory factor (MIF), allowing the adhesion to the trophoblast (Ferro et al., 2008). Moreover, the adhesion of infected immune cells can also induce local destruction of the trophoblast (Juliano et al., 2006) through the induction of pro-inflammatory cytokine-dependent apoptosis (Garcia-Lloret et al., 2000; Juliano et al., 2006). It has also been proposed that the accumulation of leukocytes is responsible for placental villitis (Yavuz et al., 2006).
The ECM degradation is another strategy used by T. gondii to cross and damage the placental barrier (Wang and Lai, 2013; Liempi et al., 2020; Figure 3A.4). Placental infections increase the expression and secretion of MMPs, MMP-2, MMP-9, and MMP-12 in the VS, causing ECM degradation (Wang and Lai, 2013) particularly of fibronectin, collagen I and IV, and laminin (Chen and Aplin, 2003).
Immune Response Against Toxoplasma gondii
Toxoplasma gondii infection in the placenta induces an inflammatory response characterized by a robust production of pro-inflammatory cytokines and chemokines (Sasai et al., 2018; Sasai and Yamamoto, 2019).
Toll-like receptors are critical in initiating defense against T. gondii; they recognize in the parasite glycosylphosphatidylinositol (GPI) anchored proteins and lipid anchors, heat shock protein 70 (TgHSP70), and profilin-like protein (TgPF) (Yarovinsky and Sher, 2006; Denkers, 2010; Yarovinsky, 2014; Sasai et al., 2018). GPI, are recognized primarily by TLR-2, TL-4 and TL2/TL1/6 heterodimers (Debierre-Grockiego et al., 2007) while TgPF is recognized by the TLR-5, 11 and -12 (Andrade et al., 2013; Salazar Gonzalez et al., 2014; Yarovinsky et al., 2005). On the other hand, parasite’s RNA and DNA are recognized by the endosomal TLR-7 and -9 (Yarovinsky, 2014). TLR activation leads to an increase of IL-12, IL-6, IL-8, and TNF-α cytokines, chemokines (CCL5, CCL12, and XCL1), interferons (IFNs), and other effector molecules such as NO, which favors infection control (Yarovinsky and Sher, 2006; Wujcicka et al., 2013).
Haplotype and single nucleotide polymorphisms (SNPs) of tlr genes can influence susceptibility to parasitic diseases (Wujcicka et al., 2013, 2017; Turkey et al., 2019). Thus, SNPs residing in TLR2, TLR4, and TLR9 genes increase susceptibility and development of toxoplasmosis in pregnancy (Wujcicka et al., 2013, 2014, 2017; Turkey et al., 2019). This susceptibility is attributed to the influence of SNPs in the receptor dimerization and the recruitment of adapter proteins involved in TLR signaling and the decreased synthesis of some cytokines (Wujcicka et al., 2017; Turkey et al., 2019), such as INF-γ and TNF-α.
We have shown previously that during ex vivo infection of human placental explants (HPE), T. gondii increases the expression of TLR-9, but not of TLR-2 and -4. However, inhibition of TLR-4 and -9 increases T. gondii DNA load in the explants. Notably, only IL-8 secretion was increased (Muñoz et al., 2016; Castillo et al., 2017a; Figure 3B); even the protective and pro-inflammatory cytokines INF-γ and TNF-α did not show a significant change in response to the parasite (Castillo et al., 2017a). This result could explain, at least partially, the susceptibility of the placenta to T. gondii infection compared to other parasites (Castillo et al., 2017a). In addition, it has been suggested that IL-8 promotes the proliferation and differentiation of the infected host cell, and subsequently, the intracellular multiplication of parasites (Milian et al., 2019), promoting, therefore, congenital transmission (Gómez-Chávez et al., 2019, 2020). Moreover, T. gondii Macrophage Migration Inhibitory Factor (TgMIF) cytokine-like, an analog to the host MIF, promotes IL-8 secretion and subsequent recruitment of immune cells to the site of infection (Sommerville et al., 2013) causing the above-described effects (Lambert et al., 2006).
On the other hand, the low INF-γ expression (Muñoz et al., 2016; Castillo et al., 2017a) could also be a placental mechanism to limit infection. Both INF-γ and TNF-α promote the adhesion of infected immune cells to the trophoblast through surface overexpression of ICAM-1 (Pfaff et al., 2005; Ferro et al., 2008). Therefore, it is possible that the downregulation of these pro-inflammatory cytokines, in addition to benefiting the maintenance of pregnancy, also limits the transfer of the parasite through the placental barrier. Furthermore, this could be related to the production of CCL22 chemoattractant in response to T. gondii (Ander et al., 2018) since CCL22 can replace the effect of IFN-γ and recruit Tregs modulating the inflammatory response. However, this increased T-reg recruitment may also be a parasite strategy to promote its persistence (Rowe et al., 2011). Furthermore, MIF is also overexpressed in responses to T. gondii infection (de Oliveira Gomes et al., 2011; Figure 3B), promoting the adhesion of infected immune cells to the trophoblast, activating the ERK1/2 MAPK pathway and subsequent production of prostaglandin E, favoring parasite proliferation and persistence through inhibition of IL-6, IL17, and TNF-α production (Kelly et al., 2005; Passos et al., 2010; Barbosa et al., 2014; Qiu et al., 2016).
The presence of T. gondii causes an imbalance of the Treg/T-helper type 17 (Th17) cells response inducing deleterious alterations in the placenta that can lead to abortion (Zhang et al., 2012). T. gondii, as mentioned above, promotes the production of IL-12, CCL-2, and CCL-22 activating naïve T cells. In addition, activated T cells secrete IL-2, INF-γ, and TNF-α, developing a pro-inflammatory environment that stimulates MΦ and B cells (Gómez-Chávez et al., 2020) and decreases Tregs and TGF-β levels (Figure 3B). The decrease of Tregs may be due to various factors, including apoptosis (Ge et al., 2008), limitation of proliferation (Oldenhove et al., 2009), and an increase in differentiation into Th17 cells (Zhang et al., 2012).
Importantly, all the factors involved in decreasing Tregs have in common the activation of multiple pathways, including the pro-inflammatory ones (Ge et al., 2008; Oldenhove et al., 2009; Zhang et al., 2012). The increased differentiation of TH17 cells is associated with altered expression of TGF-β, IL-17A, and IL-6 cytokines, transcription factors [i.e., Forkhead box-p3 (Foxp3)], and receptors [i.e., Retinoic acid-related orphan receptor γt (RORγt)], keys in cell differentiation and regulating Treg/Th17 balance (Zhang et al., 2012). Although it is unclear how T. gondii regulates the expression of TFG-B and Foxp3, it influences suppressive functions of Tregs in the placenta (Zhang et al., 2012), resulting in an exacerbated inflammatory reaction, causing placental damage and favors the transmission of the parasite to the fetus.
Congenital Neosporosis
Congenital neosporosis affects mainly dogs and cattle, but occasionally also horses, sheep, and deer (Dubey, 2003), leading to abortion, stillbirth, and weak offspring (Mesquita et al., 2018). In sheep, congenital transmission rates are high, reaching values between 66.7 and 93% (González-Warleta et al., 2018). Transplacental transmission can be exogenous or endogenous. Exogenous transmission occurs following ingestion of sporulated oocysts by pregnant females, while endogenous one occurs as the reactivation of a pre-existent infection. The latter is the primary mechanism responsible for maintaining the parasite within livestock populations. The presence of the parasite induces an inflammatory response leading to placental damage and consequent abortion (Cantón et al., 2014b; Al-Shaeli et al., 2020).
Thus, multiple necrotic lesions, thickening of the chorionic plate, and mineralization of necrotic foci can be observed in the infected ovine placenta. Notably, both the oocysts (sexual state) and tachyzoites (rapidly dividing parasite stage) of N. caninum can be detected in placental tissue (Mesquita et al., 2018; Al-Shaeli et al., 2020; Figure 4A). In the fetus, N. caninum causes severe damage to the brain, heart, lung, and liver (Al-Shaeli et al., 2020).
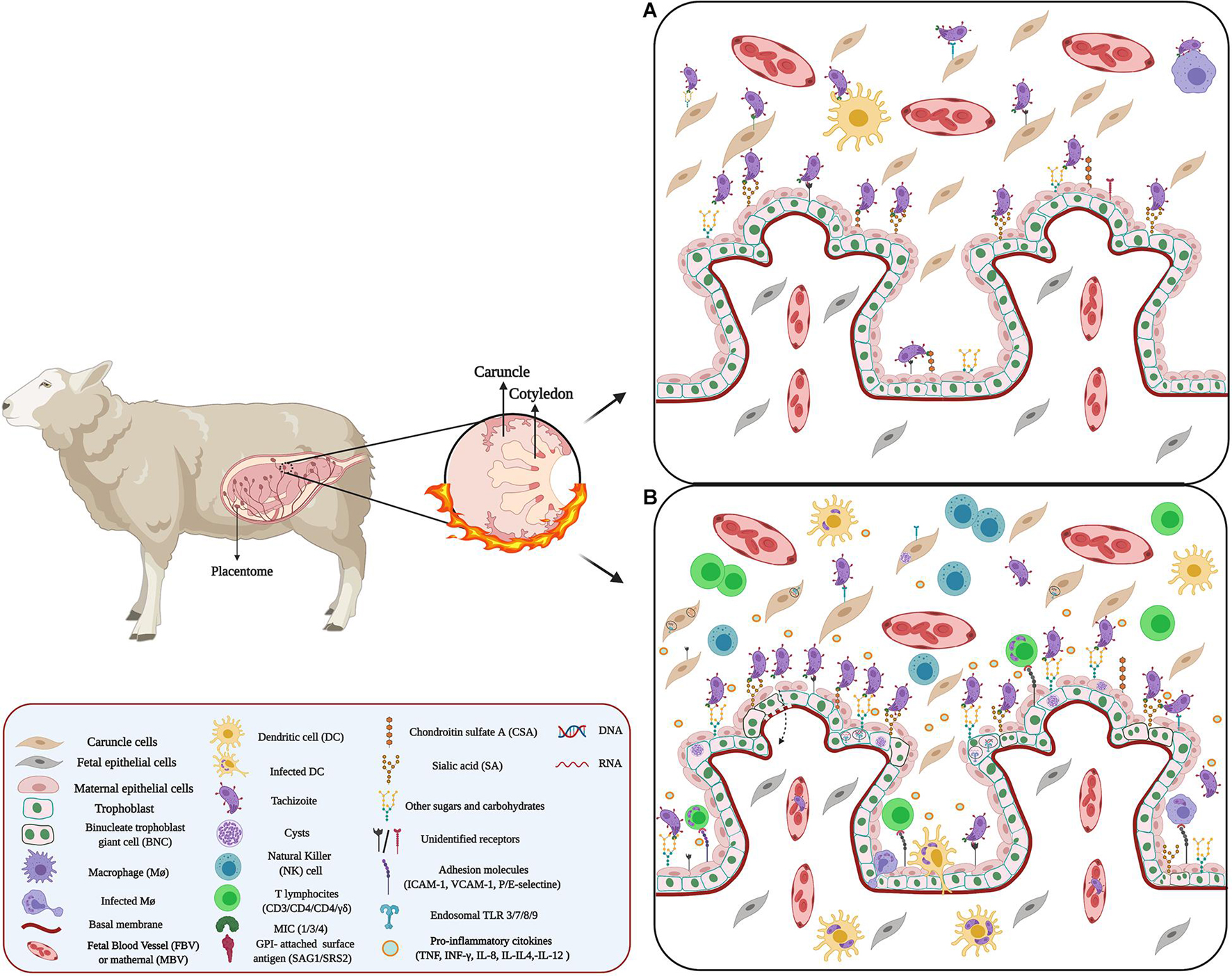
Figure 4. Host–pathogen interaction between Neospora and placenta: (A) Adhesion of Neospora. Neospora adhesion to the placenta can occur through proteins belonging to the SAG, SRS, and MIC families. These proteins expressed on the surface of the parasite act as surface antigens and ligands for molecules present on the surface of placental cells. Possibly, these protein proteins could bind to proteoglycans and placental adhesion molecules. (B) Microenvironment and inflammatory response against Neospora. Inflammation in the IVS and caruncle could be attributed to chemokines and cytokines secreted by maternal Mo, T cell, as well as ST and caruncle cells. Many of these cytokines and chemokines promote cell recruitment. Additionally, infection of some cells of the maternal immune system, such DCs, and MΦ, by Neospora alters the migration of these cells, using them as Trojan horses.
Although N. caninum is not considered a zoonotic parasite, serological studies indicate the possible capacity of N. caninum to infect humans, particularly immunocompromised individuals (Tranas et al., 1999; Lobato et al., 2006; Oshiro et al., 2015) and pregnant women. Recent reports revealed N. caninum in samples of umbilical cord blood and placental tissue in pregnant women. The seroprevalence in these women can vary between 8 and 24% (Ibrahim et al., 2009; Duarte et al., 2020) and is associated with the presence of domestic animals (Duarte et al., 2020). The biological similarities between N. caninum and T. gondii suggest the possibility of N. caninum transmission between humans (Petersen et al., 1999). Moreover, in studies where pregnant monkeys were inoculated with N. caninum, the transplacental transmission was evident; in the fetus, similar lesions as those caused by T. gondii can be observed (Barr et al., 1994; Ho et al., 1997). In mice, the congenital transmission of N. caninum occurs in 85% of the litters born from infected mothers (Cole et al., 1995).
Adhesion of N. caninum to the Placenta
Neospora caninum can actively invade a large variety of nucleated cells in vitro and in vivo conditions (Naguleswaran et al., 2002; Hemphill et al., 2004; Lei et al., 2005); its mechanisms are very similar to those proposed for T. gondii, where a first stage of low-affinity adhesion and a second stage of firm apical attachment can be observed (Hemphill et al., 2004; Carruthers and Boothroyd, 2007).
The initial adhesion is mediated by tachyzoite surface antigens belonging to the SAG-related sequence proteins (SRS) family and homologous to the T. gondii SAGs family (Howe et al., 1998). SAG1 and SRS2 are two main immuno-dominant antigens of N. caninum tachyzoites (Dong et al., 2012; Sinnott et al., 2017) that binds to host cells (Hemphill et al., 2013). In addition, SRS allows parasite adhesion through binding to GAGs expressed on the host cell’s surface, especially CSA and SA (Dubey et al., 2017). The latter is crucial for self-recognition by complement factor H (Varki and Gagneux, 2012) and fetal defense against maternal complement attack (Abeln et al., 2019).
