 Jason E. Rowntree1*
Jason E. Rowntree1* Paige L. Stanley2
Paige L. Stanley2 Isabella C. F. Maciel1
Isabella C. F. Maciel1 Mariko Thorbecke3
Mariko Thorbecke3 Steven T. Rosenzweig4
Steven T. Rosenzweig4 Dennis W. Hancock5Aidee Guzman2
Dennis W. Hancock5Aidee Guzman2 Matt R. Raven6
Matt R. Raven6- 1Department of Animal Science, Michigan State University, East Lansing, MI, United States
- 2Department of Environmental Science, Policy, and Management, University of California, Berkeley, Berkeley, CA, United States
- 3Quantis International, Boston, MA, United States
- 4General Mills Inc., Golden Valley, MN, United States
- 5Crop and Soil Sciences Department, University of Georgia, Athens, GA, United States
- 6Department of Community Sustainability, Michigan State University, East Lansing, MI, United States
Regenerative agriculture is a newly codified approach to agriculture that emphasizes reducing reliance on exogeneous inputs, as well as restoring and enhancing ecosystem services such as soil carbon (C) sequestration. These regenerative agriculture principles suggest that modern livestock systems can be redesigned to better capitalize on animals' ecological niche as biological up cyclers and may be necessary to fully regenerate some landscapes. One example is a multispecies pasture rotation (MSPR) system, which symbiotically stacks multiple animal production enterprises (i.e., chickens, cattle, sheep, and pigs) on one landscape. We conducted a whole-farm life cycle assessment (LCA) of an MSPR in the southeastern United States that was originally converted from degraded cropland. We compared the production outputs, greenhouse gas (GHG) emissions, land footprints, and soil health outcomes to a conventional, commodity (COM) production system of each respective species. Our 20-year MSPR chronosequence of soil C and other soil health indicators shows dramatic improvement since establishment, sequestering an average of 2.29 Mg C ha−1 yr−1. Incorporation of soil C sequestration into the LCA reduced net GHG emissions of the MSPR by 80%, resulting in a footprint 66% lower than COM. However, when comparing required land between the two systems for food production, MSPR required 2.5 times more land when compared to COM. Thus, while our model indicates that MSPR can simultaneously produce protein while regenerating land, a considerably greater land area is needed when compared to COM. Our results present an important yet paradoxical conclusion on land and food production balance. Should society prioritize an input-intensive, COM system that produces more food from a smaller yet degrading land base? Or, alternatively, should systems such as MSPR that produce less food on a larger, but more ecologically functional landscape be more highly prioritized? These complexities must be considered in the global debate of agricultural practice and land. Our results indicate MSPRs are a useful model for alternative livestock production systems with improved environmental outcomes, but in this study may present considerable land-use tradeoffs.
Introduction
Livestock are often considered agriculture's key greenhouse gas (GHG) emitter, contributing more than one-third of agricultural emissions (EPA, 2019). Typically, livestock production in the United States is highly specialized and intensified and is often cited as having both lower GHG [on a per carcass weight (CW) basis] and land-use footprints than pasture-based livestock systems. Alternatively, pasture-based systems often have less GHG intensity from a land use basis (Cardoso et al., 2016). However, current studies neither robustly consider complexity in diversified pasture-based livestock systems, nor consider the role of soil carbon (C) in GHG flux as well as land-use tradeoffs. This study aimed to contribute to this gap, in part, by quantifying GHG emissions, soil C sequestration, soil health, and land footprint of a farm using a diversified, multispecies pasture rotation (MSPR) in Clay County, Georgia, USA. We then compared emissions and land use to conventional, commodity (COM) production systems for beef, pork, and poultry.
Diversified farms supply 60 and 75% of the world's meat and dairy, respectively (Herrero et al., 2010; FAO, 2014). Expanding the use of diversified farming methods for animal production (including integrated crop-livestock systems, carefully managed grazing, and MSPRs) can lead to improved environmental outcomes and beneficial ecosystem services (e.g., wildlife and pollinator habitat, improved nutrient cycling) in addition to food production (Russelle et al., 2007; Kremen et al., 2012; Rivera-Ferre et al., 2016; Kremen and Merenlender, 2018; Kumar et al., 2019). Importantly, MSPRs take advantage of an “agromutualism” that builds symbiotic relationships between enterprises that lead to ecological and economic benefits. These production systems differ from industrial methods in focusing on biodiversity and mimicking natural ecological mechanisms (e.g., enhancing soil C sequestration through rotational grazing on rangelands and improving water and nutrient cycling through improved soil health), rather than specialization and intensification, albeit with considerably less overall production. However, few studies have explored such diversified livestock production systems in the United States, instead focusing mostly on very extensively (e.g., pastoralism) and intensively (e.g., feedlot) managed systems.
Livestock GHG footprints are calculated using life cycle assessment (LCA), which is an accounting approach that reports emissions resulting from all inputs and outputs of a production system on a per kg of CW of meat produced (kg CO2-e kg CW−1). LCA methodologies are often based on generally accepted Intergovernmental Panel on Climate Change (IPCC) calculations to estimate system GHG fluxes for processes such as enteric fermentation (enteric CH4), manure management, and feed production. These calculations rely on metadata accumulated over time and often from scientific literature. While these accounting principles and approaches are useful, supported by scientific literature, and give broad-based estimations on the impact of a system, they often do not account for the complexity of on-farm management and commonly trade-off regional specificity for global or national generalizations. Further, very complex diversified livestock systems are scientifically underrepresented in the literature compared to simplified animal production systems, and scientific studies of extensive systems often reduce complexity to regimented management practices designed to reduce the very complexity that farmers and ranchers face daily (Teague et al., 2013).
Recent studies show that livestock-induced soil C changes can have large impacts on the GHG balance of these production systems (Beauchemin et al., 2011; Teague et al., 2016; Stanley et al., 2018). Grazing lands are one of the most significant reservoirs of soil organic carbon (SOC) (Conant et al., 2017), containing more than 30% of total global SOC (Follett et al., 2000; Lal, 2002; Schuman et al., 2002; Derner and Schuman, 2007). Livestock are the primary users of this extensive land base and are an important management tool for mediating increased soil C sequestration (Liebig et al., 2010; Teague et al., 2011; McSherry and Ritchie, 2013; Machmuller et al., 2015; Wang et al., 2015; Griscom et al., 2017). Although our knowledge of management impacts on soil C sequestration is expanding, LCAs consistently omit it from GHG analysis (Rotz et al., 2019). Soil C has been historically excluded from LCA for a number of reasons, including lack of data on soil C sequestration, to provide conservative GHG estimates (Rotz et al., 2019), and an assumption that soils, without additional carbon inputs, are in long-term equilibrium. However, globally grass and cropland soils are highly degraded and thus have a long-term sequestration potential (Cotrufo et al., 2019; Yang et al., 2019; Lavallee et al., 2020). Some studies have shown that when including soil C changes to LCA parameters, the overall CO2-e can decrease considerably (Pelletier et al., 2010b; Stanley et al., 2018). Thus, changes in soil C could possibly be the greatest opportunity for reducing beef's carbon footprint.
In addition to GHG emissions and soil C sequestration, land use is a key evaluation metric of livestock systems. A growing global population and per-capita meat demand have increased the impetus for more efficient, and thus higher intensity, meat production. However, there are tradeoffs to extensive vs. intensive livestock production systems. For example, although overall land use is often lower in intensive systems, they often use a higher percentage of arable cropland suitable for other uses than extensive systems, which rely primarily on marginal lands. The MSPR examined in this study is an interesting case that is neither extensive nor intensive. Rather, it is a stacked-enterprise system in which animal stock density and rotational management are characteristically “intensive,” but taking place on an “extensive,” low-input, pasture-based landscape. We examined the total land-use tradeoffs for this system compared to conventional production systems for each animal species.
We hope to, in part, fill these gaps in the literature through this study in two ways: (1) by conducting a comparative analysis of an MSPR and a conventional US animal production system, thereby addressing the extensive–intensive dichotomy, and (2) using soil C sequestration and land-use trade-offs as additional comparative metrics in addition to GHG emissions.
Methods
Site Description
The USDA-certified organic farm, White Oak Pastures (WOP), is in Clay County, GA, and spans 1,214 ha of land. The prevailing soil types are Faceville, Marlboro, and Greenville fine sandy loam. Average annual rain is 1,342 mm yr−1, and mean high and low annual temperatures are 26 and 12°C, respectively (University of GA Environmental Monitoring Network 1957–2016).
Clay County, GA, was a historical scrubland/oak savanna, but agriculture has been and is currently the predominant land use (River Valley Regional Commission, 2014). Agriculture in the region most commonly employs a general crop rotation of cereal grains, corn, soybeans, cotton, and peanuts. Alternatively, WOP produces five red meat and five poultry species (including eggs)—totaling 142,935 animals annually—which are managed together on the same landscape. WOP acquires degraded croplands and converts them to MSPRs with a 3-year regeneration strategy. In years 1–3, cow–calf pairs are placed on the land at daily stock densities of 23–46 Mg ha−1 and fed hay throughout the winter (mean daily intake: 10 kg per animal). This supplies additional manure and organic matter (OM) from unconsumed hay to the soil, which is incorporated into the soil via animal impact. Bahiagrass (Paspalum notatum) is then aerial seeded and allowed to germinate. WOP is certified USDA Organic and thus does not apply chemical fertilizer or herbicides. However, residual chemicals from the transitioning degraded cropland pose a challenge to the farm. This transition process is illustrated in Figure 1.
