 Selvin Z. Reyes-Garcia1,2
Selvin Z. Reyes-Garcia1,2 Antônio-Carlos Guimarães de Almeida3
Antônio-Carlos Guimarães de Almeida3 Nancy N. Ortiz-Villatoro1
Nancy N. Ortiz-Villatoro1 Fulvio A. Scorza1
Fulvio A. Scorza1 Esper A. Cavalheiro1
Esper A. Cavalheiro1 Carla A. Scorza1*
Carla A. Scorza1*- 1Disciplina de Neurociência, Departamento de Neurologia e Neurocirurgia, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil
- 2Departamento de Ciencias Morfológicas, Facultad de Ciencias Médicas, Universidad Nacional Autónoma de Honduras, Tegucigalpa, Honduras
- 3Laboratório de Neurociência Experimental e Computacional, Departamento de Engenharia de Biossistemas, Universidade Federal de São João del-Rei, São João del-Rei, Brazil
Background: Diverse forms of long-term potentiation (LTP) have been described, but one of the most investigated is encountered in the glutamatergic synapses of the hippocampal cornu Ammonis (CA1) subfield. However, little is known about synaptic plasticity in wildlife populations. Laboratory animals are extremely inbred populations that have been disconnected from their natural environment and so their essential ecological aspects are entirely absent. Proechimys are small rodents from Brazil’s Amazon rainforest and their nervous systems have evolved to carry out specific tasks of their unique ecological environment. It has also been shown that long-term memory duration did not persist for 24-h in Proechimys, in contrast to Wistar rats, when both animal species were assessed by the plus-maze discrimination avoidance task and object recognition test.
Methods: In this work, different protocols, such as theta burst, single tetanic burst or multiple trains of high frequency stimulation (HFS), were used to induce LTP in hippocampal brain slices of Proechimys and Wistar rats.
Results: A protocol-independent fast decay of early-phase LTP at glutamatergic synapses of the CA1 subfield was encountered in Proechimys. Long-term depression (LTD) and baseline paired-pulse facilitation (PPF) were investigated but no differences were found between animal species. Input/output (I/O) relationships suggested lower excitability in Proechimys in comparison to Wistar rats. Bath application of d-(-)-2-amino-5-phosphonopentanoicacid (D-AP5) and CNQX prevented the induction of LTP in both Proechimys and Wistar. However, in marked contrast to Wistar rats, LTP induction was not facilitated by the GABAA antagonist in the Amazon rodents, even higher concentrations failed to facilitate LTP in Proechimys. Next, the effects of GABAA inhibition on spontaneous activity as well as evoked field potentials (FPs) were evaluated in CA1 pyramidal cells. Likewise, much lower activity was detected in Proechimys brain slices in comparison to those of the Wistar rats.
Conclusions: These findings suggest a possible high inhibitory tone in the CA1 network mediated by GABAA receptors in the Amazon rodents. Currently, neuroscience research still seeks to reveal molecular pathways that control learning and memory processes, Proechimys may prove useful in identifying such mechanisms in complement to traditional animal models.
Introduction
Synaptic transmission is the hub of neural communication. Essential for several brain functions, synaptic plasticity, a key attribute of synaptic transmission, is a dynamic modification in the efficacy of synaptic strength, converting information processing into neural circuits (Vitureira and Goda, 2013). By controlling the synaptic strength, glutamatergic and GABAergic neurotransmission systems are very plastic and ensure brain homeostatic regulation. In this context, it has remained an open question how one does conciliate the properties of a particular and accurately wired brain with the competence to acquire knowledge, learning and memory. The pioneer in introducing the concept of synapse specificity was Ramón y Cajal (1894) who proposed that learning would be the result of the modifications in the connection strength. The concept was later refined by Hebb (1949) and is frequently shortened to “neurons wire together if they fire together” (Löwel and Singer, 1992). The discovery of the phenomenon of long-lasting potentiation by Bliss and Lømo (1973) was the first description of synaptic plasticity. Since then, long-lasting types of synaptic plasticity discriminated as Hebbian plasticity (when presynaptic activity is strictly followed by postsynaptic activity), comprising, among others, long-term potentiation (LTP), have been largely investigated as attractive cellular mechanisms underlying some forms of learning and memory (Bliss and Lømo, 1973; Ito and Kano, 1982; Bliss and Collingridge, 1993; Collingridge et al., 2010; Bliss and Cooke, 2011; Lüscher and Malenka, 2012; Mayford et al., 2012). Diverse forms of LTP have been described, but one of the most investigated is encountered in the glutamatergic synapses of the hippocampal cornu Ammonis (CA1) subfield. However, little is known about synaptic plasticity in wildlife populations (Amrein, 2015). Laboratory animals are extremely inbred populations that have been disconnected from their natural environment and so their essential ecological aspects are entirely absent (Klaus and Amrein, 2012; Keifer and Summers, 2016). Comparative studies may add to our comprehension of the basic functions of the brain so the use of non-standard populations can be instructive in addition to traditional animal models (Keifer and Summers, 2016).
Proechimys are small rodents from Brazil’s Amazon rainforest and their nervous systems have evolved to carry out specific tasks of their unique ecological environment (Carlson, 2012; Keifer and Summers, 2016). Previous work of our group has showed that these wild rodents display a fast decaying LTP and poor long-lasting memory process (Guimarães Marques et al., 2018). It has also been shown that long-term memory duration did not persist for 24-h in Proechimys, in contrast to Wistar rats, when both animal species were assessed by the plus-maze discrimination avoidance task and object recognition test (Guimarães Marques et al., 2018). In this line, studies have suggested a link between the maintenance of LTP and long-term memory maintenance (Pastalkova et al., 2006; Sacktor, 2008). The decay of LTP is still a poorly understood process but dynamic network inhibition has been implicated in the loss of synaptic potentiation over time (Dong et al., 2015). In the present work, using different standard protocols for inducing LTP, we confirmed the protocol-independent rapid decay of the early-phase LTP in Proechimys’s glutamatergic synapses of the CA1 hippocampal subfield. Furthermore, our findings from the use of modulators of GABAergic and glutamatergic transmission suggested a high inhibitory tone in this CA1 network mediated by GABAA receptors in the wild Amazon rodents, which could play a role in the fast decay of LTP encountered in this study.
Materials and Methods
Animals
Male rodents derived from wild Proechimys trapped at the Brazilian Amazon forest were bred in a colony established at the Neuroscience Laboratory’s facility (Federal Technical Register-IBAMA number 1561643) of Escola Paulista de Medicina/Universidade Federal de São Paulo (EPM/UNIFESP). Male Wistar rats were bred in the biotery of the University EPM/UNIFESP (CEDEME). All adult animals used in the experiments, weighing 250–300 g, were housed under environmentally controlled conditions (22°C ± 1°C), 12/12 h light-dark cycle (lights on at 7 a.m), with water and food ad libitum. All animal procedures were carried out in accordance with the ethical and practical principles of the use of laboratory animals with the approval of the ethical committee of EPM/UNIFESP (CEUA 9108060315). Precautions were taken to minimize the number of animals used in the experiments.
Brain Slice Preparation
Brain slices were prepared as described by Guimarães Marques et al. (2018). Briefly, 1% isoflurane in 70% N2O and 30% O2 was used to anesthetize animals. Next, adult Wistar and Proechimys rodents were decapitated and brains immediately extracted. Brain were sliced in ice-cold artificial cerebrospinal fluid (aCSF) at 4°C ± 0.5°C temperature composed by (in mM) NaCl 129, NaHCO3 21, KCl 3, CaCl2 1.6, MgSO4 1.8, NaH2PO4 1.25 and glucose 10, saturated with 95% O2 and 5% CO2. Horizontal hippocampal slices were cut at 400 μm thick using a vibratome (LEICA VT 1200S). Slices were immediately transferred to an interface chamber perfused with aCSF at 36°C ± 0.5°C (flow rate: 1.5–2.0 ml/min, pH 7.4, osmolarity: 295–300 mOsmol/L) and recordings started 2–3 h after slice recovering.
