 Qian Li
Qian Li Qi Li
Qi Li Rancheng Xu1,4
Rancheng Xu1,4 Yuanqing Wang
Yuanqing Wang- 1Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China
- 2CAS Center for Excellence in Life and Paleoenviroment, Beijing, China
- 3Centre for Vertebrate Evolutionary Biology, Yunnan University, Kunming, China
- 4College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing, China
Rodents are an important component in Paleogene terrestrial ecosystems. Their evolution and faunal turnover have likely been influenced and triggered by global climate change. Here, we compiled rodent faunas from the early Eocene to the early Oligocene in Asia to discuss rodent faunal turnover and its correlation with paleogeographic and paleoclimate changes. Successive rodent faunas from the early Eocene to the early Oligocene are recorded in East Asia, and rodent faunal turnover is obviously affected by paleoclimatic changes. During the Ulangochuian (after the middle Eocene climatic optimum), when temperatures declined slowly, the East Asian rodent fauna showed a clear decline in generic diversity and a transformation from ctenodactyloid-dominant to cricetid–dipodid-dominant faunas. During the Eocene–Oligocene transition (EOT) and global cooling, the East Asian rodent fauna exhibited a considerably high generic diversity of cricetids, dipodids, and ctenodactylids. The low temperatures during the Ulangochuian and Ergilian in East Asia served as a habituation ground for the cricetids and dipodids, which became preadapted for the EOT, successfully helping rodents become dominant faunas after EOT. The rodent faunas in South Asia formed a relatively unique group in much of the Eocene and early Oligocene; sometimes, they were closer to European or African than to Asian rodents. The greatly different paleoenvironment probably caused striking differences between the rodent faunas in South Asia and East Asia.
Introduction
Today, rodents are the most diverse and abundant terrestrial mammals. Rodents constitute over half the species of living mammals, and their fossil forms are also extremely diverse. Rodents are found worldwide, except for Antarctica and some oceanic islands, and they exhibit a number of different lifestyles and locomotor behaviors. While generalized rodent adaptation is terrestrial, various forms are more specialized for climbing, gliding, swimming, or digging. Because of their large populations and short gestation periods, rodents have more advantages than other mammals in fauna in terms of biostratigraphic comparison and the relationship between mammal evolution and climate change.
The rodent fossil record has expanded at a significant pace, and fossils are increasingly discovered in regions that have been previously less known for fossils. In Asia, the complex rodent evolutionary history began during the early Eocene, from which the oldest fossils have been found (Li et al., 1989; Meng and Li, 2010; Li and Meng, 2015). The Paleogene is a key period for the origin and evolution of rodents, and the paleoclimate had multiple changes during the Paleogene (Westerhold et al., 2020), such as the global cooling event during the Eocene–Oligocene transitional period (Meng and McKenna, 1998; Kraatz and Geisler, 2010; Zhang et al., 2012; Sun et al., 2014); thus, we focus on rodent fossil records in Asia from the early Eocene to the early Oligocene. Furthermore, by taking advantage of an updated Cenozoic timescale in Asia (Romer, 1966; Luterbacher et al., 2004; Vandenberghe et al., 2012; Wang et al., 2019; Speijer et al., 2020), we use all available rodent faunal data and analyze rodent composition and diversity at the genus level, noting the different rodent assemblages and faunal turnovers. We discuss the possible relationships between the rodent evolution and faunal turnover and paleoclimatic changes. In addition, because different rodent assemblages also appeared in different areas of Asia during the same period, we consider the possible relationships between faunal dispersal and paleogeographic transformation.
Methods
Biochronology
Following the principle of North American land mammal ages, Romer (1966) proposed some related Asian Paleogene land mammal ages on the basis of mammalian faunas from different stratigraphic levels, which have formed the foundation for further Asian Paleogene chronostratigraphic study (Wang et al., 2019). These land mammal ages have often been used in discussing intercontinental biostratigraphic correlation and mammalian dispersals. Now, the Bumbanian, Arshantan, Irdinmanhan, Sharamurunian, Ulangochuian, and Ergilian ages in the Eocene, and the Hsandagolian age in the Oligocene have been adopted as the Asian Paleogene land mammal ages in the geologic time scales (Luterbacher et al., 2004; Vandenberghe et al., 2012; Speijer et al., 2020).
For many years, the Ulangochuian land mammal age that is currently used in the Chinese Paleogene land mammal age sequence was equivalent to both the Ulangochuian and Ergilian Asian land mammal ages (Tong et al., 1995; Vandenberghe et al., 2012). A recent investigation in the Erlian Basin, Nei Mongol, revealed that deposits between those with typical Ulangochuian and early Oligocene mammal faunas contain fossil mammals that are correlated to the Mongolian Ergilian fauna; thus, the Baiyinian land mammal age was proposed as the equivalent of the Ergilian in the Chinese Paleogene land mammal age sequence (Wang et al., 2019).
Here, these Asian Paleogene land mammal ages are used to indicate the evolution and turnover of rodent faunas in more detail. The ages of fossil localities are inferred based on the latest biostratigraphic, magnetostratigraphic, and radiometric dating.
Distribution of localities
Known rodent fossils are widely distributed in different areas of Asia, and the different mammalian fossil compositions may have been caused by paleogeographic patterns or may indicate different habitat conditions. The present study considers 35 localities that are biostratigraphically dated from the early Eocene to the early Oligocene (Appendix Tables 1–3) and assembled into three regions of Asia: East Asia (China and Mongolia), Central Asia (Kyrgyzstan and Kazakhstan), and South Asia (India, Pakistan, Thailand, and Myanmar) (Figure 1).
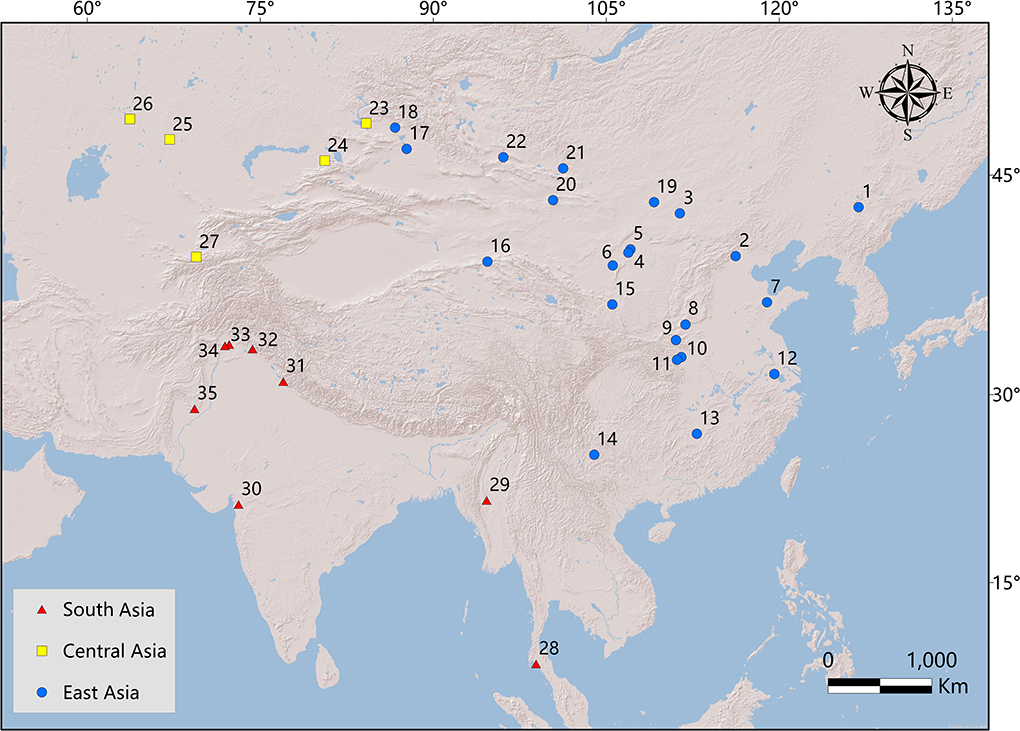
Figure 1. Principal Paleogene rodent localities of Asia are drawn in ArcMap10.3. Blue circles represent localities of East Asia, yellow squares represent localities of Central Asia, and red triangles represent localities of South Asia. Localities 1-17 are from China: 1. Huadian; 2. Changxindian; 3. Erlian Basin; 4. Qianlishan; 5. Saint Jacques; 6. Ulantatal; 7. Wutu; 8. Yuanqu; 9. Lushi; 10. Liguanqiao; 11. Danjiangkou; 12. Shanghuang; 13. Lingcha; 14. Caijiachong; 15. Haiyuan; 16. Danghe; 17 Ulungur River area; and 18. Burqin. Localities 19-22 are from Mongolia: 19. eastern Gobi Desert; 20. Nemegt Basin; 21. Valley of Lakes; and 22. Shine Us. Localities 23-26 are from Kazakhstan: 23. Zaysan; 24. Shinzhaly; 25. Kyzyl-Kak; and 26. Chelkar-Teniz; 27. Andarak 2 (Kyrgyzstan); 28. Krabi (Thailand); and 29. Pondaung (Myanmar). Localities 30–32 are from India: 30. Vastan; 31. Subathu; and 32. Kalakot. Localities 33–35 are from Pakistan: 33. Ganda Kas; 34. Chorlakki; and 35. Bugti Hills.
Taxonomy
Known rodent fossils were classified into different rodent families, and the taxonomic system of the family that was adopted in our analysis is as follows: Alagomyidae, Orogomyidae (Dashzeveg, 1990b), Aplodontidae (Wood, 1980), Anomaluroidea (Marivaux et al., 2005), Archetypomyidae (Meng et al., 2007), Castoridae, Sciuridae (McKenne and Bell, 1997), Ctenodactylidae (Wang, 1997a, 2001b), Cricetidae (Wang et al., 2020), Cylindrodontidae (Wood, 1980; Wang, 2019), Diatomyidae (Mein and Ginsburg, 1997), Dipodidae (Qiu and Wang, 2019), Eomyidae (Fejfar et al., 1998), Ischromyidae (Anderson, 2008), Tsaganomyidae (Wang, 2001a), and Zelomyidae (Dawson et al., 2003a).
Regarding ctenodactyloids, recent phylogenetic analysis indicated that some families or subfamilies, such as Cocomyinae, Advenimurinae (Dashzeveg, 1990a), Tamquammyidae (Averianov, 1996; Tong, 1997), Yuomyidae (Dawson et al., 1984; Tong, 1997), and Chapattimyidae (Averianov, 1996), are probably paraphyletic (Li and Meng, 2015). Because of the poor preservation of some early ctenodactyloid rodents and the complicated history of taxonomic studies on the group, it is extremely difficult to conduct a thorough phylogenetic analysis of ctenodactyloids. Some researchers have suggested the use of Ctenodactyloidea without reference to a specific family for the Eocene genera Dashzeveg and Meng, 1998; Li and Meng, 2015). Here, we also followed this opinion; most Eocene genera of ctenodactyloids included Ctenodactyloidea. Ctenodactylidae as a monophyletic is a generally accepted view; here, late Eocene Karakoromys and Oligocene ctenodactylids were included in Ctenodactylidae (Wang, 1997a, 2001b).
Indeterminate taxon identifications and taxonomic modifications, such as “cf.” or “?”, or some cases of “sp.”, were included in our analyses as a record.
Diversity and sampling biases
The simplest method to count the number of taxa to measure biodiversity may produce biases that distort the diversity pattern due to the different preservation and sampling intensities of various fossils. The rarefaction method is widely used in paleontology for removing biases (Gotelli and Colwell, 2001; Shen et al., 2004; Huang, 2012); here, paleobiological biodiversity curves were corrected using rarefaction by PAST software (Hammer et al., 2001).
