 Wanrui Fu
Wanrui Fu Guzailinuer Kadeer
Guzailinuer Kadeer- Department of Nutrition, Huadong Hospital Affiliated to Fudan University, Shanghai, China
Background: Mitochondrial dysfunction is a significant contributor to sarcopenia, but the mechanism remains unclear.
Methods: In the present study, we downloaded GSE117525 and GSE8479 datasets from Gene Expression Omnibus (GEO), then the weighted correlation network analysis (WGCNA) was used to construct scale-free co-expression networks respectively. The key genes of aging muscle were obtained by overlapping key modules of two networks. Receiver operating characteristic (ROC) curve was drawn to explore the diagnostic efficacy of key genes. Finally, a transcription factor-key gene network was constructed based on ChEA3 platform and hTFtarget database, and a miRNA-key gene network was constructed using starBase and the multimiR R package.
Results: The most positively or negatively correlated modules of the two datasets were identified, and genes related to oxidative phosphorylation and mitochondrial ribosomal proteins were identified as key genes. The diagnostic values were confirmed with ROC curves by self-verification (GSE117525 and GSE8479) and external verification (GSE47881). Then, Yin Yang 1 (YY1) was identified as the most important transcription factor of the transcription factor-key gene network. In addition, miRNAs related to key genes were also predicted.
Conclusion: The findings of the present study provide a novel insight into the pathological mechanism of sarcopenia.
Introduction
Sarcopenia is defined as a progressive loss of skeletal muscle mass, strength, and function with advancing age (Papadopoulou, 2020). It occurs in approximately 5%–13% of older adults aged 60–70 years, and 11%–50% among those aged 80 or above (von Haehling et al., 2010). Meanwhile, the risk of weakness, falls, frailty, and fractures are increased in elderly individuals suffering from sarcopenia (He et al., 2021). Unfortunately, the etiology and pathogenesis of sarcopenia are still not fully elucidated, which reduces the efficacy and effectiveness of pharmacological interventions (Morley, 2016), and elucidation of the underlying pathological mechanisms may provide guidelines for sarcopenia prevention and therapy.
So far, a number of pathogenetic factors, such as mitochondrial dysfunction (Yang et al., 2021), inflammation (Schaap et al., 2009), denervation (Xu et al., 2021), and insulin resistance (González-Hedström et al., 2021) serve important roles in the development of sarcopenia. Among these mechanisms, it is believed that mitochondrial dysfunction plays a central role in the signaling networks of sarcopenia (Karam et al., 2017). As the metabolic hub for energy production, mitochondria provide adenosine triphosphate (ATP) for skeletal muscle contraction through oxidative phosphorylation (OXPHOS). However, in the process of cellular senescence, there are numerous changes in mitochondria, such as decreased mitochondrial membrane potential, increased proton leakage, and reactive oxygen species production (Amorim et al., 2022), which have been proven to have an important role in inflammatory responses (Marchi et al., 2022), insulin resistance (Hayden, 2022), as well as aging (Miwa et al., 2022). Mitochondrial dysfunction could lead to the onset and progression of sarcopenia, a better understanding of the regulation factors controlling mitochondrial gene expression level and driving mitochondrial dysfunction in sarcopenia is therefore needed.
For maintaining mitochondrial structure and function, normal gene expression encoding for mitochondrial proteins are required, which is controlled by transcriptional regulators, especially transcription factors (TFs) (Taylor and Bishop, 2022) and microRNAs (miRNAs or miRs) (Giordani et al., 2021). TFs act at the transcriptional level, while miRNAs act post-transcriptionally. TFs are proteins that bind to the upstream regulatory elements of genes (enhancers and silencers) to increase or decrease gene transcription and protein synthesis. Several nuclear transcription factors, such as cAMP response element-binding protein (CREB) (Lee et al., 2005) and signal transducer and activator of transcription 3 (STAT3) (Macias et al., 2014), have been identified to participate in the regulation of mitochondrial transcription. Additionally, miRNAs are short single-stranded noncoding RNAs with ∼22 nucleotides, and they can regulate the gene expression at the posttranscriptional level. Research has found that miRNAs have a role to play in affecting mitochondrial dynamics and bioenergetics and regulating the mitochondrial homeostasis, which is important for fine-tuning the muscle atrophy program (Soares et al., 2014; Cannataro et al., 2021). Therefore, elucidating the relationships between TFs and miRNAs as well as related networks in sarcopenia may provide new ideas for the development of clinical drug target and the prevention of disease.
With the development of gene chip technology, the weighted gene co-expression network analysis (WGCNA) method, a strong clustering algorithm, is widely used to detect gene co-expression modules. In this study, we employed WGCNA and other bioinformatic analyses for young vs. healthy older individuals based on Gene Expression Omnibus (GEO) database. Our results revealed that mitochondrial function genes coding for nicotinamide adenine dinucleotide (NADH) dehydrogenase subunits played an important role in skeletal muscle aging, and regulatory networks of TFs and miRNAs could be potential therapeutic targets in sarcopenia.
Materials and methods
Data collection and data preprocessing
Microarray data of GSE117525 and GSE8479 datasets were downloaded from GEO database (https://www.ncbi.nlm.nih.gov/geo/), and GPL20880 [HuGene-1_1-st] Affymetrix Human Gene 1.1 ST Array [HuGene11st_Hs_ENTREZG_18.0.0] and GPL2700 Sentrix HumanRef-8 Expression BeadChip platform was applied to analysis (Melov et al., 2007; Hangelbroek et al., 2016). GSE117525 dataset provided microarray data from vastus lateralis muscle biopsies of 53 young and 73 healthy older adults, and the GSE8479 dataset contained samples from the same position of 25 healthy older and 26 younger adults. In addition, the raw data were subjected to background correction and quantile data normalization using the Limma package in Bioconductor (http://bioconductor.org/packages/release/bioc/html/limma.html). The median absolute deviation (MAD) top 80% genes were selected for co-expression network construction.
Co-expression network construction
The scale-free co-expression network was constructed using the weighted gene co-expression network analysis (WGCNA) package in R (Langfelder and Horvath, 2008). First, the goodSamplesGenes method of R software package WGCNA was used to remove outlier genes and samples. Then, according to the SoftThreshold, the appropriate power was used to generate a scale-free topology overlap matrix (TOM), and the corresponding dissimilarity (1-TOM) was also calculated. Next, the linkage hierarchical clustering was established based on TOM with a minimum size of 100. The Dynamic Tree-Cut algorithm was used to classify similar genes into one gene module.
Identification of key modules and hub genes
First, module eigengenes (MEs) were the major components of a particular gene module, and the correlation between MEs and age was analyzed by Pearson Correlation Coefficient to distinguish the co-expression modules that presented high associations with skeletal muscle aging. Second, as a persuasive indicator for selecting a significant module, module significance (MS) was calculated according to the average value of gene significance (GS). Eventually, the co-expression modules having the highest associations with age were regarded as the key modules. The jVenn tool (Bardou et al., 2014) was adopted to identify the intersecting genes of key modules in GSE117525 and GSE8479 to obtain a set of hub genes.
Functional and pathway enrichment analysis
To further explore the biological significance of hub genes on the pathogenesis of skeletal muscle aging, we executed gene ontology (GO) functional enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis by clusterProfiler R package (http://www.bioconductor.org/packages/release/bioc/html/clusterProfiler.html). An adjusted p < 0.01 was considered to be significantly enriched.
