 Alba Vicario
Alba Vicario Loreta Medina
Loreta Medina- Department of Experimental Medicine, Laboratory of Evolutionary Developmental Neurobiology, Lleida's Institute for Biomedical Research-Dr. Pifarre Foundation (IRBLleida), University of Lleida, Lleida, Spain
A Corrigendum on
Genetic Identification of the Central Nucleus and Other Components of the Central Extended Amygdala in Chicken During Development
by Vicario, A., Abellán, A., Desfilis, E., and Medina, L. (2014). Front. Neuroanat. 8:90. doi: 10.3389/fnana.2014.00090
In the original article, there is an error related to the identification of corticotropin-releasing factor, as explained next.
When trying to continue our work on the chicken extended amygdala, it came to our attention an error on the identification of chicken ‘Corticotropin-releasing factor' (abbreviated CRF or cCRF) in our previous publication. In particular, the sequence we used to synthetize riboprobes in order to analyze brain expression, Genbank accession no. NM_204454, does not correspond to chicken CRF but to its receptor 2 (CRFR2). Therefore, the text mentioning cCRF should refer to cCRFR2. The rest of the results of the article on Pax6, Islet1, Nkx2.1, pENK, SOM, and TH, as well as results on connections, are not affected by this correction.
Below we provide a list of the specific sections that need correction (cCRF should be cCRFR2):
A correction has been made to Results, Expression of cCRFR2, cSOM, and cTH, and Comparison to Other Markers, Paragraph 2. The corrected paragraph is shown below.
At early embryonic stages, there was no expression of cCRFR2 in the subpallium, although this gene was strongly expressed in the medial pallium (not shown). cCRFR2 only started to be weakly expressed in the striatal mantle by E14, and by E18 weak to moderate expression could be appreciated in the Ceov and the peri-INP island field (Figure 8B). The cCRFR2 expression in the striatum, peri INP island field and Ceov largely overlapped with that of cIslet1 (compare Figures 8B,C). By E19, a few cells expressing cCRFR2 were also seen in the BSTLd and a moderate number of them were found in SpAr (Figure 5J and Table 3).
Corrections have also been made to the captions for Figures 5 and 8. The corrected captions are shown below.
Figure 5. Expression of cIslet1, cPax6 and other genes in the embryonic telencephalon of chicken, at the level of the INP and the peri-INP island field. (A–K) Digital images of frontal sections through the telencephalon of chicken embryos (from E14 to E19) hybridized for cIslet1, cPax6, or cNkx2.1 (A–D, H–I) or for the phenotype markers genes cpENK, cCRFR2, or cTH (E–G,J,K). Some of the hybridized sections are also immunostained (brown staining) for calbindin (D) or Nkx2.1 (E). The sections shown are at the level of the INP or the peri-INP island field (pINP). (A–J) are high magnification images of the sections shown in (A′-J′), respectively. (K,K′) show details cTH-expressing cells in the striatal capsule (StC) and the rostral part of the subpallial extended amygdala (SpAr). See text for more details. For abbreviations, see list. The arrows in (C,D) point to a bridge of pallidal cells extending into the globus pallidus, and traversing the INP (this cell bridge expresses Nkx2.1, but is negative for Islet1). The arrows in (G,H) point to bridges of striatal cells extending from the lateral striatum into islands of the pINP, and traversing the gobus pallidus. The asterisks in (F,H–H′) indicate artifacts in the tissue. Scale bars: A = 300 μm (applies to A–J); A′ = 1 mm (applies to A′–J′); K = 400 μm; K′ = 200 μm.
Figure 8. Expression of cpENK, cCRFR2, cSOM, and cTH in the central extended amygdala of chicken embryos at intermediate or late stages. (A–F) Digital images of frontal sections of the telencephalon of chicken embryos (from E14 to E19), hybridized for the phenotype markers cpENK, cCRFR2, cTH, or cSOM. (A–F) are high magnification images of the sections shown in (A′-F′), respectively. All the sections are at the level of the Ceov and surrounding areas. See text for more details. For abbreviations, see list. Scale bars: A = 300 μm (applies to A–F); A′ = 1 mm (applies to A′-F′).