Results
Bumbanian rodent assemblage
Early Eocene Bumbanian rodent fossil assemblages are present in China, Mongolia, and India. In China, the Bumbanian rodent fossil localities are present in multiple regions, including Hunan, Hubei, Nei Mongol, Shandong, and Henan (Li, 1963; Dawson et al., 1984; Li et al., 1989, 2018; Hu, 1995; Tong, 1997; Guo et al., 2000; Wible et al., 2005; Tong and Wang, 2006; Meng and Li, 2010; Li and Meng, 2015). In addition, abundant rodent fossils are found in the Bumban Member of the Naran-Bulak Formation of the Tsagan Khushu locality in the Nemegt Basin of Mongolia (Shevyreva, 1989; Dashzeveg, 1990a,b), and an ischyromyid rodent, Meldimys, is recorded in the lower Eocene Vastan lignite mine of western India (Rana et al., 2008); this occurrence is the oldest record of a rodent from India.
The Bumbanian rodent assemblages from China and Mongolia are similar in the dominance of original ctenodactyloids, and original rodents such as Alagomys are also present (Figure 2) (Appendix Table 1). Ctenodactyloid rodents in Bumbanian, as the oldest known rodents with modern features in Asia, already have a high diversity and are represented by different ctenodactyloid genera known primarily from China and Mongolia.
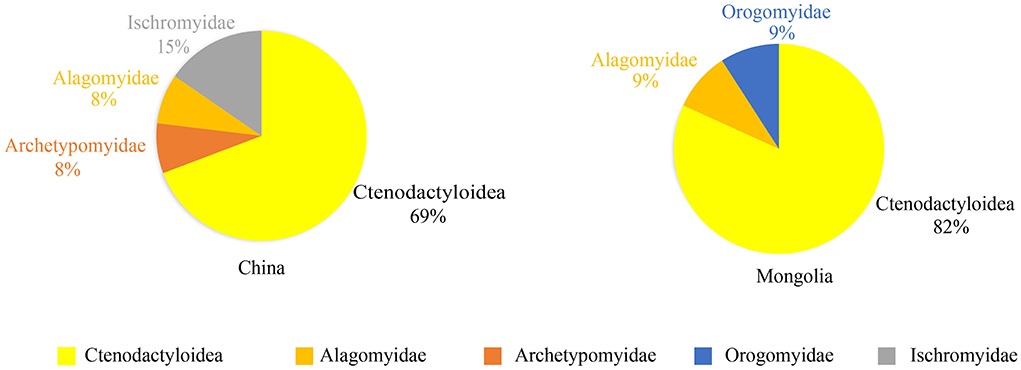
Figure 2. Diagrams of the Bumbanian rodent composition from China and Mongolia in East Asia (in percentage of genera).
Except for ctenodactyloids, other basal rodent families are present in China and Mongolia. Alagomys is from the Erlian Basin of Nei Mongol of China and the Nemegt Basin of Mongolia. However, the validity of the familial association of Alagomys is still unknown. Meng and Wyss (2001) excluded Alagomys from the Rodentia, but others pointed out that Alagomys is close to true rodents and is generally considered the most primitive rodent (Dawson, 2015). Based on the similarities of Alagomys and the original ctenodactyloids and cylindrodontids, we also adopted Alagomys in Rodentia and Alagomyidae (Dashzeveg, 1990b). Archetypomys is a primitive rodent and occupies an intermediate phylogenetic position between alagomyids and rodents of modern features. Primitive ischyromyids, such as Actioparamys and Taishanomys, are also present in the Bumbanian rodent assemblage from China. Orogomys from the Nemegt Basin of Mongolia was classified as Orogomyidae (Dashzeveg, 1990b).
Arshantan rodent assemblage
The Arshantan rodent assemblages are known in East, Central, and South Asia. In East Asia, Arshantan rodents appeared in the Erlian Basin of Nei Mongol and in the Junggar Basin of Xinjiang, China (Meng et al., 2001; Li and Meng, 2010, 2015; Li et al., 2018; Li, 2019). The ~2,900 Arshantan rodent specimens collected from the Erlian Basin represent at least 323 individuals, as counted by the minimum number of individuals (MNI), and rodents from the Erlian Basin have an absolute advantage in both number and species diversity. In Central Asia, abundant rodents are known at the Andarak 2 locality in Kyrgyzstan (Averianov, 1996) and the Zaysan Depression in eastern Kazakhstan (Shevyreva, 1984, 1996). The South Asian Arshantan rodents were described in the lower middle part of the Subathu Formation of the type area in Himachal Pradesh, northwestern sub-Himalaya, India (Gupta and Kumar, 2015).
In the Arshantan rodents, whether in East Asia, Central Asia, or South Asia, ctenodactyloids are a dominant element (Figure 3). In China, ctenodactyloids contain the primitive ctenodactyloid Tamquammys, Chenomys, Simplicimys, and Advenimus. Among them, Tamquammys, Advenimus, and Chenomys were present in the Bumbanian and in the Arshantan assemblage.
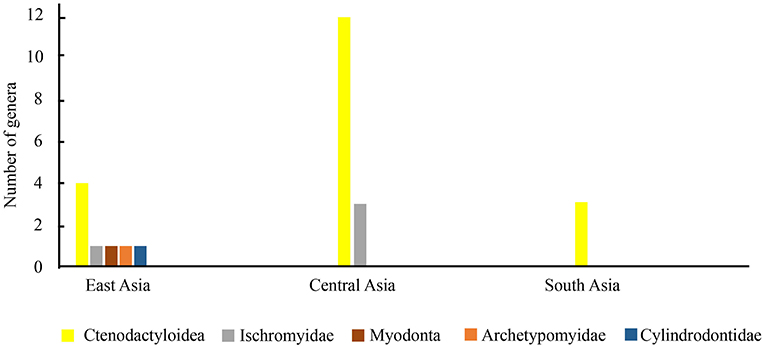
Figure 3. Diagram of the Arshantan rodent compositions in East, Central, and South Asia.
In Central Asia, ctenodactyloid rodents have high generic richness and are clearly different from East and South Asian ctenodactyloids in genera (Appendix Tables 1–3). Only Advenimus is present in East and Central Asia, which may suggest that some faunal exchanges occurred between East and Central Asia during the Arshantan. Only ctenodactyloids Subathumys, Birbalomys, and Chapattimys are present in South Asia. Except for various ctenodactyloids, in East Asia, the primitive myodont Erlianomys (Li and Meng, 2010), the basal cylindrodontid Gobiocylindrodon (Li et al., 2019), the primitive rodent Archetypomys, and some ischyromyid fossils (Li, 2016) from the Arshantan rodent assemblage are present (Figure 3). In Central Asia, the ischyromyids Mneiomys, Paramys, and Abrosomys were found in the Zaysan depression in Kazakhstan (Shevyreva, 1984, 1996).
The ctenodactyloids in Central Asia were classified as Chapattimyidae or Tamquammyidae by Averianov (1996) and Shevyreva (1984, 1989), and South Asian ctenodactyloids were included in Chapattimyidae (Gupta and Kumar, 2015). The phylogenetic analysis of these families indicated that they are not monophyletic (Li and Meng, 2015). Resolving phylogenetic problems is difficult and beyond the scope of this study. However, the P4/p4 morphology, either molariform or nonmolariform, has been frequently emphasized as an important characteristic in ctenodactyloid rodents. The ctenodactyloids classified as yuomyids and chapattimyids are distinguished from other ctenodactyloids by their molariform P4/p4 morphology. Recently, the characteristics of the molariform P4/p4 morphology have also been further clarified by Li and Meng (2015). According to the P4/p4 morphology, in the Arshantan, most East Asian ctenodactyloids present nonmolariform premolars, and the ctenodactyloids with molariform premolars are more commonly found in Central and South Asia. Gupta and Kumar (2015) described Subathymys from the Subathu Formation of India and pointed out that chapattimyids originated in the sub-Himalayan region during the Ypresian. According to the aforementioned description, ctenodactyloids with molariform premolars quite possibly originated from South Asia; they moderately diversified in Arshantan and spread rather quickly into Central Asia.
In summary, the Arshantan rodent assemblages in East, Central, and South Asia are distinct, and they rarely have the same genera.
Irdinmanhan rodent assemblage
The middle Eocene Irdinmanhan rodent assemblage is found in China in East Asia, eastern Kazakhstan in Central Asia, and Pakistan and India in South Asia. In China, the Irdinmanhan rodent fossil localities are known in Nei Mongol, Henan, Jiangsu, and Shanxi (Li, 1963, 2012; Tong, 1997; Dawson et al., 2003a, 2010; Li and Meng, 2013, 2015; Li et al., 2016, 2018; Fostowicz-Frelik et al., 2021). Irdinmanhan rodents are found in the Kolpak Formation of the Shinzhaly River in eastern Kazakhstan (Emry et al., 1998b). In South Asia, Irdinmanhan rodents have been recorded in the upper part of the Subathu Formation in India (Sahni and Khare, 1973; Sahni and Srivastava, 1976; Kumar et al., 1997a,b; Gupta and Kumar, 2015), the Kuldana Formation of the Ganda Kas area, and the Mami Khel Formation of the Chorlakki area in Pakistan (Hussain et al., 1978; Hartenberger, 1982; Dawson et al., 1984; Thewissen et al., 2001).
In East Asia, the Irdinmanhan rodent assemblage has more derived members than the Bumbanian and Arshantan rodent assemblages, such as cricetids, dipodids, zelomyids, and ? diantomyids (Appendix Table 1). The rodent faunas in some localities of the Erlian Basin in Nei Mongol and the Shipigou locality in Henan have been studied in detail based on the MNI (Tong, 1997; Li, 2016). The generic rodent number in Henan is the highest in China. Ctenodactyloids are major components in Henan and Nei Mongol, and cricetids, ischyromyids, and cylindrodontids are moderately diversified in the Irdinmanhan rodent assemblage (Figure 4). The few fossils of Zelomyidae and ? Diatomyidae appeared in the fissure D in the Shanghuang fissure in Liyang, Jigansu, and Huoshipou of the Yuanqu Basin, Shanxi (Dawson et al., 2003a, 2010).
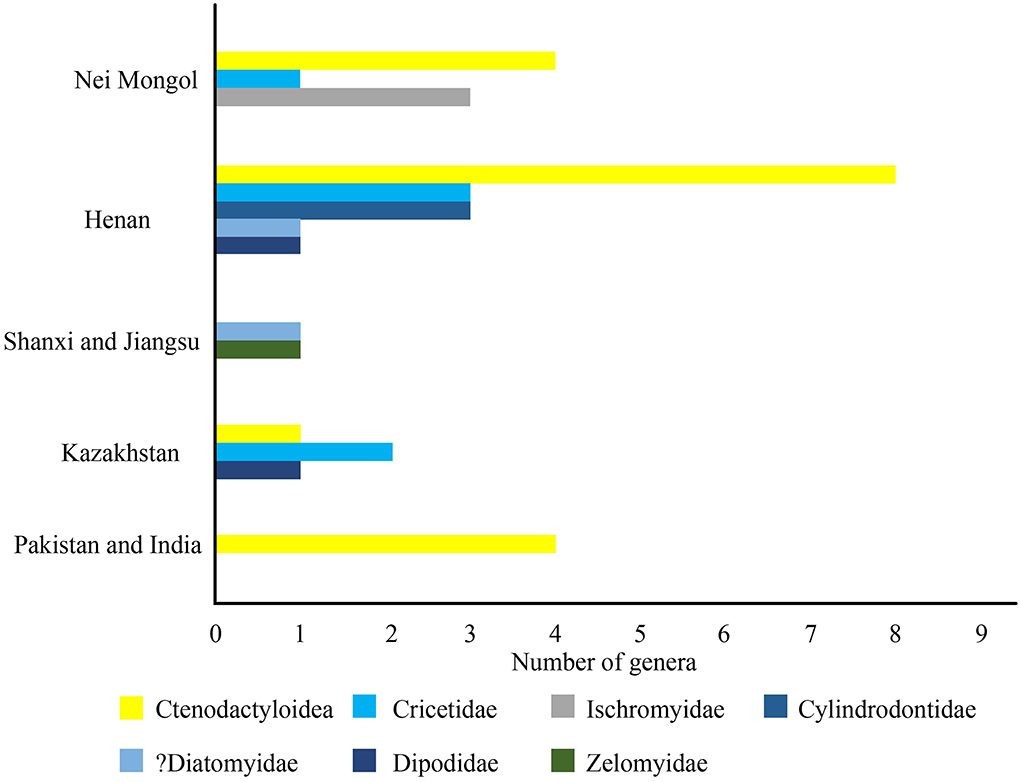
Figure 4. Diagram of the Irdinmanhan rodent compositions in East, Central, and South Asia.