Protein-protein interaction network analysis
Protein-protein interaction (PPI) information was accessed using the Search Tool for the Retrieval of Interacting Genes Database (STRING) (Szklarczyk et al., 2021) (https://www.string-db.org/), and a medium confidence score (≥0.4) was set as cutoff criteria. The results were converted visually by Cytoscape (version 3.7.1) software. The CytoHubba (version 0.1) and Molecular Complex Deletion (MCODE version 1.6.1) plug-ins for Cytoscape were employed to distinguish the top-ranked genes/nodes and modules with a cutoff MCODE score of >5.
Validation of key genes
Genes in the module sifted out by MCODE were regarded as key genes. Next, key genes self-verification (GSE117525 and GSE8479) and external verification (GSE47881) were evaluated by area under the curve (AUC) of the receiver operating characteristic (ROC) curve by the pROC R package. In the GSE47881 dataset, 44 baseline muscle samples were obtained from healthy adults aged 18 to 75.
Transcription factor enrichment analysis
ChIP-X Enrichment Analysis 3 (ChEA3) platform (Keenan et al., 2019) (https://maayanlab.cloud/chea3/) and hTFtarget database (http://bioinfo.life.hust.edu.cn/hTFtarget#!/target) were used for TF prediction of key genes respectively, and the common TFs and their targets were identified by the jVenn tool.
Construction of microRNAs-mRNA network
The online database starBase (Li et al., 2014) and the multimiR R package (Ru et al., 2014) were used to predict the miRNAs, and the miRNA-mRNA network was constructed according to the common results.
Results
Weighted gene co-expression network analysis
The WGCNA method was used to investigate the correlation between age and key genes. Before network construction, the outliers, GSM3302451 and GSM3302359 in GSE117525, as well as A49, A45, A2, and A20 in GSE8479, were removed (Supplementary Figure S1). The remaining samples were shown in Figures 1A,B. For the construction of the scale-free network, β = 7 and β = 6 were identified as the optimal soft-power value for GSE117525 and GSE8479 (Figures 1C,D). Then, we identified 13 modules of co-expressed genes in GSE117525 and 16 modules in GSE8479. Afterward, the correlations between modules and age were calculated. The green module had the strongest positive correlation with aging (r = 0.85), while the turquoise module had the strongest negative correlation (r = −0.64) in the GSE117525 database (Figure 1E). Meanwhile, the green module showed the strongest positive correlation (r = 0.77), and the black module had the strongest negative correlation (r = −0.89) in the GSE8479 database (Figure 1F). A set of hub genes were obtained by intersecting genes of key modules with the strongest relationship in GSE117525 and GSE8479 (Figures 1G,H).
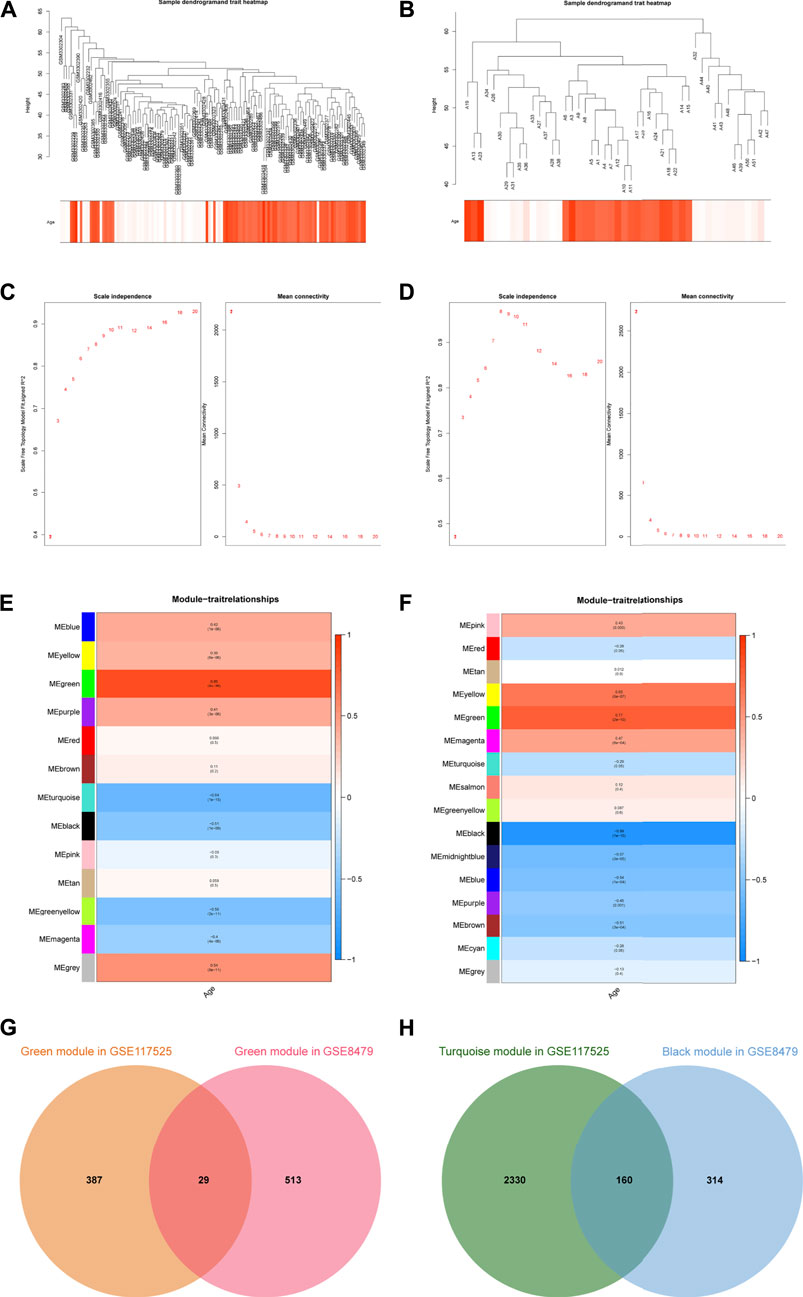
FIGURE 1. The key modules and hub genes identified by WGCNA. (A,B) Sample clustering after outliers were removed in GSE117525 (A) and GSE8479 (B). (C,D) The soft-threshold power selecting processes. The β = 7 in GSE117525 (C) and β = 6 in GSE8479 (D) were selected when R2 reached 0.85. (E,F) Heatmaps of the correlation between module eigengenes and age in GSE117525 (E) and GSE8479 (F). The corresponding correlations and p-values were presented. (G) The overlapped genes between the green modules. (H) The overlapped genes between the turquoise module and black module.
Identification of the shared pathways
Using the GO and KEGG enrichment analyses, we further explored the common regulatory pathways that 29 positively-regulating and 160 negatively-regulating genes screened by WGCNA respectively. The GO analysis showed that the positively-regulating genes might be related to the cell junction disassembly and positive regulation of cell-substrate junction organization (Figure 2B). However, there was no KEGG pathway enriched in positively-regulating genes because the number of genes was too small. As for the negatively-regulating genes, the GO analysis showed that biological processes (BP) were enriched in cellular respiration, ATP metabolic process, mitochondrial electron transport, NADH to ubiquinone as well as fatty acid metabolic process, and cellular composition (CC) terms were enriched in mitochondrial matrix and NADH dehydrogenase complex, and molecular functions (MF) were enriched in electron transfer activity, and NADH dehydrogenase activity (Figure 2A). The KEGG analysis showed that these genes might be correlated with carbon metabolism, citrate cycle (TCA cycle), OXPHOS, and chemical carcinogenesis-reactive oxygen species (ROS) (Figure 2C).