Then the apical attachment is mediated mainly by MIC (Lovett et al., 2000; Naguleswaran et al., 2002; Reid et al., 2012; Wang J. et al., 2017), MIC2 and MIC19 are unique to N. caninum (Reid et al., 2012). N. caninum MIC proteins have domains similar to those found in vertebrate ECM proteins, including thrombospondin (TPS) -like, integrin-like domain (Pereira et al., 2011) and SA- binding adhesive repeat- (MAR)-like domain (Friedrich et al., 2010). Thus, MICs can bind through GAGs to various receptors present on the surface of target cells (Dubey et al., 2017), and mediate cell adhesion (Keller et al., 2002; Wang J. et al., 2017). MIC1, MIC3, and MIC4 bind to host cells through sulfated proteoglycans (Naguleswaran et al., 2002; Keller et al., 2004; Friedrich et al., 2010). N. caninum proliferation varies between the different cells of the placenta (Gibney et al., 2008). Studies in bovine placenta cell lines have shown that caruncular cells appear to be more resistant to Neospora infection than trophoblasts due to their expression of adhesion molecules (Jiménez-Pelayo et al., 2017, 2019) and phagocytic activity (Machado et al., 2007; Jiménez-Pelayo et al., 2017). However, this resistance also depends on the virulence of the isolate (Jiménez-Pelayo et al., 2017). On the other hand, N. caninum shows a predilection for the fetal chorionic epithelium and placental blood vessels (Buxton et al., 1998).
There are few reports related to the adhesion molecules that participate in the placenta and N. caninum interaction. However, SRS2 participates in the adhesion of N. caninum to the trophoblast (Figure 4A) and the development of an effective immune response against transplacental transmission (Nishikawa et al., 2001; Haldorson et al., 2005, 2006), but the receptor for SRS2 has not yet been identified (Naguleswaran et al., 2002; Figure 4A). Nevertheless, the presence of SA and glycoproteins expressed on the surface of binucleate trophoblast giant cells (BNCs) in the ovine placenta (Jones et al., 1997; Klisch et al., 2006) could be involved in N. caninum adhesion to the placenta (Figure 4A).
Modulation of gene expression in the host cell is also a mechanism for N. caninum to invade and cross the placental barrier (Horcajo et al., 2017; Dorsch et al., 2019). Thus, infection of immortalized bovine trophoblasts with N. caninum induces the expression MMPs involved in ECM degradation (Cui et al., 2017; Horcajo et al., 2017). Additionally, infection by N. caninum alters the glycosylation pattern in the glycocalyx and apical cytoplasm of trophoblast (Dorsch et al., 2019; Figure 4A). The BNCs and the uterine epithelium also shows changes in sugars, including α-D-GalNAc, α-D-Man, β-D-Gal, α-D-Gluc, and NeuNac (Vonlaufen et al., 2004; Dorsch et al., 2019; Figure 4A). The expression of the adhesion molecules E-selectin, P-selectin, VCAM-1, and ICAM-1 is also altered, possibly through paracrine activation, through the pro-inflammatory cascade activation, which involves the release of IL-1b, IL-8, and MCP-1 leading to cell activation and subsequent expression of the adhesion molecules (Taubert et al., 2006).
Neospora caninum can also hijack immune cells to circumvent biological barriers. Thus, infection of DCs by tachyzoites enhances the translocation of parasites across cell monolayers (Collantes-Fernandez et al., 2012; Figure 4A). Furthermore, similar to T. gondii (Lambert et al., 2006; Lambert and Barragan, 2010), infection of DCs by N. caninum induce a migratory phenotype in these cells, enhancing its dissemination. In addition, N. caninum can infect MΦ and NK, cells and take advantage of their basal or inducible mobile properties (Boysen et al., 2006; Dion et al., 2011) (Figures 4A,B).
Immune Response Against Neospora
The immune response at the maternal-fetal interface against N. caninum is characterized by a mixture of Th1 and Th2 responses, implying the presence of pro-and anti-inflammatory cytokines (Cantón et al., 2014b; Hecker et al., 2015; Arranz-Solís et al., 2016; Gutiérrez-Expósito et al., 2020; Figure 4B). Although these responses play a vital role in controlling parasite multiplication, they are responsible for placental damage that might lead to abortion (Maley et al., 2006; Rosbottom et al., 2011; Cantón et al., 2014a; Gutiérrez-Expósito et al., 2020).
The role of the innate immune response in the placenta against N. caninum has been studied mainly in cattle. The parasite is recognized by endosomal TLRs (3, 7, 8, and 9), which are upregulated in response to Neospora infection (Figure 4B; Marin et al., 2017).
Similar to T. gondii infection, TLR 3/7 and 8 is associated with the control of N. caninum intracellular infection. TLR7 activation leads to the development of a protective Th1 response (Andrade et al., 2013; Castillo et al., 2018) through MyD88-dependent signaling (Mineo et al., 2009; Miranda et al., 2019) inducing the secretion of pro-inflammatory cytokines including IFN-γ, IL-12, IL-18, and TNF-α, and promotes recruitment of T lymphocyte (CD3, CD4, CD8, and γδ) subsets, MΦ, NK cells (NKp46 subpopulation) (Maley et al., 2006; Cantón et al., 2014a, b; Hecker et al., 2015; Arranz-Solís et al., 2016; Gutiérrez-Expósito et al., 2020; Figure 4B).
In addition, the Th2 (IL-4) and regulatory (IL-10) cytokines also show alterations during N. caninum infection (López-Pérez et al., 2010; Rosbottom et al., 2011). Thus, the increase of IL-4 attenuates the effects of the pro-inflammatory response and enhances susceptibility to N. caninum favoring congenital transmission (López-Pérez et al., 2010). On the other hand, IL-10 has a regulatory effect on IFN-γ and TNF-α mediated response (Eperon et al., 1999; Quinn et al., 2004) and might also promote the invasion and intracellular replication of N. caninum and consequently its transmission.
Conclusion
Apicomplexa are a large group of intracellular parasites that are distributed globally. Some of those parasites pose significant risks to pregnancy in humans and animals. The interaction between the parasites and hosts is the most critical factor in determining the success or failure of infection. During congenital infection, the parasites must break through the placental barrier and modulate host defenses. Apicomplexan parasites, despite having some differences during their life cycle, these parasites use very similar adhesion mechanisms to the placenta, based fundamentally on the alteration and modulation of the expression of molecules present on the surface of placental cells, including the trophoblast. Additionally, the presence of these parasites induces alterations in the immune response and a change in the Th1/Th2 balance, promoting the activation of defense mechanisms in the placenta based fundamentally on cellular immunity. This cellular immunity, mediated by different populations of T lymphocytes, secretion of chemokines, and pro-inflammatory cytokines, negatively affect the placental function and fetal growth that might ultimately cause fetal death and abortion.
Thus, better knowledge about changes in host gene expression, parasite strategies for modulating the host’s immune system, among others, should clarify the mechanisms of congenital transmission of those parasites.
Author Contributions
MR-P prepared the draft of the manuscript the figures. LM, MR, AL, CC, EP-P, JG-M, and SA corrected the draft and figures and participated in the different rounds of correcting the manuscript. UK corrected the manuscript into its final version for submission.
Funding
MR-P was funded by a postdoctoral fellowship from the Dirección de Investigación, Pontificia Universidad Católica de Valparaíso, Chile. ERANET-LAC grant ERANET17/HLH-0142 (UK), and “Fondo Nacional de Desarrollo Científico y Tecnológico” (FONDECYT, Chile) grants 1190341 (UK) and 3180452 (CC).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abbasi, M., Kowalewska-Grochowska, K., Bahar, M. A., Kilani, R. T., Winkler-Lowen, B., and Guilbert, L. J. (2003). Infection of placental trophoblasts by Toxoplasma gondii. J. Infect. Dis. 188, 608–616. doi: 10.1086/377132
Abeln, M., Albers, I., Peters-Bernard, U., Flächsig-Schulz, K., Kats, E., Kispert, A., et al. (2019). Sialic acid is a critical fetal defense against maternal complement attack. J. Clin. Invest. 129, 422–436. doi: 10.1172/JCI99945
Abrams, E. T., Brown, H., Chensue, S. W., Turner, G. D. H., Tadesse, E., Lema, V. M., et al. (2003). Host response to malaria during pregnancy: placental monocyte recruitment is associated with elevated beta chemokine expression. J. Immunol. 170, 2759–2764. doi: 10.4049/jimmunol.170.5.2759
Aderinboye, O., and Syed, S. S. (2010). Congenital babesiosis in a four-week-old female infant. Pediatr. Infect. Dis. J. 29:188. doi: 10.1097/INF.0b013e3181c3c971
Aikawa, M., Pongponratn, E., Tegoshi, T., Nakamura, K.-I., Nagatake, T., Cochrane, A., et al. (1992). A study on the pathogenesis of human cerebral malaria and cerebral babesiosis. Memórias Instituto Oswaldo Cruz 87, 297–301. doi: 10.1590/S0074-02761992000700051
Allred, D. R. (2019). Variable and Variant Protein Multigene Families in Babesia bovis Persistence. Pathogens 8:athogens8020076. doi: 10.3390/pathogens8020076
Allred, D. R., Carlton, J. M., Satcher, R. L., Long, J. A., Brown, W. C., Patterson, P. E., et al. (2000). The ves multigene family of B. bovis encodes components of rapid antigenic variation at the infected erythrocyte surface. Mol. Cell 5, 153–162. doi: 10.1016/s1097-2765(00)80411-6
Al-Shaeli, S. J. J., Ethaeb, A. M., and Gharban, H. A. J. (2020). Molecular and histopathological identification of ovine neosporosis (Neospora caninum) in aborted ewes in Iraq. Vet. World 13, 597–603. doi: 10.14202/vetworld.2020.597-603
Ander, S. E., Rudzki, E. N., Arora, N., Sadovsky, Y., Coyne, C. B., and Boyle, J. P. (2018). Human Placental Syncytiotrophoblasts Restrict Toxoplasma gondii Attachment and Replication and Respond to Infection by Producing Immunomodulatory Chemokines. mBio 9, 1678–1617. doi: 10.1128/mBio.01678-17
Andrade, W. A., Souza, M., do, C., Martinez, E. R., Nagpal, K., Dutra, M. S., et al. (2013). Combined action of Nucleic Acid-Sensing Toll-Like Receptors (TLRs) and TLR11/TLR12 heterodimers imparts resistance to Toxoplasma gondii in mice. Cell Host Microbe 13, 42–53. doi: 10.1016/j.chom.2012.12.003
Arranz-Solís, D., Benavides, J., Regidor-Cerrillo, J., Horcajo, P., Castaño, P., del Carmen, et al. (2016). Systemic and local immune responses in sheep after Neospora caninum experimental infection at early, mid and late gestation. Vet. Res. 47:0290. doi: 10.1186/s13567-015-0290-0
Arranz-Solís, D., Mukhopadhyay, D., and Saeij, J. J. P. (2021). Toxoplasma Effectors that Affect Pregnancy Outcome. Trends Parasitol. 37, 283–295. doi: 10.1016/j.pt.2020.10.013
Autino, B., Corbett, Y., Castelli, F., and Taramelli, D. (2012). Pathogenesis of Malaria in Tissues and Blood. Mediterr. J. Hematol. Infect. Dis. 4:061. doi: 10.4084/MJHID.2012.061
Azimi-Nezhad, M. (2014). Vascular endothelial growth factor from embryonic status to cardiovascular pathology. Rep. Biochem. Mol. Biol. 2, 59–69.
Azzouz, N., Kamena, F., Laurino, P., Kikkeri, R., Mercier, C., Cesbron-Delauw, M.-F., et al. (2013). Toxoplasma gondii secretory proteins bind to sulfated heparin structures. Glycobiology 23, 106–120. doi: 10.1093/glycob/cws134
Baba, M., Batanova, T., Kitoh, K., and Takashima, Y. (2017). Adhesion of Toxoplasma gondii tachyzoite-infected vehicle leukocytes to capillary endothelial cells triggers timely parasite egression. Sci. Rep. 7:5675. doi: 10.1038/s41598-017-05956-z
Bachmeyer, C., Mouchnino, G., Thulliez, P., and Blum, L. (2006). Congenital toxoplasmosis from an HIV-infected woman as a result of reactivation. J. Infect. 52, e55–e57. doi: 10.1016/j.jinf.2005.05.004
Barbosa, B. F., Paulesu, L., Ietta, F., Bechi, N., Romagnoli, R., Gomes, A. O., et al. (2014). Susceptibility to Toxoplasma gondii proliferation in BeWo human trophoblast cells is dose-dependent of macrophage migration inhibitory factor (MIF), via ERK1/2 phosphorylation and prostaglandin E2 production. Placenta 35, 152–162. doi: 10.1016/j.placenta.2013.12.013
Barboza, R., Lima, F. A., Reis, A. S., Murillo, O. J., Peixoto, E. P. M., Bandeira, C. L., et al. (2017). TLR4-Mediated Placental Pathology and Pregnancy Outcome in Experimental Malaria. Sci. Rep. 7:8623. doi: 10.1038/s41598-017-08299-x
Barboza, R., Reis, A. S., da Silva, L. G., Hasenkamp, L., Pereira, K. R. B., Câmara, N. O. S., et al. (2014). MyD88 signaling is directly involved in the development of murine placental malaria. Infect. Immun. 82, 830–838. doi: 10.1128/IAI.01288-13
Barfod, L., Dalgaard, M. B., Pleman, S. T., Ofori, M. F., Pleass, R. J., and Hviid, L. (2011). Evasion of immunity to Plasmodium falciparum malaria by IgM masking of protective IgG epitopes in infected erythrocyte surface-exposed PfEMP1. PNAS 108, 12485–12490. doi: 10.1073/pnas.1103708108
Barr, B. C., Conrad, P. A., Sverlow, K. W., Tarantal, A. F., and Hendrickx, A. G. (1994). Experimental fetal and transplacental Neospora infection in the nonhuman primate. Lab. Invest. 71, 236–242.