The Irdinmanhan rodents in Kazakhstan are similar to those in China, and they also contain ctenodactyloids, dipodids, and cricetids. Ctenodactyloid Saykanomys and cricetid Pappocricetodon are present in China and Kazakhstan. In South Asia, the Irdinmanhan rodent assemblage is distinctly different from the rodent assemblage in East and Central Asia and is characterized by only ctenodactyloids that were classified as Chapattimyidae (Gupta and Kumar, 2015). The Irdinmanhan rodents in India and Pakistan are very similar; the dominant elements in these areas are Birbalomys and Chapattimys, which first appeared in the Arshantan rodent assemblage in India (Gupta and Kumar, 2015), extended to the Irdinmanhan assemblage and showed high species diversity.
Sharamurunian rodent assemblage
The middle Eocene Sharamurunian rodent fossil localities are in Nei Mongol, Shanxi, Jilin, Jiangsu, and Henan in China (Dawson, 1968; Wang and Dawson, 1994; Tong, 1997; Wang et al., 1998; Meng et al., 1999; Dawson et al., 2003a; Li et al., 2016; Li, 2018, 2019). The Sharamurunian of the Asian Paleogene land mammal age proposed by Romer (1966) was derived from the Paleogene mammal fauna found in the Ula Usu locality of the Erlian Basin, Inner Mongolia. Studies on Sharamurunian rodent fossils in the classic locality are very limited (Li, 1975). Recently, abundant rodent fossils were found in the Ula Usu locality, and a detailed study of these fossils has been carried out.
In known localities, many rodent fossils of the Shanghe-Zhaili fauna in the Yuanqu Basin of Shanxi and the Erlian Basin of Nei Mongol are present, showing a variety of genera and species. The Sharamurunian rodent assemblages in the Yuanqu and Erlian Basins show similarities in family composition; they all contain ctenodactyloids, cricetids, dipodids, and ischyromyids, and many genera are present in both localities (Table 1). However, zelomyids and cylindrodontids are present in the Yuanqu and Erlian basins, respectively. The ctenodactyloids in the Yuanqu Basin show higher generic richness than those in the Erlian Basin, and Gobiomys is the dominant ctenodactyloid genus in the Erlian Basin. Detailed analysis of the rodent fauna of the Sharamurunian in the Yuanqu and Erlian basins has been carried out based on the MNI. The results showed that cricetids and dipodids are dominant in the Yuanqu Basin, but in the Erlian Basin, ctenodactyloids and dipodids are dominant and secondary, respectively (Tong, 1997).
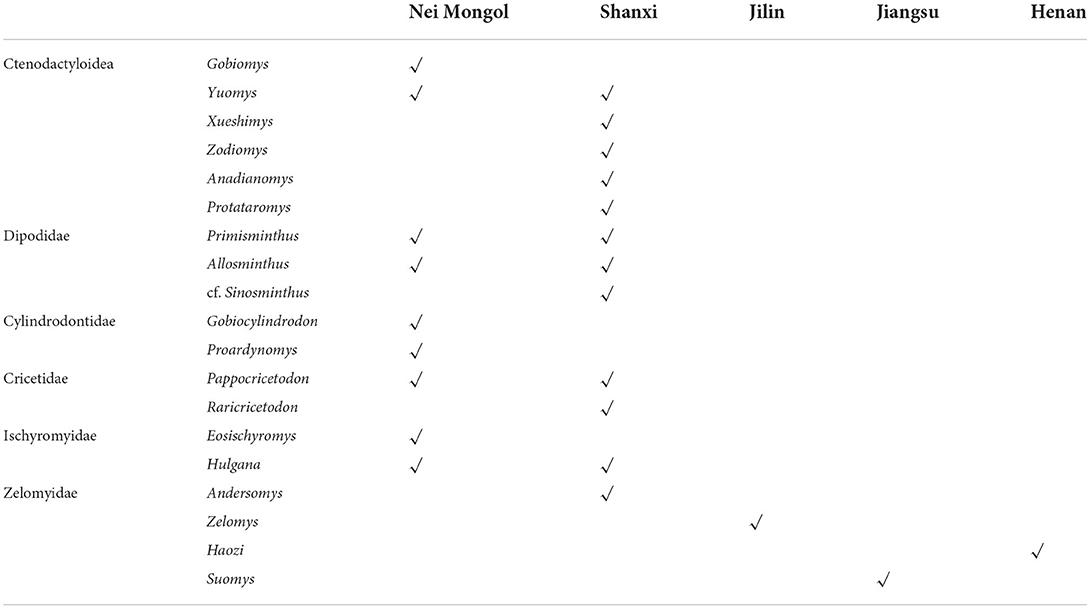
Table 1. Sharamurunian rodent assemblages in China.
In other localities, such as Huadian in Jilin, Shanghuang fissure fillings in Jiangsu, and Lushi in Henan, only zelomyids of the Sharamurunian rodent fauna have been discovered in these areas (Dawson et al., 2003a).
In addition to China, a few Sharamurunian rodent materials have been found in South Asia. The Pondaung Formation of central Myanmar has yielded one of the most diversified Eocene land mammal faunas in South Asia. Geochronological evidence has definitely established a late middle Eocene age for the mammal-bearing strata of the Pondaung Formation, and both fission track analyses of zircon grains and paleomagnetic studies of strata from the Pondaung area suggest an age of approximately 40 Ma (Tsubamoto et al., 2002; Khin et al., 2014; Jaeger et al., 2019). Therefore, the age of the Pondaung fauna corresponds to the Sharamurunian, based on biostratigraphy, nannoplankton assemblage, and magnetostratigraphic and radiometric dating. Some rodent specimens from the Pondaung Formation have been reported and included in the rodent superfamily Anomaluroidea, which is currently limited to tropical and subtropical forests of western and central Africa (Tsubamoto et al., 2000; Dawson et al., 2003b; Marivaux et al., 2005). The rodent Pondaungimys from the Pondaung Formation is similar to Nementchamys, an anomaluroid from the late middle Eocene strata of Algeria. The close relationship between Pondaungimys and Nementchamys emphasizes the widespread South Asian–North African distribution of anomaluroid rodents, thus suggesting communication between South Asia and Africa during the Sharamurunian (Marivaux et al., 2005).
Ulangochuian rodent assemblage
Of the reported Ulangochuian rodents in East Asia, localities restricted to the redefined Ulangochuian land mammal age mainly include Nei Mongol and Yunnan in China (Wang, 1985, 2001a; Wang and Meng, 1986; Li, 2018, 2019, 2021).
In Nei Mongol, Ulangochuian rodents were reported in the Lower White to Middle White beds of the Erden Obo section in the Erlian basin, Nei Mongol (Li, 2018, 2019, 2021). The classic Houldjin site in the Erlian Basin is another important locality. A recent investigation showed that the reported Houldjin fauna was represented by a mixed assemblage due to reworking (Wang, 2008; Wang et al., 2009); thus, here, we do not include these fossils. The rodent in the lower part of the Chijiachong Formation of Qujing, Yunnan, was considered as the Ulangochuian rodent (Wang, 1985, 2001a; Wang and Meng, 1986; Maridet and Ni, 2013).
The Ulangochuian rodent assemblage in Yunnan and Nei Mongol contains ctenodactyloid, cricetid, and dipodid rodents, and the cylindrodontid rodents are present only in Nei Mongol (Figure 5). In both localities, ctenodactyloids are moderately diversified and show different generic compositions, but Gobiomys in Nei Mongol and Youngomys in Yunnan are a sister group in the phylogenetic analysis (Wang, 2001c). However, some genera appear in both localities during the Ulangochuian, such as cricetid Eocricetodon, dipodid Allosminthus, Sinosminthus, and Heosminthus.
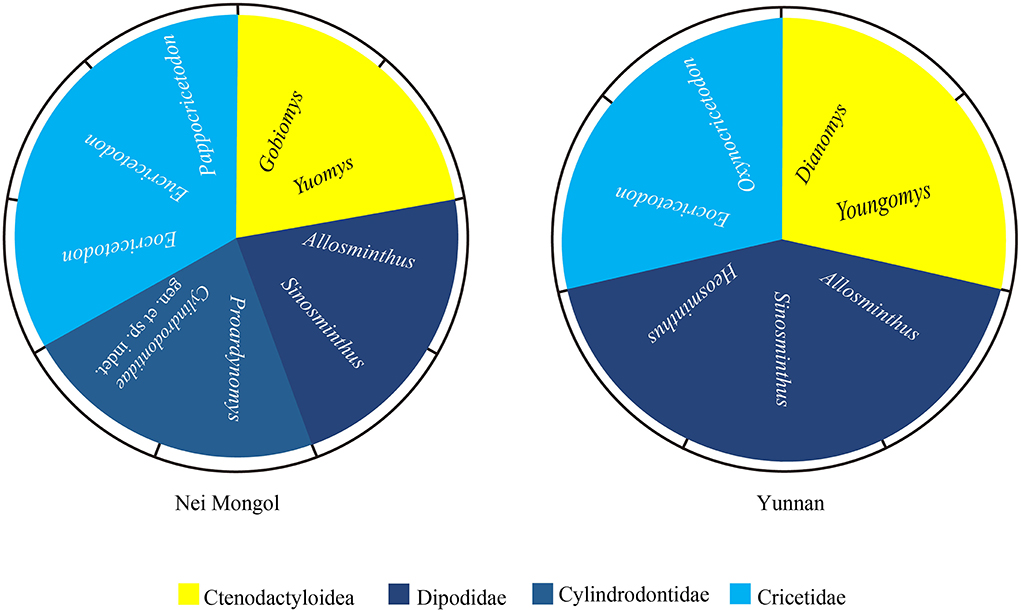
Figure 5. Diagram of the Ulangochuian rodent compositions in Nei Mongol and Yunnan in China.
Ergilian rodent assemblage
Late Eocene Ergilian rodent faunas are found in China and Mongolia in East Asia, Kazakhstan in Central Asia, and Thailand in South Asia.
Based on the biostratigraphic correlation, the age of the Upper Red beds of the Erden Obo section in the Erlian Basin, Nei Mongol, China, is the Ergilian land mammal age (Li, 2018). Ulantatal in Nei Mongol is another important site. Recently, Wasiljeff et al. (2020) derived a chronostratigraphic framework for the Ulantatal sequence, using paleomagnetic reversal stratigraphy and biostratigraphic correlation, and the time spanned by the strata was 35 to 27 Ma (from the latest Eocene to late Oligocene). The rodents in the lower part of the Kekeamu section in Ulantatal were representative of the latest Eocene (Wasiljeff and Zhang, 2022). In Mongolia, the Ergilian rodents were reported in the Ergilin Dzo Formation in the eastern Gobi Desert of Mongolia (Dashzeveg, 1993). The Zaysan Basin, eastern Kazakhstan, has a sedimentary section that includes strata spanning the Eocene–Oligocene boundary. The upper part of the Aksyir svita contains mammals, including abundant rodents that typically occur in the Ergilin Dzo fauna of Mongolia (Emry et al., 1998a).
The East and Central Asian Ergilian assemblage consists of ctenodactyloids, cricetids, dipodids, and cylindrodontids, and cricetids and dipodids are dominant. The rodent genera from Kazakhstan are all present in China and Mongolia in East Asia (Figure 6). In East Asia, the composition and dominant taxa of the Ergilian rodent assemblage are also similar to those of the Ulangochuian rodent faunas (Appendix Table 1). Ctenodactyloid Gobiomys, dipodid Allosminthus, Heosminthus, and Sinosminthus are present in the Ulangochuian and in the Ergilian assemblages.
