 Max C. Keuken1,3
Max C. Keuken1,3 Harry B. M. Uylings2
Harry B. M. Uylings2 Stefan Geyer3
Stefan Geyer3 Andreas Schäfer3
Andreas Schäfer3 Robert Turner3
Robert Turner3 Birte U. Forstmann1*
Birte U. Forstmann1*- 1Cognitive Science Center Amsterdam, University of Amsterdam, Amsterdam, Netherlands
- 2Department of Anatomy and Neuroscience, VU University Medical Center, Amsterdam, Netherlands
- 3Department of Neurophysics, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany
The prevailing academic opinion holds that the subthalamic nucleus (STN) consists of three parts, each anatomically distinct and selectively associated with cognitive, emotional, or motor functioning. We independently tested this assumption by summarizing the results from 33 studies on STN subdivisions in human and nonhuman primates. The studies were conducted from 1925 to 2010 and feature three different techniques: electrical lesions, anterograde and retrograde tracers, and classical cytoarchitectonics. Our results reveal scant evidence in support of a tripartite STN. Instead, our results show that the variability across studies is surprisingly large, both in the number of subdivisions and in their anatomical localization. We conclude that the number of subdivisions in the STN remains uncertain, and that academic consensus in support of a tripartite STN is presently unwarranted.
Introduction
The subthalamic nucleus (STN) is a small but vitally important structure in the basal ganglia. Because of its central role in motor control, the STN is the target of deep-brain stimulation that alleviates severe motor symptoms in many levodopa-resistant patients with Parkinson's disease. Unfortunately, deep-brain stimulation of the STN also comes at a cost. In particular, some patients with Parkinson's disease experience a decline in cognitive functioning, whereas others may become depressed, experience hypersexuality, hypomania, and some even commit suicide (Burkhard et al., 2004; Temel et al., 2005).
These undesirable and perhaps unexpected side-effects of deep-brain stimulation suggest that the STN regulates not only motor behavior but also cognitive and emotional processes, a suggestion consistent with one of the most influential theories of the STN (Alexander and Crutcher, 1990). How does the STN carry out these separate duties? A simple and widely held assumption is that the STN is comprised of three anatomically distinct subparts, each of which is selectively connected to cognitive, limbic, and motor areas. An assumption, that seems to be supported by recent electrophysiological recordings in humans (Mallet et al., 2007; Greenhouse et al., 2011). To underscore the popularity of this assumption, we reviewed all 16 STN review articles published since 1985 (DeLong et al., 1985; Alexander et al., 1986, 1990; Alexander and Crutcher, 1990; Parent, 1990; Parent and Hazrati, 1993, 1995a,b; Joel and Weiner, 1997; Nakano, 2000; Rodriguez-Oroz et al., 2001; Hamani et al., 2004; Temel et al., 2005; Obeso et al., 2008; Rodriguez-Oroz et al., 2009; Krack et al., 2010) and found that 12 (75%) of these articles suggested that the STN is a tripartite system (DeLong et al., 1985; Parent, 1990; Parent and Hazrati, 1995a,b; Joel and Weiner, 1997; Nakano, 2000; Rodriguez-Oroz et al., 2001, 2009; Hamani et al., 2004; Temel et al., 2005; Obeso et al., 2008; Krack et al., 2010).
However, the assumption of a tripartite STN was at odds with our initial impression of the empirical literature. To test the empirical support for a tripartite STN, we therefore, conducted an independent and comprehensive literature review. In the first section below we summarize the results on the number of subdivisions in the STN, in the second section we also summarize the results on the presumed anatomical connectivity and localization of these subdivisions.
Number of Subdivisions in the STN
Following an extensive literature review we initially identified 43 studies, published between 1925 and 2010, that investigated anatomical connections via the use of tracers or lesions in the STN for human and nonhuman primates or used cytoarchitectonic studies of the STN for human and nonhuman primates (Foix and Nicolesco, 1925; Kodama, 1928; Whittier and Mettler, 1949; Nauta and Mehler, 1966; Carpenter and Strominger, 1967; Fussenich, 1967; Carpenter et al., 1968, 1981a,b; Petras, 1968; Carpenter and Peter, 1972; Kim et al., 1976; Kuzemenský, 1976; Rafols and Fox, 1976; Monakow et al., 1978; Nauta and Cole, 1978; DeVito et al., 1980; DeVito and Anderson, 1982; Parent et al., 1984, 1989; Smith and Parent, 1986; Parent and Smith, 1987; Mizuno et al., 1988; Nakano et al., 1990; Smith et al., 1990, 1994; Hazrati and Parent, 1991, 1992a,b; Sadikot et al., 1992; Lavoie and Parent, 1994; Nambu et al., 1996, 1997; Shink et al., 1996; Takada et al., 1996, 2001; Inase et al., 1999; Sato et al., 2000; Karachi et al., 2004; Kelly and Strick, 2004; Miyachi et al., 2006; Bostan et al., 2010; Rico et al., 2010).
With respect to the animal studies, we included studies only on nonhuman primates: just as in humans, the STN in nonhuman primates is a closed nucleus (i.e., dendrites are restricted to the nucleus), in contrast for example to the rat STN (Smith et al., 1990; Marani et al., 2008). With respect to the human studies, we chose only to include studies using cytoarchitectonics since this is still the golden standard in anatomy (Amunts et al., 2007) and seems to be more sensitive than cytochemical stainings in detecting different borders generally (Uylings et al., 2010; van de Werd et al., 2010).