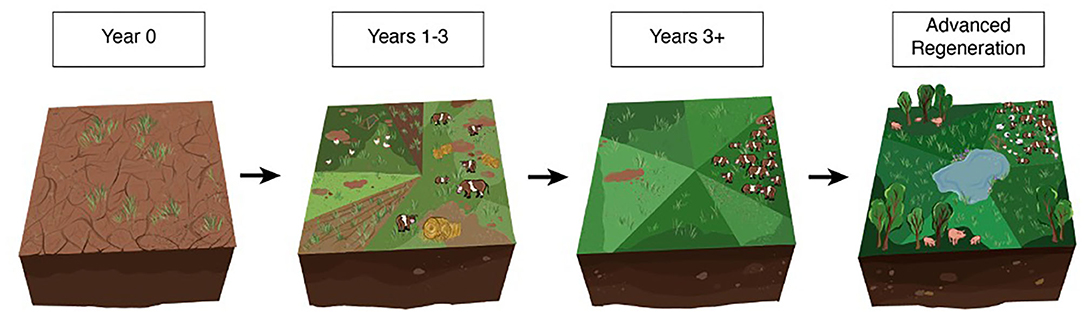
Figure 1. The regeneration process employed by White Oak Pastures. Year 0: Degraded cropland is acquired; Years 1–3: Hay is fed to cattle grouped in moderate densities, compost is applied, grass is seeded, and cattle and poultry are grazed at low stock densities; Years 3+: Animal stock densities are increased (25 to 50 Mg ha−1 daily), and holistic planned grazing (HPG) is implemented, where animals are rotated often and land is rested between grazing events; Advanced Regeneration: Represents a regenerative landscape (no seedings, added hay or compost since year 3) including rotations of diverse animal species with improved soil health and water cycling.
A combination of fertility practices is used to provide additional nutrients to the soil, including 1-cm compost application (produced and sourced on farm), and the addition of pastured layers or broiler chickens supplemented with feed. As conditions improve and forage quantity increases (years 4 and beyond), compost application is ceased, and cattle are then grazed using holistic planned grazing methodology (Savory and Butterfield, 2016). Holistic planned grazing (HPG) is a grazing process that entails high animal stock densities, division of the land into temporary small subunits (paddocks), and carefully planned herd movements that act in concert with forage availability and seasonality. Land managers use HPG with varying degrees of paddock “rest and recovery” periods to meet goals such as land improvement, increased livestock productivity, and maintenance of seasonal wildlife habitats. The manager at WOP uses livestock to defoliate plants at high stock densities (25–50 Mg ha−1 daily) and then quickly moves them off the grazed paddock daily to allow the grazed plants to enter full recovery. All beef cattle are in one single herd as opposed to the conventional practice of grouping animals by cow–calf, yearlings, and bulls. The final MSPR includes cattle, small ruminants (sheep and goats), poultry species (laying hens, guinea fowl, turkeys, ducks, and geese), swine, and rabbits, which are moved together in various herd combinations across the farm.
Clovers, forbs, and nut (primarily pecan) bearing trees are also introduced into the farm landscape to increase native plant diversity and to replicate historic oak-savanna silvopastoral conditions. These silvopastoral landscapes are also used for on-farm hog production, which is one of several other enterprises including USDA-certified organic produce, agritourism, and an on-farm restaurant.
Life Cycle Assessment
All emissions were calculated using a deterministic environmental impact model created in MS Excel with standard IPCC GHG inventory methodologies (IPCC, 2006). Face-to-face meetings, farm records, and a semistructured in-person interview with the farmer yielded model inputs and outputs. Questions included farm size and management practices (both spatial and temporal), number of animal units for each livestock category, exogenous input amounts and sources, production indicators, packing plant throughput, and quantification of animals not grown on-farm, but harvested at the on-farm USDA-inspected abattoir. Subsequent composting methods and application data were also collected. All major GHGs [methane (CH4), carbon dioxide (CO2), and nitrous oxide (N2O)] from direct and indirect sources were calculated using either Tier 1 (soil CH4 and N2O) or 2 (enteric CH4, Ym = 6.0) (IPCC, 2006) methodologies. Other emissions including feed production and transport, on-farm and abattoir energy use, and compost production were calculated (EPA, 2020). Emissions from energy used for equipment manufacture were excluded based on their minor contribution (< ~3%) (Lupo et al., 2013). All gasses were converted to CO2 equivalents (CO2-e) using current 100-year global warming potentials (CO2 = 1, CH4 = 30.5, N2O = 265). We defined the functional unit for this model as kg of CO2-e per kg of meat on a CW basis (kg CO2-e kg CW−1).
Soil Sampling and Analyses
To estimate soil C sequestration rate and changes in other soil health indicators, soils were sampled along a 20-year degraded cropland to MSPR chronosequence. The chronosequence consisted of a currently cultivated cropland (year 0) and fields converted from cropland to pasture 1, 3, 5, 8, 13, and 20 years ago. Year 0 represents land that has been continuously farmed for a minimum of a decade with rotations of corn (Zea mays), peanuts (Arachis hypogaea), wheat (Triticum), and soybeans (Glycine max). The land was routinely tilled, and chemical fertilizer and herbicides were applied annually. Initial land transformation began in year 1 when off-farm hay was applied across the degraded land and then fed to cattle grouped in relatively high stock densities (25–50 Mg ha−1 daily). This helps to both break up capped soil and more evenly disperse nutrients back into the soil from manure, urine, and residual hay. The following spring, grass was aerial seeded onto the land. In years 1–3, these fields are minimally grazed and receive 1 cm of compost ha−1 yr−1. After year 3, exogenous inputs (hay and compost) were ceased, and the regeneration strategy shifted toward an animal-only approach, whereby animals were the primary mechanism of improving the land. This was done by increasing grazing exposure, introducing multiple livestock species including pastured poultry into the MSPR, and continually rotating animals across the land using HPG. Year 20 represents a grassland site that did not receive compost or poultry impact, only planned beef cattle grazing.
In the spring of 2018, soil samples were collected from each field. Our objective was to find a site that had no animal impact for the year 0 chronosequence site. However, this location had received one instance of animal impact via hay feeding at the time of sampling. Therefore, we chose to resample at a newly acquired location that had received no animal impact and was more indicative of a true year 0. We then chose to use data from the newly acquired site as year 0, and the data from the originally sampled site as year 1. We set out to collect a minimum of four soil cores at intervals spaced 10 m apart along set transects. However, because of dry conditions, we were able to collect only one intact soil core from the year 0 site. Although there was very little difference in soil C stock between year 0 and year 1, we elected to include this in the model as a true year 0 site. We also experienced dry, difficult sampling conditions in year 13, enabling collection of two intact soil cores.
Each field was sampled within the dominant soil type according to Web Soil Survey, which was either a Faceville, Marlboro, or Greenville sandy loam in each location. At each sampling location, four 1-m soil cores were sampled (although soil conditions prevented all four samples at the 50- to 100-cm depth from being collected at some sites) using a 5.7-cm diameter Giddings probe (Windsor, CO) for soil C analysis, and eight 10-cm soil cores were collected using a 3.2-cm-diameter hand probe for soil health analysis. Meter-deep intact soil cores were separated into 0- to 10-, 10- to 30-, 30- to 50-, and 50- to 100-cm depths and sieved to 8 mm. Samples from each location were analyzed by depth for bulk density (20-g subsamples were weighed, dried at 105°C, and reweighed to determine the mass of dry soil per unit volume) and soil C [soils were ground on a ball mill and analyzed using a CN analyzer (LECO CHN-2000 autoanalyzer)], and later averaged to obtain field-level means. We used the minimum equivalent mass (Lee et al., 2009) to convert C concentrations to C stocks (Mg C ha−1).
Hand cores (10-cm depth) were placed on ice the evening of collection and delivered overnight to Cornell University. Samples were analyzed by sampling location for the Comprehensive Assessment of Soil Health, which is a suite of soil tests including texture by hydrometer, pH, wet aggregate stability, permanganate oxidizable (active) C (POXC), microbial respiration via 4-day incubation, autoclave citrate-extractable (ACE) soil protein, and available water-holding capacity (AWC); (Moebius-Clune et al., 2016). Soil health analyses were not performed on the year 0 site.
Soil clay contents ranged from 5 to 20%. Least-squares means of equivalent mass carbon stocks, wet aggregate stability, active C, ACE soil protein, and microbial respiration were calculated to account for clay content as a covariate where clay was significant (α = 0.05). Clay was not a significant covariate for water-holding capacity. Soil C sequestration rate was calculated using linear regression on least-squares means of carbon stocks. All statistical analyses were completed using RStudio Team 2019 with the package lsmeans (Boston, MA).