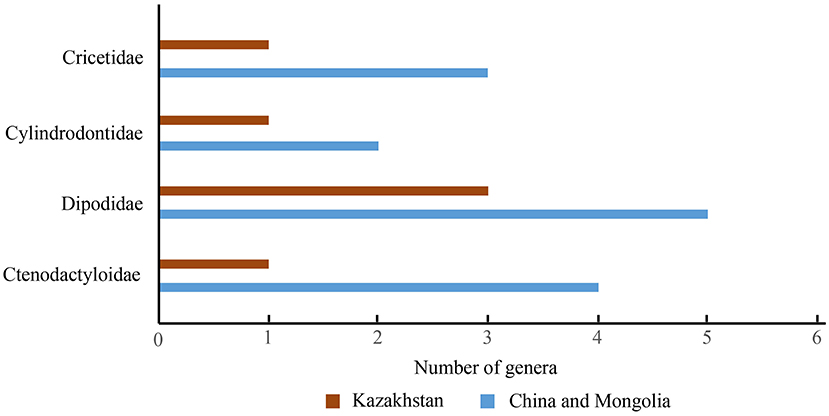
Figure 6. Diagram of the Ergilian rodent compositions in China and Mongolia in East Asia, and Kazakhstan in Central Asia.
In South Asia, Baluchimys krabiense has been found in the late Eocene strata in the Krabi Basin, Thailand. The striking affinities between B. krabiense and the late Eocene African P. algeriensis suggest faunal exchanges between South Asia and Africa during the Eocene (Marivaux et al., 2000).
Hsandagolian rodent assemblage
The Hsandagolian age corresponds to the Rupelian Stage and the lower part of the Chattian Stage (Speijer et al., 2020); thus, Hsandagolian includes the early Oligocene and part of the late Oligocene. Hsandagolian mammalian fossils are distributed in China, Mongolia, and Pakistan in Asia. In China, the Hsandagolian rodent mammals are found in the Ulantatal area, the “Upper White” beds of the Erlian Basin, Saint Jacques area and Qianlishan district in Nei Mongol (Wang, 1987; Wang and Emry, 1991; Wang and Qiu, 2003; Gomes Rodrigues et al., 2014; Li, 2018, 2021), the Burqin Basin in the northern area of Xinjiang (Wu et al., 2004; Ye et al., 2005), the Qingshuiying Formation in Haiyuan in Ningxia (Wang et al., 1994), the lower part of the Paoniuquan Formation in the Danghe Basin of Gansu (Wang and Qiu, 2004), and the upper part of the Caijiachong Formation in Qujing in Yunnan (Maridet and Ni, 2013; Li et al., 2017). The Oligocene deposits of the Ulantatal area in Nei Mongol contain rich rodent fossils, and a review of the rodent list was provided (Gomes Rodrigues et al., 2014). Based on earlier research in the area, the Ulantatal sequence was considered to be constrained to the Oligocene (Wang, 1997b; Gomes Rodrigues et al., 2014). Recently, Wasiljeff et al. (2020) redefined the stratigraphy and suggested that the Ulantatal Formation spans from the latest Eocene to the late Oligocene. Here, the Hsandagolian rodent assemblage in the Ulantatal area has been revised based on the latest work. The abundant Hsandagolian rodent fossils are mainly distributed in northern China, but few materials have been reported in South China.
Cenozoic sediments of the Valley of Lakes are rich in fossils that provide unique evidence of mammal evolution in Mongolia. The strata are intercalated with basalt flows, and 40Ar/39Ar data of the basalts frame the time of sediment deposition and mammal evolution and enable a composite age chronology for the Valley of Lakes area. Based on the radiometric ages of basalt and magnetostratigraphic data, the ages of Mongolian biozones A-C range from 33 to 25.6 Ma and correspond to the Hsandagolian (Daxner-Höck et al., 2010; Kraatz and Geisler, 2010). A total of 70 fossil beds in the Taatsiin Gol and Taatsiin Tsagaan Nuur areas in the Valley of Lakes yielded more than 19,000 mammal fossils, and rodent species composed 50% of the 175 mammal species in the study area (Daxner-Höck et al., 2017). Recently, a rich and diverse fossil assemblage in the Shine Us locality in southwestern Mongolia was described (Daxner-Höck et al., 2019).
The compositions of the Hsandagolian rodent assemblage in China and Mongolia are similar, and they include diverse small- to middle-sized rodents, such as dipodids, cricetids, ctenodactylids, cylindrodontids, eomyids, and large body-sized rodent tsaganomyids (Figure 7). Many genera in the Hsandagolian rodent assemblage are present in both Mongolia and China. In addition, in the Hsandagolian assemblage, castorids are found in China, and scuirids are present in Mongolia. In the Hsandagolian rodent assemblage in East Asia, cricetids, dipodids, and ctenodactylids are dominant, and some rodents are the earliest records of their family in China or Mongolia, such as Tsaganomyidae, Castoridae, and Aplodontidae.
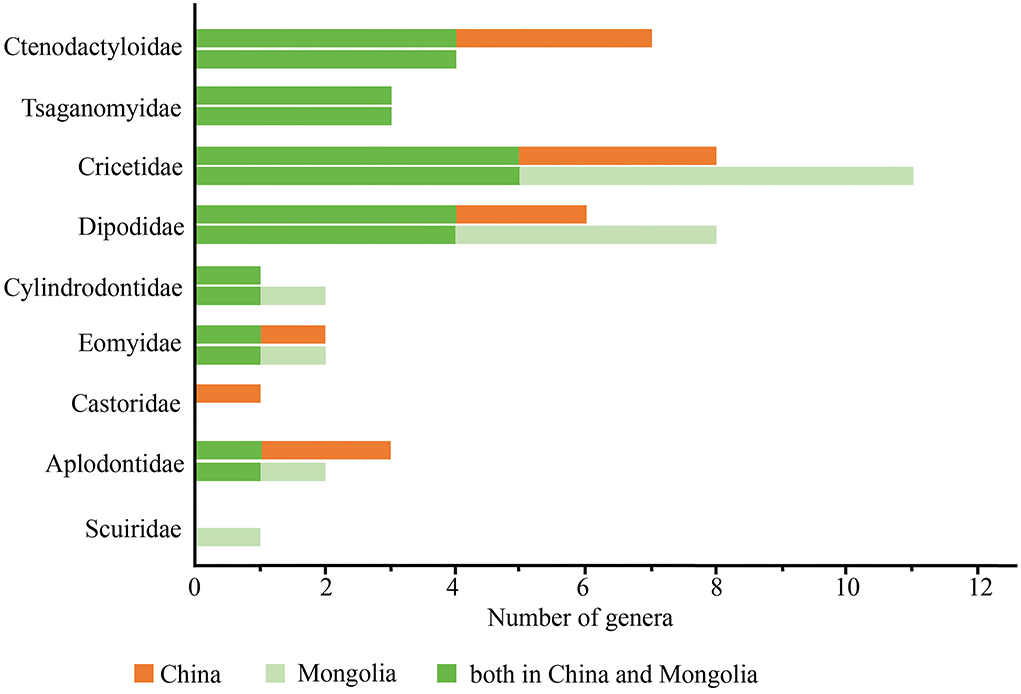
Figure 7. Diagrams of the Hsandagolian rodent composition in China and Mongolia.
The Asian record of the Dipodidae and Cricetidae dates back to the middle Eocene, where those families diversified and played a dominant role in Oligocene rodent communities. Recently, some dipodids and cricetids have been reported in European Oligocene strata (de Bruijn et al., 2019; Wessels et al., 2020), and these materials are the nearest fossil relatives to those in Asian. As a result, dipodids and cricetids immigrated from Asia to Europe in the early Oligocene, and rodent faunal exchange over a large area was possible during the period, such as Dipodidae and cricetid Pappocricetodontinae.
A few rodent materials in Kazakhstan have been reported by Shevyreva (1971), and they are described as Karakoromys and Prosciurus. In the southwestern Sulaiman geological province (Balochistan, Pakistan), terrestrial detrital facies from the Bugti Hills region have yielded rich tertiary vertebrate faunas. The age of the lower Chitarwata Formation in the Bugti Hills is still controversial (Métais et al., 2017), but the rodents and the perissodactyls from the lower Chitarwata Formation clearly suggest an early Oligocene age (Marivaux et al., 1999; Welcomme et al., 2001; Marivaux and Welcomme, 2003; Métais et al., 2009; Antoine et al., 2013), which is correlative with the MP21-22 bio-horizons in Europe. The Bugti rodent fauna did not consist only of buluchimyines that were initially interpreted as a subfamily of the ctenodactyloid Chappatimyidae (sciurognathous) by Flynn et al. (1986) and yielded dipodids, anomaluroids, sciuroids, diatomyids, and typical early Oligocene cricetids (Appendix Table 3). Recently, a phylogenetic analysis showed no support for the monophyly of a baluchimyine clade, so “baluchimyines” were reinterpreted as Hystricognathi incertae sedis, and the phylogenetic results supported that “baluchimyines” and tsaganomyids were representatives of an initial phase of the diversification of hystricognathous rodents in Asia (Marivaux et al., 2002). The Pakistan Hsandagolian rodent fauna is significantly different from that in China and Mongolia; only Cricetidae and Dipodidae at the family level are common elements in East and South Asian rodent faunas, but the genera of these two families are different.
Discussion
South Asian rodent faunas
Using the updated Paleogene timescale in Asia (Wang et al., 2019; Speijer et al., 2020), we compiled a list of Asian rodent genera from the early Eocene to the early Oligocene (Appendix Tables 1–3). In general, the most complete rodent record from the early Eocene to the early Oligocene in Asia is recorded in East Asia, followed by South Asia, and there are fewer Central Asian rodent records. Central Asia and East Asia share many common elements at the levels of rodent families and genera, but the rodent faunas in South Asia show unique characteristics.
In the Bumbanian ischyromyid Meldimys from India, which is close to the European ischormyids, the chapattimyids of Ctenodactyloidea are the only rodent group during the Arshantan and Irdinmanhan (Thewissen et al., 2001; Gupta and Kumar, 2015). In recent years, studies have suggested that some mammals from the Indian subcontinent and Asia mainland, such as perissodactyls and primates, have close phylogenetic connections at the genus level (Rose et al., 2009; Missiaen and Gingerich, 2012, 2014). These facts indicate that there was no impassable geographic barrier for the early Eocene mammals on the two patches of land. Ni et al. (2020) also pointed out that before the Sharamurunian of the Eocene, the mammalian faunas on the Indian subcontinent formed a relatively isolated group and were different from those on the Asian mainland. A recent study suggested the probable presence of strong seasonal rainfall similar to the modern South Asian monsoon in early Eocene India (Spicer et al., 2016; Ding et al., 2017). Obviously, the paleoenvironment in South Asia was different from the environment in East and Central Asia during the early Eocene. Therefore, before the Sharamurunian of the Eocene, the rodent faunas in South Asia formed a relatively isolated group and included some endemic species probably because the difference was caused by different paleoenvironments.
In the Sharamurunian assemblage, the anomaluroid Pondaungimys was found in Myanmar (Dawson et al., 2003b; Marivaux et al., 2005), and Hystricognathi Baluchimys was reported in the Ergilian in Thailand (Marivaux et al., 2000). They had close phylogenetic relationships with rodents from Africa and suggest the occurrence of faunal exchanges between South Asia and Africa during the Sharamurunian and Ergilian. Recent research has shown that since the Sharamurunian of the Eocene, the mammalian faunas in South and East Asia have started to mix together (Ni et al., 2020), but rodent faunas have not exhibited this situation. Rodent fossils from South Asia are present only in limited locations, and only a few specimens are present, so more complete and numerous specimens are needed to assess the rodent relationship between South and East Asia.
