 Pamela S. Douglas
Pamela S. Douglas- 1Transforming Maternity Care Collaborative, Griffith University, Brisbane, QLD, Australia
- 2Discipline of General Practice, The University of Queensland, Brisbane, QLD, Australia
Autism spectrum disorders (ASD) are an emergent public health problem, placing significant burden upon the individual, family and health system. ASD are polygenetic spectrum disorders of neural connectome development, in which one or more feedback loops amplify small genetic, structural, or functional variations in the very early development of motor and sensory-motor pathways. These perturbations trigger a ‘butterfly effect’ of unpredictable cascades of structural and functional imbalances in the global neuronal workspace, resulting in atypical behaviors, social communication, and cognition long-term. The first 100 days post-term are critically neuroplastic and comprise an injury-sensitive developmental window, characterized by a neural biomarker, the persistence of the cortical subplate, and a behavioral biomarker, the crying diathesis. By the time potential diagnostic signs are identified, from 6 months of age, ASD neuropathy is already entrenched. The International Society for Autism Research Special Interest Group has called for pre-emptive intervention, based upon rigorous theoretical frames, and real world translation and evaluation. This paper responds to that call. It synthesizes heterogenous evidence concerning ASD etiologies from both psychosocial and biological research literatures with complexity science and evolutionary biology, to propose a theoretical framework for pre-emptive intervention. This paper hypothesizes that environmental factors resulting from a mismatch between environment of evolutionary adaptedness and culture initiate or perpetuate early motor and sensory-motor lesions, triggering a butterfly effect of multi-directional cascades of atypical developmental in the complex adaptive system of the parent and ASD-susceptible infant. Chronic sympathetic nervous system/hypothalamic-pituitary-adrenal axis hyperarousal and disrupted parent-infant biobehavioral synchrony are the key biologic and behavioral mechanisms perpetuating these atypical developmental cascades. A clinical translation of this evidence is proposed, for application antenatally and in the first 6 months of life, as pre-emptive intervention for ASD.
Background
What Is Autism?
Autism spectrum disorders (ASD) are a group of complex and heterogenous neurodevelopmental conditions, with life-long presentations in behavior, communication, cognition, and mental health. They are defined behaviorally, by early onset of persistent difficulties in social communication and interaction, sensory atypicalities, and repetitive, restricted interests and activities causing impairment in daily life. Due to the multiple and interdependently variable traits that pertain to autism, Autism-Related Disorders has been recently proposed as a more accurate clinical descriptor (Greenspan, 2018). ASD presents differently in males and females, in a ratio of 4:1. Traits of autism are continuously distributed in the general population and overlap with clinical phenotypes.
Autism spectrum disorders is diagnosed in one in 68, or 0.7%, of Australians. Prevalence has increased across all age groups between 2009 and 2015 in Australia, most markedly in the 5–14 years age group, consistent with international trends. This rapid increase is attributed to increased awareness and diagnosis. Although signs may emerge much earlier, most children are not diagnosed until at least 2 or 3 years of age, and the mean age of diagnosis in Australia is 4.5 years (Synergies Economic Consulting, 2011).
The total direct and indirect cost of ASD annually was estimated at $5.8 billion in Australia in 2010, with the high cost of treating ASD and its associated co-morbidities subject to intense media scrutiny during the initial roll out of the National Disability Insurance Scheme (Synergies Economic Consulting, 2011). The cost to the health system over the life-span for each individual diagnosed with ASD has been estimated at $2.4 million in the United States, or £1.5 million in the United Kingdom, higher than for asthma or diabetes (Buescher et al., 2014).
Etiology
Autism spectrum disorder are polygenic syndromes, associated with 102 genes in a 2018 genetic sequencing study, and also with de novo mutations (De Rubeis and Buxbaum, 2015; Satterstrom et al., 2018). However, any individual’s complex genetic susceptibility is impacted by a myriad of environmental factors in intra-uterine and early life, which alter epigenomic regulation and phenotype expression (Tordjman et al., 2014; Keating, 2016; Mandy and Lai, 2016; Matas et al., 2016; Ismail et al., 2017; Barker et al., 2018).
Autism spectrum disorder is best conceptualized, applying a dynamic systems or complexity science framework, as a spectrum disorder of connectome development, that is, of the neural wiring of the brain, in which one or more feedback loops amplify small variations in very development. Initial lesions trigger a ‘butterfly effect’ of unpredictable cascades of structural and functional imbalances in the global neuronal workspace, which dynamically interact with and impact upon the infant’s social and non-social environmental experiences, amplifying feedback loops and affecting behaviors, cognition, and social communication long-term (Fields and Glazebrook, 2017).
The initial lesions which cause feedback loop disruptions may be structural, either genomic or due to injury; or functional, for example, from changes in the monoaminergic system. Variable phenotypic expressions emerge out of the compensations of the child’s neural networks in response to very early lesions, as myriad feedback loops in the complex adaptive system of the global neuronal workspace compensate for deficiencies and maintain the best possible functional stability. This may occur at the expense of, or with unusual development of and compensation by, higher order cognitive functions like memory, attention and executive functions. The heterogeneity of ASD reflects the multiple different disturbances that can occur along any one of multiple pathways. Dysregulation in any one neural or physiological pathway causes a cascade of events culminating in a cluster of symptoms (Fields and Glazebrook, 2017).
At the cellular level, feedback loop imbalances sculpt neuron morphology and synaptogenesis and alter synaptic transmissions by excitatory or inhibitory neurons. By the time a child is diagnosed with ASD, neuroanatomic patterns of excessive short-range connections and weakened long-range connections have emerged in vulnerable parts of the brain, including in parts of the prefrontal cortex associated with attention, social interaction, emotions, and executive control, and in a decreased density of axons below limbic cortices such as the anterior cingulate cortex (Zikopoulos and Barbas, 2010, 2013; Garcia-Cabezas et al., 2018; Zikopoulos et al., 2018; Trutzer et al., 2019).
Comorbidity of ASD with other medical disorders, including neurodevelopmental, psychiatric, and physical disorders, is common in both children and adults, further supporting the hypothesis that impaired neural and physiological developmental cascades occur in response to very early structural or functional neural lesions. Resultant neural and physiological morbidities then interact with co-occurring behavioral morbidities (Tye et al., 2019).
High-Risk Siblings
An infant who has an older sibling diagnosed with ASD is referred to in this paper as a high-risk sibling. An estimated 1 in 5 high-risk siblings will develop ASD (Ozonoff et al., 2011). High-risk siblings who do not develop ASD are nevertheless at risk of suboptimal developmental outcomes (Landa et al., 2012). High-risk siblings who develop ASD comprise only a small proportion of children diagnosed with ASD, and it is unclear to what extent findings from high-risk sibling studies can be generalized to ASD children from simplex families. When 244 two-year-olds diagnosed with ASD were compared, those with an older ASD sibling showed improved cognitive abilities relative to those from simplex families, perhaps because multiplex parents modify their parenting in response to their previous ASD experience (Dissanayake et al., 2019).
Autism Prodrome
Research in recent years has focused on identification of an autism prodrome, with the aim of developing effective screening and early intervention. Signs that may be diagnostic of ASD, including repetitive behaviors, delayed motor and language skills, reduced gesturing, and impaired eye gaze patterns, emerge from 12 months of age (Bradshaw et al., 2015; Pierce et al., 2016; Petinou and Minaidou, 2017; Shen and Piven, 2017; Whitehouse, 2017; French and Kennedy, 2018; Green and Garg, 2018; Vivanti et al., 2018). From 6 months of age, high-risk siblings are more likely to show reduced attentiveness to parent, reduced affective signaling, poorer co-ordination of communication, and less use of gestures (Wan et al., 2018). These and other soft signs of neurodevelopmental vulnerability have been identified from 6 months of age in children who are later diagnosed with ASD, and may be evident even earlier (Bolton et al., 2012).
However, it’s not possible to distinguish infants with behavioral signs of neurodevelopmental vulnerability who will be later diagnosed with ASD, from those with behavioral signs of neurodevelopmental vulnerability who will develop other neurodevelopmental disorders (Levit-Binnun and Golland, 2012). Because of the extreme heterogeneity of ASD, the non-specific nature of early signs of neurodevelopmental vulnerability, and the significant risks associated with early diagnosis, the search for clinically useful biomarkers of ASD in the first 6 months of life may not prove realistic (Goldani et al., 2014).
Pre-emptive and Early Intervention
This paper defines very early life as the first 100 days post-term, and early life as the first 12 months post-term, distinguishing these periods from intra-uterine life. Early intervention is defined here as an intervention for infants showing signs of neurodevelopmental vulnerability or ASD that is implemented between 6 to 18 months of age, and pre-emptive intervention as an intervention for infants less than 6 months of age, whether or not they are showing soft signs of neurodevelopmental vulnerability.
The International Society for Autism Research Calls for Rigorous Theoretical Framing and Real World Methodologies
In a 2019 discussion of gaps in ASD research, Bailey laments that:
… Genetic studies continue to search for risk variants of small effect in complex groups, biological research proceeds in its attempt to find unitary underlying commonalities in members of the ASD category, and early detection research continues to search in vain for markers with clinically useful specificities and sensitivities.
In this context, Dawson calls for “new and paradigm-shifting interventions and research methodologies which embrace heterogeneity,” and Charman highlights the need to “[transfer] new knowledge about risk factors, biomarkers, and behavioral signs into practical applications” for health professionals (Amaral et al., 2019).
In an era of big data, theoretical framing is key to ongoing knowledge development and cost-effective, sustainable innovation (Campbell et al., 2007; Fox et al., 2014; Ioannadis, 2014). Between 2015 and 2017, the International Society for Autism Research sponsored a Special Interest Group on Implementing and Evaluating Community-based Early Intervention. The Special Interest Group calls for:
• Carefully developed theoretical frames to guide early intervention development, evaluation, and implementation;
• An understanding of factors that make interventions more likely to be adopted and implemented in the real world;
• Rapid progression from treatment theory to community-based effectiveness trials, with community-partnered participatory research;
• An iterative approach consistent with the principles of real world research, with multidirectional knowledge exchange occurring between different phases of the research (Vivanti et al., 2018).
Green and Garg observe that large amounts of public and private money are currently being invested internationally in interventions for ASD which lack rigorous theoretical frames or an evidence base, and call for integration of psychosocial and biological intervention models, in order to eludicate potential synergies between promising psychological and biological intervention mechanisms in ASD (Green and Garg, 2018). Green asks that:
… prodromal intervention should not be tied to legacy theories of the past [but] be responsive to emerging results from basic science to get a more profound understanding of early developmental process and how best to intervene (Green, 2019).
Here, I aim to respond to these calls, as a primary care medical practitioner who specializes both clinically and as a researcher in the care of parents with infants. This paper integrates existing evidence across multiple disciplines and etiological models, in order to explicate the theoretical frames which underlie a proposed interdisciplinary pre-emptive intervention for ASD, applied from the antenatal period and throughout the first 6 months of life.
The International Society for Autism Research Special Interest Group also calls for isolation and evaluation of ‘active ingredients’ of ASD interventions (Vivanti et al., 2018). But from a complexity science perspective, epigenomic regulation of heterogenous ASD genetic susceptibility is impacted by a myriad of interacting and co-evolving environmental variables (Tordjman et al., 2014; Keating, 2016; Matas et al., 2016; Mandy and Lai, 2016; Ismail et al., 2017; Barker et al., 2018). A reductive approach which aims to identify a small number of active ingredients is, applying complexity science, predicted to have limited real world application. This is because any ingredient or mix of specific selected ingredients (that is, simplified, linear interventions) risk unintended outcomes when applied to complex adaptive systems (Campbell et al., 2007; Braithwaite et al., 2018). This may be one reason why guidelines and interventions designed in university or tertiary settings often lack relevance in community settings (Steel et al., 2014).
A complexity or whole systems approach demands development of a comprehensive and integrated pre-emptive intervention, translated from a rigorous theoretical frame, to address the many environmental factors which co-evolve together and interact with the epigenome, to protect against or mitigate expression of ASD genetic susceptibility. That is, holistic pre-emptive intervention is applied flexibly in order to stabilize as many of the myriad feedback loops within the complex adaptive system of each unique infant’s global neuronal workspace and within the complex adaptive system of the infant’s unique family as early as possible, in order to have the greatest chance of success (Marin, 2016; Green, 2019).
Leading primary care researchers have developed innovative methodologies suitable for the evaluation of complex interventions, since community-based healthcare is typically confronted by complex problems requiring early, complex, cost effective, preventive interventions (Plsek and Greenhalgh, 2001; Campbell et al., 2007; Greenhalgh et al., 2015; Braithwaite et al., 2018).
Why Pre-emptive Intervention?
To date, early intervention studies for ASD have been conducted after 6 months of age, either with high-risk siblings or with infants showing signs of neurodevelopmental vulnerability. They derive from applied behavior analysis and social communication models of ASD etiology, applying parent-mediated enrichment of the social interactive environment, and demonstrate preliminary positive outcomes on parent-infant interactions. In particular, evaluation of these early interventions suggest that undemanding synchronous parent behavior, which notices and responds to the child’s focus without suggesting a new action, is associated with stronger joint attention and language skills throughout childhood (Steiner, 2013; Rogers et al., 2014; Baker et al., 2015; Baranek et al., 2015; Bradshaw et al., 2015; Green, 2017; Petinou and Minaidou, 2017; French and Kennedy, 2018; Green and Garg, 2018; Wan et al., 2018; Whitehouse et al., 2019).
However, functional neuroimaging of high-risk siblings who later develop ASD demonstrates altered cortico-cortical connectivity from 6 months of age (Lewis et al., 2017; French and Kennedy, 2018). Regional MRI volumes across the whole brain in 4–6 month old high-risk siblings show larger cerebellar and subcortical volumes than in low risk infants, linked to more repetitive behaviors at 36 months (Pote et al., 2019). These findings suggest that initial changes in the global neuronal workspace occur either prenatally or in very early life. By the time an infant is 6 months old, the effects of very early structural or functional lesions in the global neural workspace are already entrenched, and developmental cascades of multi-directional perturbations in motor, sensory, and social interactions are underway. That is, early interventions evaluated to date have been applied to an already fundamentally altered neural landscape.
Studies of pre-emptive intervention for ASD, as defined in this paper, have not yet appeared in the scientific literature. This paper integrates current understandings of neural network developmental dynamics to support the hypothesis that ASD course alteration is possible, beginning with antenatal anticipatory guidance, followed by intervention in the first months of life.
The First 100 Days Post-Term: Window of Critically Injury-Sensitive Neuroplasticity
Neural and Behavioral Biomarkers
Neural network development activity peaks between the last three months in utero and the first three months post-birth, spanning from the beginning of cortical subplate diminution until the subplate disappears as permanent circuitries in the primary motor, somatosensory, and visual cortex take over. Until then, sensory-motor information continues to be relayed through subplate neurons, which sculpt permanent cortical templates. Cortical subplate neurons are known to be selectively sensitive to injury, and very early sensory-motor defects due to subplate neuronal injury have been implicated in neurodevelopmental disorders, including ASD (Kanold and Luhmann, 2010; Wess et al., 2017; Hadders-Algra, 2018a).
From 10 weeks post-conception, the brainstem and spinal cord central pattern generator initiates general movements. These disappear by three to 5 months post-birth, as fidgety movements and goal directed, affect-driven movements of the limbs emerge and the cortical subplate disappears. Hadders-Algra notes that the emergence, blooming, and eventual disappearance of general movements coincides temporally with the emergence, dominance, and disappearance of subplate synaptic activity. She hypothesizes that general movement complexity and variability are modulated by the cortical subplate and mediated by subplate motor efferents. In her model, abnormal general movements result from lesions in either the subplate or in subplate motor efferent connections within the periventricular white matter (De Graaf-Peters and Hadders-Algra, 2006; Hadders-Algra, 2007, 2017, 2018a).
Significantly, at the time of birth and in the middle of the period of peak neural network development, noradrenergic alpha 2 receptors and glutamatergic N-methyl-D-aspartate receptors are temporarily overexpressed, with high serotonergic innervation and dopaminergic turnover. Hadders-Algra hypothesizes that monoaminergic exuberance from birth until three months of age is associated with increased neural excitability, including of motoneurons. She speculates that monoaminergic exuberance offers evolutionary advantage, and is expressed in the writhing character of general movements. This over-expression resolves as the permanent cortical circuitries take over from the subplate (De Graaf-Peters and Hadders-Algra, 2006; Hadders-Algra, 2007, 2017, 2018a).