Barragan, A., Brossier, F., and Sibley, L. D. (2005). Transepithelial migration of Toxoplasma gondii involves an interaction of intercellular adhesion molecule 1 (ICAM-1) with the parasite adhesin MIC2. Cell. Microbiol. 7, 561–568. doi: 10.1111/j.1462-5822.2005.00486.x
Baruch, D. I., Gormely, J. A., Ma, C., Howard, R. J., and Pasloske, B. L. (1996). Plasmodium falciparum erythrocyte membrane protein 1 is a parasitized erythrocyte receptor for adherence to CD36, thrombospondin, and intercellular adhesion molecule 1. Proc. Natl. Acad. Sci. U S A. 93, 3497–3502. doi: 10.1073/pnas.93.8.3497
Beeson, J. G., Amin, N., Kanjala, M., and Rogerson, S. J. (2002). Selective Accumulation of Mature Asexual Stages of Plasmodium falciparum-Infected Erythrocytes in the Placenta. Infect. Immun. 70, 5412–5415. doi: 10.1128/IAI.70.10.5412-5415.2002
Beeson, J. G., and Brown, G. V. (2004). Plasmodium falciparum-infected erythrocytes demonstrate dual specificity for adhesion to hyaluronic acid and chondroitin sulfate A and have distinct adhesive properties. J. Infect. Dis. 189, 169–179. doi: 10.1086/380975
Beeson, J. G., Brown, G. V., Molyneux, M. E., Mhango, C., Dzinjalamala, F., and Rogerson, S. J. (1999). Plasmodium falciparum Isolates from Infected Pregnant Women and Children Are Associated with Distinct Adhesive and Antigenic Properties. J. Infect. Dis. 180, 464–472. doi: 10.1086/314899
Benavente, E. D., Oresegun, D. R., de Sessions, P. F., Walker, E. M., Roper, C., Dombrowski, J. G., et al. (2018). Global genetic diversity of var2csa in Plasmodium falciparum with implications for malaria in pregnancy and vaccine development. Sci. Rep. 8:15429. doi: 10.1038/s41598-018-33767-3
Benirschke, K., Kaufmann, P., and Baergen, P. (2012). Pathology of the Human Placenta, 6 th Edn. New York, NY: SpringerLink.
Berger, S. S., Turner, L., Wang, C. W., Petersen, J. E. V., Kraft, M., Lusingu, J. P. A., et al. (2013). Plasmodium falciparum expressing domain cassette 5 type PfEMP1 (DC5-PfEMP1) bind PECAM1. PLoS One 8:e69117. doi: 10.1371/journal.pone.0069117
Bhatia, R., Rajwaniya, D., and Agrawal, P. (2016). Congenital Malaria due to Plasmodium Vivax Infection in a Neonate. Case Rep. Pediatr. 2016:1929046. doi: 10.1155/2016/1929046
Blader, I. J., and Saeij, J. P. (2009). Communication between Toxoplasma gondii and its host: impact on parasite growth, development, immune evasion, and virulence. APMIS 117, 458–476. doi: 10.1111/j.1600-0463.2009.02453.x
Blumenschein, T. M. A., Friedrich, N., Childs, R. A., Saouros, S., Carpenter, E. P., Campanero-Rhodes, M. A., et al. (2007). Atomic resolution insight into host cell recognition by Toxoplasma gondii. EMBO J. 26, 2808–2820. doi: 10.1038/sj.emboj.7601704
Bosman, A.-M., Oosthuizen, M. C., Venter, E. H., Steyl, J. C., Gous, T. A., and Penzhorn, B. L. (2013). Babesia lengau associated with cerebral and haemolytic babesiosis in two domestic cats. Parasit. Vectors 6:128. doi: 10.1186/1756-3305-6-128
Boysen, P., Klevar, S., Olsen, I., and Storset, A. K. (2006). The Protozoan Neospora caninum Directly Triggers Bovine NK Cells To Produce Gamma Interferon and To Kill Infected Fibroblasts. Infect. Immun. 74, 953–960. doi: 10.1128/IAI.74.2.953-960.2006
Briand, V., Le Hesran, J.-Y., Mayxay, M., Newton, P. N., Bertin, G., Houzé, S., et al. (2016). Prevalence of malaria in pregnancy in southern Laos: a cross-sectional survey. Malaria J. 15:436. doi: 10.1186/s12936-016-1492-2
Buxton, D., Maley, S. W., Wright, S., Thomson, K. M., Rae, A. G., and Innes, E. A. (1998). The pathogenesis of experimental neosporosis in pregnant sheep. J. Comparat. Pathol. 118, 267–279. doi: 10.1016/S0021-9975(07)80003-X
Cantón, G. J., Katzer, F., Maley, S. W., Bartley, P. M., Benavides-Silván, J., Palarea-Albaladejo, J., et al. (2014a). Cytokine expression in the placenta of pregnant cattle after inoculation with Neospora caninum. Vet. Immunol. Immunopathol. 161, 77–89. doi: 10.1016/j.vetimm.2014.07.004
Cantón, G. J., Katzer, F., Maley, S. W., Bartley, P. M., Benavides-Silván, J., Palarea-Albaladejo, J., et al. (2014b). Inflammatory infiltration into placentas of Neospora caninum challenged cattle correlates with clinical outcome of pregnancy. Vet. Res. 45:11. doi: 10.1186/1297-9716-45-11
Capellini, I., Nunn, C. L., and Barton, R. A. (2015). Microparasites and Placental Invasiveness in Eutherian Mammals. PLoS One 10:e0132563. doi: 10.1371/journal.pone.0132563
Carlier, Y., Truyens, C., Deloron, P., and Peyron, F. (2012). Congenital parasitic infections: A review. Acta Tropica 121, 55–70. doi: 10.1016/j.actatropica.2011.10.018
Carruthers, V., and Boothroyd, J. C. (2007). Pulling together: an integrated model of Toxoplasma cell invasion. Curr. Opin. Microbiol. 10, 83–89. doi: 10.1016/j.mib.2006.06.017
Castillo, C., Liempi, A., Medina, L., and Kemmerling, I. C. (2018). Antiparasitic Mechanisms of the Human Placenta. Chagas Dis. Basic Investigat. Challenges 2018:73569. doi: 10.5772/intechopen.73569
Castillo, C., Muñoz, L., Carrillo, I., Liempi, A., Gallardo, C., Galanti, N., et al. (2017a). Ex vivo infection of human placental chorionic villi explants with Trypanosoma cruzi and Toxoplasma gondii induces different Toll-like receptor expression and cytokine/chemokine profiles. Am. J. Reprod. Immunol. 78:12660. doi: 10.1111/aji.12660
Castillo, C., Muñoz, L., Carrillo, I., Liempi, A., Medina, L., Galanti, N., et al. (2017b). Toll-like receptor-2 mediates local innate immune response against Trypanosoma cruzi in ex vivo infected human placental chorionic villi explants. Placenta 60, 40–46. doi: 10.1016/j.placenta.2017.10.005
Chakraborty, A. (2016). Understanding the biology of the Plasmodium falciparum apicoplast; an excellent target for antimalarial drug development. Life Sci. 158, 104–110. doi: 10.1016/j.lfs.2016.06.030
Chakraborty, S., Roy, S., Mistry, H. U., Murthy, S., George, N., Bhandari, V., et al. (2017). Potential Sabotage of Host Cell Physiology by Apicomplexan Parasites for Their Survival Benefits. Front. Immunol. 8:01261. doi: 10.3389/fimmu.2017.01261
Chan, J.-A., Fowkes, F. J. I., and Beeson, J. G. (2014). Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cell. Mol. Life Sci. 71, 3633–3657. doi: 10.1007/s00018-014-1614-3
Chau, K., Hennessy, A., and Makris, A. (2017). Placental growth factor and pre-eclampsia. J. Hum. Hypertens 31, 782–786. doi: 10.1038/jhh.2017.61
Chen, C.-P., and Aplin, J. D. (2003). Placental extracellular matrix: gene expression, deposition by placental fibroblasts and the effect of oxygen. Placenta 24, 316–325. doi: 10.1053/plac.2002.0904
Chêne, A., Briand, V., Ibitokou, S., Dechavanne, S., Massougbodji, A., Deloron, P., et al. (2014). Placental cytokine and chemokine profiles reflect pregnancy outcomes in women exposed to Plasmodium falciparum infection. Infect. Immun. 82, 3783–3789. doi: 10.1128/IAI.01922-14
Chesnokov, O., Merritt, J., Tcherniuk, S. O., Milman, N., and Oleinikov, A. V. (2018). Plasmodium falciparum infected erythrocytes can bind to host receptors integrins αVβ3 and αVβ6 through DBLδ1_D4 domain of PFL2665c PfEMP1 protein. Sci. Rep. 8:17871. doi: 10.1038/s41598-018-36071-2
Chishti, A. H. (2015). Malaria selectively targets pregnancy receptors. Blood 125, 217–218. doi: 10.1182/blood-2014-11-610634
Chotivanich, K., Udomsangpetch, R., Suwanarusk, R., Pukrittayakamee, S., Wilairatana, P., Beeson, J. G., et al. (2012). Plasmodium vivax Adherence to Placental Glycosaminoglycans. PLoS One 7:0034509. doi: 10.1371/journal.pone.0034509
Clark, R. L. (2019). Genesis of placental sequestration in malaria and possible targets for drugs for placental malaria. Birth Defects Res. 111, 569–583. doi: 10.1002/bdr2.1496
Coban, C., Ishii, K. J., Kawai, T., Hemmi, H., Sato, S., Uematsu, S., et al. (2005). Toll-like receptor 9 mediates innate immune activation by the malaria pigment hemozoin. J. Exp. Med. 201, 19–25. doi: 10.1084/jem.20041836
Cole, R. A., Lindsay, D. S., Blagburn, B. L., and Dubey, J. P. (1995). Vertical Transmission of Neospora caninum in Mice. J. Parasitol. 81, 730–732. doi: 10.2307/3283962
Collantes-Fernandez, E., Arrighi, R. B. G., Álvarez-García, G., Weidner, J. M., Regidor-Cerrillo, J., Boothroyd, J. C., et al. (2012). Infected Dendritic Cells Facilitate Systemic Dissemination and Transplacental Passage of the Obligate Intracellular Parasite Neospora caninum in Mice. PLoS One 7:e32123. doi: 10.1371/journal.pone.0032123
Collins, W. E., and Jeffery, G. M. (2005). Plasmodium ovale: parasite and disease. Clin. Microbiol. Rev. 18, 570–581. doi: 10.1128/CMR.18.3.570-581.2005
Conroy, A. L., McDonald, C. R., Silver, K. L., Liles, W. C., and Kain, K. C. (2011). Complement activation: a critical mediator of adverse fetal outcomes in placental malaria? Trends Parasitol. 27, 294–299. doi: 10.1016/j.pt.2011.02.005
Cooke, B. M., Mohandas, N., Cowman, A. F., and Coppel, R. L. (2005). Cellular adhesive phenomena in apicomplexan parasites of red blood cells. Vet. Parasitol. 132, 273–295. doi: 10.1016/j.vetpar.2005.07.002
Cornett, J. K., Malhotra, A., and Hart, D. (2012). Vertical Transmission of Babesiosis From a Pregnant, Splenectomized Mother to Her Neonate. Infect. Dis. Clin. Pract. 20, 408–410. doi: 10.1097/IPC.0b013e31825b20c1
Cox, F. E. G. (1982). “Babesiosis in Rodents and Humans,” in Animal Models in Parasitology: A Symposium held at the Royal Zoological Society, Regents Park, London, in March 1981, ed. D. G. Owen (London: Macmillan Education UK), 83–91. doi: 10.1007/978-1-349-06136-5_5
Crawford, J., Grujic, O., Bruic, E., Czjzek, M., Grigg, M. E., and Boulanger, M. J. (2009). Structural Characterization of the Bradyzoite Surface Antigen (BSR4) from Toxoplasma gondii, a Unique Addition to the Surface Antigen Glycoprotein 1-related Superfamily. J. Biol. Chem. 284, 9192–9198. doi: 10.1074/jbc.M808714200
Cui, N., Hu, M., and Khalil, R. A. (2017). Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 147, 1–73. doi: 10.1016/bs.pmbts.2017.02.005
de Oliveira Gomes, A., de Oliveira Silva, D. A., Silva, N. M., de Freitas Barbosa, B., Franco, P. S., Angeloni, M. B., et al. (2011). Effect of Macrophage Migration Inhibitory Factor (MIF) in Human Placental Explants Infected with Toxoplasma gondii Depends on Gestational Age. Am. J. Pathol. 178, 2792–2801. doi: 10.1016/j.ajpath.2011.02.005
De Silva, D. H. G., Mendis, K. N., Premaratne, U. N., Jayatilleke, S. M. D., and Soyza, P. E. (1982). Congenital malaria due to Plasmodium vivax: a case report from Sri Lanka. Transact. R. Soc. Trop. Med. Hygiene 76, 33–35. doi: 10.1016/0035-9203(82)90011-6
Debierre-Grockiego, F., Campos, M. A., Azzouz, N., Schmidt, J., Bieker, U., Resende, M. G., et al. (2007). Activation of TLR2 and TLR4 by Glycosylphosphatidylinositols Derived from Toxoplasma gondii. J. Immunol. 179, 1129–1137.