In vitro Electrophysiology—Stimulation and Recording
Extracellular field potentials (FPs) were obtained from the stratum radiatum (field excitatory postsynaptic potentials, fEPSPs) and pyramidale population spike (PS) of CA1 subfield. Using a stimulating bipolar twisted electrode (Ni-Cr wire with 50 μm diameter tip) positioned in the stratum radiatum of the CA1 hippocampal region, orthodromic stimuli were delivered in the Schaffer collateral axons. The recording electrode was made by a chlorinated silver wire placed into a glass capillar filled with 154 mM NaCl (5–10 MΩ). Low-pass filtered (3 kHz) data were digitized at 10 KHz then stored on computer disk through a CED 1401 interface for off-line analysis utilizing Spike 2 v6.09 (CED-1401, Cambridge, UK). Prior to the LTP/long-term depression (LTD) induction, input/output (I/O) relationships of stimulus intensity against to fEPSPs magnitude were carried out to obtain the maximal amplitude subsequently to uniform rises of stimulus intensity up to no further augmentation of the fEPSP amplitude. Stimulus intensities were adjusted to elicit a fEPSP slope of 50%–60% of the maximum accessed from the I/O. I-O curves for fEPSP slope, fiber volley (FV) amplitude and PS amplitude were recorded before and after the induction of LTP by four high frequency stimulation (4HFS). Field EPSP slope was measured after the FV with the intention of avoiding the impact of other sources of current flow. Paired pulse facilitation (PPF) was assessed at interstimulus intervals (ISIs) of 20, 40, 80, 160 and 320 ms in both groups of animals. To PPF, 10 trials at each interval were assessed. PPF (%) was determined as the ratio between the second pulse-evoked of fEPSP and the first one. LTP was defined as alterations in fEPSP slope higher than 20%. The high frequency stimulus was applied after 20 min of stable baseline recording. LTP was induced by: (1) a single train of HFS, 100 Hz in one second; (2) by four trains of HFS with 20 s inter-train interval; and (3) theta burst stimulation (TBS) protocol consisted of 10 bursts repeated at 200 ms intervals, with four pulses at 100 Hz for each burst. All protocols were delivered to the Schaffer collaterals and responses were recorded for 80 min. LTP induction was assessed in different set of experiments by observing modifications in fEPSP after adding the NMDA receptor blocker d-(-)-2-amino-5-phosphonopentanoicacid (D-AP5; 100 μM; Tocris, Bristol, UK), CNQX disodium salt (10 μM; Tocris, Bristol, UK) or the GABAA receptor blockers bicuculline methiodide (10 μM, 30 μM, 60 μM; Sigma-Aldrich, St. Louis, MO, USA) and picrotoxin (0.1 mM, Research Biochemicals International, Natick, MA, USA) or the GABAA receptor positive allosteric modulator diazepam (1 μM, Compaz®, Cristália, Brazil), in the perfusion bath, 10 min before until 20 min after LTP induction. In the experiments with bicuculline and picrotoxin, the concentrations of Ca2+ and Mg2+ were increased to 4 mM to minimize polysynaptic and burst discharges (Chapman et al., 1998). In the experiments with CNQX, magnesium-free aCSF were used (Muller et al., 1988). Additionally, spontaneous as well as evoked field activity were both investigated through the effect of GABAA antagonism on brain slices using bicuculline (10 μM, 30 μM and 60 μM).
Statistical Analysis
Data were analyzed using custom-made scripts (©Jan-Oliver Hollnagel, MATLAB R2013b; Salar et al., 2016). The fEPSP slope was measured between 20% and 80% of its maximal amplitude, the amplitude of the PS was examined by measuring the difference between the negative peak and the middle point located between the line connecting the first and second positive peaks. In order to verify the potentiation after HFS, fEPSP slopes were normalized relative to the averaged baseline response. For statistical analysis, all data were reported as mean ± standard error of the mean (SEM). Statistical significance was assessed by non-parametric Wilcoxon Signed Rank test or Mann–Whitney U test. For PPF Friedman’s two-way analysis of variance with repeated measures was used. Sets of electrophysiological experiments with three groups were analyzed using Kruskal-Wallis test, followed by Dunn’s post hoc multiple comparisons test, p < 0.05 (*) was considered to indicate a significant difference.
Results
Hippocampal Synaptic Plasticity
Figure 1 shows the I/O relationships in the hippocampal CA1 area of the Proechimys (9/4, slices/animals) and Wistar (8/4, slices/animals) rodents. The I/O relationships were evaluated before and after the induction of LTP with 4HFS protocol. Measuring the FV amplitude and fEPSP slope at different stimulus intensity allows an evaluation of the presynaptic vs. postsynaptic response (Figures 1A,B). The FV magnitude represents the quantity of the activated presynaptic axons of the Schaffer collateral and so infers the strength of the afferent input while the fEPSP is representative of the postsynaptic depolarizing activity. Indeed, the ratio of fEPSP slope to FV amplitude at each input stimulation level enables to quantify the synaptic strength connection. When comparing this synaptic strength connection before and 80-min after 4HFS, Wistar rats (Figure 1B) presented potentiation starting at 20% of the maximal stimulus contrasting to Proechimys rodents (Figure 1A) that showed increased synaptic transmission only after 40% of the maximal stimulus, suggesting that stronger stimulation is required for synaptic excitatory transmission efficacy in the Neotropical rodents. As the stimulus intensity increased, in Proechimys, the fEPSP slope and PS amplitude were increased only after 40%–60% and 60%–80% of the maximal stimulus intensities, respectively (Figures 1C,E); however, in Wistar rats, the increases in the fEPSP slopes were observed for all intensities of stimulation whilst increases in the PS amplitude occurred only after 40%–60% of the maximal stimulus intensities (Figures 1D,F). By assuming these parameters as a measure of excitability, our data suggests lower excitability in the CA1 network of the Proechimys. In order to compare different animal species, a useful approach is to assess the synaptic strength connection (ratio of fEPSP slope to FV amplitude; Sweatt, 2010; Figures 1G,H). To do so, same collection of data used on Figures 1A,B were used to perform comparative analysis that produced the Figures 1G,H. The only difference between Proechimys and Wistar rats was observed at maximum stimulus intensity after HFS, suggesting an augmentation of synaptic transmission in Wistar rats (Figure 1H). Detailed statistical data are showed in the legend of Figure 1. The PPF was assessed in slices of both animal species. Friedman’s two-way analysis of variance with repeated measures was used on the ISIs showed significant main effects of intervals, but neither differences nor interactions between groups were found. In both animal species, the maximum PPF was observed at an interval of 40 ms, 162.7 ± 2.5% in Proechimys and 159.7 ± 2.4% in Wistar rats (Figure 2A).