The Hsandagolian rodents from East Asia are mainly concentrated in the northern areas of East Asia (northern China and southern Mongolia). The Hsandagolian rodent faunas in South Asia are significantly different from those in China and Mongolia. Ni et al. (2016) suggested that the Indian subcontinent and Yunnan maintained large areas of tropical jungle habitats in the early Oligocene (Ni et al., 2016). Pound and Salzmann (2017) used an extensive palynological dataset and presented global vegetation and terrestrial temperature reconstructions for the Eocene–Oligocene transition (EOT). As a result, at the boundary of the Eocene and Oligocene, the reconstructed terrestrial temperatures showed a decline in the mean annual temperature ranges in the area that included northern China and southern Mongolia; by contrast, no change in temperature was reconstructed for India and tropical Asia. In addition, recent paleoclimate research has suggested that the Central Asian (Mongolia and northern China) steppe desert has existed since at least the Eocene (Barbolini et al., 2020). Obviously, during the early Oligocene, the paleoenvironment was different between South Asia and northern China and southern Mongolia, and the striking difference in the paleoenvironment probably caused the difference in rodent faunas.
East Asian rodent succession from the early Eocene to the early Oligocene and responses to paleoclimatic changes
In general, from the early Eocene to the early Oligocene, rodent faunas in East Asia have obvious succession, and generic rodent diversity fluctuated in relation to paleoclimatic changes. At the beginning of the early Eocene, ctenodactyloid rodents were the dominant elements and moderately diverged as early as the earliest Eocene during the Paleocene–Eocene thermal maximum (PETM) (Figures 8, 9, Table 2). An abrupt increase in diversity during the Irdinmanhan and Sharamurunian is likely related to the rising temperatures of the middle Eocene climatic optimum. Cricetids and dipodids were first found in the Irdinmanhan and Sharamurunian and had some diversity.
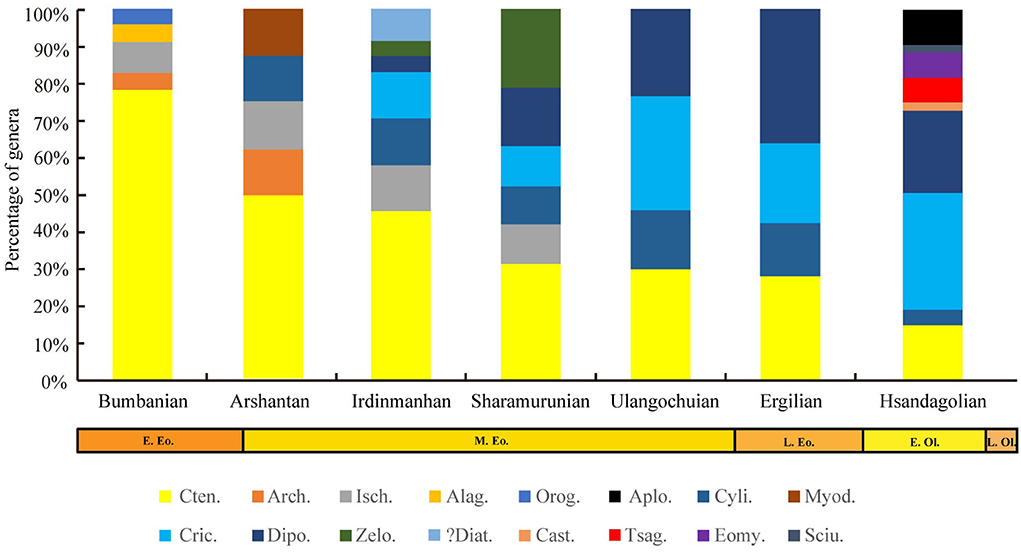
Figure 8. East Asian rodent composition from the early Eocene to the early Oligocene. Aplo., Aplodontidae; Arch., Archetypomyidae; Alag., Alagomyidae; Cast., Castoridae; Cten., Ctenodactyloidea; Cric., Cricetidae; Cyli., Cylindrodontidae; Diat., Diatomyidae; Dipo., Dipodidae; Eomy., Eomyidae; Isch., Ischromyidae; Myod., Myodonta; Sciu., Sciuridae; Tsag., Tsaganomyidae; Orog., Orogomyidae; Zelo., Zelomyidae.
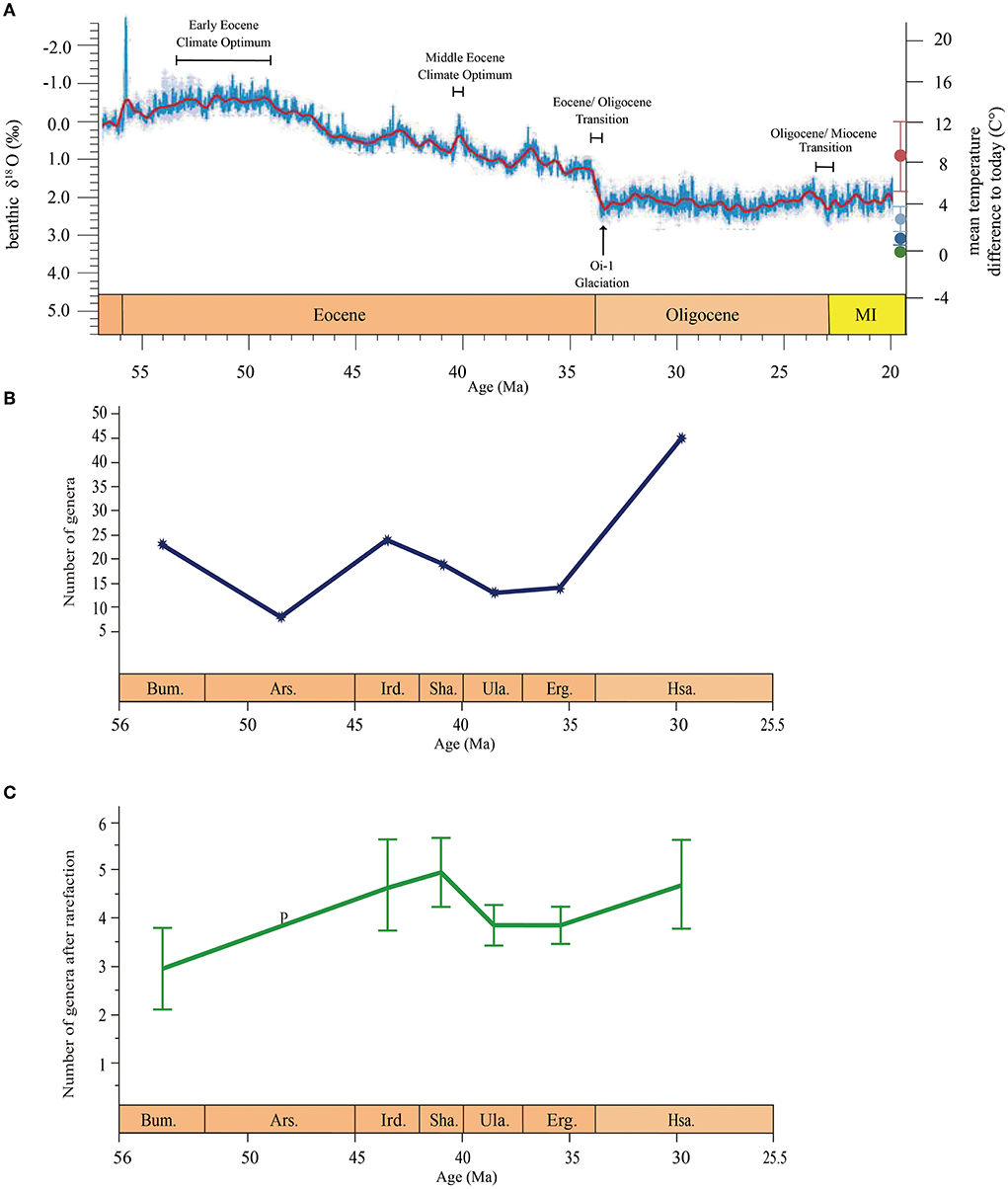
Figure 9. (A,B) East Asian rodent genus-level diversity from the early Eocene to the early Oligocene in relation to global climatic change (modified from Westerhold et al., 2020). (C) Rodent diversity trajectory based on rarefaction estimates of genus richness for each series at a sample size of five genus occurrences. Error bars depict 95% confidence intervals around the estimates. Bum., Bumbanian; Ars., Arshantan; Ird., Irdinmanhan; Sha., Sharamurunian; Ula., Ulangochuian; Erg., Ergilian; Hsa., Hsandagolian.
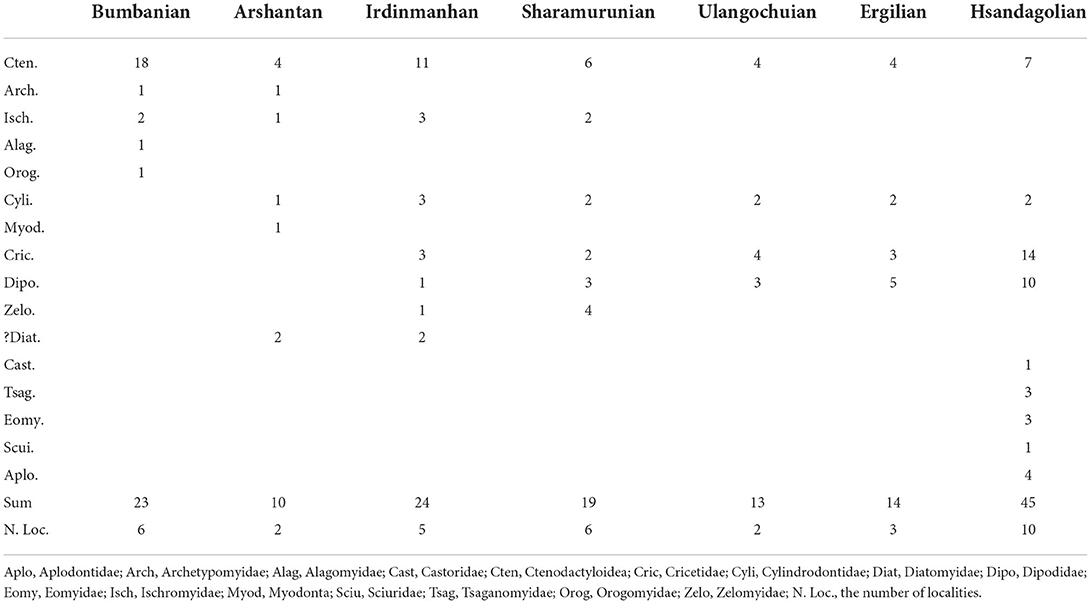
Table 2. Generic numbers of different rodent groups from early Eocene to early Oligocene in East Asia.
A conspicuous event occurred between the Sharamurunian and the Ulangochuian; the generic diversity of rodents was reduced (Figure 9), and the ctenodactyloid-dominant rodent faunas were gradually replaced by the cricetid–dipodid-dominant faunas (Figure 8). Based on the MNI, the rodent assemblages of the Erden Obo section in Nei Mongol also showed a transformation from ctenodactyloid-dominant to cricetid–dipodid-dominant faunas in the Eocene Ulangochuian (Li, 2018). Similarly, perissodactyls and the entire mammalian fauna from China showed a similar abrupt decrease after the Sharamurunian in terms of the number of both species and genera (Wang et al., 2007; Bai et al., 2020). Bai et al. (2020) named the event the “Ulan Gochu Decline”, and the event was comparable to the contemporaneous post-Uintan decline of the North American land fauna (Berggren and Prothero, 1992) and probably related to the sustained cooling following the middle Eocene climatic optimum (MECO).
The Asian mammalian faunal turnover during the EOT was known as the “Mongolian remodeling”, with the perissodactyl-dominant fauna replaced by the rodent/lagomorph-dominant fauna, and it was a turnover that was attributed to the dramatic drop in temperature at the end of the Eocene (Meng and McKenna, 1998). Perissodactyls in China showed a decrease in terms of generic number during the EOT, but the decline was not as obvious as in the “Ulan Gochu Decline” (Bai et al., 2020). However, during the EOT, the generic diversity of rodents obviously increased (Figure 9), cricetid–dipodids were still dominant elements in rodent faunas (Figure 8), and the generic diversity of cricetids and dipodids also obviously increased compared with than that in the Ulangochuian and Ergilian.