Comparison to COM Animal Production
To understand the relative emissions and land use of the MSPR examined in this analysis, we compared beef, pork, and poultry results of this LCA to COM agricultural production of beef (Rotz et al., 2019), pork, and poultry (Gerber et al., 2013).
We retrospectively determined land needed to grow feed (for pork, poultry, and feedlot beef) or graze and grass-finish beef cattle based on the CW output of the WOP MSPR and the Georgia crop and hay production averages (USDA NASS, 2018). For the non-ruminant diets, we used an 80% corn, 20% soybean meal diet per COM standard production. Importantly, pork and poultry finishing diets are more variable than our standard ration and can include dried distiller's grains and synthetic amino acids among other feedstuffs. Because of the difficulty of accounting for these differences across a large geographical context, we chose a standard baseline for diet comparison.
For the beef cattle land comparison, we first used the number of cow–calf pairs necessary to produce the annual beef output (268,777 kg yr−1) at WOP for 1 year (n = 992). Stocking rate for the system was calculated based on existing Georgia recommendations (0.81 ha per cow; D. Hancock, personal communication, 2019). Total land needed for grazing and hay was calculated at 0.66 ha per grass-finished steer in the MSPR. Because beef grown in feedlots are considerably heavier and require less land for feed, we used beef CWs and land use data from Stanley et al. (2018) to adjust cows and land needed for feed production. We also calculated the additional hay needed for supplementation in the COM system using the Stanley et al. (2018) feedlot diets and then divided by the mean hay production per acre in Georgia (USDA NASS, 2018).
Results
Meat Production and Emissions
Overall animal productivity and GHG emissions of the MSPR system are reported in Tables 1, 2. Beef, poultry, and swine comprise 96% of the overall production on a CW basis. Each year, the MSPR at WOP (including all animals) harvests 143,372 animals, totaling 637,910 kg of total CW. Summing all animals in the MSPR, the farm produces 525 kg CW ha−1. Thus, the overall productivity of the total MSPR is substantially higher when compared to grass-finished beef only (221 kg CW ha−1).
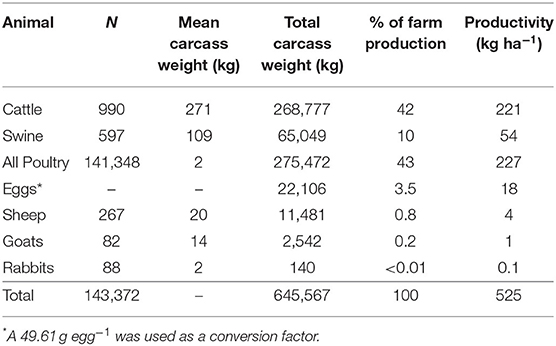
Table 1. Overall market animal production and carcass output.
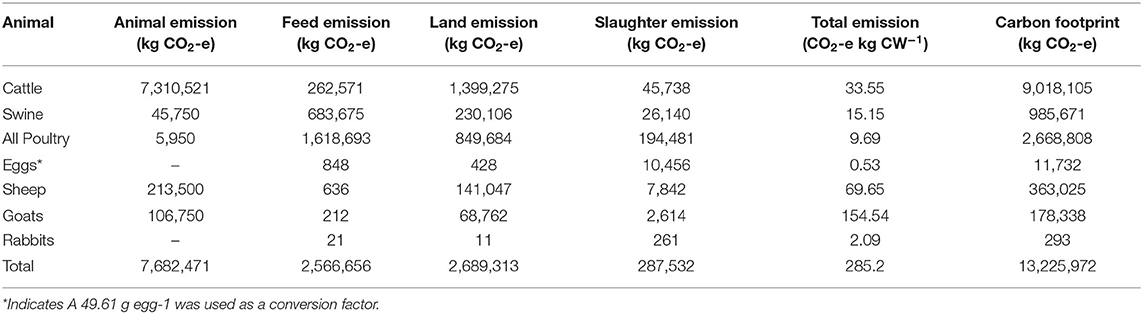
Table 2. Overall farm emissions by animal species.
While beef cattle comprise 42% of overall CW production, their emission on a CO2-e kg CW basis is higher than in other systems. Cattle contribute 33.55 kg CO2-e kg CW−1, whereas swine and all poultry contribute 15.15 and 9.69 kg CO2-e kg CW−1, respectively. The beef cattle contribute 68% of total farm emissions, totaling 9,018,105 kg CO2-e. Poultry was the second greatest contributor to overall emissions at 20%, while contributing 43% to the overall farm production. Emissions from swine production align evenly with productivity, totaling 7% of the farm GHG footprint and 10% of farm production. Eggs and all other species, primarily sheep and goats, contribute <1% of the overall farm GHG footprint.
Total farm emissions categorized by animal production, feed, land, and slaughter vary by species. Beef cattle account for about 95% of animal and 52% of land emissions. Poultry production, the second largest contributor to on-farm productivity, is responsible for 63% of total feed emissions and 68% of total slaughter emissions. The MSPR total carbon footprint was 13,225,972 kg CO2-e, with animals as the greatest emissions category (58%), followed by land (20%) and feed (19%).
Soil Parameters
We observed substantial increases across a suite of soil health indicators over the 20-year chronosequence (Table 3). Wet aggregate stability increased from 0 to 53% over the chronosequence, with a 5-fold increase between years 3 and 20 (p = 0.02). Microbial respiration increased from 0 to 0.56 mg CO2 day−1 by year 3 and 1.16 mg CO2 day−1 by year 20 (p = 0.03), whereas POXC increased 10-fold across the chronosequence (p < 0.01). ACE protein, which estimates the amount of mineralizable organic N, increased from 0 to 23 mg g−1 over the chronosequence, with a 4-fold increase from year 3 to year 20 (p < 0.01). There was no observable increase in AWC.
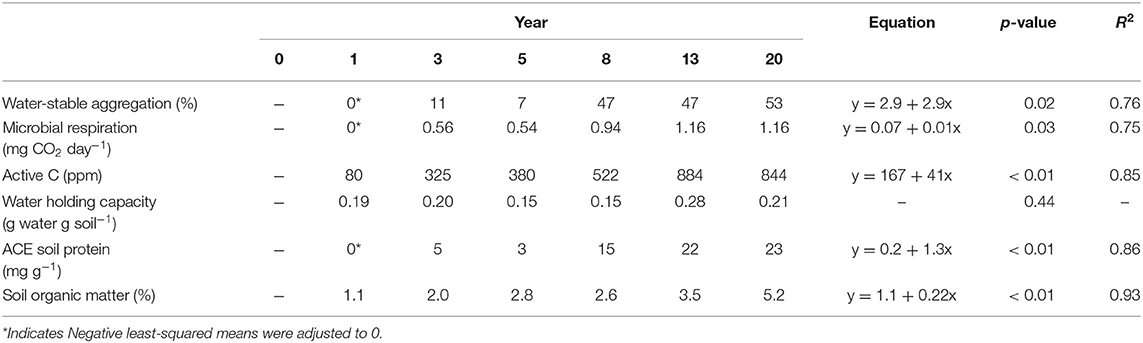
Table 3. Soil indicators.
Soil Carbon Sequestration
In addition to soil health indicators, we also measured SOC stock from year 0, prior to MSPR initiation, to year 20. Initially, SOC stocks were ~10 Mg C ha−1 and increased to 50 Mg C ha−1 in year 20, a 5-fold increase across 20 years of management. The highest measured soil C stock was in year 13, measuring 65 Mg C ha−1. Importantly, the year 20 site received no compost applications or poultry disturbance and reflected only the impact of grazing and perennial conversion from annual cropland. Soil carbon stocks at equivalent minimum mass increased linearly at a rate of 2.29 Mg C ha−1 yr−1 (p = 0.04, R2 = 0.60; Figure 2). Field-level standard errors for each soil depth is given in Supplemental Info (Supplementary Table 1). Soil OM (SOM; Table 3) concentration reflected comparable increases at the surface from 1 to 5% in years 0 and 20, respectively. Overall, the transition from a conventional row crop model to MSPR improved soil physical and biological attributes and consequently significantly improved soil C stocks.
