






- 1 HAS Research Group for Cortical Microcircuits, Department of Comparative Physiology, University of Szeged, Hungary
- 2 Department of Neurosurgery, University of Szeged, Hungary
Neurogliaform cells in the rat elicit combined GABAA and GABAB receptor-mediated postsynaptic responses on cortical pyramidal cells and establish electrical synapses with various interneuron types. However, the involvement of GABAB receptors in postsynaptic effects of neurogliaform cells on other GABAergic interneurons is not clear. We measured the postsynaptic effects of neurogliaform cells in vitro applying simultaneous whole-cell recordings in human and rat cortex. Single action potentials of human neurogliaform cells evoked unitary IPSPs composed of GABAA and GABAB receptor-mediated components in various types of inteneuron and in pyramidal cells. Slow IPSPs were combined with homologous and heterologous electrical coupling between neurogliaform cells and several human interneuron types. In the rat, single action potentials in neurogliaform cells elicited GABAB receptor-mediated component in responses of neurogliaform, regular spiking, and fast spiking interneurons following the GABAA receptor-mediated component in postsynaptic responses. In conclusion, human and rat neurogliaform cells elicit slow IPSPs and reach GABAA and GABAB receptors on several interneuron types with a connection-specific involvement of GABAB receptors. The electrical synapses recorded between human neurogliaform cells and various interneuron types represent the first electrical synapses recorded in the human cortex.
Introduction
Neurogliaform cells were first described by Ramon y Cajal as spiderweb cells according to their distinctive morphology: multiple dendrites of these cells spread radially from a small, round soma in all directions and their dense axonal plexus composed of very fine branches with small, frequently occurring boutons resembled webs of spiders (Cajal, 1904 ; Kawaguchi and Kubota, 1997 ; Kawaguchi et al., 2006 ; Kisvarday et al., 1990 ; Krimer et al., 2005 ). The occurrence of neurogliaform cells is reported in all layers of cerebral cortex (Hestrin and Armstrong, 1996 ; Kawaguchi and Kubota, 1997 ; Kisvarday et al., 1990 ; Krimer et al., 2005 ; Sancesario et al., 1998 ). Golgi studies confirmed the presence of neurogliaform cells in samples of human association cortex (Kisvarday et al., 1990 ). Neurogliaform cells were found in dorsolateral prefrontal cortex of monkeys (Krimer et al., 2005 ) and in various cerebral structures of rats including stiatum (Sancesario et al., 1998 ), CA1 area of hippocampus (Price et al., 2005 ; Zsiros and Maccaferri, 2005 ), and visual (Hestrin and Armstrong, 1996 ) and frontal cortex (Kawaguchi and Kubota, 1997 ).
Neurogliaform cells have a unique position among cortical interneurons because they establish electrical synapses not only with each other but also with other interneuron types in the neocortex and in the CA1 region of the hippocampus (Price et al., 2005 ; Simon et al., 2005 ; Zsiros and Maccaferri, 2005 ). Thus, neurogliaform cells link multiple networks of interneurons and were suggested to play a central role in generating and shaping synchronized activity of neuronal circuits (Simon et al., 2005 ; Zsiros and Maccaferri, 2005 ). All but one type of interneuron trigger fast inhibitory postsynaptic potentials (IPSPs) in their postsynaptic target cells mediated by GABAA receptors (Buhl et al., 1994 ; Gupta et al., 2000 ; Tamas et al., 2003 ). By contrast, neurogliaform cells are the only known type of interneuron capable of eliciting slow, long-lasting IPSPs through a combined activation of GABAA and GABAB receptors (Tamas et al., 2003 ). To date, this effect of neurogliaform cells has been demonstrated only on postsynaptic pyramidal cells (Tamas et al., 2003 ) and a relatively small contribution of GABAB receptors to neurogliaform cell evoked responses was demonstrated on neurogliaform cells in the CA1 area of the hippocampus (Price et al., 2005 ). The expression pattern and immunocytochemical localization suggest that GABAB receptors can be found in the neocortex on postsynaptic pyramidal cells and on interneurons (Lopez-Bendito et al., 2002 ). Electrophysiological experiments also suggest the presence of GABAB receptors on GABAergic neurons (Martina et al., 2001 ; Mott et al., 1999 ), but the cellular components of the neocortical microcircuit reaching the GABAB receptors on interneurons are not known.
To investigate the role of neurogliaform cells in human cerebral cortex and their postsynaptic effects on interneurons in rat cerebral cortex, we performed simultaneous dual, triple, and quadruple recordings of neurons in layers I–IV of human associational cortices and rat somatosensory cortex in vitro with correlated light microscopic analysis.
Materials and Methods
Electrophysiology
All procedures on human tissue were performed with the approval of the Ethical Committee of the University of Szeged and in accordance with the Declaration of Helsinki. Human slices were derived from association cortices which had to be removed to gain access for the surgical treatment of deeper brain areas with written informed consent of the patients (aged 18–73 years) prior to surgery. Anaesthesia was induced with intravenous midazolam and fentanyl (0.03 mg∕kg, 1–2 µg∕kg respectively). A bolus dose of propofol (1–2 mg∕kg) was administered intravenously. To facilitate endotracheal intubation, the patients received 0.5 mg∕kg rocuronium. After 120 second the trachea was intubated and the patients were ventilated with a mixture of O2 − N2O at a ratio of 1:2. Anaesthesia was maintained with sevoflurane at MAC volume of 1.2–1.5. Slices (350 µm thick) were prepared with a Microm (HM 650 V) vibratome, they were incubated at room temperature for 1 hour in a solution composed of (in mM) 130 NaCl, 3.5 KCl, 1 NaH2PO4, 24 NaHCO3, 1 CaCl, 3 MgSO4, 10 d()-glucose, saturated with 95% O2 and 5% CO2. The solution to be used during recordings differed only in that it contained 3 mM CaCl2 and 1.5 mM MgSO4. Micropipettes (5–7 MΩ) were filled with (in mM) 126 K-gluconate, 4 KCl, 4 ATP-Mg, 0.3 GTP-NA2, 10 HEPES, 10 kreatin-phosphate and 8 biocytin (pH 7.25; 300 mOsm). Somatic whole-cell recordings were obtained at ∼36°C from concomitantly recorded triplets and quadruplets of interneurons and pyramidal cells visualized in layers 2∕3 by infrared differential interference contrast videomicroscopy (Zeiss Axioskop microscope, Hamamatsu CCD camera, Luigs & Neumann Infrapatch set-up and two HEKA EPC 9∕double patch-clamp amplifiers). Signals were filtered at 8 kHz, digitized at 16 kHz and analyzed with PULSE software (HEKA). We identified the recorded cells according their membrane and firing properties (Cauli et al., 2000 ). After identification of the cell types, all possible permutations of connection were checked between the simultaneously recorded cells. Unless specified, traces shown are averages of 30–100 episodes. Presynaptic neurogliaform cells were stimulated to elicit action potentials with brief (2 ms) suprathreshold pulses at >90 second intervals to avoid exhaustion of transmission; other cell types were stimulated at 0.1 Hz. Postsynaptic cells were held at −51 ± 4 mV membrane potential. Rise times of postsynaptic potentials were measured at 10–90% of amplitudes of IPSPs. The decay of evoked IPSPs could not be fitted with single or double exponential functions, we thus measured the half-width of IPSPs for statistical comparison. The amplitude of postsynaptic events was defined as the difference between the peak amplitude and the baseline value measured before the postsynaptic potential onset. In all experiments the contribution of the different types of GABA receptor were tested by applying 10 µM gabazine, a GABAA receptor antagonist and 10 µM CGP35348, a GABAB receptor antagonist in the recording solution. In all cases, standard deviations are indicated at statistical data.