Behaviorally, persistence of the cortical subplate and increased monoaminergic excitability correspond post-birth with the infant crying diathesis. The crying diathesis can be viewed, then, as a behavioral biomarker, and the cortical subplate as a neural biomarker, of the critically injury-sensitive neuroplasticity of an infant’s first 100 days of life.
Stress Response Settings
Neural templates for emotional and stress regulation are laid down in the cortical subplate and cortex, and in subcortical structures such as the basal ganglia and the amygdala, during the developmentally critical first 100 days post-birth. These templates form the basis for secure psychological attachment and mental health life-long. As originally proposed by Greenspan in the Affect Diathesis Model, this paper hypothesizes that chronic sympathetic nervous system and hypothalamic-pituitary-adrenal axis (SNS-HPA) hyperarousal has monoaminergic impacts on the brainstem and cortical subplate during the critical window of monoaminergic excitability and injury-sensitive neuroplasticity, and is a key physiological mechanism disrupting capacity for attention on developmental tasks, mediating atypical developmental cascades (Schore, 2001; Greenspan, 2002; Lydon et al., 2016; Marin, 2016; Mandy and Lai, 2016; Ismail et al., 2017; Patel et al., 2018).
Although genetic susceptibility to long-term HPA axis dysregulation varies, painful perceptual experiences in very early life, including fear or chronic stress (experienced physiologically as chronic SNS-HPA hyperarousal), risk lifetime changes in SNS-HPA axis settings and dopaminergic, serotonergic, or noradrenergic circuitries. The HPA axis does not return to its ‘unstressed’ state, normal patterns of cortisol release fail to emerge, and behaviorally, infants show difficulty moving out of hyper-aroused or hypo-aroused states.
Stress also increases the permeability of the blood brain barrier, allowing penetration of pro-inflammatory cytokines. The immune dysregulation associated with chronic stress and increased allostatic load has widespread effect on physiological systems underlying developmental processes and mental and physical health, including on growth, metabolism, immunity, and cognition (Marin, 2016; Barker et al., 2018; Koss and Gunnar, 2018).
Executive functions and the development of sustained intention, including joint attention, are particularly sensitive to stress. In 12 months old neurotypical infants, those with high autonomic reactivity to stressors show short attention durations, and those with lower autonomic reactivity show longer attention durations (Wass et al., 2018). In older children diagnosed with ASD, stress triggers worsened signs and symptoms, impairing the capacity to achieve developmental tasks, which further upregulates the stress response, perpetuating multi-directional developmental cascades. This paper proposes that the same chronic SNS-HPA hyperarousal known to impact negatively on capacity to perform developmental tasks in the older child similarly impedes an infant’s capacity to perform the vital motor and sensory-motor developmental tasks of very early life.
Chronic SNS-HPA Hyperarousal in Very Early Life Is Modifiable by Environmental Factors
The infant crying diathesis is characterized by high levels of cry initiation, which are stable across cultures. In high income countries, the crying diathesis is characterized by durations of cry that average about 2 h a day for the first 6 weeks, decreasing to 72 min a day at 12 weeks, and has resolved, for most, by about 16 weeks post-birth (Wolke et al., 2017). Twenty percent of families report excessive crying, of over 3 h a day, for at least 3 days a week, and many more report cry-fuss problems (Wake et al., 2006). But importantly, infant crying is modifiable according to socioculturally determined environmental factors. Cultures more likely to breastfeed, offer physical contact, and provide cued care have the same high frequency of cry initiation in this period, but at least halved cry durations (St James-Roberts et al., 2006; Wolke et al., 2017).
This paper proposes pre-emptive intervention for ASD which integrates the neurobiological model of cry-fuss problems (Table 1). In the complex adaptive system of parent and infant, problems of crying and fussing, feeds, and sleep often interact and co-evolve, and may be referred to as ‘regulatory problems,’ or ‘dysregulation.’ The neurobiological model of cry-fuss problems conceptualizes problem crying as a behavioral biomarker of chronic SNS-HPA hyperarousal, that is, of stress in very early life (Douglas et al., 2011; Douglas and Hill, 2013a). Crying and fussing, or chronic SNS-HPA hyperarousal, emerge when the parent-infant complex adaptive system is unable to stabilize multiple feedback-loop disruptions arising from interacting and co-evolving environmental factors (Douglas et al., 2011).

Table 1. Current explanatory models for cry-fuss problems.
The neurobiological model of cry-fuss behaviors identifies three key environmental causes of chronic SNS-HPA hyperarousal in the first 100 days post-term:
(1) Suboptimal environmental stimulation
(2) First wave behavioral (FWB) approaches to infant sleep
(3) Unidentified and unmanaged feeding problems, either
(a) Poor satiety
(i) due to compromised milk transfer in breastfeeding, typically due to suboptimal fit and hold (‘latch and positioning’), and resultant low supply
(ii) due to feed-spacing in either breast or formula-fed infants
(b) Functional lactose overload in breastfed infants
(c) Conditioned SNS-HPA hyperarousal with feeds, most often due to positional instability at the breast, or coercive practices during either breast or bottle feeds (Douglas and Hill, 2013a).
In the neurobiological model of cry-fuss problems, these disruptive environmental factors interact unpredictably in the context of monoaminergic excitability in the first 100 days of life and predispose to:
(1) High levels of SNS-HPA arousal, which temporarily trigger more SNS-HPA arousal, in a positive feedback loop of inconsolable crying (correlating with monoaminergic excitability);
(2) Development of conditioned SNS-HPA hyperarousal in response to particular triggers e.g., conditioned SNS-HPA hyperarousal with breastfeeding or bottle-feeding;
(3) Temporary re-setting of the baby’s ‘stress thermostat’ in response to chronically high levels of SNS-HPA arousal (also known as ‘difficulty shifting arousal states’), which resolves at the end of the crying period;
(4) Permanent re-calibration of the ‘stress thermostat’ in biologically susceptible infants (that is, permanent alteration of neural connectivity), due to:
(a) Alteration of the infant’s stress response settings, and/or
(b) Cascade effects of altered parent-infant interactions;
(5) Maternal chronic SNS-HPA hyperarousal in response to infant crying, feed, and sleep problems, which increases the risk of:
(a) Postnatal anxiety and/or depression
(b) Avoidance behaviors (Douglas and Hill, 2013a).
Chronic SNS-HPA Hyperarousal in Very Early Life Permanently Alters Stress Response Settings in Susceptible Infants
Multiple studies link infant regulatory problems in the domains of feed, cry-fuss behaviors and sleep with impaired developmental and behavioral outcomes in later childhood. This association is strongest if there is more than one regulatory problem, or if the regulatory problem persists beyond 4–5 months of age (DeGangi et al., 2000; Desantis et al., 2004; Von Kries et al., 2006; Schmid and Wolke, 2014; Winsper and Wolke, 2014; Santos et al., 2015; Laetitia et al., 2016; Sidor et al., 2017; Williams et al., 2017; Bilgin et al., 2018; Breeman et al., 2018; Cook et al., 2019). Regulatory problems in infancy have also been linked with avoidant adult personality traits (Bauml et al., 2018). Multiple persistent regulatory problems are associated with attention problems in later childhood, and trajectories of attention problems from childhood to adulthood (Bilgin et al., 2018). A comparative study which claims to show no link between regulatory problems in the first 3 months of life and behavior problems at 2–3 years of age retrospectively excludes infants whose crying was found to persist at 6 months of age (Bell et al., 2018). Yet it is not possible to predict which infants with cry-fuss problems in very early life will have persistent crying at 5 or 6 months of age, and intervention should be offered promptly to all parents reporting problem crying.
More ‘cued care’ at term predicts fewer regulatory problems at three months, but after the first highly neuroplastic 100 days, patterns of chronic SNS-HPA hyperarousal may become entrenched in susceptible infants, since cued care beyond this age does not result in less regulatory problems. Biological vulnerabilities such as preterm birth provide more significant prediction of long-term regulatory problems (Bilgin and Wolke, 2017). But upregulated infant behavior, or crying and fussing, also predisposes to negative affect and mood problems in parents (Vik et al., 2009; Cook et al., 2017), which disrupt the parent’s capacity to engage and lengthen reciprocity chains. This bi-directional disruption to parent-infant biobehavioral synchrony adds to allostatic load in infants.
These findings corroborate the neurobiological model of infant crying, which argues that while most families are resilient, and suffer no long-term effects of excessive infant crying, cry-fuss problems are a behavioral biomarker of infant stress and disrupted parent–infant biobehavioral synchrony due to modifiable environmental factors, and affect the infant’s capacity to attend to developmental tasks. In a small but important subset of genetically susceptible infants or psychosocially vulnerable families, cry-fuss problems are then associated with long-term SNS-HPA axis disruption and developmental impairment (DeGangi et al., 2000; Desantis et al., 2004; Von Kries et al., 2006; Schmid and Wolke, 2014; Winsper and Wolke, 2014; Santos et al., 2015; Laetitia et al., 2016; Sidor et al., 2017; Williams et al., 2017; Bilgin et al., 2018; Breeman et al., 2018; Cook et al., 2019).
Primacy of Motor Lesions in Development of ASD
Motor, sensory, cognitive, and social communication development are not independent phenomena occurring in sequential order or in discrete neuroanatomic locations, but are complex functions which dynamically co-evolve within the global neuronal workspace of the complex adaptive systems of the developing fetus and infant (Whyatt and Craig, 2013; Estes et al., 2015; Fields and Glazebrook, 2017).
Up to 60% of children with ASD have motor deficits, including impaired postural control, impaired motor planning and sequencing, which affects gesture planning and imitation, and low motor tone (Paquet et al., 2016; Paquet et al., 2019). Prospective or feedforward mechanisms of motor timing, also referred to as sensory-motor intentionality, are typically disrupted in ASD, as is perceptual awareness of others’ intentions conveyed in body movement or eye gaze. Torres et al. argue that motor disruption is a core feature of ASD, requiring assessment methods suitable for inclusion in clinical diagnostic criteria (Torres et al., 2013).
This paper draws on the hypotheses concerning the primacy of either in utero, intra-partum, or very early life motor lesions in ASD etiology, as detailed in ground-breaking syntheses by Torres et al. (2013), Trevarthen and Delafield-Butt (2013), Delafield-Butt and Trevarthan (2017), Dadalko and Travers (2018), and Delafield-Butt et al. (2018). These researchers propose that a structural (genetic or injury) or functional (monoaminergic) motor lesion is sustained in the brainstem systems or cortical subplate in the critically neuroplastic window in utero, intra-partum, or in very early life, which impairs prospective, affect-driven movement. Cascades of imbalance between local and global connectivity emerge, resulting in atypical neurological, psychological, and behavioral development. Chronic SNS-HPS hyperarousal and disrupted parent-infant biobehavioral synchrony are key physiological and behavioral mechanisms which either predispose to very early motor lesions, or perpetuate the effects of very early motor lesions.
Infants require complex and unpredictable postural variability from birth in order to optimize postural control strategies; they also require rich and active movement experiences in order to learn to perceive visual and tactile stimuli (Hadders-Algra, 2010; Dusing, 2016; Hadders-Algra, 2017; Hadders-Algra, 2018b). Reduced movement complexity and variability, that is, reduced movement repertoire, associated with decreased affect-driven prospective movement from birth, impairs sensory feedback and parent response, limiting capacity to process sensory information (Delafield-Butt et al., 2018). In the phase of secondary variability of general movements, these infants have further difficulty selecting an appropriately adapted strategy from of their repertoire, due to limited variability (Hadders-Algra, 2010; Dadalko and Travers, 2018; Delafield-Butt et al., 2018; Hadders-Algra, 2018b). Shafer et al. (2017) propose that motor stereotypy is a downstream manifestation of low motor complexity. The reduced exploratory motor drive evident in many ASD children is hypothesized to begin with these deficits in primary and secondary variability of movement, which cascade to motor coordination difficulties and impaired complex motor sequencing (Denisova and Zhao, 2017) (Box 1).
BOX 1. Selected studies corroborating the hypothesis that very early life motor lesions initiate the atypical developmental trajectories of ASD.
See Torres et al. (2013), Trevarthen and Delafield-Butt (2013), Delafield-Butt and Trevarthan (2017), Dadalko and Travers (2018), Delafield-Butt et al. (2018) for comprehensive reviews.
A retrospective 2008 study of videos of 20 children diagnosed with ASD demonstrated impaired complexity and variability of general movements and absent or abnormal fidgety movements in infancy (Phagava et al., 2008). A 2013 prospective, longitudinal study of 235 low and high-risk infants aged 6–36 months demonstrated developmental differences from 6 months of age, and poorer fine motor skills were evident in the high-risk infants by 14 months of age (Landa et al., 2013; Bradshaw et al., 2018). In a 2014 study of 158 high-risk siblings and low risk infants, repetitive and stereotyped motor behavior was observed as early as 12 months of age in high-risk siblings who are were diagnosed with ASD at 24 months of age (Elison et al., 2014). A 2017 comparative study of 71 low and high-risk infants showed that high-risk siblings who later develop learning delays have less variety of general movements in the first 8 weeks post-birth, and less variety of movements in response to language, suggesting less flexible sensory-motor systems (Denisova and Zhao, 2017). A 2018 study comparing 86 high-risk siblings with 113 low risk infants showed that high-risk siblings who walked independently at 12 months of age had superior social-communication skills relative to high-risk siblings who weren’t walking. A 2019 prospective study of 437 high-risk siblings and 188 low risk infants were assessed at 6 and 36 months. High-risk siblings have poorer fine motor skills and delayed motor development at 6 months, though these findings are not predictive of later diagnosis of ASD (Iverson et al., 2019).
The social communication developmental pathway depends throughout early life on motor competency and capacity for motor synchrony with another, including mutual gaze, joint attention and shared positive affect, attention disengagement, gesture, touch, and language learning (Yu and Smith, 2017). For example, from birth multiple sequential motor and sensory-motor patterns lead to joint attention. By three months of age, social gaze is the primary modality of coordinated interactions, occurring 30–50% of the time in low risk infants. But attention to eyes declines between 2 and 6 months in infants who later develop ASD (Jones and Klin, 2013), and high-risk siblings perform lower on visual attention tasks at 2 and 3 months of age compared to low-risk infants (Bradshaw et al., 2019).
Eye contact, joint attention, and touch synchrony are driven from birth by the motor-emotional system for the enjoyment of shared experience (Greenspan, 2002; Trevarthen and Delafield-Butt, 2013). Early motor and coordination deficits result in atypical control of eye movements, delays in development of gestures such as pointing, and associated impairment of joint attention (Landa et al., 2013). In neurotypical infants, touch synchrony, the coordination of affectionate touch with episodes of shared gaze, increases significantly from 3 to 9 months with the development of fine-motor skills. In this time, episodes of shared gaze decrease to about a third of the time, while shared attention to objects increases dramatically. This emphasizes the dynamic relationship between downstream development in the motor domain, which allows infants to crawl, grasp, and manipulate objects, and development of social competencies. ‘Sticky’ attention to objects is one of the earliest biomarkers of ASD, evident from 7 to 14 months. Difficulty disengaging attention interferes with social orienting and impairs social communication skills.
Speech comprehension relies on multi-sensory integration, predominantly auditory, enhanced by concomitant visual information; speech capacity relies on motor competence (McCleery et al., 2013; Akhtar et al., 2016; Stevenson et al., 2018).
The relationship between motor development in infancy and later cognition correlates neurally with the involvement of extensive cortico-subcortical networks and structures such as the dorsolateral prefrontal cortex and the neocerebellum in both motor and cognitive functions (Heineman et al., 2018).
Up to 87% of individuals with ASD demonstrate atypical sensory processing, often categorized as hyper-sensitive, hypo-sensitive, or sensory-seeking behaviors. From 6 months of age, signs of sensory processing abnormality are predictive of social communication deficits and repetitive behaviors in childhood, and ASD diagnosis (Robertson and Baron-Cohen, 2017).
Synchronous multisensory experiences, including proprioceptive, haptic, visual, and auditory, enhance neural connectivity and sensory-motor processing (Werchan et al., 2018). This paper hypothesizes that impoverishment of environmental stimulation, both social and non-social, chronic SNS-HPA hyperarousal, and disrupted parent-infant biobehavioral synchrony, result in trajectories of compensatory sensory hyper- or hypo-sensitivity and sensory seeking behaviors. In the same way that short-sightedness is a neuronal consequence of prolonged exposure to interior environments with suboptimal opportunities for long-distance focus, this paper proposes that the three variations of sensory processing difficulty are compensations within the global neuronal workspace for suboptimal motor and sensory-motor enrichment in very early life. Compensatory trajectories vary according to the environmental experience, predispositions, and feedback loops activated within that individual infant.