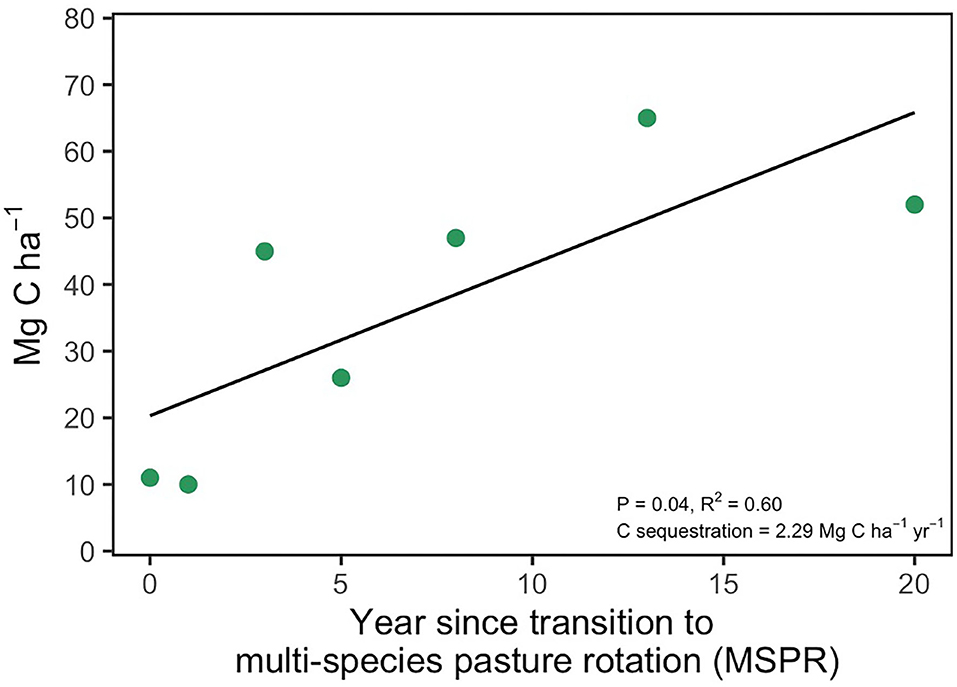
Figure 2. Soil carbon stock at equivalent soil mass of 9,900 Mg/ha. Points represent least-squared means adjusted for soil clay content generated from 4 in-field replicate soil samples.
The overall MSPR beef footprint totaled 33.55 kg CO2-e kg CW−1 and was 36.5% greater compared to the COM beef GHG footprint (21.3 kg CO2-e kg CW−1). The greatest emission disparity between production methods was observed in pork, where MSPR pork was 3-fold greater compared to a COM production footprint (15.15 vs. 4.6 kg CO2-e kg CW−1 for MSPR and COM pork, respectively). The MSPR poultry was over twice that of COM poultry but in each production system represented the least emission intensity of all species analyzed in the model (Figure 3).
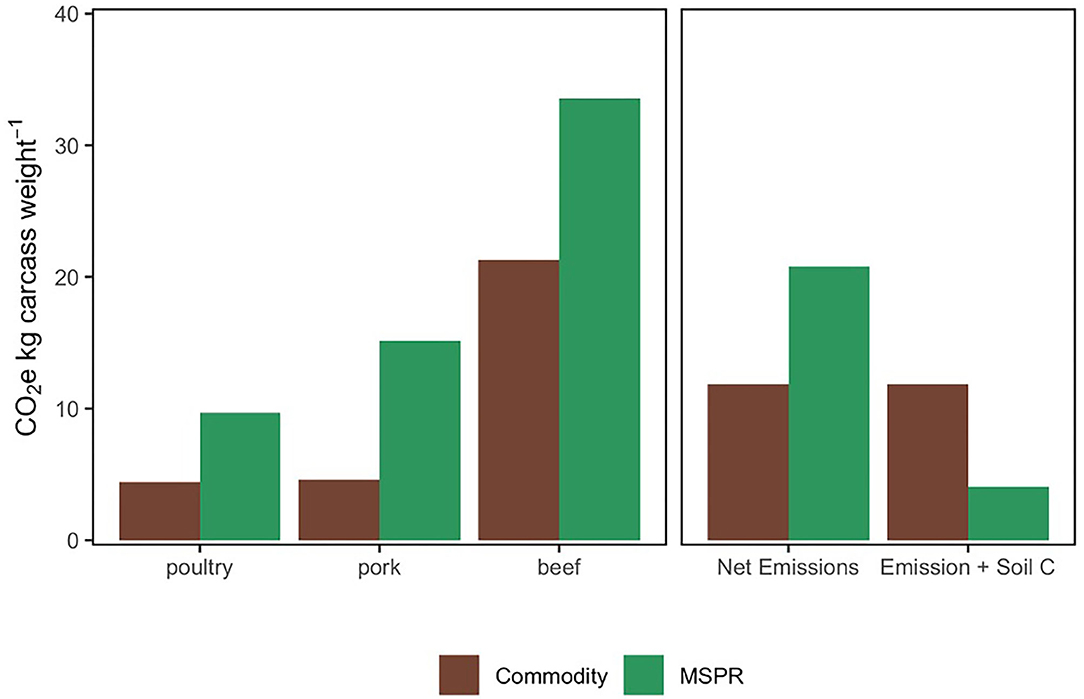
Figure 3. Comparison of a Commodity and Multi-Species Pasture Rotation (MSPR) CO2-e on a kg CW basis by specie (left) and then aggregated for the mean overall net farm emission with and without soil C sequestration (right).
We next totaled all emissions in each species production category and present the overall net emission for the MSPR as compared to COM. The overall MSPR carbon footprint for poultry, pork, and beef produced on farm totaled 20.8 kg CO2-e kg CW−1, 44% greater than COM, which totaled 11.9 kg CO2-e kg CW−1 for all livestock species produced.
We integrated measured soil C sequestration (Figure 2) into the net emissions from MSPR and COM. We used mean soil C sequestration of 2.29 Mg C ha−1 yr−1 for MSPR and considered COM to be in a soil C dynamic equilibrium. Incorporation of soil C sequestration as a GHG sink in the MSPR system reduced emissions from 20.8 to 4.1 kg CO2-e kg CW−1 representing an ~5-fold drop in emission intensity. The resulting 4.1 kg CO2-e kg CW−1 of net MSPR emissions then become 7.8 kg CO2-e kg CW−1 lower than COM. These results point to the dramatic changes that can occur in animal protein LCA when accounting for changes in soil C stocks over time. Importantly, if we were to attribute the soil C sequestration across the chronosequence to only cattle, MSPR beef produced in this system would be a net sink of −4.4 kg CO2-e kg CW−1 annually.
Finally, in Figure 4, we calculated the land required to produce all proteins in the COM and MSPR models. The required land to graze beef and supply feed for each species (poultry, pork, and beef) is considerably greater for the MSPR system than COM. The MSPR required 2.5 times more land when compared to COM to produce the same amount of CW. Thus, while our model indicates that MSPR can simultaneously produce protein while increasing soil health indicators and soil C stock, a considerably greater land area is needed when compared to COM.
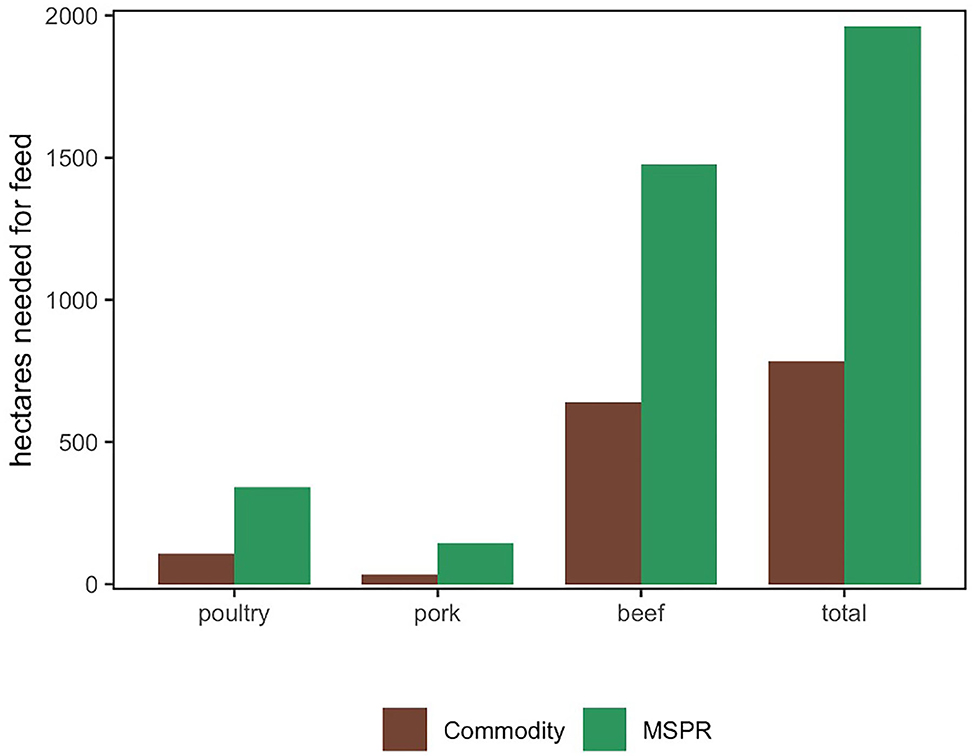
Figure 4. Comparison of a Commodity and Multi-Species Pasture Rotation (MSPR) for land needed to graze beef and supply feed for poultry (275,242 kg), pork (65,049 kg) and beef (268,777 kg) similar to outputs of monitored MSPR farm.
Discussion
Meat Production and Emissions
We report animal production and resulting emission metrics of an MSPR production system whose owner's primary goal is to farm regeneratively. Gosnell et al. (2019) define regenerative agriculture as an “alternative” form of food and fiber production oriented toward enhancing resilience and ecological health.