Rat experiments were performed with the approval of the University of Szeged and in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals. Young (P20–30) Wistar rats were anaesthetized by the intraperitoneal injection of ketamine (30 mg∕kg) and xylazine (10 mg∕kg), and following decapitation coronal slices (350 µm thick) were prepared from their somatosensory cortex. Further experimental procedures on rat tissue were performed as detailed above for the human.
Histology
Visualization of biocytin was performed as described previously (Tamas et al., 1997 ). Three-dimensional light microscopic reconstructions were performed using Neurolucida (MicroBrightField, Williston, VT) with 100 × objective. Correlated light microscopy were performed as described previously (Szabadics et al., 2001 ; Tamas et al., 1997 ).
Results
Similar to our earlier experiments, the postsynaptic IPSPs elicited by neurogliaform cells were highly sensitive to the firing rate of the presynaptic cells (Tamas et al., 2003 ). We activated the presynaptic neurogliaform cells with single action potentials delivered at various intervals, and stable amplitude of postsynaptic responses could only be achieved if the interval between presynaptic spikes was at least 1.5 minute. Accordingly, all single action potential-evoked responses were collected at especially low presynaptic firing rates (one spike in 90 second).
Human experiments
We recorded 13 connections in layers 2∕3 in slices derived from human association cortices in which the presynaptic neurons were neurogliaform cells. Like the neurogliaform cells identified previously (Simon et al., 2005 ; Tamas et al., 2003 ), all neurogliaform cells showed similar anatomical features (Figures 1 Bc and 2 A Their somata were relatively small and mostly spherical. Multiple dendrites radiated from all parts of the soma and were arranged to a relatively small dendritic field. Small boutons frequently occurred along thin axonal collaterals, forming a dense meshwork within the compact axonal field.
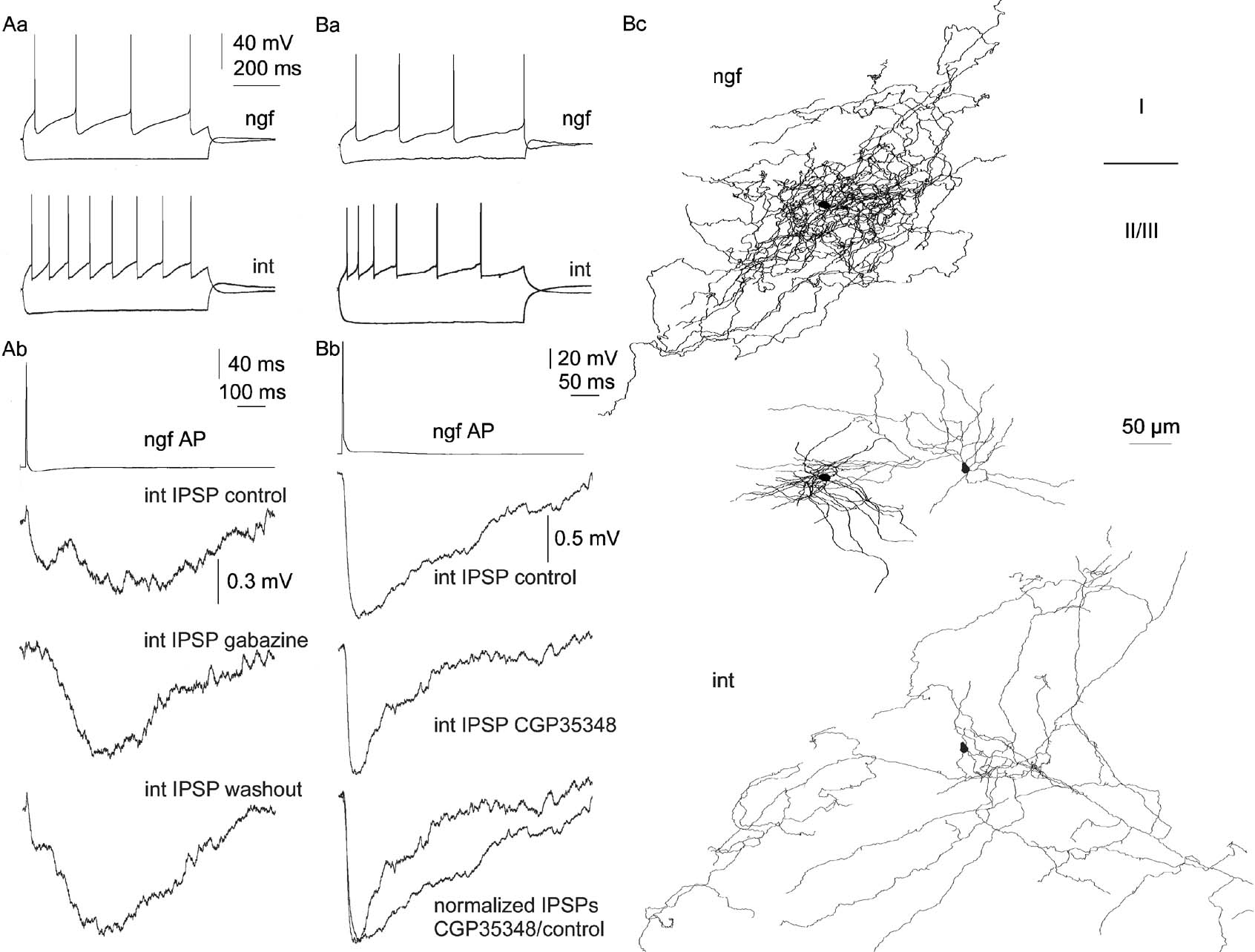
Figure 1. The effect of human neurogliaform cells is mediated by GABAA and GABAB receptors on postsynaptic interneurons. (Aa) Firing pattern of a human neurogliaform cell (ngf) and a postsynaptic interneuron (int). (Ab) Single action potentials of the neurogliaform cell elicited biphasic IPSPs in the interneuron in control conditions (top). The initial component of the IPSP was reversibly blocked by the GABAA receptor antagonist gabazine (10 µM, middle and bottom). (Ba) Firing properties of a presynaptic neurogliaform cell (ngf) and postsynaptic interneuron (int) recorded in human association cortex. (Bb) The postsynaptic cell responded with a long-lasting IPSP to single action potentials evoked in the neurogliaform cell (top). The decay of IPSP could be shortened by the GABAB receptor blocker CGP35348 (60 µM, middle and bottom). (Bc) Three-dimensional light microscopic reconstructions of the presynaptic neurogliaform cell (black) and the postsynaptic interneuron (gray). Axonal (top and bottom) and dendritic (middle) reconstructions are separated for clarity, the relative position of the two cells is shown by the dendritic arborizations.