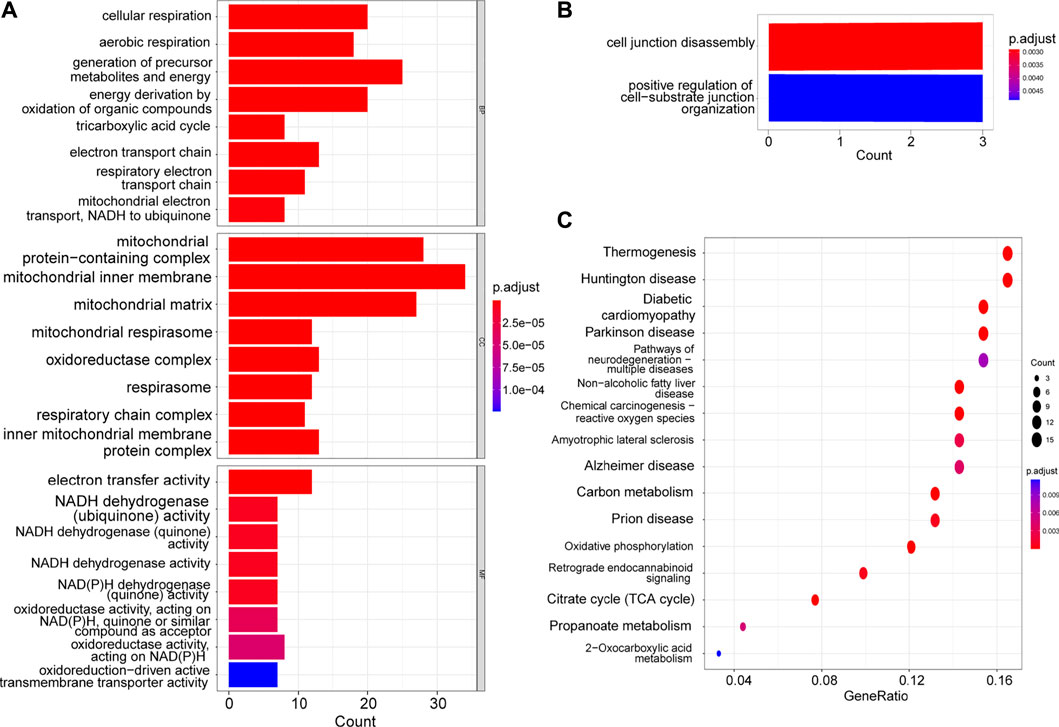
FIGURE 2. GO and KEGG enrichment of hub genes. (A) The GO analysis of the negatively-regulating genes. (B) The GO analysis of the positively-regulating genes. (C) The KEGG analysis of the negatively-regulating genes.
Construction and analysis of the protein-protein interaction network
To explore the interactions among negatively-regulating genes in depth, we constructed a PPI network using the STRING online database and Cytoscape software (Figure 3A). The core PPI network consisted of 110 nodes and 349 edges with a medium confidence score. Using Cytoscape MCODE, we found a significant module (score = 12.5) from the PPI network complex, and the degree algorithm in CytoHubba also showed genes in the module had top degrees (Figure 3B). Therefore, genes in this module (Supplementary Table S1) were regarded as key genes.
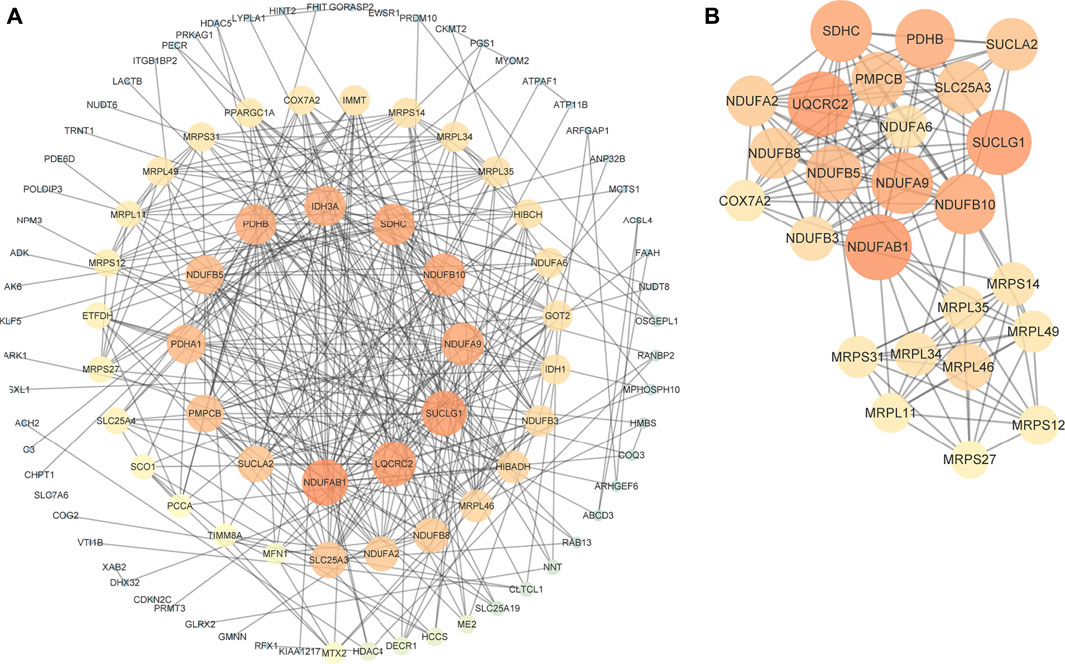
FIGURE 3. PPI network of negatively-regulating hub genes. (A) The core PPI network with a medium confidence score. (B) A significant module screened by MCODE. Different colors and sizes of nodes represent different degrees. The larger and brighter the node becomes, the greater the degree is.
Validation of key genes
Then, the diagnostic values of key genes were confirmed by ROC curve analysis and the AUC value. As shown in Table 1 and Supplementary Figures S2–S4, the AUC of 10 key genes was greater than 0.75 among three datasets, and the maximum AUC was 0.9646 for NADH:ubiquinone oxidoreductase subunit B8 (NDUFB8) in GSE8479. The results indicated that the key genes could play a good accuracy in the early diagnosis of sarcopenia.
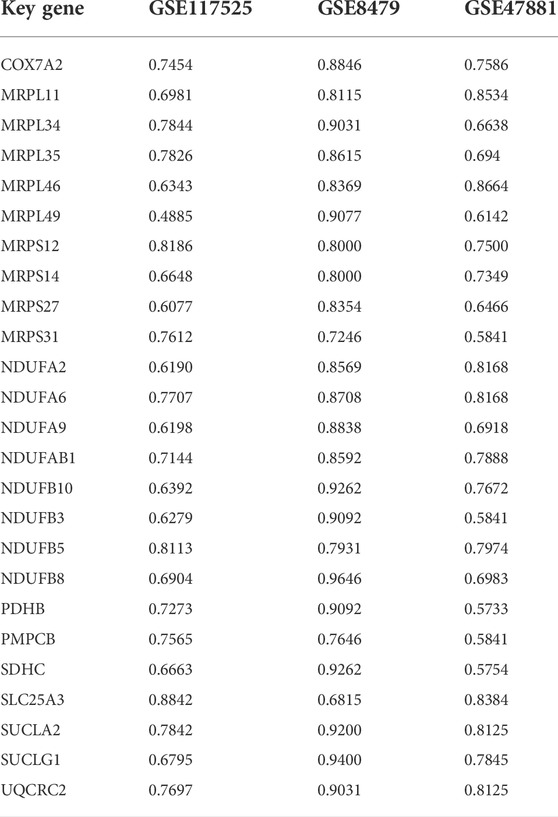
TABLE 1. The AUC value of key genes in the three datasets.
