
- 1 Unité de Génétique Moléculaire Animale-INRA UMR 06, University of Limoges, Limoges, France
- 2 Department of Human Anatomy and Psychobiology and CIBER en Enfermedades Raras, Unit 736, Medical School, University of Murcia, Murcia, Spain
Sonic hedgehog (SHH) is a secreted signaling factor that is implicated in the molecular patterning of the central nervous system (CNS), somites, and limbs in vertebrates. SHH has a crucial role in the generation of ventral cell types along the entire rostrocaudal axis of the neural tube. It is secreted early in development by the axial mesoderm (prechordal plate and notochord) and the overlying ventral neural tube. Recent studies clarified the impact of SHH signaling mechanisms on dorsoventral patterning of the spinal cord, but the corresponding phenomena in the rostral forebrain are slightly different and more complex. This notably involves separate Shh expression in the preoptic part of the forebrain alar plate, as well as in the hypothalamic floor and basal plates. The present work includes a detailed spatiotemporal description of the singular alar Shh expression pattern in the rostral preoptic forebrain of chick embryos, comparing it with FoxG1, Dlx5, Nkx2.1, and Nkx2.2 mRNA expression at diverse stages of development. As a result of this mapping, we report a subdivision of the preoptic region in dorsal and ventral zones; only the dorsal part shows Shh expression. The positive area impinges as well upon a median septocommissural preoptic domain. Our study strongly suggests tangential migration of Shh-positive cells from the preoptic region into other subpallial domains, particularly into the pallidal mantle and the intermediate septum.
Introduction
The central nervous system (CNS) is regionalized along anteroposterior (AP) and dorsoventral (DV) axes. According to the prosomeric model, the secondary prosencephalon (rostral forebrain; Puelles and Rubenstein, 2003; Puelles et al., 2004, 2007) includes the entire telencephalon as an evaginated dorsal part of the alar plate; on the basis of shared molecular specification properties it is currently thought that the telencephalic subpallium includes the non-evaginated preoptic area (POA). Underneath the telencephalic stalk lies the hypothalamus (rest of the alar plate and subjacent basal and floor plates). The subpallium is divided structurally in four distinct parallel territories disposed parallel to the septoamygdaloid dimension (Puelles et al., 2000). These are known as striatum, pallidum, anterior entopeduncular area and preoptic area (St, Pal, AEP, POA) and adopt a nested configuration (Puelles et al., 2000; Puelles and Rubenstein, 2003). In agreement with a recent terminological proposal of Puelles in the Allen Developmental Atlas of the Mouse Brain1 we will use here the term “diagonal domain” (Dg) instead of the confusing “anterior entopeduncular area” term formerly employed in all versions of the prosomeric model. The Dg/AEP concept essentially refers to a full radial histogenetic domain intercalated at the telencephalic stalk between Pal and POA. It extends from the neighborhood of the medial septum back to the subpallial amygdala. The classic innominate area encompassing the basal magnocellular cell population forms its intermediate stratum, and the diagonal band nuclei together with the associated tract occupy its superficial stratum. At periventricular levels, the Dg contains a medial part of the complex of the bed nucleus of the stria terminalis (Bst), while the corresponding lateral part belongs to the adjacent pallidal domain.
The entire subpallial territory is often referred to as “basal forebrain,” although it belongs embryologically to the alar plate. Indeed, as shown by fate mapping at neural plate stage (Cobos et al., 2001), the four subpallial territories extend dorsally (topologically) into the septum, identified at the rostral rim of the neural plate, whose median fused commissural part represents, after closure of the rostral neuropore, a rostral part of the telencephalic roof plate, rostral to the telencephalic choroidal tela (Bulfone et al., 1993; Puelles et al., 2000; Cobos et al., 2001; Marín and Rubenstein, 2002; Puelles and Rubenstein, 2003; Pombero and Martinez, 2009).
A detailed recent analysis of molecularly distinct progenitor areas within the mouse subpallium identified 16 differentially specified units, each theoretically capable of producing unique cell types (Flames et al., 2007). The Dg domain was listed there tentatively as a part of the pallidal complex, namely the pMGE5 domain. Because of such structural complexity and the variety of migratory movements of postmitotic neurons born at a given progenitor area that migrate tangentially to neighboring or distant telencephalic domains, the developmental derivation of many specific types of neurons within the subpallial domain remains still uncertain.
The ventricular zone of the Dg was reported to be one of the subpallial sources of calcium binding protein-containing neurons destined to the pallial amygdala (Legaz et al., 2005). It is also the major source of basal cholinergic neurons (Marín and Rubenstein, 2002; Zhao et al., 2003; Mori et al., 2004), of striatal interneurons that express the NKX2.1 homeodomain protein (Marín et al., 2000) and of some GABAergic cortical interneurons (Taglialatela et al., 2004). On the other hand, early oligodendrocyte progenitors were reported to arise in the Pal and Dg under the influence of Sonic hedgehog (SHH) signals (Perez-Villegas et al., 1999; Olivier et al., 2001; Cobos et al., 2001; Spassky et al., 2001; Tekki-Kessaris et al., 2001; Kessaris et al., 2006).
The complex anatomical organization of the brain is anticipated by molecular regionalization of the neuroepithelium at early developmental stages, a patterning process that precedes differential genoarchitectonic development of the mantle layer. Combined expression of diverse transcription factors in the neural plate starts to create a grid of molecularly distinct neuroepithelial progenitor regions, which soon acquire specific developmental fates (Lumsden and Krumlauf, 1996; Rubenstein and Beachy, 1998; Rubenstein et al., 1998; Puelles et al., 2005; Flames et al., 2007; Sánchez-Arrones et al., 2009). Some aspects of neural patterning are controlled by secreted extracellular signaling molecules that spread over variable distances across the neuroepithelium, notably SHH, of the Hedgehog family (HH), and members of the Bone Morphogenetic Protein family (BMP) (Patten and Placzek, 2000; Briscoe and Ericson, 2001; Ulloa and Briscoe, 2007). Interactions between SHH and BMPs are important for the specification of dorsal and intermediate DV cell types. The activities of these signals are integrated spatiotemporally with other signals (e.g., FGFs, Wnts, retinoic acid) to determine the specific combinations of transcription factors that are activated in distinct AP and DV compartments of the brain.
Sonic hedgehog is widely held to act principally as a ventralizing signal and BMPs are considered to be dorsalizing signals. However, BMPs also modulate ventral patterning effects in the hypothalamus (Ohyama et al., 2008), and SHH released from the diencephalic zona limitans intrathalamica (an alar interprosomeric landmark) is known to exert patterning effects upon the thalamic and prethalamic alar plate domains (Kiecker and Lumsden, 2004; Vieira et al., 2005; Scholpp et al., 2006; Szabó et al., 2009; Vue et al., 2009). This indicates that, in the forebrain, these morphogens occasionally may also act outside the standard DV context.
Functional studies indicate that SHH is a widespread diffusible signal that is implicated in both short- and long-range interactions related to patterning in the CNS, somite and limbs (Wagner et al., 1990; Krauss et al., 1993; Riddle et al., 1993; Chang et al., 1994; Echelard et al., 1994; Roelink et al., 1994). In the CNS, diffusing SHH signals emanating from the underlying notochord induce the development in the overlying neural ectoderm of a specialized population of ventral midline cells, the floor plate, which ulteriorly functions as well as a source of ventralizing effects upon the rest of the neural wall (Yamada et al., 1991; Placzek et al., 1993).
In the spinal cord, a ventrodorsal concentration gradient of SHH directly organizes the development of the ventral part of the neuroepithelium. Differential steplike cellular interpretation of these signals subdivides the lateral wall of the spinal cord in multiple DV domains with characteristic homeodomain transcription factor “codes” (Briscoe et al., 2000). Analogous DV subdivisions based on a code of transcription factors have been studied in the hindbrain, midbrain, pretectum and the subpallium (subpallial progenitor domains) (Agarwala et al., 2001; Ferran et al., 2007; Flames et al., 2007; Sieber et al., 2007; Kala et al., 2009). Most of these domains seem to be patterned via an integration of BMP, FGF8, and SHH signals.
The precise role of SHH in subpallial telencephalic patterning remains so far unclear. We know that its preoptic expression domain appears later than its expression in the hypothalamic floor and basal domains (Martí et al., 1995). Moreover, this delayed expression depends on the normal development of the prechordal plate up to stage HH6 (García-Calero et al., 2008). Existing descriptions of the location and time of appearance of preoptic (subpallial) SHH signaling are clearly insufficiently detailed. The aim of the present work is to resolve this problem, examining the spatiotemporal expression of Shh in the alar plate of the secondary prosencephalon in the chick.
It was previously suggested that the Dg (old AEP) is the main telencephalic domain expressing Shh in both the chick and the mouse (Shimamura et al., 1995; Shimamura and Rubenstein, 1997; Perez-Villegas et al., 1999; Puelles et al., 2004). According to present immunohistochemical and in situ hybridization data, we arrived at the conclusion that the ventricular zone expression of Shh and SHH actually does not occur in the Dg, but characterizes selectively a dorsal part of the POA and a related sector of the median commissural septum. This agrees with recent data in the mouse (Flames et al., 2007; García-López et al., 2008). Analysis of different developmental stages indicated that Shh-expressing cells subsequently observed in the mantle zone of the telencephalic vesicle probably migrate tangentially out of this POA/septum domain, mainly invading the evaginated subpallial domains, notably the pallidum and the striatum.
Materials and Methods
Animals
Care and use of laboratory animals conformed to normatives of the European Community (86/609/EEC) and the Spanish Government (Royal Decree 1201/2005).
Commercial fertilized chicken eggs were incubated at 37.5°C in a forced-draft incubator until the desired embryonic stages. The embryos were staged according to Hamburger and Hamilton (1951) and immersion- or perfusion-fixed (+postfixed) according to their stage with 4% paraformaldehyde in phosphate-buffered saline (PBS) at 4°C for 48 h. The brains were dissected free, washed in PBS, embedded in 4% low-melting point agarose and sectioned 100 μm thick with a vibratome. Subsequent processing was performed on floating sections in six-well plastic dishes for in situ hybridization and/or immunohistochemistry. Others brains were cryoprotected in 30% sucrose, embedded in gelatin, sectioned with a cryostat 18 μm thick and mounted on Super Frost slides.
Immunocytochemistry
Following several rinses in 0.1 M PBS with 0.75% Triton X-100 (PBT), sections were treated with 0.3% hydrogen peroxide in PBT for 15 min in the dark to inactivate endogenous peroxidase activity. After several washes in PBT, they were blocked with 0.2% gelatin (PBTG) and 0.1 M lysine for 1–4 h. The primary antibody – diluted in PBTG with 0.1% sodium azide – was applied at 4°C with moderate shaking for 48 h. We employed a rabbit polyclonal anti-calbindin serum [SWant, Bellinzona, Switzerland, #CB-38a (new batch), Lot No.: 9.03; 1:3000] prepared against recombinant rat calbindin D-28 K (all CB-positive structures described here showed no cross- reactivity with SWant polyclonal anti-calretinin antibody, #7696, Lot No.: 25392; 1:3000) and an affinity-purified rabbit polyclonal IgG prepared against a synthetic peptide representing amino acids 111–120 of the rat NKX2.1 protein (TTF-1, Biopat Immunotechnologies; Piedimonte Matese, Caserta, Italy, #PA0100; 1:2000). Western blots performed for the anti-calbindin antiserum by Suarez et al. (2006) checked that it recognizes the chicken protein.
After washes in PBT and PBTG, the tissues were incubated with goat anti-rabbit IgG (1:200 in PBT, 1.5 h; Vector Laboratories, Burlingame, CA, USA), passing thereafter to streptavidin/HRP complex (Vectastain-ABC kit; Vector Laboratories, 1:350, 1 h). The peroxidase brown reaction was developed for 15 min with 3,3′-diaminobenzidine (Sigma, St Louis, MO, USA; 50 mg/100 ml) plus 0.03% hydrogen peroxide in 0.05 M Tris–HCl buffer (pH 7.6) for 5–10 min. The reaction was stopped with 0.05 M Tris buffer. The sections were dehydrated in alcohol and xylol and coverslipped for single labeled sections.
In Situ mRNA Hybridization
Whole mount dissected embryonic brains and floating 100-μm-thick vibratome sections were hybridized in situ following the protocol described by Shimamura et al. (1994). Gelatin-embedded and 18-μm-thick cryostat-sectioned material were processed for in situ hybridization according to a protocol previously described by Schaeren-Wiemers and Gerfin-Moser (1993) and modified by Hidalgo-Sánchez et al. (2005). Digoxigenin-UTP-labeled sense and antisense riboprobes for chicken Shh (99% of similarity with Mus musculus homologous sequence NM_009170, positions 339–931; C. Tabin), Dlx5 (accession number: U25274, positions 496–1095; P. Gruss), Nkx2.1 (accession number: NM_204616.1, positions 89–1777; J.L.R. Rubenstein), and Nkx2.2 (accession number: AF110994.1, positions 1–759; J.L.R. Rubenstein), FoxG1 (accession number: NM_205193.1, positions 1153–1786; C. Stern) were prepared according to instructions of the digoxigenin manufacturer (Roche Diagnostics S.L, Applied Science; Barcelona). In each case the cDNAs were sequenced (SAI, University of Murcia) and their specificity was checked using the BLAST (NCBI) or BLAT (Ensembl) engines, corroborating that the nucleotide sequences employed are specific for the respective mRNAs. As general ISH controls, sense and antisense probes were applied to adjacent representative sections (in every case the signal was present only with the antisense probe), and some sections were processed without either sense or antisense probes to check for background due to the other reactives used in the ISH procedure.