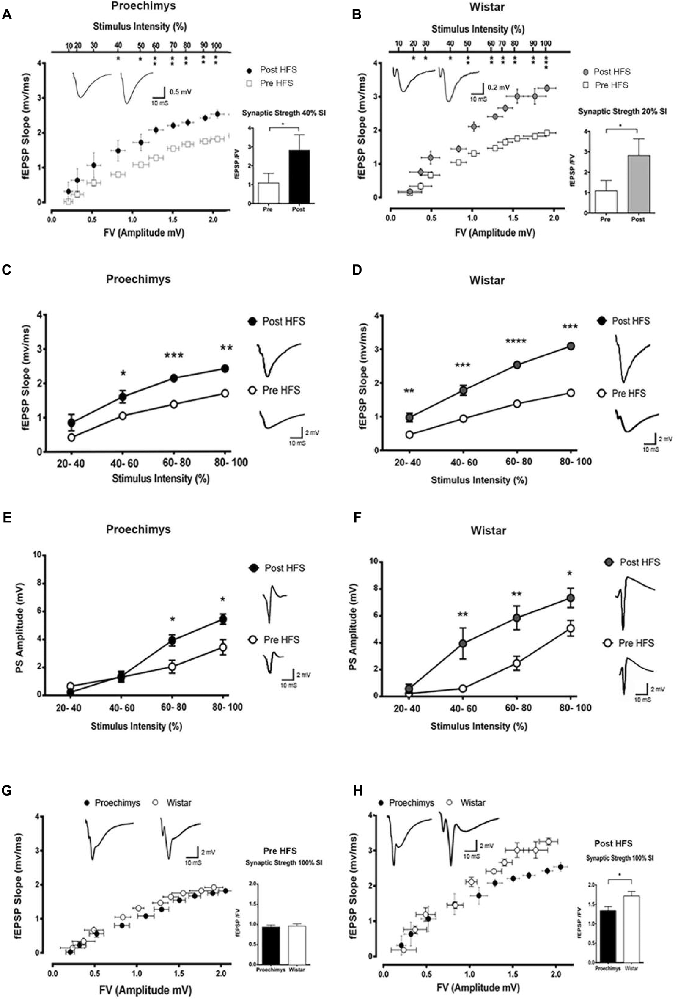
Figure 1. Input/output (I/O) relationships in Proechimys and Wistar rodents before and after four high frequency stimulation (4HFS). (A,B) Analysis of fiber volley (FV) amplitude vs. field excitatorypostsynaptic potential (fEPSP) slope at each level of stimulus intensity (% of the maximum). Insets, average and recordings of synaptic strength connection (fEPSP/FV amplitude). (A) In Proechimys, potentiation started at 40% of maximal stimulus intensity (MSI; W = 16, *p = 0.0469). (B) In Wistar, at 20% of MSI, (W = 21, *p = 0.0156). (C,D) Stimulus intensity vs. fEPSP slope. (C) In Proechimys, 40%–100% of MSI resulted in enhancement of fEPSP slope after 4HFS; for example, for 40%–60% of MSI: 1.048 ± 0.09 mV/ms pre-4HFS and 1.604 ± 0.18 mV/ms post-4HFS (W = 34, *p = 0.0078). (D) In Wistar, 20%–100% of MSI resulted in enhancement of fEPSP slope after 4HFS, example at 20%–40% of MSI: 0.47 ± 0.09 mV/ms pre-4HFS and 0.98 ± 0.13 mV/ms post-4HFS (W = 50, **p = 0.0237). Insets in (C,D) are examples of fEPSP recorded at 40%–60% of stimulus intensity pre and post 4HFS. (E,F) Stimulus intensity vs. population spike (PS) amplitude. (E) In Proechimys, enhancement in pyramidal cell firing was found at 60%–100% of MSI, example of amplitude at 60%–80% of MSI: 2.05 ± 0.46 mV pre-4HFS and 3.93 ± 0.39 mV post-4HFS, Wilcoxon test (W = 58, *p = 0.0105). (F) In Wistar, differences were found at 40%–100% of MSI, example at 40%–60% of MSI: 0.59 ± 0.20 mV pre-4HFS and 3.94 ± 1.16 mV post-4HFS, Wilcoxon test (W = 51, **p = 0.0059). Insets are example or PS recordings at 60%–80% of MSI. (G,H) Comparison of synaptic strength between both animal species. (G) No differences were encountered between Proechimys and Wistar before tetanic stimulation. (H) After 4HFS, higher enhancement was found only at 100% of MSI when comparing Proechimys and Wistar: 1.34 ± 0.10 and 1.72 ± 0.11 mV/ms.mV, respectively (U = 0, *p = 0.0153). Inset, the average of strength connection at 100% of stimulus intensity and the correspondent fEPSP. Time and amplitude of recordings are given by calibration bars on the right. *< 0.05; **≤ 0.01; ***≤ 0.001; ****≤ 0.0001.
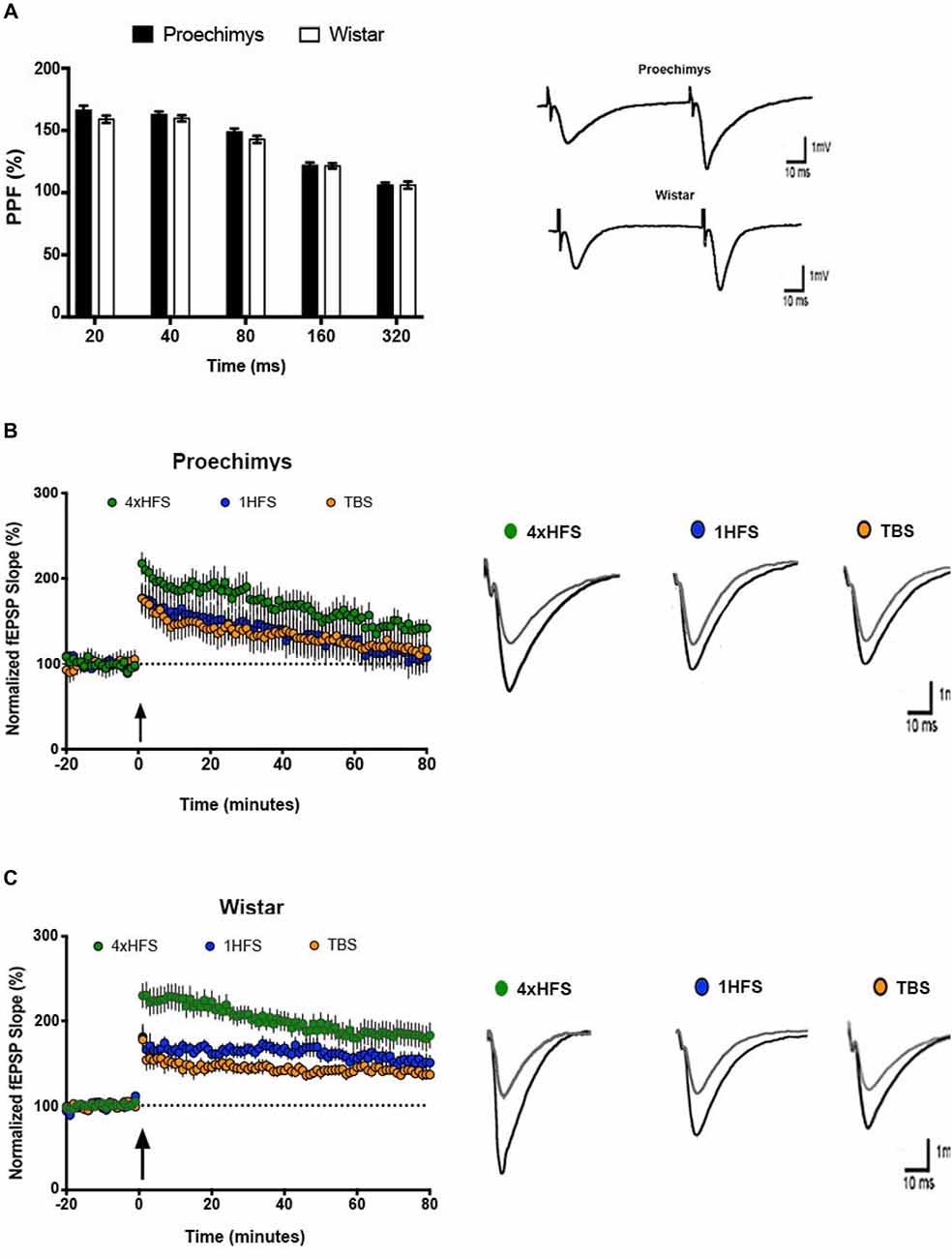
Figure 2. Short and long-term plasticity in Proechimys and Wistar rodents. (A) Paired pulse facilitation (PPF) at intervals of 20–320 ms. The maximum PPF was observed at an interstimulus intervals (ISIs) of 40 ms in both animal species, but neither differences nor interactions between groups were found. Insets are example or PPF recordings at 40 ms of MSI. White bars, PPF in Wistar group; black bars, PPF in Proechimys group at each ISI. Inset representative evoked field potentials (FPs) at interval of 40 ms. Time and amplitude of recordings are given by calibration bars on the right. (B,C) Long-term potentiation (LTP) was induced with three different tetanic stimulation protocols theta burst stimulation (TBS; 1HFS and 4HFS) delivered after 20 min of stable baseline recordings. (B) Time evolution of fEPSP slopes of Proechimys. Note that fEPSP slope potentiation decays through the time reaching baseline levels around 60-min after 1HFS and TBS. The gradual decay of potentiation was also observed after 4HFS, however this last stimulus elicited higher enhancement of potentiation and baseline levels were never reached during the 80-min recorded in our study. (C) Wistar showed enhanced and sustained potentiation with all protocols. Insets, representative superimposed recordings of averaged fEPSP before (gray trace) and 30 min after (black trace) tetanic stimulations in Proechimys (B) and Wistar (C). Time and amplitude of recordings are given by calibration bars on the right.
