 Daniela A. Pereira1†
Daniela A. Pereira1† Marcelo R. Luizon1,2*†‡
Marcelo R. Luizon1,2*†‡ Ana C. Palei3
Ana C. Palei3 José E. Tanus-Santos4
José E. Tanus-Santos4 Ricardo C. Cavalli5
Ricardo C. Cavalli5 Valeria C. Sandrim2*‡
Valeria C. Sandrim2*‡- 1Department of Genetics, Ecology and Evolution, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais, Brazil
- 2Department of Biophysics and Pharmacology, Institute of Biosciences, Universidade Estadual Paulista (UNESP), Botucatu, Brazil
- 3Department of Surgery, University of Mississippi Medical Center, Jackson, MS, United States
- 4Department of Pharmacology, Ribeirao Preto Medical School, University of Sao Paulo, Ribeirão Preto, Brazil
- 5Department of Gynecology and Obstetrics, Ribeirao Preto Medical School, University of Sao Paulo, Ribeirão Preto, Brazil
Impaired nitric oxide (NO) formation may be associated with endothelial dysfunction and increased cardiovascular disease risk in preeclampsia (PE). Functional single-nucleotide polymorphisms (SNPs) of nitric oxide synthase 3 (NOS3) (rs3918226) and guanylate cyclase 1, soluble, alpha 3 (GUCY1A3) (rs7692387) increase susceptibility to the adverse consequences due to inadequate generation of NO by the endothelium. However, no previous study has examined whether these SNPs affect NO formation in healthy pregnancy and in gestational hypertension (GH) and PE. Here, we compared the alleles and genotypes of NOS3 (rs3918226) and GUCY1A3 (rs7692387) SNPs in normotensive pregnant women (NP, n = 153), in GH (n = 96) and PE (n = 163), and examined whether these SNPs affect plasma nitrite concentrations (a marker of NO formation) in these groups. We further examined whether the interaction among SNP genotypes is associated with GH and PE. Genotypes were determined using TaqMan allele discrimination assays, and plasma nitrite concentrations were determined by an ozone-based chemiluminescence assay. Multifactor dimensionality reduction was used to examine the interactions among SNP genotypes. Regarding NOS3 rs3918226, the CT genotype (p = 0.046) and T allele (p = 0.020) were more frequent in NP than in GH, and GH patients carrying the CT+TT genotypes showed lower nitrite concentrations than NP carrying the CT+TT genotypes (p < 0.05). Regarding GUCY1A3 rs7692387, the GA genotype (p = 0.013) and A allele (p = 0.016) were more frequent in PE than in NP, and NP women carrying the GG genotype showed higher nitrite concentrations than GH or PE patients carrying the GG genotype (p < 0.05). However, we found no significant interactions among genotypes for these functional SNPs to be associated with GH or PE. Our novel findings suggest that NOS3 rs3918226 and GUCY1A3 rs7692387 may affect NO formation and association with hypertensive disorders of pregnancy.
1 Introduction
Hypertensive disorders of pregnancy affect up to 10% of pregnancies (Hutcheon et al., 2011), which include gestational hypertension (GH) and preeclampsia (PE). PE is defined as new-onset hypertension after 20 weeks of pregnancy that occurs along with proteinuria or other indications of renal insufficiency, thrombocytopenia, liver dysfunction, pulmonary edema, and cerebral disturbances (ACOG, 2013). Therefore, PE is a complex multisystem disease and a major cause of maternal and perinatal mortality and morbidity (Chappell et al., 2021). Moreover, an increased risk of future cardiovascular disease has been observed in women with GH (Lo et al., 2020) and in women with a history of PE (Wu et al., 2017; Veiga et al., 2021), which can be explained by endothelial dysfunction during PE (Ahmed et al., 2014).
Widespread vascular endothelial dysfunction observed in PE is caused by the release of antiangiogenic factors into maternal circulation as a result of placental ischemia and hypoxia, including the soluble fms-like tyrosine kinase-1 (sFlt-1) (Maynard et al., 2003; Powe et al., 2011). It is to be noted that previous studies conducted by our research group found sFlt-1 to be inversely related to nitric oxide (NO) formation in PE, which is characterized by reduced bioavailability of NO (Sandrim et al., 2008b). In addition, we found that single-nucleotide polymorphisms (SNPs) of the endothelial NO synthase 3 (NOS3) gene may affect plasma and blood nitrite concentrations (a marker of endogenous NO formation) in healthy subjects (Metzger et al., 2013) and in patients with PE (Sandrim et al., 2010), and these SNPs of NOS3 were found to be associated with susceptibility to GH or PE (Sandrim et al., 2008a; Muniz et al., 2012).
NO signaling pathway plays key roles in the regulation of vascular tone and platelet activation (Emdin et al., 2018). Notably, NO produced by the NOS3 enzyme is an effective vasodilator, with a key role in the control of blood pressure (Vanhoutte, 2003; Moncada and Higgs, 2006). NO activates soluble guanylyl cyclase, a heterodimeric protein with a subunit encoded by the guanylate cyclase 1, soluble, alpha 3 (GUCY1A3) gene (Forstermann et al., 1994; Mergia et al., 2006). Soluble guanylyl cyclase then produces cyclic guanosine monophosphate (cGMP), which activates downstream signaling molecules that lead to vasodilation, inhibition of platelet aggregation, and lowering of blood pressure (Denninger and Marletta, 1999). Therefore, NOS3 and GUCY1A3 are key mediators of NO signaling and its downstream effects (Emdin et al., 2018).