Hybridized products were detected immunocytochemically using anti-digoxigenin antiserum coupled to alkaline phosphatase. The standard visualization procedure with NBT/BCIP solution as chromogenic alkaline phosphatase substrate gave a visible dark blue reaction product. The sections were dehydrated and coverslipped, and the whole mounts were transferred to 50% glycerol.
Imaging
Microphotographs were captured with an Axiocam digital camera, and brightness and contrast were adjusted with Adobe Photoshop 7.0.1. Representative images were used as imported templates within Canvas 9.0 software (Deneba) to draw structural schemata.
Results
Preliminary Remarks about Telencephalic Subpallial Nomenclature
In chick embryos, the telencephalic vesicle starts to evaginate at stages HH16/17 (Puelles et al., 1987a) and already expresses widely the FoxG1 transcription factor from HH14 onwards (Figures 1C,H,N). This primordium includes the prospective pallial and subpallial regions, which are later distinguished by specific gene expression patterns (Puelles et al., 2000, 2004, 2007; Puelles and Rubenstein, 2003). Dlx family genes represent generic markers of the subpallium. The best known part of the subpallium is represented by the pallidal and striatal domains, but generic subpallial markers such as the transcription factor Dlx5 also extend continuously their expression into the diagonal domain (Dg; old AEP), found at the stalk of the hemisphere, and the adjoining, non-evaginated POA. These domains also express the generic telencephalic marker FoxG1, and accordingly are considered now to represent non-evaginated telencephalic parts, consistently with the classical concept of “unpaired telencephalon” (Puelles et al., 2000, 2004, 2007; Puelles and Rubenstein, 2003). In the prosomeric model the entire telencephalon is understood as forming part of the forebrain alar plate (Puelles and Rubenstein, 2003).
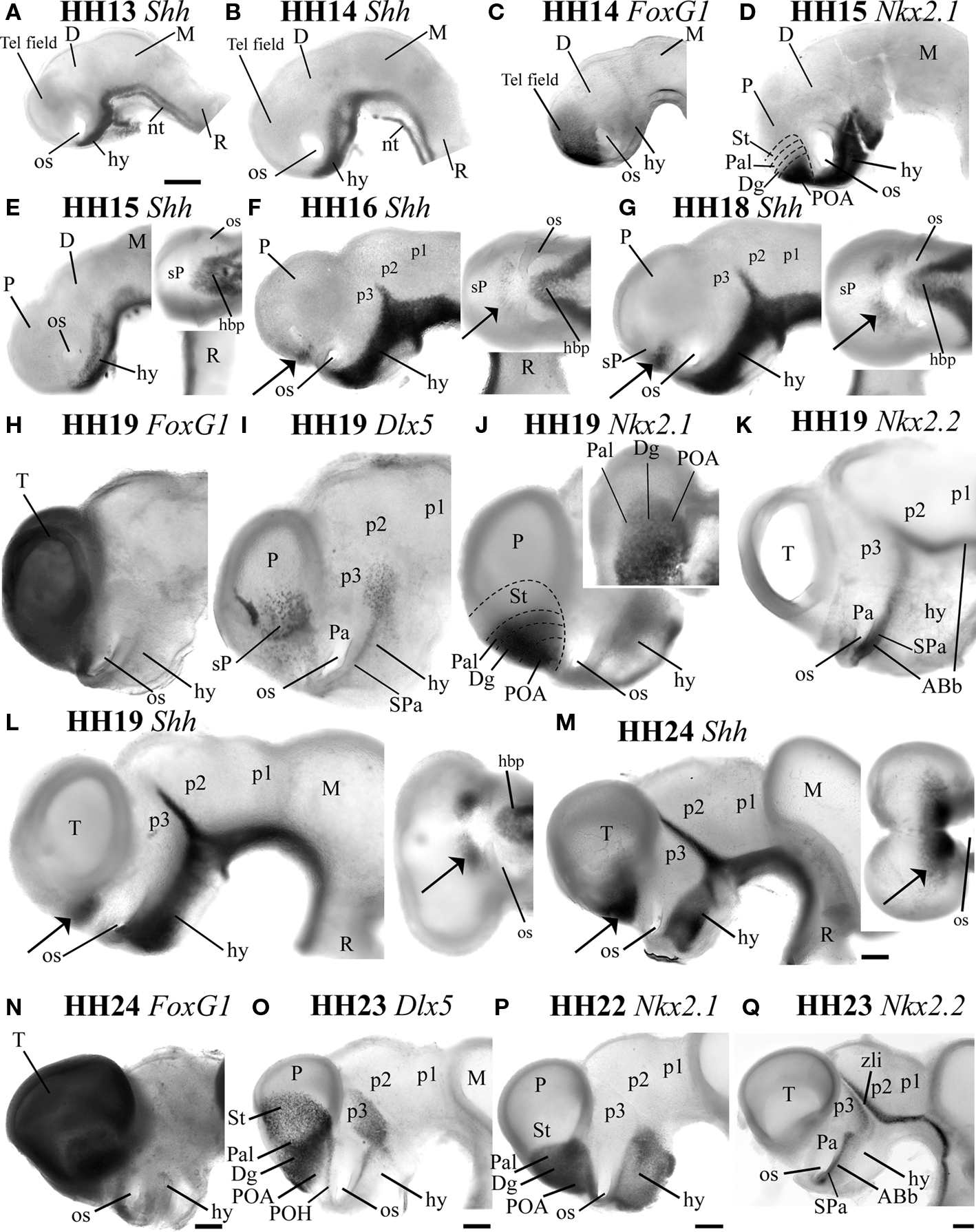
Figure 1. The Shh-positive domain appears in the subpallium at HH16, being strictly separate from the Shh-positive basal plate domain. Whole mount in situ hybridization for Shh in the early developing embryo in lateral views at HH13 (A), HH14 (B), HH15 (E), HH16 (F), HH18 (G), HH19 (L), HH24 (M) are compared with whole mount in situ hybridization for the telencephalic marker FOXG1 at HH14 (C), HH19 (H), and HH24 (N), the subpallial marker Dlx5 at HH19 (I) and HH23 (O) and the pallidal marker Nkx2.1 at HH15 (D), HH19 (J), and HH22 (P). The alar-basal boundary was visualized with in situ hybridization for Nkx2.2 at HH19 (K) and HH23 (Q). The insets at the top right corner of (E), (F), (G), (J), (L), and (M) are ventral views at the same magnification as the respective lateral views. The arrows in (F), (G), (L), (M) indicate in lateral or ventral views the Shh-positive domain detected in the subpallial preoptic alar region. Scale bar in (A) = 0.2 mm applies to (A)–(L), whereas the bar = 0.3mm in (M)–(Q).
In agreement with Puelles et al. (2007), Flames et al. (2007) and García-López et al. (2008), we identify here the newly recognized septocommissural preoptic area (SCPo; Figures 3A,C,D,I–K). The rationale for this new concept relates to the observation that all molecularly characterized subpallial domains extend into the septum (Puelles et al., 2000), and the POA, now understood as a fourth subpallial domain, also visibly does so, reaching with the dorsal tip of the median preoptic nucleus the median septal locus occupied by the crossing anterior commissure (Figure 7). The septal region in general originates from the fusion of the bilateral rim of the neural plate during neurulation, and the SCPo, with the bed of the anterior commissure, corresponds to the rostral end of the forebrain roof plate (Puelles et al., 1987b). It thus represents the rostromedian roof plate overlying the rostralmost forebrain alar plate, which includes the POA (Puelles et al., 1987b, 2007; Puelles, 1995; Shimamura et al., 1995; Cobos et al., 2001; García-López et al., 2008).
The Onset of Alar Shh Expression in the Embryonic Chick Forebrain
The expression of Shh in ventral parts of the CNS (e.g., floor and basal plate of the forebrain) begins already at stage HH5, as soon as gastrulation begins (Sánchez-Arrones et al., 2009). This early floor plate expression domain expands into the adjoining basal plate zone across forebrain and midbrain shortly afterwards, (Martí et al., 1995). In contrast, alar expression of Shh in the forebrain occurs independently from the floor and basal domains of expression. Figures 1A,B,E–G,L,M display Shh mRNA expression in forebrain whole mounts between HH13 and HH24, focusing on the secondary prosencephalon. At HH13 and HH14, Shh expression extends along the entire floor plate of the brain as well as the underlying notochord (nt; Figures 1A,B). In the secondary prosencephalon, Shh expression already has expanded from the floor plate into the right and left basal plates, whose rostral ends are continuous across the midplane, ventral to the optic stalks and the prospective chiasmatic area (hy, os; Figures 1A,B). At HH13–14, Shh-positive cells are absent within the FoxG1-positive telencephalic field, which is still wholly unevaginated (compare Tel field in Figures 1B,C). The transcription factor Nkx2.1 is first expressed in the subpallium between HH12 and HH13 (not shown; Pera and Kessel, 1998; Crossley et al., 2001). At HH15, the alar expression domain of Nkx2.1 transiently covers mainly the presumptive territory of the POA, but is as yet excluded from the prospective diagonal, pallidal, and striatal subpallial areas (compare Figure 1D with Shh-positive POA at HH16 in Figure 1F; note the diagonal and pallidal areas do express Nkx2.1 at HH19 – see Figure 1J). Shh mRNA is clearly undetectable in the entire subpallium at HH15 (Figure 1E, see a ventral view in the inset). At HH16, Shh begins to be downregulated in a median floor area of the hypothalamus that extends from the retromammillary area to the median eminence (Ohyama et al., 2005, 2008; Manning et al., 2006; igure 1F). This negative area expands thereafter, practically coming to restrict ventral Shh expression to the hypothalamic basal plate (Figures 1F,G,L,M and their insets).
At HH16, Shh-positive cells first appear in the alar telencephalic field (Figure 1F, arrow; ventral view in the inset). This occurs in bilateral patches observed near the incipient telencephalic stalk; these patches are clearly separated from the hypothalamic Shh-positive basal plate domain by the negative optic stalk and optic chiasma areas, both at this stage and at all subsequent ones (see arrow, os, hbp in Figures 1F,G,L,M). Given its relative position inside the larger Nkx2.1-expressing part of the subpallium (compare Figure 1D), this Shh-positive alar domain apparently corresponds to a dorsal part of the prospective POA. Ventral views shown in Figures 1F,G,L,M (insets) illustrate the lack of continuity between the basal and alar Shh domains in the forebrain. In fact, we know from other studies that the alar Shh-positive patches are separated from the local Shh-positive basal plate by at least two different longitudinal alar hypothalamic domains: (1) the Dlx- and Shh-negative supraopto-paraventricular area, which lies under the border of the FoxG1-positive telencephalic field (Pa in Figures 1I,K) and (2) the subparaventricular area, which lies under it, adjacent to the Shh-positive basal plate, and is positive for Dlx5 and Nkx2.2 (SPa; Figures 1I,K). The Pa area is positive for the transcription factor Otp (not shown; Bardet, 2007; Bardet et al., 2008).
At HH19, FoxG1 is still expressed in the whole telencephalon (T, Figure 1H), whereas Dlx5 expression characterizes the whole subpallium (sP, Figure 1I), and encloses the Pal/Dg/POA area sharing now Nkx2.1 signal (Pal, Dg, POA; Figure 1J). All these subpallial domains, are located in the alar plate, consistently with the more ventral linear expression of the marker Nkx2.2, held to overlap the alar-basal boundary (ABb; both the alar subparaventricular band and a small part of underlying basal plate). This extends into a rostromedian crossing under the optic stalks and the chiasma (Puelles and Rubenstein, 1993; see ABb, SPa; Figure 1K; compare Figures 1H–L). The Shh-positive preoptic domain has expanded somewhat toward the midline (Figure 1L, inset), but does not occupy the whole POA; there is a small ventral part of the preoptic domain, just dorsal to the optic stalk, that is negative for Shh, whereas it is positive for FoxG1, Dlx5, and Nkx2.1 (Figures 1H–L).
From stage HH19 onwards, the intensity and extent of the Shh signal in the dorsal POA increases (Figures 1L,M versus Figures 1F,G). At HH24, the Shh-positive dorsal preoptic domain has expanded toward the preoptic midline, reaching close to the lamina terminalis, without entering it properly yet (compare insets of Figures 1L,M; also LT in Figure 2C and inset). The Shh signal observed in whole mounts at this stage also appears scattered peripherally into the neighboring part of the telencephalic vesicles, corresponding – we think – to tangentially migrating positive cells in the mantle (see below; arrow in inset, Figure 1M). At HH24, the alar Shh domain is still enclosed by the nested telencephalic domains expressing FoxG1- (compare Figures 1M,N), Dlx5- (compare Figures 1M,O), and Nkx2.1-positive cells (compare Figures 1M,P). The postulated ABb, roughly visualized in Figure 1Q by means of the linear Nkx2.2 signal, corroborates also at this stage the lack of contiguity between the alar subpallial territory expressing Shh and the Shh-positive hypothalamic basal plate (Figure 1M).