del Campo, J., Heger, T. J., Rodríguez-Martínez, R., Worden, A. Z., Richards, T. A., Massana, R., et al. (2019). Assessing the Diversity and Distribution of Apicomplexans in Host and Free-Living Environments Using High-Throughput Amplicon Data and a Phylogenetically Informed Reference Framework. Front. Microbiol. 10:02373. doi: 10.3389/fmicb.2019.02373
Denkers, E. Y. (2010). Toll-Like Receptor Initiated Host Defense against Toxoplasma gondii. J. Biomed. Biotechnol. 2010:737125. doi: 10.1155/2010/737125
Diallo, M., Aldebert, D., Moreau, J.-C., Ndiaye, M., and Jambou, R. (2008). Decrease of lymphoid dendritic cells in blood from malaria-infected pregnant women. Int. J. Parasitol. 38, 1557–1565. doi: 10.1016/j.ijpara.2008.04.013
Dion, S., Germon, S., Guiton, R., Ducournau, C., and Dimier-Poisson, I. (2011). Functional activation of T cells by dendritic cells and macrophages exposed to the intracellular parasite Neospora caninum. Int. J. Parasitol. 41, 685–695. doi: 10.1016/j.ijpara.2011.01.008
Dobaño, C., Berthoud, T., Manaca, M. N., Nhabomba, A., Guinovart, C., Aguilar, R., et al. (2018). High production of pro-inflammatory cytokines by maternal blood mononuclear cells is associated with reduced maternal malaria but increased cord blood infection. Malar. J. 17:177. doi: 10.1186/s12936-018-2317-2
Dobbs, K. R., and Dent, A. E. (2016). Plasmodium malaria and antimalarial antibodies in the first year of life. Parasitology 143, 129–138. doi: 10.1017/S0031182015001626
Dombrowski, J. G., de Souza, R. M., Silva, N. R. M., Barateiro, A., Epiphanio, S., Gonçalves, L. A., et al. (2018). Malaria during pregnancy and newborn outcome in an unstable transmission area in Brazil: A population-based record linkage study. PLoS One 13:e0199415. doi: 10.1371/journal.pone.0199415
Donahoe, S. L., Lindsay, S. A., Krockenberger, M., Phalen, D., and Šlapeta, J. (2015). A review of neosporosis and pathologic findings of Neospora caninum infection in wildlife. Int. J. Parasitol. Parasites Wildl. 4, 216–238. doi: 10.1016/j.ijppaw.2015.04.002
Dong, J., Otsuki, T., Kato, T., and Park, E. Y. (2012). Development of a diagnostic method for neosporosis in cattle using recombinant Neospora caninum proteins. BMC Biotechnol. 12:19. doi: 10.1186/1472-6750-12-19
Dorsch, M. A., de Yaniz, M. G., Fiorani, F., Hecker, Y. P., Odeón, A. C., Morrell, E. L., et al. (2019). A Descriptive Study of Lectin Histochemistry of the Placenta in Cattle following Inoculation of Neospora caninum. J. Comparat. Pathol. 166, 45–53. doi: 10.1016/j.jcpa.2018.10.172
Dos Santos, T. R., Faria, G., da, S. M., Guerreiro, B. M., Dal Pietro, N. H. P., da, S., et al. (2016). Congenital Toxoplasmosis in Chronically Infected and Subsequently Challenged Ewes. PLoS One 11:e0165124. doi: 10.1371/journal.pone.0165124
Duarte, P. O., Oshiro, L. M., Zimmermann, N. P., Csordas, B. G., Dourado, D. M., Barros, J. C., et al. (2020). Serological and molecular detection of Neospora caninum and Toxoplasma gondii in human umbilical cord blood and placental tissue samples. Sci. Rep. 10, 65991–65991. doi: 10.1038/s41598-020-65991-1
Duaso, J., Rojo, G., Cabrera, G., Galanti, N., Bosco, C., Maya, J. D., et al. (2010). Trypanosoma cruzi induces tissue disorganization and destruction of chorionic villi in an ex vivo infection model of human placenta. Placenta 31, 705–711. doi: 10.1016/j.placenta.2010.05.007
Dubey, J. P. (2003). Review of Neospora caninum and neosporosis in animals. Kor. J. Parasitol. 41, 1–16. doi: 10.3347/kjp.2003.41.1.1
Dubey, J. P., Graham, D. H., Dahl, E., Sreekumar, C., Lehmann, T., Davis, M. F., et al. (2003). Toxoplasma gondii isolates from free-ranging chickens from the United States. J. Parasitol. 89, 1060–1062. doi: 10.1645/GE-124R
Dubey, J. P., Hemphill, A., Calero-Bernal, R., and Schares, G. (2017). Neosporosis in animals, 1st Edn. Florida, FL: CRC Press.
Dubey, J. P., Schares, G., and Ortega-Mora, L. M. (2007). Epidemiology and Control of Neosporosis and Neospora caninum. Clin. Microbiol. Rev. 20, 323–367. doi: 10.1128/CMR.00031-06
Dubey, J. P., Verma, S. K., Ferreira, L. R., Oliveira, S., Cassinelli, A. B., Ying, Y., et al. (2014). Detection and survival of Toxoplasma gondii in milk and cheese from experimentally infected goats. J. Food Prot. 77, 1747–1753. doi: 10.4315/0362-028X.JFP-14-167
Duncanson, P., Terry, R. S., Smith, J. E., and Hide, G. (2001). High levels of congenital transmission of Toxoplasma gondii in a commercial sheep flock. Int. J. Parasitol. 31, 1699–1703. doi: 10.1016/S0020-7519(01)00282-X
Efstratiou, A., Galon, E. M. S., Wang, G., Umeda, K., Kondoh, D., Terkawi, M. A., et al. (2020). Babesia microti Confers Macrophage-Based Cross-Protective Immunity Against Murine Malaria. Front. Cell. Infect. Microbiol. 10:00193. doi: 10.3389/fcimb.2020.00193
Eperon, S., Brönnimann, K., Hemphill, A., and Gottstein, B. (1999). Susceptibility of B-cell deficient C57BL/6 (microMT) mice to Neospora caninum infection. Parasite Immunol. 21, 225–236. doi: 10.1046/j.1365-3024.1999.00223.x
Esernio-Jenssen, D., Scimeca, P. G., Benach, J. L., and Tenenbaum, M. J. (1987). Transplacental/perinatal babesiosis. J. Pediatr. 110, 570–572. doi: 10.1016/s0022-3476(87)80552-8
Ferro, E. A. V., Mineo, J. R., Ietta, F., Bechi, N., Romagnoli, R., Silva, D. A. O., et al. (2008). Macrophage Migration Inhibitory Factor Is Up-Regulated in Human First-Trimester Placenta Stimulated by Soluble Antigen of Toxoplasma gondii, Resulting in Increased Monocyte Adhesion on Villous Explants. Am. J. Pathol. 172, 50–58. doi: 10.2353/ajpath.2008.070432
Flick, K., Scholander, C., Chen, Q., Fernandez, V., Pouvelle, B., Gysin, J., et al. (2001). Role of nonimmune IgG bound to PfEMP1 in placental malaria. Science 293, 2098–2100. doi: 10.1126/science.1062891
Fortier, B., Aïssi, E., Ajana, F., Dieusart, P., Denis, P., Martin, et al. (1991). Spontaneous abortion and reinfection by Toxoplasma gondii. Lancet 338:444. doi: 10.1016/0140-6736(91)91064-2
Fried, M., Domingo, G. J., Gowda, C. D., Mutabingwa, T. K., and Duffy, P. E. (2006). Plasmodium falciparum: chondroitin sulfate A is the major receptor for adhesion of parasitized erythrocytes in the placenta. Exp. Parasitol. 113, 36–42. doi: 10.1016/j.exppara.2005.12.003
Fried, M., Muga, R. O., Misore, A. O., and Duffy, P. E. (1998). Malaria elicits type 1 cytokines in the human placenta: IFN-gamma and TNF-alpha associated with pregnancy outcomes. J. Immunol. 160, 2523–2530.
Friedrich, N., Santos, J. M., Liu, Y., Palma, A. S., Leon, E., Saouros, S., et al. (2010). Members of a novel protein family containing microneme adhesive repeat domains act as sialic acid-binding lectins during host cell invasion by apicomplexan parasites. J. Biol. Chem. 285, 2064–2076. doi: 10.1074/jbc.M109.060988
Fuentes, I., Rubio, J. M., Ramírez, C., and Alvar, J. (2001). Genotypic Characterization of Toxoplasma gondii Strains Associated with Human Toxoplasmosis in Spain: Direct Analysis from Clinical Samples. J. Clin. Microbiol. 39, 1566–1570. doi: 10.1128/JCM.39.4.1566-1570.2001
Furtado, G. C., Cao, Y., and Joiner, K. A. (1992). Laminin on Toxoplasma gondii mediates parasite binding to the beta 1 integrin receptor alpha 6 beta 1 on human foreskin fibroblasts and Chinese hamster ovary cells. Infect. Immun. 60, 4925–4931. doi: 10.1128/IAI.60.11.4925-4931.1992
Gabrielli, S., Bellina, L., Milardi, G. L., Katende, B. K., Totino, V., Fullin, V., et al. (2016). Malaria in children of Tshimbulu (Western Kasai, Democratic Republic of the Congo): epidemiological data and accuracy of diagnostic assays applied in a limited resource setting. Malar. J. 15:81. doi: 10.1186/s12936-016-1142-8
Gallego-Lopez, G. M., Cooke, B. M., and Suarez, C. E. (2019). Interplay between Attenuation- and Virulence-Factors of Babesia bovis and Their Contribution to the Establishment of Persistent Infections in Cattle. Pathogens 8:athogens8030097. doi: 10.3390/pathogens8030097
Garcia-Lloret, M. I., Winkler-Lowen, B., and Guilbert, L. J. (2000). Monocytes adhering by LFA-1 to placental syncytiotrophoblasts induce local apoptosis via release of TNF-alpha. A model for hematogenous initiation of placental inflammations. J. Leukoc. Biol. 68, 903–908.
Gaw, S. L., Hromatka, B. S., Ngeleza, S., Buarpung, S., Ozarslan, N., Tshefu, A., et al. (2019). Differential Activation of Fetal Hofbauer Cells in Primigravidas Is Associated with Decreased Birth Weight in Symptomatic Placental Malaria. Malar. Res. Treat. 2019:1378174. doi: 10.1155/2019/1378174
Ge, Y. Y., Zhang, L., Zhang, G., Wu, J. P., Tan, M. J., Hu, E., et al. (2008). In pregnant mice, the infection of Toxoplasma gondii causes the decrease of CD4+CD25+-regulatory T cells - GE - 2008 - Parasite Immunology - Wiley Online Library. Parasite Immunol. 30, 471–481.
Geleta, G., and Ketema, T. (2016). Severe Malaria Associated with Plasmodium falciparum and P. vivax among Children in Pawe Hospital. Northwest Ethiopia. Malar. Res. Treat. 2016:1240962. doi: 10.1155/2016/1240962
Gibney, E. H., Kipar, A., Rosbottom, A., Guy, C. S., Smith, R. F., Hetzel, U., et al. (2008). The extent of parasite-associated necrosis in the placenta and foetal tissues of cattle following Neospora caninum infection in early and late gestation correlates with foetal death. Int. J. Parasitol. 38, 579–588. doi: 10.1016/j.ijpara.2007.09.015
Gilbert, R. E., Freeman, K., Lago, E. G., Bahia-Oliveira, L. M. G., Tan, H. K., Wallon, M., et al. (2008). Ocular sequelae of congenital toxoplasmosis in Brazil compared with Europe. PLoS Negl. Trop. Dis. 2:e277. doi: 10.1371/journal.pntd.0000277
Gillrie, M. R., Avril, M., Brazier, A. J., Davis, S. P., Stins, M. F., Smith, J. D., et al. (2015). Diverse functional outcomes of Plasmodium falciparum ligation of EPCR: potential implications for malarial pathogenesis. Cell. Microbiol. 17, 1883–1899. doi: 10.1111/cmi.12479
Gimenez, G., Magalhães, K. G., Belaunzarán, M. L., Poncini, C. V., Lammel, E. M., Gonzalez Cappa, S. M., et al. (2010). Lipids from attenuated and virulent Babesia bovis strains induce differential TLR2-mediated macrophage activation. Mol. Immunol. 47, 747–755. doi: 10.1016/j.molimm.2009.10.014
Gómez-Chávez, F., Cañedo-Solares, I., Ortiz-Alegría, L. B., Flores-García, Y., Figueroa-Damián, R., and Luna-Pastén, H. et al. (2020). A Proinflammatory Immune Response Might Determine Toxoplasma gondii Vertical Transmission and Severity of Clinical Features in Congenitally Infected Newborns. Molecules 26:4955.
Gómez-Chávez, F., Cañedo-Solares, I., Ortiz-Alegría, L. B., Flores-García, Y., Luna-Pastén, H., Figueroa-Damián, R., et al. (2019). Maternal Immune Response During Pregnancy and Vertical Transmission in Human Toxoplasmosis. Front. Immunol. 10:00285. doi: 10.3389/fimmu.2019.00285
González-Warleta, M., Castro-Hermida, J. A., Calvo, C., Pérez, V., Gutiérrez-Expósito, D., Regidor-Cerrillo, J., et al. (2018). Endogenous transplacental transmission of Neospora caninum during successive pregnancies across three generations of naturally infected sheep. Vet. Res. 49:106. doi: 10.1186/s13567-018-0601-3
Griffiths, S. K., and Campbell, J. P. (2015). Placental structure, function and drug transfer. Contin. Educat. Anaesthes. Crit. Care Pain 15, 84–89. doi: 10.1093/bjaceaccp/mku013
Gude, N. M., Roberts, C. T., Kalionis, B., and King, R. G. (2004). Growth and function of the normal human placenta. Thromb. Res. 114, 397–407. doi: 10.1016/j.thromres.2004.06.038
Gutiérrez-Expósito, D., González-Warleta, M., Espinosa, J., Vallejo-García, R., Castro-Hermida, J. A., Calvo, C., et al. (2020). Maternal immune response in the placenta of sheep during recrudescence of natural congenital infection of Neospora caninum. Vet. Parasitol. 285:109204. doi: 10.1016/j.vetpar.2020.109204
Haldorson, G. J., Mathison, B. A., Wenberg, K., Conrad, P. A., Dubey, J. P., Trees, A. J., et al. (2005). Immunization with native surface protein NcSRS2 induces a Th2 immune response and reduces congenital Neospora caninum transmission in mice. Int. J. Parasitol. 35, 1407–1415. doi: 10.1016/j.ijpara.2005.05.013
Haldorson, G. J., Stanton, J. B., Mathison, B. A., Suarez, C. E., and Baszler, T. V. (2006). Neospora caninum: antibodies directed against tachyzoite surface protein NcSRS2 inhibit parasite attachment and invasion of placental trophoblasts in vitro. Exp. Parasitol. 112, 172–178. doi: 10.1016/j.exppara.2005.11.004
Hamann, L., Bedu-Addo, G., Eggelte, T. A., Schumann, R. R., and Mockenhaupt, F. P. (2010). The toll-like receptor 1 variant S248N influences placental malaria. Infect. Genet. Evolut. 10, 785–789. doi: 10.1016/j.meegid.2010.05.005
Harker, K. S., Ueno, N., Wang, T., Bonhomme, C., Liu, W., and Lodoen, M. B. (2013). Toxoplasma gondii modulates the dynamics of human monocyte adhesion to vascular endothelium under fluidic shear stress. J. Leukoc. Biol. 93, 789–800. doi: 10.1189/jlb.1012517
He, X., Grigg, M. E., Boothroyd, J. C., and Garcia, K. C. (2002). Structure of the immunodominant surface antigen from the Toxoplasma gondii SRS superfamily. Nat. Struct. Biol. 9, 606–611. doi: 10.1038/nsb819
Hecker, Y. P., Cantón, G., Regidor-Cerrillo, J., Chianini, F., Morrell, E., Lischinsky, L., et al. (2015). Cell mediated immune responses in the placenta following challenge of vaccinated pregnant heifers with Neospora caninum. Vet. Parasitol. 214, 247–254. doi: 10.1016/j.vetpar.2015.10.015
Hemphill, A., Debache, K., Monney, T., Schorer, M., Guionaud, C., Alaeddine, F., et al. (2013). Proteins mediating the Neospora caninum-host cell interaction as targets for vaccination. Front. Biosci. 5:23–36. doi: 10.2741/e593
Hemphill, A., Vonlaufen, N., Naguleswaran, A., Keller, N., Riesen, M., Guetg, N., et al. (2004). Tissue culture and explant approaches to studying and visualizing Neospora caninum and its interactions with the host cell. Microsc. Microanal. 10, 602–620. doi: 10.1017/S1431927604040930
Hide, G. (2016). Role of vertical transmission of Toxoplasma gondii in prevalence of infection. Expert Rev. Anti Infect. Ther. 14, 335–344. doi: 10.1586/14787210.2016.1146131
Ho, M. S., Barr, B. C., Tarantal, A. F., Lai, L. T., Hendrickx, A. G., Marsh, A. E., et al. (1997). Detection of Neospora from tissues of experimentally infected rhesus macaques by PCR and specific DNA probe hybridization. J. Clin. Microbiol. 35, 1740–1745.