Long-Term Plasticity
Multiple protocols are available to induce LTP, 1HFS and TBS are commonly used. The potentiation at 30 min after 1HFS was 46.6 ± 16.3% and 67.8 ± 4.6% above baseline levels for Proechimys and Wistar, respectively. However, in Proechimys (8/4, slices/animals), fEPSP slope potentiation decayed over time reaching basal levels at 75-min after 1HFS stimulation (Figure 2B), contrasting to Wistar rats (8/4, slices/animals), in which potentiation was maintained during recordings (Figure 2C). Likewise, at 30-min after TBS, the potentiation in Proechimys was 34.3 ± 8.3% and in Wistar 42.7 ± 6.4%. Moreover, LTP decay was observed only in Proechimys. Therefore, we applied a stronger induction protocol (4HFS) in order to compare the magnitude of LTP produced in CA1 area of the hippocampal slices (4HFS vs. 1HFS/TBS; Figures 2B,C). The protocol of 4HFS induced a robust potentiation and it was chosen for all the experiments in this work. The post-tetanic potentiation after 30-min of 4HFS was 91.6 ± 21.3% and 107 ± 15.4% above baseline levels for Proechimys and Wistar, respectively. Our results showed that stronger stimulation was required to LTP maintenance in the wild rodents. Recordings were more stable over time in Wistar rats, however, in Proechimys the fEPSP slope dropped gradually even with 4HFS but LTP was maintained during the 80-min of experiment.
Effects of D-AP5, CNQX, Bicuculline, Picrotoxin and Diazepam on LTP
Since the decay of potentiation over time was observed in the slices of Proechimys, modulators of synaptic transmission were assessed only in LTP experiments. The D-AP5 was used to assess the effects of the NMDA-receptor mediated activity on LTP in Proechimys (12/5, slices/animals) and Wistar (12/5, slices/animals). Bath application of D-AP5 prevented the induction of LTP in both animal species, for example, fEPSP slope changes from baseline at 20 min after tetanic stimulation were 8.5 ± 4.6% in Proechimys and 12.4 ± 4.6% in Wistar (Figures 3A,B).
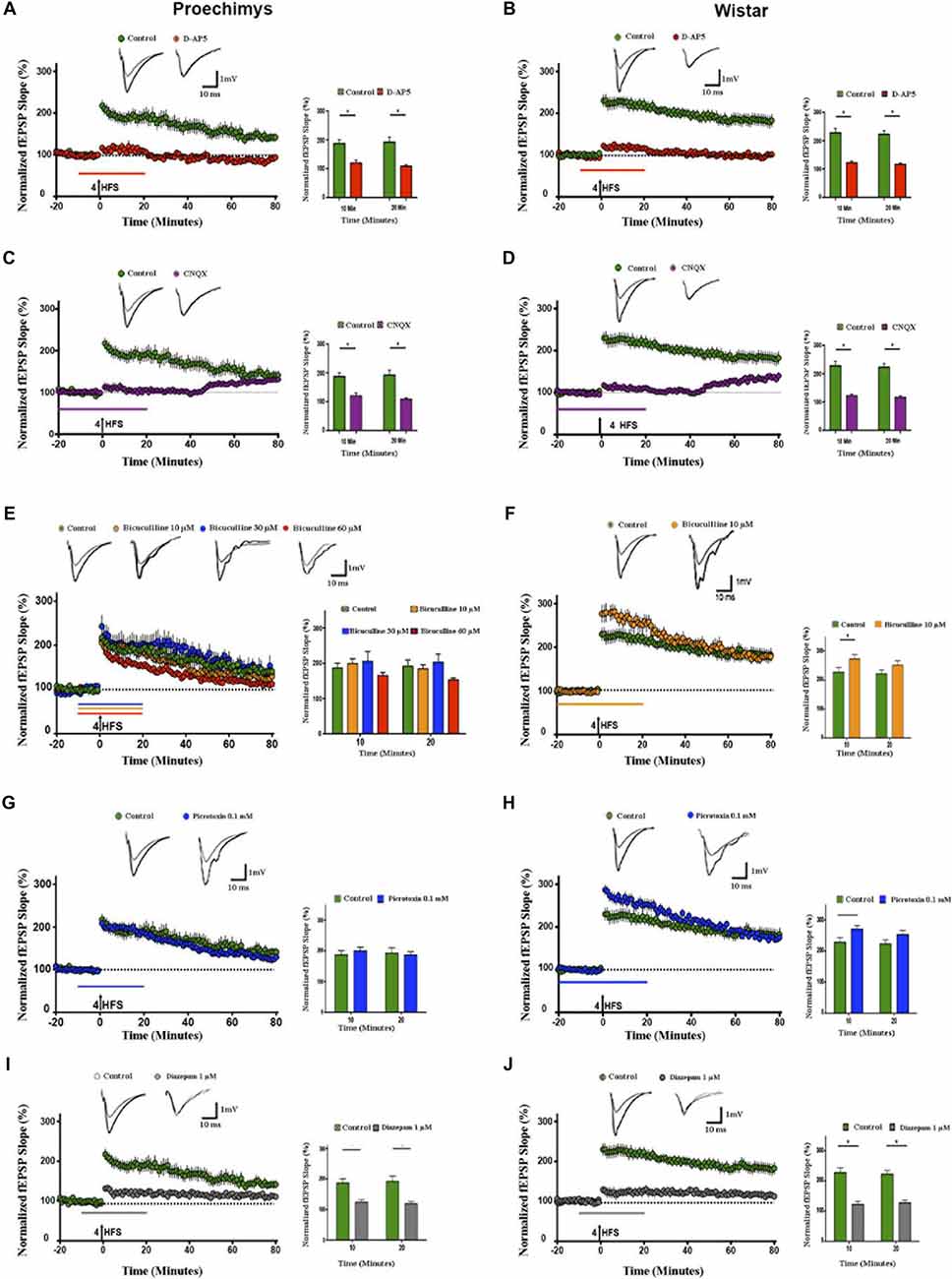
Figure 3. Effects of d-(-)-2-amino-5-phosphonopentanoicacid (D-AP5), CNQX, bicuculline, picrotoxin and diazepam on LTP. Slices of both species were bathed with D-AP5 (100 μM), CNQX (10 μM) bicuculline (10, 30 and 60 μM), picrotoxin (0.1 mM) and diazepam (1 μM) before and during 20 min after tetanic stimulation; see the horizontal lines insets above x-axis representing time periods. (A,B) D-AP5 and (C,D) CNQX prevented the LTP induction in slices of both animal species (red and purple circles, respectively). However, bicuculline (10 μM; orange circles), bicuculline (30 μM; blue circles) bicuculline (60 μM; red circles) did not enhance fEPSP slope after 4HFS in Proechimys (E), but LTP induction was facilitated only in Wistar’s slices with bicuculline (10 μM; F). Recordings of control slices are represented by green circles. The graph insets represent the average of fEPSP slope at 10 and 20 min after tetanic stimulation in control slices and with bicuculline application. For Wistar, the facilitation was found only at 10 min after 4HFS (inset F). Superimposed recordings of average fEPSP slope: from baseline (gray trace) and after 20 min of 4HFS (black trace) for D-AP5, CNQX, control and bicuculline with different doses concentration. (G,H) Time evolution of fEPSP slopes of Proechimys and Wistar bathed with picrotoxin (0.1 mM), in Proechimys, fEPSP slope was not facilitated during GABAA receptor antagonist (G). In Wistar, fEPSP slope of slices bathed with picrotoxin is increased in comparison to control (H). Inset, superimposed recordings of average fEPSP slope: baseline (gray trace) and after 20 min of 4HFS (black trace), for picrotoxin. Note that in the experiments of Wistar under GABAA antagonist, the period of application of the drug was increased to allow a stable baseline before the induction of LTP. (I,J) Time evolution of fEPSP slopes during diazepam application. In both animal species, diazepam prevented the LTP induction (gray circles), recordings of control slices (green circles). Inset, superimposed recordings of average fEPSP slope: baseline (gray trace) and after 20 min (black trace). Graphs represent the average of fEPSP slope at the different times assessed. *< 0.05.