Construction of the transcription factor-key gene network
We used ChEA3 platform and hTFtarget database to predict the key transcription factor of key genes. As a result, there were 75 intersecting transcription factors enriched (Supplementary Table S2). As shown in Figure 4, twenty genes in all 25 key genes were predicted to be regulated by Yin Yang 1 (YY1).
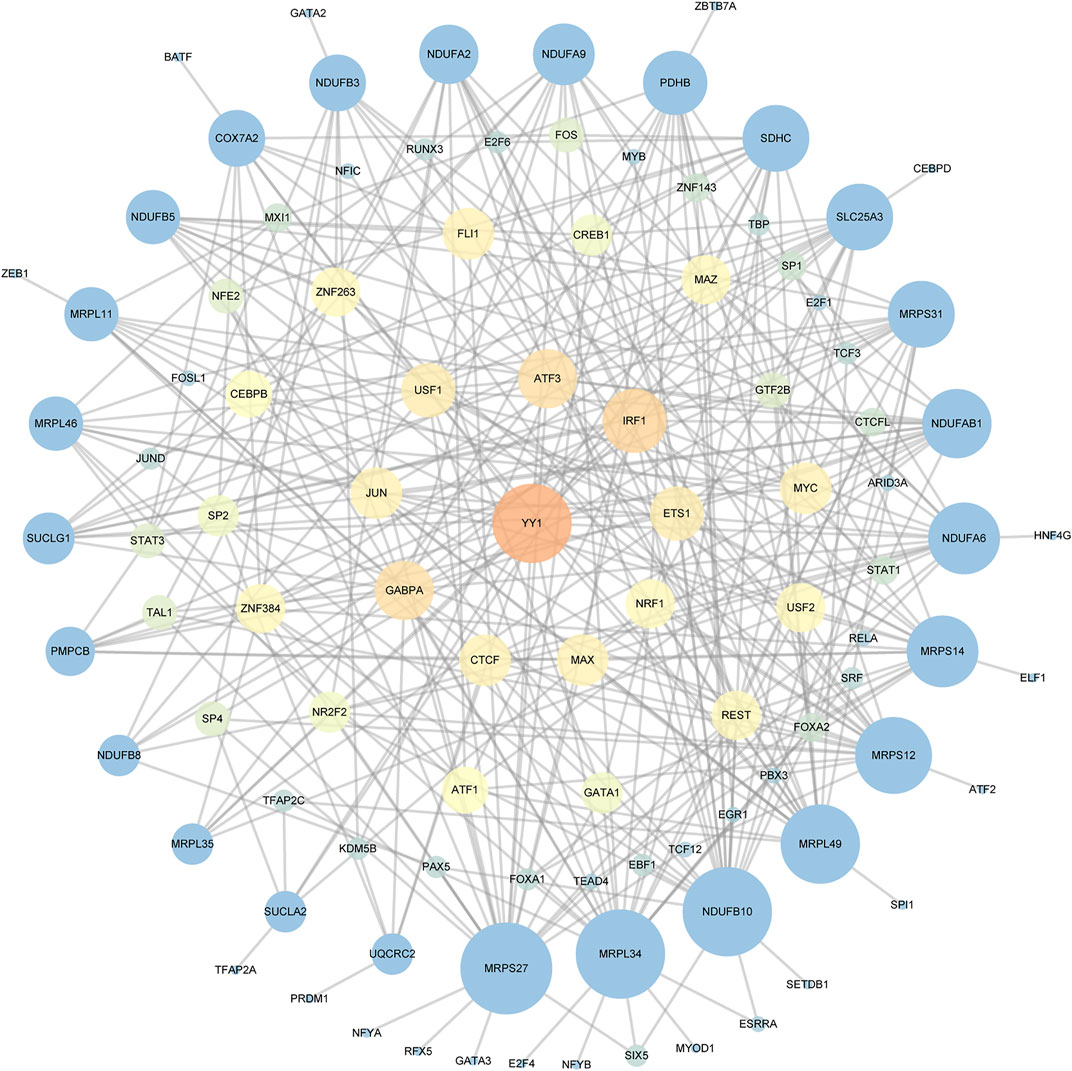
FIGURE 4. The TF-key Gene Network. Different colors and sizes of nodes represent different degrees. The larger and brighter the node becomes, the greater the degree is.
Construction of the microRNA-key gene network
The miRNAs of key genes were identified using the online database starBase and the multimiR R package respectively, and 493 miRNAs were identified as candidates by starBase, as well as 583 miRNAs were identified by multimiR. To increase the accuracy, we selected the intersection of the results of each method (Supplementary Table S3). It should be noted that there was no corresponding miRNA for mitochondrial ribosomal protein S31 (MRPS31), NADH:ubiquinone oxidoreductase subunit A9 (NDUFA9), NDUFB8, and peptidase, mitochondrial processing subunit beta (PMPCB) after intersection. In the end, one hundred and thirty-nine miRNAs in total were obtained and the miRNA-key Gene Network was constructed as shown in Figure 5. In addition, the flowchart shows all essential and significant procedures in our study (Figure 6), and the abbreviations are provided in the supplementary abbreviation list (Supplementary Table S4).