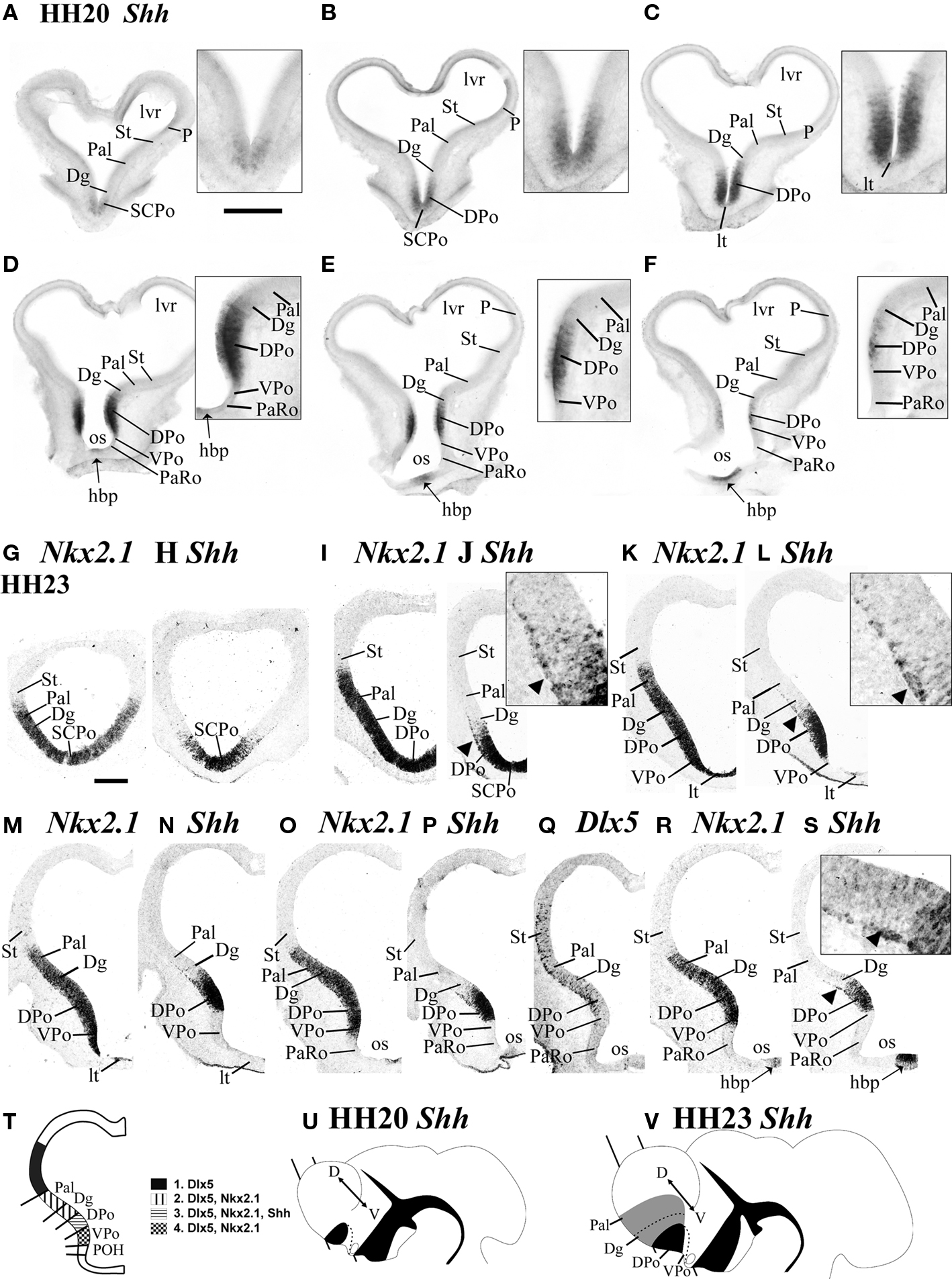
Figure 2. Shh is expressed in the ventricular zone of a dorsal preoptic subdomain at HH20 and HH23. Emergence of the first tangentially migrating Shh-positive cellsoccurs at HH23. (A–F) Transverse 100 μm vibratome sections cut through the chicken forebrain at HH20 and processed by in situ hybridization for Shh are arranged from rostral to caudal [see section plane in (U)]. The insets at the right top corners of (A)–(F) show enlarged detail views of the Shh-positive DPo ventricular zone. (G–S) Transversel 16-μm-thick adjacent cryostat sections at HH23 [see section plane in (V)] labeled by in situ hybridization for Shh (H, J, L, N, P, S), Nkx2.1 (G, I, K, M, O, R) and Dlx5 (Q). The insets at the right top corners of (J), (L), (S) show enlarged views of the Shh-positive subpial cells migrating out of DPo (arrowheads mark the limits between neighboring subpallial domains. In (T), the main ventricular domains revealed by Dlx5, Nkx2.1 and Shh expression are summarized in a drawing of a transverse section [similar to (R)]. (U) and (V) are schematic lateral views of chick embryos at HH20 and HH23, respectively, representing the areas of ventricular (black) and mantle (gray) expression of Shh; the bar symbolizes the section plane used in (A)–(F) (HH20) and (G)–(S) (HH23). Scale bar in (A) = 0.4 mm [applies to (A)–(F)] and = 0.2 mm for the respective magnified views. Scale bar in (G) = 0.2 mm [applies to (G)–(S)] and = 0.05 mm for the corresponding magnified views.
Ventricular Zone Expression of Shh Divides the Preoptic Region into Dorsal and Ventral Parts
At stages HH22–24, the ventral part of the POA, adjacent to the optic stalk, does not display Shh signal (which appears only in the dorsal POA), but expresses weakly FoxG1 and Dlx5, and strongly Nkx2.1, (VPo, DPo; Figures 1M–P). Apart of these results, observations on brain sections also clearly disclosed two parts in the POA, specifically as regards ventricular Shh expression. Figure 2 shows coronal sections through the POA and the evaginated telencephalic vesicle at HH20 and HH23 (see explanatory schema and section planes in Figures 2T–V).
At HH20, the majority of the Shh-positive preoptic tissue consists only of ventricular cells (Figures 2A–F,U). The dorsalmost expression of Shh is found at the SCPo, the prospective bed of the anterior commissure (SCPo; Figures 2A,B). In sections through the conventional POA, which displays the lamina terminalis at the midline, the dorsal preoptic ventricular zone continues strongly Shh positive, whereas the lamina terminalis (DPo; Figures 2B–F; LT in Figure 2C and inset) and the small ventral preoptic subdomain lack Shh altogether (VPo, Figures 2D–F). The Shh-positive DPo and the Shh-positive hypothalamic basal plate (hbp), are separated by the unlabeled VPo, as well as by the likewise negative paraventricular and subparaventricular areas of the hypothalamic alar plate, also related at the midline to the prospective optic chiasma (the later starts to receive retinal fibers at HH24).
Shh-Positive Postmitotic Cells Spread Out in the Subpallial Mantle as Far as the Striatopallidal Border
We compared Shh expression at HH23 with the pallidal Nkx2.1 and striatal Dlx5 markers (Figures 2G–S,V). The SCPo part of the prospective septum, site of the future anterior commissure, and the underlying alar DPo still have a Shh-positive ventricular zone, including the commissural midline (Figures 2H,J). Note that this commissural anlage is the rostral end of the forebrain roof plate (Cobos et al., 2001), and as such represents a part of the septal commissural plate The alar lamina terminalis starts underneath the SCPo, is thinner than it, and in contrast is Shh-negative (LT; Figures 2K–N). Shh is not expressed in the lateral wall of the ventral preoptic area (VPo, Figures 2K–P,R,S), though Nkx2.1 signal is present there. Preoptic ventricular Shh signal is accordingly restricted to the dorsal POA and the SCPo (DPo, SCPo; Figures 2L,N,P,S).
In addition, an incipient subpial stream of Shh-positive cells is observed in the DPo mantle, extending tangentially away into the neighboring diagonal and pallidal domains of the subpallium (arrowheads and inserts, Figures 2J,L,S). At this stage they already practically reach the striatopallidal limit (note the intratelencephalic border of Nkx2.1 expression – indicating the striatopallidal border – roughly corresponds with the farthest Shh-positive superficial cells).
There is no correlative Shh expression in the ventricular zone of these invaded subpallial domains (Figure 2T). The tangential arrangement of these subpial Shh-positive cells suggests that they constitute a migratory stream of cells originating in the DPo. Such cells are less apparent at the SCPo (compare inset of Figure 1M). We did not observe Shh-positive cells inside the striatal Dlx5-positive and Nkx2.1-negative territory at HH23 (St, Figures 2Q–S).
Follow-Up of Ventricular Shh Expression in the Dorsal POA and SCPo, and Migratory Dispersion of Shh Cells in the Subpallium at HH28–34
At stages HH28–34, the expression of Shh is still strong in the ventricular zone at the DPo and SCPo areas (Figures 3A,C,D,F,I–K, 4A–C, 5B,C,F,L,M and 6B,C). Horizontal sections through the subpallium (section plane in Figure 3A) that were reacted with ISH for Shh, or Dlx5 (for reference), show an increased number of Shh-positive cells in the subpallial mantle (Figures 3B–N). The pattern observed changes between the different subpallial sectors and is clearer in coronal sections at HH29 (Figures 3G–N).
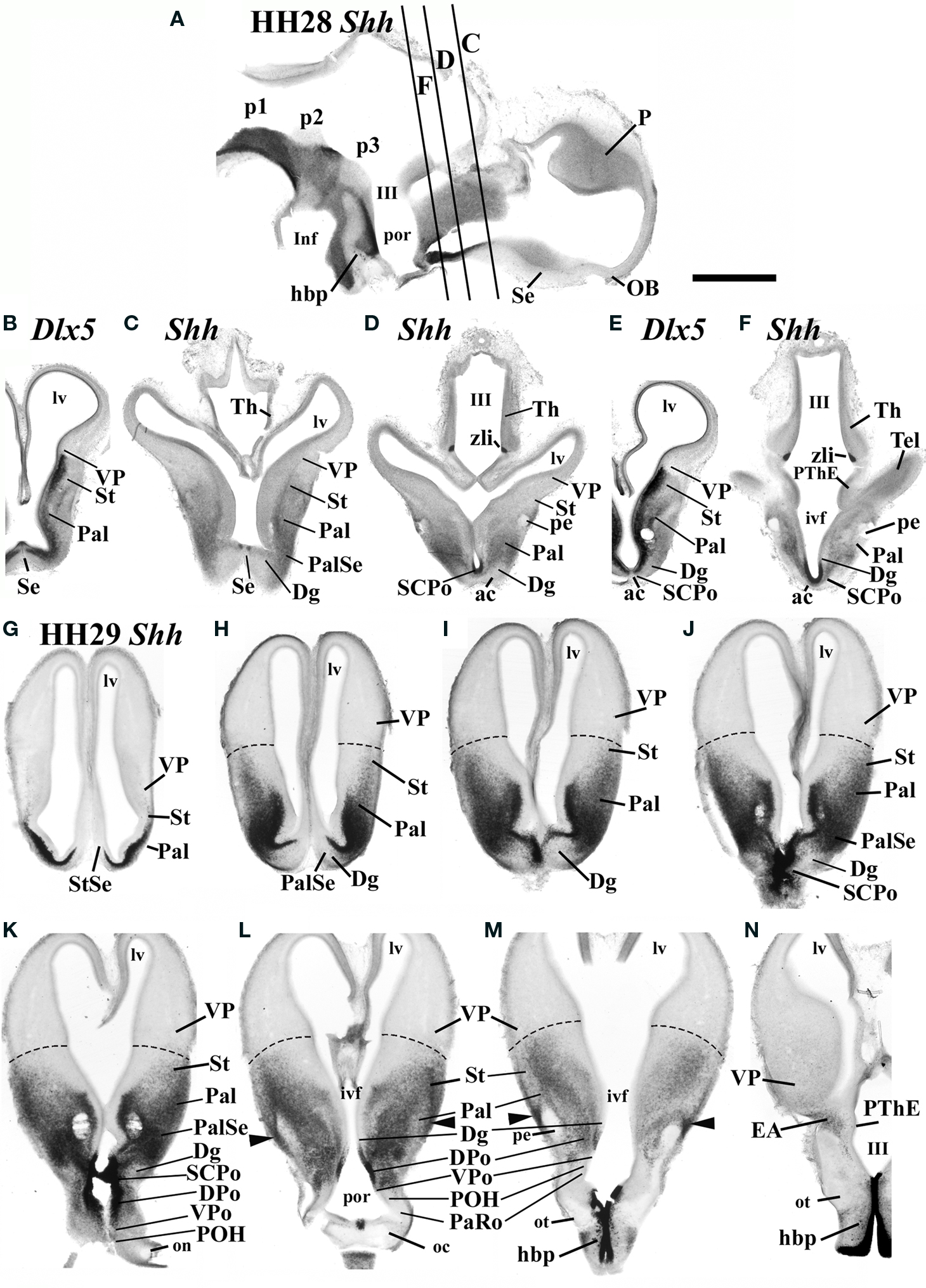
Figure 3. (A–F) At HH28, Shh-positive cells are spread out in the subpallium, principally in the pallidum (Pal), the pallidoseptal area (PalSe), and the SCPo. In situ hybridization for Shh on 100-μm-thick vibratome sections, performed on sagittal (A) and transverse sections (C, D, F) arranged from rostral to caudal, and compared with similar sections (though in a slightly different section plane) through an embryo processed by in situ hybridization for the subpallial marker Dlx5 (B, E). (G–N) In situ hybridization for Shh was performed at HH29 on 100-μm-thick coronal vibratome sections through the forebrain, and shown from rostral to caudal. Scale bar in (A) = 1 mm applies to all images.