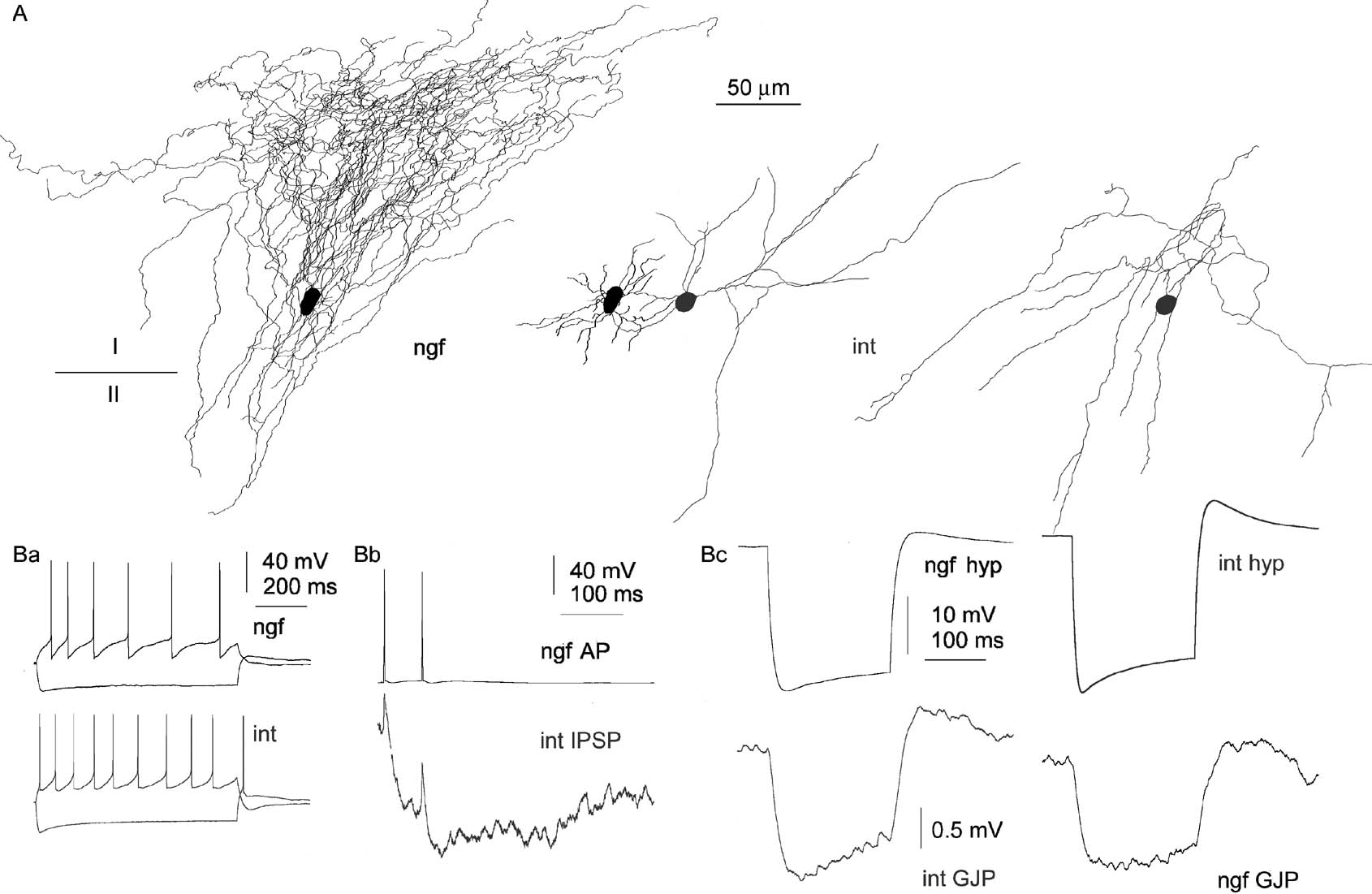
Figure 2. Human neurogliaform cells are involved in electrical coupling. (A) Three-dimensional light microscopic reconstructions of an electrically coupled neurogliaform cell (ngf, black) and a different type of interneuron (int, gray) in layer I of the human cerebral cortex. Axonal (left and right) and dendritic (middle) reconstructions are separated for clarity, the dendritic arborizations show the original position of the interneurons. (Ba) Firing patterns of the neurogliaform cell and the interneuron. (Bb) Action potentials in the neurogliaform cell triggered postsynaptic spikelets in the interneuron followed by long-lasting IPSPs. (Bc) Reciprocal transmission of hyperpolarizing current pulses injected into the neurogliaform cell (left) or the interneuron (right) confirmed electrical coupling between the two cells.
In general, firing parameters of human neurogliaform cells were similar to what was described previously in the rat. Neurogliaform cells in the human sample responded to long (800 ms) suprathreshold current injections with late spiking firing pattern or with an accelerating train of spikes when activated from resting membrane potential (−62.4 ± 4.7 mV, Figures 1 Aa, 1 Ba, and 2 Ba). This firing behavior was changed to a decelerating pattern when applying larger current pulses. The time constant and the input resistances of human neurogliaform cells were 8.7 ± 1.5 ms and 219.1 ± 55.1 MΩ, respectively. Human neurogliaform cells showed a characteristic sag in response to hyperpolarizing current injections, which was not typical of neurogliaform cells in rat cerebral cortex (Figures 1 Aa, 1 Ba, and 2 Ba). Postsynaptic cells were pyramidal cells (n = 4) and various interneurons (n = 7) in the human sample. We identified these cells based on firing characteristics and dendritic and axonal morphology. All four postsynaptic pyramidal cells showed regular spiking firing pattern, their input resistances and membrane time constants were 150.5 ± 42.1 MΩ) and 11.9 ± 1.5 ms, respectively. Electrophysiological properties of postsynaptic interneurons were variable; input resistances and time constants were 255.3 ± 88.2 MΩ) and 13.3 ± 6.9 ms, respectively (Figures 1 Aa, 1 Ba, and 2 Ba). Five out of seven postsynaptic human interneurons responded with a sag to hyperpolarizing current injections and, in two cells showed rebound spikes following the hyperpolarization (Figure 2 Ba). The sag and rebound spiking was apparent in postsynaptic human interneurons exhibiting various firing behavior (i.e., fast spiking and accommodating) in response to depolarizing current injections.