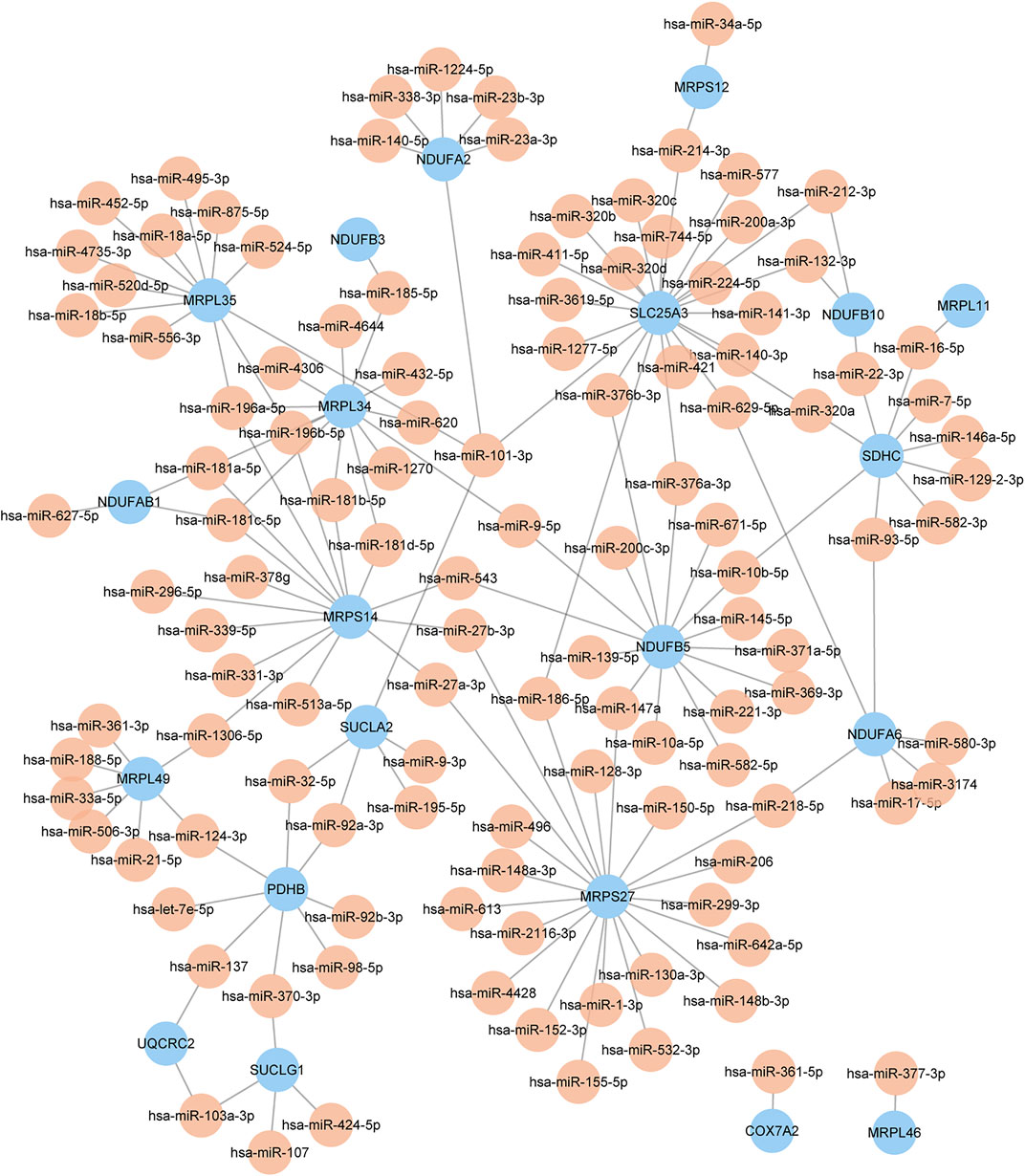
FIGURE 5. The miRNA-key gene network. Different colors of nodes represent different genes or miRNAs. The color yellow represents miRNA, and the color blue represents key genes.
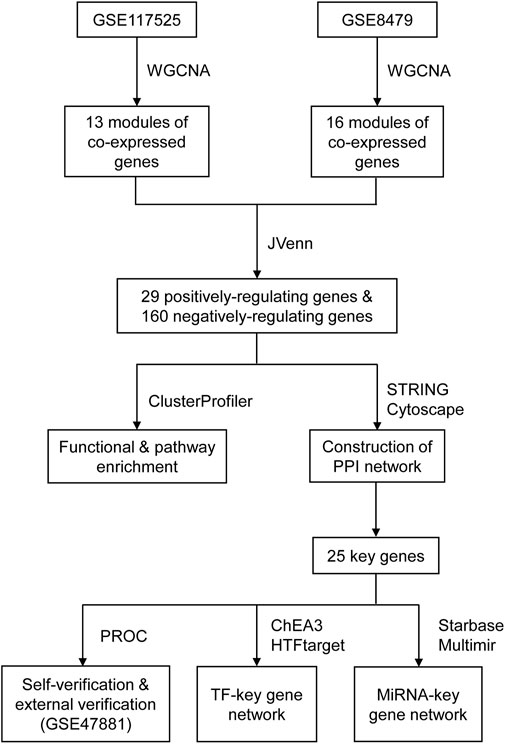
FIGURE 6. The flowchart of the analysis.
Discussion
Sarcopenia is very common in older adults. It is characterized by a progressive and generalized degenerative loss of skeletal muscle mass, quality, and strength with normal aging (Wiedmer et al., 2021). The pathogenesis of sarcopenia is complex, and it appears to be difficult the identification of novel specific molecular markers that can be accurately and effectively used in the diagnosis, treatment, or prognosis evaluation of sarcopenia. With the development of omics techniques and bioinformatics technology, new opportunities are offered to identify new targets to help us understand the pathophysiology of sarcopenia.
In this study, we constructed WGCNA to identify co-expression genes and possible biological pathways of sarcopenia based on the GSE117525 and GSE8479 datasets. The most positively and negatively correlated modules with aging were green and turquoise module in GSE117525 and green and black modules in GSE8479. The function analysis revealed that cellular respiration-related biological processes and pathways, including NADH to ubiquinone, thermogenesis, TCA cycle, and OXPHOS, were significantly involved in the overlapped genes between the most negatively correlated modules of the two datasets. Then, genes related to OXPHOS and mitochondrial ribosomal proteins were identified as key genes. YY1 along with the other 75 transcription factors was likely to play an important role in regulating the expression of key genes. In addition, as important regulators in the post-transcriptional regulation of gene expression, miRNAs in connection with key genes were predicted and the relevant miRNA-key gene network may help to explore the pathological mechanism of sarcopenia.
Our GO/KEGG results demonstrated that mitochondrial dysfunction played a significant role in the pathological process of sarcopenia. With advancing age, the mitochondrial capacity to produce energy decreases in skeletal muscle, particularly when maximal performance is required (Gonzalez-Freire et al., 2018). NAD decline has been recognized as one possible mechanism. As a coenzyme for redox reactions, NAD is at the center of energy metabolism, because NAD along with NADH participates in the TCA cycle and the electron-transporter channel, and an optimal NAD/NADH ratio is needed for efficient mitochondrial metabolism (Navas and Carnero, 2021). The decline in NAD levels is linked to numerous aging-associated diseases, including cognitive decline, metabolic disease, sarcopenia, and frailty (Covarrubias et al., 2021). Research has found that mitochondrial oxidative capacity and NAD biosynthesis are reduced in human sarcopenia across ethnicities (Migliavacca et al., 2019), and reducing NAD levels by ablating nicotinamide phosphoribosyltransferase (NAMPT)-mediated NAD salvage could lead to progressive muscle degeneration in the adult mouse (Frederick et al., 2016). Moreover, raising NAD (+) levels in old mice restores mitochondrial function to that of a young mouse in a sirtuin-1 (SIRT1)-dependent manner (Gomes et al., 2013). Therefore, targeting NAD metabolism has emerged as a potential therapeutic approach to ameliorate sarcopenia, and extend the human health-span and lifespan.
As the first and largest complex of the mitochondrial respiratory chain, NADH-ubiquinone oxidoreductase, or complex I, plays an important role in mitochondrial ATP synthesis. It could drive proton translocation across the inner mitochondrial membrane by liberating and transferring electrons from NADH to ubiquinone. Interestingly, some members of the NADH-ubiquinone oxidoreductase (NDUF) family were identified as key genes in our study, including NADH: ubiquinone oxidoreductase subunit A2 (NDUFA2), NADH: ubiquinone oxidoreductase subunit A6 (NDUFA6), NDUFA9, NADH: ubiquinone oxidoreductase subunit AB1 (NDUFAB1), NADH: ubiquinone oxidoreductase subunit B10 (NDUFB10), NADH: ubiquinone oxidoreductase subunit B3 (NDUFB3), NADH: ubiquinone oxidoreductase subunit B5 (NDUFB5), and NDUFB8, making this gene family into an important candidate for future study and biomarker development. Mutations in these genes could lead to complex I deficiency, which is one of the most frequent defects of the OXPHOS system. Complex I deficiency could result in the impairment of organs with a high-energy demand and participate in the pathogenesis of cancer, aging, and neurodegenerative conditions (Fiedorczuk and Sazanov, 2018). A study found that NDUFAB1 is a crucial regulator of mitochondrial energy and ROS metabolism as it could effectively enhance mitochondrial bioenergetics while limiting ROS production (Hou et al., 2019). In addition, the aging-associated reduction in expression of NDUFA6, NDUFA9, NDUFB5, NDUFB8, and NADH: ubiquinone oxidoreductase core subunit S2 (NDUFS2) was correlated with the decline in activity of OXPHOS in the senescent heart (Emelyanova et al., 2018). In conclusion, our findings indicated that genes coding for complex I were essential for a normal OXPHOS process, and they could be novel mitochondrial targets to prevent sarcopenia.