With respect to on-farm production, 42% of the overall farm CW was produced from cattle, where the mean grass-finished beef CW was 271 kg hd−1. Most beef LCAs measure productivity on an animal performance basis vs. actually indicating a CW (Pelletier et al., 2010b; Lupo et al., 2013) or determine the amount of animals necessary to produce a certain amount of beef (Capper, 2012). However, we were only able to collect CWs from the packing facility, making comparisons back to a live weight productivity difficult. However, our reported MSPR finished beef CW closely aligns with Stanley et al. (2018), who reported a 280 kg hd−1 in an adaptive multipaddock (AMP) grazing system—a similar management strategy, but using beef cattle only. Importantly, these results show that grass-finished CWs are ~33% lower than existing grain-finishing beef LCAs (Pelletier et al., 2010b; Lupo et al., 2013; Stanley et al., 2018).
The interview with the owner of WOP indicated that the average age of slaughtered beef cattle was between 20 and 22 months of age (data not shown). This is considerably less than a recent study by Heflin et al. (2019), who modeled a grass finishing system in the lower Southern Plains and indicated a time to slaughter of 30 months with an average CW 40 kg greater than our MSPR system. However, that the WOP MSPR system reached similar CWs 10 months sooner, comparatively, than other grass-finished beef systems is an important improvement that both reduced GHG emissions over a shorter lifetime while simultaneously producing other food animal proteins.
Life cycle emissions for beef cattle in the MSPR were 33.55 kg CO2-e kg CW−1. This is 30% higher than the most current models evaluating business-as-usual beef cattle production systems (with grain finishing) in the United States (Rotz et al., 2019). This is due to the widely accepted fact that grass-finished cattle have a higher enteric CH4 footprint than those finished on grain because of differences in feed digestibility. In our study, 81% of beef cattle footprint is attributed to enteric CH4. However, the proportional trade-offs of specific GHGs in each production system are also important to consider. For example, while enteric CH4 in the MSPR was proportionately high, CH4 is a short-lived climate pollutant where C is contained in existing biomass and cycled quickly through the atmosphere, lasting on average 10 years before being oxidized (Lynch et al., 2020; Thompson, 2020). Alternatively, although overall emissions in grain-finished beef systems are lower, the portion of fossil-fuel derived emissions is higher, including CO2 and N2O (lasting 1,000 and 100 years on average, respectively) arising from fertilizer production and application for fodder crops and fossil fuel–derived energy use (Picasso et al., 2014). Pierrehumbert and Eshel (2015) also report less overall climate impact of pastured-beef systems with no or minimal fertilization, despite greater enteric CH4 emissions compared to feedlot systems. Recent IPCC estimates show that global CO2 and N2O concentrations have been rising more rapidly than CH4, which has been plateauing (IPCC, 2014). The shorter life span of CH4 in the atmosphere, however, also makes it an attractive target for near-term GHG mitigation.
The beef cattle in the MSPR represent the largest emission source in the production system for three reasons: they make up the largest group of animals in the system, they produce high CWs, and they contribute more CO2-e per kg of CW compared with other livestock categories. Studies have demonstrated that intensive feeding and management of beef cattle in grain-fed system result in higher CW and lowest overall CO2-e kg−1 emission (Heflin et al., 2019; Kamilaris et al., 2020). However, they generally lack a systems perspective to net GHG fluxes, thereby omitting soil carbon sequestration, which has great potential to mitigate GHG emissions for grass-fed systems (Liebig et al., 2010; Pelletier et al., 2010a). Inclusion of field-measured soil C sequestration (as a CO2-e sink) has been shown to completely mitigate the C footprint of intensively managed grass-finished cattle in some specific cases (Stanley et al., 2018) and drastically lower (but not neutralize it) in others (Machmuller et al., 2015; Wang et al., 2015; Hillenbrand et al., 2019). Although few exist, these cases present a unique nexus that (a) alleviates the pressure to use input and fossil-fuel intensive production systems to maximize cattle gains and lower per-kg CO2-e and (b) maximizes biological ecosystem functions to reduce net GHG emissions while maintaining productivity.
Less dichotomy exists in comparing the MSPR pork and poultry outputs to generally accepted COM pork and poultry outputs. More difficulty arises when comparing pastured poultry models in the literature. For pastured broiler only models, recommendations range from 500 to as much as 1,000 broilers 0.40 ha−1, with almost all nutrients coming from cropland derived feed instead of pasture (Meeh et al., 2014). At WOP, 445,182 eggs were produced using the MSPR, weighing an average of 49.6 g (data not shown) totaling 22,106 kg of eggs.
Although poultry production, including eggs, represents 46.5% of the total carcass weight in this system, they contribute only 20% of total carbon footprint. Feed production for poultry was the largest impact category (Tongpool et al., 2012; MacLeod et al., 2013), mainly comprising energy- and protein-rich ingredients (more than 60%). In our system, emissions from feed totaled 60.6% of the total poultry carbon footprint. For eggs, however, emissions associated with slaughter (processing and transport) outpaced those from feed production, contributing 89 and 7%, respectively. Poultry-meat produces a greater emissions footprint than eggs partly because rations for broiler chickens, on average, include a higher share of soybean products, which are sourced from areas where land-use change is taking place (MacLeod et al., 2013).
Feed production was proportionally the greatest emission source for both poultry and swine, whose diets consisted of primarily corn (80%) and soybean (20%) products. These results are generally consistent across the literature, although COM swine production systems often have larger GHG footprints associated with facilities (Pelletier et al., 2010a; Eshel et al., 2014; Kebreab et al., 2016; Tallaksen et al., 2020).
Soil Parameters
We observed large increases across the suite of soil health indicators examined in the MSPR chronosequence, indicating improvements in soil function as a result of perennial establishment and regenerative MSPR management at WOP. SOM is related to nearly every soil-related ecosystem service including water and nutrient cycling, habitat for biodiversity, and erosion control (Wall et al., 2012). Observed increases in SOM were likely mediated by greater aggregation, as aggregation is one of the primary mechanisms of SOM stabilization via physical protection and microbial habitat (Tisdall and Oades, 1982; Gupta and Germida, 2015), and we observed a 5-fold increase in both SOM and wet aggregate stability over the chronosequence. Additionally, higher wet aggregate stability indicates an improved ability to maintain soil structural integrity in the face of events such as extreme precipitation, leading to greater water infiltration and reduced erosion (Franzluebbers, 2002). We expected these increases in aggregation and SOM to translate to greater AWC. However, we did not observe an increase in AWC across the chronosequence, further adding to literature suggesting the link between SOM and AWC is not as pronounced as previously thought (Minasny and Mcbratney, 2017).
Rapid responses in microbial respiration (2-fold increase), ACE protein (5-fold increase), and active C (10-fold increase) during the chronosequence indicate the enhancement of soil C and N cycling with MSPR. Microbial activity in annual cropland soils is often limited by C availability (Schimel, 1986), and the increase in active C and microbial respiration observed within the first several years of the chronosequence reflects the alleviation of C limitation via greater C inputs in the perennial MSPR system. Additionally, increased ACE protein reflects a growing pool of readily mineralizable organic N as a result of greater plant inputs, animal manures, and additions of other organic materials such as compost. Large reservoirs of organic N coupled with an active microbial community are critical for efficiently meeting plant N needs in agroecosystems, as organic N released slowly through mineralization is more efficiently utilized than pulse additions of inorganic N via synthetic fertilizer (Gardner and Drinkwater, 2009). Together, the improvements across the suite of soil health properties measured here indicate that the building blocks for a growing microbial community in soils under MSPR management were met, ultimately contributing to the increased soil C pool and more efficient N cycling.
Soil Carbon Sequestration
Over the 20-year chronosequence, the MSPR system at WOP sequestered an average of 2.29 Mg C ha−1 yr−1 (Figure 2). However, the oldest location in the chronosequence received grazing only rather than all management practices applied to younger sites (e.g., compost). Thus, the average C sequestration rate may have been higher if this site were more representative of the entire chronosequence.
Compared to other literature, our estimated soil C sequestration rate lies toward the higher end—both above (Wang et al., 2015; Conant et al., 2017) and below (Stanley et al., 2018) reported values from others. It is important to note that each system is unique and that resulting soil C sequestration with the application of a management system like the MSPR employed by WOP will differ based on land use history (e.g., conversion from cropland or overgrazed pasture), time since adoption (discussed more in detail below), and changing weather conditions (e.g., drought) among other factors. For example, Stanley et al. (2018) reported average SOC sequestration after five-year conversion from continuous haying and grazing to AMP grazing (analogous to the HPG system used at WOP, but with cattle grazing only). Our system reflects a longer transition, over 20 years, which may explain the lower average sequestration rate, comparatively. Alternatively, the MSPR system in this study was employed after conversion from degraded cropland, was combined with compost application, and was conducted in a non-arid ecosystem. These practices explain the higher relative sequestration rates compared to some others (Wang et al., 2015; Conant et al., 2017).