Human neurogliaform cells evoked long-lasting inhibition in postsynaptic cells (Figures 1 Ab, 1 Bb, and 2 Bb). The rise times and half-width of IPSPs in these connections were 16.5 ± 8.5 and 184.9 ± 96.5 ms, respectively. When comparing IPSPs arriving to postsynaptic pyramidal cells (not shown) and interneurons in the human sample, we could not detect significant differences in 10–90% rise times and half-widths (20.1 ± 9.8 ms and 12.7 ± 6.8; 230.4 ± 64.6 ms and 148.6 ± 108.4, respectively). For comparison, 10–90% rise times and half-width of IPSPs evoked by neurogliaform cells in rat pyramidal cells were 23.4 ± 9.8 ms and 183.9 ± 82.5 (Tamas et al., 2003 ). The maximal amplitudes of IPSPs evoked by human neurogliaform cells were 0.94 ± 0.88 mV.
We applied pharmacological blockers to dissect the contribution of the two GABA receptor subtypes to the inhibition evoked by human neurogliaform cells. Addition of the GABAB receptor blocker CGP35348 (60 µM) into the bath solution could shorten the duration of slow IPSPs evoked by neurogliaform cells (n = 2; postsynaptic cells were interneurons, Figure 1 Bb. When testing the connections by applying the GABAA receptor blocker gabazine (10 µM), we could isolate the slow component of the IPSPs (n = 2; postsynaptic cells were interneurons, Figure 1 Ab).
In addition to chemical neurotransmission elicited by human neurogliaform cells, we detected homologous electrical connections from neurogliaform cells to other neurogliaform cells (n = 2, not shown) and a heterologous electrical connection between a neurogliaform cell and a different type of interneuron which was combined with a slow IPSP triggered by the neurogliaform cell (Figure 2 ). Electrical synapses were detected as spikelets in the postsynaptic cells in response to presynaptic action potentials (Figure 2 Bb). When injecting hyperpolarizing current pulses into either of the connected cells, the other cell responded with an electrical coupling potential of similar polarity (Figure 2 Cc) confirming electrical interaction. The strength of electrical coupling was symmetrical in the homologous and heterologous electrical synapses showing subthreshold coupling coefficients of 0.036 ± 0.004.
Rat experiments
Following our findings in the human cortex based on a relatively limited sample, we performed supporting experiments in slices taken from the rat cerebral cortex. We recorded 48 pairs of neurons in which neurogliaform cells established chemical connections with various types of interneuron (neurogliaform cells, regular spiking cells, fast spiking basket cells, fast spiking axo-axonic cells, and unclassified interneurons). We identified these cells based on their firing characteristics and by light microscopic evaluation of dendritic and axonal morphology.
The input resistances of postsynaptic neurogliaform cells (n = 12) were 182.7 ± 55.2 MΩ) (Figure 3 A). The time constants of neurogliaform cells in our sample were 9.9 ± 1.1 ms. There was no significant difference between time constants of human and rat neurogliaform cells (p = 0.792). Input resistance in rat neurogliaform cells was significantly lower than that of in the human (p < 0.01). Neurogliaform cells responded with long-lasting IPSPs to single action potentials of presynaptic neurogliaform cells (Figure 3 B). The rise times and half-width of IPSPs of neurogliaform cells were 6.5 ± 1.5 and 44.4 ± 10.6 ms. To determine the extent of GABAB receptor contribution to IPSPs elicited by neurogliaform cells, we blocked GABAB receptor-mediated component with CGP35348 in three neurogliaform cell to neurogliaform cell connections (Figures 3 B and 3 C). The decay phase of neurogliaform cell to neurogliaform cell IPSPs were shortened by CGP35348 resulting in a 16 ± 7.9% decrease in the half-width of IPSPs showing the involvement of GABAB receptors in these connections.
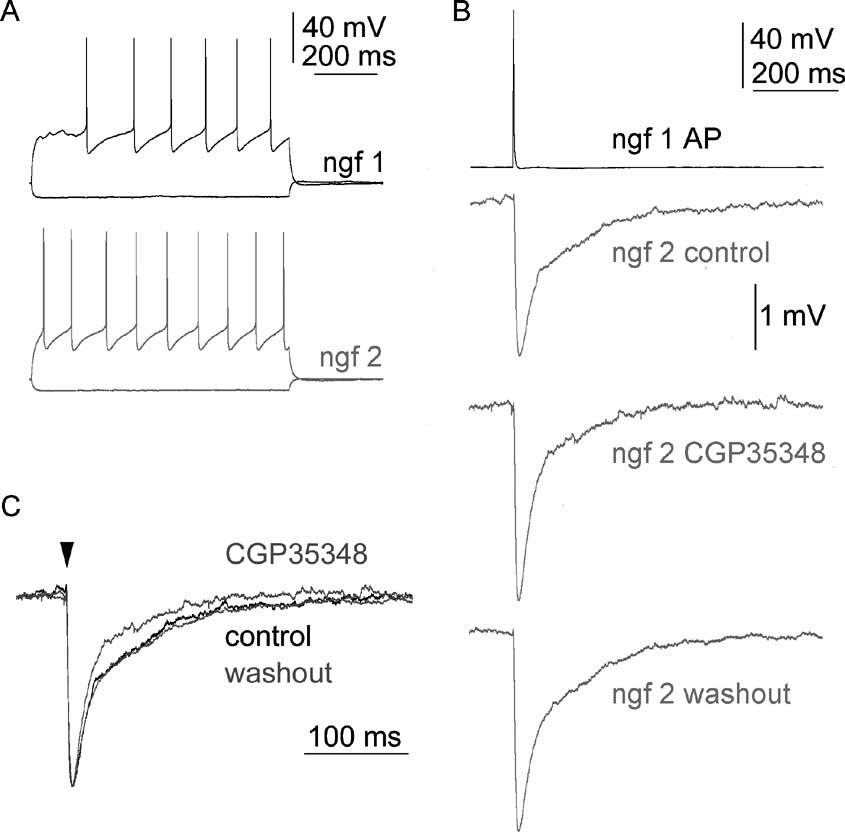
Figure 3. GABAB receptors contribute to IPSPs between neurogliaform cells in the rat cortex. (A) Firing patterns of two neurogliaform cells (ngf 1 and ngf 2). (B) Single action potentials in the presynaptic neurogliaform cell elicited slow IPSPs in the postsynaptic neurogliaform cell. The decay of the IPSP could be shortened by the applicaion of the GABAB receptor blocker CGP35348 (60 µM). The late component of IPSP showed recovery after 30 minute of washout. (C) Superimposed traces are shown normalized to the control IPSP.