From the initial group of 43 studies, we focused on a subset of 33 studies that explicitly consider the number of subdivisions in the STN (Foix and Nicolesco, 1925; Kodama, 1928; Whittier and Mettler, 1949; Nauta and Mehler, 1966; Fussenich, 1967; Kim et al., 1976; Kuzemenský, 1976; Rafols and Fox, 1976; Monakow et al., 1978; Nauta and Cole, 1978; Carpenter et al., 1981a,b; DeVito and Anderson, 1982; Parent et al., 1984, 1989; Smith and Parent, 1986; Parent and Smith, 1987; Nakano et al., 1990; Smith et al., 1990; Sadikot et al., 1992; Lavoie and Parent, 1994; Nambu et al., 1996, 1997; Shink et al., 1996; Inase et al., 1999; Sato et al., 2000; Takada et al., 2001; Karachi et al., 2004; Kelly and Strick, 2004; Miyachi et al., 2006; Bostan et al., 2010; Rico et al., 2010). These 33 studies were assigned to one of three categories, (A) 25 studies used anterograde and retrograde tracers to investigate possible STN subdivisions and their anatomical location in old-world and new-world monkeys (Carpenter et al., 1968; Kim et al., 1976; Monakow et al., 1978; Nauta and Cole, 1978; Carpenter et al., 1981a,b; DeVito and Anderson, 1982; Parent et al., 1984, 1989; Smith and Parent, 1986; Parent and Smith, 1987; Nakano et al., 1990; Smith et al., 1990; Sadikot et al., 1992; Lavoie and Parent, 1994; Nambu et al., 1996, 1997; Shink et al., 1996; Inase et al., 1999; Sato et al., 2000; Takada et al., 2001; Karachi et al., 2004; Kelly and Strick, 2004; Miyachi et al., 2006; Bostan et al., 2010; Rico et al., 2010), (B) two studies used electrical lesions in old-world monkeys (Nauta and Mehler, 1966; Carpenter et al., 1968), (C) six studies used classical cytoarchitectonics to study STN subdivisions by examining STN cell type, cell size, and cell density in human and nonhuman primates (Foix and Nicolesco, 1925; Kodama, 1928; Whittier and Mettler, 1949; Fussenich, 1967; Kuzemenský, 1976; Rafols and Fox, 1976). For each of these 33 studies, we summarized the data as supporting none, two, three, or four subdivisions in the STN. This classification reflected the opinion of the original authors of the studies we reviewed, and we did not attempt to reinterpret data or alter their conclusions in any way.
Table 1 summarizes the results. Overall, more than half of the studies we reviewed concluded that the STN has two subdivisions (61%), 21% concluded that the STN has no subdivisions, 12% concluded that the STN has three subdivisions, and 6% concluded that the STN has four subdivisions. These results contrast sharply with the popular academic opinion that the STN is a tripartite system. In addition, although the data most support the hypothesis that the STN has two subdivisions, the extent of this preference (i.e., 61% vs. 39% based on 33 studies) is not conclusive and the results appear to vary substantially from one study to the next.
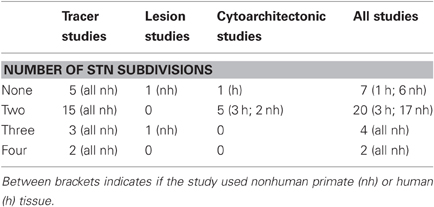
Table 1. Summary of 33 studies that examined the number of subdivisions in the STN for human and nonhuman primates.
The variability across studies may be partly due to methodological differences. In particular, it could be that results vary as a result of the specific tracer injection sites. It is also possible, however, that the general summary of results in Table 1 hides additional variability due to more detailed information regarding location and connectivity. For example, when researcher X argues that the STN has two subdivisions, one lateral and one medial, this claim is still inconsistent with that of researcher Y, who may also argue that the STN has two subdivisions, but believes that one is dorsal and the other ventral. The next section examines the connection profiles and STN subdivisions in relative detail.
Anatomical Connectivity and Localization of Subdivisions in the STN
We separately analyzed the results from each of 27 studies that used either tracing or electrical lesions to construct connection profiles and localize subdivisions in the STN. See mckitchen.files.wordpress.com/2010/01/supplementary-table-stn.pdf for a detailed overview for the methods, the number of animals and species used in the studies included in this review.
The results of these studies are summarized in Figure 1 and ordered by the number of subdivisions reported and the localization of the subdivisions within the STN. The subdivisions are represented on three schematic slices of the STN in rostro-caudal direction. This was done because individual studies could not be registered in one common anatomical space. Note that subdivisions were determined based both on statements derived from the individual studies and on figures contained in the articles.