Apart from the NDUF family, many mitochondrial ribosomal proteins (MRPs) were also identified as key genes in this study. Mitochondrial ribosomes (mitoribosomes) consist of a small 28S subunit encoded by 30 MRP genes, and a large 39S subunit encoded by 52 MRP genes in human. They are implicated in the synthesis of all 13 mitochondrial DNA (mtDNA) -encoded protein subunits of the human OXPHOS system. Dysfunction of these proteins not only impairs mitochondrial protein translation but also causes primary mitochondrial respiratory chain activity deficiencies and clinical diseases, such as neurodegenerative diseases (Lopez Sanchez et al., 2021), inflammatory disorders (Gopisetty and Thangarajan, 2016) as well as aging (Molenaars et al., 2020). A study has found that MRPL49 shows altered proteolytic processing by dopamine treatment in Parkinson’s disease (Lualdi et al., 2019). In addition, decreased expression of MRPL12 could reduce mitochondrial OXPHOS in proximal tubular epithelial cells (Gu et al., 2021). In a word, MRPs are indispensable for respiration, but their biological functions are not yet clear and further research is needed.
In the present study, we constructed a TF-mRNA network to understand the potential mechanisms of sarcopenia. As a result, YY1 was screened out as the most important TF. When YY1 was deleted in skeletal muscle satellite cells, the acute damage-induced muscle repair was blocked, resulting in muscular dystrophy (Chen et al., 2019). And in another study, the muscle-specific knockout of Yy1 could lead to defective mitochondrial morphology and oxidative function (Blättler et al., 2012). Mechanistically, loss of YY1 or its repressive acetylation at K392/K393 may affect energy metabolism as mitochondrial complex I genes are bound directly by YY1 (Perekatt et al., 2014; Addicks et al., 2022). Notably, a study has found that YY1 is significantly upregulated in senior groups (Zampieri et al., 2015), and YY1 has been regarded as both a transcriptional repressor and activator (Kawamura et al., 2021). In the proliferative stage, YY1 is likely to inhibit the function of mitochondria, but in the differentiation stage, it switches to firing up mitochondria and OXPHOS (Blättler et al., 2012). The function of YY1 remains unclear, but the two-way regulation of YY1 on mitochondria during different periods may be an important mechanism for regulating the development and aging of muscle. The other TFs, such as interferon regulatory factor 1 (IRF1) (Baron et al., 2011), activating transcription factor 3 (ATF3) (Lim et al., 2006), CREB1 (Franko et al., 2008), also has been proved to participate in regulating the expression of genes encoding mitochondrial proteins during myogenesis and mitochondrial homeostasis.
MiRNAs are gene regulators that post-transcriptionally regulate specific genes. A growing stream of research suggests that miRNAs have a big role in skeletal muscle physiology by targeting and preventing the translation of specific mRNAs. Research has found that miR-127 and miR-434-3p were generally downregulated in most aged mouse muscles (Lee and Kang, 2022). Additionally, miR-101 was associated with frailty syndrome (Carini et al., 2022) and Alzheimer’s disease (Barbato et al., 2020). A study has found that miR-101a/b could suppress the p38 mitogen activated protein kinase (MAPK), interferon-gamma (IFN-gamma), and Wnt pathways and enhance the CCAAT/enhancer binding protein (C/EBP) pathway, which led to the inhibition of myoblast differentiation (Liu et al., 2021). Moreover, miR-101-3p has a regulative effect on mtDNA transcription and the assembly of the succinate dehydrogenase complex subunit C (SDHC) subunit, which is important for complex II biogenesis (Ziemann et al., 2022). In conclusion, miRNAs play vital roles in the regulation of OXPHOS and mitochondrial metabolism, providing new insights into deciphering the key molecular pathways related to sarcopenia and age-associated disorders.
Finally, the present study had several limitations that should be acknowledged: 1) Our study is limited to the processing of previous data, requiring in vivo and in vitro experiments to verify the results. 2) Due to the small number of overlapping positively-regulating genes, we could not identify KEGG pathways and key genes of them.
Conclusion
In this study, genes related to OXPHOS and MRPs were regarded as the ones that had the strongest association with skeletal muscle aging. The relevant TFs and miRNAs were also identified and they could participate in the pathogenesis and treatment of sarcopenia by regulating the expression of mitochondria-related genes. In conclusion, our results will be helpful for better understanding the role of mitochondria in the pathophysiology of sarcopenia. Further mechanistic studies and more evidence are needed to validate these results and prove the mechanism.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
WF contributed to data acquisition and article drafting; GK and YH provided technical support; YF contributed to study design and supervision. All authors approved the manuscript.