Neurogliaform cell innervated postsynaptic interneurons (n = 12) that responded to suprathreshold current injections with a regular-spiking firing pattern with accomodating train of spikes (Figure 4 B). The input resistance and membrane time constant of regular spiking nonpyramidal cells was 249.1 ± 31.2 MΩ) and 13.7 ± 10.4 ms, respectively. Their anatomical features were similar to regular spiking nonpyramidal cells identified by previous studies (Kawaguchi and Kubota, 1997 ; Krimer et al., 2005 ; Szabadics et al., 2001 ). The morphology of sparsely spiny dendrites and individual axonal branches of these cells with relatively large boutons were in line with their physiological similarity (Figure 4 A). The IPSPs from neurogliaform cells to regular spiking interneurons were the longest among neurogliaform to interneuron connections (p < 0.002 for neurogliaform to neurogliaform, p < 0.001 for neurogliaform to fast spiking cell connections, see below; Figure 4 C). The differences detected between the duration of IPSPs triggered by neurogliaform cells on regular spiking, neurogliaform and fast spiking cells could not be accounted for differences in postsynaptic membrane time constants. The rise time and half-width of IPSPs in neurogliaform to regular spiking nonpyramidal cell connections were 16.6 ± 10.8 and 135.7 ± 51.1 ms. Application of the GABAA receptor blocker gabazine (n = 4) and the GABAB receptor blocker CGP35348 (n = 2) showed that neurogliaform to regular spiking nonpyramidal cell connections were composed of GABAA and GABAB receptor-mediated components (Figure 4 C). When GABAA receptors were blocked, we could detect residual hyperpolarizing responses with a late onset (41.8 ± 2 ms) and slow decay times. Blockade of GABAB receptors could shorten the duration of IPSPs by 42.9 ± 21.1%.
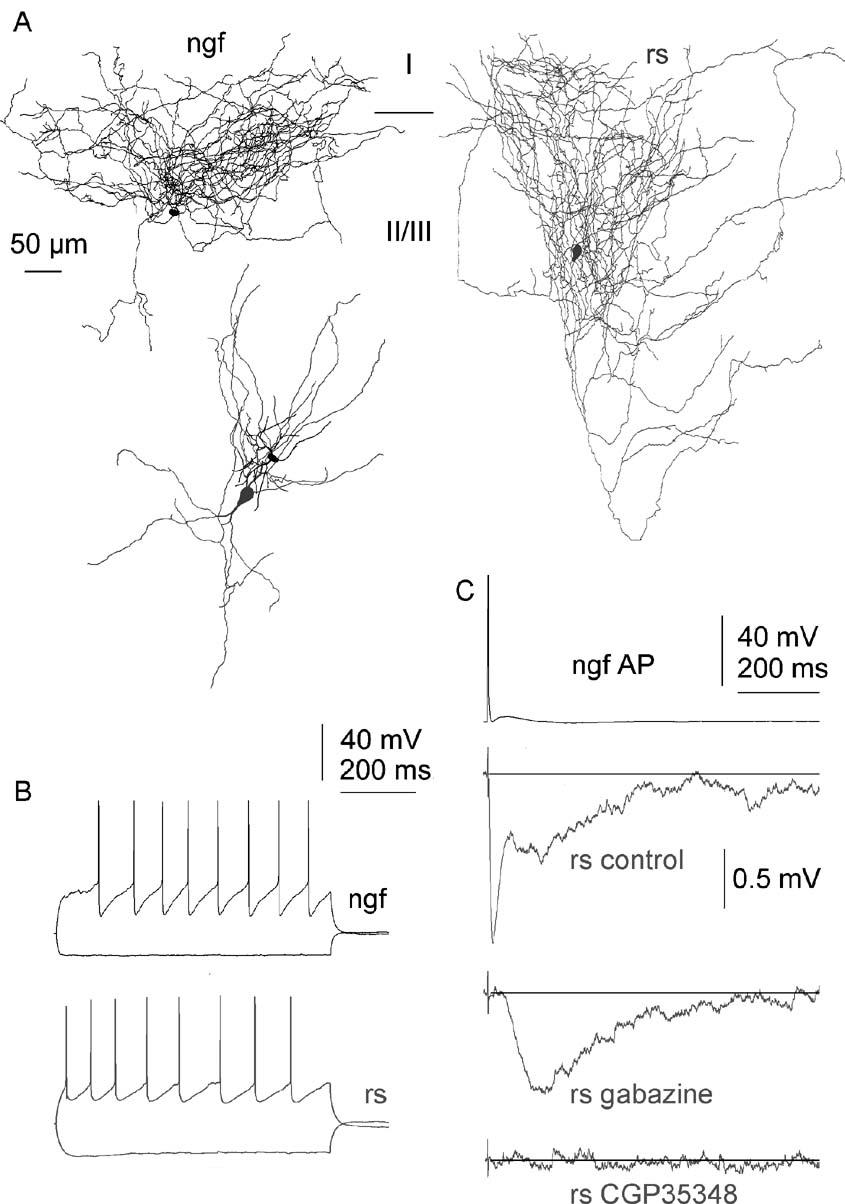
Figure 4. GABAA and GABAB receptors mediate neurogliaform cells (ngfc) to regular spiking interneuron (rs) connections. (A) Three-dimensional light microscopic reconstructions of the presynaptic neurogliaform cell (black) and the postsynaptic regular spiking interneuron (gray). Axonal (left and right) and dendritic (middle) reconstructions are separated for clarity, the relative position of the two cells is shown by the dendritic arborizations. (B) Firing patterns of the two cells. (C) Single action potentials in the neurogliaform cell elicited slow IPSPs with two components in the regular spiking interneuron. The early component could be blocked by the GABAA receptor blocker gabazine (10 µM) and the late phase of IPSP was abolished by the subsequent addition of CGP35348 (60 µM).
Postsynaptic fast spiking cells responded with high frequency, nonaccomodating trains of spikes to depolarizing current pulses (Figure 5 A). Axonal terminals of eight postsynaptic fast spiking cells showed close appositions on unlabeled somata, thus we classified these neurons as basket cells (Karube et al., 2004 ). In the remaining two cases, axon terminals of the postsynaptic cells were arranged into cartridges or candles characteristic to chandelier or axo-axonic cells. Input resistance and membrane time constants of basket and axo-axonic fast spiking interneurons were similar, thus we pooled the data (110.8 ± 25.2 and 7.1 ± 1.2 ms, respectively). The kinetics of neurogliaform to basket and neurogliaform to axo-axonic cell connections were also similar and showed relatively small slow component (Figure 5 B). The rise times and half-width of IPSPs elicited by neurogliaform cells in fast spiking cells were 3.5 ± 0.9 and 25.8 ± 14.6 ms. The IPSPs from neurogliaform cells to fast spiking interneurons were the shortest among neurogliaform to interneuron connections (p < 0.03 for neurogliaform to neurogliaform, p < 0.001 for neurogliaform to regular spiking cell connections). Pharmacological dissection could be performed in three neurogliaform to basket cell connections. A small residual IPSP component was detected during GABAA receptor blockade with gabazine (n = 2, 10 µM, Figure 5 B) and, when blocking GABAB receptor activation with CGP35348 (n = 1, 60 µM), the half-width of response changed only slightly (from 72 to 62.3 ms).