A remarkable study explored >335,000 participants in the UK Biobank genotyped using Affymetrix arrays, along with analysis from seven large-scale GWAS including nearly 1.5 million subjects, and found that variants that increase NO and NO-mediated cGMP synthesis are associated with decreased mean arterial pressure and reduced risk of coronary heart disease, peripheral arterial disease, and stroke (Emdin et al., 2018). Notably, common functional SNPs of NOS3 (rs3918226) and GUCY1A3 (rs7692387) were highlighted among these several array SNPs, which confirmed the adverse consequences of inadequate NO generation by the endothelium (Emdin et al., 2018; Loscalzo, 2018). However, no previous study has examined whether these functional SNPs of NOS3 and GUCY1A3 affect NO formation in healthy pregnancy and in GH or PE, including their association with GH or PE. Moreover, since NOS3 and GUCY1A3 are key genes that mediate NO signaling, they may interact to generate NO. However, no previous study has examined whether the interaction among their genotypes is associated with GH and PE.
In this study, we examined the association of these functional SNPs of NOS3 (rs3918226) and GUCY1A3 (rs7692387) with susceptibility to GH and PE and whether they affect NO formation in normotensive pregnant (NP) women and in patients with GH and PE. Moreover, we examined whether the interaction among genotypes for these functional SNPs is associated with GH and PE.
2 Methods
2.1 Study population
All volunteers were enrolled at the Department of Obstetrics and Gynecology, University Hospital at Ribeirao Preto Medical School of University of Sao Paulo (FMRP/USP). The Institutional Review Board at FMRP/USP approved this study. We included normotensive women with uncomplicated pregnancies (NP, n = 153) and patients with GH (n = 96) and PE (n = 163). GH was defined as pregnancy-induced hypertension (≥140 mmHg systolic or ≥90 mmHg diastolic blood pressure on two or more measurements at least 6 h apart) after 20 weeks of gestation and returning to normal by 12 weeks post-partum. PE was defined as GH along with significant proteinuria (≥0.3 g/L per 24 h) or with renal insufficiency, thrombocytopenia, liver dysfunction, pulmonary edema, and cerebral or visual symptoms (ACOG, 2013). The exclusion criteria of the study were women with pre-existing hypertension, with/without superimposed PE, and women with gestational diabetes. All individuals who were enrolled in this study are unrelated.
Written informed consent was provided by volunteers during clinic attendance. Maternal venous blood samples were collected into tubes containing EDTA and heparin, which were used for DNA extraction and to measure nitrite, respectively. Plasma samples were obtained after centrifugation at 1000 g for 10 min and stored at −70°C until assayed. Genomic DNA was extracted using a salting-out method from the cellular fraction of 1 mL of whole blood and stored at −20°C until analyzed.
2.2 Genotyping
Genotypes for NOS3 rs3918226 and GUCY1A3 rs7692387 were determined using TaqMan allelic discrimination assays (IDs C__30245515_10, and C__29125113_10, respectively; Applied Biosystems, Carlsbad, CA, USA). Real-time PCR was performed in a total volume of 12 μL (3 ng of template DNA, 1x TaqMan Genotyping Master Mix (Life Technologies Corporation, Grand Island, NY, USA) and 1x TaqMan allelic discrimination assay). Thermal cycling was performed in standard conditions, fluorescence was recorded by StepOnePlus Real-Time PCR equipment (Applied Biosystems, Carlsbad, CA, USA), and the results were obtained using the manufacturer’s software.
2.3 Measurement of nitrite concentrations
Nitrite concentrations were measured using an ozone-based chemiluminescence assay, as previously described (Sandrim et al., 2008b). Briefly, 200 μL of plasma aliquots analyzed in triplicate was injected into a solution of acidified triiodide, purging with nitrogen in-line with a gas-phase chemiluminescence NO analyzer (Sievers Model 280 NO Analyzer, General Electric Company, Boulder, CO, USA). Approximately 8 mL of triiodide solution (2.0 g of potassium iodide and 1.3 g of iodine dissolved in 40 mL of water with 140 mL of acetic acid) was placed in the purging vessel to which plasma samples were added. The triiodide solution reduced nitrite to NO gas, which was detected by the NO analyzer.
2.4 Statistical analysis
The characteristics of NP and GH and PE patients were compared using Student’s unpaired t-test, Mann–Whitney U test, or x2 test as appropriate. The effects of different genotypes for NOS3 and GUCY1A3 SNPs on circulating nitrite concentrations among and within study groups were compared using one-way ANOVA (Kruskal–Wallis) with post-hoc tests (Dunn’s multiple comparison). Distribution of genotypes was assessed for deviation from Hardy–Weinberg equilibrium using Hardy–Weinberg exact tests for each locus in each population using the “web version of Genepop” available at https://genepop.curtin.edu.au/genepop_op1.html, which generates the estimation of exact p-values by the Markov chain method. We have used the suboptions “For each locus in the population” and “3. Probability test,” and the Markov chain parameters (default values): dememorization: 1000; batches: 100; iterations per batch: 1000 (Rousset, 2008) and differences in genotype and allele frequencies were assessed using x2 tests. The difference in plasma nitrite concentrations among the study groups was obtained using one-way ANOVA followed by Bonferroni’s post-hoc test. The relationships between plasma nitrite concentrations and fasting glucose were analyzed using Spearman’s correlation (r and p values). A value of p < 0.05 was considered the level of statistical significance. Power calculation was performed using QUANTO version 1.2.4 (Gauderman, 2002). Given the sample size of the study, the power was 0.824 considering an alpha of 0.05 and the frequency of the minor allele of SNP GUCY1A3 (rs7692387, G>A), which was found to be significantly associated with PE. The power was 0.721 considering an alpha of 0.05 and the frequency of the minor allele of SNP NOS3 (rs3918226, C>T), which was found to be significantly more frequent in NP than in GH.