In this study, SOC sequestration is estimated via a space-for-time substitution rather than directly measuring SOC change over time. While it has limitations, without baseline SOC data for each field, the chronosequence approach is the best alternative for measuring temporal SOC changes with differing management when compared within soil types and has been used widely throughout the ecological sciences (Walker et al., 2010).
Our estimated SOC sequestration rate (2.29 Mg C ha−1 yr−1) is an average over 20 years. To better assess temporal dynamics of SOC sequestration, we can also analyze how the sequestration rate changes over time. It is commonly assumed that there is a finite capacity of soils to store C and that sequestration rates will slow over time as soils come closer to a “saturation” point. Our results indicate a sharp increase in SOC stocks from years 1 to 3, with slower increases from years 5 to 13. Soil C stock at the oldest MSPR site (at 20 years) indicated a slightly lower soil C stock than the 13-year site, which may suggest a peak soil C accumulation at ~13 years since establishment of the MSPR. However, we do not believe this is indicative of a declining sequestration rate due to proximity to saturation. Rather, we posit that this is an artifact of management differences between the sites, as the 20-year site received grazing only, rather than the entire suite of management interventions (i.e., compost, poultry manure) that were applied to all other MSPR sites across the farm.
Further, carbon stock alone does not allow us to make conclusions about soil C storage capacity, which can be better informed by the relative distribution of soil C between mineral-associated OM (MAOM) and particulate OM (POM) (Cotrufo et al., 2019). These authors also showed that soil C in grasslands is contained mostly in the MAOM fraction, which is often microbially processed and high in N, making it highly persistent and stable in soils (Lavallee et al., 2020). MAOM also saturates in soils because of the finite availability of mineral surfaces to sorb OM. However, the authors also suggest that POM can be indefinitely accrued in soils irrespective of MAOM saturation and, further, that most grassland soils are unlikely to be “saturated” with respect to MAOM-C. We did not fractionate SOM into MAOM and POM pools in this analysis; however, given the large increases in soil aggregation, it is likely that POM is increasing with MSPR adoption in this system, because POM persistence is largely dependent on aggregation. Further, results presented by Cotrufo et al. (2019) and others (West and Six, 2007; Jagadamma et al., 2014; Nicoloso et al., 2018) lead us to question the certainty of soil C saturation in grassland soils.
The results of other soil parameters in addition to the SOC sequestration in this study allow us to infer management drivers as well as functional changes in the soil. In general, soil C stock can be increased by (a) increasing C inputs to the soil or (b) reducing the relative rate of loss (as CO2) via decomposition or stabilization, which reduces emissions to the atmosphere that would otherwise occur (Conant et al., 2017). In our MSPR system, C inputs were increased in three ways, by the increase of native and perennial plant diversity under MSPR as clovers, forbs, and nut bearing trees; by the addition of compost and manure from livestock; and via exogeneous poultry feed. Further, the short-duration, high-intensity grazing (otherwise termed HPG; as well as rotation with other animals) used in this system has been shown to maximize plant residue left in the pasture and improve below-ground soil C allocation via plant roots (Teague et al., 2011).
Soil C sequestration is a vital ecosystem function to mitigate climate change. Here, we demonstrate that land restoration using MSPR is an important regenerative agricultural tool to support this effort.
Net GHG Footprint and Land Use
While the GHG footprint of the MSPR is considerably greater than corresponding COM estimates (Figure 3), the reverse is true after incorporation of the on-farm soil C sequestration as a GHG sink. When considering only standard LCA boundaries on a kg CO2-e kg CW−1, animals in the COM system are more efficient—gaining more weight in less time and thus contributing 43% fewer GHGs (MSPR: 20.8 kg CO2-e kg CW−1 vs. COM: 11.9 kg CO2-e kg CW−1). However, our on-farm analysis of soil C accrual at WOP revealed a sequestration rate of 2.29 Mg C ha−1 yr−1, on average, over 20 years of MSPR adoption. After incorporating this into our LCA boundaries, this reduced the GHG footprint of the MSPR system by 80% (from 20.8 to 4.1 kg CO2-e kg CW−1), ultimately finishing at 66% lower than comparative COM production.
However, when comparing required land between the two food production systems, MSPR required 2.5 times more land than COM production. Thus, while our model indicates that MSPR can simultaneously produce protein while regenerating land and can contribute other ecosystem services, a considerably greater land area is needed when compared to COM. However, MSPR is well-suited for more marginal lands while requiring fewer exogenous inputs such as feed stocks. Consequently, increased implementation of MSPR on marginal lands, including degraded cropland, could free up more productive land for production of higher value and more nutrient dense crops. Theoretically, this trade-off in land use could also, to an extent, partially mitigate the greater land area needed for MSPR livestock production vs. COM.
Our results present an important yet paradoxical conclusion on land and food production balance in the face of climate change. Should society prioritize an input-intensive, COM system that produces more food from a smaller, yet degrading land base with externalized societal costs? Or, alternatively, should systems such as MSPR that produce less food on a larger, but more ecologically functional and diverse landscape be more highly valued? These complexities must be considered in the global debate of agricultural practice and land, as land-management strategies that prioritize soil health to regenerate agroecosystems are increasingly needed to meet the needs of a growing population.
Regardless of the starting point on any farm or ranch, we hope to emphasize the importance of diversifying as a process to provide and enhance ecosystem services that are becoming increasingly important in addition to food production, such as resiliency and adaptive capacity to extreme weather, nutrient cycling, water retention, and climate change mitigation. Teague et al. (2016) provides a cadre of tools to improve ecosystem services in both cropping and grazing systems, which can be implemented by farms and ranches of all production types. For example, reducing and eliminating tillage, maintaining soil cover with cover crops, increasing biodiversity and nutrient cycling via integrated crop-livestock systems, and maximizing rest periods in grazing-only systems are all tangible actions for regenerating agroecosystems (Brewer and Gaudin, 2020). The WOP MSPR examined in this study exemplifies a farm using a highly evolved production system at the far end of the diversification spectrum.
Lastly, although we highlight the need for more research on diversified livestock production systems, the benefits of diversified agroecological production systems for the provisioning of ecosystem services are well-established. The results of this research point us to other important and timely questions of farmer practice adoption, payment for ecosystem services (PES), and other incentivization mechanisms (Gosnell et al., 2020). Currently, underdeveloped PES and carbon markets present major challenges to the adoption of regenerative agricultural practices in the United States. As it becomes increasingly clear that deployment of carbon capture and storage (CCS) technologies (potentially via PES), in addition to GHG mitigation measures, will be necessary to meeting our climate goals, regenerative agriculture is arising as a practice with clear CCS potential. Thus, we recommend that federal monetization strategies be developed to increase adoption of regenerative agricultural practices simultaneously to ongoing research, rather than sequentially.
Conclusions and Implications
Diversified livestock production systems are highly underrepresented in scientific literature, despite evidence of widespread global use (Robinson et al., 2011). We present, to our knowledge, the most robust analysis of an MSPR system in the scientific literature comprising beef, pork, and poultry. In addition to business-as-usual LCA methodology, we also incorporated measured on-farm soil health parameters, including soil C sequestration. Most often, animal production LCAs are generated for one species of livestock and likewise are analyzed with broad-based formulas generated from empirical models across large geographical contexts. Our study provides unique model parameters for an actual farm in the United States, populated with on-farm generated vs. literature derived production metrics with actual soil C and subsequent soil health data across time and space.
This study provides interesting new context to current agricultural debates, including those surrounding land-sharing vs. land-sparing, sustainable intensification, and the use of regenerative agriculture to sequester soil C. WOP is a USDA Organic MSPR employing principles of regenerative agriculture through holistic management. As defined by Gosnell et al. (2019), regenerative agriculture “focuses on enhancing and restoring holistic, regenerative, resilient systems supported by functional ecosystem processes and healthy, organic soils capable of producing a full suite of ecosystem services, among them soil carbon sequestration and improved soil water retention.” Our results indicate that this system does, in fact, regenerate ecological function including soil health, resilience, GHG mitigation, and biodiversity. It accomplishes this by managing animals intensively (not to be confused with input-intensity) in an otherwise extensive system (no chemical fertilizers, biocides, tillage, etc.). When comparing this approach to a business-as-usual COM-based approach, and including soil C sequestration, the overall emission footprint of the regenerative agriculture approach was 3-fold less. Adoption of practices such as the MSPR investigated in this study should be incentivized at a greater scale while concomitantly investigating technologies and approaches that can reduce the necessary land needed to produce the regenerative proteins.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
JR led overall project management, data analysis, and writing. PS contributed in project conception, data management, and writing. IM oversaw dataset management for production and emissions of all livestock in the White Oak Pastures production system and also assisted with writing. MT oversaw life cycle assessment model development, and assisted with writing. SR oversaw all soil data collection, laboratory analysis, and statistical analysis of data and assisted with writing. DH assisted with chronsequence development, provided overall guidance on the farm site, and assisted with writing. AG developed and created the figures. MR aided in project conception and assisted with writing. All authors contributed to the article and approved the submitted version.
Funding
General Mills Inc funded the project in entirety.