Parent–Infant Biobehavioral Synchrony Optimizes Reciprocity Chains and Protects Stress Response Settings
Biobehavioral Synchrony Integrates Psychosocial and Biological Models of Parent–Infant Interaction
Feldman’s theory of parent-infant biobehavioral synchrony integrates psychosocial and biological models of parent-infant co-regulation, in order to describe interactions which optimize secure psychological attachment and other long-term developmental outcomes (Feldman, 2007, 2016). This paper uses the term biobehavioral synchrony to refer to reciprocal motor and multi-sensory parent–infant exchanges, integrating behavioral observation and parent-reported psychosocial experience with neural and physiological correlates. Biobehavioral synchrony significantly expands upon the concept of synchrony used in social communication models of ASD etiology (Green et al., 2015). Biobehavioral synchrony stabilizes the myriad physiological, neural, and behavioral feedback loops that operate within the complex adaptive system of parent and infant.
To give one example, biobehavioral synchrony between a parent and newborn occurs during contact with a carer’s body and skin. This early proximity, known as skin-to-skin contact, comprises rich and complex sensory-motor stimulation for the infant, and supports both positive parent affect and early infant experience of secure psychological attachment. Skin-to-skin contact improves glucose and oxygen levels, downregulates the infant’s autonomic nervous system, including heart and respiratory rates, and improves breastfeeding outcomes due to sensory-motor activation of mammalian reflexes (Moore et al., 2016).
This paper offers an interdisciplinary synthesis of heterogenous studies investigating infant dysregulation, stress, and parent-infant biobehavioral synchrony. Infant dysregulation may be caused by, and may cause, disrupted parent–infant biobehavioral synchrony. Both parent and infant stress may disrupt parent–infant biobehavioral synchrony, and each may also result from disrupted parent–infant biobehavioral synchrony. In this way, disruptions to parent–infant biobehavioral synchrony in very early life may trigger a butterfly effect of multi-directional cascades of impaired cognitive, social, emotional and self-regulatory skills in ASD susceptible children.
Cued Care
In very early life, an infant signals or cues his or her biological needs, e.g., for milk, or for richer sensory-motor experience, through affect-driven motor behaviors and autonomic cues. Motor behaviors include spinal extension or writhing spinal movements, postural changes, grimaces, and non-speech-like vocalizations, including groans and cries. Autonomic cues include reddened face, increased respiratory rate, and tremors. An infant increases intensity of signaling along a spectrum of SNS-HPA arousal, from mild agitation to screaming.
‘Cued care’ has also been known as ‘responsive care’ or ‘attunement,’ and refers to parenting behaviors which respond to infant cues. Cued care is the foundational behavioral mechanism underlying parent-infant biobehavioral synchrony (Shonkoff and Phillips, 2000; Mansfield and Cordova, 2007; Fogel et al., 2008; Swain et al., 2013; Matas et al., 2016). The care-giver aims to downregulate the infant’s SNS-HPA axis and meet the infant’s needs for safety, nutrition, and loving human-mediated sensory-motor nourishment, by providing eye-contact, touch, motor interaction, and sounds, including speech and song, and also by providing non-human sensory-motor nourishment, such as opportunities for experience of the complex non-human natural environment. Milk and sensory-motor enrichment upregulate the parasympathetic nervous system (PNS), which downregulates the SNS. Downregulation of the SNS-HPA axis facilitates learning and joint attention, as well as cognitive and social processes (Greenspan, 2002).
Extinction of cues may occur when the infant does not receive adequate responses to his or her communications, a process elucidated in the operant conditioning principles of the first wave of the school of behaviorism (FWB), which has been widely applied in infant-care from the mid-twentieth century. The rate at which infant cues are extinguished may also depend on an infant’s underlying genetic or biological susceptibility.
Behaviorally hypo-aroused infants offer minimal cues and are at particular risk of cue extinction. That is, they may quickly respond to SNS-HPA hyperarousal with ‘learned helplessness.’ Minimal cues result in less parental engagement, shortened reciprocity chains, and less opportunities for joint attention, reinforcing the extinction of cues. For these behaviorally hypo-aroused infants, cued care from birth protects against cascades of disrupted development (Greenspan, 2002).
Reciprocity Chains
Reciprocity chains refer to back-and-forth exchanges which incorporate infant responses to parent-initiated communication as well as parental responses to infant-initiated cues. Behavioral reciprocity chains build on cued care, resulting in parent–infant biobehavioral synchrony. Cues from the infant, including affect-driven movements toward the parent, and sensory-motor and speech responses from the parent, including co-ordination of gaze, downregulate and organize the infant’s autonomic, motor, and attentive states. Reciprocity chains also facilitate lactation and downregulate parental stress responses, and are driven by the powerful evolutionary drive for mutual enjoyment.
Between the age of 2–3 months, specific combinations of interactive behaviors become more frequent and turn into constellations of behaviors. From a complexity perspective, these are ‘attractor states,’ recurring patterns that shape neural networks. In the second half of the first year of life, sensory-motor reciprocity chains are increasingly organized, symbolic, and complex, evolving into protophones and verbal communications, although the capacity to perceive and respond to complex non-verbal sensory-motor communication remains foundationally important for social communication throughout life.
Parent–infant biobehavioral synchrony, or enjoyment of increasingly long and complex sensory-motor reciprocity chains, is necessary from birth in order to protect cognitive, language, and social communication development (Britto et al., 2017). This paper proposes that very early life motor or sensory-motor lesions disrupt biobehavioral synchrony, either resulting in, caused by, or perpetuated by chronic SNS-HPA hyperarousal. The resultant shortened, less complex reciprocity chains trigger cascades of atypical sensory processing, motor skills, language, and cognition typical of ASD.
For example, at 4 months of age, the optimal interactive structure comprises a predominance of ‘infant-leads-parent-follows,’ but this is less likely to be found with high-risk siblings, even though maternal responsivity, scaffolding, and linguistic input are the same. By 6 months of age, high-risk siblings demonstrate attenuated social attention and fewer speech-like vocalizations. Although some parents may respond in the second half of the first year of life to minimal infant cuing with disengagement and decreased gaze, many parents of high-risk siblings become directive, in an effort to impose attentional shift and scaffold communicative behaviors. Unfortunately, either pattern may perpetuate a cascade of impaired social development in susceptible infants. ‘Undemanding synchrony,’ a pattern of noticing and responding to the infant’s cues without attempting to control, is the optimal parent response (Green et al., 2017).
Chronic SNS-HPA Hyperarousal in Very Early Life Triggers Atypical Developmental Trajectories in ASD Susceptible Infants
The Avon Longitudinal Study of Parents and Children, a prospective longitudinal cohort study of the offspring of 14,541 pregnant women, demonstrates that high levels of irritable or unsettled infant behavior in very early life predicts autistic traits at 6 months and 2 years of age (Bolton et al., 2012). This association is corroborated by other longitudinal and retrospective studies (Box 2).
BOX 2. Studies demonstrating links between unsettled behavior in very early life and ASD.
In a 2013 audit of 208 preschool children with ASD, 50% of whom had intellectual disability, child health records demonstrated significantly more consultations for crying, feeding or sleeping problems regulatory problems in the first 18 months of life for children who were later diagnosed with ASD (Barnevik et al., 2013).
A 2017 retrospective study investigating 80 monozygotic twin pairs and 46 dizoygotic twin pairs with autistic traits and ASD found that early medical events, such as low birth weight or prenatal valproate exposure, and poor sleep, feeding problems, frequent vomiting, and crying in the first year of life, were associated with an increased risk of autistic traits and ASD. Cumulative load of risk factors correlated with severity of ASD. The authors concluded that non-shared environmental stressors, or allostatic load, resulted in epigenomic expression of autistic traits and ASD in individuals with genetic vulnerability (Willfors et al., 2017).
A 2018 analysis of the Danish National Birth Cohort showed that parents of the 973 children later diagnosed with ASD reported the same rates of periods of crying for over half an hour in the first 6 months of life as non-ASD children. However, when infants who had excessive crying in both groups were compared, those later diagnosed with ASD had cried longer per day and more days per week in the first 6 months than their excessively crying controls. Mothers who reported difficulty caring for their infant when interviewed at 6 months and 18 months post-birth were more likely to have a child later diagnosed with ASD (Lemcke et al., 2018).
In a 2018 retrospective comparative study of 200 children, parent-reported persistent infant crying, and excessive crying with long duration, that is, higher allostatic load, was associated with later diagnosis of ASD (Bag et al., 2018).
In a 2019 longitudinal sample of 282 high-risk siblings compared with 114 low risk infants, assessed at 6, 12, and 24 months of age, 6-month-old high-risk siblings later diagnosed with ASD exhibited poorer parent-reported regulatory capacity and less surgency compared to high-risk siblings and low risk infants not later diagnosed with ASD. Approach behaviors such as smiling and vocal reactivity were less. At 12 months, these temperament differences persisted, with high-risk siblings who were later diagnosed with ASD demonstrating decreased surgency or approach, impaired regulatory capacity, and increased negativity compared with typically developing infants (Paterson et al., 2019).
Motor development, as this paper argues, is primary for healthy development of sensory, social, and cognitive skills, and is facilitated by rich sensory-motor experience, including irregular postural variation. However, the benefits of rich sensory-motor experience occur when the newborn is in a quiet, alert state and able to integrate sensory feedback from primary and secondary generalized movements and other environmental experience (Hadders-Algra, 2018b). This paper hypothesizes that chronic SNS-HPA hyperarousal in very early life, most commonly associated with unsettled infant behavior, but sometimes with hypoaroused behaviors, as discussed below, disrupts capacity to maintain necessary attention on sensory feedback to achieve developmentally appropriate motor and sensory-motor learning tasks, including auditory and visual tasks, joint attention, and motor planning and sequencing (Bilgin et al., 2018; Breeman et al., 2018). These disruptions further perpetuate chronic SNS-HPA axis hyperarousal and add to allostatic load as the child continues to struggle with ongoing developmental tasks, until ASD is diagnosed.
The hypothesis that chronic SNS-HPA hyperarousal disrupts the capacity to attend and develop motor skills is corroborated by the Danish National Cohort study. Although this cohort showed no link between transient ‘infantile colic’ and developmental coordination disorder in later childhood, infants with persistent crying were at increased risk of developmental coordination disorder (Milidou et al., 2015). A prospective study of 37 infants with colic found transient developmental lags in both fine and gross motor skills at 6 months. They were less responsive and less able to maintain optimal functioning for months after resolution of the crying, though these subtle impacts resolved by 12–18 months of age (Sloman et al., 1990).
Corroborating the hypothesis that chronic SNS-HPA hyperarousal plays a key role in perpetuating disrupted feedback loops in ASD etiology, 4-month-old infants later diagnosed with ASD show altered respiratory sinus arrhythmia response to a social stressor compared to non-autistic infants. Similarly, toddlers later diagnosed with ASD have less variable respiratory sinus arrhythmia at 18 months of age (Sheinkopf et al., 2019). ASD adolescents report high levels of anger-focused rumination (Patel et al., 2018).
Hypoaroused Behavior
Although many children diagnosed with ASD are retrospectively reported to have had regulatory problems in the domains of feeds, sleep and cry-fuss problems in very early life, other children diagnosed with ASD are retrospectively characterized as ‘easy babies,’ whose behavioral regulation difficulties emerged in toddlerhood or later.
The Affect Diathesis model proposes that these behaviorally hypoaroused ASD-susceptible infants also experience chronic SNS-HPA hyperarousal, which is subjected to PNS override (Greenspan, 2002). In behaviorally hypoaroused babies, affect-driven sensory-motor cues are partially extinguished, either due to biological propensity or early ‘learned helplessness’ or both, resulting in less parental engagement and shortened reciprocity chains. These infants may be particularly vulnerable to the developmental impacts of suboptimal environmental stimulation and disrupted parent–infant biobehavioral synchrony (Middlemiss et al., 2012).
Atypical Crying Acoustics
One month old infants later diagnosed with ASD are reported by parents to have louder, more distressed, less typical cries, as if in pain (English et al., 2018). At 6 months of age, parents report that high-risk siblings produce cries with higher pitch and greater variability than low risk infants, as if in pain. High-risk siblings later diagnosed with ASD produced the widest frequency range and most poorly phonated cries. Parents report difficulty understanding the causes of crying episodes in the early life of infants later diagnosed with ASD (Esposito et al., 2017). Paradoxically, by 12 months of age, high-risk infants and infants later diagnosed with ASD have shorter cry durations, with higher frequency and decreased number of pauses compared to low-risk or neurotypical toddlers (Unwin et al., 2017). This paper proposes that environmental factors, that is, one or more problems of feeds, sleep and unmet needs for richer sensory-motor and social stimulation, may induce cries at the extreme end of the spectrum of distress in very early life, triggering aversive or avoidant responses in the care-giver, and finally, extinction of cries, associated with cascades of atypical development (Bag et al., 2018). In very early life, infant crying is commonly inappropriately attributed to gut pain (Bergmann et al., 2014; Gieruszczak-Bialek et al., 2015; Gordon et al., 2018).
Suboptimal Sensory-Motor Stimulation
The neurobiological model of infant cry-fuss problems argues that infants in a low sensory environment cry because of unmet needs for sensory-motor enrichment, in a biological bid to optimize development of neural circuitries during a window of critically sensitive neuroplasticity and monoaminergic excitability (Douglas and Hill, 2013a). The human infant’s biological requirement for rich postural variability and rich and complex sensory-motor reciprocity chains for normal motor development has been discussed previously. Interestingly, a study of infant macaques found that rich social environments in the first week of the macaques’ lives improved gaze-following and social skills at 7 months of age (Simpson et al., 2016).
Regulatory difficulties in very early life are typically interpreted, through a linear causative theoretical frame, as an early sign of sensory processing deficit in infants who later develop ASD. By 12 months, when neural changes are entrenched, sensory processing and regulatory reactivity are known to predict parent-reported executive function deficits in children later diagnosed with ASD (Robertson and Baron-Cohen, 2017; Stephens et al., 2018). But this paper applies the neurobiological model of cry-fuss problems to argue that infant dysregulation in very early life is less likely to be a sign of pre-existing sensory processing problems, and more likely to be a behavioral biomarker of chronic SNS-HPA hyperarousal resulting from environmental factors, including suboptimal sensory-motor stimulation, which predispose to sensory processing problems.
Feeding Dysregulation
Children diagnosed with ASD are 5 times more likely to have a feeding problem than neurotypical children (Peverill et al., 2019). In very early life, ‘feeding problems’ refer to fussing at the breast or bottle, and breast or bottle refusal. Infant feeding problems often result in a cascade of worsening parental anxiety, controlling parental feeding practices, and conditioned infant withdrawal or distress (Estrem et al., 2016). The latter is referred to in the neurobiological model of cry-fuss problems as ‘conditioned SNS-HPA hyperarousal’ with feeds, and may result in temporary re-setting of the HPA axis in the critically sensitive neural networks of very early life (Douglas and Hill, 2013a). Feeding-related signs such as refluxing or back-arching and fussing with the breast or bottle are often inappropriately medicalized, and underlying causes, such as positional instability with breastfeeds or conditioned SNS-HPA hyperarousal with breast or bottle feeds, remain untreated (Douglas, 2013). Feeding problems may escalate into a developmental trajectory of dysregulated feeding behavior into childhood (Winsper and Wolke, 2014). In the older child, the term ‘feeding problems’ encompasses a range of concerns, including food selectivity, problematic mealtime behavior, and oromotor challenges, and are most commonly the downstream effect of early life feeding problems. Children with feeding problems may withdraw from or reject their mother’s touch; their mothers touch them less; and shared eye contact is reduced (Estrem et al., 2016).
Feeding problems in children diagnosed with ASD decrease from time of diagnosis to clinically insignificance by school age, the same pattern followed by feeding problems in neurotypical children. However, ASD children with higher levels of anxiety or mood difficulties and externalizing behavioral challenges are most at risk of a chronic course of feeding problems, with bi-directional negative effects on family interactions, emphasizing the role of chronic SNS-HPA hyperarousal in perpetuation of atypical developmental trajectories (Peverill et al., 2019).
Sleep Dysregulation
There are strong links between sleep disruptions in the first months of life and impaired cognitive, sleep and behavioral outcomes at school-age (Williams et al., 2016, 2017). However, a prospective birth cohort of 5151 children in the Netherlands found that reports of sleep problems co-occur with autistic traits in early childhood beginning from 18 months of age, when parental reports of sleep problems are resolving for neurotypical infants (Verhoeff et al., 2018).