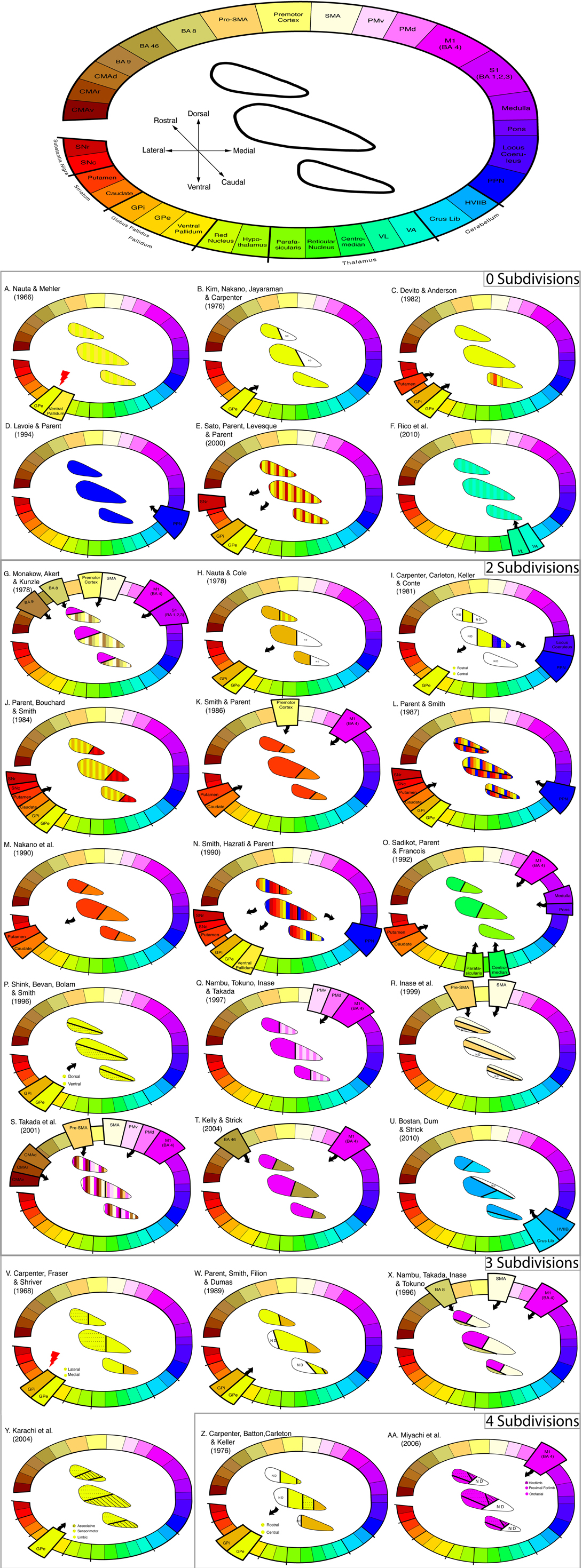
Figure 1. Connection profile and subdivisions of the STN show highly variable results. (Top of figure) Color-coded lesion, anterograde, and retrograde tracer injection sites including cortical, subcortical, and brainstem regions. In the middle, the anatomical orientation of the STN template is displayed. Three schematic slices from caudal to rostral are shown. Anatomical abbreviations: BA, Brodmann Area; CMAd, Cingulate Motor Area dorsal; CMAr, Cingulate Motor Area rostral; CMAv, Cingulate Motor Area ventral; GPe, Globus Pallidus external segment; GPi, Globus Pallidus internal segment; HVIIB, Gracial Lobule; M1, Primary Motor cortex, PMd, Pre-Motor dorsal; PMv, Pre-Motor ventral; PPN, Pedunculopontine Nucleus; Pre-SMA, Pre-Supplementary Motor Area; S1, Primary Sensory cortex; SMA, Supplementary Motor Area; SN, Substantia Nigra compacta; SNr, Substantia Nigra reticularis; VA, Ventral Anterior Nucleus; VL, Ventral Lateral Nucleus. (A–F) Six studies arguing for zero subdivisions. The author and publication year is shown in the left upper corner. (G–U) Fifteen studies arguing for two subdivisions. The author and publication year is shown in the left upper corner. (V–Y) Four studies arguing for three subdivisions. The author and publication year is shown in the left upper corner. (Z–AA) Two studies arguing for four subdivisions. The author and publication year is shown in the left upper corner. Black arrows denote the direction of connection. The red arrow denotes the location of a lesion. Localization of subdivisions within the STN are color coded according to the injection or lesion site.