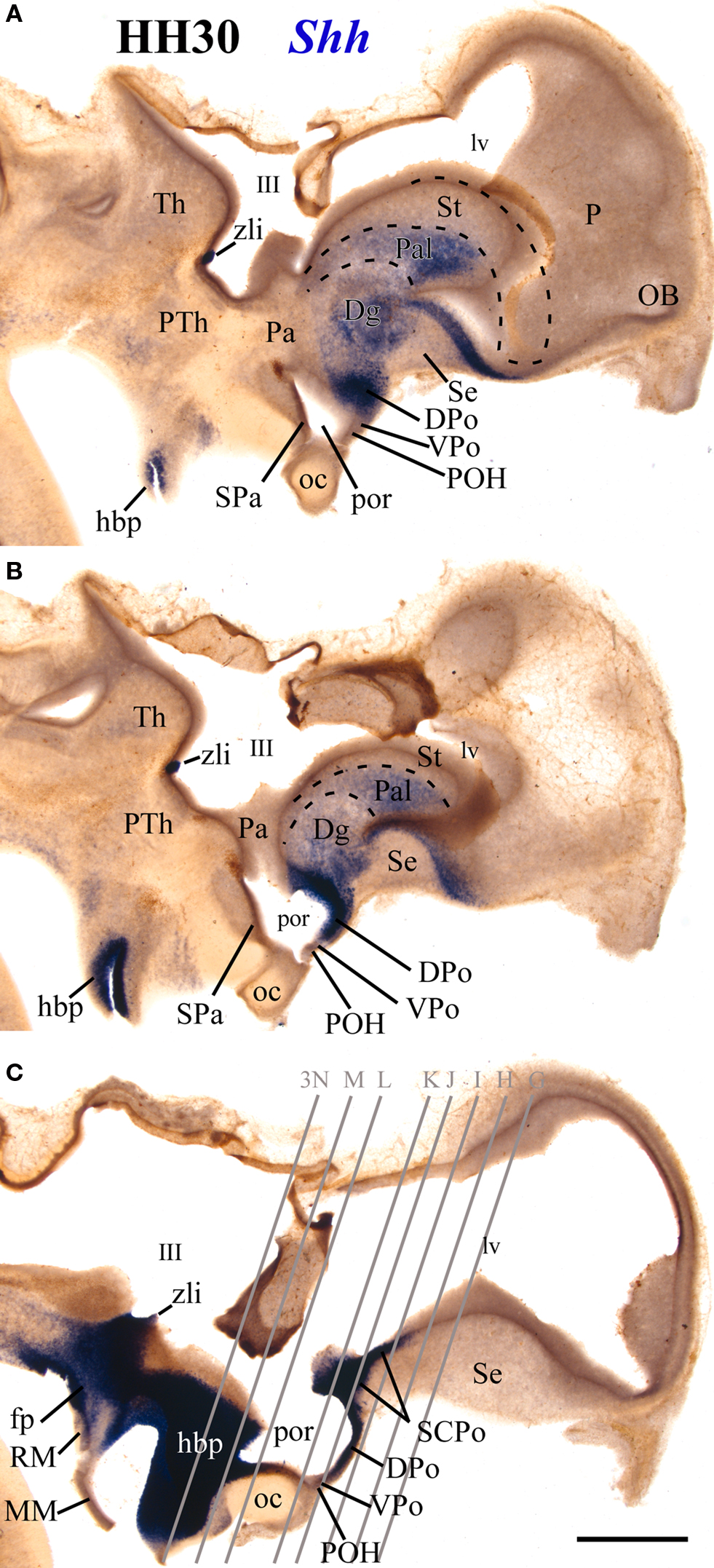
Figure 4. 100-μm-thick vibratome sagittal sections of an HH30 embryo processed for in situ hybridization for Shh, arranged from lateral (A) to medial (C). Scale bar in (C) = 1 mm.
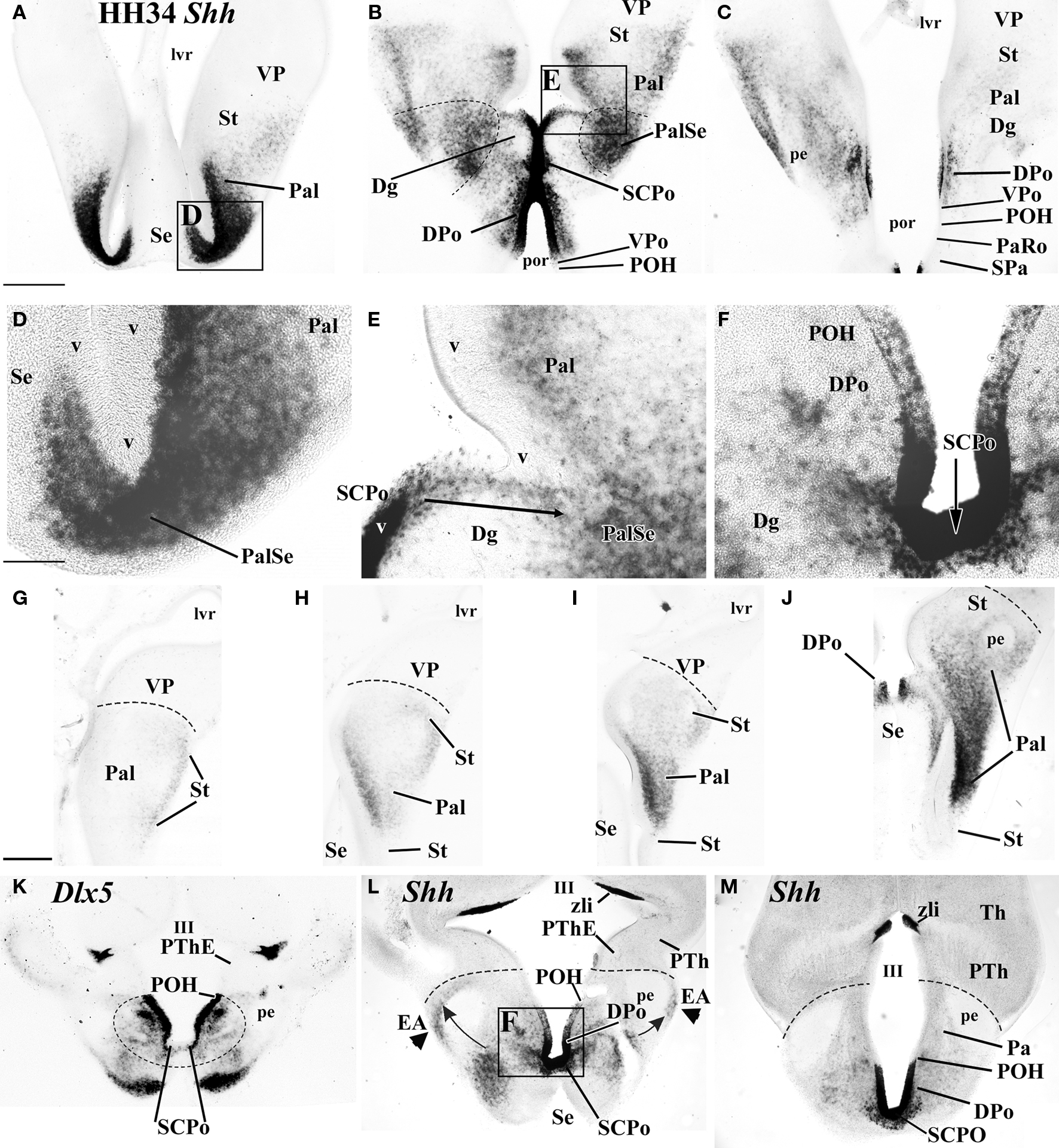
Figure 5. In situ hybridized 100-μm-thick vibratome transverse (A–E) and horizontal sections (F–M) cut through chicken forebrain at HH34 and processed for either Shh (A–E, F–J, L, M) or Dlx5 (K). These two sectioning planes display the topography of the Shh-positive territories, especially the preoptic area. Scale bar in (A) applies to (A)–(C) = 0.5 mm, scale bar in (D) applies to (D)–(F) = 0.125 mm, scale bar in (G) applies to (G)–(L) = 0.5 mm.
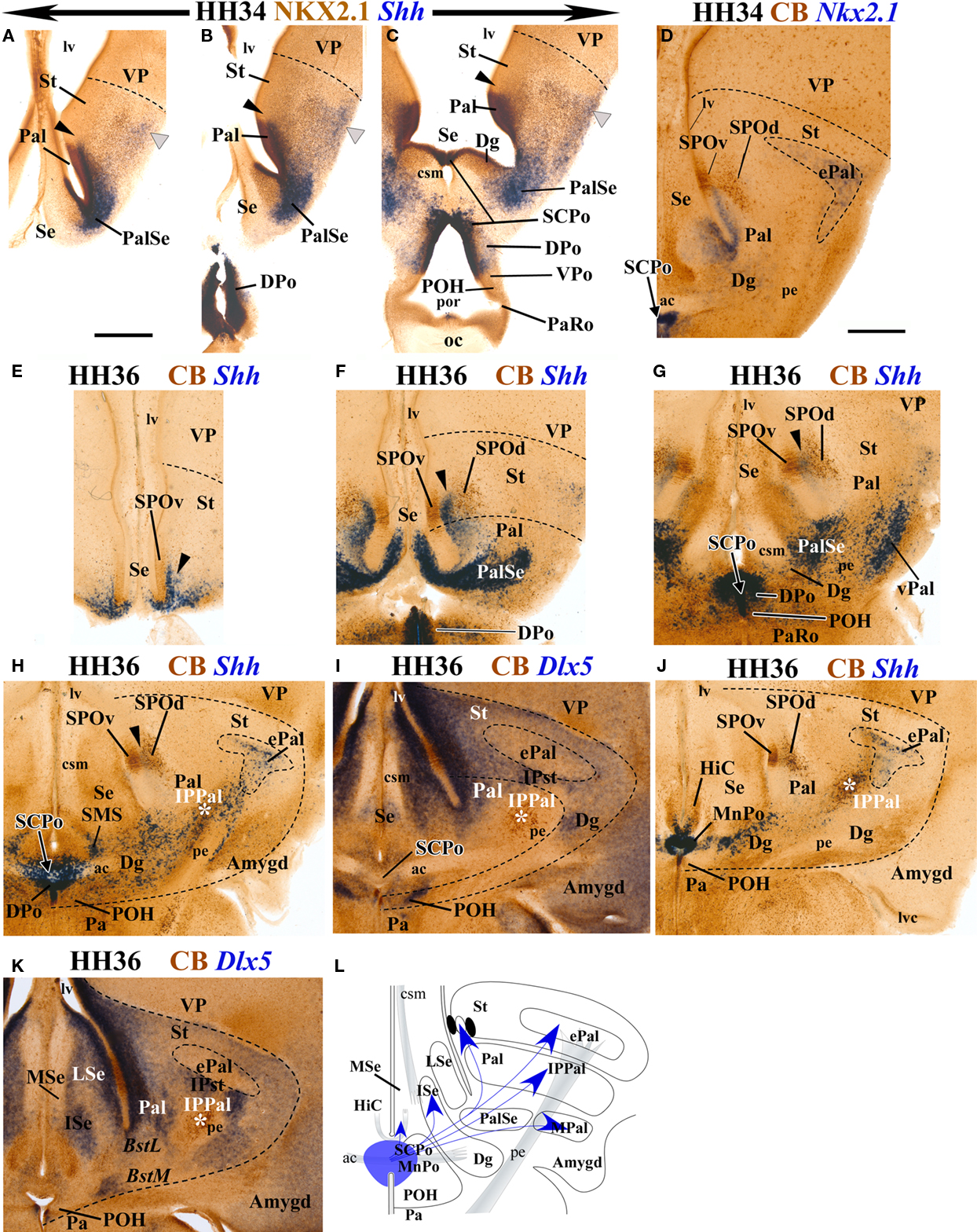
Figure 6. Transverse sections through chicken forebrain at HH34 (A–D) and HH36 (E–K), double-labeled with NKX2.1 immunohistochemistry and in situ hybridization for Shh (A–C), or double-labeled for CB immunohistochemistry and in situ hybridization for Nkx2.1 (D), Shh (E–H, J, L) or Dlx5 (I, K). (E–K) the order goes from rostral to caudal, illustrating the topography of Shh-positive populations in comparison with the general subpallial marker Dlx5 (I, K) and with the telencephalic striatopallidal CB-positive circumventricular organ, which is continuous with the preoptohypothalamic transition zone [SPO and POH (E–K)]. (L) Summary schema of the apparent ventricular preoptic source of Shh-positive cells which spread out from the SCPo and DPo to Pal, PalSe. ePal, IPPal, and MAm. Scale bar in (A) applies to (A)–(C) = 0.5 mm, scale bar in (D) applies to (D)–(K) = 0.2 mm.
At the median unevaginated telencephalon, the neuroepithelium of the SCPo is strongly Shh positive (SCPo; Figures 3D,J,K, 4C and 5B,F,L,M). Ventral to it, Shh is also expressed at these stages in the dorsal preoptic ventricular zone, found under the anterior commissure and the SCPo (DPo, Figures 3D,F,J–L, 4A,B and 5B,C,L,M). The mantle domain of the DPo contains dispersed Shh-positive cells, particularly periventricularly (DPo; Figures 3K,L, 4A, 5B,C,L,M and 6C). The VPo remains largely Shh-negative, though some positive DPo cells may have dispersed tangentially into it (VPo; Figures 3K,L, 5B,C and 6C).
Within the diagonal domain (Dg), the labeled cells appear mainly within the deep periventricular stratum, or in a thin subpial stream of cells, with sparse labeled cells at intermediate radial levels (Dg; Figures 3H–M, 4A,B, 5B,E and 6C). In contrast, the pallidum (Pal), shows a thick, densely populated periventricular stratum, continuous with that of the Dg, but there are also numerous positive cells in medium-dense intermediate and superficial populations, thus allowing the visualization of a distinct molecular boundary with the Dg, particularly at the pallidoseptal transition area (Pal, PalSe; Figures 3G–K, 4A,B, 5A,B,D,E and 6A–C). The striatal area contained a less important and more dispersed Shh-positive population, which was largely restricted to superficial and intermediate levels of the mantle, and did not reach the pallio-subpallial boundary (St; Figures 3H–M, 5G–J and 6A–D). We believe that this population invading the striatum is related to the well-known partial intrusion of pallidal structures into the striatal domain of birds (not so in mammals), building there an ectopic pallidum area (ePal in Figure 6D; Reiner et al., 1984; Puelles et al., 2007).