Children with ASD show more frequent ‘bedtime resistance,’ sleep anxiety, and longer durations of night waking than children with other development disabilities (Cohen et al., 2014; Valicenti-McDermott et al., 2019). A study of 1201 children with ASD demonstrates links between aggressive behavior and sleep disruption, and proposes that this link is mediated by chronic SNS-HPA hyperarousal, noting that children with ASD experience a great deal of stress resulting from impairments in their ability to communicate with others or to understand the world around them (Shui et al., 2018). Sleep difficulties worsen behavior and cognition problems for the ASD child, and impact negatively on his or her family’s communications.
This paper proposes that sleep problems in children with ASD are a consequence, first, of elevated levels of anxiety or chronic SNS-HPA hyperarousal, and, second, disrupted circadian rhythms, both worsened by the widespread application of ‘sleep training’ or FWB sleep strategies for children with ASD, which paradoxically disrupt the circadian clock.
Adults with ASD have dampened cortisol secretion and subjective somatic physiological arousal, associated with increased sleep onset latency, poorer sleep efficiency, and increased waking after sleep onset. This corroborates the hypothesis that dysregulation of the SNS-HPA is a key mechanism underlying phenotypic expression of ASD (Baker et al., 2019).
Chronic SNS-HPA Hyperarousal Affects the Gut Microbiome and Metabolic and Immune Settings in ASD
The gut is a major microbial–host interface, and a dominant immune organ. Immune function, HPA axis regulation, and the microbiota-gut-brain axis interact and co-evolve in the mother-infant complex adaptive system, and are each affected by stress. The infant gut microbiome in very early life has life-long effect on the settings of metabolism, immune, endocrine, and gut health. Gut dysbiosis is associated with impaired gut barrier integrity, inflammation and autoimmune disease (Cenit et al., 2017; Warner, 2018).
Mothers of children with ASD are twice as likely to report at least one gastrointestinal symptom in their child between 6 and 36 months. High-risk siblings have a greater prevalence of gastrointestinal symptoms. Young children diagnosed with ASD demonstrate significant gut dysbiosis (Coretti et al., 2018). Chronic constipation is the most common gastrointestinal problem in children with ASD. Many children with ASD have functional gastrointestinal tract disorders relating to selective eating, medications, and differences in sensory processing, which are associated with increased anxiety (Tye et al., 2019). Gastrointestinal problems may worsen developmental cascades of child behavior problems and disrupted family interactions. Studies show a strong correlation between gastrointestinal dysfunction and autism severity, across all domains including speech, social and behavioral (Ding et al., 2017). Early clinical trials suggest that by targeting the gut ecosystem, both ASD and gastrointestinal tract symptoms can be impacted, suggesting potential shared mechanisms (Tye et al., 2019).
However, this paper proposes that widespread inappropriate medicalization of infant behavioral cues, usually as signs of gut problems, results in a failure to identify and manage the underlying environmental factors which precipitate chronic SNS-HPA hyperarousal in very early life. Inappropriate medicalization of upregulated behavior and the failure to identify and manage underlying clinical problems have deleterious effects on gut health long-term. Instead of applying a linear and causative disease model which assumes that gut dysbiosis causes excessive infant crying, the neurobiological model of cry-fuss problems proposes that chronic SNS-HPA hyperarousal and feeding problems, and also the widely prescribed proton pump inhibitors, interact in the complex adaptive system of the mother and infant, out of which gut dysbiosis emerges (Douglas P.S. and Hill P.S., 2011; Douglas and Hill, 2013a; Castellani et al., 2017; Rhoads et al., 2018). This paper proposes that these and other environmental factors disrupt parent-infant biobehavioral synchrony, precipitating multiple trajectories of atypical development in ASD susceptible infants.
Chronic inflammation is emerging as the critical pathophysiological feature of mental disorders generally, associated with dysregulation of normal microglial synaptic pruning (Cenit et al., 2017). Siniscalco hypothesizes that pro-inflammatory processes and immune alterations in very early life are etiological events for autism (Siniscalco, 2015) (Box 3). ASD has been linked with:
• Increased pro-inflammatory cytokines in the cerebro-spinal fluid
• Acquired mitochondrial dysfunction, an early sign of neurodegeneration
• Decreased antioxidants in urine
• Higher plasma GABA levels
• Amino acid and neuropeptide disruptions
• Increased autoimmune antibodies targeting central nervous system proteins.
BOX 3. Evidence demonstrating that ASD is a pro-inflammatory state.
Many autism susceptibility genes are localized in the immune system and related to immune or infection pathways (Carter, 2019), and ASD is associated with an increased prevalence of specific immune-related conditions (Tye et al., 2019). Endoplasmic reticulum stress, oxidative stress, and apoptosis have been proposed as molecular mechanisms underlying autism (Dong et al., 2018). The oxidative and integrated stress responses are upregulated in the autism brain and may contribute to myelination problems (Carter, 2019). Increased autoantibodies directed toward central nervous system proteins have been observed in children with ASD, which may signal heightened inflammatory processes or an autoimmune component that could decrease the integrity of the mucosal barrier (Tye et al., 2019). Cytokines are proteins produced by neurons that regulate immune responses including hematopoiesis, inflammation, immune cell proliferation and differentiation.
Pro-inflammatory cytokines are found in higher levels in children with more severe ASD symptoms compared to ASD children with milder symptom presentation (Tye et al., 2019). A recent meta-analysis of cytokine levels in unmedicated individuals with ASD, mostly children, confirmed an abnormal cytokine profile, characterized by elevations in proinflammatory cytokines and reduced levels of anti-inflammatory cytokines (Masi et al., 2014).
ASD is associated with altered expression of genes associated with blood-brain barrier integrity, increased neuroinflammation, and possibly impaired gut barrier integrity. Immunological abnormalities affect both the gastrointestinal system and microglial cells of the brain and CNS. Monocytes, the precursors for macrophages, dendritic and microglial cells, show significant pro-inflammatory dysfunctions in ASD children. Individuals with ASD have upregulated inflammatory cytokines which induce blood brain barrier disruption. Altered blood brain barrier permeability directly influences neural plasticity, connectivity and function, and is hypothesized to underlie impairments in social interaction, communication behavior (Siniscalco, 2015).
Chronic SNS-HPA hyperarousal releases cortisol, which alters intestinal motility, gut epithelial permeability, and induces changes in gut microbial composition. This paper argues that the pro-inflammatory state of children with ASD arises from chronic SNS-HPA hyperarousal, or stress and anxiety, with long-term multi-directional impacts upon gut health, behavior and immune trajectories (De Palma et al., 2014; Gottfried et al., 2015; Siniscalco, 2015; Ding et al., 2017; Carter, 2019).
Excessive infant crying, or chronic SNS-HPA hyperarousal, is a pro-inflammatory state, impacting on gut microbiome and permeability, with long-term effects on metabolic, immune and mental health in susceptible infants (Partty et al., 2017; Rhoads et al., 2018). The Affect Diathesis model theorizes that behaviorally hypo-aroused infants also experience chronic SNS-HPA hyperarousal, with PNS override, which would be expected to similarly predispose to pro-inflammatory states (Greenspan, 2002). Anxiety disorder is common in adults with ASD, and symptoms of anxiety cause substantial functional impairment in ASD adults more broadly (Tye et al., 2019). This paper proposes that chronic anxiety and gastrointestinal problems in children and adults diagnosed with ASD are a downstream effect of chronic SNS-HPA axis dysregulation that begins in very early life.
Dysregulated Parent Mood Increases the Risk of ASD in Susceptible Children
A meta-analysis of nine observational studies shows increased ASD risk in children exposed to parental affective, depressive, and bipolar disorders (Ayano et al., 2019). Previous findings of links between maternal antenatal antidepressant use and ASD are attributed to the confounder of maternal mood disorder (Sujan et al., 2019). But there is, in particular, an increased risk of ASD in children of mothers who experience affective and depressive disorders, and this can be attributed to the developmental impacts of disrupted parent–infant biobehavioral synchrony in susceptible infants.
Depressed mothers with a 6-month-old baby show lower baseline vagal tone, linked with poorer emotional regulation and sensitivity to stress; less vagal brake, linked with poorer emotional regulation; less joint attention; and less initiation of interaction including touch and vocalization with the infant. Depressed mothers are less likely to touch the child, take significantly longer to respond to changes in their infant’s behavior, and have less capacity to repair interactive errors. A cycle of mutual disengagement occurs, as the infant develops flat affect and makes fewer bids for joint gaze and response. Matched states are more likely to be negative, in distress or anger. Difficulties in mother-infant biobehavioral synchrony are detected not only in cases of full-blown clinical depression but among mothers with chronic sub-clinical symptomatology (Tietz et al., 2014). An anxious mother may direct very active communications toward her baby, which are not attuned to the infant’s cues. The resultant shortened, less complex reciprocity chains between both highly anxious and depressed mothers and their infants in very early life predict the child’s emotional dysregulation in later life (Feldman, 2007).
Biology-Culture Mismatch in Very Early Life Generates Environmental Factors Which Increase the Risk of ASD in Susceptible Infants
A susceptible infant experiences biology-culture mismatch as adversity. Although the human infant is highly adaptive across a wide variety of culturally determined infantcare practices, a significant gap between cultural practices and evolutionary expectation may result in chronic SNS-HPA hyperarousal in very early life, in addition to other risks (Barr, 1990, 1999; Hofer, 2002; Douglas, 2005; McKenna et al., 2007; Ball, 2008; Gettler and McKenna, 2011; Koss and Gunnar, 2018; Renz-Poster and De Bock, 2018). This paper proposes that the initial motor and sensory-motor neural lesions of ASD are more likely in susceptible children when environmental factors are mismatched with biological expectations during the critically neuroplastic, injury-sensitive first 100 days post-birth, resulting in chronic SNS-HPA hyperarousal which either triggers or perpetuates multi-directional cascades of atypical development.
In very early life, the infant evolved, in the Homo sapien’s environment of evolutionary adaptedness, to expect:
(1) Rich environmental stimulation, including
(a) Prolonged physical contact with older children and adults, including co-sleeping and diverse and frequent social sensory-motor enrichment
(b) High levels of postural variability
(c) Multi-centric social interactions
(d) Complex non-social environmental stimulation, e.g., outdoors.
(2) Affect-driven, increasingly long, sensory-motor reciprocity chains with caring older children and adults. Mutual positive affect between adults and infants, or enjoyment and delight, has ensured Homo sapiens’ evolutionary survival.
(3) Human milk transferred directly from the lactating breast to the infant gut, optimizing the gut microbiome, and metabolic, endocrine, and immune protection. Frequent and flexible breastfeeding facilitates increasingly long and complex motor and sensory-motor reciprocity chains.
Currently, in very early life, parents receive a great deal of conflicting advice from health professionals concerning breastfeeding, infant sensory needs, unsettled infant behavior and parent-infant sleep, and consult with multiple providers (McCallum et al., 2011; Schmied et al., 2011). There are serious gaps in health professional training across disciplines in management of breastfeeding and infant behavior problems (Renfrew et al., 2012; Rimer and Hiscock, 2014; Kirby et al., 2015; Gavine et al., 2017); widespread inappropriate medicalization of infant behavior, risking worsened outcomes; and substantial evidence that popularly applied approaches to breastfeeding and infant regulatory problems do not help parents and babies, or make breastfeeding, crying and sleep problems worse (Douglas, 2013; Svensson et al., 2013; Bergmann et al., 2014; Gieruszczak-Bialek et al., 2015; Kempler et al., 2016; Thompson et al., 2016; Woods et al., 2016; NHMRC, 2017; O’Shea et al., 2017; Gordon et al., 2018).
That is, the gap between biological expectation and socioculturally determined advice concerning infantcare is profound in contemporary societies, placing susceptible infants at neurodevelopmental risk, and increasing the risk of maternal postnatal anxiety and depression (Stein et al., 2014; Dias and Figueiredo, 2015; Tsivos et al., 2015). Infant cry-fuss, feeding, and sleep problems are often highly stressful for parents, and predispose to maternal postnatal depression (Dorheim et al., 2009; Vik et al., 2009; Dias and Figueiredo, 2015).
This paper hypothesizes that three key environmental factors emerge from biology-culture mismatch, and interact in the complex adaptive system of the parent and infant, increasing the risk of ASD in susceptible infants: suboptimal environmental stimulation, disruption of parent-infant biobehavioral synchrony, and gut dysbiosis or feeding problems.
Suboptimal Environmental Stimulation
In Australia and in many countries today, parents continue to be advised that sleep training or FWB approaches are necessary for optimal developmental outcomes and good sleep habits (Table 3). Infant sleep training emerged in the 1950s and 1960s when the first wave of the school of behaviorism (FWB) in psychology was applied to infantcare. Yet high level evidence demonstrates no decreased night waking or reliably improved maternal mood scores as a result of FWB interventions, and no improvement in developmental outcomes (Price et al., 2012; Bryanton et al., 2013; Douglas and Hill, 2013b; Mindell and Lee, 2015; Price et al., 2015; Kempler et al., 2016; NHMRC, 2017; Pennestri et al., 2018).
This paper proposes that application of FWB approaches to infant sleep in very early life impacts negatively on neurodevelopmental outcomes in ASD-susceptible infants, because FWB approaches decrease environmental stimulation in four ways:
(1) Parents are advised to teach the infant ‘good sleep habits’ by having the infant sleep in a cot or on an immobile surface, often in a quiet dim room with deliberately minimized visual stimulation, iteratively throughout the day. This regular recourse to a low sensory interior environment impoverishes sensory-motor experience, both social and non-social, and fails to offer susceptible infants adequate task demands for optimal development of skills in interpretation of sensory information, adaptation of movements in response to external stimuli, and organization of postural control.
(2) Stimulation is problematized. Parents are advised to avoid social and non-social ‘overstimulation.’
(3) Inadequate sensory-motor stimulation results in SNS-HPA arousal (crying and fussing), which parents are advised to interpret as ‘tired signs’ or ‘overstimulation,’ triggering more attempts to put the baby to sleep.
(4) Patterns of inadequate environmental enrichment may result in chronic infant SNS-HPA hyperarousal (Douglas and Hill, 2013a; Whittingham and Douglas, 2014; Ball et al., 2018).
Disruption of Parent–Infant Biobehavioral Synchrony
Our environment of evolutionary adaptedness offered infants rich opportunities for increasingly long and complex motor and sensory-motor reciprocity chains across the two dominant sites of parent–infant transaction in very early life, feeds and sleep. Patterns of cued care are associated in cross-cultural studies with downregulation of both infant and parents’ SNS-HPA axes (Hamilton, 1981; Barr et al., 1991; St James-Roberts et al., 2006; Wolke et al., 2017). Parent–infant biobehavioral synchrony is disrupted by problems of crying and fussing, breastfeeding, and sleep.
Inappropriate Medicalisation of Infant Cry-Fuss Behaviors
The neurobiological model of cry-fuss problems proposes that chronic SNS-HPA hyperarousal, manifesting as crying and fussing in otherwise well infants, emerges out of a mismatch between environment of evolutionary adaptedness and various socioculturally determined environmental factors in contemporary life.
But overdiagnosis and overtreatment, including in children, is a growing global concern, and occurs commonly in very early life, with deleterious effects (Coon et al., 2014; Brownlee et al., 2017). Parents are often taught that their infant’s cry results from physical pain, e.g., from reflux, aerophagia-induced reflux, food allergies or intolerances, lactose intolerance, or from oral connective tissue restrictions (Bergmann et al., 2014; Gieruszczak-Bialek et al., 2015; O’Shea et al., 2017; Gordon et al., 2018; Kapoor et al., 2018). The resultant inappropriate medicalization risks unintended outcomes and perpetuates disruption to parent-infant biobehavioral synchrony (Table 2).