Funding
This research was funded by the Shanghai Municipal Health System Important and Weak Discipline (Clinical Nutrition) Project (No. 2019ZB0102).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2022.975886/full#supplementary-material
References
Addicks, G. C., Zhang, H., Ryu, D., Vasam, G., Green, A. E., Marshall, P. L., et al. (2022). GCN5 maintains muscle integrity by acetylating YY1 to promote dystrophin expression. J. Cell. Biol. 221 (2), e202104022. doi:10.1083/jcb.202104022
Amorim, J. A., Coppotelli, G., Rolo, A. P., Palmeira, C. M., Ross, J. M., and Sinclair, D. A. (2022). Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat. Rev. Endocrinol. 18 (4), 243–258. doi:10.1038/s41574-021-00626-7
Barbato, C., Giacovazzo, G., Albiero, F., Scardigli, R., Scopa, C., Ciotti, M. T., et al. (2020). Cognitive decline and modulation of Alzheimer's disease-related genes after inhibition of MicroRNA-101 in mouse hippocampal neurons. Mol. Neurobiol. 57 (7), 3183–3194. doi:10.1007/s12035-020-01957-8
Bardou, P., Mariette, J., Escudié, F., Djemiel, C., and Klopp, C. (2014). jvenn: an interactive Venn diagram viewer. BMC Bioinforma. 15, 293. doi:10.1186/1471-2105-15-293
Baron, D., Magot, A., Ramstein, G., Steenman, M., Fayet, G., Chevalier, C., et al. (2011). Immune response and mitochondrial metabolism are commonly deregulated in DMD and aging skeletal muscle. PloS One 6 (11), e26952. doi:10.1371/journal.pone.0026952
Blättler, S. M., Verdeguer, F., Liesa, M., Cunningham, J. T., Vogel, R. O., Chim, H., et al. (2012). Defective mitochondrial morphology and bioenergetic function in mice lacking the transcription factor Yin Yang 1 in skeletal muscle. Mol. Cell. Biol. 32 (16), 3333–3346. doi:10.1128/MCB.00337-12
Cannataro, R., Carbone, L., Petro, J. L., Cione, E., Vargas, S., Angulo, H., et al. (2021). Sarcopenia: Etiology, nutritional approaches, and miRNAs. Int. J. Mol. Sci. 22 (18), 9724. doi:10.3390/ijms22189724
Carini, G., Mingardi, J., Bolzetta, F., Cester, A., Bolner, A., Nordera, G., et al. (2022). miRNome profiling detects miR-101-3p and miR-142-5p as putative blood biomarkers of frailty syndrome. Genes. 13 (2), 231. doi:10.3390/genes13020231
Chen, F., Zhou, J., Li, Y., Zhao, Y., Yuan, J., Cao, Y., et al. (2019). YY1 regulates skeletal muscle regeneration through controlling metabolic reprogramming of satellite cells. EMBO J. 38 (10), e99727. doi:10.15252/embj.201899727
Covarrubias, A. J., Perrone, R., Grozio, A., and Verdin, E. (2021). NAD+ metabolism and its roles in cellular processes during ageing. Nat. Rev. Mol. Cell. Biol. 22 (2), 119–141. doi:10.1038/s41580-020-00313-x
Emelyanova, L., Preston, C., Gupta, A., Viqar, M., Negmadjanov, U., Edwards, S., et al. (2018). Effect of aging on mitochondrial energetics in the human atria. J. Gerontol. A Biol. Sci. Med. Sci. 73 (5), 608–616. doi:10.1093/gerona/glx160
Fiedorczuk, K., and Sazanov, L. A. (2018). Mammalian mitochondrial complex I structure and disease-causing mutations. Trends Cell. Biol. 28 (10), 835–867. doi:10.1016/j.tcb.2018.06.006
Franko, A., Mayer, S., Thiel, G., Mercy, L., Arnould, T., Hornig-Do, H.-T., et al. (2008). CREB-1alpha is recruited to and mediates upregulation of the cytochrome c promoter during enhanced mitochondrial biogenesis accompanying skeletal muscle differentiation. Mol. Cell. Biol. 28 (7), 2446–2459. doi:10.1128/MCB.00980-07
Frederick, D. W., Loro, E., Liu, L., Davila, A., Chellappa, K., Silverman, I. M., et al. (2016). Loss of NAD homeostasis leads to progressive and reversible degeneration of skeletal muscle. Cell. Metab. 24 (2), 269–282. doi:10.1016/j.cmet.2016.07.005
Giordani, C., Silvestrini, A., Giuliani, A., Olivieri, F., and Rippo, M. R. (2021). MicroRNAs as factors in bidirectional crosstalk between mitochondria and the nucleus during cellular senescence. Front. Physiol. 12, 734976. doi:10.3389/fphys.2021.734976
Gomes, A. P., Price, N. L., Ling, A. J. Y., Moslehi, J. J., Montgomery, M. K., Rajman, L., et al. (2013). Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 155 (7), 1624–1638. doi:10.1016/j.cell.2013.11.037
Gonzalez-Freire, M., Scalzo, P., D'Agostino, J., Moore, Z. A., Diaz-Ruiz, A., Fabbri, E., et al. (2018). Skeletal muscle ex vivo mitochondrial respiration parallels decline in vivo oxidative capacity, cardiorespiratory fitness, and muscle strength: The Baltimore Longitudinal Study of Aging. Aging Cell. 17 (2), e12725. doi:10.1111/acel.12725
González-Hedström, D., Priego, T., Amor, S., de la Fuente-Fernández, M., Martín, A. I., López-Calderón, A., et al. (2021). Olive leaf extract supplementation to old wistar rats attenuates aging-induced sarcopenia and increases insulin sensitivity in adipose tissue and skeletal muscle. Antioxidants (Basel, Switz. 10 (5), 737. doi:10.3390/antiox10050737
Gopisetty, G., and Thangarajan, R. (2016). Mammalian mitochondrial ribosomal small subunit (MRPS) genes: A putative role in human disease. Gene 589 (1), 27–35. doi:10.1016/j.gene.2016.05.008
Gu, X., Liu, Y., Wang, N., Zhen, J., Zhang, B., Hou, S., et al. (2021). Transcription of MRPL12 regulated by Nrf2 contributes to the mitochondrial dysfunction in diabetic kidney disease. Free Radic. Biol. Med. 164, 329–340. doi:10.1016/j.freeradbiomed.2021.01.004
Hangelbroek, R. W. J., Fazelzadeh, P., Tieland, M., Boekschoten, M. V., Hooiveld, G. J. E. J., van Duynhoven, J. P. M., et al. (2016). Expression of protocadherin gamma in skeletal muscle tissue is associated with age and muscle weakness. J. Cachexia Sarcopenia Muscle 7 (5), 604–614. doi:10.1002/jcsm.12099
Hayden, M. R. (2022). The mighty mitochondria are unifying organelles and metabolic hubs in multiple organs of obesity, insulin resistance, metabolic syndrome, and type 2 diabetes: An observational ultrastructure study. Int. J. Mol. Sci. 23 (9), 4820. doi:10.3390/ijms23094820
He, Y., Xie, W., Li, H., Jin, H., Zhang, Y., and Li, Y. (2021). Cellular senescence in sarcopenia: Possible mechanisms and therapeutic potential. Front. Cell. Dev. Biol. 9, 793088. doi:10.3389/fcell.2021.793088
Hou, T., Zhang, R., Jian, C., Ding, W., Wang, Y., Ling, S., et al. (2019). NDUFAB1 confers cardio-protection by enhancing mitochondrial bioenergetics through coordination of respiratory complex and supercomplex assembly. Cell. Res. 29 (9), 754–766. doi:10.1038/s41422-019-0208-x
Karam, C., Yi, J., Xiao, Y., Dhakal, K., Zhang, L., Li, X., et al. (2017). Absence of physiological Ca2+ transients is an initial trigger for mitochondrial dysfunction in skeletal muscle following denervation. Skelet. Muscle 7 (1), 6. doi:10.1186/s13395-017-0123-0
Kawamura, K., Higuchi, T., and Fujiwara, S. (2021). YAF2-Mediated YY1-sirtuin6 interactions responsible for mitochondrial downregulation in aging tunicates. Mol. Cell. Biol. 41 (7), e0004721. doi:10.1128/MCB.00047-21
Keenan, A. B., Torre, D., Lachmann, A., Leong, A. K., Wojciechowicz, M. L., Utti, V., et al. (2019). ChEA3: Transcription factor enrichment analysis by orthogonal omics integration. Nucleic Acids Res. 47, W212–W224. doi:10.1093/nar/gkz446