Then, isolated NMDA receptor was assessed using AMPA/kainate receptors antagonist (CNQX 10 μM) combined with magnesium-free aCSF. The effects of bath application with CNQX were measured in Proechimys (13/6 slices/animals) and Wistar (12/5 slices/animals). The potentiation of fEPSP slope were prevented in both animal species; for example, at 20 min after LTP induction, fEPSP slope changes were 6.4 ± 5.8% in Proechimys and 9.0 ± 5.7% in Wistar (Figures 3C,D).
Next, using bicuculline, a GABAA receptor antagonist, we evaluated the effects of GABAA inhibition on LTP. Interestingly, contrasting to Wistar rats (16/8, slices/animals), LTP induction was not facilitated by bicuculline in the Proechimys rodents (30/10, slices/animals). For example, using bicuculline, fEPSP slope changes at 10-min after tetanic stimulation in Proechimys were 99.1 ± 13.3 (bicuculline 10 μM, 15/10, slices/animals) and 97.8 ± 18.6% (control, 8/4, slices/animals; U = 46, p = 0.1780; Figure 3E). In Wistar rats, at 10-min after HFS: 172.9 ± 14.6% (bicuculline 10 μM, 16/8, slices/animals) and 126.0 ± 22.5% (control, 8/4, slices/animals; U = 42, *p = 0.0149; Figure 3F). Although the lack of fEPSP potentiation in the presence of bicuculline (10 μM) may point to a possible inhibitory network modulation of the synaptic transmission in the Neotropical rodents, another hypothesis would be that the regulatory mechanism by inhibitory network is similar in both animal species, however GABAA receptors sensitivity to bicuculline is distinct, in this case GABAA receptors sensitivity of Proechimys would be lower than that of Wistar. To test this hypothesis, experiments using bicuculline 30 μM as well as 60 μM were performed. However, no differences in fEPSP slope were encountered among all bicuculline concentrations, for example at 10-min after HFS: 99.1 ± 13.3 (bicuculline 10 μM, 15/10, slices/animals), 105.9 ± 26.6 (bicuculline 30 μM, 13/10, slices/animals), 65.3 ± 9.1% (bicuculline 60 μM, 13/10, slices/animals) and 97.8 ± 18.6% (control, 8/4, slices/animals; H = 3.024; p = 0.3880; Figure 3E). Since these findings suggested that sensitivity of the GABAA receptor to bicuculline is not a crucial issue, picrotoxin (0.1 mM), a non-competitive antagonist of GABAA receptors was tested on LTP induction. At 10-min after tetanic stimulation, changes in fEPSP slope in Proechimys (10/8, slices/animals) were 99.7 ± 12.1% and 97.8 ± 18.6% (control; U = 53, p = 0.1478; Figure 3G); in Wistar rats (16/8, slices/animals) at 10 min after HFS, fEPSP slope changes of 170.2 ± 11.9% and 126.0 ± 22.5% (control; U = 66, *p = 0.0067; Figure 3H). Bicuculline is a potent competitive antagonist of GABAA receptors, but its binding action can be reverted by increasing the amount of substrate near to the receptor. Picrotoxin is a non-competitive antagonist of GABAA receptors which binds to the allosteric site of the receptor, leading to distortion in the configuration of the active site, causing substrate inability to attach GABA to the receptor. Since neither of these two approaches were satisfactory to promote LTP potentiation, these results suggested a possible role of the GABAergic network modulation during LTP induction in the Neotropical rodents. Next, the effects of the GABAA receptor positive allosteric modulator on LTP induction were assessed. Diazepam (1 μM) effectively prevented the LTP induction in slices of both animal species, for example, in Proechimys (10/5, slices/animals) fEPSP slope changes at 20-min after tetanic stimulation were 19.1 ± 7.7% and 97.8 ± 18.6% in control (Figure 3I; U = 10, **p = 0.0016); in Wistar rats (10/5, slices/animals) 27.5 ± 9.2% and 122. ± 12.9% in control (U = 4, ***p = 0.0007; Figure 3J).
Effects of GABAA Receptor Blockade by Bicuculline on Field Responses
The effects of GABAA inhibition on spontaneous FPs were evaluated in CA1 pyramidal cells by adding bicuculline in the perfusion bath medium in Proechimys (18/6, slices/animals; bicuculline 10 μM, 20 μM and 60 μM) and Wistar rats (10/3, slices/animals; bicuculline 10 μM). With 10 μM of bicuculline, slices of Wistar rats exhibited spontaneous activity in 100% of the slices (for this reason, Wistar rats were studies only with 10 μM of bicuculline). On the other hand, only 50% of Proechimys’s slices bathed with 10 μM of bicuculline showed spontaneous activity (x2 = 5.268, *p = 0.0220), but with 30 μM and 60 μM of bicuculline, 100% of slices showed spontaneous activity. However, number of spontaneous events were extremely different between the two species of rodents. Using 10 μM, number of events per minute was 0.38 ± 0.1 in Proechimys vs. 6.14 ± 0.3 in Wistar (U = 0.0, *p = 0.0143). In Proechimys, the number of events per minute was increased when analyzing all the three different concentrations (H = 8.138, *p = 0.0106), but the difference was observed between 10 μM and 30 μM as well as between 10 μM and 60 μM, but not between 30 μM and 60 μM (Dunn’s multiple comparison test; mean of events per minute, 10 μM = 0.38 ± 0.1; 30 μM = 1.84 ± 0.43; 60 μM = 1.81 ± 0.44). Note in Figures 4A,B the higher spontaneous activity in the Wistar rats than that of the Proechimys. Note the single recurrent PS and single fEPSP in Proechimys’s slices (Figures 4C1,C2) in presence of bicuculline, contrasting with the multiple bursts in Wistar recordings (Figures 4D1,D2).
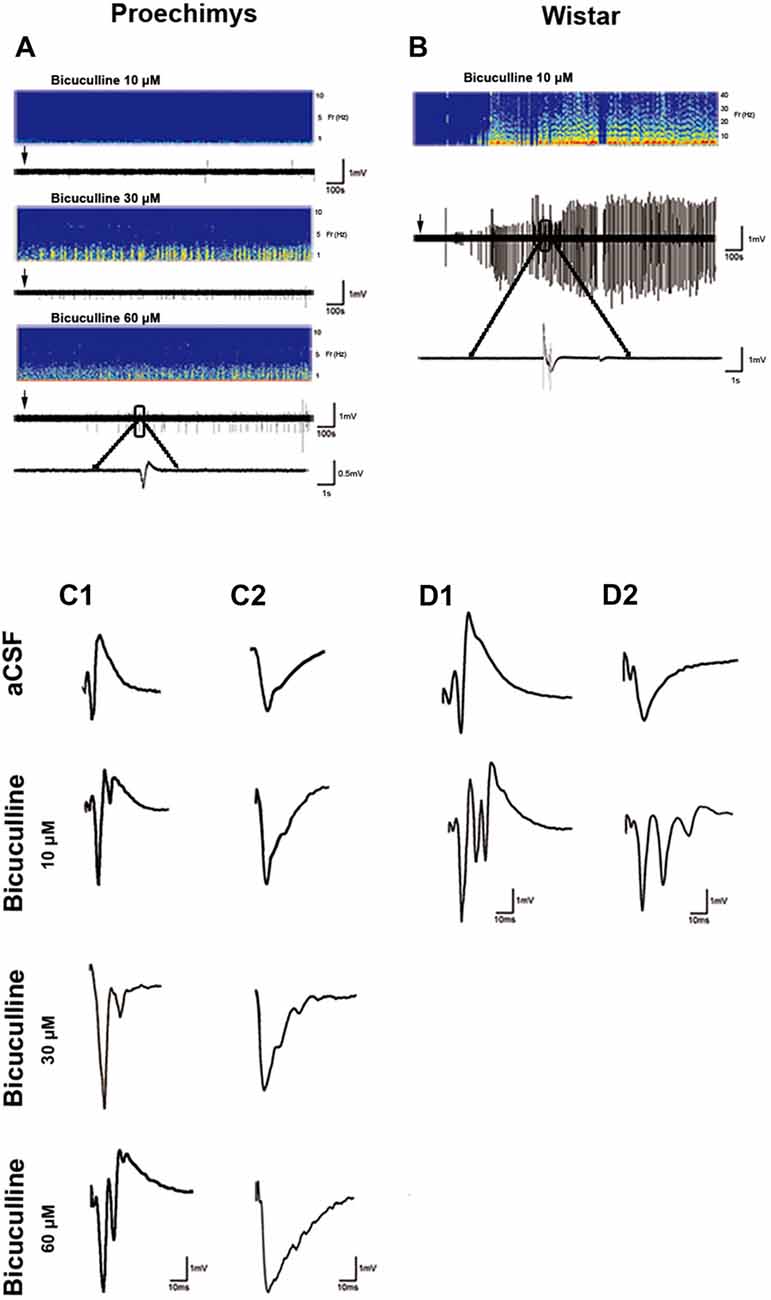
Figure 4. Effects of GABAA receptor blockade by bicuculline on field responses. (A,B) show the recording of spontaneous activity after bath-applied bicuculline. Note that spontaneous activity is remarkably lower in Proechimys, even during increase of bicuculline concentration (10, 30 and 60 μM; A). For Wistar, only was assessed spontaneous activity with bicuculline at 10 μM (B). Arrows indicate when bicuculline was applied. Representative spectrograms from two animals showing the corresponding burst of spontaneous activity during bicuculline bath. Panels (C1,D1) are samples of evoked-PS. Panels (C2,D2) represent evoked-fEPSP. Both were recorded in artificial cerebrospinal fluid (aCSF) and in presence of bicuculline 10 μM, 30 μM and 60 μM. Note the single recurrent PS and single fEPSP in Proechimys’s slices in presence of bicuculline, contrasting with the multiple bursts in Wistar recordings.