Horcajo, P., Jiménez-Pelayo, L., García-Sánchez, M., Regidor-Cerrillo, J., Collantes-Fernández, E., Rozas, D., et al. (2017). Transcriptome modulation of bovine trophoblast cells in vitro by Neospora caninum. Int. J. Parasitol. 47, 791–799. doi: 10.1016/j.ijpara.2017.08.007
Howe, D. K., Crawford, A. C., Lindsay, D., and Sibley, L. D. (1998). The p29 and p35 immunodominant antigens of Neospora caninum tachyzoites are homologous to the family of surface antigens of Toxoplasma gondii. Infect. Immun. 66, 5322–5328. doi: 10.1128/IAI.66.11.5322-5328.1998
Hsieh, F.-L., Turner, L., Bolla, J. R., Robinson, C. V., Lavstsen, T., and Higgins, M. K. (2016). The structural basis for CD36 binding by the malaria parasite. Nat. Commun. 7:12837. doi: 10.1038/ncomms12837
Hutchings, C. L., Li, A., Fernandez, K. M., Fletcher, T., Jackson, L. A., Molloy, J. B., et al. (2007). New insights into the altered adhesive and mechanical properties of red blood cells parasitized by Babesia bovis. Mol. Microbiol. 65, 1092–1105. doi: 10.1111/j.1365-2958.2007.05850.x
Hutson, S. L., Wheeler, K. M., McLone, D., Frim, D., Penn, R., Swisher, C. N., et al. (2015). Patterns of Hydrocephalus Caused by Congenital Toxoplasma gondii Infection Associate With Parasite Genetics. Clin. Infect. Dis. 61, 1831–1834. doi: 10.1093/cid/civ720
Ibrahim, H. M., Huang, P., Salem, T. A., Talaat, R. M., Nasr, M. I., Xuan, X., et al. (2009). Short report: prevalence of Neospora caninum and Toxoplasma gondii antibodies in northern Egypt. Am. J. Trop. Med. Hyg. 80, 263–267.
Jackson, A. P., Otto, T. D., Darby, A., Ramaprasad, A., Xia, D., Echaide, I. E., et al. (2014). The evolutionary dynamics of variant antigen genes in Babesia reveal a history of genomic innovation underlying host-parasite interaction. Nucleic Acids Res. 42, 7113–7131. doi: 10.1093/nar/gku322
Jacquet, A., Coulon, L., De Nève, J., Daminet, V., Haumont, M., Garcia, L., et al. (2001). The surface antigen SAG3 mediates the attachment of Toxoplasma gondii to cell-surface proteoglycans. Mol. Biochem. Parasitol. 116, 35–44. doi: 10.1016/s0166-6851(01)00297-3
Jeninga, M. D., Quinn, J. E., and Petter, M. (2019). ApiAP2 Transcription Factors in Apicomplexan Parasites. Pathogens 8:athogens8020047. doi: 10.3390/pathogens8020047
Jiménez-Pelayo, L., García-Sánchez, M., Regidor-Cerrillo, J., Horcajo, P., Collantes-Fernández, E., Gómez-Bautista, M., et al. (2017). Differential susceptibility of bovine caruncular and trophoblast cell lines to infection with high and low virulence isolates of Neospora caninum. Parasit. Vectors 10:463. doi: 10.1186/s13071-017-2409-9
Jiménez-Pelayo, L., García-Sánchez, M., Regidor-Cerrillo, J., Horcajo, P., Collantes-Fernández, E., Gómez-Bautista, M., et al. (2019). Immune response profile of caruncular and trophoblast cell lines infected by high- (Nc-Spain7) and low-virulence (Nc-Spain1H) isolates of Neospora caninum. Parasites Vectors 12:218. doi: 10.1186/s13071-019-3466-z
Jones, C. J., Dantzer, V., Leiser, R., Krebs, C., and Stoddart, R. W. (1997). Localisation of glycans in the placenta: a comparative study of epitheliochorial, endotheliochorial, and haemomonochorial placentation. Microsc. Res. Tech. 38, 100–114. doi: 10.1002/(SICI)1097-0029(19970701/15)38:1/2<100::AID-JEMT11<3.0.CO;2-T
Joseph, J. T., Purtill, K., Wong, S. J., Munoz, J., Teal, A., Madison-Antenucci, S., et al. (2012). Vertical transmission of Babesia microti, United States. Emerging Infect. Dis. 18, 1318–1321. doi: 10.3201/eid1808.110988
Juliano, P. B., Blotta, M. H. S. L., and Altemani, A. M. A. (2006). ICAM-1 is overexpressed by villous trophoblasts in placentitis. Placenta 27, 750–757. doi: 10.1016/j.placenta.2005.07.008
Jung, C., Lee, C. Y.-F., and Grigg, M. E. (2004). The SRS superfamily of Toxoplasma surface proteins. Int. J. Parasitol. 34, 285–296. doi: 10.1016/j.ijpara.2003.12.004
Katz, L. M., and Dodd, R. Y. (2019). “Transfusion-Transmitted Diseases,” in Transfusion Medicine and Hemostasis, eds B. H. Shaz and C. D. Hillyer (Amsterdam: Elsevier), 437–453. doi: 10.1016/B978-0-12-813726-0.00073-8
Keller, N., Naguleswaran, A., Cannas, A., Vonlaufen, N., Bienz, M., Björkman, C., et al. (2002). Identification of a Neospora caninum Microneme Protein (NcMIC1) Which Interacts with Sulfated Host Cell Surface Glycosaminoglycans. Infect. Immun. 70, 3187–3198. doi: 10.1128/IAI.70.6.3187-3198.2002
Keller, N., Riesen, M., Naguleswaran, A., Vonlaufen, N., Stettler, R., Leepin, A., et al. (2004). Identification and Characterization of a Neospora caninum Microneme-Associated Protein (NcMIC4) That Exhibits Unique Lactose-Binding Properties. Infect. Immun. 72, 4791–4800. doi: 10.1128/IAI.72.8.4791-4800.2004
Kelly, M. N., Kolls, J. K., Happel, K., Schwartzman, J. D., Schwarzenberger, P., Combe, C., et al. (2005). Interleukin-17/Interleukin-17 Receptor-Mediated Signaling Is Important for Generation of an Optimal Polymorphonuclear Response against Toxoplasma gondii Infection. Infect. Immun. 73, 617–621. doi: 10.1128/IAI.73.1.617-621.2005
Kemmerling, U., Osuna, A., Schijman, A. G., and Truyens, C. (2019). Congenital Transmission of Trypanosoma cruzi: A Review About the Interactions Between the Parasite, the Placenta, the Maternal and the Fetal/Neonatal Immune Responses. Front. Microbiol. 10:1854. doi: 10.3389/fmicb.2019.01854
Khan, A., Shaik, J. S., Sikorski, P., Dubey, J. P., and Grigg, M. E. (2020). Neosporosis: An Overview of Its Molecular Epidemiology and Pathogenesis. Engineering 6, 10–19. doi: 10.1016/j.eng.2019.02.010
Khangura, R. K., Williams, N., Cooper, S., and Prabulos, A.-M. (2019). Babesiosis in Pregnancy: An Imitator of HELLP Syndrome. AJP Rep. 9, e147–e152. doi: 10.1055/s-0039-1687873
Kim, K., Weiss, L. M., and Tanowitz, H. B. (2016). “Parasitic Infections,” in Murray and Nadel’s Textbook of Respiratory Medicine, eds R. J. Mason, A. Slutsky, J. F. Murray, J. A. Nadel, M. Gotway, and V. C. Broaddus (Amsterdam: Elsevier), 682.e–698.e. doi: 10.1016/B978-1-4557-3383-5.00039-7
Klisch, K., Boos, A., Friedrich, M., Herzog, K., Feldmann, M., Sousa, N., et al. (2006). The glycosylation of pregnancy-associated glycoproteins and prolactin-related protein-I in bovine binucleate trophoblast giant cells changes before parturition. Reproduction 132, 791–798. doi: 10.1530/REP-06-0040
Krause, P. J., Daily, J., Telford, S. R., Vannier, E., Lantos, P., and Spielman, A. (2007). Shared features in the pathobiology of babesiosis and malaria. Trends Parasitol. 23, 605–610. doi: 10.1016/j.pt.2007.09.005
Krishnegowda, G., Hajjar, A. M., Zhu, J., Douglass, E. J., Uematsu, S., Akira, S., et al. (2005). Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: cell signaling receptors, glycosylphosphatidylinositol (GPI) structural requirement, and regulation of GPI activity. J. Biol. Chem. 280, 8606–8616. doi: 10.1074/jbc.M413541200
Lachenmaier, S. M., Deli, M. A., Meissner, M., and Liesenfeld, O. (2011). Intracellular transport of Toxoplasma gondii through the blood–brain barrier. J. Neuroimmunol. 232, 119–130. doi: 10.1016/j.jneuroim.2010.10.029
Lachkhem, A., Galal, L., Lahmar, I., Passebosc, K., Riahi, H., Plault, N., et al. (2021). First isolation and genotyping of Toxoplasma gondii strains from domestic animals in Tunisia. Sci. Rep. 11:9328. doi: 10.1038/s41598-021-88751-1
Laha, R., Das, M., and Sen, A. (2015). Morphology, epidemiology, and phylogeny of Babesia: An overview. Trop. Parasitol. 5, 94–100. doi: 10.4103/2229-5070.162490
Lahmar, I., Lachkhem, A., Babba, O., Slama, D., Trabelsi, A., Passebosc-Faure, K., et al. (2020). First isolation and molecular characterization of Toxoplasma gondii strains from human congenital toxoplasmosis cases in Monastir, Tunisia. Sci. Rep. 10:1963. doi: 10.1038/s41598-020-59060-w
Lambert, H., and Barragan, A. (2010). Modelling parasite dissemination: host cell subversion and immune evasion by Toxoplasma gondii. Cell. Microbiol. 12, 292–300. doi: 10.1111/j.1462-5822.2009.01417.x
Lambert, H., Hitziger, N., Dellacasa, I., Svensson, M., and Barragan, A. (2006). Induction of dendritic cell migration upon Toxoplasma gondii infection potentiates parasite dissemination. Cell. Microbiol. 8, 1611–1623. doi: 10.1111/j.1462-5822.2006.00735.x
Lei, Y., Davey, M., and Ellis, J. T. (2005). Attachment and invasion of Toxoplasma gondii and Neospora caninum to epithelial and fibroblast cell lines in vitro. Parasitology 131, 583–590. doi: 10.1017/S0031182005008310
Lekutis, C., Ferguson, D. J., Grigg, M. E., Camps, M., and Boothroyd, J. C. (2001). Surface antigens of Toxoplasma gondii: variations on a theme. Int. J. Parasitol. 31, 1285–1292. doi: 10.1016/s0020-7519(01)00261-2
Lennartz, F., Smith, C., Craig, A. G., and Higgins, M. K. (2019). Structural insights into diverse modes of ICAM-1 binding by Plasmodium falciparum-infected erythrocytes. PNAS 116, 20124–20134. doi: 10.1073/pnas.1911900116
Liempi, A., Castillo, C., Carrillo, I., Muñoz, L., Droguett, D., Galanti, N., et al. (2016). A local innate immune response against Trypanosoma cruzi in the human placenta: The epithelial turnover of the trophoblast. Microb. Pathog. 99, 123–129. doi: 10.1016/j.micpath.2016.08.022
Liempi, A., Castillo, C., Medina, L., Galanti, N., Maya, J. D., Parraguez, V. H., et al. (2020). Comparative ex vivo infection with Trypanosoma cruzi and Toxoplasma gondii of human, canine and ovine placenta: Analysis of tissue damage and infection efficiency. Parasitol. Int. 76:102065. doi: 10.1016/j.parint.2020.102065
Liempi, A., Castillo, C., Medina, L., Rojas, M., Maya, J. D., Parraguez, V. H., et al. (2019). Ex vivo infection of human placental explants with Trypanosoma cruzi and Toxoplasma gondii: Differential activation of NF kappa B signaling pathways. Acta Trop. 199:105153. doi: 10.1016/j.actatropica.2019.105153
Lobato, J., Silva, D. A. O., Mineo, T. W. P., Amaral, J. D. H. F., Segundo, G. R. S., Costa-Cruz, J. M., et al. (2006). Detection of Immunoglobulin G Antibodies to Neospora caninum in Humans: High Seropositivity Rates in Patients Who Are Infected by Human Immunodeficiency Virus or Have Neurological Disorders. Clin. Vaccine Immunol. 13, 84–89. doi: 10.1128/CVI.13.1.84-89.2006
López-Pérez, I. C., Collantes-Fernández, E., Rojo-Montejo, S., Navarro-Lozano, V., Risco-Castillo, V., Pérez-Pérez, V., et al. (2010). Effects of Neospora caninum Infection at Mid-Gestation on Placenta in a Pregnant Mouse Model. J. Parasitol. 96, 1017–1020.