To exemplify the kind of information visualized in Figure 1, consider the work by DeVito and Anderson (1982), who state: “Silver grains were distributed diffusely over the STN, with no apparent pattern to reflect an underlying anatomical organization (p. 110)”; as another example, Karachi et al. (2004) mention: “In summary, the pallidosubthalamic projection appears to be topographically arranged, with the sensorimotor part of the STN occupying its dorsolateral half, and the limbic part being restricted to its most anterior and medioventral portion (p. 178)”; finally, Miyachi et al. (2006) provide a figure (i.e., their Figure 6B) that presents the layout of subdivisions (see Table 2 for all the statements used to visualize all the included studies).
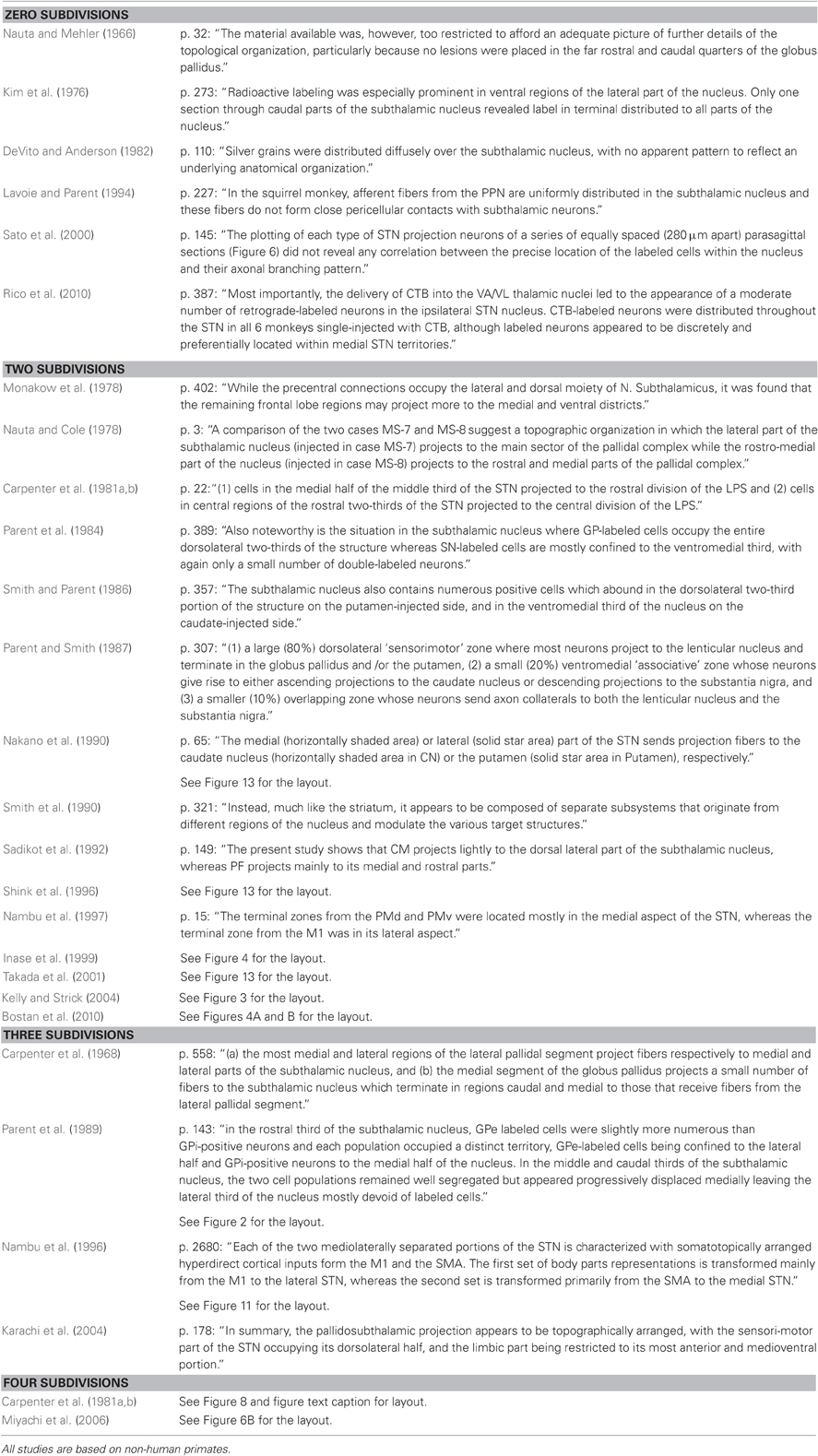
Table 2. The exact statement or figure number referring to the number of subdivisions derived from all tracings and lesions studies displayed in Figure 1.
One might argue that a temporal bias may be present, i.e., a lower number of subdivisions could have been found due to older methods and that more recent studies using advanced methodologies converge to a specific number of subdivisions. Figure 2A shows that this is not the case. Another possible bias is the species, i.e., new- or old-world monkey, and the type of tracer used. Again, no clear evidence is found for such a bias (Figure 2B).
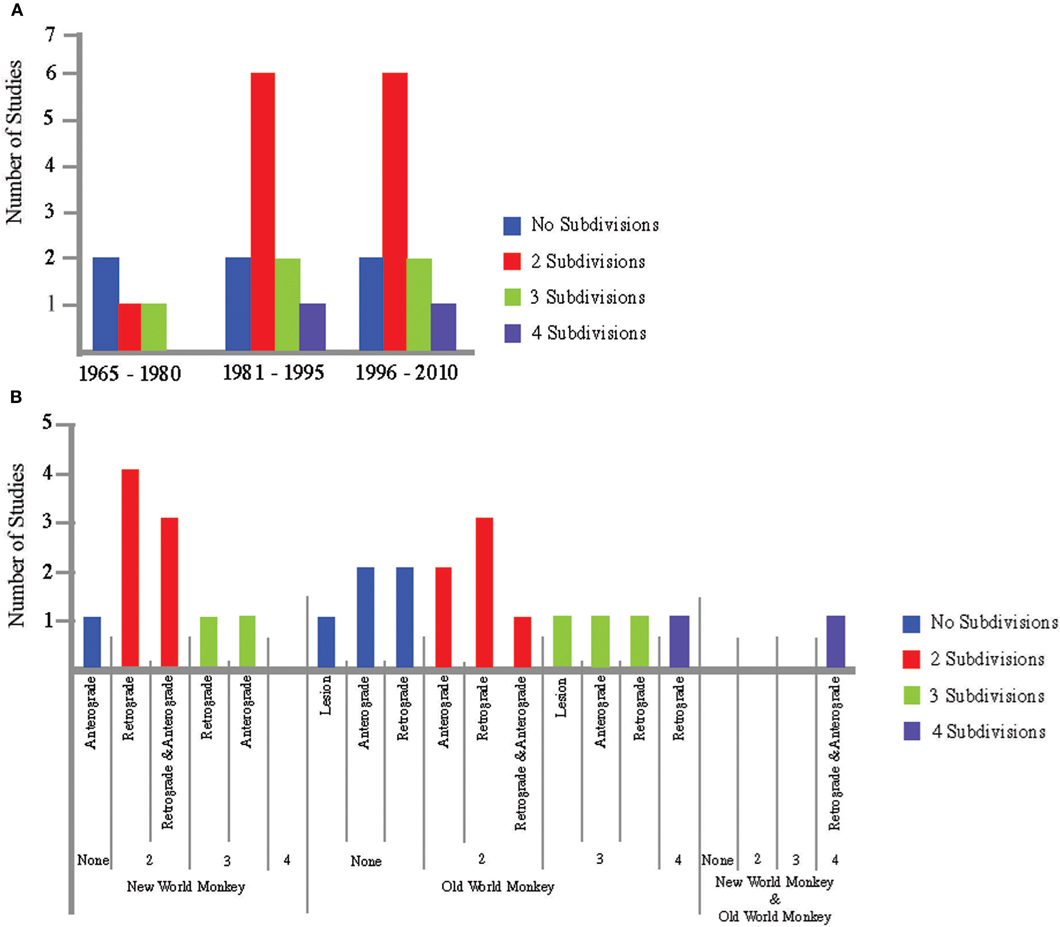
Figure 2. Reported number of subdivisions sorted by the year of publication, method and species. (A) Histogram including studies that argue for subdivisions in the STN published during the past 45 years, summarized in time-windows of 15 years. (B) Histogram including studies that argue for subdivisions in the STN separated for (a) the type of tracer used, and (b) the species under investigation. Color indicates the number of subdivisions reported. Blue bars indicate no subdivisions, red bars indicate two subdivisions, green bars indicate three subdivisions and purple bars indicate four subdivisions.