Discussion
Little is known about synaptic plasticity in wild Neotropical rodents. The present findings draw attention to the protocol-independent fast decay of the early-phase LTP and to the robust dynamic network inhibition assessed in the hippocampal CA1 area of the Proechimys rodents. The comparison between LTP expression in the presence and absence of a GABAA receptor antagonist offers an indirect mensuration of alterations in inhibition and the consequent impact on excitatory responses since the inhibitory inputs and associated modifications should be inhibited by the antagonist. In Wistar rats, our results are consistent with previous findings that have shown that relief from GABAA inhibition has facilitated LTP induction (Wigström and Gustafsson, 1983; Grover and Yan, 1999). In contradiction, our findings suggest that fEPSP of hippocampal CA1 pyramidal cells are not modulated by GABAA inhibition during LTP induction in Proechimys since LTP induction was not facilitated by the presence of both bicuculline and picrotoxin.
A plausible hypothesis to explain our data would be given by differences in GABAA quantification and distribution. Rocha et al. (2006) performed quantitative receptor autoradiography to assess the distribution of high-affinity GABAA receptors through the powerful GABAA agonist [3H] muscimol and reported no differences between Proechimys and Wistar rats in GABAA binding quantification in the hippocampal subfields. On the other hand, it has been reported that the effect of inhibition on LTP seems to be stimulation-dependent (Chapman et al., 1998), thus the pattern of stimulation used in this study may play a role in the inhibition of population EPSP in the presence of bicuculline/picrotoxin.
Efficient synaptic transmission involves the control of brain excitability through inhibitory signaling which fundamentally occurs through GABAergic neurotransmission via ionotropic GABAA receptors and the output of the adult hippocampal pyramidal neurons is strongly regulated by the activity of GABAergic cells (Bernard et al., 2000; Lee and Maguire, 2014). It seems that Proechimys’ brain exerts a very particular control of inhibitory modulation. For example, when 43 Proechimys rodents were submitted to electrical Kindling process, only three of them reached stage 5 of Racine scale and among the remaining 40 animals, 16 of them remained at stage 1, 14, 7 and 3 of them did not get beyond stages 2, 3 and 4, respectively (Racine, 1972; Arida et al., 2005). These findings suggest that Proechimys rodents seem incapable to consolidate an epileptic circuitry which could reflect functional alterations in inhibitory and facilitatory processes in the brain of these animals. Indeed, it has been suggested that brain stimulation (e.g., HFS) used to induce kindling are similar to LTP since both result in synaptic structural alterations and facilitation (McEachern and Shaw, 1999; Meador, 2007). Following this reasoning, previous reports described that in Proechimys hippocampus, parvalbumin (PV)-expressing interneurons are generally distributed and predominate in the stratum oriens, pyramidale and alveus (Fabene et al., 2001; Scorza et al., 2010). Furthermore, after an acute epileptogenic insult, Fos immediate early genes induction occurred in almost all the hippocampal PV-expressing interneurons up to 24 h after status epilepticus in Proechimys, contrasting to the lower proportion of the double labeled PV-Fos cells found in Wistar rats, thus suggesting differential mechanisms of GABAergic response between the two animals species (Fabene et al., 2004).
Interneurons regulate the firing rate of neurons to modulate circuitry activity. The CA1 hippocampal subfield possess a huge diversity of interneurons (Klausberger and Somogyi, 2008), thus it is possible to speculate that an inhibitory interneuron, which was innervating another inhibitory interneuron, is now inhibited in the presence of GABAA antagonist and this second interneuron, that is the source of feed forward inhibition onto neurons of CA1 pyramidal layer, is now released from inhibition and hence, is able to project an inhibitory input onto CA1 pyramidal neuron. Therefore, the disinhibition implemented in the local inhibitory circuit could account for our findings in Proechimys but remains to be investigated.
It’s worth mentioning that LTP induction was protocol-independent in Proechimys, since TBS, 1HFS and 4HFS protocols reliably induced LTP but fEPSP slopes dropped gradually in all protocols, short-lived LTP was encountered after TBS/1HFS however LTP maintenance was effective after 4HFS during 80-min of recordings. As suggested by previous studies performed in guinea pigs and laboratory rats (Buzsáki and Eidelberg, 1982; Stelzer et al., 1994), it is possible that synaptic strength inhibition might be enhanced during LTP maintenance in Proechimys. Furthermore, the I/O relationship for fEPSP vs. FV magnitude points to lower excitability in the CA1 area of Proechimys (pre × post HFS). Likewise, utilizing an in vitro model of bicuculline (10 μM), which is broadly used to assess enhancement of electrical activity, Wistar slices exposed to bicuculline generated intense activity, in accordance with previous reports (Li et al., 2001). Notwithstanding, it is striking that, even increasing the bicuculline concentration up to 60 μM, the electrical activity was not as intense in Proechimys as in Wistar rats. High concentrations of bicuculline (60 microMol) decrease LTP expression at longer delay in Proechimys compared to control. Early-phase LTP in Proechimys decays very rapidly (10, 30 and 60 microMol). Previous studies reported that even weak protocols of induction can induce LTP that lasts several hours in rodents (Lu et al., 2011; Dong et al., 2015). Why early LTP in Proechimys decays very rapidly remains unknown. In our previous study (Guimarães Marques et al., 2018), fEPSP showed that LTP decays quickly over time reaching baseline at 90 min after TBS in Proechimys, in opposition to the stable LTP encountered in the Wistar rodents during 3-h period.
Previous studies in the laboratory rodents asserted that NMDA receptor activation is essential for induction of LTP (Collingridge et al., 1983; Muller et al., 1988; Murphy et al., 1997; Gruart et al., 2006). In agreement with those reports, we found the failure of LTP induction in the presence of D-AP5 and in both Wistar and Proechimys rodents. In the case of CNQX, HFS did not generate robust LTP. However, when the drug was removed, the potentiation effect started to appear. Therefore, NMDA receptor activation elicited LTP in both animal species, in which non-NMDA receptors were needed for the LTP expression.