Lovett, J. L., Howe, D. K., and Sibley, L. D. (2000). Molecular characterization of a thrombospondin-related anonymous protein homologue in Neospora caninum. Mol. Biochem. Parasitol. 107, 33–43. doi: 10.1016/S0166-6851(99)00228-5
Lucchi, N. W., Peterson, D. S., and Moore, J. M. (2008). Immunologic activation of human syncytiotrophoblast by Plasmodium falciparum. Malaria J. 7:42. doi: 10.1186/1475-2875-7-42
Lucchi, N. W., Sarr, D., Owino, S. O., Mwalimu, S. M., Peterson, D. S., and Moore, J. M. (2011). Natural hemozoin stimulates syncytiotrophoblast to secrete chemokines and recruit peripheral blood mononuclear cells. Placenta 32, 579–585. doi: 10.1016/j.placenta.2011.05.003
Machado, R. Z., Mineo, T. W. P., Landim, L. P., Carvalho, A. F., Gennari, S. M., and Miglino, M. A. (2007). Possible role of bovine trophoblast giant cells in transplacental transmission of Neospora caninum in cattle. Rev. Bras. Parasitol. Vet. 16, 21–25.
Maciel, E., Siqueira, I., Queiroz, A. C., and Melo, A. (2000). Toxoplasma gondii myelitis in a patient with adult T-cell leukemia-lymphoma. Arquivos Neuro Psiquiatria 58, 1107–1109. doi: 10.1590/S0004-282X2000000600019
Maldonado, Y. A., Read, J. S., and Committee on Infectious Diseases. (2017). Diagnosis, Treatment, and Prevention of Congenital Toxoplasmosis in the United States. Pediatrics 139:3860. doi: 10.1542/peds.2016-3860
Maley, S. W., Buxton, D., Macaldowie, C. N., Anderson, I. E., Wright, S. E., Bartley, P. M., et al. (2006). Characterization of the Immune Response in the Placenta of Cattle Experimentally Infected with Neospora caninum in Early Gestation. J. Comparat. Pathol. 135, 130–141. doi: 10.1016/j.jcpa.2006.07.001
Manger, I. D., Hehl, A. B., and Boothroyd, J. C. (1998). The Surface of Toxoplasma Tachyzoites Is Dominated by a Family of Glycosylphosphatidylinositol-Anchored Antigens Related to SAG1. Infect. Immun. 66, 2237–2244.
Marin, M. S., Hecker, Y. P., Quintana, S., Pérez, S. E., Leunda, M. R., Cantón, G. J., et al. (2017). Toll-like receptors 3, 7 and 8 are upregulated in the placental caruncle and fetal spleen of Neospora caninum experimentally infected cattle. Vet. Parasitol. 236, 58–61. doi: 10.1016/j.vetpar.2017.02.002
Martínez-Ocampo, F. (2018). Genomics of Apicomplexa. Farm Anim. Dis. Recent Omic Trends New Strateg. Treat. 2018:72633. doi: 10.5772/intechopen.72633
Marty, P., Bongain, A., Rahal, A., Thulliez, P., Wasfi, D., Lambert, J. C., et al. (1994). Prenatal diagnosis of severe fetal toxoplasmosis as a result of toxoplasmic reactivation in an HIV-1 seropositive woman. Prenat. Diagn. 14, 414–415. doi: 10.1002/pd.1970140514
Masocha, W., and Kristensson, K. (2012). Passage of parasites across the blood-brain barrier. Virulence 3, 202–212. doi: 10.4161/viru.19178
Matejevic, D., Neudeck, H., Graf, R., Müller, T., and Dietl, J. (2001). Localization of hyaluronan with a hyaluronan-specific hyaluronic acid binding protein in the placenta in pre-eclampsia. Gynecol. Obstet. Invest. 52, 257–259. doi: 10.1159/000052986
Maubert, B., Guilbert, L. J., and Deloron, P. (1997). Cytoadherence of Plasmodium falciparum to intercellular adhesion molecule 1 and chondroitin-4-sulfate expressed by the syncytiotrophoblast in the human placenta. Infect. Immun. 65, 1251–1257.
McAuley, J. B. (2014). Congenital Toxoplasmosis. J. Pediatric. Infect. Dis. Soc. 3, S30–S35. doi: 10.1093/jpids/piu077
McLean, A. R., Ataide, R., Simpson, J. A., Beeson, J. G., and Fowkes, F. J. (2015). Malaria and immunity during pregnancy and postpartum: a tale of two species. Parasitology 142, 999–1015. doi: 10.1017/S0031182015000074
Melamed, J., Eckert, G. U., Spadoni, V. S., Lago, E. G., and Uberti, F. (2010). Ocular manifestations of congenital toxoplasmosis. Eye 24, 528–534. doi: 10.1038/eye.2009.140
Mesquita, L. P., Costa, R. C., Nogueira, C. I., Abreu, C. C., Orlando, D. R., Ascari Junior, I., et al. (2018). Placental lesions associated with abortion and stillbirth in goats naturally infected by Neospora caninum. Pesquisa Veterinária Brasileira 38, 444–449. doi: 10.1590/1678-5150-pvb-4598
Metwally, N. G., Tilly, A.-K., Lubiana, P., Roth, L. K., Dörpinghaus, M., Lorenzen, S., et al. (2017). Characterisation of Plasmodium falciparum populations selected on the human endothelial receptors P-selectin, E-selectin, CD9 and CD151. Sci. Rep. 7:4069. doi: 10.1038/s41598-017-04241-3
Milian, I. C. B., Silva, R. J., Manzan-Martins, C., Barbosa, B. F., Guirelli, P. M., Ribeiro, M., et al. (2019). Increased Toxoplasma gondii Intracellular Proliferation in Human Extravillous Trophoblast Cells (HTR8/SVneo Line) Is Sequentially Triggered by MIF, ERK1/2, and COX-2. Front. Microbiol. 10:852. doi: 10.3389/fmicb.2019.00852
Mineo, T. W. P., Benevides, L., Silva, N. M., and Silva, J. S. (2009). Myeloid differentiation factor 88 is required for resistance to Neospora caninum infection. Vet. Res. 40:2009015. doi: 10.1051/vetres/2009015
Miranda, V. D. S., França, F. B. F., da Costa, M. S., Silva, V. R. S., Mota, C. M., Barros, P., et al. (2019). Toll-Like Receptor 3-TRIF Pathway Activation by Neospora caninum RNA Enhances Infection Control in Mice. Infect. Immun. 87, 739–718. doi: 10.1128/IAI.00739-18
Mondragon, R., Howe, D. K., Dubey, J. P., and Sibley, L. D. (1998). Genotypic analysis of Toxoplasma gondii isolates from pigs. J. Parasitol. 84, 639–641.
Moormann, A. M., Sullivan, A. D., Rochford, R. A., Chensue, S. W., Bock, P. J., Nyirenda, T., et al. (1999). Malaria and pregnancy: placental cytokine expression and its relationship to intrauterine growth retardation. J. Infect. Dis. 180, 1987–1993. doi: 10.1086/315135
Moraes, L. V., de, Tadokoro, C. E., Gómez-Conde, I., Olivieri, D. N., and Penha-Gonçalves, C. (2013). Intravital Placenta Imaging Reveals Microcirculatory Dynamics Impact on Sequestration and Phagocytosis of Plasmodium-Infected Erythrocytes. PLoS Pathog. 9:e1003154. doi: 10.1371/journal.ppat.1003154
Mosqueda, J., Olvera-Ramírez, A., Aguilar-Tipacamú, G., and Cantó, G. (2012). Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 19, 1504–1518. doi: 10.2174/092986712799828355
Multhaup, A., Huppertz, B., Göhner, C., Böhringer, M., Mai, M., Markert, U., et al. (2018). N-cadherin knockdown leads to disruption of trophoblastic and endothelial cell interaction in a 3D cell culture model - New insights in trophoblast invasion failure. Cell Adh. Migr. 12, 259–270. doi: 10.1080/19336918.2017.1386822
Muñoz, L., Castillo, C., Carrillo, I., Salinas, A., Liempi, A., Droguett, D., et al. (2016). The ex vivo infection of human placental chorionic villi explants with Trypanosoma cruzi and Toxoplasma gondii is mediated by different toll-like receptors. Placenta 45:86. doi: 10.1016/j.placenta.2016.06.089
Muthusamy, A., Achur, R. N., Bhavanandan, V. P., Fouda, G. G., Taylor, D. W., and Gowda, D. C. (2004). Plasmodium falciparum-infected erythrocytes adhere both in the intervillous space and on the villous surface of human placenta by binding to the low-sulfated chondroitin sulfate proteoglycan receptor. Am. J. Pathol. 164, 2013–2025. doi: 10.1016/S0002-9440(10)63761-3
Naguleswaran, A., Cannas, A., Keller, N., Vonlaufen, N., Björkman, C., and Hemphill, A. (2002). Vero cell surface proteoglycan interaction with the microneme protein NcMIC(3) mediates adhesion of Neospora caninum tachyzoites to host cells unlike that in Toxoplasma gondii. Int. J. Parasitol. 32, 695–704. doi: 10.1016/s0020-7519(02)00014-0
New, D. L., Quinn, J. B., Qureshi, M. Z., and Sigler, S. J. (1997). Vertically Transmitted Babesiosis. J. Pediatr. 131, 163–164. doi: 10.1016/S0022-3476(97)70143-4
Ngô, H. M., Zhou, Y., Lorenzi, H., Wang, K., Kim, T.-K., Zhou, Y., et al. (2017). Toxoplasma Modulates Signature Pathways of Human Epilepsy, Neurodegeneration & Cancer. Sci. Rep. 7:11496. doi: 10.1038/s41598-017-10675-6
Nishikawa, Y., Xuan, X., Nagasawa, H., Igarashi, I., Fujisaki, K., Otsuka, H., et al. (2001). Prevention of vertical transmission of Neospora caninum in BALB/c mice by recombinant vaccinia virus carrying NcSRS2 gene. Vaccine 19, 1710–1716. doi: 10.1016/s0264-410x(00)00407-2
Nunes, M. C., and Scherf, A. (2007). Plasmodium falciparum during pregnancy: a puzzling parasite tissue adhesion tropism. Parasitology 134, 1863–1869. doi: 10.1017/S0031182007000133
O’Connor, R. M., and Allred, D. R. (2000). Selection of Babesia bovis-infected erythrocytes for adhesion to endothelial cells coselects for altered variant erythrocyte surface antigen isoforms. J. Immunol. 164, 2037–2045. doi: 10.4049/jimmunol.164.4.2037
Okamgba, O. C., Ifeanyichukwu, M. O., Ilesanmi, A. O., and Chigbu, L. N. (2018). Variations in the leukocyte and cytokine profiles between placental and maternal circulation in pregnancy-associated malaria. Res. Rep. Trop. Med. 9, 1–8. doi: 10.2147/RRTM.S137829
Oldenhove, G., Bouladoux, N., Wohlfert, E. A., Hall, J. A., Chou, D., Dos santos, L., et al. (2009). Decrease of Foxp3+ Treg Cell Number and Acquisition of Effector Cell Phenotype during Lethal Infection. Immunity 31, 772–786. doi: 10.1016/j.immuni.2009.10.001
Olupot-Olupot, P., Eregu, E. I. E., Naizuli, K., Ikiror, J., Acom, L., and Burgoine, K. (2018). Neonatal and congenital malaria: a case series in malaria endemic eastern Uganda. Malar. J. 17:171. doi: 10.1186/s12936-018-2327-0
Ordi, J., Ismail, M. R., Ventura, P. J., Kahigwa, E., Hirt, R., Cardesa, A., et al. (1998). Massive chronic intervillositis of the placenta associated with malaria infection. Am. J. Surg. Pathol. 22, 1006–1011. doi: 10.1097/00000478-199808000-00011
Ortiz-Alegría, L. B., Caballero-Ortega, H., Cañedo-Solares, I., Rico-Torres, C. P., Sahagún-Ruiz, A., Medina-Escutia, M. E., et al. (2010). Congenital toxoplasmosis: candidate host immune genes relevant for vertical transmission and pathogenesis. Genes Immun. 11, 363–373. doi: 10.1038/gene.2010.21
Oshiro, L. M., Motta-Castro, A. R. C., Freitas, S. Z., Cunha, R. C., Dittrich, R. L., Meirelles, A. C. F., et al. (2015). Neospora caninum and Toxoplasma gondii serodiagnosis in human immunodeficiency virus carriers. Revista Sociedade Brasileira Med. Trop. 48, 568–572. doi: 10.1590/0037-8682-0151-2015
Ossani, R. A., Borges, H. A. T., Souza, A. P., Sartor, A. A., Miletti, L. C., Federle, M., et al. (2017). Toxoplasma gondii in milk of naturally infected dairy ewes on west mesoregion of Santa Catarina state, Brazil. Arq. Bras. Med. Vet. Zootec. 69, 1294–1300. doi: 10.1590/1678-4162-9177
Ouédraogo, A., Tiono, A. B., Diarra, A., Bougouma, E. C. C., Nébié, I., Konaté, A. T., et al. (2012). Transplacental Transmission of Plasmodium falciparum in a Highly Malaria Endemic Area of Burkina Faso. J. Trop. Med. 2012:109705. doi: 10.1155/2012/109705
Ozarslan, N., Robinson, J. F., and Gaw, S. L. (2019). Circulating Monocytes, Tissue Macrophages, and Malaria. J. Trop. Med. 2019:3720838. doi: 10.1155/2019/3720838
Ozawa, S., Haynie, D. G., Bessias, S., Laing, S. K., Ngamasana, E. L., Yemeke, T. T., et al. (2019). Modeling the Economic Impact of Substandard and Falsified Antimalarials in the Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 100, 1149–1157. doi: 10.4269/ajtmh.18-0334
Pappas, G., Roussos, N., and Falagas, M. E. (2009). Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 39, 1385–1394. doi: 10.1016/j.ijpara.2009.04.003
Parekh, F. K., Davison, B. B., Gamboa, D., Hernandez, J., and Branch, O. H. (2010). Placental Histopathologic Changes Associated with Subclinical Malaria Infection and Its Impact on the Fetal Environment. Am. J. Trop. Med. Hyg. 83, 973–980. doi: 10.4269/ajtmh.2010.09-0445
Parroche, P., Lauw, F. N., Goutagny, N., Latz, E., Monks, B. G., Visintin, A., et al. (2007). Malaria hemozoin is immunologically inert but radically enhances innate responses by presenting malaria DNA to Toll-like receptor 9. Proc. Natl. Acad. Sci. U S A. 104, 1919–1924. doi: 10.1073/pnas.0608745104
Parrodi, F., Wright, I. G., Kerr, J. D., and Dobson, C. (1990). In vitro adherence of erythrocytes infected with Babesia bigemina and Babesia rodhainito thrombospondin. Int. J. Parasitol. 20, 899–903. doi: 10.1016/0020-7519(90)90028-L
Passos, S. T., Silver, J. S., O’Hara, A. C., Sehy, D., Stumhofer, J. S., and Hunter, C. A. (2010). IL-6 promotes NK cell production of IL-17 during toxoplasmosis. J. Immunol. 184, 1776–1783. doi: 10.4049/jimmunol.0901843
Pedroni, M. J., Sondgeroth, K. S., Gallego-Lopez, G. M., Echaide, I., and Lau, A. O. (2013). Comparative transcriptome analysis of geographically distinct virulent and attenuated Babesia bovis strains reveals similar gene expression changes through attenuation. BMC Genomics 14:763. doi: 10.1186/1471-2164-14-763
Pehrson, C., Mathiesen, L., Heno, K. K., Salanti, A., Resende, M., Dzikowski, R., et al. (2016). Adhesion of Plasmodium falciparum infected erythrocytes in ex vivo perfused placental tissue: a novel model of placental malaria. Malaria J. 15:292. doi: 10.1186/s12936-016-1342-2
Penazzato, M., Rampon, O., Canale, E. D., Rossi, A. D., Mazza, A., D’Elia, R., et al. (2007). Congenital Plasmodium ovale malaria in an infant born to HIV positive mother. J. Pediatr. Infect. Dis. 2, 167–169.