Multifactor dimensionality reduction (MDR) characterizes the interactions of genotypes for their ability to be classified into different groups through cross-validation (CV) steps and permutation testing (Ritchie and Motsinger, 2005; Pander et al., 2010). The interaction model with the highest testing score and CV consistency (CVC) was considered the best interaction model, and its statistical significance was determined using permutation testing (Ritchie and Motsinger, 2005; Luizon et al., 2014; Luizon et al., 2017).
3 Results
The characteristics of subjects enrolled in this study are shown in Table 1. NP and GH and PE patients showed similar ethnicity, current smokers, heart rate, hemoglobin, hematocrit, creatinine, primiparity, and gestational age at sampling (all p > 0.05). As expected, PE and GH patients had higher systolic and diastolic blood pressure than NP patients (both p < 0.05), despite that most of these patients were receiving antihypertensive drugs. Patients with PE and GH were older than those with NP (p < 0.05). Increased body mass index and fasting glucose were found in both PE and GH compared with NP patients (both p < 0.05). Gestational age at delivery and newborn weights were lower in PE patients than in GH patients and NP (all p < 0.05). Significant proteinuria was found in PE.
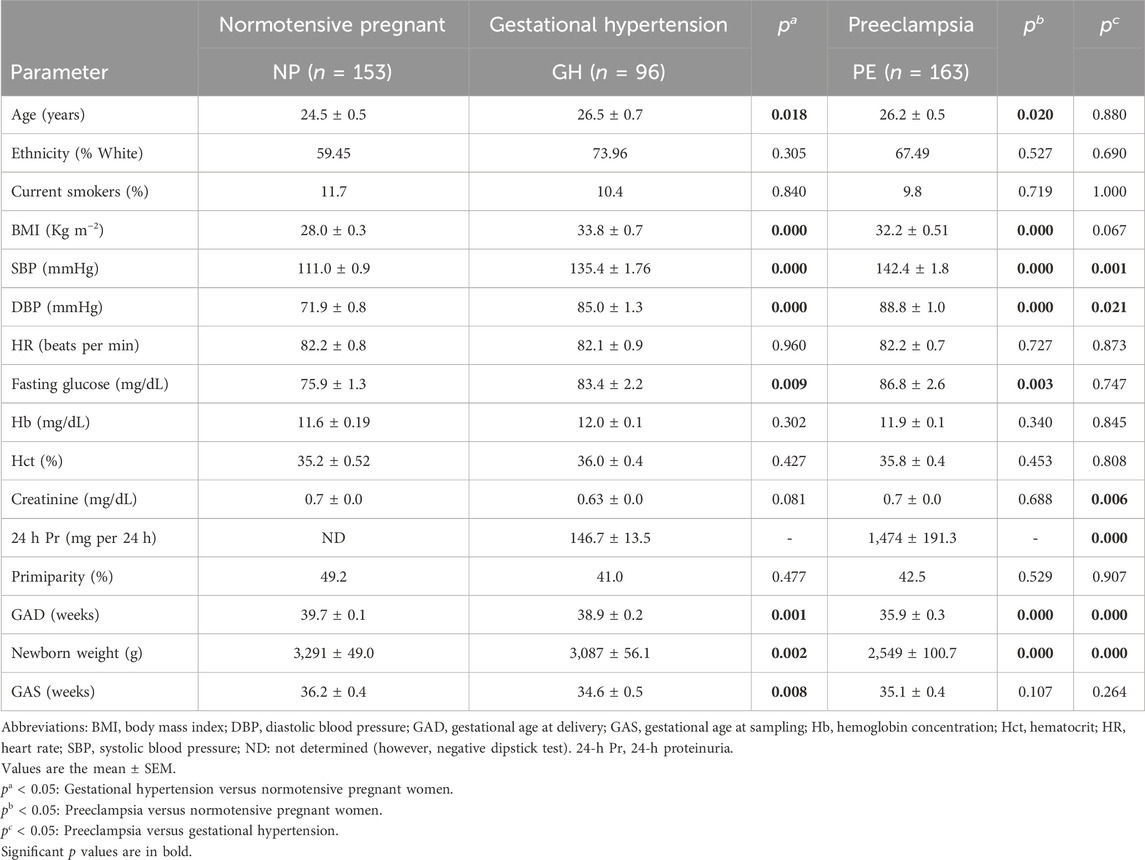
TABLE 1. Clinical and demographic characteristics of the study subjects.
Genotype and allele distributions for NP, GH, and PE are shown in Table 2. The distribution of genotypes for each SNP in each of the study groups showed no deviation from the Hardy–Weinberg equilibrium (all p > 0.05). Regarding the NOS3 rs3918226 SNP, the CT genotype and the T allele were more frequent in NP than in GH patients (both p < 0.05, and odds ratio (OR) = 0.435 and 0.407, respectively; Table 2). Considering the dominant model, the CT+TT genotypes were more frequent in NP than in GH patients (p = 0.032, OR = 0.407; Table 2). Regarding the GUCY1A3 rs7692387 SNP, the GA genotype and the A allele were more frequent in PE patients than in NP patients (p < 0.05, OR = 1.991 and OR = 1.742, respectively; Table 2). Considering the dominant model, the GA+AA genotypes were more frequent in PE patients than in NP patients (p = 0.009, OR = 1.955; Table 2). We examined the effects of NOS3 and GUCY1A3 genotypes on circulating nitrite concentrations. Due to no availability of plasma samples, circulating nitrite concentrations were measured only in a small number of subjects, NP (n = 95), GH (n = 41), and PE (n = 69). Plasma nitrite concentrations in the three study groups are shown in Supplementary Figure S1, which were found to be lower in both groups of hypertensive disorders of pregnancy (GH and PE) than in NP (both p < 0.05), as previously reported (Sandrim et al., 2008b). Regarding NOS3 rs3918226, GH patients carrying both the CC and CT+TT genotypes showed lower nitrite concentrations than NP patients carrying the CC and CT+TT genotypes (p < 0.05, Figure 1A). Regarding GUCY1A3 rs7692387, NP patients carrying the GG genotype showed higher nitrite concentrations than GH and PE patients carrying the GG genotype (p < 0.05, Figure 1B). However, we found no significant effects of different genotypes for both SNPs on nitrite concentrations within each of the study groups (p > 0.05).