In summary, the Eocene rodent faunas in East Asia showed two different changes in diversity, which may be related to global climatic declines. The first change occurred after the MECO when temperatures declined slowly and manifested as a clearly decreased diversity of rodents and a transformation from ctenodactyloid-dominant to cricetid–dipodid-dominant faunas. The second change occurred at the EOT and may have been a response to the sudden global drop in temperature; the change was manifested as a high generic diversity of cricetids, dipodids, and ctenodacylids and the appearance of rodent families such as Tsaganomyidae, Castoridae, and Eomyidae.
Based on the aforementioned discussion, after the dramatic drop in temperature, rodents became the dominant element in the Oligocene mammal fauna, but not overnight. The low temperatures during the Ulangochuian and Ergilian in East Asia served as a habituation ground for the cricetids and dipodids, which became preadapted for the EOT, successfully helping the rodent become the dominant fauna after the EOT.
Conclusion
Successive Asian rodent faunas from the early Eocene to the early Oligocene are recorded in East Asia, and the rodent records in Central and South Asia are incomplete. Central Asia and East Asia share many common elements at the rodent family and genus levels. The rodent faunal turnover in East Asia was obviously affected by paleoclimatic changes. During the Irdinmanhan and Sharamurunian, East Asian rodent faunas showed an increase in the diversity of genera and families, which may be related to the rising temperatures of the mid-Eocene climatic optimum. In the Ulangochuian (after the MECO), when temperatures declined slowly, the rodent fauna showed a clear decline in the diversity at the genus level and a transformation from ctenodactyloid-dominant to cricetid–dipodid-dominant faunas. During the Eocene–Oligocene transition (EOT) and global cooling, the East Asian rodent fauna is shown by the considerably high generic diversity of cricetids, dipodids, and ctenodactylids. The low temperatures during the Ulangochuian and the Ergilian in East Asia served as a habituation ground for the cricetids and dipodids, which became preadapted for the EOT, successfully helping the rodent as the dominant fauna after the EOT.
Before the Sharamurunian of the Eocene and the early Oligocene, the South Asian rodent faunas showed unique characteristics, and the striking differences between the rodent faunas in South Asia and those in East Asia were probably the result of different paleoenvironmental habitats. In the Sharamurunian and Ergilian, rodents in South Asia have close phylogenetic relationships with rodents from Africa and suggest the occurrence of faunal exchanges between South Asia and Africa.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
QiaL designed the study and wrote the manuscript. QiL and RX analyzed the data. YW helped revise the manuscript and gave some useful suggestions. All authors contributed to the manuscript and approved the submitted version.
Funding
This research was funded by the National Natural Science Foundation of China (Grant 42072023) and the Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000). The fieldwork was also supported by the Special Fund for Fossil Excavation and Preparation, CAS.
Acknowledgments
We thank Jin Meng, Bin Bai, Xun Jin, Ping Li, K. C. Beard, D. L. Gebo, Xijun Ni, Wei Chen, Wei Zhou, Wei Gao, and Shijie Li for their assistance in the fieldwork. Wang Bian helped improve English in the text. We are grateful to the editors and reviewers for their constructive comments on our manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.955779/full#supplementary-material
References
Anderson, D. (2008). “Ischyromidae,” in Evolution of Tertiary Mammals of North American, eds. C. M. Janis, G. F. Gunnell, and M. D. Uhen (New York: Cambridge University press) 311–325.
Antoine, P. O., Métais, G., Orliac, M. J., Crochet, J. Y., Flynn, L. J., Marivaux, L., et al. (2013). “Mammalian Neogene biostratigraphy of the Sulaiman Province, Pakistan,” in Fossil Mammals of Asia, eds. X. Wang, L. J. Flynn, and M. Fortelius (San Francisco: California University Press), 400–422. doi: 10.7312/columbia/9780231150125.003.0016
Averianov, A. O. (1996). Early Eocene Rodentia of Kyrgyzstan. Bull. Mus. Natl. Hist. Nat. 18C, 663–671. doi: 10.1127/njgpm/1996/1996/739
Bai, B., Meng, J., Janis, C. M., Zhang, Z. Q., and Wang, Y. Q. (2020). Perissodactyl diversities and responses to climate changes as reflected by dental homogeneity during the Cenozoic in Asia. Ecol. Evol. 00, 1–23. doi: 10.1002/ece3.6363
Barbolini, N., Woutersen, A., Dupont-Nivet, G., Silvestro, D., Tardif, D., Coster, P. M. C., et al. (2020). Cenozoic evolution of the steppe-desert biome in Central Asian. Sci. Adv. 6, eabb8227. doi: 10.1126/sciadv.abb8227
Berggren, W. A., and Prothero, D. R. (1992). “Eocene-Oligocene climatic and biotic evolution: An overview.” in Eocene–Oligocene climatic and biotic evolution, eds. W. A. Berggren, and D. R. Prothero (Princeton, NJ: Princeton University Press), 1–28. doi: 10.1515/9781400862924.1
Dashzeveg, D. (1990a). The earliest rodents (Rodentia, Ctenodactyloidea) of Central Asia. Acta Zool. Cracov. 33, 11–35.
Dashzeveg, D. (1990b). New trends in adaptive radiation of Early Tertiary rodents (Rodentia, Mammalia). Acta Zool. Cracov. 33, 37–44.
Dashzeveg, D. (1993). A synchronism of the main mammalian faunal events near the Eocene–Oligocene boundary. Ter. Res. 14, 141–149.
Dashzeveg, D., and Meng, J. (1998). New Eocene ctenodactyloid rodents from the eastern Gobi desert of Mongolia and a phylogenetic analysis of ctenodactyloids based on dental features. Am. Muse. Novit. 3246, 1–20.
Dawson, M. R. (1968). Oligocene rodents (Mammalia) from East Mesa, Inner Mongolia. Am. Muse. Novit. 2324, 1–12.
Dawson, M. R. (2015). “Emerging perspectives on some Paleogene sciurognath rodents in Laurasia: the fossil record and its interpretation,” in Evolution of the Rodents: Advances in Phylogeny, Functional Morphology and Development, eds. P. G. Cox and L. Hautier (Cambridge: Cambridge University Press), 70–86. doi: 10.1017/CBO9781107360150.004
Dawson, M. R., Huang, X. S., Li, C. K., and Wang, B. Y. (2003a). Zelomyidae, a new family of Rodentia (Mammalia) from the Eocene of Asia. Vert. PalAsiat. 41, 249–270.
Dawson, M. R., Li, C. K., and Qi, T. (1984). Eocene ctenodactyloid rodents (Mammalia) of eastern and central Asia. Carnegie Mus. Na.t Hist. Special Publ. 9, 138–150.
Dawson, M. R., Li, C. K., and Qi, T. (2010). The diatomyidae (Mammalia, Rodentia) and bilophodonty in Middle Eocene Asian rodents. Vert. PalAsiat. 48, 328–335.
Dawson, M. R., Tsubamoto, T., Takai, M., Egi, N., Tun, S. T., and Sein, C. (2003b). Rodents of the family Anomaluridae (Mammalia) from Southeast Asia (middle Eocene, Pondaung Formation, Myanmar). Ann. Carnegie Mus. 72, 203–213. doi: 10.5962/p.215090
Daxner-Höck, G., Badamgarav, D., Barsbold, R., Bayarmaa, B., Erbajeva, M., Göhlich, U. B., et al. (2017). Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Palaeobio. Palaeoenv. 97, 111–218. doi: 10.1007/s12549-016-0257-9
Daxner-Höck, G., Badamgarav, D., and Erbajeva, M. (2010). Oligocene stratigraphy based on a sediment-basalt association in Central Mongolia (Taatsiin Gol and Taatsiin Tsagaan Nuur Area, Valley of Lakes): review of a Mongolian-Austrian project. Vert. PalAsiat. 48, 348–366.
Daxner-Höck, G., Erbajeva, M. A., Göhlich, U. B., Lopez-Guerrero, P., Narantsetseg, T., Mennecart, B., et al. (2019). The Oligocene vertebrate assemblage of Shine Us (Khaliun Basin, south western Mongolia). Ann. Naturhist. Mus. Wien, Serie A. 121, 195–256.
de Bruijn, H., Marković, Z., Wessels, W., and van de Weerd, A. A. (2019). Pappocricetodontinae (Rodentia, Muridae) from the Paleogene of south-east Serbia. Palaeobio. Paleoenv. 99, 511–526. doi: 10.1007/s12549-018-0343-2
Ding, L., Spice, R. A., Yang, J., Xu, Q., Cai, F., Li, S., et al. (2017). Quantifying the rise of the Himalaya orogen and implications for the South Asian monsoon. Geology. 45:215–218. doi: 10.1130/G38583.1
Emry, R. J., Lucas, S. G., Tyutkova, L. A., and Wang, B. Y. (1998a). The Ergilian-Shandgolian (Eocene-Oligocene) transition in the Zaysan Basin, Kazakstan. Bull. Carnegie Mus. Nat. Hist. 34, 298–312.
Emry, R. J., Tyutkova, L. A., Lucas, S. G., and Wang, B. Y. (1998b). Rodents of the middle Eocene Shinzhaly fauna of Eastern Kazakstan. J. Vert. Pale. 18, 218–227. doi: 10.1080/02724634.1998.10011045
Fejfar, O., Rummel, M., and Tomida, Y. (1998). “New eomyid genus and species from the Early Miocene (MN zones 3-4) of Europe and Japan related to Apeomys (Eomyidae, Rodentia, Mammalia).” In: Advances in Vertebrate Paleontology and Geochronology, eds. Tomida, Y., Flynn, L. J., Jacpbs, I. I. (Tokyo: Nat Sci Mus Monographs), 14, 123–143.
Flynn, L. J., Jacobs, L. L., and Cheema, I. U. (1986). Baluchimyinae. A new Ctenodactyloid Rodent subfamily from the Miocene of Baluchistan. Am. Muse. Novit. 2841, 1–58.
Fostowicz-Frelik, L., López-Torres, S., and Li, Q. (2021). Tarsal morphology of ischyromyids rodents from the middle Eocene of China gives an insight into the group's diversity in Central Asia. Sci. Rep. 11:11543. doi: 10.1038/s41598-021-90796-1
Gomes Rodrigues, H., Marivaux, L., and Vianey-Liaud, M. (2014). Rodent paleocommunities from the Oligocene of Ulantatal (Inner Mongolia, China). Palaeovertebrata. 38, 1–12. doi: 10.18563/pv.38.1.e3
Gotelli, N. J., and Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 4, 379–391. doi: 10.1046/j.1461-0248.2001.00230.x
Guo, J. W., Wang, Y., and Yang, X. A. (2000). A new early Eocene ctenodactyloid rodent (Rodentia, Mammalia) and the associated mammalian fossils from Danjiangkou, Hubei. Vert. PalAsiat. 38, 303–313.
Gupta, S., and Kumar, K. (2015). Early Eocene rodents (Mammalia) from the Subathu Formation of type area (Himachal Pradesh), NW sub-Himalaya, India: Palaeobiogeographic implications. J. Earth Syst. Sci. 124, 1201–1221. doi: 10.1007/s12040-015-0607-4
Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: Palaeontological statistics software package for education and data analysis. Pala. Elec. 4, 1–9.
Hartenberger, J. L. (1982). A review of the Eocene rodents of Pakistan. Contrib. Mus. Paleontol. Univ. Michigan. 26, 19–35.
Hu, Y. M. (1995). New late early Eocene ctenodactyloid rodents (Rodentia, Mammalia) from Danjiangkou, Hubei. Vert. PalAsiat. 33, 24–38.
Huang, B. (2012). Rarefaction and its application to the study of diversity of palaeocommunities. Acta Palae. Sin. 51, 200–208.