Pereira, L. M., Candido-Silva, J. A., De Vries, E., and Yatsuda, A. P. (2011). A new thrombospondin-related anonymous protein homologue in Neospora caninum (NcMIC2-like1). Parasitology 138, 287–297. doi: 10.1017/S0031182010001290
Petersen, E., Lebech, M., Jensen, L., Lind, P., Rask, M., Bagger, P., et al. (1999). Neospora caninum infection and repeated abortions in humans. Emerg. Infect. Dis. 5, 278–280.
Pfaff, A. W., Georges, S., Abou-Bacar, A., Letscher-Bru, V., Klein, J.-P., Mousli, M., et al. (2005). Toxoplasma gondii regulates ICAM-1 mediated monocyte adhesion to trophoblasts. Immunol. Cell Biol. 83, 483–489. doi: 10.1111/j.1440-1711.2005.01356.x
PLoS Neglected Tropical Disease (2020). PLoS Neglected Tropical Diseases. Available online at: https://journals.plos.org/plosntds/s/journal-information.%20Accessed%201%20April%202020. (accessed August 21, 2020).
Pollard, A. M., Onatolu, K. N., Hiller, L., Haldar, K., and Knoll, L. J. (2008). Highly polymorphic family of glycosylphosphatidylinositol-anchored surface antigens with evidence of developmental regulation in Toxoplasma gondii. Infect. Immun. 76, 103–110. doi: 10.1128/IAI.01170-07
Pons, J. C., Sigrand, C., Grangeot-Keros, L., Frydman, R., and Thulliez, P. (1995). [Congenital toxoplasmosis: transmission to the fetus of a pre-pregnancy maternal infection]. Presse. Med. 24, 179–182.
Qiu, J., Wang, L., Zhang, R., Ge, K., Guo, H., Liu, X., et al. (2016). Identification of a TNF-α inducer MIC3 originating from the microneme of non-cystogenic, virulent Toxoplasma gondii. Sci. Rep. 6:39407. doi: 10.1038/srep39407
Quinn, H. E., Miller, C. M. D., and Ellis, J. T. (2004). The cell-mediated immune response to Neospora caninum during pregnancy in the mouse is associated with a bias towards production of interleukin-4. Int. J. Parasitol. 34, 723–732. doi: 10.1016/j.ijpara.2004.01.007
Ralph, S. A., van Dooren, G. G., Waller, R. F., Crawford, M. J., Fraunholz, M. J., Foth, B. J., et al. (2004). Tropical infectious diseases: metabolic maps and functions of the Plasmodium falciparum apicoplast. Nat. Rev. Microbiol. 2, 203–216. doi: 10.1038/nrmicro843
Recht, J., Siqueira, A. M., Monteiro, W. M., Herrera, S. M., Herrera, S., and Lacerda, M. V. G. (2017). Malaria in Brazil, Colombia, Peru and Venezuela: current challenges in malaria control and elimination. Malaria J. 16:273. doi: 10.1186/s12936-017-1925-6
Reichel, M. P., Alejandra Ayanegui-Alcérreca, M., Gondim, L. F. P., and Ellis, J. T. (2013). What is the global economic impact of Neospora caninum in cattle - the billion dollar question. Int. J. Parasitol. 43, 133–142. doi: 10.1016/j.ijpara.2012.10.022
Reid, A. J., Vermont, S. J., Cotton, J. A., Harris, D., Hill-Cawthorne, G. A., Könen-Waisman, S., et al. (2012). Comparative genomics of the apicomplexan parasites Toxoplasma gondii and Neospora caninum: Coccidia differing in host range and transmission strategy. PLoS Pathog. 8:e1002567. doi: 10.1371/journal.ppat.1002567
Reis, A. S., Barboza, R., Murillo, O., Barateiro, A., Peixoto, E. P. M., Lima, F. A., et al. (2020). Inflammasome activation and IL-1 signaling during placental malaria induce poor pregnancy outcomes. Sci. Adv. 6:eaax6346. doi: 10.1126/sciadv.aax6346
Rico-Torres, C. P., Vargas-Villavicencio, J. A., and Correa, D. (2016). Is Toxoplasma gondii type related to clinical outcome in human congenital infection? Systematic and critical review. Eur. J. Clin. Microbiol. Infect. Dis. 35, 1079–1088. doi: 10.1007/s10096-016-2656-2
Ripoll, J. G., Rizvi, M. S., King, R. L., and Daniels, C. E. (2018). Severe Babesia microti infection presenting as multiorgan failure in an immunocompetent host. BMJ Case. Rep. 2018:224647. doi: 10.1136/bcr-2018-224647
Robbins, J. R., Zeldovich, V. B., Poukchanski, A., Boothroyd, J. C., and Bakardjiev, A. I. (2012). Tissue barriers of the human placenta to infection with Toxoplasma gondii. Infect. Immun. 80, 418–428. doi: 10.1128/IAI.05899-11
Robert-Gangneux, F., and Dardé, M.-L. (2012). Epidemiology of and Diagnostic Strategies for Toxoplasmosis. Clin. Microbiol. Rev. 25, 264–296. doi: 10.1128/CMR.05013-11
Robert-Gangneux, F., Murat, J.-B., Fricker-Hidalgo, H., Brenier-Pinchart, M.-P., Gangneux, J.-P., and Pelloux, H. (2011). The placenta: a main role in congenital toxoplasmosis? Trends Parasitol. 27, 530–536. doi: 10.1016/j.pt.2011.09.005
Robinson, D. P., and Klein, S. L. (2012). Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm. Behav. 62, 263–271. doi: 10.1016/j.yhbeh.2012.02.023
Rosbottom, A., Gibney, H., Kaiser, P., Hartley, C., Smith, R. F., Robinson, R., et al. (2011). Up Regulation of the Maternal Immune Response in the Placenta of Cattle Naturally Infected with Neospora caninum. PLoS One 6:e15799. doi: 10.1371/journal.pone.0015799
Rowe, J. H., Ertelt, J. M., Aguilera, M. N., Farrar, M. A., and Way, S. S. (2011). Foxp3(+) regulatory T cell expansion required for sustaining pregnancy compromises host defense against prenatal bacterial pathogens. Cell Host Microbe 10, 54–64. doi: 10.1016/j.chom.2011.06.005
Saetre, K., Godhwani, N., Maria, M., Patel, D., Wang, G., Li, K. I., et al. (2018). Congenital Babesiosis After Maternal Infection With Borrelia burgdorferi and Babesia microti. J. Pediat. Infect. Dis. Soc. 7, e1–e5. doi: 10.1093/jpids/pix074
Safeukui, I., Buffet, P. A., Deplaine, G., Perrot, S., Brousse, V., Sauvanet, A., et al. (2018). Sensing of red blood cells with decreased membrane deformability by the human spleen. Blood Adv. 2, 2581–2587. doi: 10.1182/bloodadvances.2018024562
Salazar Gonzalez, R. M., Shehata, H., O’Connell, M. J., Yang, Y., Moreno-Fernandez, M. E., Chougnet, C. A., et al. (2014). Toxoplasma gondii-Derived Profilin Triggers Human Toll-Like Receptor 5-Dependent Cytokine Production. J. Innate Immun. 6, 685–694. doi: 10.1159/000362367
Salomaki, E. D., and Kolisko, M. (2019). There Is Treasure Everywhere: Reductive Plastid Evolution in Apicomplexa in Light of Their Close Relatives. Biomolecules 9:biom9080378. doi: 10.3390/biom9080378
Sanecka, A., and Frickel, E.-M. (2012). Use and abuse of dendritic cells by Toxoplasma gondii. Virulence 3, 678–689. doi: 10.4161/viru.22833
Sartelet, H., Garraud, O., Rogier, C., Milko-Sartelet, I., Kaboret, Y., Michel, G., et al. (2000). Hyperexpression of ICAM-1 and CD36 in placentas infected with Plasmodium falciparum: a possible role of these molecules in sequestration of infected red blood cells in placentas. Histopathology 36, 62–68. doi: 10.1046/j.1365-2559.2000.00742.x
Sasai, M., and Yamamoto, M. (2019). Innate, adaptive, and cell-autonomous immunity against Toxoplasma gondii infection. Exp. Mol. Med. 51, 1–10. doi: 10.1038/s12276-019-0353-9
Sasai, M., Pradipta, A., and Yamamoto, M. (2018). Host immune responses to Toxoplasma gondii. Int. Immunol. 30:dxy004. doi: 10.1093/intimm/dxy004
Schetters, T. (2019). Mechanisms Involved in the Persistence of Babesia canis Infection in Dogs. Pathogens 8:athogens8030094. doi: 10.3390/pathogens8030094
Sethi, S., Alcid, D., Kesarwala, H., and Tolan, R. W. (2009). Probable Congenital Babesiosis in Infant, New Jersey, USA. Emerg. Infect. Dis. 15, 788–791. doi: 10.3201/eid1505.070808
Sharma, L., and Shukla, G. (2017). Placental Malaria: A New Insight into the Pathophysiology. Front. Med. 4:00117. doi: 10.3389/fmed.2017.00117
Shoda, L. K. M., Kegerreis, K. A., Suarez, C. E., Roditi, I., Corral, R. S., Bertot, G. M., et al. (2001). DNA from Protozoan Parasites Babesia bovis, Trypanosoma cruzi, and T. brucei Is Mitogenic for B Lymphocytes and Stimulates Macrophage Expression of Interleukin-12, Tumor Necrosis Factor Alpha, and Nitric Oxide. Infect. Immun. 69, 2162–2171. doi: 10.1128/IAI.69.4.2162-2171.2001
Shoda, L. K., Palmer, G. H., Florin-Christensen, J., Florin-Christensen, M., Godson, D. L., and Brown, W. C. (2000). Babesia bovis-stimulated macrophages express interleukin-1beta, interleukin-12, tumor necrosis factor alpha, and nitric oxide and inhibit parasite replication in vitro. Infect. Immun. 68, 5139–5145. doi: 10.1128/iai.68.9.5139-5145.2000
Sinnott, F. A., Monte, L. G., Collares, T. F., Silveira, R. M., and Borsuk, S. (2017). Review on the immunological and molecular diagnosis of neosporosis (years 2011-2016). Vet. Parasitol. 239, 19–25. doi: 10.1016/j.vetpar.2017.04.008
Skariah, S., Arnaboldi, P., Dattwyler, R. J., Sultan, A. A., Gaylets, C., Walwyn, O., et al. (2017). Elimination of Babesia microti Is Dependent on Intraerythrocytic Killing and CD4 + T Cells. J.I. 199, 633–642. doi: 10.4049/jimmunol.1601193
Smith, J. D. (2014). The role of PfEMP1 adhesion domain classification in Plasmodium falciparum pathogenesis research. Mol. Biochem. Parasitol. 195, 82–87. doi: 10.1016/j.molbiopara.2014.07.006
Sommerville, C., Richardson, J. M., Williams, R. A. M., Mottram, J. C., Roberts, C. W., Alexander, J., et al. (2013). Biochemical and Immunological Characterization of Toxoplasma gondii Macrophage Migration Inhibitory Factor. J. Biol. Chem. 288, 12733–12741. doi: 10.1074/jbc.M112.419911
Sondgeroth, K. S., McElwain, T. F., Allen, A. J., Chen, A. V., and Lau, A. O. (2013). Loss of neurovirulence is associated with reduction of cerebral capillary sequestration during acute Babesia bovis infection. Parasites Vect. 6:181. doi: 10.1186/1756-3305-6-181
Soulard, V., Amadoudji Zin, M., Fitting, C., Ibitokou, S., Oesterholt, M., Luty, A. J. F., et al. (2011). Placental Malaria-Associated Suppression of Parasite-Specific Immune Response in Neonates Has No Major Impact on Systemic CD4 T Cell Homeostasis ∇. Infect. Immun. 79, 2801–2809. doi: 10.1128/IAI.00203-11
Stelzer, S., Basso, W., Benavides Silván, J., Ortega-Mora, L. M., Maksimov, P., Gethmann, J., et al. (2019). Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 15:e00037. doi: 10.1016/j.fawpar.2019.e00037
Sugiyama, T., Cuevas, L. E., Bailey, W., Makunde, R., Kawamura, K., Kobayashi, M., et al. (2001). Expression of Intercellular Adhesion Molecule 1 (ICAM-1) in Plasmodium falciparum -infected Placenta. Placenta 22, 573–579. doi: 10.1053/plac.2001.0692
Suguitan, A. L., Leke, R. G. F., Fouda, G., Zhou, A., Thuita, L., Metenou, S., et al. (2003). Changes in the Levels of Chemokines and Cytokines in the Placentas of Women with Plasmodium falciparum Malaria. J. Infect. Dis. 188, 1074–1082. doi: 10.1086/378500
Sun, Y., Li, S.-G., Jiang, J.-F., Wang, X., Zhang, Y., Wang, H., et al. (2014). Babesia venatorum Infection in Child. China. Emerg. Infect. Dis. 20, 896–897. doi: 10.3201/eid2005.121034
Surra, N. D., and Jesus, J. E. (2015). The anemic and thrombocytopenic febrile neonate. J. Emerg. Med. 48, 675–678. doi: 10.1016/j.jemermed.2014.11.026
Taubert, A., Krüll, M., Zahner, H., and Hermosilla, C. (2006). Toxoplasma gondii and Neospora caninum infections of bovine endothelial cells induce endothelial adhesion molecule gene transcription and subsequent PMN adhesion. Vet. Immunol. Immunopathol. 112, 272–283. doi: 10.1016/j.vetimm.2006.03.017
Tegegne, D., Kelifa, A., Abdurahaman, M., and Yohannes, M. (2016). Seroepidemiology and associated risk factors of Toxoplasma gondii in sheep and goats in Southwestern Ethiopia. BMC Vet. Res. 12:280. doi: 10.1186/s12917-016-0906-2
Tegegne, Y., Asmelash, D., Ambachew, S., Eshetie, S., Addisu, A., and Jejaw Zeleke, A. (2019). The Prevalence of Malaria among Pregnant Women in Ethiopia: A Systematic Review and Meta-Analysis. J. Parasitol. Res. 2019:8396091. doi: 10.1155/2019/8396091
Terkawi, M. A., Cao, S., Herbas, M. S., Nishimura, M., Li, Y., Moumouni, P. F. A., et al. (2015). Macrophages Are the Determinant of Resistance to and Outcome of Nonlethal Babesia microti Infection in Mice. Infect. Immun. 83, 8–16. doi: 10.1128/IAI.02128-14
Tołkacz, K., Bednarska, M., Alsarraf, M., Dwużnik, D., Grzybek, M., Welc-Falȩciak, R., et al. (2017). Prevalence, genetic identity and vertical transmission of Babesia microti in three naturally infected species of vole, Microtus spp. (Cricetidae). Parasites Vect. 10:66. doi: 10.1186/s13071-017-2007-x
Tomavo, S. (1996). “The Major Surface Proteins of Toxoplasma gondii: Structures and Functions,” in Toxoplasma gondii Current Topics in Microbiology and Immunology, ed. U. Gross (Berlin: Springer), 45–54. doi: 10.1007/978-3-642-51014-4_4
Torgerson, P. R., and Mastroiacovo, P. (2013). The global burden of congenital toxoplasmosis: a systematic review. Bull. World Health Organ. 91, 501–508. doi: 10.2471/BLT.12.111732
Tranas, J., Heinzen, R. A., Weiss, L. M., and McAllister, M. M. (1999). Serological Evidence of Human Infection with the Protozoan Neospora caninum. Clin. Diagn. Lab. Immunol. 6, 765–767.