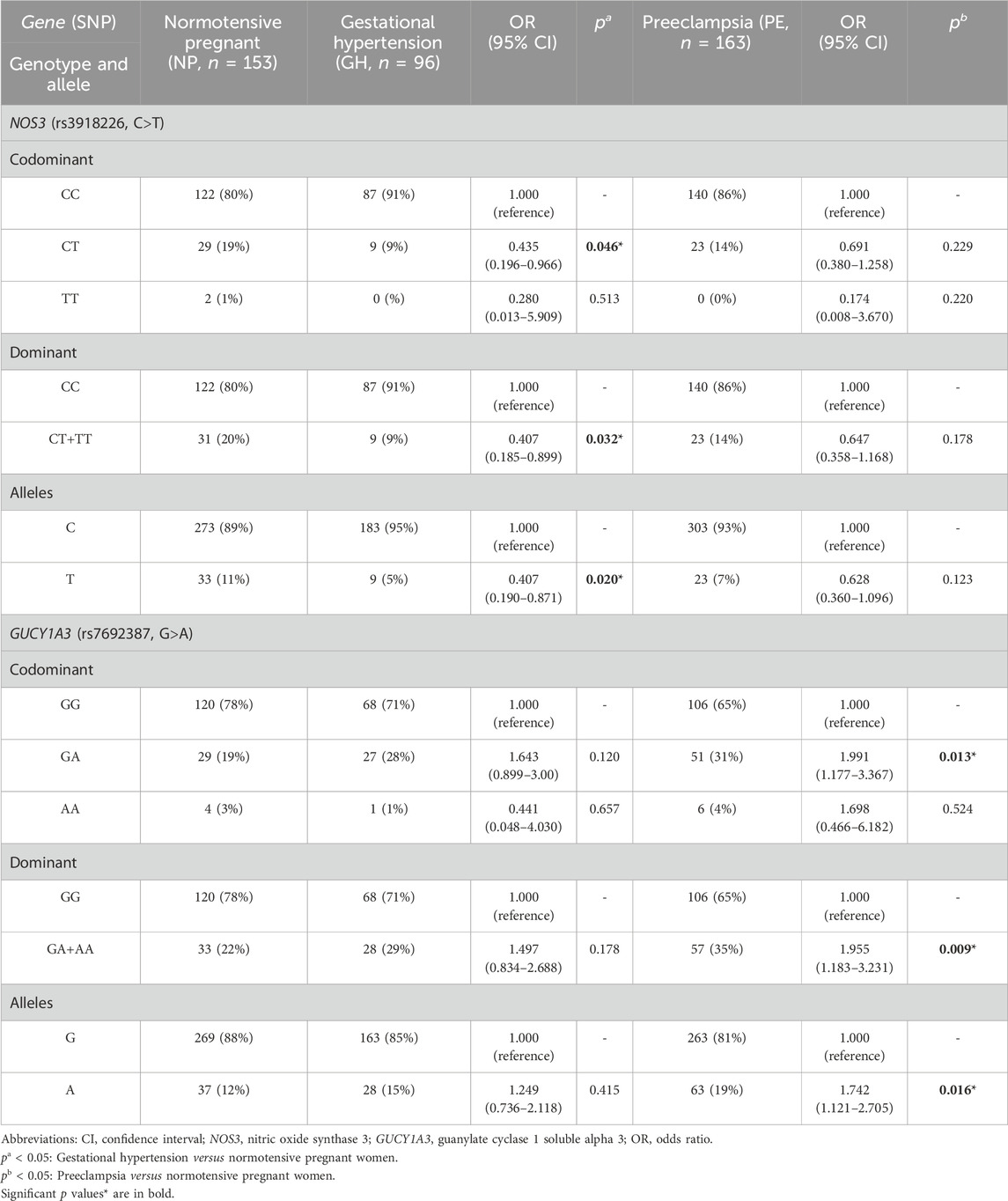
TABLE 2. Genotype and allele frequencies for NOS3 and GUCY1A3 polymorphisms in normotensive pregnant women and in patients with gestational hypertension and preeclampsia.
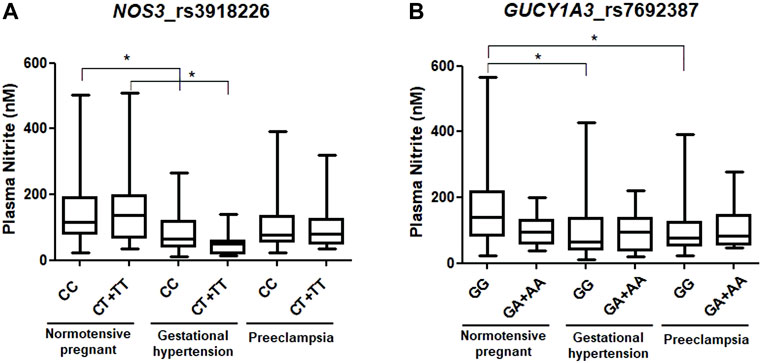
FIGURE 1. Effects of genotypes of (A) NOS3 rs3918226 C>T and (B) GUCY1A3 rs7692387 G>A SNPs on circulating nitrite concentrations in normotensive pregnant women and in patients with gestational hypertension and preeclampsia. *p < 0.05.
We further examined the relationships between plasma nitrite concentrations and fasting glucose in the study groups (Supplementary Figure S2); however, we found no significant correlations.