Corrections have also been made to Tables 1–3. The row ‘cCRF' has been amended to state ‘cCRFR2.' The corrected tables are shown below.

Table 1. Expression of several genes in the central extended amygdala and some surrounding areas of chicken at E9-E10.
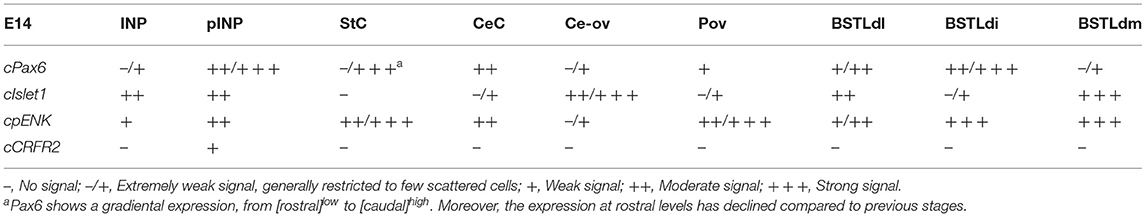
Table 2. Expression of several genes in the central extended amygdala and some surrounding areas of chicken at E14.
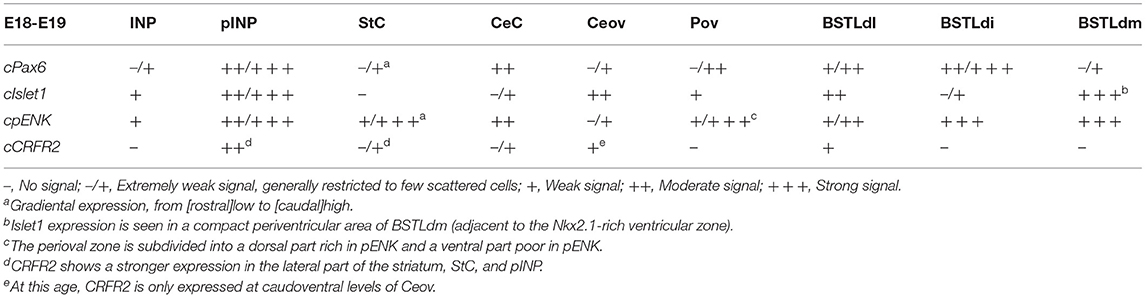
Table 3. Expression of several genes in the central extended amygdala and some surrounding areas of chicken at E18-E19.
Lastly, corrections have been made to Discussion, All Paragraphs. Any time we refer to our results on ‘CRF' in the Discussion section, this should be written as ‘cCRFR2' instead.
We sincerely apologize for the inconveniences created by this mistake. This correction only affects minimally the conclusions of the paper, which are mostly based on expression of the transcription factors Pax6, Islet1, and Nkx2.1, and further supported by the expression of pENK and SOM plus some connectivity patterns which are highly conserved between chicken and mouse. Moreover, in a previous study on the distribution of CRF immunoreaction in the chicken and quail brains, by Richard et al. (2004), CRF immunoreactive cells were found in some of the same subpallial areas where we described the CRFR2 (in particular, the striatum and the part of the extended amygdala identified by us as the oval central amygdalar nucleus or Ceov). The finding of CRFR2 expression in areas of the chicken brain comparable to the central amygdala is relevant in order to understand the function of CRF related systems and their evolution. In vertebrates, there are two receptors, which bind CRF and urocortin peptides with different affinities (Chang and Hsu, 2004). In mammals, both receptors show different expression patterns in the brain and outside the brain, and play different roles in stress, anxiety, and motivation (Chalmers et al., 1995; Steckler and Holsboer, 1999; Kishimoto et al., 2000; Henckens et al., 2016). Interestingly, the expression of CRFR2 in the extended amygdala differs between rodents (with variations between species) and non-human primates: in some rats, the expression is high the amygdalostriatal transition area and the medial bed nucleus of the stria terminalis, moderate in the medial amygdala, and low in the central amygdala (Chalmers et al., 1995; Van Pett et al., 2000; Coen et al., 2015); in contrast, in macaques, the expression is high in the central amygdala, moderate in the medial amygdala, and low in the bed nucleus of the stria terminalis (Sánchez et al., 1999). In view of the inter-species variations, it is necessary to carry out a thorough analysis of the expression of different CRF receptors and their ligands in the brain of more species of mammals and non-mammals, which will help to understand possible differences in the stress response between species.