Tufts, D. M., and Diuk-Wasser, M. A. (2018). Transplacental transmission of tick-borne Babesia microti in its natural host Peromyscus leucopus. Parasit. Vectors 11:286. doi: 10.1186/s13071-018-2875-8
Tuikue Ndam, N. G., Salanti, A., Bertin, G., Dahlbäck, M., Fievet, N., Turner, L., et al. (2005). High Level of var2csa Transcription by Plasmodium falciparum Isolated from the Placenta. J. Infect. Dis. 192, 331–335. doi: 10.1086/430933
Turkey, S. A., Abbas, A. A.-H., Hathal, H. D., and Abdulrasul, E. A. (2019). The Role of TLR4 Gene Polymorphism and Haplotypes in the Susceptibility to Toxoplasmosis in Iraqi Aborted Women. J. Pure Appl. Microbiol. 13, 1151–1157.
Umbers, A. J., Boeuf, P., Clapham, C., Stanisic, D. I., Baiwog, F., Mueller, I., et al. (2011). Placental Malaria-Associated Inflammation Disturbs the Insulin-like Growth Factor Axis of Fetal Growth Regulation. J. Infect. Dis. 203, 561–569. doi: 10.1093/infdis/jiq080
Valdès, V., Legagneur, H., Watrin, V., Paris, L., and Hascoët, J. M. (2011). Congenital toxoplasmosis due to maternal reinfection during pregnancy. Arch. Pediatr. 18, 761–763.
van Duivenvoorde, L. M., Voorberg-van, der Wel, A., van der Werff, N. M., Braskamp, G., Remarque, E. J., et al. (2010). Suppression of Plasmodium cynomolgi in rhesus macaques by coinfection with Babesia microti. Infect. Immun. 78, 1032–1039. doi: 10.1128/IAI.00921-09
Vannier, E., and Krause, P. J. (2012). Human babesiosis. N. Engl. J. Med. 366, 2397–2407. doi: 10.1056/NEJMra1202018
Varki, A., and Gagneux, P. (2012). Multifarious roles of sialic acids in immunity. Ann. N Y. Acad. Sci. 1253, 16–36. doi: 10.1111/j.1749-6632.2012.06517.x
Vásquez, A. M., Segura, C., and Blair, S. (2013). Induction of pro-inflammatory response of the placental trophoblast by Plasmodium falciparum infected erythrocytes and TNF. Malar. J. 12:421. doi: 10.1186/1475-2875-12-421
Vogel, N., Kirisits, M., Michael, E., Bach, H., Hostetter, M., Boyer, K., et al. (1996). Congenital toxoplasmosis transmitted from an immunologically competent mother infected before conception. Clin. Infect. Dis. 23, 1055–1060.
Vonlaufen, N., Guetg, N., Naguleswaran, A., Müller, N., Björkman, C., Schares, G., et al. (2004). In Vitro Induction of Neospora caninum Bradyzoites in Vero Cells Reveals Differential Antigen Expression, Localization, and Host-Cell Recognition of Tachyzoites and Bradyzoites. Infect. Immun. 72, 576–583. doi: 10.1128/IAI.72.1.576-583.2004
Vottier, G., Arsac, M., Farnoux, C., Mariani-Kurkdjian, P., Baud, O., and Aujard, Y. (2008). Congenital malaria in neonates: two case reports and review of the literature. Acta Paediatrica 97, 505–508. doi: 10.1111/j.1651-2227.2008.00690.x
Votýpka, J., Modrý, D., Oborník, M., Šlapeta, J., and Lukeš, J. (2017). “Apicomplexa,” in Handbook of the Protists, eds J. M. Archibald, A. G. B. Simpson, C. H. Slamovits, L. Margulis, M. Melkonian, D. J. Chapman, et al. (Cham: Springer International Publishing), 1–58. doi: 10.1007/978-3-319-32669-6_20-1
Wahlgren, M., Goel, S., and Akhouri, R. R. (2017). Variant surface antigens of Plasmodium falciparum and their roles in severe malaria. Nat. Rev. Microbiol. 15, 479–491. doi: 10.1038/nrmicro.2017.47
Wang, J., Tang, D., Li, W., Xu, J., Liu, Q., and Liu, J. (2017). A new microneme protein of Neospora caninum, NcMIC8 is involved in host cell invasion. Exp. Parasitol. 175, 21–27. doi: 10.1016/j.exppara.2017.01.004
Wang, M. F., and Lai, S. C. (2013). Fibronectin degradation by MMP-2/MMP-9 in the serum of pregnant women and umbilical cord with Toxoplasma gondii infection. J. Obstet. Gynaecol. 33, 370–374. doi: 10.3109/01443615.2013.769501
Wang, X., Xiao, Y.-P., Bouchut, A., Al-Khedery, B., Wang, H., and Allred, D. R. (2012). Characterization of the Unusual Bidirectional ves Promoters Driving VESA1 Expression and Associated with Antigenic Variation in Babesia bovis. Eukaryotic Cell 11, 260–269. doi: 10.1128/EC.05318-11
Wang, Z.-D., Liu, H.-H., Ma, Z.-X., Ma, H.-Y., Li, Z.-Y., Yang, Z.-B., et al. (2017). Toxoplasma gondii Infection in Immunocompromised Patients: A Systematic Review and Meta-Analysis. Front. Microbiol. 8:00389. doi: 10.3389/fmicb.2017.00389
WHO (2019). World malaria report 2019. Geneva: WHO. Available online at: https://www.who.int/publications-detail-redirect/9789241565721 (accessed August 18, 2020)
Wobeser, B. K., Godson, D. L., Rejmanek, D., and Dowling, P. (2009). Equine protozoal myeloencephalitis caused by Neospora hughesi in an adult horse in Saskatchewan. Can. Vet. J. 50, 851–853.
Wright, I. G., Goodger, B. V., McKenna, R. V., and Mahoney, D. F. (1979). AcuteBabesia bovis infection: A study of the vascular lesions in kidney and lung. Z. Parasitenkd. 60, 19–27. doi: 10.1007/BF00928968
Wujcicka, W., Wilczyński, J., and Nowakowska, D. (2013). SNPs in toll-like receptor (TLR) genes as new genetic alterations associated with congenital toxoplasmosis? Eur. J. Clin. Microbiol. Infect. Dis. 32, 503–511. doi: 10.1007/s10096-012-1763-y
Wujcicka, W., Wilczyński, J., and Nowakowska, D. (2014). Do the placental barrier, parasite genotype and Toll-like receptor polymorphisms contribute to the course of primary infection with various Toxoplasma gondii genotypes in pregnant women? Eur. J. Clin. Microbiol. Infect. Dis. 33, 703–709. doi: 10.1007/s10096-013-2017-3
Wujcicka, W., Wilczyński, J., and Nowakowska, D. (2017). Genetic alterations within TLR genes in development of Toxoplasma gondii infection among Polish pregnant women. Adv. Med. Sci. 62, 216–222. doi: 10.1016/j.advms.2017.02.002
Xiao, J., Garcia-Lloret, M., Winkler-Lowen, B., Miller, R., Simpson, K., and Guilbert, L. J. (1997). ICAM-1-mediated adhesion of peripheral blood monocytes to the maternal surface of placental syncytiotrophoblasts: implications for placental villitis. Am. J. Pathol. 150, 1845–1860.
Yager, P. H., Luginbuhl, L. M., and Dekker, J. P. (2014). Case records of the Massachusetts General Hospital. Case 6-2014. A 35-day-old boy with fever, vomiting, mottled skin, and severe anemia. N. Engl. J. Med. 370, 753–762. doi: 10.1056/NEJMcpc1208155
Yarovinsky, F. (2014). Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 14, 109–121. doi: 10.1038/nri3598
Yarovinsky, F., and Sher, A. (2006). Toll-like receptor recognition of Toxoplasma gondii. Int. J. Parasitol. 36, 255–259. doi: 10.1016/j.ijpara.2005.12.003
Yarovinsky, F., Zhang, D., Andersen, J. F., Bannenberg, G. L., Serhan, C. N., Hayden, M. S., et al. (2005). TLR11 activation of dendritic cells by a protozoan profilin-like protein. Science 308, 1626–1629.
Yavuz, E., Aydin, F., Seyhan, A., Topuz, S., Karagenc, Y., Tuzlali, S., et al. (2006). Granulomatous villitis formed by inflammatory cells with maternal origin: a rare manifestation type of placental toxoplasmosis. Placenta 27, 780–782. doi: 10.1016/j.placenta.2005.07.002
Yi, W., Bao, W., Rodriguez, M., Liu, Y., Singh, M., Ramlall, V., et al. (2018). Robust adaptive immune response against Babesia microti infection marked by low parasitemia in a murine model of sickle cell disease. Blood Adv. 2, 3462–3478. doi: 10.1182/bloodadvances.2018026468
Young, K. M., Corrin, T., Wilhelm, B., Uhland, C., Greig, J., Mascarenhas, M., et al. (2019). Zoonotic Babesia: A scoping review of the global evidence. PLoS One 14:e0226781. doi: 10.1371/journal.pone.0226781
Zeldovich, V. B., Clausen, C. H., Bradford, E., Fletcher, D. A., Maltepe, E., Robbins, J. R., et al. (2013). Placental syncytium forms a biophysical barrier against pathogen invasion. PLoS Pathog. 9:e1003821. doi: 10.1371/journal.ppat.1003821
Zhang, H., Hu, X., Liu, X., Zhang, R., Fu, Q., and Xu, X. (2012). The Treg/Th17 imbalance in Toxoplasma gondii-infected pregnant mice. Am. J. Reprod. Immunol. 67, 112–121. doi: 10.1111/j.1600-0897.2011.01065.x
Zhao, L., Jiang, R., Jia, N., Ning, N., Zheng, Y., Huo, Q., et al. (2020). Human Case Infected With Babesia venatorum: A 5-Year Follow-Up Study. Open Forum Infect. Dis. 7:ofaa062. doi: 10.1093/ofid/ofaa062
Keywords: Apicomplexa, congenital transmission, placenta, infection-immunology, host–parasite interactions
Citation: Rojas-Pirela M, Medina L, Rojas MV, Liempi AI, Castillo C, Pérez-Pérez E, Guerrero-Muñoz J, Araneda S and Kemmerling U (2021) Congenital Transmission of Apicomplexan Parasites: A Review. Front. Microbiol. 12:751648. doi: 10.3389/fmicb.2021.751648
Received: 01 August 2021; Accepted: 01 September 2021;
Published: 29 September 2021.
Edited by:
George Grant, University of Aberdeen, United KingdomReviewed by:
Antonio Ortega-Pacheco, Universidad Autónoma de Yucatán, MexicoHem Chandra Jha, Indian Institute of Technology Indore, India
Copyright © 2021 Rojas-Pirela, Medina, Rojas, Liempi, Castillo, Pérez-Pérez, Guerrero-Muñoz, Araneda and Kemmerling. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ulrike Kemmerling, ukemmerling@uchile.cl