Two important conclusions can be drawn from this overview: (1) claims about the number of subdivisions in the STN do not depend on injection site in a clear and systematic way; (2) claims about the localization of subdivisions within the STN are highly variable. The variability of the results is clear from a visual inspection of Figure 1, but it is also apparent from other considerations; for instance, studies using the same subcortical injection sites such as the Globus Pallidus (Parent et al., 1989; Karachi et al., 2004) yield different localizations of subdivisions within the STN.
Based on this overview, there is a discrepancy between the empirical work and the general conclusions of the reviews published since 1985, as 75% of those reviews argue that there are three subdivisions, 6% conclude that there are four subdivisions, 13% state that there are five subdivisions, and finally 6% conclude that there are subdivisions in the STN but do not specify the number. Notably, during the past 26 years, no review paper has been published arguing in favor of none or two subdivisions in the STN despite the empirical studies that show, if at all, more evidence for two than three subdivisions.
It is also noteworthy, that the misconception of a tripartite STN could have been caused by misquotations. For instance, two review articles (Alexander et al., 1986, 1990) argue for five subdivisions, however, they have frequently been cited by others in favor of three subdivisions (e.g., Parent and Hazrati, 1995a,b). Such quotations are prone to yield a scientific consensus despite a yet to be answered question about the number of subdivisions in the STN.
Classical Cytoarchitectonic Studies in Human and Non-Human Primates
The classical cytoarchitectonic studies (i.e., studies depending on measures such as cell type, cell size, and cell density) are also inconsistent with the tripartite STN (Figure 3). We found six articles, including four with human specimens and two with old-world monkeys, published between 1925 and 1976. Five of these six studies reported two subdivisions in the STN (Foix and Nicolesco, 1925; Kodama, 1928; Whittier and Mettler, 1949; Fussenich, 1967; Rafols and Fox, 1976), clearly arguing for a heterogeneous neuronal distribution and one study with a human specimen arguing for no subdivisions (Kuzemenský, 1976). Although these results are less variable than the results of the tracer studies (80% of the cytoarchitectonic studies report the same number of subdivisions compared to 55% in the tracer and lesion studies), considerable variability regarding the localization of the subdivisions remains, as only three cytoarchitectonic studies show a comparable localization of the subdivisions within the STN.
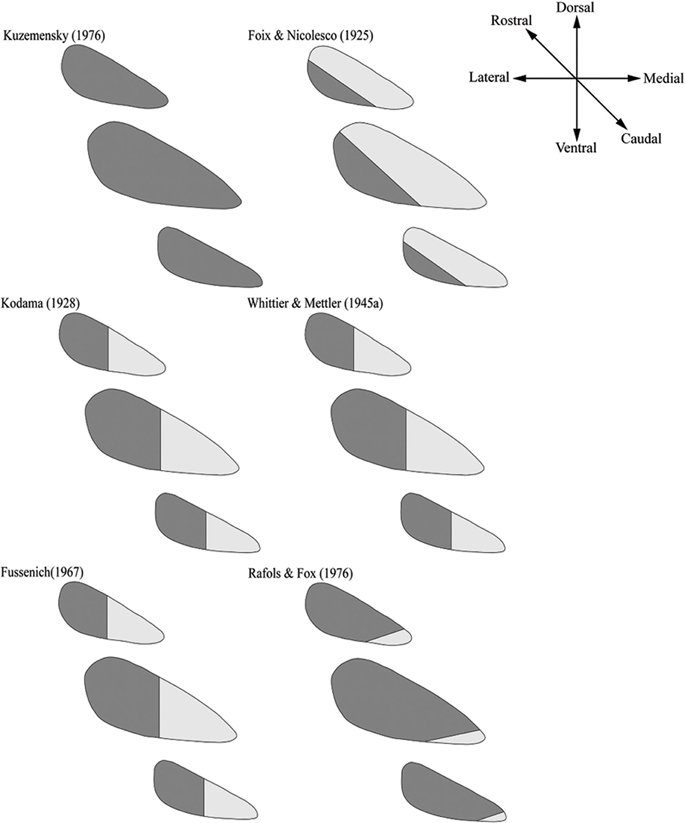
Figure 3. The variable location of subdivisions within the STN based on six individual studies using classical cytoarchitectonic features. Six human and nonhuman studies are shown that argue for either zero or two subdivisions. For each study, three schematic slices from caudal to rostral are displayed. Shaded areas refer to different subdivisions in the STN.
Conclusion
Our review of the literature sharply contrasts with the popular belief that the STN is a tripartite system (see Figure 1). In addition, our review shows that the variability across studies is surprisingly large—studies that support the same number of subdivisions almost always disagree on the localization of these subdivisions. A finding that cannot be explained by a potential bias regarding the publication year of the studies, methodological differences between studies, and the species under investigation (see Figure 2).
In the face of these results, it is difficult to explain why the belief in a tripartite STN continues to be as popular as it is. Moreover, Alexander et al. (1990) considered 5 parallel loop circuits in the basal ganglia and the present view considers 10 or more parallel circuits (e.g., Middleton and Strick, 2001) involving the STN. Therefore, one might speculate that the STN consist of more than three subdivisions. In sum, our review suggests that to better understand the anatomical structure of the STN further systematic studies are needed. We hope that future postmortem and in vivo studies using ultra-high resolution magnetic resonance imaging and histology will provide more definitive evidence regarding STN subdivisions and will help answer some important questions (Cho et al., 2010; Forstmann et al., 2010; Turner, 2011). Based on the empirical data that are presently available, firm conclusions about the number and localization of the STN are presently not warranted.