Since NOS3 and GUCY1A3 are key genes that mediate NO signaling, it is possible that they may interact to generate NO. Therefore, we further examined whether the possible interaction among their genotypes was associated with susceptibility to GH and PE. However, we found no model of interaction among genotypes of NOS3 (rs3918226) and GUCY1A3 (rs7692387) SNPs to be significantly associated with GH or with PE (p > 0.05, Table 3).
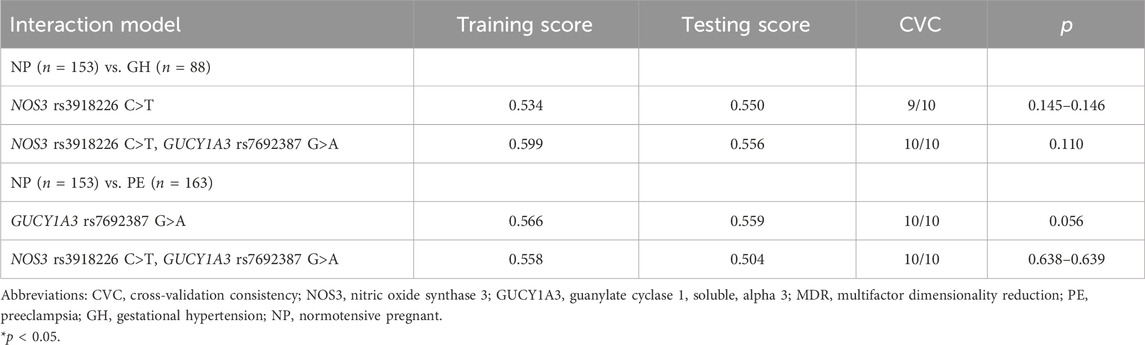
TABLE 3. MDR interaction models among NOS3 and GUCY1A3 polymorphisms in normotensive pregnant (NP) compared with both gestational hypertension (GH) and with preeclampsia (PE).
4 Discussion
The main novel findings reported here are that the CT genotype and T allele for NOS3 rs3918226 were more frequent in NP than in GH patients, and GH patients carrying the CT+TT genotypes showed lower nitrite concentrations than NP patients carrying the CT+TT genotypes. Regarding GUCY1A3 rs7692387, the GA genotype and A allele were more frequent in PE patients than in NP, and NP patients carrying the GG genotype showed higher nitrite concentrations than GH or PE patients carrying the GG genotype. However, no significant model of interaction among genotypes of these functional NOS3 and GUCY1A3 SNPs was associated with GH or with PE.
Common variants in the NOS3 promoter (rs3918226) and GUCY1A3 intron (rs7692387) were associated with increased NOS3 and GUCY1A3 expression and with reduced mean arterial pressure (Emdin et al., 2018). The effects of NOS3 rs3918226 on NO formation were previously examined in healthy subjects (Luizon et al., 2012), and no significant differences were found when comparing the nitrite concentrations in the CC genotype with the CT+TT genotypes (Luizon et al., 2012). However, the NOS3 rs3918226 was previously associated with NO production in children and adolescents (de Miranda et al., 2015). rs3918226 is located next to a potential transcription factor-binding site for the Ets family domain, and it may affect transcription factor-binding affinity (Salvi et al., 2012). Indeed, it was previously shown to affect NOS3 expression using luciferase assays in HeLa and HEK293T cells transfected with the NOS3 promoter carrying either the T or the C allele (Salvi et al., 2013). The T allele was associated with reduced NOS3 transcriptional activity compared with the C allele of the rs3918226 located at the NOS3 promoter (Salvi et al., 2013). Moreover, the TT homozygosity was associated with lesser transcription and higher hypertension risk (Salvi et al., 2013). In the Genome Tissue Expression (GTEx) project database (Consortium, 2015), the C allele was associated with increased NOS3 expression in the lung tissue. In the present study, we find that GH patients carrying the CT+TT genotypes showed lower nitrite concentrations than NP patients carrying the CT+TT genotypes. However, we found no difference within each of the study groups.
To our knowledge, no previous study has examined the association of GUCY1A3 polymorphisms with PE. We showed for the first time that the GA genotype and A allele for the GUCY1A3 rs7692387 were more frequent in PE patients than in NP. Consistent with this finding, the A allele of GUCY1A3 rs7692387 was associated with hypertensive disorders of pregnancy (OR = 0.90 [95% CI, 0.84–0.97]; p = 0.004) (Honigberg et al., 2020). However, this study used phenotypic data from the UK Biobank, but it has not defined the pregnant women who had PE (Honigberg et al., 2020). Nevertheless, no previous study has examined the effects of GUCY1A3 polymorphisms on NO formation in hypertensive disorders of pregnancy. The minor allele A of the rs7692387 polymorphism was associated with increased GUCY1A3 expression in the GTEx portal (Consortium, 2015). Accordingly, the G allele of GUCY1A3 rs7692387 has been characterized to reduce the expression of GUCY1A3 via disruption of a ZEB1 transcription factor site (Kessler et al., 2017). However, we found no significant effects for the different GUCY1A3 rs7692387 genotypes on circulating nitrite concentrations within each of the study groups. Despite that, NP patients carrying the GG genotype showed higher nitrite levels than those of GH and PE carrying the same genotype. The protective effect of variation on GUCY1A3 rs7692387 against hypertensive disorders of pregnancy and the underlying pathways relative to blood pressure lowering requires further studies.
Since plasma nitrite levels were previously associated with body mass index (Ghasemi et al., 2008), fasting blood glucose, and systolic blood pressure (Li et al., 2004), we examined the potential relationships between plasma nitrite concentrations and fasting glucose in the three study groups. However, we found no significant correlations between plasma nitrite concentrations and fasting glucose in our study, which could be due to the small number of subjects with measurements for nitrite concentrations for NP and GH and PE. Further studies are needed to explore the correlations of plasma nitrite concentrations with fasting glucose in hypertensive disorders of pregnancy.