Conflict of Interest
The authors declare receiving funding from General Mills Inc. The funders had the following involvement in the study: funding for the project design, soil and data collection, soil analysis and overall life cycle assessment.
This project began under the auspices of collecting data solely for the internal use of General Mills Inc. After data collection and analysis, General Mills Inc. gave the authors permission to publish the project results in manuscript form.
SR was an employee of General Mills Inc. MT was an employee of Quantis International at the time of study completion.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank the WOP Farm: Will Harris, Jenni Harris and Jodi Harris Benoit and the all staff who provided information for this study. Secondly Jeff Hanratty and Jerry Lynch with General Mills Inc. were instrumental in funding this project. Equally supportive, Jon Dettling from Quantis provided important input and guidance for the project. Finally, Taylor Collins and Katie Forrest from Epic Provisions were instrumental in advocating and supporting this project.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2020.544984/full#supplementary-material
References
Beauchemin, K. A., Janzen, H. H., Little, S. M., McAllister, T. A., and McGinn, S. M. (2011). Mitigation of greenhouse gas emissions from beef production in western Canada–evaluation using farm-based life cycle assessment. Anim. Feed Sci. Technol. 166–167, 663–677. doi: 10.1016/j.anifeedsci.2011.04.047
Brewer, K. M., and Gaudin, A. C. (2020). Potential of crop-livestock integration to enhance carbon sequestration and agroecosystem functioning in semi-arid croplands. Soil Biol. Biochem. 149:107936. doi: 10.1016/j.soilbio.2020.107936
Capper, J. L. (2012). Is the grass always greener? comparing the environmental impact of conventional, natural and grass-fed beef production systems. Animals 2, 127–143. doi: 10.3390/ani2020127
Cardoso, A. S., Berndt, A., Leytem, A., Alves, B. J. R., de Carvalho, I., das, N. O., et al. (2016). Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use. Agric. Syst. 143, 86–96. doi: 10.1016/j.agsy.2015.12.007
Conant, R. T., Cerri, C. E. P., Osborne, B. B., and Paustian, K. (2017). Grassland management impacts on soil carbon stocks: a new synthesis. Ecol. Appl. 27, 662–668. doi: 10.1002/eap.1473
Cotrufo, M. F., Ranalli, M. G., Haddix, M. L., Six, J., and Lugato, E. (2019). Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 12, 989–994. doi: 10.1038/s41561-019-0484-6
Derner, J. D., and Schuman, G. E. (2007). Carbon sequestration and rangelands: a synthesis of land management and precipitation effects. J. Soil Water Conserv. 62, 77–85. Available online at: https://www.jswconline.org/content/62/2/77
EPA (2019). Inventory of U.S. Greenhouse Gas Emissions and Sinks 1990-2017. U.S. Environmental Protection Agency.
EPA (2020). Greenhouse Gases Equivalencies Calculator-Calculations and References [WWW Document]. Energy and the Environment. Available online at: https://www.epa.gov/energy/greenhouse-gases-equivalencies-calculator-calculations-and-references (accessed March17, 2020).
Eshel, G., Shepon, A., Makov, T., and Milo, R. (2014). Land, irrigation water, greenhouse gas, and reactive nitrogen burdens of meat, eggs, and dairy production in the United States. Proc. Natl. Acad. Sci. U.S.A. 111, 11996–12001. doi: 10.1073/pnas.1402183111
Follett, R., Kimble, J., and Lal, R., (eds.). (2000). “The potential of U.S. grazing lands to sequester soil carbon,” in The Potential of U.S. Grazing Lands to Sequester Carbon and Mitigate the Greenhouse Effect, (Boca Raton, FL: CRC Press), 401–427. doi: 10.1201/9781420032468.sec5
Franzluebbers, A. J. (2002). Water infiltration and soil structure related to organic matter and its stratification with depth. Soil Tillage Res. 66, 197–205. doi: 10.1016/S0167-1987(02)00027-2
Gardner, J. B., and Drinkwater, L. E. (2009). The fate of nitrogen in grain cropping systems: a meta-analysis of 15N field experiments. Ecol. Appl. 19, 2167–2184. doi: 10.1890/08-1122.1
Gerber, P. J., Steinfeld, H., Henderson, B., Mottet, A., Opio, C., Dijkman, J., et al. (2013). Tackling Climate Change Through Livestock; A Global Assessment of Emissions and Mitigation Opportunities. Rome: FAO.
Gosnell, H., Charnley, S., and Stanley, P. (2020). Climate change mitigation as a co-benefit of regenerative ranching: insights from Australia and the United States. Interface Focus 10:20200027. doi: 10.1098/rsfs.2020.0027
Gosnell, H., Gill, N., and Voyer, M. (2019). Transformational adaptation on the farm: processes of change and persistence in transitions to ‘climate-smart’ regenerative agriculture. Glob. Environ. Change 59:101965. doi: 10.1016/j.gloenvcha.2019.101965
Griscom, B. W., Adams, J., Ellis, P. W., Houghton, R. A., Lomax, G., Miteva, D. A., et al. (2017). Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650. doi: 10.1073/pnas.1710465114
Gupta, V. V. S. R., and Germida, J. J. (2015). Soil aggregation: influence on microbial biomass and implications for biological processes. Soil Biol. Biochem. 80, A3–A9. doi: 10.1016/j.soilbio.2014.09.002
Heflin, K. R., Parker, D. B., Marek, G. W., Auvermann, B. W., and Marek, T. H. (2019). Greenhouse-gas emissions of beef finishing systems in the Southern High Plains. Agric. Syst. 176:102674. doi: 10.1016/j.agsy.2019.102674
Herrero, M., Thornton, P. K., Notenbaert, A. M., Wood, S., Msangi, S., Freeman, H. A., et al. (2010). Smart investments in sustainable food production: revisiting mixed crop-livestock systems. Science 327, 822–825. doi: 10.1126/science.1183725
Hillenbrand, M., Thompson, R., Wang, F., Apfelbaum, S., and Teague, R. (2019). Impacts of holistic planned grazing with bison compared to continuous grazing with cattle in South Dakota shortgrass prairie. Agric. Ecosyst. Environ. 279, 156–168. doi: 10.1016/j.agee.2019.02.005
IPCC (2014). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva: IPCC. p. 151.
Jagadamma, S., Mayes, M. A., Zinn, Y. L., Gísladóttir, G., and Russell, A. E. (2014). Sorption of organic carbon compounds to the fine fraction of surface and subsurface soils. Geoderma 213, 79–86. doi: 10.1016/j.geoderma.2013.07.030
Kamilaris, C., Dewhurst, R. J., Sykes, A. J., and Alexander, P. (2020). Modelling alternative management scenarios of economic and environmental sustainability of beef finishing systems. J. Clean. Prod. 253:119888. doi: 10.1016/j.jclepro.2019.119888
Kebreab, E., Liedke, A., Caro, D., Deimling, S., Binder, M., and Finkbeiner, M. (2016). Environmental impact of using specialty feed ingredients in swine and poultry production: a life cycle assessment. J. Anim. Sci. 94, 2664–2681. doi: 10.2527/jas.2015-9036
Kremen, C., Iles, A., and Bacon, C. (2012). Diversified farming systems: an agroecological, systems-based alternative to modern industrial agriculture. Ecol. Soc. 17:44. doi: 10.5751/ES-05103-170444
Kremen, C., and Merenlender, A. M. (2018). Landscapes that work for biodiversity and people. Science 362:eaau6020. doi: 10.1126/science.aau6020
Kumar, S., Sieverding, H., Lai, L., Thandiwe, N., Wienhold, B., Redfearn, D., et al. (2019). Facilitating crop–livestock reintegration in the northern great plains. Agron. J. 111, 2141–2156. doi: 10.2134/agronj2018.07.0441
Lal, R. (2002). Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 116, 353–362. doi: 10.1016/S0269-7491(01)00211-1
Lavallee, J. M., Soong, J. L., and Cotrufo, M. F. (2020). Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Change Biol. 26, 261–273. doi: 10.1111/gcb.14859
Lee, J., Hopmans, J. W., Rolston, D. E., Baer, S. G., and Six, J. (2009). Determining soil carbon stock changes: Simple bulk density corrections fail. Agric. Ecosyst. Environ. 134, 251–256. doi: 10.1016/j.agee.2009.07.006
Liebig, M. A., Gross, J. R., Kronberg, S. L., Phillips, R. L., and Hanson, J. D. (2010). Grazing management contributions to net global warming potential: a long-term evaluation in the Northern Great Plains. J. Environ. Qual. 39, 799–809. doi: 10.2134/jeq2009.0272
Lupo, C. D., Clay, D. E., Benning, J. L., and Stone, J. J. (2013). Life-cycle assessment of the beef cattle production system for the Northern Great Plains, USA. J. Environ. Qual. 42, 1386–1394. doi: 10.2134/jeq2013.03.0101
Lynch, J., Cain, M., Pierrehumbert, R., and Allen, M. (2020). Demonstrating GWP*: a means of reporting warming-equivalent emissions that captures the contrasting impacts of short-and long-lived climate pollutants. Environ. Res. Lett. 15:044023. doi: 10.1088/1748-9326/ab6d7e
Machmuller, M. B., Kramer, M. G., Cyle, T. K., Hill, N., Hancock, D., and Thompson, A. (2015). Emerging land use practices rapidly increase soil organic matter. Nat. Commun. 6:6995. doi: 10.1038/ncomms7995
MacLeod, M., Gerber, P., Mottet, A., Tempio, G., Falcucci, A., Opio, C., et al. (2013). Greenhouse gas emissions from pig and chicken supply chain - a global life cycle assessment . Rome: Food and Agriculture Organization of the United Nations.