Ventrally to the POA there appears at some distance the rostral tip of the hypothalamic basal plate, whose ventricular zone expresses strongly Shh (hbp; Figures 3L–N); this domain is still well separated from the Shh-positive alar domain by Shh-negative parts of the preoptic and hypothalamic alar plate. The Shh-positive basal plate ends in a median spike closely underneath the optic chiasm (hbp; Figures 3A,L–N and 4C).
We also noted at HH29 and HH34 a small stream of Shh-positive cells that runs bilaterally at the back of the brain and ends close to the prethalamic eminence, passing always superficial to the peduncular tract; these cells apparently come from the neighboring subpallial telencephalic mantle in more rostral coronal sections (arrowheads in Figures 3K–M) and may represent a migration of dorsal preoptic derivatives along or into the extended amygdala (EA; Figures 3N and 5L).
Establishment of the Definitive Regional Distribution of Shh Populations in the Subpallial Mantle
Data obtained at HH36 (10 days of incubation) revealed a conspicuous, largely superficial striatal population of Shh-expressing cells which overlaps the ectopic pallidal regions, whereas strictly striatal regions (labeled strongly by Dlx5 expression – see Figures 6I,K – and not positive for Nkx2.1; see Figure 6D) are devoid of such cells (Figures 6G–K). The pallidum, which previously (e.g., HH34) could be sharply delimited from the striatum by the association of its Shh-positive periventricular cells to Nkx2.1-expressing ventricular and periventricular strata (Pal; Figures 6A–C) shows now some changes in this Shh cell population with regard to the calbindin-immunoreactive striatopallidal circumventricular organ recently described by
Bardet et al. (2006), present on the other side of the striatopallidal boundary (SPO; Figure 6D). The earlier dense periventricular stratum of the pallidum is reduced gradientally within the pallidum itself (Pal; Figure 6F), and shows a denser tip that insinuates itself inside the striatum, specifically inside the SPO, occupying precisely the space that separates its ventricular and distal CB-positive cell populations (arrowhead; SPOv, SPOd; Figures 6E–H,J). This new Shh-positive SPO subpopulation lies thus strictly within the striatum and is most voluminous in sections passing through the base of the septum (Figure 6F) and becomes tenuous more rostrally (Figure 6E) or caudally (Figures 6G,H,J). This finding suggests that the periventricular cell stream previously observed advancing through the lateral Dg and Pal domains may be largely destined to form this intermediate stratum of the striatopallidal organ (Bardet et al., 2006).
In contrast, at HH36, the Shh-positive pallidoseptal transitional region of the pallidum, which borders the Dg, condensates without further displacement into a very compact periventricular cell population, the apparent anlage of the PalSe area distinguished by Puelles et al. (2007). This population expands superficially and caudalwards around the peduncle, producing a related, slightly less compact local superficial formation whose position might be compared conceptually with that of the mammalian ventral pallidum (PalSe; pe; VPal; Figures 6F,G). Moreover, the dense PalSe area is continuous at the medial pallidoseptal transition zone with Shh-positive cells which already formed at HH34 a thin but dense periventricular stratum entering the septum (Figures 6A,B); this stratum is now much better developed at HH36 (Se; Figure 6F). It is less prominent in rostral sections (Se, Figures 5A and 6E). It apparently characterizes the postulated transition of the pallidum into the pallidal septum (Puelles et al., 2000), just caudal to the striatal accumbens formation (the latter only contains Shh cells within the SPO; present results; Bardet et al., 2006; Puelles et al., 2007). This Shh-positive pallidoseptal transitional region limits caudomedially with the diagonal domain, which still represents an histogenetic area with sparse Shh-positive cells, which lies intercalated between the PalSe and DPo regions(Dg; Figures 6G,H).
At the telencephalic midline, the SCPo ventricular zone is still strongly positive for Shh, as is that of the contiguous dorsal preoptic area (SCPo, DPo, Figures 6G–J). The sparsely populated positive mantle of the SCPo appears now traversed by the anterior commissure tract (ac; Figures 6H,I). Positive cells apparently streamed out of the SCPo mantle stratum seem to populate the submedial septal nucleus (SMS; Figure 6H; see Puelles et al., 2007) and also form Shh-positive cellular streams that penetrate the nucleus of the hippocampal commissure (HiC; Figure 6J; note the latter was formerly known as “pallial commissure”; see Puelles et al., 2007). The Shh-positive neuroepithelium of the DPo ends ventrally overlapping partially the CB-positive circumventricular preoptic organ (Bardet et al., 2006), next to the Dlx5-positive and Nkx2.1- and Shh-negative preoptohypothalamic zone (POH, Figures 1O,P and 6G,H,J; see Puelles et al., 2007).
At the transition between the SCPo and the Dg, Shh-positive cells were found to penetrate tangentially the deep stratum of the Dg and adjacent PalSe (SCPo; Dg, Figures 5F and 6H,J). Laterally dispersed Shh-positive cells colonize in general the pallidal domain and partly also a calbindin-immunoreactive and Dlx5-negative formation, called here tentatively “pallidal intrapeduncular nucleus” (IPPal), found interstitial to the peduncle before the latter enters the better known striatal intrapeduncular nucleus (IPSt) (asterisk; IPPal; IPSt; Figures 6H–K). This pallidal IPPal formation was first observed, but not named, in calbindin-immunoreacted material by Bardet et al. (2006) and Bardet (2007).
Some Shh-positive cells reach laterally the subpallial extended amygdala, as mentioned above, and partly aggregate just lateral to the lateral forebrain bundle, possibly contributing as well to the medial amygdala (Figures 6H–L).
We summarize in Figure 6L the main chicken subpallial areas displaying a Shh-positive cell population at stage HH36, and we tentatively show with arrows the apparent routes by which Shh cells seem to “escape” the DPo or SCPo sources as they distribute into different subpallial regions.
Discussion
Evolution Over Time of the Definitions of the Preoptic Area and Diagonal Domain
It seems convenient to start this section by discussing the recently changed concepts of the relevant histogenetic domains found in the neighborhood of the preoptic region, particularly the adjacent telencephalic stalk area, which abuts the septum medially and the pallidum laterally. The pioneering genoarchitectonic analysis of the preopto-subpallial region performed by Bulfone et al. (1993) and Puelles and Rubenstein (1993), who used Dlx2, Nkx2.1, and Gbx2 gene expression patterns to distinguish local subdivisions and relationships in mouse embryos, led to the tentative proposal of an anterior POA intercalated dorsoventrally between the diagonal domain (identified in those reports as AEP area) and the posterior POA (Figure 7 in Bulfone et al., 1993; Figure 4 in Puelles and Rubenstein, 1993). The AEP, which we now prefer to identify as diagonal domain or Dg, (see Allen Developmental Mouse Brain Atlas2) was already conceived as lying intercalated between the medial ganglionic eminence (prospective pallidum) and the POA. Puelles et al. (2000) proposed that the Dg (old AEP) reaches medially the septal region, side-by-side with the pallidal and striatal portions of the subpallium. It seemed at this time that the POA did not include itself a septal extension, since the adjacent Dg was held to end at the median commissural septum (SCPo), encompassing the anterior commissure. The Dg domain was subsequently also thought to be characterized by Shh expression, as judged mainly by the presence of Shh-positive cells in its mantle zone (Figure 3 in Puelles and Rubenstein, 2003). Recently, however, Flames et al. (2007) performed a detailed analysis of the ventricular expression patterns of multiple genes in the mouse subpallium at E13.5, revealing the existence of a multiplicity of differentially specified progenitor domains. They defined the mouse dorsal POA by its strong and selective ventricular Shh expression, and proposed that this characteristic area actually has a diminutive median septal component – called the SCPo (Figure 7; Bardet et al., 2006; Bardet, 2007; García-López et al., 2008 later identified it as preopto-commissural area or POC).
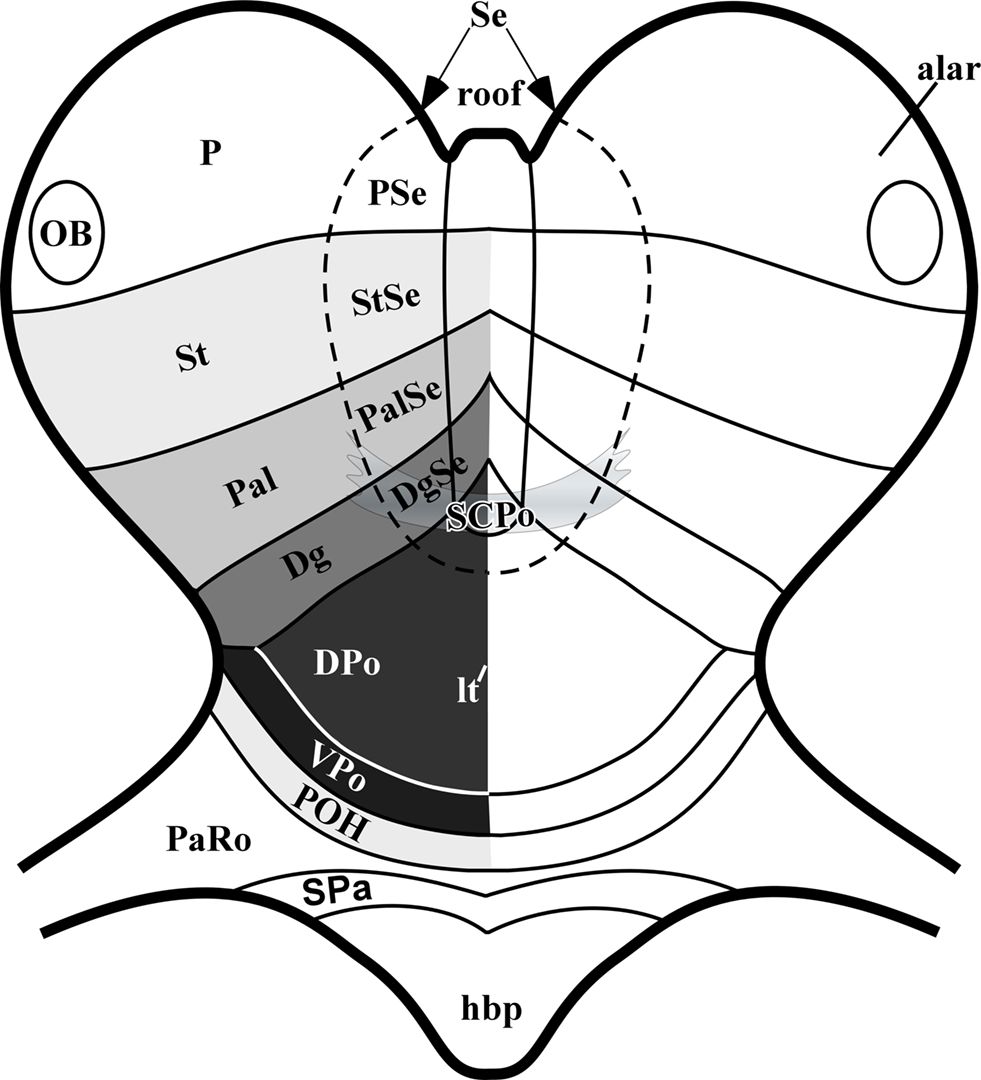
Figure 7. Schematic frontal view of the early developing forebrain, centered on the alar plate and the median roof plate. The septum participates both in the roof and alar plates, and is proposed to be divided into subpallial domains corresponding and continuing those of the lateral wall of the telencephalic vesicles (StSe/striatum, PalSe/pallidum, DgSe/diagonal domain, SCPo/preoptic area), with the addition of a small pallial septal domain – PSe – caudally (site of the hippocampal commissure, and eventually also of the corpus callosum in mammals). The preoptic area is divided into the SCPo at the septal roof plate (bed of the anterior commissure) and the alar Shh-positive DPo and Shh-negative VPo.
The existence of such a median SCPo domain clearly excludes the Dg from reaching the commissural midline, pushing its septal end to a paramedian septal position adjacent to the SCPo; the Dg domain was classified by Flames et al. (2007) as a pallidal partition (pMGE5) associated to production of cholinergic neurons. The POA was divided into molecularly distinct dorsal and ventral parts (their pPOA1 and pPOA2 areas), plus a POH area comparable to ours, limiting with the hypothalamus. García-López et al. (2008) also studied at several stages of mouse development the expression of various genes in this area, including Shh. They concluded that the POA, defined by them by ventricular Shh expression, apart the other subpallial markers, includes a median septal subdomain encompassing the anterior commissure (their POC). They referred to the Dg domain at the hemispheric stalk region as AEP, concluding that it accompanies the SCPo into the septum. Other recent work performed in chick embryos similarly identified adjacent Shh-negative Dg (AEP) and Shh-positive POA territories (Bardet et al., 2006; Bardet, 2007; Abellan and Medina, 2009). These last authors distinguished preopto-commissural and preopto-basal parts within the POA, which roughly correspond to our dorsal-plus-SCPo and ventral parts of POA, respectively. We think that the “commissural” adjective only applies to the thin SCPo part of the POA that constitutes the local roof plate (this is the rostral end of that longitudinal zone, according to fate mapping by Cobos et al., 2001), whereas the rest of the Shh-expressing POA belongs to the alar plate. We think that SCPo and DPo are best kept separate. We also find it is confusing to call “-basal” a portion of the alar plate, and thus prefer our VPo term for this area (Figure 7).