These studies show that SNPs of NOS3 and GUCY1A3 may individually be relevant to the susceptibility of PE, with a focus on the role of NO bioavailability in the physiopathology of PE. However, epistasis may also be relevant, and gene–gene interactions have been used before to detect susceptibility to complex diseases and response to drugs, including PE, (Gui et al., 2011; Williams and Broughton Pipkin, 2011; Singhal et al., 2023) and response to antihypertensive therapy (Luizon et al., 2018). Given that the final products of NOS3 and GUCY1A3 genes interact together in the same biological pathway (Wobst et al., 2018), the NO produced by NOS3 activates soluble guanylyl cyclase, which has a subunit encoded by GUCY1A3 (Forstermann et al., 1994; Mergia et al., 2006). Thus, it is of relevance to explore the interaction among its polymorphisms and how it could affect the susceptibility to hypertensive disorders of pregnancy. In this context, we examined whether the interactions among genotypes for the functional SNPs of NOS3 (rs3918226) and GUCY1A3 (rs7692387) were associated with GH and with PE. Although we found no significant model of interaction, further studies should be performed in order to examine the interactions among other polymorphisms in other candidate genes that are relevant to the physiopathology of PE.
The present study has limitations. The circulating nitrite concentrations were measured only in a small number of subjects enrolled due to no availability of plasma samples and technical reasons. Nevertheless, we were able to found significant effects of different genotypes for NOS3 rs3918226 and GUCY1A3 rs7692387 on nitrite concentrations in hypertensive disorders of pregnancy. Importantly, our findings must be replicated on different populations by further studies, which should consider an increase in the sample size. It is to be noted that allele frequencies for both SNPs analyzed (NOS3, rs3918226 and GUCY1A3, rs7692387) do not show important differences among Europeans, Africans, and Asians, according to the dbSNP database (Sherry et al., 2001). Finally, an increase in the number of the studied SNPs, at least in the NOS3 gene, could also be worthy for examining the potential association of haplotypes with nitrite concentrations.
In conclusion, we found that the CT genotype and the T allele for NOS3 rs3918226 were more frequent in NP than in patients with GH, while the GA genotype and A allele for GUCY1A3 rs7692387 were more frequent in patients with PE than NP. Moreover, patients with GH carrying the CT+TT genotypes for NOS3 rs3918226 showed lower nitrite concentrations than those with NP carrying the CT+TT genotypes, and NP patients carrying the GG genotype for GUCY1A3 rs7692387 showed higher nitrite concentrations than patients with GH or with PE carrying the same GG genotype. However, we found no significant model of interaction among genotypes for these functional SNPs to be associated with GH or with PE. Our novel findings suggest that functional SNPs of NOS3 (rs3918226) and GUCY1A3 (rs7692387) may affect NO formation and be associated with hypertensive disorders of pregnancy.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by the Institutional Review Board at University Hospital at the Ribeirao Preto Medical School of University of Sao Paulo (FMRP/USP). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
DP: formal analysis, investigation, methodology, writing–original draft, and writing–review and editing. ML: formal analysis, investigation, methodology, writing–original draft, and writing–review and editing. AP: data curation, funding acquisition, methodology, project administration, writing–original draft, and writing–review and editing. JT-S: data curation, funding acquisition, methodology, project administration, and writing–review and editing. RC: conceptualization, data curation, funding acquisition, methodology, project administration, and writing–review and editing. VS: conceptualization, data curation, funding acquisition, methodology, project administration, writing–original draft, and writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the National Council for Scientific and Technological Development (CNPq/Brazil; grants #314486/2023-2 and #308504/2021-6), the São Paulo Research Foundation (FAPESP/Brazil; grants #2019/07230-8 and #2021/12010-7), the Coordination for the Improvement of Higher Education Personnel (CAPES/Brazil) (finance code 001), and the National Institutes of Health (NIH/USA; grants K01HL159032, R01HL148191, and U54GM115428).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2024.1293082/full#supplementary-material
References
Acog (2013). Hypertension in pregnancy. Report of the American college of obstetricians and gynecologists' task force on hypertension in pregnancy. Obstet. Gynecol. 122 (5), 1122–1131. doi:10.1097/01.AOG.0000437382.03963.88
Ahmed, R., Dunford, J., Mehran, R., Robson, S., and Kunadian, V. (2014). Pre-eclampsia and future cardiovascular risk among women: a review. J. Am. Coll. Cardiol. 63, 1815–1822. doi:10.1016/j.jacc.2014.02.529
Chappell, L. C., Cluver, C. A., Kingdom, J., and Tong, S. (2021). Pre-eclampsia. Lancet 398, 341–354. doi:10.1016/S0140-6736(20)32335-7