This study described a rapid loss of synaptic potentiation over time and an a possibly high inhibitory tone in the CA1 network mediated by GABAA receptors in the Proechimys rodents. Although bicuculine and picrotoxin did not modify LTP values in the Proechimys rodents, other aspects argue in favor of a high inhibitory tone mediated by GABAA receptors, such as the lower spontaneous FPs under GABAA antagonists as well the lower excitability observed in the I/O relationships evaluated before and after the induction of LTP. Nevertheless, other mechanisms could be involved such as functional saturation of the CA3-CA1 synapses through LTP induction evoked by HFS, a process that could modulate the activity-dependent synaptic plasticity and prevent acquisition of associative learning in behaving mice (Gruart et al., 2006). The inhibitory control carried by GABAB receptors is another possibility to explain our findings. GABAB receptors are pivotal regulators of neuronal excitability and their activation is crucial for associative learning in behaving animals (Jurado-Parras et al., 2016). Experiments assessing the roles of GABAB receptor remain to be addressed in Proechimys. Mice lacking the GABAB subunit isoform exhibited larger in vivo LTP than wild type (WT) mice but operant learning task impairments (Jurado-Parras et al., 2016). A contrast between increased LTP and lack of learning in operant tasks was also reported in mutant mice with deficits in GABAergic septohippocampal projections, which exhibited delayed acquisition and lower performance in the medial septum self-stimulation task, suggesting a distinctive susceptibility of the hippocampus to GABAergic inputs and cognitive processes (Vega-Flores et al., 2014). Therefore, the incorporation of robust inhibition into the hippocampal excitatory circuitry is a key ingredient in network dynamics that may crucially affect learning and memory. Previous investigation showed the poor performance of Proechimys rodents in plus-maze discrimination avoidance task and object recognition test since long-term memory did not last 24-h in these rodents, contrasting with the preserved 24-h memory of Wistar rats (Guimarães Marques et al., 2018). Current studies satisfy a number of the criteria needed to link learning/memory and hippocampal LTP as well as to link the conservation of long-lasting memory and LTP maintenance (Pastalkova et al., 2006; Whitlock et al., 2006; Sacktor, 2008). This study suggests a lower excitability in the hippocampus of Proechimys in comparison to Wistar rats. The transformation of decaying to non-decaying LTP is a process resulting from active inhibition and crucial for the transformation of short-term to long-term memory (Nguyen et al., 1994; Lu et al., 2011; Dong et al., 2015). The excitation-inhibition ratio known as E/I balance is widely accepted as a critical measure for evaluating fitness of any brain and memory preservation is costly and demands energy (Hardt et al., 2013). The brains of these rodents from Amazon rainforest may express signatures of their challenging wild ecology. At this point, a causal relation between the robust inhibitory tone in Proechimys and rapid early-LTP decay/poor long-term memory is unknown, and several fundamental questions arise from this investigation which remain to be addressed: (1) How the present findings effectively impact learning and memory of the Proechimys? (2) What are the behavioral advantages? (3) Manipulation of inhibitory network will be able to down-regulate this early-phase decay LTP in Proechimys? (4) How to increase the longevity of LTP and long-term memory? and (5) What are the physiological implications? Currently, neuroscience research still seeks to reveal molecular pathways that control learning and memory processes, Proechimys may prove useful in identifying such mechanisms in complement to traditional animal models.
Author Contributions
All experiments were performed in the Department of Neurology/Neurosurgery, Universidade Federal de São Paulo. SR-G and NO-V together carried out the experiments and wrote the article. SR-G, A-CA, FS and EC prepared the figures and wrote the article. SR-G and CS designed the investigation and wrote the article. All persons designated as authors qualify for authorship. The authors have read and approved the final version of the article.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brazil (CAPES)—Finance Code 001, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; 2016/06879-2), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), FAPESP/CNPq/MCT-Instituto Nacional de Ciência e Tecnologia de Neurociência Translacional (573604/2008-8) and Universidad Nacional Autónoma de Honduras (CU-O-041-05-2014).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Jan-Oliver Hollnagel for technical assistance with Matlab programming and Christophe Bernard for his comments on the manuscript.
References
Amrein, I. (2015). Adult hippocampal neurogenesis in natural populations of mammals. Cold Spring Harb. Perspect. Biol. 7:a021295. doi: 10.1101/cshperspect.a021295
Arida, R. M., Scorza, F. A., de Amorim Carvalho, R., and Cavalheiro, E. A. (2005). Proechimys guyannensis: an animal model of resistance to epilepsy. Epilepsia 46, 189–197. doi: 10.1111/j.1528-1167.2005.01065.x
Bernard, C., Cossart, R., Hirsch, J. C., Esclapez, M., and Ben-Ari, Y. (2000). What is GABAergic inhibition? How is it modified in epilepsy? Epilepsia 41, S90–S95. doi: 10.1111/j.1528-1157.2000.tb01564.x
Bliss, T. V. P., and Collingridge, G. L. (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361, 31–39. doi: 10.1038/361031a0
Bliss, T. V. P., and Cooke, S. F. (2011). Long-term potentiation and long-term depression: a clinical perspective. Clinics 66, 3–17. doi: 10.1590/s1807-59322011001300002
Bliss, T. V. P., and Lømo, T. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. J. Physiol. 232, 331–356. doi: 10.1113/jphysiol.1973.sp010273
Buzsáki, G., and Eidelberg, E. (1982). Direct afferent excitation and long-term potentiation of hippocampal interneurons. J. Neurophysiol. 48, 597–607. doi: 10.1152/jn.1982.48.3.597
Carlson, B. A. (2012). Diversity matters: the importance of comparative studies and the potential for synergy between neuroscience and evolutionary biology. Arch. Neurol. 69, 987–993. doi: 10.1001/archneurol.2012.77
Chapman, C. A., Perez, Y., and Lacaille, J. C. (1998). Effects of GABAA inhibition on the expression of long-term potentiation in CA1 pyramidal cells are dependent on tetanization parameters. Hippocampus 8, 289–298. doi: 10.1002/(sici)1098-1063(1998)8:3<289:aid-hipo10>3.0.co;2-x
Collingridge, G. L., Kehl, S. J., and McLennan, H. (1983). Excitatory amino acids in synaptic transmission in the Schaffer collateral-commissural pathway of the rat hippocampus. J. Physiol. 334, 33–46. doi: 10.1113/jphysiol.1983.sp014478
Collingridge, G. L., Peineau, S., Howland, J. G., and Wang, Y. T. (2010). Long-term depression in the CNS. Nat. Rev. Neurosci. 11, 459–473. doi: 10.1038/nrn2867
Dong, Z., Han, H., Li, H., Bai, Y., Wang, W., Tu, M., et al. (2015). Long-term potentiation decay and memory loss are mediated by AMPAR endocytosis. J. Clin. Invest. 125, 234–247. doi: 10.1172/JCI77888
Fabene, P. F., Andrioli, A., Priel, M. R., Cavalheiro, E. A., and Bentivoglio, M. (2004). Fos induction and persistence, neurodegeneration, and interneuron activation in the hippocampus of epilepsy-resistant versus epilepsy-prone rats after pilocarpine-induced seizures. Hippocampus 14, 895–907. doi: 10.1002/hipo.20003
Fabene, P. F., Correia, L., Carvalho, R. A., Cavalheiro, E. A., and Bentivoglio, M. (2001). The spiny rat Proechimys guyannensis as model of resistance to epilepsy: chemical characterization of hippocampal cell populations and pilocarpine-induced changes. Neuroscience 104, 979–1002. doi: 10.1016/s0306-4522(01)00138-5
Grover, L. M., and Yan, C. (1999). Blockade of GABAA receptors facilitates induction of NMDA receptor-independent long-term potentiation. J. Neurophysiol. 81, 2814–2822. doi: 10.1152/jn.1999.81.6.2814
Gruart, A., Muñoz, M. D., and Delgado-García, J. M. (2006). Involvement of the CA3-CA1 synapse in the acquisition of associative learning in behaving mice. J. Neurosci. 26, 1077–1087. doi: 10.1523/JNEUROSCI.2834-05.2006
Guimarães Marques, M. J., Reyes-Garcia, S. Z., Marques-Carneiro, J. E., Lopes-Silva, L. B., Andersen, M. L., Cavalheiro, E. A., et al. (2018). Long-term potentiation decay and poor long-lasting memory process in the wild rodents Proechimys from brazil’s amazon rainforest. Front. Behav. Neurosci. 12:2. doi: 10.3389/fnbeh.2018.00002
Hardt, O., Nader, K., and Wang, Y.-T. (2013). GluA2-dependent AMPA receptor endocytosis and the decay of early and late long-term potentiation: possible mechanisms for forgetting of short- and long-term memories. Philos. Trans. R. Soc. B Biol. Sci. 369:20130141. doi: 10.1098/rstb.2013.0141
Hebb, D. O. (1949). The Organization of Behavior: A Neuropsychological Theory. New York, NY: Wiley & Sons.