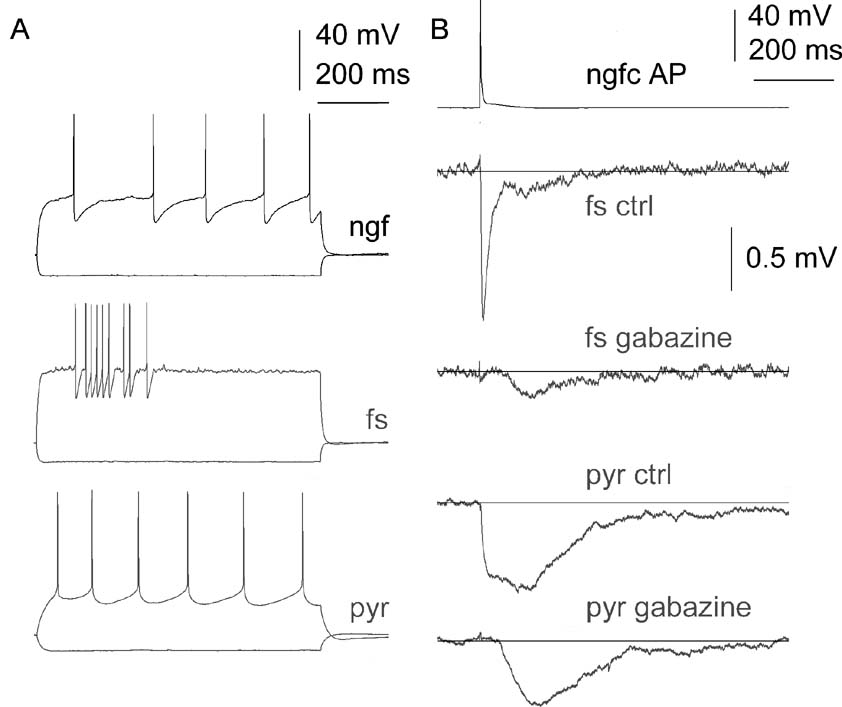
Figure 5. Divergent connections of a neurogliaform cell (ngfc) to postsynaptic fast spiking (fs) and pyramidal (pyr) cells. (A), Firing properties of the recorded cells. (B), Single action potentials in the neurogliaform cell elicited IPSPs with complex kinetics in the fast spiking as well as in the pyramidal cell. A residual slow component was detectable in both postsynaptic neurons during application of the GABAA receptor blocker gabazine (10 µM).
Discussion
The first simultaneous multiple recordings of human neurogliaform cells and their postsynaptic targets revealed that single spikes in neurogliaform cells elicit long-lasting unitary IPSPs composed of GABAA and GABAB receptor-mediated components in various types of interneuron. We confirmed these results in the rat cortex and, moreover, our experiments showed that human neurogliaform cells, similar to those tested in the rat (Tamas et al., 2003 ), evoke long-lasting IPSPs in pyramidal cells. These results suggest that neurogliaform cells represent a widespread source for slow IPSPs targeting multiple cell types in microcircuits of the human and rat neocortex and are in line with earlier experiments showing a relatively small GABAB receptor component in unitary connections between neurogliaform cells of the hippocampus (Price et al., 2005 ).
The effect of neurogliaform cells in the rat (Tamas et al., 2003 ; Vida et al., 1998 ) and human (Kisvarday et al., 1990 ) cerebral cortex is targeted towards the dendritic compartment of the postsynaptic cells arriving to dendritic spines, spine necks, and dendritic shafts but some evidence suggests that postsynaptic somata might also receive synapses from neurogliaform cells (Kawaguchi and Kubota, 1997 ). Dendritic spines receive the majority of glutamatergic inputs arriving to cortical pyramidal cells and simulations suggested that inhibition arriving to spines is more effective through GABAB than GABAA receptors in reducing EPSPs through powerful a hyperpolarizing effect (Qian and Sejnowski, 1990 ; Tamas et al., 2003 ). Metabotropic GABAB receptors are coupled to G-protein-coupled inwardly rectifying K+ channels (Kir3 channels) (Calver et al., 2002 ). Quantitative analysis of immunogold particles labeling Kir3.2 revealed an enrichment of the protein around putative glutamatergic synapses on dendritic spines, similar to that of GABAB receptors and a high degree of co-clustering of Kir3.2 and GABA(B1) was revealed around excitatory synapses by freeze-fracture replica immunolabeling (Kulik et al., 2006 ). Interestingly, GABAB receptors and Kir3.2 channels were found to be mainly segregated on dendritic shafts (Kulik et al., 2006 ) and thus the postsynaptic machinery of neurogliaform input on dendritic shafts of pyramidal cells and on smooth dendrites of GABAergic interneurons known to express GABAB receptors (Kulik et al., 2003 ; Lopez-Bendito et al., 2002 ) is not clear. However, more recent experiments provide evidence for a differential molecular composition of presynaptic and postsynaptic GABAB receptors, but dendritic receptors were found to contain the GABAB1b isoform in the hippocampus and in the neocortex (Perez-Garci et al., 2006 ; Vigot et al., 2006 ) suggesting a similar mechanism of action on dendritic shafts and spines for neurogliaform cells. Postsynaptic GABAB receptor-mediated inhibition can powerfully depress Ca2+-spikes in apical dendrites of neocortical pyramidal cells (Perez-Garci et al., 2006 ). Back-propagating action potentials modulate Ca2+-signaling in hippocampal and neocortical interneurons (Goldberg et al., 2003a ; Goldberg et al., 2003b ; Kaiser et al., 2001 ; Saraga et al., 2003 ; Wang and Kelly, 2001 ) and thus GABAB receptor-mediated inhibition of neocortical interneurons by neurogliaform cells can interact with dendritic Ca2+-dynamics in GABAergic cells.
The cooperation between GABAB receptors and dendritic excitability might be influenced by the differential availability of GABAB receptors on postsynaptic GABAergic dendrites. The differences detected between the duration of IPSPs triggered by neurogliaform cells on regular spiking, neurogliaform, and fast spiking cells suggest a minor relative contribution of GABAB receptors on fast spiking basket and axo-axonic cells compared to postsynaptic regular spiking and neurogliaform cells. Heterogeneous GABAB receptor involvement in IPSCs from various types of GABAergic cells were recorded in response to a molecular layer stimulus in hippocampal dentate-hylus border interneurons (Mott et al., 19999 ). In the neocortex, synaptically released and exogenous GABA evoked IPSPs were recorded without a detectable GABAB component in fast spiking cells whereas compound GABAA and GABAB receptor-mediated responses were found in regular spiking cells (Martina et al., 2001 ). Although our method of recording unitary IPSPs elicited by neurogliaform cells allowed the identification of GABAB receptors expressed by fast spiking basket cells, it appears that the actual ratio of GABAA and GABAB receptor involvement in the output of neurogliaform cells could be connection specific and primarily determined by the postsynaptic cell type.