McSherry, M. E., and Ritchie, M. E. (2013). Effects of grazing on grassland soil carbon: a global review. Glob. Change Biol. 19, 1347–1357. doi: 10.1111/gcb.12144
Meeh, D. C., Rowntree, J. E., and Hamm, M. W. (2014). Feeding a population with smaller scale and alternate system production: an examination of farm requirements with a multi-species pasture system to feed 10 million people. RAF 29, 176–185. doi: 10.1017/S1742170513000070
Minasny, B., and Mcbratney, A. B. (2017). Limited effect of organic matter on soil available water capacity. Eur. J. Soil Sci. 69, 39–47. doi: 10.1111/ejss.12475
Moebius-Clune, B. N., Moebius-Clune, D. J., Gugino, B. K., Idowu, O. J., Schindelbeck, R. R., Ristow, A. J., et al. (2016). Comprehensive Assessmentof Soil Health-The Cornell Framework (Manual). Geneva, NY: Cornell University.
Nicoloso, R. S., Rice, C. W., Amado, T. J. C., Costa, C. N., and Akley, E. K. (2018). Carbon saturation and translocation in a no-till soil under organic amendments. Agric. Ecosyst. Environ. 264, 73–84. doi: 10.1016/j.agee.2018.05.016
Pelletier, N., Lammers, P., Stender, D., and Pirog, R. (2010a). Life cycle assessment of high- and low-profitability commodity and deep-bedded niche swine production systems in the Upper Midwestern United States. Agric. Syst. 103, 599–608. doi: 10.1016/j.agsy.2010.07.001
Pelletier, N., Pirog, R., and Rasmussen, R. (2010b). Comparative life cycle environmental impacts of three beef production strategies in the Upper Midwestern United States. Agric. Syst. 103, 380–389. doi: 10.1016/j.agsy.2010.03.009
Picasso, V. D., Modernel, P. D., Becoña, G., Salvo, L., Gutiérrez, L., and Astigarraga, L. (2014). Sustainability of meat production beyond carbon footprint: a synthesis of case studies from grazing systems in Uruguay. Meat Sci. 98, 346–354. doi: 10.1016/j.meatsci.2014.07.005
Pierrehumbert, R. T., and Eshel, G. (2015). Climate impact of beef: an analysis considering multiple time scales and production methods without use of global warming potentials. Environ. Res. Lett. 10:085002. doi: 10.1088/1748-9326/10/8/085002
River Valley Regional Commission (2014). Clay County Comprehensive Plan Update 2014. River Valley Regional Commission.
Rivera-Ferre, M. G., López-i-Gelats, F., Howden, M., Smith, P., Morton, J. F., and Herrero, M. (2016). Re-framing the climate change debate in the livestock sector: mitigation and adaptation options. WIREs Clim. Change 7, 869–892. doi: 10.1002/wcc.421
Robinson, T. P., Thorton, P. K., Franceschini, G., Kruska, R. L., Chiozza, F., Notenbaert, A., et al. (2011). Global Livestock Production Systems. Rome: Food and Agriculture Organization of the United Nations (FAO) and International Livestock Research Institute (ILRI).
Rotz, C. A., Asem-Hiablie, S., Place, S., and Thoma, G. (2019). Environmental footprints of beef cattle production in the United States. Agric. Syst. 169, 1–13. doi: 10.1016/j.agsy.2018.11.005
Russelle, M. P., Entz, M. H., and Franzluebbers, A. J. (2007). Reconsidering integrated crop–livestock systems in north america. Agron. J. 99, 325–334. doi: 10.2134/agronj2006.0139
Savory, A., and Butterfield, J. (2016). Holistic Management, Third Edition: A Commonsense Revolution to Restore Our Environment. Washington, DC: Island Press.
Schimel, D. S. (1986). Carbon and nitrogen turnover in adjacent grassland and cropland ecosystems. Biogeochemistry 2, 345–357. doi: 10.1007/BF02180325
Schuman, G. E., Janzen, H. H., and Herrick, J. E. (2002). Soil carbon dynamics and potential carbon sequestration by rangelands. Environ. Pollut. 116, 391–396. doi: 10.1016/S0269-7491(01)00215-9
Stanley, P. L., Rowntree, J. E., Beede, D. K., DeLonge, M. S., and Hamm, M. W. (2018). Impacts of soil carbon sequestration on life cycle greenhouse gas emissions in Midwestern USA beef finishing systems. Agric. Syst. 162, 249–258. doi: 10.1016/j.agsy.2018.02.003
Tallaksen, J., Johnston, L., Sharpe, K., Reese, M., and Buchanan, E. (2020). Reducing life cycle fossil energy and greenhouse gas emissions for Midwest swine production systems. J. Clean. Prod. 246:118998. doi: 10.1016/j.jclepro.2019.118998
Teague, R., Provenza, F., Kreuter, U., Steffens, T., and Barnes, M. (2013). Multi-paddock grazing on rangelands: why the perceptual dichotomy between research results and rancher experience? J. Environ. Manage. 128, 699–717. doi: 10.1016/j.jenvman.2013.05.064
Teague, W. R., Apfelbaum, S., Lal, R., Kreuter, U. P., Rowntree, J., Davies, C. A., et al. (2016). The role of ruminants in reducing agriculture's carbon footprint in North America. J. Soil Water Conserv. 71, 156–164. doi: 10.2489/jswc.71.2.156
Teague, W. R., Dowhower, S. L., Baker, S. A., Haile, N., DeLaune, P. B., and Conover, D. M. (2011). Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ. 141, 310–322. doi: 10.1016/j.agee.2011.03.009
Thompson, L. R. (2020). Invited review: methane sources, quantification, andmitigation in grazing systems. Appl. Anim. Sci. 36, 556–573. doi: 10.15232/aas.2019-01951
Tisdall, J. M., and Oades, J. M. (1982). Organic matter and water-stable aggregates in soils. J. Soil Sci. 33, 141–163. doi: 10.1111/j.1365-2389.1982.tb01755.x
Tongpool, R., Phanichavalit, N., Yuvaniyama, C., and Mungcharoen, T. (2012). Improvement of the environmental performance of broiler feeds: a study via life cycle assessment. J. Clean. Prod. 35, 16–24. doi: 10.1016/j.jclepro.2012.05.007
USDA NASS (2018). USDA/NASS 2019 State Agriculture Overview for Georgia [WWW Document]. 2019 State Agriculture Overview. Available online at: https://www.nass.usda.gov/Quick_Stats/Ag_Overview/stateOverview.php?state=GEORGIA (accessed March 18, 2020).
Walker, L. R., Wardle, D. A., Bardgett, R. D., and Clarkson, B. D. (2010). The use of chronosequences in studies of ecological succession and soil development. J. Ecol. 98, 725–736. doi: 10.1111/j.1365-2745.2010.01664.x
Wall, D. H., Behan-Pelletier, V., Ritz, K., Herrick, J. E., Jones, T. H., Six, J., et al. (2012). Soil Ecology and Ecosystem Services. Oxford: Oxford University Press.
Wang, T., Teague, W., Park, S., and Bevers, S. (2015). GHG mitigation potential of different grazing strategies in the united states southern great plains. Sustainability 7, 13500–13521. doi: 10.3390/su71013500
West, T. O., and Six, J. (2007). Considering the influence of sequestration duration and carbon saturation on estimates of soil carbon capacity. Clim. Change 80, 25–41. doi: 10.1007/s10584-006-9173-8
Keywords: regenerative agriculture, soil carbon (C) sequestration, life cycle (impact) assessment, multi-species grazing, holistic planned grazing™
Citation: Rowntree JE, Stanley PL, Maciel ICF, Thorbecke M, Rosenzweig ST, Hancock DW, Guzman A and Raven MR (2020) Ecosystem Impacts and Productive Capacity of a Multi-Species Pastured Livestock System. Front. Sustain. Food Syst. 4:544984. doi: 10.3389/fsufs.2020.544984
Received: 23 March 2020; Accepted: 22 October 2020;
Published: 04 December 2020.
Edited by:
Juan Jose Villalba, Utah State University, United StatesReviewed by:
Kathryn Ann Slebodnik, Utah State University, United StatesJasmine A. Dillon, Colorado State University, United States
Copyright © 2020 Rowntree, Stanley, Maciel, Thorbecke, Rosenzweig, Hancock, Guzman and Raven. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jason E. Rowntree, cm93bnRyZTFAbXN1LmVkdQ==