Outstanding Questions
- Are there subdivisions in the STN? If so, how many?
- What are the clinical implications of subdivisions in the STN? To what extent are nonhuman primate STN subdivisions comparable to in vivo human STN subdivisions?
- Are there individual differences in STN structural connectivity that predict differences in cognition?
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by the Max-Planck Society (Max C. Keuken, Stefan Geyer, Andreas Schäfer, Robert Turner, Birte U. Forstmann), by a VENI grant from the Netherlands Organization for Scientific Research (Birte U. Forstmann). This paper is part of the Research Priority Program “Brain and Cognition” at the University of Amsterdam (Max C. Keuken, Birte U. Forstmann). We thank E. J. Wagenmakers for constructive and detailed comments on a previous version of the manuscript.
References
Alexander, G. E., and Crutcher, M. D. (1990). Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci. 13, 266–271.
Alexander, G. E., Crutcher, M. D., and DeLong, M. R. (1990). Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. Prog. Brain Res. 85, 119–146.
Alexander, G. E., DeLong, M. R., and Strick, P. L. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9, 357–381.
Amunts, K., Schleicher, A., and Zilles, K. (2007). Cytoarchitecture of the cerebral cortex- more than localization. Neuroimage 37, 1061–1065.
Bostan, A. C., Dum, R. P., and Strick, P. L. (2010). The basal ganglia communicate with the cerebellum. Proc. Natl. Acad. Sci. U.S.A. 107, 8452–8456.
Burkhard, P. R., Vingerhoets, F. J. G., Berney, A., Bogousslavsky, J., Villemure, J.-G., and Ghika, J. (2004). Suicide after successful deep brain stimulation for movement disorders. Neurology 63, 2170–2172.
Carpenter, M. B., Batton, R. R., Carleton, S. C., and Keller, J. T. (1981a). Interconnections and organization of pallidal and subthalamic nucleus neurons in the monkey. J. Comp. Neurol. 197, 579–603.
Carpenter, M. B., Carleton, S. C., Keller, J. T., and Conte, P. (1981b). Connections of the subthalamic nucleus in the monkey. Brain Res. 224, 1–29.
Carpenter, M. B., Fraser, R. A. R., and Shriver, J. E. (1968). The organization of pallidosubthalamic fibers in the monkey. Brain Res. 11, 522–559.
Carpenter, M. B., and Peter, P. (1972). Nigrostriatal and nigrothalamic fibers in the rhesus monkey. J. Comp. Neurol. 144, 93–115.
Carpenter, M. B., and Strominger, N. L. (1967). Efferent fibers of the subthalamic nucleus in the monkey. A comparison of the efferent projections of the subthalamic nucleus, substantia nigra and globus pallidus. Am. J. Anat. 121, 41–72.
Cho, Z. H., Min, H. K., Oh, S. H., Han, J. Y., Park, C. W., Chi, J. G., Kim, Y. B., Paek, S. H., Lozano, A. M., and Lee, K. H. (2010). Direct visualization of deep brain stimulation targets in Parkinson disease with the use of 7-tesla magnetic resonance imaging. J. Neurosurg. 113, 639–647.
DeLong, M. R., Crutcher, M. D., and Georgopoulos, A. P. (1985). Primate globus pallidus and subthalamic nucleus: functional organization. J. Neurophysiol. 53, 530–543.
DeVito, J., and Anderson, M. (1982). An autoradiographic study of efferent connections of the Globus pallidus in Macaca mulatta. Exp. Brain Res. 46, 107–117.
DeVito, J. L., Anderson, M. E., and Walsh, K. E. (1980). A horseradish peroxidase study of afferent connections of globus pallidus in Macaca mulatta. Exp. Brain Res. 38, 65–73.
Foix, C., and Nicolesco, J. (1925). Les noyaux gris centraux et la région mesencéphalo-sous-optique. Paris: Masson.
Forstmann, B. U., Anwander, A., Schafer, A., Neumann, J., Brown, S., Wagemakers, J.-E., Bogacz, R., and Turner, R. (2010). Cortico-striatal connections predict control over speed and accuracy in perceptual decision making. Proc. Natl. Acad. Sci. U.S.A. 107, 15916–15920.
Fussenich, M. (1967). Vergleichend anatomische studien uber den nucleus subthalamicus (corpus Luys) bei primaten. Ph.D. thesis, University of Freiburg.
Greenhouse, I., Gould, S., Houser, M., Hicks, G., Gross, J., and Aron, A. R. (2011). Stimulation at dorsal and ventral electrode contacts targeted at the subthalamic nucleus has different effects on motor and emotion functions in Parkinson's Disease. Neuropsychologia 49, 528–534.
Hamani, C., Saint-Cyr, J. A., Fraser, J., Kaplitt, M., and Lozano, A. M. (2004). The subthalamic nucleus in the context of movement disorders. Brain 127, 4–20.
Hazrati, L. N., and Parent, A. (1991). Projection from the external pallidum to the reticular thalamic nucleus in the squirrel monkey. Brain Res. 550, 142–146.
Hazrati, L. N., and Parent, A. (1992a). Differential patterns of arborization of striatal and subthalamic fibers in the two pallidal segments in primates. Brain Res. 598, 311–315.
Hazrati, L. N., and Parent, A. (1992b). Convergence of subthalamic and striatal efferents at pallidal level in primates: an anterograde double-labeling study with biocytin and PHA-L. Brain Res. 569, 336–340.