Our experiments represent the first electrical synapses recorded electrophysiologically in the human cerebral cortex. Although electrical coupling is firmly established between cortical interneurons in juvenile animals (Bennett and Zukin, 2004 ; Connors and Long, 2004 ; Galarreta and Hestrin, 2001 ; Sohl et al., 2005 ), electrophysiological evidence for electrical synapses from adult cerebral cortex is very limited (Galarreta et al., 2004 ) due to technical difficulties, i.e., the quality of images using infrared videomicroscopy is influenced by axonal myelination resulting in a decreased transparency of slices. Nonpyramidal cells form gap junctions in the adult hippocampus (Kosaka and Hama, 1985 ) and neocortex (Peters, 1980 ; Sloper, 1972 ; Tamas et al., 2000 ), but the probability of dye coupling and gap junctions is reportedly affected by development (Connors et al., 1983 ; Peinado et al., 1993 ). Our recordings demonstrate that simultaneous quadruple recordings from visually identified neurons can be performed in the adult human cortex and that electrical coupling is present between mature human interneurons. The human recordings are in line with data obtained in the rat showing that human neurogliaform cells form homologous electrical synapses with other neurogliaform cells (Chu et al., 2003 ; Price et al., 2005 ; Simon et al., 2005 ) and, moreover, human neurogliaform cells can establish heterologous electrical coupling with other types of interneuron (Simon et al., 2005 ; Zsiros and Maccaferri, 2005 ). We provide evidence that long-lasting IPSPs are combined with homologous and heterologous electrical coupling between neurogliaform cells and several human and rat interneuron types. Synaptic potentials propagate through electrically coupled networks of interneurons (Zsiros et al., 2007 ), and the slow kinetics of neurogliaform cell triggered IPSPs are particularly well suited for effectively spreading in the network through electrical synapses which act as low pass filters. Neurogliaform cells represent a unique element in human and rat microcircuits of the cerebral cortex. Embedded into an extensive network of homologous and heterologous electrical synapses linking several interneuron classes, neurogliaform cells are able to monitor the sub- and suprathreshold activity of coupled neurons and can transform this activity to long-lasting chemical signaling through metabotropic GABAB receptors on multiple neuron populations.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgements
This study was funded by the European Young Investigator Awards, National Institutes of Health NS-35915, the Wellcome Trust, the Hungarian Academy of Sciences, the Hungarian National Office for Research and Technology Grants KFKT-1-2006-009 and RET008/2004, the Hungarian Scientific Research Fund Grants T049535 and TS049868, Howard Hughes Medical Institute Grant 55005625, and Boehringer Ingelheim Fonds.
References
Bennett, M. V., and Zukin, R. S. (2004). Electrical coupling and neuronal synchronization in the mammalian brain. Neuron 41, 495–511.
Buhl, E. H., Halasy, K., and Somogyi, P. (1994). Diverse sources of hippocampal unitary inhibitory postsynaptic potentials and the number of synaptic release sites. Nature 368, 823–828.
Cajal, S. R. y. (1904). Textura del Systema Nervioso del Hombre y los Vertebrados (Madrid, Spain, Moya), Vol. 2.
Calver, A. R., Davies, C. H., and Pangalos, M. (2002). GABA(B) receptors: from monogamy to promiscuity. Neurosignals 11, 299–314.
Cauli, B., Porter, J. T., Tsuzuki, K., Lambolez, B., Rossier, J., Quenet, B., and Audinat, E. (2000). Classification of fusiform neocortical interneurons based on unsupervised clustering. Proc. Natl. Acad. Sci. USA 97, 6144–6149.
Chu, Z., Galarreta, M., and Hestrin, S. (2003). Synaptic interactions of late-spiking neocortical neurons in layer 1. J. Neurosci. 23, 96–102.
Connors, B. W., Benardo, L. S., and Prince, D. A. (1983). Coupling between neurons of the developing rat neocortex. J. Neurosci. 3, 773–782.
Connors, B. W., and Long, M. A. (2004). Electrical synapses in the mammalian brain. Annu. Rev. Neurosci. 27, 393–418.
Galarreta, M., Erdelyi, F., Szabo, G., and Hestrin, S. (2004). Electrical coupling among irregular-spiking GABAergic interneurons expressing cannabinoid receptors. J. Neurosci. 24, 9770–9778.
Galarreta, M., and Hestrin, S. (2001). Electrical synapses between GABA-releasing interneurons. Nat. Rev. Neurosci. 2, 425–433.
Goldberg, J. H., Tamas, G., Aronov, D., and Yuste, R. (2003a). Calcium microdomains in aspiny dendrites. Neuron 40, 807–821.
Goldberg, J. H., Yuste, R., and Tamas, G. (2003b). Ca2+ imaging of mouse neocortical interneurone dendrites: Contribution of Ca2+-permeable AMPA and NMDA receptors to subthreshold Ca2+dynamics. J. Physiol. (Lond.) 551, 67–78.
Gupta, A., Wang, Y., and Markram, H. (2000). Organizing principles for a diversity of GABAergic interneurons and synapses in the neocortex. Science 287, 273–278.
Hestrin, S., and Armstrong, W. E. (1996). Morphology and physiology of cortical neurons in layer I. J. Neurosci. 16, 5290–5300.
Kaiser, K. M., Zilberter, Y., and Sakmann, B. (2001). Back-propagating action potentials mediate calcium signalling in dendrites of bitufted interneurons in layer 2∕3 of rat somatosensory cortex. J. Physiol. 535, 17–31.
Karube, F., Kubota, Y., and Kawaguchi, Y. (2004). Axon branching and synaptic bouton phenotypes in GABAergic nonpyramidal cell subtypes. J. Neurosci. 24, 2853–2865.
Kawaguchi, Y., Karube, F., and Kubota, Y. (2006). Dendritic branch typing and spine expression patterns in cortical nonpyramidal cells. Cereb. Cortex 16, 696–711.
Kawaguchi, Y., and Kubota, Y. (1997). GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cereb. Cortex 7, 476–486.
Kisvarday, Z. F., Gulyas, A., Beroukas, D., North, J. B., Chubb, I. W., and Somogyi, P. (1990). Synapses, axonal and dendritic patterns of GABA-immunoreactive neurons in human cerebral cortex. Brain 113, 793–812.
Kosaka, T., and Hama, K. (1985). Gap junctions between non-pyramidal cell dendrites in the rat hippocampus (CA1 and CA3 regions): a combined Golgi-electron microscopy study. J. Comp. Neurol. 231, 150–161.