The authors apologize for this error and state that this does not change the scientific conclusions of the article in any way. The original article has been updated.
References
Chalmers, D. T., Lovenberg, T. W., and De Souza, E. B. (1995). Localization of novel corticotropin-releasing factor receptor (CRF2) mRNA expression to specific subcortical nuclei in rat brain: comparison with CRF1 receptor mRNA expression. J. Neurosci. 15, 6340–6350. doi: 10.1523/JNEUROSCI.15-10-06340.1995
Chang, C. L., and Hsu, S. Y. T. (2004). Ancient evolution of stress-regulating peptides in vertebrates. Peptides 25, 1681–1688. doi: 10.1016/j.peptides.2004.05.022
Coen, C. W., Kalamatianos, T., Oosthuizen, M. K., Poorun, R., Faulkes, C. G., and Bennett, N.C. (2015). Sociality and the telencephalic distribution of corticotrophin-releasing factor, urocortin 3, and binding sites for CRF type 1 and type 2 receptors: a comparative study of eusocial naked mole-rats and solitary Cape mole-rats. J. Comp. Neurol. 523, 2344–2371. doi: 10.1002/cne.23796
Henckens, M. J., Deussing, J. M., and Chen, A. (2016). Region-specific roles of the corticotropin-releasing factor-urocortin system in stress. Nat. Rev. Neurosci. 17, 636–651. doi: 10.1038/nrn.2016.94
Kishimoto, T., Radulovic, J., Radulovic, M., Lin, C. R., Schrick, C., Hooshmand, F., et al. (2000). Deletion of crhr2 reveals an anxiolytic role for corticotropin-releasing hormone receptor-2. Nat. Genet. 24, 415–419. doi: 10.1038/74271
Richard, S., Martínez-García, F., Lanuza, E., and Davies, D. C. (2004). Distribution of corticotropin- releasing factor-immunoreactive neurons in the central nervous system of the domestic chicken and Japanese quail. J. Comp. Neurol. 469, 559–580. doi: 10.1002/cne.11023
Sánchez, M. M., Young, L. J., Plotsky, P. M., and Insel, T. R. (1999). Autoradiographic and in situ hybridization localization of corticotropin-releasing factor 1 and 2 receptors in nonhuman primate brain. J. Comp. Neurol. 408, 365–377.
Steckler, T., and Holsboer, F. (1999). Corticotropin-releasing hormone receptor subtypes and emotion. Biol. Psychiatry. 46, 1480–1508. doi: 10.1016/s0006-3223(99)00170-5
Keywords: Islet1, Pax6, enkephalin, corticotropin releasing factor, bed nucleus of the stria terminalis, fear responses, evolution
Citation: Vicario A, Abellán A, Desfilis E and Medina L (2021) Corrigendum: Genetic Identification of the Central Nucleus and Other Components of the Central Extended Amygdala in Chicken During Development. Front. Neuroanat. 15:671725. doi: 10.3389/fnana.2021.671725
Received: 24 February 2021; Accepted: 30 March 2021;
Published: 19 May 2021.
Edited by:
Alberto Munoz, Complutense University of Madrid, SpainReviewed by:
Fernando Martinez-Garcia, University of Jaume I, SpainCopyright © 2021 Vicario, Abellán, Desfilis and Medina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Loreta Medina, bG9yZXRhLm1lZGluYUB1ZGwuY2F0