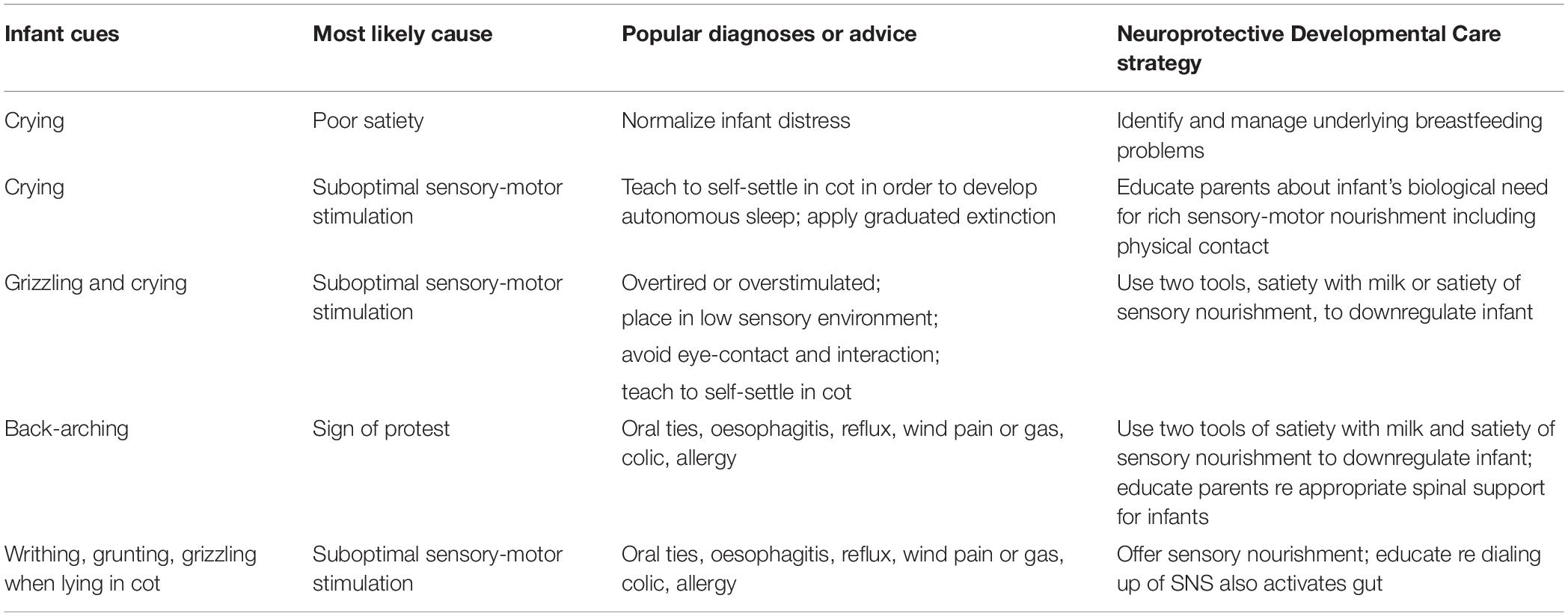
Table 2. Cry-fuss problems: popular sociocultural and clinical advice which disrupts parent–infant biobehavioral synchrony compared with Neuroprotective Developmental Care strategies which promote parent–infant biobehavioral synchrony.
First Wave Behavioral Interventions for Parent–Infant Sleep
In response to emerging neuroscience and psychological attachment research, FWB approaches have adopted a discourse which emphasizes the importance of cued care for secure attachment and optimal mental health outcomes. Yet paradoxically, FWB sleep advice continues to actively disrupt cued care and biobehavioral synchrony, by advising parents to behave in directive and non-contingent ways, iteratively, day and night (Table 3). More extreme FWB strategies, such as minimizing eye contact and interaction at sleep-time throughout the days and nights, iteratively present a ‘still face’ to the infant, which is known to result in greater efforts by the infant to engage at first, before he or she withdraws from cueing (Bertin, 2006). Standard FWB approaches advise parents to not respond to SNS-HPA arousal, or to delay responses, or to respond but not as they believe the baby intends. Parents are advised to iteratively override the powerful biological cues of sleepiness after feeds by applying behaviors such as burping, holding upright, or wrapping, at the same time as they are instructed to achieve prescribed nap frequencies and durations.
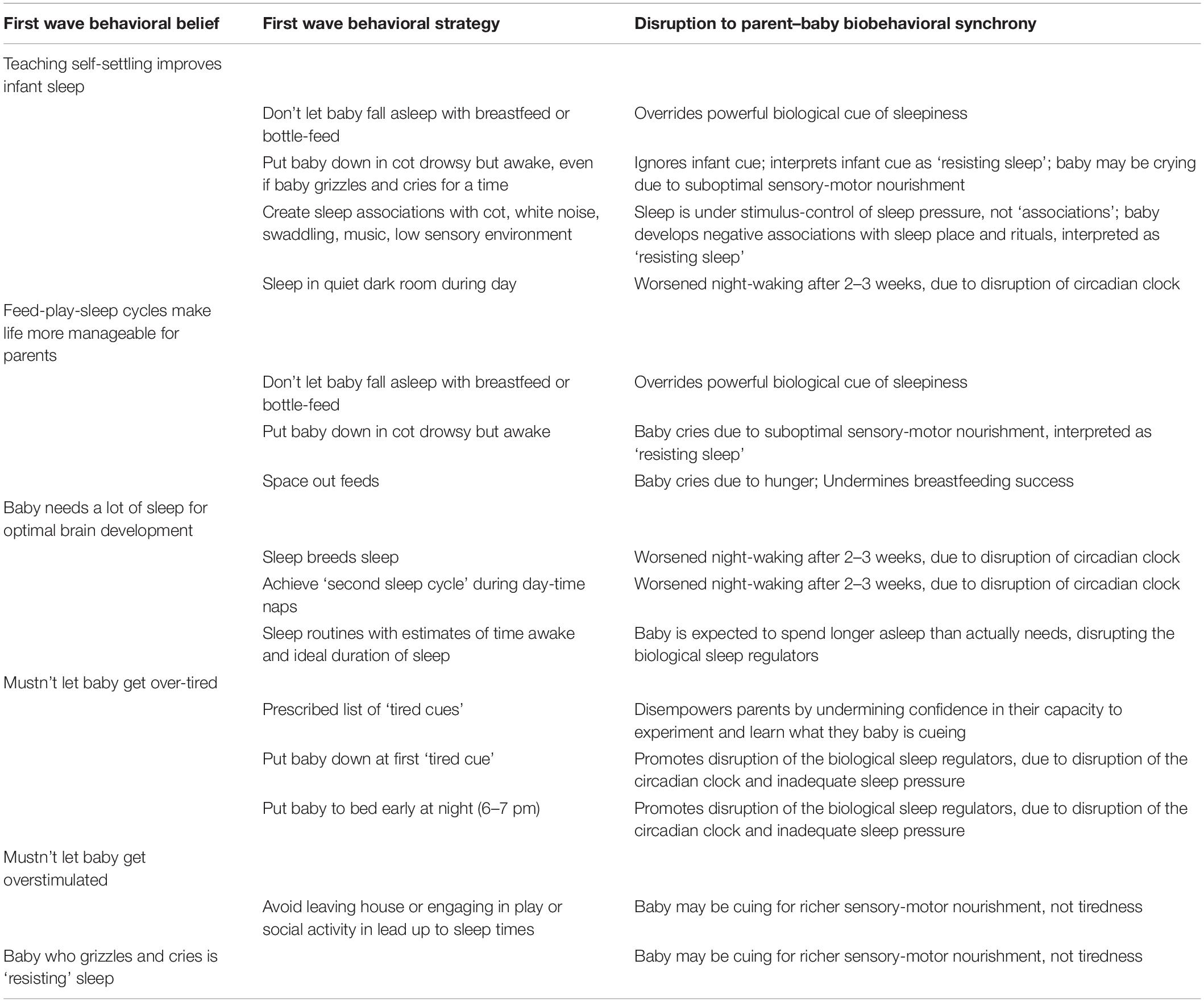
Table 3. Sleep problems: popular sociocultural and clinical advice which disrupts parent–infant biobehavioral synchrony.
FWB approaches may exacerbate parental anxiety, which predisposes to poor sleep efficiency and postnatal depression (Blunden et al., 2016; Etherton et al., 2016). The parent trying to enforce sleep because she believes it necessary for her baby’s healthy development is likely to feel anxious and distressed when the baby ‘resists sleep’ multiple times a day; the infant ‘resisting sleep’ (that is, whose sleep pressure is not yet high enough for easy sleep) is repeatedly subject to the biological stress of a low sensory environment, resulting in chronic SNS-HPA hyperarousal and increased allostatic load. This cycle may place behaviorally hypo-aroused infants at particular risk.
The link between sleep problems in very early life and behavioral and sleep problems in later childhood may be, paradoxically, mediated by the widespread sociocultural and clinical application of sleep training or FWB strategies, resulting in cascades of both parent and infant chronic SNS-HPA hyperarousal (Simard et al., 2017; Williams et al., 2017; Ball et al., 2018). FWB approaches also disrupt the dyadic synchrony of the circadian clock, by promoting long blocks of sleep during the day (Thomas et al., 2014).
Unidentified and Unmanaged Breastfeeding and Unsettled Infant Behavior Problems Increase Risk of Maternal Postnatal Depression
Cry-fuss problems, poor maternal sleep efficiency, and breastfeeding problems are key modifiable risk factors for post-natal depression (Vik et al., 2009; Stein et al., 2014; Dias and Figueiredo, 2015; Tsivos et al., 2015; Werner et al., 2015). Each of these may arise from, or be exacerbated by, a mismatch between popular sociocultural and clinical approaches, and infant biology. Health professionals report inadequate training in management of breastfeeding and unsettled infant behavior problems, and often recommend approaches which have been shown not to help, or may even worsen these problems, with associated deleterious effects on parent–infant biobehavioral synchrony (Price et al., 2012; Bryanton et al., 2013; Douglas and Hill, 2013b; Rimer and Hiscock, 2014; Mindell and Lee, 2015; Price et al., 2015; Blunden et al., 2016; Etherton et al., 2016; Kempler et al., 2016; Thompson et al., 2016; Gavine et al., 2017; NHMRC, 2017; Pennestri et al., 2018).
There is widespread recognition of the importance of prevention of, or early detection and treatment of, perinatal anxiety and depression, the most common mental health condition post-birth. Winnicott proposed that a state of maternal preoccupation or heightened sensitivity develops toward the end of pregnancy and lasts throughout the postpartum period. From an evolutionary perspective, this heightened state supports a woman’s ability to anticipate her infant’s needs and to respond to her infant’s unique cues, and may include anxious, hypervigilant and intrusive thoughts about the infant. In contemporary contexts, often characterized by minimal social support, the same heightened state increases her risk of postnatal anxiety and depression if infant behavior problems emerge.
Gut Dysbiosis and Feeding Problems
In the Homo sapiens environment of evolutionary adaptedness, the complex behavior of breastfeeding is the dominant site of parent-infant motor and sensory-motor interaction in very early life. The infant gut is part of the enteromammary immune system, an extension of the maternal gastrointestinal and immune systems facilitated by breastfeeding. Breastfeeding offers powerful protection of the gut microbiome, with positive impact upon immune and metabolic health.
Successful breastfeeding is linked with increased amounts of cued care (Brown and Lee, 2013), which leads to longer and more complex reciprocity chains. Early breastfeeding is driven by infant stepping, suck and swallow movements (secondary variable movements) and complex sensory-motor interactions, offering rich and diverse environmental stimulation and opportunities for affect-driven sensory-motor reciprocity chains. This may explain associations between breastfeeding and higher IQ (Belfort, 2017). Cued care also improves breastfeeding duration (Brown and Arnott, 2014). Breastfeeding difficulties, and lactation-related breast pain predispose women to postnatal depression, with an associated cascade of related crying behaviors and disruption of mother–infant biobehavioral synchrony (Howard et al., 2006; Brown et al., 2015; Dias and Figueiredo, 2015).
Breastfeeding, either exclusively or with some formula supplementation, is associated with decreased risk of ASD (Schultz et al., 2006; Boucher et al., 2017; Tseng et al., 2017; Bittker and Bell, 2018; Manohar et al., 2018). A Spanish multicenter birth-cohort study of 1,346 children showed that longer duration of breastfeeding protected against autistic traits (Boucher et al., 2017). The Danish National Birth cohort study of 76,322 mothers showed that breastfeeding for less than 6 months was associated with an increased risk of diagnosis of ASD (Lemcke et al., 2018). High-risk siblings are more likely to be weaned earlier. Weaning after 6 months and exclusive breast milk are also associated with protection against gastrointestinal tract symptoms, in particular constipation and abdominal distress, in high-risk siblings (Penn et al., 2016).
This paper proposes that unidentified clinical breastfeeding problems such as positional instability or conditioned dialing up at the breast, may cause chronic SNS-HPA hyperarousal, premature weaning, and related gut dysbiosis (Table 4). Unfortunately, popular clinical approaches often don’t help, or may even worsen breastfeeding problems (Svensson et al., 2013; Thompson et al., 2016; Woods et al., 2016; O’Shea et al., 2017). Developmental trajectories of feeding problems and disrupted parent–infant biobehavioral synchrony may result (Douglas and Hill, 2013a).
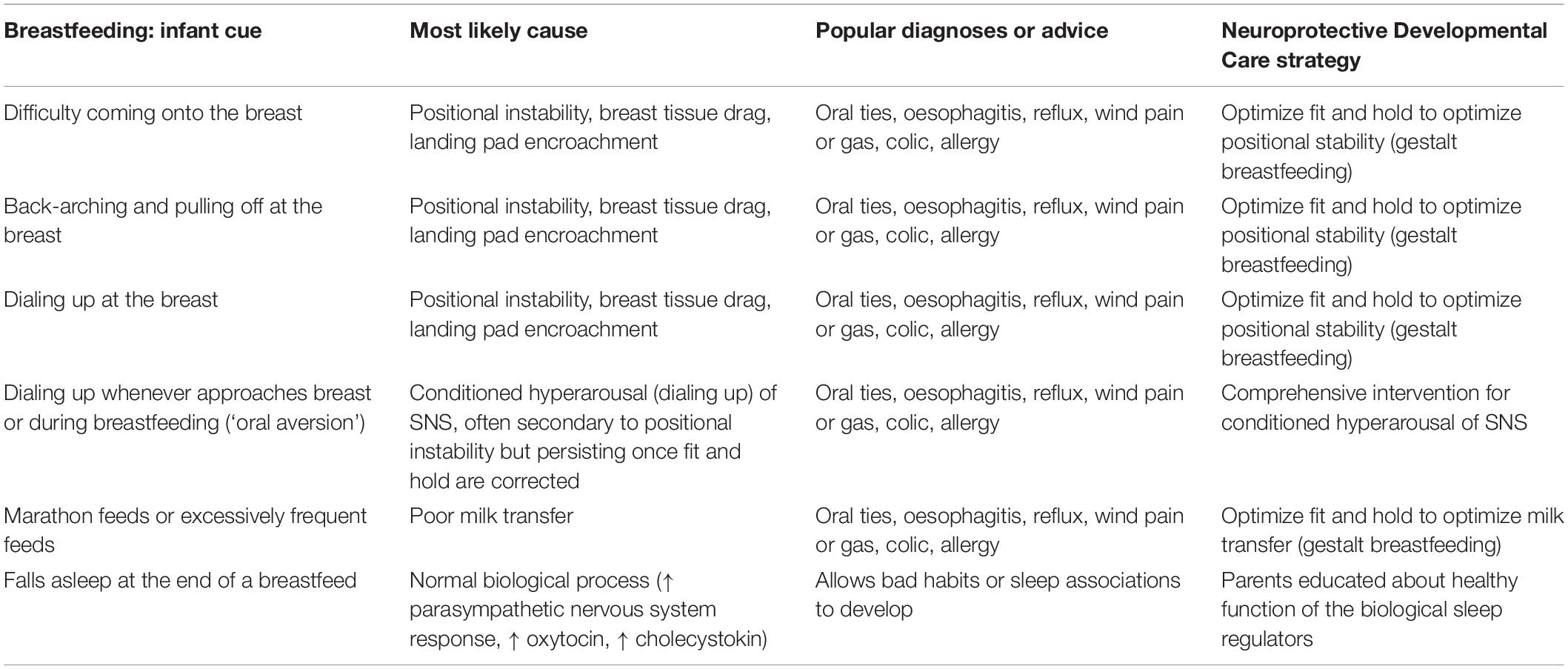
Table 4. Breastfeeding problems: popular sociocultural and clinical advice which disrupts parent-infant biobehavioral synchrony compared with Neuroprotective Developmental Care strategies which promote parent–infant biobehavioral synchrony.
Targeting Key Environmental Factors for Pre-Emptive Intervention
A prospective cohort study of 85,176 children in Norway shows that folic acid supplementation, taken by the mother from 4 weeks prior to 8 weeks after the first day of her last menstrual period, is associated with a lower risk of ASD. For this reason, folate supplementation from pre-conception is recommended as an initial step in pre-emptive intervention (Suren et al., 2013).