A gradient of SHH can be assumed to form around the SCPo at stages which significantly precede arrival of anterior commissure fibers at their crossing locus. It is conceivable that these commissural fibers grow up the concentration gradient into the SCPo. Interestingly, in mammals the fibers of the fornix tract also first converge upon the SCPo, but then pass ventralwards behind the anterior commissure, as they get detoured into their hypothalamic course. This suggests both attractive and repelling (direct or indirect) SHH effects on axonal navigation at the preoptic roof plate area (SCPo).
Our present results about ontogenetic early expression of Shh in the chicken subpallium in essence corroborate and complement these various new results obtained in mouse and chicken (the latter had been limited to data at middle and advanced stages of development), confirming that Shh characterizes in fact the dorsal POA and SCPo progenitor zones, and not the neighboring Dg, as thought before (old AEP concept), or the VPo and POH preoptic subareas. The discovery of the SCPo was crucial in this advance.
We were able to assess the specific stage at which Shh expression begins at the chicken DPo, namely stage HH16. Interestingly, Martí et al. (1995) illustrated a stage HH15 chick embryo wholly devoid of preoptic Shh signal, as well as a stage HH18 embryo clearly having it (their Figures 4B,E). This clearly corroborates our results. Adding the information obtained from previous and present data to published fate mapping results on the chick neural plate (Cobos et al., 2001; Sánchez-Arrones et al., 2009), the primordia of the four subpallial domains would seem to be arranged at neural plate stages in a topological AP sequence (with POA anteriormost and striatum caudalmost) and they all reach the prospective septal region that overlaps the median roof plate and the adjacent part of the alar plate.
One immediate consequence resulting from the corrected molecular definition of the POA and Dg subpallium domains is the need to redefine the reported AEP source of telencephalic oligodendrocytes (Timsit et al., 1995; Perez-Villegas et al., 1999; Nery et al., 2001; Olivier et al., 2001) in terms of the new Dg and POA domains. Present results and some other recent data (see below) suggest that the so-called AEP source of oligodendrocytes in fact may lie either in the POA or in the pallidal domain, rather than in the Dg.
Spatiotemporal Expression Pattern of Molecular Subpallial Markers
Our results suggest that the DPo primordium begins to be established as a Shh-expressing progenitor domain at stage HH16. Between HH19–23 a robust molecular specification code is established for most subpallial partitions (Figures 1 and 2). According to the fate map data correlated with Ganf, Pax6, and Fgf8 gene expression patterns recently reported by Sánchez-Arrones et al. (2009), the initial specification of the primordial telencephalic field apparently occurs at late neural plate stages (HH8–9). The pallio-subpallial boundary was likewise thought to be incipient at HH8–9, according to differential expression of Pax6 and Fgf8. FoxG1 (BF1), a general telencephalon marker, is expressed from HH9–10 onwards at the prospective telencephalic area (Bell et al., 2001). Pallial markers such as Emx1 and Emx2 start to be expressed in the prospective pallium at HH11–12 (Bell et al., 2001). The subpallial marker Nkx2.1 seems to be expressed initially at the POA, where it starts at stages HH12–13 (Pera and Kessel, 1998; Crossley et al., 2001; see present mapping at HH15) and its expression domain subsequently expands to include the prospective Dg and Pal, but is excluded from the striatal domain. Dlx genes become expressed throughout the subpallium from stage HH16–17 onwards (not shown, data from our collection). At HH16, Shh expression begins at the DPo, in a subarea of the Nkx2.1-expressing subpallial domain (Crossley et al., 2001; present results), and subsequently expands into the SCPo. This initial upregulation in DPo at HH16 coincides with first appearance of postmitotic neurons in this area (Puelles et al., 1987a,b).
Causes of Subpallial Shh Expression
Considering the sequential pattern of genetic regionalization in the subpallium, our next question is: how is preoptic Shh expression established?
Nkx2.1-null mice lose completely the POA Shh expression domain, which strongly suggests that Nkx2.1 function is required for Shh upregulation in the POA (Kimura, 1996; Takuma et al., 1998; Sussel et al., 1999; Sousa and Fishell, 2010). In the chick, moreover, early prechordal plate ablations at stages HH4+ to HH5+ produced embryos lacking Nkx2.1 expression in the subpallium at stages HH14–17, which were also devoid of POA Shh expression at stage HH17 (García-Calero et al., 2008). These experiments corroborate that local Nkx2.1 function precedes specification of POA Shh expression, and additionally suggest that an early prechordal induction effect – itself dependent on early Shh signaling (Kohtz et al., 1998; Gunhaga et al., 2000; Monuki and Walsh, 2001) – is necessary in order to obtain the Nkx2.1 and Shh subpallial domains. Ablations of the prechordal plate at HH6 did not any longer have effects on the subpallial expression patterns (García-Calero et al., 2008). It is still unclear precisely when occurs the prechordal signaling effect upon the prospective subpallium, since the prechordal tissue ablated in these HH4+/HH5+ experiments was placed under the hypothalamic floor and basal plate (i.e., out of range for influencing the subpallium as a source of diffusible signals). It was assumed in that study that the prechordal cells subsequently migrate to a position either contacting directly the prospective subpallium, or at least sufficiently close to make signaling via morphogen diffusion effective. The conjectured migratory phase for the potentially inducing prechordal cells may explain the time interval between the early stages mentioned, when prechordal ablation is effective, and the first appearance of dependent subpallial Nkx2.1 and Shh signals at stages HH12–13 and HH16, respectively.
Geng et al. (2008) showed an important role of Six3, an early general marker of the rostral forebrain (see Sánchez-Arrones et al., 2009), in the activation of Shh in the mouse subpallium. Thus Six3 probably acts upstream of both Nkx2.1 and Shh in the subpallium specification process.
The early distinction between the Nkx2.1-expressing MGE and non-Nkx2.1-expressing LGE domains within the subpallium appears to depend in part on a limited range of SHH signaling, either from the prechordal cells or from the incipient DPo domain, which according to present data starts expressing Nkx2.1 first. After Shh is upregulated in this area, the Nkx2.1 domain expands into Dg and Pal. Remarkably, Nkx2.1 results expressed along most of the DV dimension of the telencephalon in mutant Ptc−/− mice (Goodrich et al., 1999), suggesting that genes activated downstream of Shh itself – such as Ptc – may contribute to limit Shh effects to neighboring subpallial areas (see Ekker et al., 1995; Chiang et al., 1996; Shimamura and Rubenstein, 1997; Ohkubo et al., 2002; Fuccillo et al., 2004; Marcucio et al., 2005; Motoyama, 2006). Mice lacking Shh lose the MGE, even if some Nkx2.1 signal remains (Rallu et al., 2002).
Other collateral evidence in favor of a prechordal causal effect on the chicken subpallium derives from Pera and Kessel (1997), who placed a graft of prechordal plate lateral to the neural tube at HH5, producing an ectopic patch of Nkx2.1 expression, jointly with a downregulation of Pax6 around the graft at stage HH11. This ectopic effect occurred slightly before Nkx2.1 subpallial expression normally begins in the subpallium, but the observed parallel downregulation of Pax6 is consistent with a potential early prechordal “ventralizing” effect upon the telencephalon, which might precede the normal Nkx2.1 expression at stages HH12–13 (Pera and Kessel, 1997).
POA as a Progenitor Source for Pallial and Subpallial Migratory Streams
Our results suggest that Shh-positive cells originated from the dorsal preoptic region migrate tangentially out of this domain, translocating mainly to other Nkx2.1-expressing subpallial domains. Experimental evidence in mice raises the notion that diverse cell types may in fact migrate out from this domain. Studies centered on oligodendrocyte origins have proposed an important AEP source, which we have reinterpreted above as possibly corresponding in part, at least, to dorsal POA (Timsit et al., 1995; Perez-Villegas et al., 1999; Nery et al., 2001; Olivier et al., 2001). In the chick, expression of PLP/DM20 first appears at the “AEP” (dorsal POA) at stages HH24–26, and the relevant progenitor cells in the ventricular zone coexpress Shh and PLP (Perez-Villegas et al., 1999). Immature oligodendrocytes disseminate from there into subpallial and pallial domains, but such cells apparently do not express Shh once they enter the mantle zone (Perez-Villegas et al., 1999; Nery et al., 2001; Olivier et al., 2001). We assume from this correlation that the early Shh-positive migrating cells we report here probably are not oligodendrocytes. If oligodendrocytes start to be produced in the POA at HH24, they would represent a late wave of derivatives, possibly having differential properties relative to earlier derivatives which retain Shh expression in the mantle (i.e., starting to be produced earlier, at stage HH16).
Genetic cell lineage tracing using Shh-Cre recombination in mice (Flandin et al., 2010) recently confirmed that Olig2-positive oligodendrocytes that eventually populate commissural axon tracts in the septum derive from the POA domain. In the context of other data on oligodendrocytes mentioned above, this strongly indicates that some oligodendrocytes arise from Shh-positive POA progenitors, but they downregulate transcription of Shh as they start to differentiate in the mantle. The set of derivatives observed by Flandin et al. (2010) should include in principle all DPo and SCPo derivatives, irrespective whether they continue expressing Shh in the mantle or not. One conceivable qualification of this assumption is that we saw faint Shh expression in a few ventricular cells of the Dg domain (see Figures 2F,L). This low signal might be still able to label additionally some Dg derivatives via Shh-Cre recombination in mice. The clearest candidates to represent such a collateral labeling effect are the striatal cholinergic neurons, which are otherwise held to likely be produced at the Dg (Zhao et al., 2003; Fragkouli et al., 2005; Flames et al., 2007; Flandin et al., 2010).
Our data clearly indicate that some early derivatives of the DPo and SCPo progenitor areas continue expressing the Shh marker as they migrate lateralward in the mantle and finally incorporate into septal, normal pallidal and ectopic pallidal formations of the subpallium, as well as into a restricted intermediate stratum of the striatopallidal circumventricular organ (SPO; Bardet et al., 2006; present data) and the extended amygdala and medial amygdaloid territories. Abellan and Medina (2009) also found previously that POA-derived Shh-positive cells reach extended and medial parts of the amygdala, and García-López et al. (2008) reported similar results in the mouse.
Apart the SPO population, the definitive striatal domain results largely devoid of Shh-expressing cells in our HH36 material (see also García-López et al., 2008; Abellan and Medina, 2009). At early and intermediate stages, the outer striatal mantle stratum does show Shh-positive cells, but, as a more mature structure develops, such cells result restricted to the ectopic pallidum, which is thought to be homologous to the mammalian globus pallidus (present data; Reiner et al., 1984; Puelles et al., 2000, 2007). This is corroborated by the persistent expression of Nkx2.1 by its population (Figures 6D,H–L). In contrast, Flandin et al. (2010) found some cells derived from the dorsal POA within the mouse striatum. These were either oligodendrocytes, cholinergic neurons or GABAergic interneurons. We conjecture, because of other data on production of cholinergic cells (Zhao et al., 2003; Fragkouli et al., 2005; Abellan and Medina, 2009), and our comments above about low Shh expression in Dg ventricular cells, that the cholinergic cells labeled by Flandin et al. (2010) in the striatum may have arisen in the adjoining Dg domain. The accompanying labeled striatal oligodendrocytes and GABAergic interneurons may well have come from the POA. We accordingly cannot exclude that some migrating Shh-positive cells that penetrate the avian and mouse striatum subsequently lose their transcription of Shh and Nkx2.1.
In the mouse, there is the curious observation that, although Shh expression disappears in the re-patterned subpallial region in Nkx2.1 mutants (Sussel et al., 1999; Nery et al., 2001), it is conserved in the medial amygdala nuclear primordium (Figure 5 in Nery et al., 2001). This raises the question of where this amygdaloid population comes from, or why it does not depend on Nkx2.1 function.
Additionally, a set of POA-related postmitotic cells express Nkx5.1 in the mouse (Rinkwitz-Brandt et al., 1995). Using Nkx5.1-Cre mice crossed with a R26R-EYFP reporter strain, it was demonstrated that the POA is a source of cortical GABAergic interneurons (Gelman et al., 2009). This work documented that Nkx5.1 is expressed in the mantle layer, but the relevant population does not express Shh ( Gelman et al., 2009; their Figures 4A,B). This preoptic Nkx5-1-positive migration of GABAergic cells seems therefore quite distinct from the stream of Shh-expressing cells considered here, which clearly do not reach pallial territories. These elements might nevertheless explain the GABAergic neurons found by Flandin et al. (2010) within the mouse striatum.
Using in utero electroporation experiments targeting the POA of Nkx2.1-Cre mice at E12.5, and analyzing the position of migrated neurons at E15.5, GFP-positive cells were found in the globus pallidus (Nóbrega-Pereira et al., 2010). In the same work, fate-mapping studies in P30 mice, using a Dbx1-Cre line, revealed that a small fraction of parvalbumin expressing neurons in the globus pallidus derive from Dbx1-expressing progenitors in the POA (Nóbrega-Pereira et al., 2010). A similar study by Hirata et al. (2009) showed that a cell population migrated from the POA and expressing Dbx1 produces an inhibitory neuron subtype in the mature medial amygdala nucleus (the major inhibitory output nucleus of the amygdala). Nkx2.1-Cre fate mapping also showed that the medial amygdala is partly derived from a Nkx2.1-expressing domain (Xu et al., 2008). All these pallidal and amygdaloid cell populations derived from the POA and expressing Nkx2.1 or Dbx1 are candidates to correlate with the cells that migrate expressing Shh.
Analysis of Shh-Cre labeled derivatives at E11.5, E13.5, and P0 (genetic fate map) recently showed that the ventromedial nucleus of amygdala was populated by such cells, whereas the dorsomedial nucleus was Nkx2.1 positive, but does not derive from the Shh lineage (Flandin et al., 2010). This study also showed that around 70% of parvalbumin-positive globus pallidus neurons, as well as a small fraction of cortical and striatal GABAergic interneurons (largely parvalbumin-containing ones), apart the striatal cholinergic neurons and some Olig2+ oligodendrocytes, derive from the Shh-Cre lineage.