Consortium, G. T. (2015). Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: multitissue gene regulation in humans. Science 348, 648–660. doi:10.1126/science.1262110
De Miranda, J. A., Lacchini, R., Belo, V. A., Lanna, C. M., Sertorio, J. T., Luizon, M. R., et al. (2015). The effects of endothelial nitric oxide synthase tagSNPs on nitrite levels and risk of hypertension and obesity in children and adolescents. J. Hum. Hypertens. 29, 109–114. doi:10.1038/jhh.2014.48
Denninger, J. W., and Marletta, M. A. (1999). Guanylate cyclase and the.NO/cGMP signaling pathway. Biochim. Biophys. Acta 1411, 334–350. doi:10.1016/s0005-2728(99)00024-9
Emdin, C. A., Khera, A. V., Klarin, D., Natarajan, P., Zekavat, S. M., Nomura, A., et al. (2018). Phenotypic consequences of a genetic predisposition to enhanced nitric oxide signaling. Circulation 137, 222–232. doi:10.1161/CIRCULATIONAHA.117.028021
Forstermann, U., Closs, E. I., Pollock, J. S., Nakane, M., Schwarz, P., Gath, I., et al. (1994). Nitric oxide synthase isozymes. Characterization, purification, molecular cloning, and functions. Hypertension 23, 1121–1131. doi:10.1161/01.hyp.23.6.1121
Gauderman, W. J. (2002). Sample size requirements for matched case-control studies of gene-environment interaction. Stat. Med. 21, 35–50. doi:10.1002/sim.973
Ghasemi, A., Zahedi Asl, S., Mehrabi, Y., Saadat, N., and Azizi, F. (2008). Serum nitric oxide metabolite levels in a general healthy population: relation to sex and age. Life Sci. 83, 326–331. doi:10.1016/j.lfs.2008.06.010
Gui, J., Andrew, A. S., Andrews, P., Nelson, H. M., Kelsey, K. T., Karagas, M. R., et al. (2011). A robust multifactor dimensionality reduction method for detecting gene-gene interactions with application to the genetic analysis of bladder cancer susceptibility. Ann. Hum. Genet. 75, 20–28. doi:10.1111/j.1469-1809.2010.00624.x
Honigberg, M. C., Chaffin, M., Aragam, K., Bhatt, D. L., Wood, M. J., Sarma, A. A., et al. (2020). Genetic variation in cardiometabolic traits and medication targets and the risk of hypertensive disorders of pregnancy. Circulation 142, 711–713. doi:10.1161/CIRCULATIONAHA.120.047936
Hutcheon, J. A., Lisonkova, S., and Joseph, K. S. (2011). Epidemiology of pre-eclampsia and the other hypertensive disorders of pregnancy. Best. Pract. Res. Clin. Obstet. Gynaecol. 25, 391–403. doi:10.1016/j.bpobgyn.2011.01.006
Kessler, T., Wobst, J., Wolf, B., Eckhold, J., Vilne, B., Hollstein, R., et al. (2017). Functional characterization of the GUCY1A3 coronary artery disease risk locus. Circulation 136, 476–489. doi:10.1161/CIRCULATIONAHA.116.024152
Li, R., Lyn, D., Lapu-Bula, R., Oduwole, A., Igho-Pemu, P., Lankford, B., et al. (2004). Relation of endothelial nitric oxide synthase gene to plasma nitric oxide level, endothelial function, and blood pressure in African Americans. Am. J. Hypertens. 17, 560–567. doi:10.1016/j.amjhyper.2004.02.013
Lo, C. C. W., Lo, A. C. Q., Leow, S. H., Fisher, G., Corker, B., Batho, O., et al. (2020). Future cardiovascular disease risk for women with gestational hypertension: a systematic review and meta-analysis. J. Am. Heart Assoc. 9, e013991. doi:10.1161/JAHA.119.013991
Loscalzo, J. (2018). Nitric oxide signaling and atherothrombosis redux: evidence from experiments of nature and implications for therapy. Circulation 137, 233–236. doi:10.1161/CIRCULATIONAHA.117.032901
Luizon, M. R., Metzger, I. F., Lacchini, R., and Tanus-Santos, J. E. (2012). Endothelial nitric oxide synthase polymorphism rs3918226 associated with hypertension does not affect plasma nitrite levels in healthy subjects. Hypertension 59, e52. author reply e53. doi:10.1161/HYPERTENSIONAHA.112.194654
Luizon, M. R., Palei, A. C., Sandrim, V. C., Amaral, L. M., Machado, J. S., Lacchini, R., et al. (2014). Tissue inhibitor of matrix metalloproteinase-1 polymorphism, plasma TIMP-1 levels, and antihypertensive therapy responsiveness in hypertensive disorders of pregnancy. Pharmacogenomics J. 14, 535–541. doi:10.1038/tpj.2014.26
Luizon, M. R., Palei, A. C. T., Belo, V. A., Amaral, L. M., Lacchini, R., Duarte, G., et al. (2017). Gene-gene interactions in the NAMPT pathway, plasma visfatin/NAMPT levels, and antihypertensive therapy responsiveness in hypertensive disorders of pregnancy. Pharmacogenomics J. 17, 427–434. doi:10.1038/tpj.2016.35
Luizon, M. R., Pereira, D. A., and Sandrim, V. C. (2018). Pharmacogenomics of hypertension and preeclampsia: focus on gene-gene interactions. Front. Pharmacol. 9, 168. doi:10.3389/fphar.2018.00168
Maynard, S. E., Min, J. Y., Merchan, J., Lim, K. H., Li, J., Mondal, S., et al. (2003). Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Invest. 111, 649–658. doi:10.1172/JCI17189
Mergia, E., Friebe, A., Dangel, O., Russwurm, M., and Koesling, D. (2006). Spare guanylyl cyclase NO receptors ensure high NO sensitivity in the vascular system. J. Clin. Invest. 116, 1731–1737. doi:10.1172/JCI27657
Metzger, I. F., Luizon, M. R., Lacchini, R., Ishizawa, M. H., and Tanus-Santos, J. E. (2013). Effects of endothelial nitric oxide synthase tagSNPs haplotypes on nitrite levels in black subjects. Nitric Oxide 28, 33–38. doi:10.1016/j.niox.2012.10.002