Pre-emptive intervention for ASD then aims to promote infant neuro-resilience by preventing, or re-stabilizing, disrupted feedback loops emerging from small motor and sensory-motor structural or functional lesions in very early life. Drawing on the preceding review of heterogeneous interdisciplinary research literature, the following will require optimization:
(1) Environmental enrichment, social and non-social (that is, ‘rich motor and sensory-motor nourishment’)
(2) Parent–infant biobehavioral synchrony, which is promoted by
(a) Cued care for downregulation of the SNS-HPA axis
(b) Multi-lateral strategies for management of cry-fuss problems
(c) Avoidance of inappropriate medicalization of infant behavior
(d) Healthy regulation of parent-infant sleep
(e) Increasingly long and complex, multi-sensory, affect-driven reciprocity chains, often infant-initiated
(f) Optimal parent mental health, which requires
(i) Effective intervention for the key modifiable risk factors of breastfeeding, sleep and cry-fuss problems
(ii) Strategies for managing difficult thoughts and feelings
(3) Gut microbiota and mitigation of pro-inflammatory states, promoted by
(a) Optimizing breastfeeding
(b) Teaching paced bottle-feeding
(c) Avoiding inappropriate medical diagnoses.
Over the past 20 years, robust peer-reviewed theoretical frames have been developed for a program known as Neuroprotective Developmental Care (NDC or ‘the Possums programs’), which targets these environmental factors as anticipatory guidance in the antenatal period and as clinical and educational support throughout the first 6–12 months of life. NDC has applied an integrative, primary care generalist’s lens to heterogenous evidence from the fields of neuroscience, evolutionary biology, developmental psychology, and the medical, lactation and sleep sciences (Douglas P. and Hill P., 2011; Douglas P.S. and Hill P.S., 2011; Whittingham and Douglas, 2014; Douglas and Keogh, 2017; Douglas and Geddes, 2018). NDC is a novel, community-based, parent-mediated, early life clinical and educational intervention. The term ‘Neuroprotective Family Centered Developmental Care’ has been previously applied to Heideleise Als pioneering work in the hospital setting with prematurely born infants, demonstrating improved development outcomes in this population (Als, 2009; Altimier and Phillips, 2016).
NDC has been delivered clinically in the Australian primary care setting since 2011 and is also available as online programs and for interdisciplinary health professional education.
Environmental Enrichment
This paper accepts the primacy of motor lesions in utero, intra-partum, or in very early life in ASD etiology. Pre-emptive intervention therefore advocates adult-mediated motor and sensory-motor environmental enrichment post-birth, both socially and non-socially, if early motor deficits are to be prevented or early disruption of feedback loops stabilized. Patterns of rich postural variability, with general movements of the infant’s limbs and trunk occurring against the adult’s changing bodily configurations (rather than in the context of long periods on an immobile cot or mattress), and patterns of rich multi-centric social interactions, that is, physically interactive play and enjoyment with older children and adults, are required to optimize sensory-motor feedback (Lickliter, 2011).
The NDC sensory domain is unique amongst infant-care programs in de-problematizing sensory stimulation, and offers parents multiple practical strategies for environmental enrichment, also referred to as healthy sensory-motor nourishment. Physical contact with the infant is encouraged where this is sensibly possible, which helps address the infant’s biological need for complex and continuing postural variation. NDC supports the primary carer to leave the low sensory environment of the Western interior daily for social contact, activities, and tasks, including frequent walks, as carers focuses on creating a rich, full and meaningful life, in large part outside the home, and to have the infant sleep day or night in the vicinity of the carer.
Optimizing Parent–Infant Biobehavioral Synchrony
The first step to optimal parent–infant biobehavioral synchrony is the elimination of clinical disruptors, which allows attention to the small communications of the infant to emerge. The second step is to encourage attention to the present moment and prioritize playful enjoyment of the baby.
Multi-Domain Management of Cry-Fuss Problems
NDC applies a systematic 5-domain approach to management of cry-fuss problems in parents, developed from the neurobiological model of cry-fuss problems (Douglas and Hill, 2013a). Each of the domains (feeds, sleep, environmental enrichment, infant gut and health considerations, and parental mood) integrate multiple evidence-based strategies. Preliminary evaluation of the 5-domain approach shows halved crying and fussing durations (Douglas et al., 2013). NDC applies a ‘dialing up’ and ‘dialing down’ metaphor, referring to up-regulation and down-regulation of the SNS-HPA, to describe parents’ co-regulatory efforts as they respond to their infant’s cues (Douglas and Hill, 2013a).
Healthy Regulation of the Infant’s Circadian Clock and Sleep–Wake Homeostat
The NDC sleep domain comprises the Possums Sleep Program, which enriches environmental stimulation, both social and non-social, by offering the only existing evidence-based alternative to FWB approaches for parent-infant sleep (Whittingham and Douglas, 2014; Ball et al., 2018; Douglas, 2018). Preliminary evaluation confirms the Possums Sleep Program has high levels of acceptability to parents and improves their quality of life (Ball et al., 2018). Parents are educated about maintenance of healthy function of their baby’s sleep regulators, and offered evidence-based information which minimizes anxiety about sleep. This includes:
(1) Educating parents about normal sleep variability and normal night-waking trajectories
(2) Differentiating between excessive night waking and normal night waking
(3) Identifying circadian clock disruption or satiety problems that result in unnecessary disruption of parent-baby sleep,
(4) Removing disruptors to sleep efficiency for parents and baby
(5) Education about how to optimize the function of the two biological sleep regulators, the circadian clock and the sleep–wake homeostat.
Strategies for Managing Difficult Thoughts and Feelings
NDC aims to change parents’ relationship with difficult thoughts and feelings in order to optimize enjoyment of the baby, which results in increasingly long and complex reciprocity chains of interaction. Traditional Cognitive Behavioral Therapy, which has been extensively investigated perinatally and is demonstrated as effective, promotes disputing irrational thoughts, which in the perinatal context may paradoxically worsen the biological predisposition to maternal rumination, precipitating worsened anxiety and depression (Monteiro et al., 2018). For this reason, NDC integrates applied functional contextualism, popularly known as Acceptance and Commitment Therapy (ACT), a third wave behaviorism. ACT has a burgeoning evidence-base for depression and anxiety, overtaking traditional Cognitive Behavioral Therapy on some indicators, with preliminary positive findings for parenting (Blackledge and Hayes, 2006; Ruiz, 2010; Whittingham et al., 2014).
ACT avoids unnecessary pathologizing of mental health challenges. It helps parents clarify values, and targets rumination, which predisposes mothers to perinatal anxiety and depression and which has been thought to mediate impaired infant development outcomes (DeJong et al., 2016). ACT offers a range of strategies for cognitive defusion and expansion of attention, in order to help parents manage difficult thoughts and feelings. ACT teaches behavioral activation skills, in the service of values, demonstrated to be effective with postnatal anxiety and depression (Lavender et al., 2016; Marchesi et al., 2016), and mindfulness, or anchoring in the present moment, demonstrated to assist in anxiety and depression generally (Gotink et al., 2015). ACT offers acceptance-focused processes and self-compassion, also demonstrated to help prevent postpartum depression and anxiety (Monteiro et al., 2018). A study of 139 parents of children with ASD showed that those who demonstrated more self-compassion experienced less stress and improved quality of life (Bohadana et al., 2019).
ACT is vitally concerned with context, and is well-suited for integration into health professionals’ consultations for common infantcare problems post-birth. ACT strategies are integrated into the Possums 5-domain approach to cry-fuss problems, the Possums Sleep Program, and gestalt breastfeeding (Whittingham and Douglas, 2016; Douglas and Keogh, 2017). Within these programs, women with moderate to severe mental illness are appropriately diagnosed, treated pharmaceutically as needed, and referred for ongoing psychological support.
‘Growing Joy in Early Life’
Unlike early intervention programs derived from social communication models, NDC does not instruct parents to avoid intrusiveness or directiveness, or offer suggestions about the meaning of infant cues and how to respond, which may inadvertently communicate assumptions of parental incompetence and may also increase parental anxiety. Similarly, NDC also does not employ tools such as the Neonatal Observation Scale to teach parents about their infant’s behavioral repertoire and communication competence in the first six months of life (Nugent, 2013). Instead, NDC educates parents about the benefits of reciprocity chains, and encourages enjoyment of the baby. NDC proposes that parental competence and evolutionary drive for enjoyment of the baby will emerge in families once disruptive sociocultural and clinical advice are removed, underlying clinical problems are identified and repaired, and the importance of satisfying and socially engaged days outside the home explained.
NDC confidence in parental competence is corroborated by a study of 864 parent–newborn pairs observed spending time together as the baby lay close to the parent. Parents were given minimal instructions, but asked to interact with the baby comfortably, as they saw fit. Most of the 480 full-term newborns showed subtle affect-driven initiation of arm movements toward the parents as they interacted, though this was somewhat reduced in the prematurely born infants, and all parents engaged in quiet and supportive interaction without being intrusive (Delafield-Butt et al., 2018).
Optimizing Breastfeeding and Enjoyment of Feeds
Optimizing gut health in susceptible infants requires not only avoidance of unnecessary medicalization of unsettled infant behavior, but importantly, the support of breastfeeding success, with its known gut and immune protections.
A systematic review of ultrasound studies of breastfeeding mother-baby pairs forms the biomechanical basis for a new ‘gestalt’ model of clinical breastfeeding support, out of which the NDC gestalt breastfeeding approach is developed (Douglas and Geddes, 2018). Gestalt breastfeeding builds on the pioneering advance of baby-led breastfeeding, which emphasizes cued care by activating and responding to the baby’s mammalian reflexes, but points to the evidence showing that this is not enough to achieve pain-free effective milk transfer for many women (Svensson et al., 2013; Woods et al., 2016). The gestalt approach educates about the biomechanics of effective breastfeeding. Impaired fit and hold and the resultant infant cues of fussing at the breast, difficulty coming onto the breast, back-arching, maternal nipple pain, marathon feeds, excessively frequent feeds, excessive night waking, and poor weight gain, are addressed applying gestalt methods (Douglas and Keogh, 2017; Douglas and Geddes, 2018).
Formula, which offers the human infant unphysiological doses of cow’s milk protein, lacks a myriad immune factors, micro-nutrients, and live bacteria, predisposing to short- and long-term gut dysbiosis and increased risk of immune system alterations. DNA methylation, a key mechanism of epigenomic regulation, is highly responsive to diet. Nutrients and bioactive compounds are known to alter the expression of genes at the transcriptional level and result in long-term phenotypic changes (Barker et al., 2018).
However, women feed babies in complex contemporary sociocultural contexts, facing multiple disruptors, pain, and conflicting clinical advice. NDC is strictly non-judgmental concerning mode of infant feeding, and teaches paced bottle-feeding in order to optimize feeding-related reciprocity chains in formula or expressed-breastmilk-fed infants. NDC also identifies and manages conditioned dialing up with the bottle, which is commonly inappropriately medicalized (Table 5).
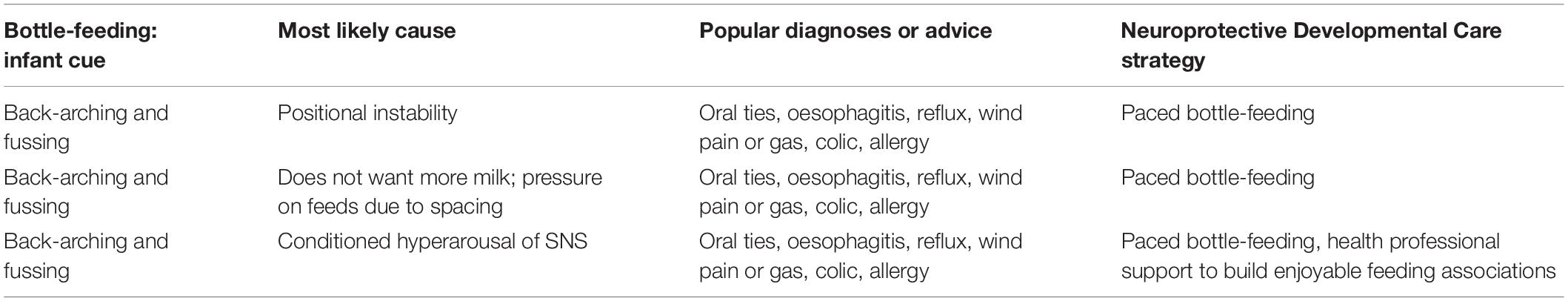
Table 5. Bottle-feeding problems: popular sociocultural and clinical advice which disrupts parent–infant biobehavioral synchrony compared with Neuroprotective Developmental Care strategies which promote parent–infant biobehavioral synchrony.
Conclusion
ASDs are complex polygenic syndromes displaying highly variable phenotypic expressions and co-morbidities. Myriad environmental factors are known to modulate epigenomic regulation of ASD phenotypic expression. By 6 months of age, when the first signs of atypical connectivity emerge in neuroimaging studies, neuropathy may be entrenched. This paper responds to a recent call from the International Society for Autism Research for the development of theoretical foundations for, and clinical translation of, pre-emptive intervention for ASD.
The first 100 days of human life are characterized by injury-sensitive neuroplasticity, correlating with in the neural biomarker of cortical subplate persistence and the behavioral biomarker of the crying diathesis. Environmental factors in these critically injury-sensitive first 100 days may either initiate or perpetuate structural or functional lesions in global neuronal workspace of susceptible infants. This paper builds on ground-breaking etiological models concerning the primacy of motor lesions in ASD etiology, to propose that chronic SNS-HPA hyperarousal and disrupted parent-infant biobehavioral synchrony are key mechanisms facilitating perpetuation of the atypical developmental trajectories of ASD, triggered by very early motor and sensory-motor lesions.
Existing clinical guidelines for protection of infant growth and development in very early life, including support for feeds and sleep, are confused and conflicting, exacerbating parental anxiety. From an evolutionary perspective, popular clinical and cultural advice often promotes biology-culture mismatch, resulting in chronic SNS-HPA hyperarousal and disrupted parent–infant biobehavioral synchrony. Health professionals are not adequately trained in the identification and management of common clinical problems arising in the domains of feeds, sleep, cry-fuss problems and parental mood, which disrupt parent–infant biobehavioral synchrony. This health system context places ASD susceptible infants, and infants susceptible to other neurodevelopmental disorders, at risk.
Any pre-emptive intervention for ASD that aims to apply a focused selection of ‘active ingredients’ risks exacerbating parental anxiety, because parents already receive conflicting and confusing advice concerning common infant-care problems. Contact with these providers will continue. This paper argues that a comprehensive, holistic, community-based approach to the care of at-risk siblings is necessary to support parents who must inevitably navigate the health system, and proposes an Australian program known as NDC.
Research funding for evaluation of primary care initiatives remains scarce and is diminishing. Evaluation of NDC as pre-emptive intervention for ASD (Box 4), using real world methodologies suitable for complex community-based interventions, is required.
BOX 4. When to apply NDC as pre-emptive intervention.
Families with the following risk factors for ASD should be offered NDC pre-emptive intervention as soon as possible, beginning with anticipatory antenatal education and throughout the first 6 months of life:
• Child diagnosed with ASD
• Parent diagnosed with ASD.
Further potential indicators for NDC pre-emptive intervention are perinatal factors associated with later diagnosis of ASD, including:
• Antenatal depression
• Pre-eclampsia
• Premature birth
• Caesarian section under general anesthesia
• Infant hypoxic-ischemic encephalopathy (Chien and Lin, 2015; Getahun et al., 2017).
The following factors that have been associated with infant neurodevelopmental vulnerability also warrant NDC as pre-emptive intervention:
• Unsettled infant behavior
• Atypical crying acoustics
• Breastfeeding or feeding difficulties
• Maternal postnatal depression
• Plagiocephaly (marker of increased risk of transient motor and developmental delay, suggesting need for enriched sensory-motor experience)
• Other signs of developmental vulnerability (Martiniuk et al., 2017; Renz-Poster and De Bock, 2018)
If efficacy of NDC is demonstrated in infants at risk, a second phase would evaluate NDC as a health system response, replacing current conflicting and confusing advice from multiple health professionals in order to offer consistent, evidence-based care which optimizes neuroprotection for all parents and infants from the antenatal period onward, universally available through usual maternal and child health services.
Author Contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Conflict of Interest
PD is the medical director of a small primary care charity, Possums Education, which sells Neuroprotective Developmental Care programs online for parents, and also as education workshops for health professionals www.possumsonline.com. All revenue raised is returned into the development of education and research.