Hussain, S. T., de Bruijn, H., and Leinders, J. M. (1978). Middle Eocene rodents from the Kala Chitta Range (Punjab, Pakistan). Proc. Konink. Nederl. Akad. Wetensch. Ser. B. 81, 74–112.
Jaeger, J.-J., Chavasseau, O., Lazzari, V., Soe, A. N., Sei, C., Maitre, A. L., et al. (2019). New Eocene primate from Myanmar shares dental characters with African Eocene crown anthropoids. Nature Comm. 10, 3511. doi: 10.1038/s41467-019-11295-6
Khin, Z., Meffre, S., Takai, M., Suzuki, H., Burret, C., Htike, T., et al. (2014). The oldest anthropoid primates in SE Asia: Evidence from LA-ICP-MS U-Pb zircon age in the Late Middle Eocene Pondaung Formation, Myanmar. Gondwana Res. 26, 122–131. doi: 10.1016/j.gr.2013.04.007
Kraatz, P., and Geisler, J. H. (2010). Eocene-Oligocene transition in Central Asia and its effects on mammalian evolution. Geology. 38, 111–114. doi: 10.1130/G30619.1
Kumar, K., Loyal, R. S., and Srivastava, R. (1997a). Eocene rodents from new localities in Himachal Pradesh, northwest Himalaya, India: Biochronologic implications. J. Geol. Soc. India. 50, 461–474.
Kumar, K., Srivastava, R., and Sahni, A. (1997b). Middle Eocene rodents from the Subathu Group, northwest Himalaya. Palaeovertebrata. 26, 83–128.
Li, C. K. (1975). Yuomys, a new ischyromyoid rodent genus from the Upper Eocene of North China. Vert. PalAsiat. 13, 58–70.
Li, C. K., Zheng, J. J., and Ting, S. Y. (1989). “The skull of Cocomys lingchaensis, an Early Eocene ctenodactyloid rodent of Asia,” in Papers on Fossil Rodents in Honor of Albert Elmer Wood, eds. C. C. Black, and M. R. Dawson (Los Angeles, NHMLAC Press), 179–192.
Li, L. Z., Ni, X. J., Lu, X. Y., and Li, Q. (2017). First record of Cricetops rodent in the Oligocene of southwestern China. Hist. Biol. 29, 488–494. doi: 10.1080/08912963.2016.1196686
Li, Q. (2012). Middle Eocene cricetids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. PalAsiat. 50, 237–244.
Li, Q. (2016). Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): Biochronological implications. Palaeoworld. 25, 95–103. doi: 10.1016/j.palwor.2015.07.001
Li, Q. (2018). Additional cricetid and dipodid rodent material from the Erden Obo section, Erlian Basin (Nei Mongol, China) and its biochronological implications. Palaeoworld. 27, 490–505. doi: 10.1016/j.palwor.2018.09.003
Li, Q. (2019). Eocene ctenodactyloid rodent assemblages and diversification from Erden Obo, Nei Mongol, China. Hist. Biol. 31, 813–823. doi: 10.1080/08912963.2017.1395422
Li, Q. (2021). Additional tsaganomyid, cylindrodontid and ctenodactyloid rodent materials from the Erden Obo section, Erlian Basin (Nei Mongol, China). Vert. PalAsiat. 59, 1–18.
Li, Q., Mao, F. Y., and Wang, Y. Q. (2018). First record of Eocene fossil rodent assemblages from the lower part of the Erden Obo section, Erlian Basin (Nei Mongol, China) and its biochronological implications. Palaeobio. Paleoenv. 98, 259–276. doi: 10.1007/s12549-017-0303-2
Li, Q., and Meng, J. (2010). Erlianomys combinatus, a primitive myodont rodent from the Eocene Arshanto Formation, Nuhetingboerhe, Nei Mongol, China. Vert. PalAsiat. 48, 133–144.
Li, Q., and Meng, J. (2013). Eocene ischyromyids (Rodentia, Mammalia) from the Erlian Basin, Nei Mongol, China. Vert. PalAsiat. 51, 289–304.
Li, Q., and Meng, J. (2015). New ctenodactyloid rodents from the Erlian Basin, Nei Mongol, China, and the phylogenetic relationships of Eocene Asian ctenodactyloids. Am. Muse. Novit. 3828, 1–58. doi: 10.1206/3828.1
Li, Q., Wang, Y. Q., and Fostowicz-Frelik, Ł. (2016). Small mammal fauna from Wulanhuxiu (Nei Mongol, China) implies the Irdinmanha-Sharamurunian (Eocene) faunal turnover. Acta Palaeontol. Pol. 61, 759–776. doi: 10.4202/app.00292.2016
Li, Q., Wang, Y. Q., Mao, F. Y., and Meng, J. (2019). A new Eocene cylindrodontid rodent from the Erlian Basin (Nei Mongol, China) and its implications for phylogeny and biochronolog. J. Vert. Paleont. 39, 5,e1680990. doi: 10.1080/02724634.2019.1680990
Luterbacher, H. P., Ali, J. R., Brinkhuis, H., Gradstein, F. M., Hooker, J. J., Monechi, S., et al. (2004). “The Paleogene Period”, in A Geological Time Scale 2004, eds. F. M. Gradstein, J. G. Ogg, and A. Smith (Cambridge: Cambridge University Press) 384–408. doi: 10.1017/CBO9780511536045.021
Maridet, O., and Ni, X. J. (2013). A new cricetid rodent from the early Oligocene of Yunnan, China, and its evolutionary implications for early Eurasian cricetids. J. Vert. Paleon. 33, 185–194. doi: 10.1080/02724634.2012.710283
Marivaux, L., Benammi, M., Ducrocq, S., Jaeger, J. J., and Chaimanee, Y. (2000). A new baluchimyine rodent from the Late Eocene of the Krabi Basin (Thailand): palaeobiogeographic and biochronologic implications. Earth Planet Sci. 331, 427–433. doi: 10.1016/S1251-8050(00)01427-0
Marivaux, L., Ducrocq, S., Jaeger, J. J., Marandat, B., Sudre, J., Chaimanee, Y., et al. (2005). New remains of Pondaungimys anomaluropsis (Rodentia, Anomaluroidae) from the latest Middle Eocene Pondaung Formation of central Myanmar. J. Vert. Paleon. 25, 214–227. doi: 10.1671/0272-4634(2005)025[0214:NROPAR]2.0.CO;2
Marivaux, L., Vianey-Liaud, M., and Welcomme, J. L. (1999). Première découverte de Cricetidae (Rodentia, Mammalia) oligocènes dans le synclinal sud de Gandoï (Bugti Hills, Balouchistan, Pakistan). CR l'Acad. Sci. Paris, Ser. IIa. 329, 839–844. doi: 10.1016/S1251-8050(00)88640-1
Marivaux, L., Vianey-Liaud, M., Welcomme, J. L., and Jaeger, J. J. (2002). The role of Asia in the origin and diversification of hystricognathous rodents. Zool. Scr. 31, 225–239. doi: 10.1046/j.1463-6409.2002.00074.x
Marivaux, L., and Welcomme, J. L. (2003). New diatomyid and baluchimyine rodents from the Oligocene of Pakistan (Bugti Hills, Balochistan): systematic and paleobiogeographic implications. J. Vertebr. Paleontol. 23, 420–434. doi: 10.1671/0272-4634(2003)023[0420:NDABRF]2.0.CO;2
McKenne, M. C., and Bell, S. K. (1997). Classification of Mammals Above the Species Level. New York: Columbia University Press.
Mein, P., and Ginsburg, L. (1997). Les mammiféres du gisement miocéne infêrieur de Li Mae Long, Thailande. Geodiversitas. 19, 783–844
Meng, J., and Li, C. K. (2010). New rodents from the earliest Eocene of Nei Mongol, China. Vert. PalAsiat. 48, 390–401.
Meng, J., Li, C. K., Ni, X. J., Wang, Y. Q., and Beard, K. C. (2007). A new Eocene rodent from the lower Arshanto Formation in the Nuhetingboerhe (Camp Margetts) area, Inner Mongolia. Am. Muse. Novit. 3569, 1–18. doi: 10.1206/0003-0082(2007)536[1:ANERFT]2.0.CO;2
Meng, J., and McKenna, M. C. (1998). Faunal turnovers of Palaeogenemammals from the Mongolian plateau. Nature. 394, 364–367. doi: 10.1038/28603
Meng, J., Wu, W. Y., and Ye, J. (2001). A new species of Advenimus (Rodentia, Mammalia) from the Eocene of northern Junggar Basin of Xinjiang, China. Vert. PalAsiat. 39, 185–196.
Meng, J., and Wyss, A. R. (2001). The morphology of Tribosphenomys (Rodentiaformes, Mammalia): phylogenetic implications for basal Glires. J. Mammal Evol. 8, 1–71. doi: 10.1023/A:1011328616715
Meng, J., Ye, J., and Huanag, X. S. (1999). Eocene mammals from the Bayan Ulan of Nei Mongol (Inner Mongolia) and comments on related stratigraphy. Vert PalAsiat. 37, 165–174.
Métais, G., Antoine, P. O., Baqri, S. R. H., Crochet, J. Y., De Franceschi, D., Marivaux, L., et al. (2009). Lithofacies, depositional environments, regional biostratigraphy and age of the chitarwata formation in the Bugti Hills, Balochistan, Pakistan. J. Asian Earth Sci. 34, 154–167. doi: 10.1016/j.jseaes.2008.04.006
Métais, G., Mennecart, B., and Roohi, G. (2017). A new assemblage of stem pecoran ruminants from the Oligocene Chitarwata Formation, Bugti Hills, Baluchistan, Pakistan: Paleoenvironmental and paleobiogeographic implications. J. Asian Earth Sci. 136, 40–49. doi: 10.1016/j.jseaes.2016.09.009
Missiaen, P., and Gingerich, P. D. (2012). New Early Eocene tapiromorph perissodactyls from the Ghazij Formation of Pakistan, with implications for mammalian biochronology in Asia. Acta Palaeontol. Pol. 57, 21–34. doi: 10.4202/app.2010.0093
Missiaen, P., and Gingerich, P. D. (2014). New basal perissodactyla (Mammalia) from the lower Eocene Ghazij Formation of Pakistan. Contrib. Muse. Paleont. Univ. Michigan 32, 139–160.
Ni, X. J., Li, Q., Li, L. Z., and Beard, K. C. (2016). Oligocene primates from China reveal divergence between African and Asian primate evolution. Science. 352(6286), 673–677. doi: 10.1126/science.aaf2107
Ni, X. J., Li, Q., Zhang, C., Samiullah, K., Zhang, L. M., Yang, Y. H. S., et al. (2020). Paleogene mammalian fauna exchanges and the paleogeographic pattern in Asia. Sci. China Earth Sci. 63, 202–211. doi: 10.1007/s11430-019-9479-1
Pound, M. J., and Salzmann, U. (2017). Heterogeneity in global vegetation and terrestrial climate change during the late Eocene to early Oligocene transition. Sci. Rep. 7, 43386. doi: 10.1038/srep43386
Qiu, Z. D., and Wang, B. Y. (2019). “Dipodidae,” in Palaeovertebrata Sinica, VIII, Basal Synapsids and Mammals, Fascicle 5(2), Glires II: Rodentia I, eds. Z. X. Qiu, and C. K. Li (Beijing: Science Press), 242–325.
Rana, R. S., Kumar, K., Escarguel, G., Sahni, A., Rose, K. D., Smith, T., et al. (2008). An ailuravine rodent from the lower Eocene Cambay Formation at Vastan, western India, and its palaeobiogeographic implications. Acta Palaeontol. Pol. 53, 1–14. doi: 10.4202/app.2008.0101
Rose, K. D., Rana, R. S., Sahni, A., Kumar, K., Missiaen, P., et al. (2009). Early Eocene Primates from Gujarat, India. J. Human Evol. 56, 366–404. doi: 10.1016/j.jhevol.2009.01.008
Sahni, A., and Khare, S. K. (1973). Additional Eocene mammals from the Subathu Formation of Jammu and Kashmir. J. Palaeontol. Soc. India. 17, 31–49.