Langfelder, P., and Horvath, S. (2008). Wgcna: an R package for weighted correlation network analysis. BMC Bioinforma. 9, 559. doi:10.1186/1471-2105-9-559
Lee, J., and Kang, H. (2022). Role of MicroRNAs and long non-coding RNAs in sarcopenia. Cells 11 (2), 187. doi:10.3390/cells11020187
Lee, J., Kim, C.-H., Simon, D. K., Aminova, L. R., Andreyev, A. Y., Kushnareva, Y. E., et al. (2005). Mitochondrial cyclic AMP response element-binding protein (CREB) mediates mitochondrial gene expression and neuronal survival. J. Biol. Chem. 280 (49), 40398–40401. doi:10.1074/jbc.C500140200
Li, J.-H., Liu, S., Zhou, H., Qu, L.-H., and Yang, J.-H. (2014). starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 42, D92–D97. doi:10.1093/nar/gkt1248
Lim, J. H., Lee, J. I., Suh, Y. H., Kim, W., Song, J. H., and Jung, M. H. (2006). Mitochondrial dysfunction induces aberrant insulin signalling and glucose utilisation in murine C2C12 myotube cells. Diabetologia 49 (8), 1924–1936. doi:10.1007/s00125-006-0278-4
Liu, S., Xie, S., Chen, H., Li, B., Chen, Z., Tan, Y., et al. (2021). The functional analysis of transiently upregulated miR-101 suggests a "braking" regulatory mechanism during myogenesis. Sci. China. Life Sci. 64 (10), 1612–1623. doi:10.1007/s11427-020-1856-5
Lopez Sanchez, M. I. G., Krüger, A., Shiriaev, D. I., Liu, Y., and Rorbach, J. (2021). Human mitoribosome biogenesis and its emerging links to disease. Int. J. Mol. Sci. 22 (8), 3827. doi:10.3390/ijms22083827
Lualdi, M., Ronci, M., Zilocchi, M., Corno, F., Turilli, E. S., Sponchiado, M., et al. (2019). Exploring the mitochondrial degradome by the TAILS proteomics approach in a cellular model of Parkinson's disease. Front. Aging Neurosci. 11, 195. doi:10.3389/fnagi.2019.00195
Macias, E., Rao, D., Carbajal, S., Kiguchi, K., and DiGiovanni, J. (2014). Stat3 binds to mtDNA and regulates mitochondrial gene expression in keratinocytes. J. Invest. Dermatol. 134 (7), 1971–1980. doi:10.1038/jid.2014.68
Marchi, S., Guilbaud, E., Tait, S. W. G., Yamazaki, T., and Galluzzi, L. (2022). Mitochondrial control of inflammation. Nat. Rev. Immunol., 25 1–15. doi:10.1038/s41577-022-00760-x
Melov, S., Tarnopolsky, M. A., Beckman, K., Felkey, K., and Hubbard, A. (2007). Resistance exercise reverses aging in human skeletal muscle. PloS One 2 (5), e465. doi:10.1371/journal.pone.0000465
Migliavacca, E., Tay, S. K. H., Patel, H. P., Sonntag, T., Civiletto, G., McFarlane, C., et al. (2019). Mitochondrial oxidative capacity and NAD+ biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 10 (1), 5808. doi:10.1038/s41467-019-13694-1
Miwa, S., Kashyap, S., Chini, E., and von Zglinicki, T. (2022). Mitochondrial dysfunction in cell senescence and aging. J. Clin. Invest. 132 (13), e158447. doi:10.1172/JCI158447
Molenaars, M., Janssens, G. E., Williams, E. G., Jongejan, A., Lan, J., Rabot, S., et al. (2020). A conserved mito-cytosolic translational balance links two longevity pathways. Cell. Metab. 31 (3), 549–563. doi:10.1016/j.cmet.2020.01.011
Morley, J. E. (2016). Pharmacologic options for the treatment of sarcopenia. Calcif. Tissue Int. 98 (4), 319–333. doi:10.1007/s00223-015-0022-5
Navas, L. E., and Carnero, A. (2021). NAD+ metabolism, stemness, the immune response, and cancer. Signal Transduct. Target. Ther. 6 (1), 2. doi:10.1038/s41392-020-00354-w
Papadopoulou, S. K. (2020). Sarcopenia: A contemporary health problem among older adult populations. Nutrients 12 (5), E1293. doi:10.3390/nu12051293
Perekatt, A. O., Valdez, M. J., Davila, M., Hoffman, A., Bonder, E. M., Gao, N., et al. (2014). YY1 is indispensable for Lgr5+ intestinal stem cell renewal. Proc. Natl. Acad. Sci. U. S. A. 111 (21), 7695–7700. doi:10.1073/pnas.1400128111
Ru, Y., Kechris, K. J., Tabakoff, B., Hoffman, P., Radcliffe, R. A., Bowler, R., et al. (2014). The multiMiR R package and database: Integration of microRNA-target interactions along with their disease and drug associations. Nucleic Acids Res. 42 (17), e133. doi:10.1093/nar/gku631
Schaap, L. A., Pluijm, S. M. F., Deeg, D. J. H., Harris, T. B., Kritchevsky, S. B., Newman, A. B., et al. (2009). Higher inflammatory marker levels in older persons: Associations with 5-year change in muscle mass and muscle strength. J. Gerontol. A Biol. Sci. Med. Sci. 64 (11), 1183–1189. doi:10.1093/gerona/glp097
Soares, R. J., Cagnin, S., Chemello, F., Silvestrin, M., Musaro, A., De Pitta, C., et al. (2014). Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions. J. Biol. Chem. 289 (32), 21909–21925. doi:10.1074/jbc.M114.561845
Szklarczyk, D., Gable, A. L., Nastou, K. C., Lyon, D., Kirsch, R., Pyysalo, S., et al. (2021). The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 49 (D1), D605–D612. doi:10.1093/nar/gkaa1074
Taylor, D. F., and Bishop, D. J. (2022). Transcription factor movement and exercise-induced mitochondrial biogenesis in human skeletal muscle: Current knowledge and future perspectives. Int. J. Mol. Sci. 23 (3), 1517. doi:10.3390/ijms23031517
von Haehling, S., Morley, J. E., and Anker, S. D. (2010). An overview of sarcopenia: Facts and numbers on prevalence and clinical impact. J. Cachexia Sarcopenia Muscle 1 (2), 129–133. doi:10.1007/s13539-010-0014-2
Wiedmer, P., Jung, T., Castro, J. P., Pomatto, L. C. D., Sun, P. Y., Davies, K. J. A., et al. (2021). Sarcopenia - molecular mechanisms and open questions. Ageing Res. Rev. 65, 101200. doi:10.1016/j.arr.2020.101200
Xu, H., Ranjit, R., Richardson, A., and Van Remmen, H. (2021). Muscle mitochondrial catalase expression prevents neuromuscular junction disruption, atrophy, and weakness in a mouse model of accelerated sarcopenia. J. Cachexia Sarcopenia Muscle 12 (6), 1582–1596. doi:10.1002/jcsm.12768
Yang, Y.-F., Yang, W., Liao, Z.-Y., Wu, Y.-X., Fan, Z., Guo, A., et al. (2021). MICU3 regulates mitochondrial Ca2+-dependent antioxidant response in skeletal muscle aging. Cell. Death Dis. 12 (12), 1115. doi:10.1038/s41419-021-04400-5
Zampieri, S., Pietrangelo, L., Loefler, S., Fruhmann, H., Vogelauer, M., Burggraf, S., et al. (2015). Lifelong physical exercise delays age-associated skeletal muscle decline. J. Gerontol. A Biol. Sci. Med. Sci. 70 (2), 163–173. doi:10.1093/gerona/glu006
Keywords: sarcopenia, mitochondria, transcription factor, miRNA, WGCNA
Citation: Fu W, Kadeer G, He Y and Feng Y (2022) The regulatory network of potential transcription factors and MiRNAs of mitochondria-related genes for sarcopenia. Front. Genet. 13:975886. doi: 10.3389/fgene.2022.975886
Received: 26 July 2022; Accepted: 22 August 2022;
Published: 12 September 2022.
Edited by:
Giuseppe Passarino, University of Calabria, ItalyReviewed by:
Erika Cione, University of Calabria, ItalyPaolina Crocco, University of Calabria, Italy
Copyright © 2022 Fu, Kadeer, He and Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ying Feng, yingfenghd@hotmail.com