Abbreviations
ASD, Autism spectrum disorder/s; FWB, First wave behaviorism/first wave behavioral; PNS, Parasympathetic nervous system; NDC, Neuroprotective Developmental Care; SNS, Sympathetic nervous system; SNS-HPA, Sympathetic nervous system and hypothalamic-pituitary-adrenal axis
References
Akhtar, N., Jaswal, V. K., Dinishak, J., and Stephan, C. (2016). On social feedback loops and cascading effects in autism: a commentary on Warlaumont, Richards, Gilkerson, and Oller (2014). Psychol. Sci. 27, 1528–1530. doi: 10.1177/0956797616647520
Als, H. (2009). Newborn individualized developmental care and assessment program (NIDCAP): new frontier for neonatal and perinatal medicine. J. Neonatal Perinatal Med. 2, 135–147.
Altimier, L., and Phillips, R. (2016). The neonatal integrative developmental care model: advanced clinical applications of the seven core measures for neuroprotective family-centred developmental care. Newborn Infant Nurs. Rev. 16, 230–244. doi: 10.1053/j.nainr.2016.09.030
Amaral, D. G., Andreson, G. M., Bailey, A., Bernie, R., Bishop, S., Blatt, G., et al. (2019). Gaps in current autism research: the thoughts of the autism research editorial board and associate editors. Autism Res. 12, 700–714. doi: 10.1002/aur.2101
Ayano, G., Maravilla, J. C., and Alati, R. (2019). Risk of autistic spectrum disorder in offspring with parental mood disorders: a systematic review and meta-analysis. J. Affect. Disord. 248, 185–197. doi: 10.1016/j.jad.2019.01.038
Bag, O., Guney, S. A., Binici, N. C., Tuncel, T., Sahin, A., Berksoy, E., et al. (2018). Infant colic or early symptom of autism spectrum disorder? Pediatr. Int. 60, 517–522. doi: 10.1111/ped.13565
Baker, E. K., Richdale, A. L., Hazi, A., and Prendergast, L. A. (2019). Assessing a hyperarousal hypothesis of insomnia in adults with autism spectrum disorder. Autism Res. 12, 897–910. doi: 10.1002/aur.2094
Baker, J. K., Fenning, R. M., Howland, M. A., Baucom, B. R., Moffitt, J., and Erath, S. A. (2015). Brief report: a pilot study of parent-child biobehavioral synchrony in autism spectrum disorder. J. Autism Dev. Disord. 45, 4140–4146. doi: 10.1007/s10803-015-2528-0
Ball, H., Douglas, P. S., Whittingham, K., Kulasinghe, K., and Hill, P. S. (2018). The Possums Infant Sleep Program: parents’ perspectives on a novel parent-infant sleep intervention in Australia. Sleep Health 4, 519–526. doi: 10.1016/j.sleh.2018.08.007
Ball, H. L. (2008). “Evolutionary paediatrics: a case study in applying Darwinian medicine,” in Medicine and Evolution: Current Applications, Future Prospects, eds S. Elton, and P. O’Higgens, (Boca Raton, FL: CRC Press), 127–152. doi: 10.1201/9781420051377.ch7
Baranek, G. T., Watson, L. R., Turner-Brown, L., Field, S. H., Crais, E. R., Wakeford, L., et al. (2015). Preliminary efficacy of adapted responsive teaching for infants at risk of autism spectrum disorder in a community sample. Autism Res. Treat. 2015:386951. doi: 10.1155/2015/386951
Barker, E. D., Walton, E., and Cecil, C. A. M. (2018). Annual Research Review: DNA methylation as a mediator in the association betwen risk exposure and child and adolescent psychopathology. J. Child Psychol. Psychiatry 59, 303–322. doi: 10.1111/jcpp.12782
Barnevik, O. M., Carlsson, L., Westerlund, J., Gillberg, C., and Fernell, E. (2013). Autism before diagnosis: crying, feeding and sleeping problems in the first two years of life. Acta Paediatr. 102, 635–639. doi: 10.1111/apa.12229
Barr, R. G. (1990). The early crying paradox: a modest proposal. Hum. Nat. 1, 355–389. doi: 10.1007/BF02734051
Barr, R. G. (1999). “Infant crying behavior and colic: an interpretation in evolutionary perspective,” in Evolutionary Medicine, eds W. R. Trevathan, E. O. Smith, and J. J. McKenna, (New York, NY: Oxford University Press), 28–51.
Barr, R. G., Konner, M., Bakeman, R., and Adamson, L. (1991). Crying in !Kung San infants: a test of the cultural specificity hypothesis. Dev. Med. Child Neurol. 33, 601–610. doi: 10.1111/j.1469-8749.1991.tb14930.x
Bauml, J. G., Baumann, N., Avram, M., Bratec, S. M., Breeman, L. D., Berndt, M., et al. (2018). The default mode network mediates the impact of infant regulatory problems on adult avoidant personality traits. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 4, 333–342. doi: 10.1016/j.bpsc.2018.11.005
Belfort, M. B. (2017). The science of breastfeeding and brain development. Breastfeed. Med. 12, 459–461. doi: 10.1089/bfm.2017.0122
Bell, G., Hiscock, H., Tobin, S., Cook, F., and Sung, V. (2018). Behavioral outcomes of infant colic in toddlerhood: a longitudinal study. J. Pedatr. 201, 154–159. doi: 10.1016/j.jpeds.2018.05.010
Bergmann, M. M., Caubet, J.-C., McLin, V., Belli, D., Schappi, M., and Eigenmann, P. (2014). Common colic, gastroesophageal reflux and constipation in infants under 6 months do not necessitate an allergy work-up. Pediatr. Allergy Immunol. 25, 410–412. doi: 10.1111/pai.12199
Bertin, E. (2006). The still-face response in newborn, 1.5-, and 3-month-old infants. Infant Behav. Dev. 29, 294–297. doi: 10.1016/j.infbeh.2005.12.003
Bilgin, A., Baumann, N., Jaekel, J., Breeman, L. D., Bartmann, P., Bauml, J. G., et al. (2018). Early crying, sleeping, and feeding problems and trajectories of attention problems from childhood to adulthood. Child Dev. doi: 10.1111/cdev.13155 [Epub ahead of print].
Bilgin, A., and Wolke, D. (2017). Development of comorbid crying, sleeping, feeding problems across infancy: neurodevelopmental vulnerability and parenting. Early Hum. Dev. 109, 37–43. doi: 10.1016/j.earlhumdev.2017.04.002
Bittker, S. S., and Bell, K. R. (2018). Acetaminophen, antibiotics, ear infection, breastfeeding, vitamin D drops, and autism: an epidemiological study. Neuropsychiatr. Dis. Treat. 14, 1399–1414. doi: 10.2147/NDT.S158811
Blackledge, J. T., and Hayes, S. C. (2006). Using acceptance and commitment training in support of parents of children diagnosed with autism. Child Family Behav. Ther. 28, 1–18. doi: 10.1300/j019v28n01_01
Blunden, S., Etherton, H., and Hauck, Y. (2016). Resistance to cry intensive sleep intervention in young children: are we ignoring children’s cries or parental concerns? Children 3:8. doi: 10.3390/children3020008
Bohadana, G., Morrissey, S., and Paynter, J. (2019). Self-compassion: a novel predictor of stress and quality of life in parents of children with autism spectrum disorder. J. Autism Dev. Disord. 49, 4039–4052. doi: 10.1007/s10803-019-04121-x
Bolton, P., Golding, J., Emond, A., and Steer, C. (2012). Autism spectrum disorder and autistic traits in the Avon Longitudinal Study of Parents and Children: precursors and early signs. J. Am. Acad. Child Adoles. Psychiatry 51, 249–260. doi: 10.1016/j.jaac.2011.12.009
Boucher, O., Julvez, J., Guxens, M., Arranz, E., Ibarluzea, J., Sanchez de Miguel, M., et al. (2017). Association between breastfeeding duration and cognitive development, autistic traits and ADHD symptoms: a multicenter study in Spain. Pediatr. Res. 81, 434–442. doi: 10.1038/pr.2016.238
Bradshaw, J., Klaiman, C., Gillespie, S., Brane, N., Lewis, M., and Saulnier, C. (2018). Walking ability is associated with social communication skills in infants at high risk for autism spectrum disorder. Infancy 23, 674–691. doi: 10.1111/infa.12242
Bradshaw, J., Klin, A., Evans, L., Klaiman, C., Saulnier, C., and McCracken, C. (2019). Development of attention from birth to 5 months in infants at risk for autism spectrum disorder. Dev. Psychopathol. doi: 10.1017/S0954579419000233 [Epub ahead of print].
Bradshaw, J., Steiner, A. M., Gengoux, G., and Koegel, L. K. (2015). Feasibility and effectiveness of very early intervention for infants at-risk for autism spectrum disorder: a systematic review. J. Autism Dev. Disord. 45, 778–794. doi: 10.1007/s10803-014-2235-2
Braithwaite, J., Churruca, K., Long, J. C., Louise, E. A., and Herkes, J. (2018). When complexity science meets implementation science: a theoretical and empirical analysis of systems change. BMC Med. 16:63. doi: 10.1186/s12916-018-1057-z
Breeman, L. D., Jaekel, J., Baumann, N., Bartmann, P., Bauml, J. G., Avram, M., et al. (2018). Infant regulatory problems, parenting quality and childhood attention problems. Early Hum. Dev. 124, 11–16. doi: 10.1016/j.earlhumdev.2018.07.009
Britto, P. R., Lye, S. J., Proulx, K., and Yousafzai, A. (2017). Nurturing care: promoting early childhood development. Lancet 389, 91–102. doi: 10.1016/S0140-6736(16)31390-3
Brown, A., and Arnott, B. (2014). Breastfeeding duration and early parenting behaviour: the importance of an infant-led, responsive style. PLoS One 9:e83893. doi: 10.1371/journal.pone.0083893
Brown, A., and Lee, M. (2013). Breastfeeding is associated with a maternal feeding style low in control from birth. PLoS One 8:e54229. doi: 10.1371/journal.pone.0054229
Brown, A., Rance, J., and Bennett, P. (2015). Understanding the relationship between breastfeeding and postnatal depression: the role of pain and physical difficulties. J. Adv. Nursing 72, 273–282. doi: 10.1111/jan.12832
Brownlee, S., Chalkidou, K., Doust, J., Elshaug, A. G., Glasziou, P., Heath, I., et al. (2017). Evidence for overuse of medical services around the world. Lancet 390, 156–168. doi: 10.1016/S0140-6736(16)32585-5
Bryanton, J., Beck, C., and Montelpare, W. (2013). Postnatal parental education for optimizing infant general health and parent-infant relationships. Cochrane Database Syst. Rev. 11:CD004068.
Buescher, A. V., Cidav, Z., Knapp, M., and Mandell, D. S. (2014). Costs of autism spectrum disorders in the United Kingdom and the United States. JAMA Pediatr. 168, 721–728. doi: 10.1001/jamapediatrics.2014.210
Campbell, N., Murray, E., Darbyshire, J., Emery, J., Farmer, A., Griffiths, F., et al. (2007). Designing and evaluating complex interventions to improve health care. Br. Med. J. 334, 455–459. doi: 10.1136/bmj.39108.379965.be
Carter, C. J. (2019). Autism genes and the leukocyte transcriptome in autistic toddlers relate to pathogen interactomes, infection and the immune system. Neurochem. Int. 126, 36–58. doi: 10.1016/j.neuint.2019.03.007
Castellani, C., Singer, G., Kashofer, K., Huber-Zeyringer, A., Flucher, C., Kaiser, M., et al. (2017). The influence of proton pump inhibitors on the fecal microbiome of infants with gastroesophageal reflux - a prospective longitudinal interventional study. Front. Cell. Infect. Microbiol. 111:444.
Cenit, M. C., Sanz, Y., and Codoner-Freanch, P. (2017). Influence of gut microbiota on neuropsychiatric disorders. World J. Gastroenterol. 23, 5486–5498. doi: 10.3748/wjg.v23.i30.5486
Chien, L.-N., and Lin, H.-C. (2015). Risk of Autism associated with general anesthesia during cesarian delivery: a population-based birth-cohort analysis. J. Autism Dev. Disord. 45, 932–942. doi: 10.1007/s10803-014-2247-y
Cohen, S., Russell, C., Lockley, S. W., Rajaratnam, S., and Cornish, K. M. (2014). The relationship between sleep and behavior in autism spectrum disorder (ASD): a review. J. Neurodeve. Disord. 6:44. doi: 10.1186/1866-1955-6-44
Cook, F., Giallo, R., Hiscock, H., Mensah, F. K., Sanchez, K., and Reilly, S. (2019). Infant regulation and child mental health concerns: a longitudinal study. Pediatics 143:e20180977.
Cook, F., Giallo, R., Petrovic, Z., Coe, A., Seymour, M., Cann, W., et al. (2017). Depression and anger in fathers of unsettled infants: a community cohort study. J. Paediatr. Child Health. 53, 131–135. doi: 10.1111/jpc.13311
Coon, E. R., Quinonez, R. A., Moyer, V. A., and Schroeder, A. R. (2014). Overdiagnosis: how our compulsion for diagnosis may be harming children. Pediatrics 134, 1–11. doi: 10.1542/peds.2014-1778
Coretti, L., Paparo, L., Riccio, M. P., Amato, F., Cuomo, M., and Natale, A. (2018). Gut microbiota features in young children with autism spectrum disorders. Front Microbiol. 9:3146.
Dadalko, O. I., and Travers, B. G. (2018). Evidence for brainstem contributions to autism spectrum disorders. Front. Integr. Neurosci. 12:47. doi: 10.3389/fnint.2018.00047
De Graaf-Peters, V. B., and Hadders-Algra, M. (2006). Ontogeny of the human central nervous system: what is happening when? Early Hum. Dev. 82, 257–266. doi: 10.1016/j.earlhumdev.2005.10.013
De Palma, G., Collins, S. M., Bercik, P., and Verdu, E. F. (2014). The microbiota-gut-brain axis in gastrointestinal disorders: stressed bugs, stressed brain or both? J. Physiol. 592, 2989–2997. doi: 10.1113/jphysiol.2014.273995
De Rubeis, S., and Buxbaum, J. D. (2015). Genetics and genomics of autism spectrum disorder: embracing complexity. Hum. Mol. Genet. 24, R24–R31. doi: 10.1093/hmg/ddv273
DeGangi, G. A., Breinbauer, C., Roosevelt, J. D., Porges, S., and Greenspan, S. (2000). Prediction of childhood problems at three years in children experiencing disorders of regulation during infancy. infant. Men. Health J. 21, 156–175. doi: 10.1002/1097-0355(200007)21:3<156::aid-imhj2>3.0.co;2-d
DeJong, H., Fox, E., and Stein, A. (2016). Rumination and postnatal depression: A systematic review and a cognitive model. Behav. Res. Ther. 82, 38–49. doi: 10.1016/j.brat.2016.05.003
Delafield-Butt, J. T., Freer, Y., Perkins, J., Skulina, D., Schogler, B., and Lee, D. N. (2018). Prospective organization of neonatal arm movements: a motor foundation of embodied agency, disrupted in premature birth. Dev. Sci. 21:e12693. doi: 10.1111/desc.12693
Delafield-Butt, J. T., and Trevarthan, C. (2017). “On the brainstem origin of autism: disruption to movements of the primary self,” in Autism: the Movement Sensing Perspective, eds E. B. Torres, and C. Whyatt, (Boca Raton, FL: Taylor and Francis CRC Press).