Flandin et al. (2010) also fate-mapped the Nkx2-1f/−;ShhCre/+ mutant strain, showing that Shh-expressing derivatives failed to populate the globus pallidus, and there was no stream of tangentially migrating Shh-expressing cells at the subventricular zone of the medial ganglionic eminence. Instead, the labeled cells accumulate in the periventricular mantle zone of the DPo progenitor domain (Flandin et al., 2010). This lineage tracing is accordingly consistent with our description of Shh-positive DPo and SCPo derivatives that migrate laterally and rostrally in the mantle zone, traversing the Dg and colonizing mainly pallidal domains. This stream apparently represents only a subset of all cell types migrated out of the POA. These seem to correspond to pallidal and septal neurons that maintain Nkx2.1 expression and later differentiate as SST+ and PV+ interneurons (Xu et al., 2005, 2010).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The present study was supported by a Spanish MEC grant BFU 2008-04156, NIH grant 1-R01-MH070370-01A2, SENECA Foundation grant 04548/GERM/06, and CIBER en Enfermedades Raras U736 (Luis Puelles, José L. E. Ferran, Luisa Sanchez-Arrones). Sylvia M. Bardet is a former MEC doctoral fellow at Murcia University, and now a postdoctoral researcher at the University Limoges; and Luisa Sanchez-Arrones is a FPI predoctoral fellow of the Spanish Ministry of Science.
Footnotes
References
Abellan, A., and Medina, L. (2009). Subdivisions and derivatives of the chicken subpallium based on expression of LIM and other regulatory genes and markers of neuron subpopulations during development. J. Comp. Neurol. 515, 465–501.
Agarwala, S., Sanders, T. A., and Ragsdale, C. W. (2001). Sonic hedgehog control of size and shape in midbrain pattern formation. Science 291, 2147–2150.
Bardet, S. M. (2007). Organización morfológica y citogenética del hipotálamo del pollo sobre base de mapas moleculares. Doctoral thesis, Neuroscience programme, University of Murcia, Murcia.
Bardet, S. M., Cobos, I., Puelles, E., Martínez-de-la-Torre, M., and Puelles, L. (2006). The chicken lateral septal organ and other circumventricular organs form in the striatal domain, abutting the molecular striatopallidal border. J. Comp. Neurol. 499, 745–767.
Bardet, S. M., Martinez-de-la-Torre, M., Northcutt, R. G., Rubenstein, J. L., and Puelles, L. (2008). Conserved pattern of OTP-positive cells in the paraventricular nucleus and other hypothalamic sites of tetrapods. Brain Res. Bull. 75, 231–235.
Bell, E., Ensini, M., Gulisano, M., and Lumsden, A. (2001). Dynamic domains of gene expression in the early avian forebrain. Dev. Biol. 236, 76–88.
Briscoe, J., and Ericson, J. (2001). Specification of neuronal fates in the ventral neural tube. Curr. Opin. Neurobiol. 11, 43–49.
Briscoe, J., Pierani, A., Jessell, T. M., and Ericson, J. (2000). A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube. Cell 101, 435–445.
Bulfone, A., Puelles, L., Porteus, M. H., Frohman, M. A., Martin, G. R., and Rubenstein, J. L. (1993). Spatially restricted expression of Dlx-1, Dlx-2 (Tes-1), Gbx-2, and Wnt-3 in the embryonic day 12.5 mouse forebrain defines potential transverse and longitudinal segmental boundaries. J. Neurosci. 13, 3155–3172.
Chang, D. T., Lopez, A., von Kessler, D. P., Chiang, C., Simandl, B. K., Zhao, R., Seldin, M. F., Fallon, J. F., and Beachy, P. A. (1994). Products, genetic linkage and limb patterning activity of a murine hedgehog gene. Development 120, 3339–3353.
Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H., and Beachy, P. A. (1996). Cyclopia and defective axial patterning in mice lacking sonic hedgehog gene function. Nature 383, 407–413.
Cobos, I., Shimamura, K., Rubenstein, J. L. R., Martínez, S., and Puelles, L. (2001). Fate map of the avian anterior forebrain at the four-somite stage, based on the analysis of quail-chick chimeras. Dev. Biol. 239, 46–67.
Crossley, P. H., Martinez, S., Ohkubo, Y., and Rubenstein, J. L. (2001). Coordinate expression of Fgf8, Otx2, Bmp4, and Shh in the rostral prosencephalon during development of the telencephalic and optic vesicles. Neuroscience 108, 183–206.
Echelard, Y., Vassileva, G., and McMahon, A. P. (1994). Cis-acting regulatory sequences governing Wnt-1 expression in the developing mouse CNS. Development 120, 2213–2224.
Ekker, S. C., Ungar, A. R., Greenstein, P., von Kessler, D. P., Porter, J. A., Moon, R. T., and Beachy, P. A. (1995). Patterning activities of vertebrate hedgehog proteins in the developing eye and brain. Curr. Biol. 5, 944–955.
Ferran, J. L., Sanchez-Arrones, L., Sandoval, J. E., and Puelles, L. (2007). A model of early molecular regionalization in the chicken embryonic pretectum. J. Comp. Neurol. 505, 379–403.
Flames, N., Pla, R., Gelman, D. M., Rubenstein, J. L., Puelles, L., and Marín, O. (2007). Delineation of multiple subpallial progenitor domains by the combinatorial expression of transcriptional codes. J. Neurosci. 27, 9682–9695.
Flandin, P., Kimura, S., and Rubenstein, J. L. (2010). The progenitor zone of the ventral medial ganglionic eminence requires nkx2-1 to generate most of the globus pallidus but few neocortical interneurons. J. Neurosci. 30, 2812–2823.
Fragkouli, A., Hearn, C., Errington, M., Cooke, S., Grigoriou, M., Bliss, T., Stylianopoulou, F., and Pachnis, V. (2005). Loss of forebrain cholinergic neurons and impairment in spatial learning and memory in LHX7-deficient mice. Eur. J. Neurosci. 21, 2923–2938.
Fuccillo, M., Rallu, M., McMahon, A. P., and Fishell, G. (2004). Temporal requirement for hedgehog signaling in ventral telencephalic patterning. Development 131, 5031–5040.
García-Calero, E., Fernández-Garre, P., Martínez, S., and Puelles, L. (2008). Early mammillary pouch specification in the course of prechordal ventralization of the forebrain tegmentum. Dev. Biol. 320, 366–377.
García-López, M., Abellán, A., Legaz, I., Rubenstein, J. L., Puelles, L., and Medina, L. (2008). Histogenetic compartments of the mouse centromedial and extended amygdala based on gene expression patterns during development. J. Comp. Neurol. 506, 46–74.
Gelman, D. M., Martini, F. J., Nobrega-Pereira, S., Pierani, A., Kessaris, N., and Marín, O. (2009). The embryonic preoptic area is a novel source of cortical GABAergic interneurons. J. Neurosci. 29, 9380–9389.
Geng, X., Speirs, C., Lagutin, O., Inbal, A., Liu, W., Solnica-Krezel, L., Jeong, Y., Epstein, D. J., and Oliver, G. (2008). Haploinsufficiency of Six3 fails to activate Sonic hedgehog expression in the ventral forebrain and causes holoprosencephaly. Dev. Cell 15, 236–247.
Goodrich, L. V., Jung, D., Higgins, K. M., and Scott, M. P. (1999). Overexpression of ptc1 inhibits induction of Shh target genes and prevents normal patterning in the neural tube. Dev. Biol. 211, 323–334.
Gunhaga, L., Jessell, T. M., and Edlund, T. (2000). Sonic hedgehog signaling at gastrula stages specifies ventral telencephalic cells in the chick embryo. Development 127, 3283–3293.
Hamburger, V., and Hamilton, H. L. (1951). A series of normal stages in the development of the chick embryo. J. Morphol. 88, 49–92.
Hidalgo-Sánchez, M., Martínez-de-la-Torre, M., Alvarado-Mallart, R. M., and Puelles, L. (2005). Distinct pre-isthmic domain, defined by overlap of Otx2 and Pax2 expression domains in the chicken caudal midbrain. J. Comp. Neurol. 483, 17–29.
Hirata, T., Li, P., Lanuza, G. M., Cocas, L. A., Huntsman, M. M., and Corbin, J. G. (2009). Identification of distinct telencephalic progenitor pools for neuronal diversity in the amygdala. Nat. Neurosci. 12, 141–149.
Kala, K., Haugas, M., Lillevali, K., Guimera, J., Wurst, W., Salminen, M., and Partanen, J. (2009). Gata2 is a tissue-specific post-mitotic selector gene for midbrain GABAergic neurons. Development 136, 253–262.
Kessaris, N., Fogarty, M., Iannarelli, P., Grist, M., Wegner, M., and Richardson, W. D. (2006). Competing waves of oligodendrocytes in the forebrain and postnatal elimination of an embryonic lineage. Nat. Neurosci. 9, 173–179.
Kiecker, C., and Lumsden, A. (2004). Hedgehog signaling from the ZLI regulates diencephalic regional identity. Nat. Neurosci. 7, 1242–1249.
Kimura, S. (1996). The T/ebp null mouse: thyroid specific enhancer-binding protein is essential for the oganogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes Dev. 10, 60–69.
Kohtz, J. D., Baker, D. P., Corte, G., and Fishell, G. (1998). Regionalization within the mammalian telencephalon is mediated by changes in responsiveness to Sonic hedgehog. Development 125, 5079–5089.
Krauss, S., Concordet, J. P., and Ingham, P. W. (1993). A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell 75, 1431–1444.
Legaz, I., Olmos, L., Real, M. A., Guirado, S., Davila, J. C., and Medina, L. (2005). Development of neurons and fibers containing calcium binding proteins in the pallial amygdala of mouse, with special emphasis on those of the basolateral amygdalar complex. J. Comp. Neurol. 488, 492–513.
Manning, L., Ohyama, K., Saeger, B., Hatano, O., Wilson, S., Logan, M., and Placzek, M. (2006). Regional morphogenesis in the hypothalamus: a BMP-Tbx2 pathway coordinates fate and proliferation through Shh downregulation. Dev. Cell 11, 873–885.
Marcucio, R. S., Cordero, D. R., Hu, D., and Helms, J. A. (2005). Molecular interactions coordinating the development of the forebrain and face. Dev. Biol. 284, 48–61.
Marín, O., Anderson, S. A., and Rubenstein, J. L. (2000). Origin and molecular specification of striatal interneurons. J. Neurosci. 20, 6063–6076.
Marín, O., and Rubenstein, J. L. R. (2002). “Patterning, regionalization, and cell differentiation in the forebrain”. in Mouse Development. Pattering, Morphogenesis, and Organogenesis, eds J. Arossant and P. P. L. Tam (San Diego: Academic Press), 75–106.
Martí, E., Takada, R., Bumcrot, D. A., Sasaki, H., and McMahon, A. P. (1995). Distribution of Sonic hedgehog peptides in the developing chick and mouse embryo. Development 121, 2537–2547.
Monuki, E. S., and Walsh, C. A. (2001). Mechanisms of cerebral cortical patterning in mice and humans. Nat. Neurosci. 4, 1199–1206.
Mori, T., Yuxing, Z., Takaki, H., Takeuchi, M., Iseki, K., Hagino, S., Kitanaka, J., Takemura, M., Misawa, H., Ikawa, M., Okabe, M., and Wanaka, A. (2004). The LIM homeobox gene, L3/Lhx8, is necessary for proper development of basal forebrain cholinergic neurons. Eur. J. Neurosci. 19, 3129–3141.
Motoyama, J. (2006). Essential roles of Gli3 and sonic hedgehog in pattern formation and developmental anomalies caused by their dysfunction. Congenit. Anom. (Kyoto) 46, 123–128.
Nery, S., Wichterle, H., and Fishell, G. (2001). Sonic hedgehog contributes to oligodendrocyte specification in the mammalian forebrain. Development 128, 527–540.
Nóbrega-Pereira, S., Gelman, D., Bartolini, G., Pla, R., Pierani, A., and Marín, O. (2010). Origin and molecular specification of globus pallidus neurons. J. Neurosci. 30, 2824–2834.
Ohkubo, Y., Chiang, C., and Rubenstein J. L. (2002). Coordinate regulation and synergistic actions of BMP4, SHH and FGF8 in the rostral prosencephalon regulate morphogenesis of the telencephalic and optic vesicles. Neuroscience 111, 1–17.
Ohyama, K., Das, R., and Placzek, M. (2008). Temporal progression of hypothalamic patterning by a dual action of BMP. Development 135, 3325–3331.
Ohyama, K., Ellis, P., Kimura, S., and Placzek, M. (2005). Directed differentiation of neural cells to hypothalamic dopaminergic neurons. Development 132, 5185–5197.
Olivier, C., Cobos Silleros, I., Perez-Villegas, E. M., Spassky, N., Zalc, B., Thomas, J. L., and Martinez, S. (2001). Monofocal origin of telencephalic oligodendrocytes in the chick embryo: the entopeduncular area. Development 128, 1757–1769.
Patten, I., and Placzek, M. (2000). The role of Sonic hedgehog in neural tube patterning. Cell. Mol. Life Sci. 57, 1695–1708.
Pera, E. M., and Kessel, M. (1997). Patterning of the chick forebrain anlage by the prechordal plate. Development 124, 4153–4162.
Pera, E. M., and Kessel, M. (1998). Demarcation of ventral territories by the homeobox gene NKX2.1 during early chick development. Dev. Genes Evol. 208, 168–171.