Krimer, L. S., Zaitsev, A. V., Czanner, G., Kroner, S., Gonzalez-Burgos, G., Povysheva, N. V., Iyengar, S., Barrionuevo, G., and Lewis, D. A. (2005). Cluster analysis-based physiological classification and morphological properties of inhibitory neurons in layers 2–3 of monkey dorsolateral prefrontal cortex. J. Neurophysiol. 94, 3009–3022.
Kulik, A., Vida, I., Fukazawa, Y., Guetg, N., Kasugai, Y., Marker, C. L., Rigato, F., Bettler, B., Wickman, K., Frotscher, M., and Shigemoto, R. (2006). Compartment-dependent colocalization of Kir3.2-containing K+ channels and GABAB receptors in hippocampal pyramidal cells. J. Neurosci. 26, 4289–4297.
Kulik, A., Vida, I., Lujan, R., Haas, C. A., Lopez-Bendito, G., Shigemoto, R., and Frotscher, M. (2003). Subcellular localization of metabotropic GABA(B) receptor subunits GABA(B1a∕b) and GABA(B2) in the rat hippocampus. J. Neurosci. 23, 11026–11035.
Lopez-Bendito, G., Shigemoto, R., Kulik, A., Paulsen, O., Fairen, A., and Lujan, R. (2002). Expression and distribution of metabotropic GABA receptor subtypes GABABR1 and GABABR2 during rat neocortical development. Eur. J. Neurosci. 15, 1766–1778.
Martina, M., Royer, S., and Pare, D. (2001). Cell-type-specific GABA responses and chloride homeostasis in the cortex and amygdala. J. Neurophysiol. 86, 2887–2895.
Mott, D. D., Li, Q., Okazaki, M. M., Turner, D. A., and Lewis, D. V. (1999). GABAB-Receptor-mediated currents in interneurons of the dentate-hilus border. J. Neurophysiol. 82, 1438–1450.
Peinado, A., Yuste, R., and Katz, L. C. (1993). Extensive dye coupling between rat neocortical neurons during the period of circuit formation. Neuron 10, 103–114.
Perez-Garci, E., Gassmann, M., Bettler, B., and Larkum, M. E. (2006). The GABA(B1b) isoform mediates long-lasting inhibition of dendritic Ca(2+) spikes in layer 5 somatosensory pyramidal neurons. Neuron 50, 603–616.
Peters, A. (1980). Morphological correlates of epilepsy: cells in the cerebral cortex. Adv. Neurol. 27, 21–48.
Price, C. J., Cauli, B., Kovacs, E. R., Kulik, A., Lambolez, B., Shigemoto, R., and Capogna, M. (2005). Neurogliaform neurons form a novel inhibitory network in the hippocampal CA1 area. J. Neurosci. 25, 6775–6786.
Qian, N., and Sejnowski, T. J. (1990). When is an inhibitory synapse effective?. Proc. Natl. Acad. Sci. USA 87, 8145–8149.
Sancesario, G., Pisani, A., D'Angelo, V., Calabresi, P., and Bernardi, G. (1998). Morphological and functional study of dwarf neurons in the rat striatum. Eur. J. Neurosci. 10, 3575–3583.
Saraga, F., Wu, C. P., Zhang, L., and Skinner, F. K. (2003). Active dendrites and spike propagation in multi-compartment models of oriens-lacunosum∕moleculare hippocampal interneurons. J. Physiol. 552, 673–689.
Simon, A., Olah, S., Molnar, G., Szabadics, J., and Tamas, G. (2005). Gap-junctional coupling between neurogliaform cells and various interneuron types in the neocortex. J. Neurosci. 25, 6278–6285.
Sloper, J. J. (1972). Gap junctions between dendrites in the primate neocortex. Brain Res. 44, 641–646.
Sohl, G., Maxeiner, S., and Willecke, K. (2005). Expression and functions of neuronal gap junctions. Nat. Rev. Neurosci. 6, 191–200.
Szabadics, J., Lorincz, A., and Tamas, G. (2001). b and g frequency synchronization by dendritic GABAergic synapses and gap junctions in a network of cortical interneurons. J. Neurosci. 21, 5824–5831.
Tamas, G., Buhl, E. H., Lorincz, A., and Somogyi, P. (2000). Proximally targeted GABAergic synapses and gap junctions synchronize cortical interneurons. Nat. Neurosci. 3, 366–371.
Tamas, G., Buhl, E. H., and Somogyi, P. (1997). Fast IPSPs elicited via multiple synaptic release sites by distinct types of GABAergic neuron in the cat visual cortex. J. Physiol. (Lond.) 500, 715–738.
Tamas, G., Lorincz, A., Simon, A., and Szabadics, J. (2003). Identified sources and targets of slow inhibition in the neocortex. Science 299, 1902–1905.
Vida, I., Halasy, K., Szinyei, C., Somogyi, P., and Buhl, E. H. (1998). Unitary IPSPs evoked by interneurons at the stratum radiatum-stratum lacunosum-moleculare border in the CA1 area of the rat hippocampus in vitro. J. Physiol. 506, 755–773.
Vigot, R., Barbieri, S., Brauner-Osborne, H., Turecek, R., Shigemoto, R., Zhang, Y. P., Lujan, R., Jacobson, L. H., Biermann, B., and Fritschy, J. M. et al. (2006). Differential compartmentalization and distinct functions of GABA(B) receptor variants. Neuron 50, 589–601.
Wang, J. H., and Kelly, P. (2001). Calcium-calmodulin signalling pathway up-regulates glutamatergic synaptic function in non-pyramidal, fast spiking rat hippocampal CA1 neurons. J. Physiol. 533, 407–422.
Zsiros, V., Aradi, I., and Maccaferri, G. (2007). Propagation of postsynaptic currents and potentials via gap junctions in GABAergic networks of the rat hippocampus. J. Physiol. 578, 527–544.
Keywords: neurogliaform, cortex, GABAA receptor, GABAB receptor, human
Citation: Szabolcs Oláh, Gergely Komlósi, János Szabadics, Csaba Varga, Éva Tóth, Pál Barzó,and Gábor Tamás (2007). Output of neurogliaform cells to various neuron types in the human and rat cerebral cortex. Front. Neural Circuits 1:4. doi: 10.3389/neuro.04/004.2007
Received: 24 August 2007;
Paper pending published: 17 September 2007;
Accepted: 15 October 2007;
Published online: 2 November 2007
Edited by:
Rafael Yuste, Columbia University, New York City, USAReviewed by:
Erin M. Schuman, Caltech/HHMI, Pasadena, USACopyright: © 2007 Oláh, Komlósi, Szabadics, Varga, Tóth, Barzó, Tamás. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Gábor Tamás, HAS Research Group for Cortical Microcircuits, Department of Comparative Physiology, University of Szeged, Közép fasor 52., Szeged, H-6726, Hungary. e-mail:Z3RhbWFzQGJpby51LXN6ZWdlZC5odQ==