Inase, M., Tokuno, H., Nambu, A., Akazawa, T., and Takada, N. (1999). Corticostriatal and corticosubthalamic input zones from the presupplementary motor area in the macaque monkey: comparison with the input zone from the supplementary motor area. Brain Res. 833, 191–201.
Joel, D., and Weiner, I. (1997). The connections of the primate subthalamic nucleus: indirect pathways and the open-interconnected scheme of basal ganglia-thalamocortical circuitry. Brain Res. Rev. 23, 62–78.
Karachi, C., Yelnik, J., Tande, D., Tremblay, L., Hirsch, E. C., and Francois, C. (2004). The pallidosubthalamic projection: an anatomical substrate for nonmotor functions of the subthalamic nucleus. Mov. Disord. 20, 172–180.
Kelly, R. M., and Strick, P. L. (2004). Macro-architecture of basal ganglia loops with the cerebral cortex: use of rabies virus to reveal multisynaptic circuits. Prog. Brain Res. 143, 449–459.
Kim, R., Nakano, K., Jayaraman, A., and Carpenter, M. B. (1976). Projections of the globus pallidus and adjacent structures: an autoradiographic study in the monkey. J. Comp. Neurol. 169, 263–289.
Kodama, S. (1928). Beitrage zur normalen anatomie des corpus luysi beim menschen. Arbeit. Anat. Inst. Sendai 13, 221–254.
Krack, P., Hariz, M. I., Baunez, C., Guridi, J., and Obeso, J. A. (2010). Deep brain stimulation: from neurology to psychiatry? Trends Neurosci. 33, 474–484.
Kuzemenský, J. (1976). Comparison of the cytoarchitectonic structure of the subthalamic nucleus in certain mammals. Folia Morphol. 24, 129–140.
Lavoie, B., and Parent, A. (1994). Pedunculopontine nucleus in the squirrel monkey: projections to the basal ganglia as revealed by anterograde tract-tracing methods. J. Comp. Neurol. 344, 210–231.
Mallet, L., Schupback, M., N'Diaye, K., Remy, P., Bardinet, E., Czerneckf, V., Welter, M.-L., Pelissolo, A., Ruberg, M., Agid, Y., and Yelnik, J. (2007). Stimulation of subterritories of the subthalamic nucleus reveals its role in the integration of emotional and motor aspects of behavior. Proc. Natl. Acad. Sci. U.S.A. 104, 10661–10666.
Marani, E., Heida, T., Lakke, E. A., and Usunoff, K. G. (2008). The Subthalamic Nucleus. Berlin: Springer.
Middleton, F. A., and Strick, P. L. (2001). “A revised neuroanatomy of frontal-subcortical circuits,” in Frontal-Subcortical Circuits in Psychiatric and Neurological Disorders, eds D. G. Lichter and J. L. Cummings (New York, NY: The Guildford Press), 44–58.
Miyachi, S., Lu, X., Imanishi, M., Sawada, K., Nambu, A., and Takada, N. (2006). Somatotopically arranged inputs from putamen and subthalamic nucleus to primary motor cortex. Neurosci. Res. 56, 300–308.
Mizuno, N., Ueyamana, T., Itoh, K., Satoda, T., Tashiro, T., and Shigemoto, R. (1988). Direct projections from the subthalamic nucleus of Luys to the spinal cord in the Japanese monkey. Neurosci. Lett. 89, 13–18.
Monakow, K. H., Akert, K., and Kunzle, H. (1978). Projections of the precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey. Exp. Brain Res. 33, 395–403.
Morel, A., Loup, F., Magnin, M., and Jeanmonod, D. (2002). Neurochemical organization of the human basal ganglia: anatomofunctional territories defined by the distributions of calcium-binding proteins and SMI-32. J. Comp. Neurol. 443, 86–103.
Nakano, K. (2000). Neural circuits and topographic organization of the basal ganglia and related regions. Brain Dev. 22, S5–S16.
Nakano, K., Hasegawa, Y., Tokushige, A., Nakagawa, S., Kayahara, T., and Mizuno, N. (1990). Topographical projections from the thalamus, subthalamic nucleus and pedunculopontine tegmental nucleus to the striatum in the Japanese monkey, Macaca fuscata. Brain Res. 537, 54–68.
Nambu, A., Takada, M., Inase, M., and Tokuno, H. (1996). Dual somatotopical representations in the primate subthalamic nucleus: evidence for ordered but reversed body-map transformations from the primary motor cortex and supplementary motor area. J. Neurosci. 16, 2671–2683.
Nambu, A., Tokuno, H., Inase, M., and Takada, M. (1997). Corticosubthalamic input zones from forelimb representations of the dorsal and ventral divisions of the premotor cortex in the macaque monkey: comparison with the input zones from the primary motor cortex and the supplementary motor area. Neurosci. Lett. 239, 13–16.
Nauta, H. J., and Cole, M. (1978). Efferent projections of the subthalamic nucleus: an autoradiographic study in monkey and cat. J. Comp. Neurol. 180, 1–16.
Nauta, W. J., and Mehler, W. R. (1966). Projections of the lentiform nucleus in the monkey. Brain Res. 1, 3–42.
Obeso, J. A., Rodriguez-Oroz, M. C., Benitez-Temino, B., Blesa, F. J., Guridi, J., Marin, C., and Rodriguez, M. (2008). Functional organization of the basal ganglia: therapeutic implications for Parkinson's disease. Mov. Disord. 23, S548–S559.
Parent, A., Bouchard, C., and Smith, Y. (1984). The striatopallidal and striatonigral projections: two distinct finber systems in primates. Brain Res. 303, 385–390.
Parent, A., and Hazrati, L. N. (1993). Anatomical aspects of information processing in primate basal ganglia. Trends Neurosci. 16, 111–116.