Ito, M., and Kano, M. (1982). Long-lasting depression of parallel fiber-Purkinje cell transmission induced by conjunctive stimulation of parallel fibers and climbing fibers in the cerebellar cortex. Neurosci. Lett. 33, 253–258. doi: 10.1016/0304-3940(82)90380-9
Jurado-Parras, M. T., Delgado-García, J. M., Sánchez-Campusano, R., Gassmann, M., Bettler, B., and Gruart, A. (2016). Presynaptic GABABreceptors regulate hippocampal synapses during associative learning in behaving mice. PLoS One 11:e0148800. doi: 10.1371/journal.pone.0148800
Keifer, J., and Summers, C. H. (2016). Putting the “Biology” back into “Neurobiology”: the strength of diversity in animal model systems for neuroscience research. Front. Syst. Neurosci. 10:69. doi: 10.3389/fnsys.2016.00069
Klaus, F., and Amrein, I. (2012). Running in laboratory and wild rodents: differences in context sensitivity and plasticity of hippocampal neurogenesis. Behav. Brain Res. 227, 363–370. doi: 10.1016/j.bbr.2011.04.027
Klausberger, T., and Somogyi, P. (2008). Neuronal diversity and temporal dynamics: the unity of hippocampal circuit operations. Science 321, 53–57. doi: 10.1126/science.1149381
Lee, V., and Maguire, J. (2014). The impact of tonic GABAA receptor-mediated inhibition on neuronal excitability varies across brain region and cell type. Front. Neural Circuits 8:3. doi: 10.3389/fncir.2014.00003
Li, J., Shen, H., Naus, C. C., Zhang, L., and Carlen, P. L. (2001). Upregulation of gap junction connexin 32 with epileptiform activity in the isolated mouse hippocampus. Neuroscience 105, 589–598. doi: 10.1016/s0306-4522(01)00204-4
Löwel, S., and Singer, W. (1992). Selection of intrinsic horizontal connections in the visual cortex by correlated neuronal activity. Science 255, 209–212. doi: 10.1126/science.1372754
Lu, Y., Ji, Y., Ganesan, S., Schloesser, R., Martinowich, K., Sun, M., et al. (2011). TrkB as a potential synaptic and behavioral tag. J. Neurosci. 31, 11762–11771. doi: 10.1523/JNEUROSCI.2707-11.2011
Lüscher, C., and Malenka, R. C. (2012). NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 4:a005710. doi: 10.1101/cshperspect.a005710
Mayford, M., Siegelbaum, S. A., and Kandel, E. R. (2012). Synapses and memory storage. Cold Spring Harb. Perspect. Biol. 4:a005751. doi: 10.1101/cshperspect.a005751
McEachern, J. C., and Shaw, C. A. (1999). The plasticity-pathology continuum: defining a role for the LTP phenomenon. J. Neurosci. Res. 58, 42–61. doi: 10.1002/(sici)1097-4547(19991001)58:1<42:aid-jnr6>3.0.co;2-l
Meador, K. J. (2007). The basic science of memory as it applies to epilepsy. Epilepsia 48, 23–25. doi: 10.1111/j.1528-1167.2007.01396.x
Muller, D., Joly, M., and Lynch, G. (1988). Contributions of quisqualate and NMDA receptors to the induction and expression of LTP. Science 242, 1694–1697. doi: 10.1126/science.2904701
Murphy, K. P. S. J., Reid, G. P., Trentham, D. R., and Bliss, T. V. P. (1997). Activation of NMDA receptors is necessary for the induction of associative long-term potentiation in area CA1 of the rat hippocampal slice. J. Physiol. 504, 379–385. doi: 10.1111/j.1469-7793.1997.379be.x
Nguyen, P., Abel, T., and Kandel, E. (1994). Requirement of a critical period of transcription for induction of a late phase of LTP. Science 265, 1104–1107. doi: 10.1126/science.8066450
Pastalkova, E., Serrano, P., Pinkhasova, D., Wallace, E., Fenton, A. A., and Sacktor, T. C. (2006). Storage of spatial information by the maintenance mechanism of LTP. Science 313, 1141–1144. doi: 10.1126/science.1128657
Racine, R. J. (1972). Modification of seizure activity by electrical stimulation: I. After-discharge threshold. Electroencephalogr. Clin. Neurophysiol. 32, 269–279. doi: 10.1016/0013-4694(72)90176-9
Ramón y Cajal, S. (1894). The croonian lecture: la fine structure des centres nerveux. Proc. R. Soc. Lond. 55, 444–468. doi: 10.1098/rspl.1894.0063
Rocha, L., Arida, R. M., Carvalho Rde, A., Scorza, F. A., Neri-Bazan, L., and Cavalheiro, E. A. (2006). GABA and opioid binding distribution in the brain of the seizure-resistant Proechimys guyannensis: an autoradiography study. Synapse 60, 392–398. doi: 10.1002/syn.20312
Sacktor, T. C. (2008). Chapter 2 PKMζ, LTP maintenance, and the dynamic molecular biology of memory storage. Prog. Brain Res. 169, 27–40. doi: 10.1016/s0079-6123(07)00002-7
Salar, S., Lapilover, E., Müller, J., Hollnagel, J. O., Lippmann, K., Friedman, A., et al. (2016). Synaptic plasticity in area CA1 of rat hippocampal slices following intraventricular application of albumin. Neurobiol. Dis. 91, 155–165. doi: 10.1016/j.nbd.2016.03.008
Scorza, C. A., Araujo, B. H. S., Arida, R. M., Scorza, F. A., Torres, L. B., Amorim, H. A., et al. (2010). Distinctive hippocampal CA2 subfield of the Amazon rodent Proechimys. Neuroscience 169, 965–973. doi: 10.1016/j.neuroscience.2010.05.079
Stelzer, A., Simon, G., Kovacs, G., and Rai, R. (1994). Synaptic disinhibition during maintenance of long-term potentiation in the CA1 hippocampal subfield. Proc. Natl. Acad. Sci. U S A 91, 3058–3062. doi: 10.1073/pnas.91.8.3058
Sweatt, J. D. (2010). “Long-term potentiation—a candidate cellular mechanism for information storage in the central,” in Mechanisms of Memory, ed. J. D. Sweatt (London: Academic Press), 150–189.
Vega-Flores, G., Rubio, S. E., Jurado-Parras, M. T., Gómez-Climent, M. Á., Hampe, C. S., Manto, M., et al. (2014). The GABAergic septohippocampal pathway is directly involved in internal processes related to operant reward learning. Cereb. Cortex 24, 2093–2107. doi: 10.1093/cercor/bht060
Vitureira, N., and Goda, Y. (2013). Cell biology in neuroscience: the interplay between Hebbian and homeostatic synaptic plasticity. J. Cell Biol. 203, 175–186. doi: 10.1083/jcb.201306030
Whitlock, J. R., Heynen, A. J., Shuler, M. G., and Bear, M. F. (2006). Learning induces long-term potentiation in the hippocampus. Science 313, 1093–1097. doi: 10.1126/science.1128134
Keywords: synaptic plasticity, long-term potentiation, CA1 region, hippocampal, GABA modulators, neural inhibition
Citation: Reyes-Garcia SZ, de Almeida A-CG, Ortiz-Villatoro NN, Scorza FA, Cavalheiro EA and Scorza CA (2018) Robust Network Inhibition and Decay of Early-Phase LTP in the Hippocampal CA1 Subfield of the Amazon Rodent Proechimys. Front. Neural Circuits 12:81. doi: 10.3389/fncir.2018.00081
Received: 09 April 2018; Accepted: 13 September 2018;
Published: 04 October 2018.
Edited by:
Matthew Charles Walker, University College London, United KingdomReviewed by:
José M. Delgado-García, Universidad Pablo de Olavide, SpainJean-Marie Billard, Institut National de la Santé et de la Recherche Médicale (INSERM), France
Copyright © 2018 Reyes-Garcia, de Almeida, Ortiz-Villatoro, Scorza, Cavalheiro and Scorza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carla A. Scorza, Y2FybGEuc2NvcnphQHVuaWZlc3AuYnI=