Moncada, S., and Higgs, E. A. (2006). The discovery of nitric oxide and its role in vascular biology. Br. J. Pharmacol. 147 (Suppl. 1), S193–S201. doi:10.1038/sj.bjp.0706458
Muniz, L., Luizon, M. R., Palei, A. C., Lacchini, R., Duarte, G., Cavalli, R. C., et al. (2012). eNOS tag SNP haplotypes in hypertensive disorders of pregnancy. DNA Cell Biol. 31, 1665–1670. doi:10.1089/dna.2012.1768
Pander, J., Wessels, J. A., Mathijssen, R. H., Gelderblom, H., and Guchelaar, H. J. (2010). Pharmacogenetics of tomorrow: the 1 + 1 = 3 principle. Pharmacogenomics 11, 1011–1017. doi:10.2217/pgs.10.87
Powe, C. E., Levine, R. J., and Karumanchi, S. A. (2011). Preeclampsia, a disease of the maternal endothelium: the role of antiangiogenic factors and implications for later cardiovascular disease. Circulation 123, 2856–2869. doi:10.1161/CIRCULATIONAHA.109.853127
Ritchie, M. D., and Motsinger, A. A. (2005). Multifactor dimensionality reduction for detecting gene-gene and gene-environment interactions in pharmacogenomics studies. Pharmacogenomics 6, 823–834. doi:10.2217/14622416.6.8.823
Rousset, F. (2008). genepop'007: a complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 8, 103–106. doi:10.1111/j.1471-8286.2007.01931.x
Salvi, E., Kutalik, Z., Glorioso, N., Benaglio, P., Frau, F., Kuznetsova, T., et al. (2012). Genomewide association study using a high-density single nucleotide polymorphism array and case-control design identifies a novel essential hypertension susceptibility locus in the promoter region of endothelial NO synthase. Hypertension 59, 248–255. doi:10.1161/HYPERTENSIONAHA.111.181990
Salvi, E., Kuznetsova, T., Thijs, L., Lupoli, S., Stolarz-Skrzypek, K., D'avila, F., et al. (2013). Target sequencing, cell experiments, and a population study establish endothelial nitric oxide synthase (eNOS) gene as hypertension susceptibility gene. Hypertension 62, 844–852. doi:10.1161/HYPERTENSIONAHA.113.01428
Sandrim, V. C., Palei, A. C., Cavalli, R. C., Araujo, F. M., Ramos, E. S., Duarte, G., et al. (2008a). eNOS haplotypes associated with gestational hypertension or preeclampsia. Pharmacogenomics 9, 1467–1473. doi:10.2217/14622416.9.10.1467
Sandrim, V. C., Palei, A. C., Metzger, I. F., Gomes, V. A., Cavalli, R. C., and Tanus-Santos, J. E. (2008b). Nitric oxide formation is inversely related to serum levels of antiangiogenic factors soluble fms-like tyrosine kinase-1 and soluble endogline in preeclampsia. Hypertension 52, 402–407. doi:10.1161/HYPERTENSIONAHA.108.115006
Sandrim, V. C., Palei, A. C., Sertorio, J. T., Cavalli, R. C., Duarte, G., and Tanus-Santos, J. E. (2010). Effects of eNOS polymorphisms on nitric oxide formation in healthy pregnancy and in pre-eclampsia. Mol. Hum. Reprod. 16, 506–510. doi:10.1093/molehr/gaq030
Sherry, S. T., Ward, M. H., Kholodov, M., Baker, J., Phan, L., Smigielski, E. M., et al. (2001). dbSNP: the NCBI database of genetic variation. Nucleic Acids Res. 29, 308–311. doi:10.1093/nar/29.1.308
Singhal, P., Verma, S. S., and Ritchie, M. D. (2023). Gene interactions in human disease studies-evidence is mounting. Annu. Rev. Biomed. Data Sci. 6, 377–395. doi:10.1146/annurev-biodatasci-102022-120818
Vanhoutte, P. M. (2003). Endothelial control of vasomotor function: from health to coronary disease. Circ. J. 67, 572–575. doi:10.1253/circj.67.572
Veiga, E. C. A., Rocha, P. R. H., Caviola, L. L., Cardoso, V. C., Costa, F. D. S., Saraiva, M., et al. (2021). Previous preeclampsia and its association with the future development of cardiovascular diseases: a systematic review and meta-analysis. Clin. (Sao Paulo) 76, e1999. doi:10.6061/clinics/2021/e1999
Williams, P. J., and Broughton Pipkin, F. (2011). The genetics of pre-eclampsia and other hypertensive disorders of pregnancy. Best. Pract. Res. Clin. Obstet. Gynaecol. 25, 405–417. doi:10.1016/j.bpobgyn.2011.02.007
Wobst, J., Schunkert, H., and Kessler, T. (2018). Genetic alterations in the NO-cGMP pathway and cardiovascular risk. Nitric Oxide 76, 105–112. doi:10.1016/j.niox.2018.03.019
Keywords: genetic polymorphisms, gestational hypertension, guanylate cyclase 1 soluble alpha 3, nitric oxide, nitric oxide synthase 3, preeclampsia, nitrite, pregnancy
Citation: Pereira DA, Luizon MR, Palei AC, Tanus-Santos JE, Cavalli RC and Sandrim VC (2024) Functional polymorphisms of NOS3 and GUCY1A3 affect both nitric oxide formation and association with hypertensive disorders of pregnancy. Front. Genet. 15:1293082. doi: 10.3389/fgene.2024.1293082
Received: 12 September 2023; Accepted: 12 February 2024;
Published: 26 February 2024.
Edited by:
Mohiuddin Mohammed Taher, Umm al-Qura University, Saudi ArabiaReviewed by:
Carlos Galaviz-Hernandez, National Polytechnic Institute (IPN), MexicoMaryam Pirhoushiaran, Tehran University of Medical Sciences, Iran
Marcos Antonio Fernandes Da Silva Gregnani, Federal University of São Paulo, Brazil
Copyright © 2024 Pereira, Luizon, Palei, Tanus-Santos, Cavalli and Sandrim. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcelo R. Luizon, mrluizon@ufmg.br; Valeria C. Sandrim, valeria.sandrim@unesp.br
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share senior authorship