Parent, A., and Hazrati, L. N. (1995a). Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Res. Rev. 20, 91–127.
Parent, A., and Hazrati, L. N. (1995b). Functional anatomy of the basal ganglia. II. The place of subthalamic nucleus and external pallidum in basal ganglia circuitry. Brain Res. Rev. 20, 128–154.
Parent, A., and Smith, Y. (1987). Differential dopaminergic innervation of the two pallidal segments in the squirrel monkey (Saimiri sciures). Brain Res. 436, 296–310.
Parent, A., Smith, Y., Filion, M., and Dumas, J. (1989). Distinct afferents to internal and external pallidal segments in the squirrel monkey. Neurosci. Lett. 96, 140–144.
Petras, J. M. (1968). Some efferent connections of the motor and somatosensory cortex simian primates and felid canid and procyonid carnivores. Ann. N.Y. Acad. Sci. 167, 469–505.
Rafols, J. A., and Fox, C. A. (1976). The neurons in the primate subthalamic nucleus: a Golgi and electron microscopic study. J. Comp. Neurol. 168, 75–111.
Rico, A. J., Barroso-Chinea, P., Conte-Perales, L., Roda, E., Gomez-Bautista, V., Gendive, M., Obeso, J. A., and Lanciego, J. L. (2010). A direct projection from the subthalamic nucleus to the ventral thalamus in monkeys. Neurobiol. Dis. 39, 381–392.
Rodriguez-Oroz, M. C., Jahanshai, M., Krack, P., Litvan, I., Macias, R., Bezard, E., and Obeso, J. A. (2009). Initial clinical manifestations of Parkinson's disease: features and pathophysiological mechanisms. Lancet Neurol. 8, 1128–1139.
Rodriguez-Oroz, M. C., Rodriguez, M., Guridi, J., Mewes, K., Chckkman, V., Vitek, J., DeLong, M. R., and Obeso, J. A. (2001). The subthalamic nucleus in Parkinson's disease: somatotopic organization and physiological characteristics. Brain 124, 1777–1790.
Sadikot, A. F., Parent, A., and Francois, C. (1992). Efferent connections of the centromedian and parafascicular thalamic nuclei in the squirrel monkey: a PHA-L study of subcortical projections. J. Comp. Neurol. 315, 137–159.
Sato, F., Parent, M., Levesque, M., and Parent, A. (2000). Axonal branching pattern of neurons of the subthalamic nucleus in primates. J. Comp. Neurol. 424, 142–152.
Shink, E., Bevan, M. D., Bolam, J. P., and Smith, Y. (1996). The subthalamic nucleus and the external pallidum: two tightly interconnected structures that control the output of the basal ganglia in the monkey. Neuroscience 73, 335–357.
Smith, Y., Hazrati, L. N., and Parent, A. (1990). Efferent projections of the subthalamic nucleus in the squirrel monkey as studied by the PHA-L anterograde tracing method. J. Comp. Neurol. 294, 306–323.
Smith, Y., and Parent, A. (1986). Differential connections of caudate nucleus and putamen in the squirrel monkey (Saimiri sciureus). Neuroscience 18, 347–371.
Smith, Y., Wichmann, T., and DeLong, M. R. (1994). Synaptic innervation of neurones in the internal pallidal segment by the subthalamic nucleus and the external pallidum in monkeys. J. Comp. Neurol. 343, 297–318.
Takada, M., Tokuno, H., Hamada, I., Inase, M., Ito, Y., Imanishi, M., Hasegawa, N., Akazawa, T., Hatanaka, N., and Nambu, A. (2001). Organization of inputs form cingulate motor areas to basal ganglia in macaque monkey. Eur. J. Neurosci. 14, 1633–1650.
Takada, M., Tokuno, H., and Inase, M. (1996). Direct projections from the presupplementary motor area (pre-SMA) to the basal ganglia in the Japanese monkey. Neuroscience 25, S179.
Temel, Y., Blokland, A., Steinbusch, H. W. M., and Visser-Vandewalle, V. (2005). The functional role of the subthalamic nucleus in cognitive and limbic circuits. Prog. Neurobiol. 76, 393–413.
Turner, R. (2011). “Applications of high-field MRI in humans,” in High-Field MR Imaging. (New York, NY: Springer), 137–149.
Uylings, H. B. M., Sanz-Arigita, E. J., de Vos, K., Pool, C. W., Evers, P., and Rajkowska, G. (2010). 3-D cytoarchitectonic parcellation of human orbitofrontal cortex correlation with postmortem MRI. Psychiatr. Res. 183, 1–20.
van de Werd, H. J. J. M., Rajkowska, G., Evers, P., and Uylings, H. B. M. (2010). Cytoarchitectonic and chemoarchitectonic characterization of the prefrontal cortical areas in the mouse. Brain Struct. Funct. 214, 339–353.
Keywords: subthalamic nucleus, subdivisions, primate
Citation: Keuken MC, Uylings HBM, Geyer S, Schäfer A, Turner R and Forstmann BU (2012) Are there three subdivisions in the primate subthalamic nucleus?. Front. Neuroanat. 6:14. doi: 10.3389/fnana.2012.00014
Received: 22 February 2012; Accepted: 24 April 2012;
Published online: 10 May 2012.
Edited by:
Jose L. Lanciego, University of Navarra, SpainReviewed by:
Richard S. Nowakowski, Florida State University, USAAtsushi Nambu, National Institute for Physiological Sciences, Japan
Copyright: © 2012 Keuken, Uylings, Geyer, Schäfer, Turner and Forstmann. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Birte U. Forstmann, Cognitive Science Center Amsterdam, University of Amsterdam, Plantage Muidergracht 24, 1018 TV Amsterdam, Netherlands. e-mail:YnVmb3JzdG1hbm5AZ21haWwuY29t