Sahni, A., and Srivastava, V. C. (1976). Eocene rodents and associated reptiles from the Subathu Formation of northwestern India. J. Paleontol. 50, 922–928.
Shen, S. Z., Zhang, H., and Li, W. Z. (2004). An introduction of methods for removing biases in establishing biodiversity patterns from fossil records. Acta Palae. Sin. 43, 433–441.
Shevyreva, N. S. (1971). New Middle Oligocene rodents of Kazakhstan and Mongolia. Bull. Aca. Sci. SSR.130, 70–86.
Shevyreva, N. S. (1984). “New rodents from the Early Eocene of the Zaisan Depression,” in Floras and Faunas of the Zaisan Depression, eds. L. S. Daweta, and S. Wele (AH GSSR), 77–114 (in Russian)
Shevyreva, N. S. (1989). New rodents (Ctenodactyloidea, Rodentia, Mammalia) from the Lower Eocene of Mongolia. J. Paleontolo. 3, 60–72.
Shevyreva, N. S. (1996). New rodents (Rodentia, Mammalia) from the Lower Eocene of the Zaisan Depression (Eastern Kazakhstan). J. Paleontolo. 30, 81–94.
Speijer, R. P., Pälike, H., Hollis, C. J., Hooker, J. J., and Ogg, J. G. (2020). “The Paleogene Period,” in Geologic Time Scale 2020, eds. F. M. Gradstein, J.G. Ogg, M.D. Schmitz, and G.M. Ogg (Oxford: Elsevier BV), 1087–1140. doi: 10.1016/B978-0-12-824360-2.00028-0
Spicer, R. A., Yang, J., Herman, A. B., Kodrul, T., Maslova, N., Spicer, T. E. V., et al. (2016). Asian Eocene monsoons as revealed by leaf architectural signatures. Earth Planet. Sc. Lett. 449, 61–68. doi: 10.1016/j.epsl.2016.05.036
Sun, J. M., Ni, X. J., Bi, S. D., Wu, W. Y., Ye, J., Meng, J., et al. (2014). Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene boundary in Asia. Sci. Rep. 4, 7463. doi: 10.1038/srep07463
Thewissen, J. G. M., Williams, E. M., and Hussain, S. T. (2001). Eocene mammal faunas from northern Indo-Pakistan. J. Vert. Paleontol. 21, 347–366. doi: 10.1671/0272-4634(2001)021[0347:EMFFNI]2.0.CO;2
Tong, Y. S. (1997). Middle Eocene small mammals from Liguanqiao Basin of Henan Province and Yuanqu Basin of Shanxi Province, Central China. Paleont. Sin. New Ser. C. 26, 1–256.
Tong, Y. S., and Wang, J. W. (2006). Fossil Mammals from the Early Eocene Wutu Formation of Shandong Province, Central China. Paleont. Sin. New Ser. C. 28, 1–195.
Tong, Y. S., Zheng, S. H., and Qiu, Z. D. (1995). Cenozoic mammal ages of China. Vert. PalAsiat. 33, 290–314.
Tsubamoto, T., Egi, N., Takai, M., Shigehara, N., Aung, A. K., Thein, T., et al. (2000). A preliminary report on the Eocene mammals of the Pondaung fauna, Myanmar. Asian Paleoprimatology. 1, 29–101.
Tsubamoto, T., Takai, M., Shigehara, N., Egi, N., Thura, T. S., Aung, K. A., et al. (2002). Fission-track zircon age of the Eocene Pondaung Formation, Myanmar. J. Hum. Evol. 42, 361–369. doi: 10.1006/jhev.2001.0543
Vandenberghe, N., Hilgen, F. L., and Speijer, R. P. (2012). “The Paleogene Period,” in The Geologic Time Scale 2012, eds. F. M. Gradstein, J. G. Ogg, M. D. Schmitz, and G. M. Ogg (Oxford: Elsevier BV), 855–922. doi: 10.1016/B978-0-444-59425-9.00028-7
Wang, B. Y. (1985). Zapodidae (Rodentia, Mammalia) from the Lower Oligocene of Qujing, Yunnan, China. Mainz. Geowiss. Mitt. 14, 345–367.
Wang, B. Y. (1987). Discovery of Aplodontidae (Rodentia, Mammalia) from middle Oligocene of Nei Mongol, China. Vert. PalAsiat. 25(1), 32–45.
Wang, B. Y. (1997a). The Mid-Tertiary Ctenodactylidae (Rodentia, Mammalia) of eastern and central Asia. Bul. Am. Mus. Nat. Hist. 234: 1–88.
Wang, B. Y. (1997b). Chronological sequence and subdivision of Chinese Oligocene mammalian faunas. J. Stratigr. 21, 183–191.
Wang, B. Y. (2001a). Late Eocene ctenodactyloids (Rodentia, Mammalia) from Qujing, Yunnan, China. Vert. PalAsiat. 39, 24–42.
Wang, B. Y. (2001b). On Tsaganomyidae (Rodentia, Mammalia) of Asia. Am. Muse. Novit. 3317, 1–50. doi: 10.1206/0003-0082(2001)317<0001:OTRMOA>2.0.CO;2
Wang, B. Y. (2001c). Eocene ctenodactyloids (Rodentia, Mammalia) from Nei Mongol, China. Vert. PalAsiat. 39, 98–114
Wang, B. Y. (2008). Additional rodent material from Houldjin Formation of Erenhot, Nei Mongol, China. Vert. PalAsiat. 46(1), 21–30.
Wang, B. Y. (2019). “Cylindrodontidae,” in Palaeovertebrata Sinica, VIII, Basal Synapsids and Mammals, Fascicle 5(2), Glires II: Rodentia I, eds. Z. X. Qiu, and C. K. Li (Beijing: Science Press), 449–478
Wang, B. Y., and Dawson, M. R. (1994). A primitive cricetid (Mammalia: Rodentia) from the Middle Eocene of Jiangsu Provinces, China. Ann. Carnegie Mus. 63, 239–256. doi: 10.5962/p.215814
Wang, B. Y., and Emry, R. J. (1991). Eomyidae (Rodentia: Mammalia) from the Oligocene of Nei Mongol, China. J. Vert. Paleontol. 11, 370–377. doi: 10.1080/02724634.1991.10011404
Wang, B. Y., and Meng, J. (1986). Eucricetodon (Rodentia, Mammalia) from the Lower Oligocene of Qujing, Yunnan, China. Vert. PalAsiat. 24(2),110–120.
Wang, B. Y., and Qiu, Z. X. (2003). Note on Early Oligocene Ursids (Carnivora, Mammalia) from Saint Jacques, Nei Mongol, China. Bull. Am. Mus. Nat. Hist. 279, 116–124. doi: 10.1206/0003-0090(2003)279<0116:C>2.0.CO;2
Wang, B. Y., and Qiu, Z. X. (2004). Discovery of early Oligocene mammalian fossils from Danghe are, Gansu, China. Vert. PalAsiat. 42,130–143.
Wang, B. Y., Qiu, Z. X., Zhang, Q. Z., Wu, L. J., and Ning, P. J. (2009). Large mammals found from Houldjin Formation near Erenhot, Nei Mongol, China. Vert. PalAsiat. 47, 85–110.
Wang, B. Y., Wu, W. Y., and Qiu, Z. X. (2020). “Cricetidae,” in Palaeovertebrata Sinica, VIII, Basal Synapsids and Mammals, Fascicle 5(2), Glires II: Rodentia II, eds. Z. X. Qiu, and C. K. Li (Beijing: Science Press) 12–151.
Wang, B. Y., Yan, Z. Q., Lu, Y. J., and Chen, G. X. (1994). Discovery of two mid-Tertiary mammalian faunas from Haiyuan, Ningxia, China. Vert. PalAsiat. 32, 285–296.
Wang, B. Y., Zhai, R. J., and Dawson, M. R. (1998). Discovery of Ischyromyinae (Rodentia, Mammalia) from the middle Eocene of North China. Vert. PalAsiat. 36, 1–12.
Wang, Y. Q., Li, Q., Bai, B., Jin, X., Mao, F. Y., and Meng, J. (2019). Paleogene integrative stratigraphy and timescale in China. Sci. China Earth Sci. 62, 287–309. doi: 10.1007/s11430-018-9305-y
Wang, Y. Q., Meng, J., Ni, X. J., and Li, C. K. (2007). Major events of Paleogene mammal radiation in China. Geol. J. 42, 415–430. doi: 10.1002/gj.1083
Wasiljeff, J., Kaakinen, A., Salminen, J. M., and Zhang, Z. Q. (2020). Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: implications for Asian aridification and faunal turnover before the Eocene–Oligocene boundary. Earth Planet. Sci. Lett. 535, 116–125. doi: 10.1016/j.epsl.2020.116125
Wasiljeff, J., and Zhang, Z. Q. (2022). Stratigraphical significance of Ulantatal sequence (Nei Mongol, China) in refining the latest Eocene and Oligocene terrestrial regional stages. Vert PalAsiat. 60, 42–53. doi: 10.19615/j.cnki.2096-9899.210716
Welcomme, J. L., Benammi, M., Crochet, J. Y., Marivaux, L., Métais, G., Antoine, P. O., et al. (2001). Himalayan Forelands: palaeontological evidence for Oligocene detrital deposits in the Bugti Hills (Balochistan, Pakistan). Geol. Mag. 138, 397–405. doi: 10.1017/S0016756801005428
Wessels, W., van de Weerd, A. A., de Bruijn, H, and Marković, Z. (2020). Dipodidea (Mammalia, Rodentia) from the Paleogene of south-east Serbia. Palaeobio. Paleoenv. 100, 841–848. doi: 10.1007/s12549-019-00392-5
Westerhold, T., Marwan, N., Drury, A. J., Liebrand, D., Agnini, A., Anagnostou, E., et al. (2020). An astronomically dated record of Earth's climate and its predictability over the last 66 million years. Sci. 369, 1383–1387. doi: 10.1126/science.aba6853
Wible, J. R., Wang, Y. Q., Li, C. K., and Dawson, M. R. (2005). Cranial anatomy and relationships of a new ctenodactyloid (Mammalia, Rodentia) from the Early Eocene of Hubei Province, China. Ann. Carnegie Mus. 74, 91–150. doi: 10.2992/0097-4463(2005)74[91:CAAROA]2.0.CO;2
Wood, A. E. (1980). The Oligocene rodents of North America. Trans. Am. Philos. Soc. 70, 1–68. doi: 10.2307/1006314
Wu, W. Y., Meng, J., Ye, J., and Ni, X. J. (2004). Propalaeocastor (Rodentia, Mammalia) from the Early Oligocene of Burqin Basin, Xinjiang. Am Mus Novit. 3461, 1–16. doi: 10.1206/0003-0082(2004)461<0001:PRMFTE>2.0.CO;2
Ye, J., Meng, J., Wu, W. Y., and Ni, X. J. (2005). Late Eocene-early Oligocene lithological and biological Stratigraphy in the Burqin region of Xinjiang. Vert. PalAsiat. 43, 49−60.
Keywords: Paleogene, climate change, rodent fauna, East Asia, South Asia
Citation: Li Q, Li Q, Xu R and Wang Y (2022) Rodent faunas, their paleogeographic pattern, and responses to climate changes from the early Eocene to the early Oligocene in Asia. Front. Ecol. Evol. 10:955779. doi: 10.3389/fevo.2022.955779
Received: 29 May 2022; Accepted: 05 August 2022;
Published: 08 September 2022.
Edited by:
K. Christopher Beard, University of Kansas, United StatesReviewed by:
Pauline Coster, National Geologic Nature Reserve of Luberon, FranceWilma Wessels, Utrecht University, Netherlands
Copyright © 2022 Li, Li, Xu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qian Li, bGlxaWFuQGl2cHAuYWMuY24=