Perez-Villegas, E. M., Olivier, C., Spassky, N., Poncet, C., Cochard, P., Zalc, B., Thomas, J. L., and Martínez, S. (1999). Early specification of oligodendrocytes in the chick embryonic brain. Dev. Biol. 216, 98–113.
Placzek, M., Jessell, T. M., and Dodd, J. (1993). Induction of floor plate differentiation by contact-dependent, homeogenetic signals. Development 117, 205–218.
Pombero, A., and Martinez, S. (2009). Telencephalic morphogenesis during the process of neurulation: an experimental study using quail-chick chimeras. J. Comp. Neurol. 512, 784–797.
Puelles, L. (1995) A segmental morphological paradigm for understanding vertebrate forebrains. Brain Behav. Evol. 46, 319-337.
Puelles, L., Domenech-Ratto, G., and Martinez-de-la-Torre, M. (1987a). Localization of the rostral end of the longitudinal brain axis: review of an old topic in the light of marking experiments on the closing rostral neuropore. J. Morphol. 194, 163–171.
Puelles, L., Amat, J. A., and Martínez-de-la-Torre, M. (1987b). Segment-related, mosaic neurogenetic pattern in the forebrain and mesencephalon of early chick embryos. I. Topography of AChE-positive neuroblasts up to stage HH18. J. Comp. Neurol. 266, 147–268.
Puelles, L., Fernández-Garre, P., Sánchez-Arrones, L., García-Calero, E., and Rodríguez-Gallardo, L. (2005). Correlation of a chicken stage 4 neural plate fate map with early gene expression patterns. Brain Res. Brain Res. Rev. 49, 167–178.
Puelles, L., Kuwana, E., Puelles, E., Bulfone, A., Shimamura, K., Keleher, J., Smiga, S., and Rubenstein, J. L. (2000). Pallial and subpallial derivatives in the embryonic chick and mouse telencephalon, traced by the expression of the genes Dlx-2, Emx-1, Nkx-2.1, Pax-6, and Tbr-1. J. Comp. Neurol. 424, 409–438.
Puelles, S., Martinez, S., Martinez-De-La-Torre, M., and Rubenstein, J. L. R. (2004) “Gene maps and related histogenetic domains in the forebrain and midbrain,” in The Rat Nervous System (3rd Edn), ed. G. Paxinos (San Diego: Elsevier) 3–25.
Puelles, L., Martínez-de-la-Torre, M., Paxinos, G., Watson, C., and Martínez, S. (2007). The Chick Brain in Stereotaxic Coordinates. An Atlas Featuring Neuromeric Subdivisions and Mammalian Homologies. San Diego: Academic Press, Elsevier.
Puelles, L., and Rubenstein, J. L. (1993). Expression patterns of homeobox and other putative regulatory genes in the embryonic mouse forebrain suggest a neuromeric organization. Trends Neurosci. 16, 472–479.
Puelles, L., and Rubenstein, J. L. (2003). Forebrain gene expression domains and the evolving prosomeric model. Trends Neurosci. 26, 469–476.
Rallu, M., Corbin, J. G., and Fishell, G. (2002). Parsing the prosencephalon. Nat. Rev. Neurosci. 3, 943–951.
Reiner, A., Davis, B. M., Brecha, N. C., and Karten, H. J. (1984). The distribution of enkephalin-like immunoreactivity in the telencephalon of the adult and developing domestic chicken. J. Comp. Neurol. 228, 245–262.
Riddle, R. D., Johnson, R. L., Laufer, E., and Tabin, C. (1993). Sonic hedgehog mediates the polarizing activity of the ZPA. Cell 75, 1401–1416.
Rinkwitz-Brandt, S., Justus, M., Oldenettel, I., Arnold, H. H., and Bober, E. (1995). Distinct temporal expression of mouse Nkx-5.1 and Nkx-5.2 homeobox genes during brain and ear development. Mech. Dev. 52, 371–381.
Roelink, H., Augsburger, A., Heemskerk, J., Korzh, V., Norlin, S., Ruiz, I., Altaba, A., Tanabe, Y., Placzek, M., Edlund, T., and Jessell, T. M. (1994). Floor plate and motor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressed by the notochord. Cell 76, 761–775.
Rubenstein, J. L., and Beachy, P. A. (1998). Patterning of the embryonic forebrain. Curr. Opin. Neurobiol. 8, 18–26.
Rubenstein, J. L., Shimamura, K., Martinez, S., and Puelles, L. (1998). Regionalization of the prosencephalic neural plate. Annu. Rev. Neurosci. 21, 445–477.
Sánchez-Arrones, L., Ferran, J. L., Rodríguez-Gallardo, L., and Puelles, L. (2009). Incipient forebrain boundaries traced by differential gene expression and fate mapping in the chick neural plate. Dev. Biol. 335, 43–65.
Schaeren-Wiemers, N., and Gerfin-Moser, A. (1993). A single protocol to detect transcripts of various types and expression levels in neural tissue and cultured cells: in situ hybridization using digoxigenin-labeled cRNA probes. Histochemistry 100, 431–440.
Scholpp, S., Wolf, O., Brand, M., and Lumsden, A. (2006). Hedgehog signalling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon. Development 133, 855–864.
Shimamura, K., Hartigan, D. J., Martinez, S., Puelles, L., and Rubenstein, J. L. (1995). Longitudinal organization of the anterior neural plate and neural tube. Development 121, 3923–3933.
Shimamura, K., Hirano, S., McMahon, A. P., and Takeichi, M. (1994). Wnt-1-dependent regulation of local E-cadherin and (N-catenin expression in the embryonic mouse brain. Development 120, 2225–2234.
Shimamura, K., and Rubenstein, J. L. (1997). Inductive interactions direct early regionalization of the mouse forebrain. Development 124, 2709–2718.
Sieber, M. A., Storm, R., Martinez-de-la-Torre, M., Müller, T., Wende, H., Reuter, K., Vasyutina, E., and Birchmeier, C. (2007). Lbx1 acts as a selector gene in the fate determination of somatosensory and viscerosensory relay neurons in the hindbrain. J. Neurosci. 27, 4902–4909.
Sousa, V. H., and Fishell, G. (2010). Sonic hedgehog functions through dynamic changes in temporal competence in the developing forebrain. Curr. Opin. Genet. Dev. 20, 1–9.
Spassky, N., Heydon, K., Mangatal, A., Jankovski, A., Olivier, C., Queraud-Lesaux, F., Goujet-Zalc, C., Thomas, J. L., and Zalc, B. (2001). Sonic hedgehog-dependent emergence of oligodendrocytes in the telencephalon: evidence for a source of oligodendrocytes in the olfactory bulb that is independent of PDGFRalpha signaling. Development 128, 4993–5004.
Suarez, J., Dávila, J. C., Real, M. A., Guirado, S., and Medina, L. (2006). Calcium-binding proteins, neuronal nitric oxide synthase and GABA help to distinguish different pallial areas in the developing and adult chicken: I. Hippocampal formation and hyperpallium. J. Comp. Neurol. 497, 751–771.
Sussel, L., Marín, O., Kimura, S., and Rubenstein, J. L. (1999). Loss of Nkx2.1 homeobox gene function results in a ventral to dorsal molecular respecification within the basal telencephalon: evidence for a transformation of the pallidum into the striatum. Development 126, 3359–3370.
Szabó, N. E., Zhao, T., Zhou, X., and Alvarez-Bolado, G. (2009). The role of Sonic hedgehog of neural origin in thalamic differentiation in the mouse. J. Neurosci. 29, 2453–2466.
Taglialatela, P., Soria, J. M., Caironi, V., Moiana, A., and Bertuzzi, S. (2004). Compromised generation of GABAergic interneurons in the brains of Vax1−/− mice. Development 131, 4239–4249.
Takuma, N., Sheng, H. Z., Furuta, Y., Ward, J. M., Sharma, K., Hogan, B. L., Pfaff, S. L., Westphal, H., Kimura, S., and Mahon, K. A. (1998). Formation of Rathke’s pouch requires dual induction from the diencephalon. Development 125, 4835–4840.
Tekki-Kessaris, N., Woodruff, R., Hall, A. C., Gaffield, W., Kimura, S., Stiles, C. D., Rowitch, D. H., and Richardson, W. D. (2001). Hedgehog-dependent oligodendrocyte lineage specification in the telencephalon. Development 128, 2545–2554.
Timsit, S., Martinez, S., Allinquant, B., Peyron, F., Puelles, L., and Zalc, B. (1995). Oligodendrocytes originate in a restricted zone of the embryonic ventral neural tube defined by DM-20 mRNA expression. J. Neurosci. 15, 1012–1024.
Ulloa, F., and Briscoe, J. (2007). Morphogens and the control of cell proliferation and patterning in the spinal cord. Cell Cycle 6, 2640–2649.
Vieira, C., Garda, A. L., Shimamura, K., and Martinez, S. (2005). Thalamic development induced by Shh in the chick embryo. Dev. Biol. 284, 351–363.
Vue, T. Y., Bluske, K., Alishahi, A., Yang, L. L., Koyano-Nakagawa, N., Novitch, B., and Nakagawa, Y. (2009). Sonic hedgehog signaling controls thalamic progenitor identity and nuclei specification in mice. J. Neurosci. 29, 4484–4497.
Wagner, M., Thaller, C., Jessell, T., and Eichele, G. (1990). Polarizing activity and retinoid synthesis in the floor plate of the neural tube. Nature 345, 819–822.
Xu, Q., Guo, L., Moore, H., Waclaw, R. R., Campbell, K., and Anderson, S. A. (2010). Sonic hedgehog signaling confers ventral telencephalic progenitors with distinct cortical interneuron fates. Neuron 65, 328–340.
Xu, Q., Tam, M., and Anderson, S. A. (2008). Fate mapping Nkx2.1-lineage cells in the mouse telencephalon. J. Comp. Neurol. 506, 16–29.
Xu, Q., Wonders, C. P., and Anderson, S. A. (2005). Sonic hedgehog maintains the identity of cortical interneuron progenitors in the ventral telencephalon. Development 132, 4987–4998.
Yamada, T., Placzek, M., Tanaka, H., Dodd, J., and Jessell, T. M. (1991). Control of cell pattern in the developing nervous system: polarizing activity of the floor plate and notochord. Cell 64, 635–647.
Keywords: chick embryo, alar plate, forebrain, preoptic area, anterior entopeduncular area, subpallium, telencephalon, pallidum
Citation: Bardet SM, Ferran JLE, Sanchez-Arrones L and Puelles L (2010) Ontogenetic expression of Sonic hedgehog in the chicken subpallium. Front. Neuroanat. 4:28. doi: 10.3389/fnana.2010.00028
Received: 23 March 2010;
Paper pending published: 06 April 2010;
Accepted: 09 June 2010;
Published online: 21 July 2010
Edited by:
George Paxinos, University of New South Wales, AustraliaReviewed by:
Charles R. Watson, Curtin University, Australia; Prince of Wales Medical Research Institute, AustraliaTomomi Shimogori, RIKEN Brain Science Institute, Japan
Copyright: © 2010 Bardet, Ferran, Sanchez-Arrones and Puelles. This is an open-access article subject to an exclusive license agreement between the authors and the Frontiers Research Foundation, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are credited.
*Correspondence: Luis Puelles, Department of Human Anatomy and Psychobiology, Medical School, University of Murcia, Murcia E30071, Spain. e-mail:cHVlbGxlc0B1bS5lcw==
Abbreviations: ABb, alar-basal boundary; ac, anterior commissure; AEP, anterior entopeduncular area; Amygd, amygdala; AP, anteroposterior axis; BstL, bed nucleus of stria terminalis, lateral part; BstM, bed nucleus of stria terminalis, medial part; CNS, central nervous system; cp, choroid plexus; csm, cortico-septo-mesencephalic tract; D, diencephalon; Dg, diagonal domain; DgSe, diagonal septum; DPo, dorsal preoptic domain; DV, dorsoventral axis; ePal, ectopic pallidum; fp, floor plate; hbp, hypothalamic basal plate; hic, hippocampal commissure (pallial commissure); HiC, nucleus of the hippocampal commissure; hy, hypothalamus; III, third ventricle; Inf, infundibulum; ivf, interventricular foramen; IPPal, pallidal intrapeduncular nucleus; IPSt, striatal intrapeduncular nucleus; L, lateral; LSe, lateral septum; lt, lamina terminalis; lv, lateral ventricle; lvc, caudal horn of lateral ventricle; lvr, rostral horn of lateral ventricle; M, mesencephalon; MAm, medial amygdala; me, median eminence; MnPo, median preoptic nucleus; MPal, medial pallidum; MSe, medial septum; nt, notochord; OB, olfactory bulb; oc, optic chiasm; on, optic nerve; os, optic stalk; ot, optic tract; Ot, olfactory tubercle; P, pallium; p1, prosomere 1; p2, prosomere 2; p3, prosomere 3; Pa, paraventricular nucleus; Pal, pallidum; PalSe, pallidal septum; PaRo, paraventricular nucleus, rostral part; pe, telencephalic peduncle; POA, preoptic area; POH, preoptohypothalamic transition area; por, preoptic recess; PTh, prethalamus; PThE, prethalamic eminence; R, rhombencephalon; SCPo, septocommissural preoptic area; Se, septum; SI, substantia innominata; sP, subpallium; SPa, subparaventricular area; SPO, striatopallidal organ; SPOd, striatopallidal organ, distal part; SPOv, striatopallidal organ, ventricular part; St, striatum; StSe, striatal septum; T, telencephalon; Tel field, telencephalic field; Th, thalamus; v, ventricular zone; VP, ventral pallium; vPal, ventral pallidum; VPo, ventral preoptic domain; zli, zona limitans intrathalamica.