Denisova, K., and Zhao, G. (2017). Inflexible neurobiological signatures precede atypical development in infants at high risk for autism. Sci. Rep. 7:11285. doi: 10.1038/s41598-017-09028-0
Desantis, A., Coster, W., Bigsby, R., and Lester, B. (2004). Colic and fussing in infancy, and sensory processing at 3 to 8 years of age. Infant Men. Health J. 25, 522–539. doi: 10.1002/imhj.20025
Dias, C. C., and Figueiredo, B. (2015). Breastfeeding and depression: a systematic review of the literature. J. Affect. Disord. 171, 142–154. doi: 10.1016/j.jad.2014.09.022
Ding, H. T., Taur, Y., and Walkup, J. T. (2017). Gut microbiota and autism: key concepts and findings. J. Autism Dev. Disord. 2017, 480–489. doi: 10.1007/s10803-016-2960-9
Dissanayake, C., Searles, J., Barbaro, J., Sadka, N., and Lawson, L. P. (2019). Cognitive and behavioral differences in toddlers with autism spectrum disorder from multiplex and simplex families. Autism Res. 12, 682–693. doi: 10.1002/aur.2074
Dong, D., Zielke, H. R., Yeh, D., and Yang, P. (2018). Cellular stress and apoptosis contribute to the pathogenesis of autism mspectrum disorder. Autism Res. 11, 1076–1090. doi: 10.1002/aur.1966
Dorheim, S. K., Bondevik, G. T., Eberhard-Gran, M., and Bjorvatn, B. (2009). Sleep and depression in postpartum women: a population-based study. Sleep 32, 847–855. doi: 10.1093/sleep/32.7.847
Douglas, P. (2013). Diagnosing gastro-oesophageal reflux disease or lactose intolerance in babies who cry alot in the first few months overlooks feeding problems. J. Paediatr. Child Health 49, e252–e256. doi: 10.1111/jpc.12153
Douglas, P., and Hill, P. (2011). Managing infants who cry excessively in the first few months of life. BMJ 343:d7772. doi: 10.1136/bmj.d7772
Douglas, P. S., and Hill, P. S. (2013a). A neurobiological model for cry-fuss problems in the first three to four months of life. Med. Hypotheses 81, 816–822. doi: 10.1016/j.mehy.2013.09.004
Douglas, P., and Hill, P. S. (2013b). Behavioural sleep interventions in the first six months of life do not improve outcomes for mothers or infants: a systematic review. J. Dev. Behav. Pediatr. 34, 497–507. doi: 10.1097/dbp.0b013e31829cafa6
Douglas, P., Miller, Y., Bucetti, A., Hill, P. S., and Creedy, D. (2013). Preliminary evaluation of a primary care intervention for cry-fuss behaviours in the first three to four months of life (”The Possums Approach”): effects on cry-fuss behaviours and maternal mood. Aust. J. Primary Health. 21, 38–45.
Douglas, P. S. (2005). Excessive crying and gastro-oesophageal reflux disease in infants: misalignment of biology and culture. Med. Hypotheses 64, 887–898. doi: 10.1016/j.mehy.2004.12.009
Douglas, P. S. (2018). The Possums Sleep Program: supporting easy, healthy parent-infant sleep. Int. J. Birth Parent Educ. 6, 13–16.
Douglas, P. S., and Geddes, D. B. (2018). Practice-based interpretation of ultrasound studies leads the way to less pharmaceutical and surgical intervention for breastfeeding babies and more effective clinical support. Midwifery 58, 145–155. doi: 10.1016/j.midw.2017.12.007
Douglas, P. S., and Hill, P. S. (2011). The crying baby: what approach? Curr. Opin. Pediatr. 23, 523–529. doi: 10.1097/MOP.0b013e32834a1b78
Douglas, P. S., Hill, P. S., and Brodribb, W. (2011). The unsettled baby: how complexity science helps. Arch. Dis. Child 96, 793–797. doi: 10.1136/adc.2010.199190
Douglas, P. S., and Keogh, R. (2017). Gestalt breastfeeding: helping mothers and infants optimise positional stability and intra-oral breast tissue volume for effective, pain-free milk transfer. J. Hum. Lact. 33, 509–518. doi: 10.1177/0890334417707958
Dusing, S. C. (2016). Postural variability and sensorimotor development in infancy. Dev. Med. Child Neurol. 58, (Suppl. 4), 17–21. doi: 10.1111/dmcn.13045
Elison, J. T., Wolff, J. J., Reznick, J. S., Botteron, K. N., Estes, A. M., Gu, H., et al. (2014). Repetitive behavior in 12-month-olds later classified with autism spectrum disorder. J. Am. Acad. Child Adolesc. Psychiatry 53, 1216–1224. doi: 10.1016/j.jaac.2014.08.004
English, M. S., Tenenbaum, E. J., Levine, T. P., Lester, T. P., and Sheinkopf, S. J. (2018). Perception of cry characteristics in 1-month old infants later diagnosed with autism spectrum disorder. J. Autism Dev. Disord. 49, 834–844. doi: 10.1007/s10803-018-3788-2
Esposito, G., Hiroi, N., and Scattoni, M. L. (2017). Cry, baby, cry: expression of distress as a biomarker and modulator in autism spectrum disorder. Int. J. of Neuropsychopharmacol. 20, 498–503. doi: 10.1093/ijnp/pyx014
Estes, A., Zwaigenbaum, L., Gu, H., St. John, T., Paterson, S., Elison, J. T., et al. (2015). Behavioral, cognitive, and adaptive development in infants with autism spectrum disorder in the first 2 years of life. J. Neurodev. Disord. 7:24. doi: 10.1186/s11689-015-9117-6
Estrem, H. H., Pados, B. F., Park, J., Knafl, K. A., and Thoyre, S. M. (2016). Feeding problems in infancy and early childhood: evolutionary concept analysis. J. Adv. Nurs 73, 56–70. doi: 10.1111/jan.13140
Etherton, H., Blunden, S., and Hauck, Y. (2016). Discussion of extinction-based behavioral sleep interventions for young children and reasons why parents may find them difficult. J. Clin. Sleep Med. 12, 1535–1543. doi: 10.5664/jcsm.6284
Fatheree, N. Y., Liu, Y., and Taylor, C. M. (2017). Lactobacillus reuteri for infants with colic: a double-blind, placebo-controlled, randomized clinical trial. J. Pediatr. 191, 170–178.
Feldman, R. (2007). Parent-infant synchrony and the construction of shared timing: physiological precursors, developmental outcomes, and risk conditions. J. Child Psychol. Psychiatry 48, 329–354. doi: 10.1111/j.1469-7610.2006.01701.x
Feldman, R. (2016). The neurobiology of mammalian parenting and the biosocial context of human caregiving. Horm. Behav. 77, 3–17. doi: 10.1016/j.yhbeh.2015.10.001
Fields, C., and Glazebrook, J. F. (2017). Disrupted development and imbalanced function in the global neuronal workspace: a positive-feedback mechanism for the emergence of ASD in early infancy. Cogn. Neurodyn. 11, 1–21. doi: 10.1007/s11571-016-9419-8
Fogel, A., King, B. J., and Shanker, S. G. (2008). Human Development in the Twenty-First Century: Visionary Ideas From Systems Scientists. New York, NY: Cambridge University Press.
Fox, A., Gardner, G., and Osborne, S. (2014). A theoretical framework to support research of health service innovation. Aust. Health Rev. 39, 70–75. doi: 10.1071/AH1403
French, L., and Kennedy, E. M. M. (2018). Annual research review: early intervention for infants and young children with, or at-risk of, autism spectrum disorder: a systematic review. J. Child Psychol. Psychiatry 59, 444–456. doi: 10.1111/jcpp.12828
Garcia-Cabezas, M. A., Barbas, H., and Zikopoulos, B. (2018). Parallel development of chromatin patterns, neuron morphology, and connections: potential for disruption in autism. Front. Neuroanat. 12:70. doi: 10.3389/fnana.2018.00070
Gavine, A., MacGillivray, S., Renfew, M. J., Siebelt, L., Haggi, H., and McFadden, A. (2017). Education and training of healthcare staff in the knowledge, attitudes and skills needed to work effectively with breastfeeding women: a systematic review. Int. Breastfeed. J. 12:6. doi: 10.1186/s13006-13016-10097-13002
Getahun, D., Fassett, M. J., Peltier, M. R., Wing, D. A., Xiang, A. H., Chiu, V., et al. (2017). Association of perinatal risk factors with autism spectrum disorder. Am. J. Perinatol. 34, 295–304. doi: 10.1055/s-0036-1597624
Gettler, L. T., and McKenna, J. J. (2011). Evolutionary perspectives on mother-infant sleep proximity and breastfeeding in a laboratory setting. Am. J. Phys. Anthropol. 144, 454–462. doi: 10.1002/ajpa.21426
Gieruszczak-Bialek, D., Konarska, Z., Skorka, A., Vandenplas, Y., and Szajewska, H. (2015). No effect of proton pump inhibitors on crying and irritability in infants: systematic review of randomized controlled trials. J. Pediatr. 166, 767–770. doi: 10.1016/j.jpeds.2014.11.030
Goldani, A. A. S., Downs, S. R., Widjaja, F., Lawton, B., and Hendren, R. L. (2014). Biomarkers in autism. Front. Psychiatry 5:100. doi: 10.3389/fpsyt.2014.00100
Gordon, M., Biagioli, E., Sorrenti, M., Lingua, C., Moja, L., Banks, S. S., et al. (2018). Dietary modifications for infantile colic. Cochrane Database Syst. Rev. 10, CD011029. doi: 10.1002/14651858.CD14601129.pub14651852
Gotink, R. A., Chu, P., Busschbach, J. J. V., Benson, H., Fricchione, G. L. M., and Hunink, M. G. (2015). Standardised mindfulness-based interventions in healthcare: an overview of systematic reviews and meta-analyses of RCTs. Plos One 10:e0124344. doi: 10.1371/journal.pone.0124344
Gottfried, C., Bambini-Junior, V., Francis, F., Riesgo, R., and Savino, W. (2015). The impact of neuroimmune alterations in autism spectrum disorder. Hypothesis Theory. 6:121. doi: 10.3389/fpsyt.2015.00121
Green, J. (2017). Editorial: basic science and treatment innovation. J. Child Psychol. Psychiatry 58, 967–969. doi: 10.1111/jcpp.12797
Green, J. (2019). “Intervention during the prodromal stages of ASD,” in Infants With Autism, eds F. Volkmar, and K. Chawarska, (New York: Guilford Press).
Green, J., Charman, T., Pickles, A., Wan, M. W., Elsabagh, M., Slonims, V., et al. (2015). Parent-mediated intervention versus no interventon for infants at high risk of autism: a parallel, single-blind, randomised trial. Lancet 2, 133–140. doi: 10.1016/S2215-0366(14)00091-1
Green, J., and Garg, S. (2018). Annual research review: the state of autism intervention science: progress, target psychological and biological mechanisms and future prospects. J. Child Psychol. Psychiatry 59, 424–443. doi: 10.1111/jcpp.12892
Green, J., Pickles, A., Pasco, G., Bedford, R., Wan, M. W., Elsabbagh, M., et al. (2017). Randomised trial of a parent-mediated intervention for infants at high risk for autism: longitudinal outcomes to age 3 years. J. Child Psychol. Psychiatry 58, 1330–1340. doi: 10.111/jcpp
Greenhalgh, T., Wong, G., Jagosh, J., Greenhalgh, J., Manzano, A., Westhorp, G., et al. (2015). Protocol - the REMESES II study: developing guidance and reporting standards for realist evaluation. BMJ Open 5:e008567. doi: 10.1136/bmjopen-2015-008567
Greenspan, N. S. (2018). Autism, evolution, and the inadequacy of ‘spectrum’. Evol. Med. Public Health. 1, 213–218. doi: 10.1093/emph/eoy025
Greenspan, S. (2002). The Affect Diathesis Hypothesis: the role of emotions in the core deficit in autism and in the development of intelligence and social skills. J. Dev. Learn. Disord. 5, 1–44.
Hadders-Algra, M. (2007). Putative neural substrate of normal and abnormal general movements. Neurosci. Biobehav. Rev. 31, 1181–1190. doi: 10.1016/j.neubiorev.2007.04.009
Hadders-Algra, M. (2010). Variation and variability: key words in human motor development. Physical. Ther. 90, 1823–1837. doi: 10.2522/ptj.20100006
Hadders-Algra, M. (2017). Neural substrate and clinical significance of general movements: an update. Dev. Med. Child Neurol. 60, 39–46. doi: 10.1111/dmcn.13540
Hadders-Algra, M. (2018a). Early human brain development: starring the subplate. Neurosci. Biobehav. Rev. 92, 276–290. doi: 10.1016/j.neubiorev.2018.06.017
Hadders-Algra, M. (2018b). Early human motor development: from variation to the ability to vary and adapt. Neurosci. Biobehav. Rev. 90, 411–427. doi: 10.1016/j.neubiorev.2018.05.009
Hamilton, A. (1981). Nature and Nurture: Aboriginal Childrearing in North-Central Arnhem Land. Canberra: AIAS Press.
Heineman, K. R., Schendelaar, P., Van Den Heuvel, E. R., and Hadders-Algra, M. (2018). Motor development in infancy is related to cognitive function at 4 years of age. Dev. Med. Child Neurol. 60, 1149–1155. doi: 10.1111/dmcn.13761
Hofer, M. A. (2002). Unexplained infant crying: an evolutionary perspective. Acta Paediatr. 91, 491–496. doi: 10.1111/j.1651-2227.2002.tb03262.x
Howard, C., Lanphear, N., Lanphear, B., Eberly, S., and Lawrence, R. (2006). Parental responses to infant crying and colic: the effect on breastfeeding duration. Breastfeed. Med. 1, 146–155. doi: 10.1089/bfm.2006.1.146
Ioannadis, J. (2014). Increasing value and reducing waste in research design, conduct, and analysis. Lancet 383, 166–175. doi: 10.1016/s0140-6736(13)62227-8
Ismail, F. Y., Fatemi, A., and Johnston, M. V. (2017). Cerebral plasticity: windows of opportunity in the developing brain. Eur. J. Paediatr. Neurol. 21, 23–48. doi: 10.1016/j.ejpn.2016.07.007
Iverson, J. M., Shic, F., Wall, C. A., Chawarska, K., Curtin, S., Estes, A., et al. (2019). Early motor abilities in infants at heightened versus low risk for ASD: a baby siblings research consortium (BSRC) study. J. Abnorm. Child Psychol. 128, 69–80. doi: 10.1037/abn0000390
Jones, W., and Klin, A. (2013). Attention to eyes is present but in decline in 2-6-month-old infants later diagnosed with autism. Nature 504, 427–431. doi: 10.1038/nature12715
Kanold, P., and Luhmann, H. (2010). The subplate and early cortical circuits. Annu. Rev. Neurosci. 33, 23–48. doi: 10.1146/annurev-neuro-060909-153244
Kapoor, V., Douglas, P. S., Hill, P. S., Walsh, L., and Tennant, M. (2018). Frenotomy for tongue-tie in Australian children (2006-2016): an increasing problem. MJA 208, 88–89. doi: 10.5694/mja17.00438
Keating, D. P. (2016). Transformative role of epigenetics in child development research: commentary on the special section. Child Dev. 87, 135–142. doi: 10.1111/cdev.12488
Kempler, L., Sharpe, L., Miller, C. B., and Bartlett, D. J. (2016). Do psychosocial sleep interventions improve infant sleep or maternal mood in the postnatal period? A systematic review and meta-analysis of randomised controlled trials. Sleep Med. Rev. 29, 15–22. doi: 10.1016/j.smrv.2015.08.002
Kirby, C. N., Segal, A. Y., Hinds, R., Jones, K. M., and Pitterman, L. (2015). Infant gastro-oesophageal reflux disease (GORD): Australian GP attitudes and practices. J. Paediatr. Child Health 52, 47–53. doi: 10.111/jpc.12983
Koss, K. J., and Gunnar, M. R. (2018). Annual research review: early adversity, the hypothalamic-pituitary-adrenocortical axis, and child psychopathology. J. Child Psychol. Psychiatry 59, 327–346. doi: 10.1111/jcpp.12784
Laetitia, J., Smarius, C. A., Strieder, T. G. A., Loomans, E. M., Doreleijers, T. A. H., Vrijkotte, T. G. M., et al. (2016). Excessive infant crying doubles the risk of mood and behavioral problems at age 5: evidence for mediation by maternal characteristics. Eur. Child Adolesc. Psychiatry 26, 293–302. doi: 10.1007/s00787-016-0888-4
Landa, R. J., Stuart, E. A., Gross, A. L., and Faherty, A. (2012). Developmental trajectories in children with and without autism spectrum disorders: the first 3 years. Child Dev. 84, 429–442. doi: 10.1111/j.1467-8624.2012.01870.x
Landa, R. J., Stuart, E. A., Gross, A. L., and Faherty, A. (2013). Developmental trajectories in children with and without autism spectrum disorders: the first 3 years. Child Dev. 84, 429–442. doi: 10.1111/j.1467-8624.2012.01870.x
Lavender, T. J., Ebert, L., and Jones, D. (2016). An evaluation of perinatal mental health interventions: an integrative literature review. Women Birth. 29, 399–406. doi: 10.1016/j.wombi.2016.04.004
Lemcke, S., Parner, E. T., Bjerrum, M., Thomsen, P. H., and Lauritsen, M. B. (2018). Early regulation in children who are later diagnosed wtih autism spectrum disorder. A longitudinal study within the Danish national birth cohort. Infant Men. Health J. 39, 170–182. doi: 10.1002/imhj.21701
Levit-Binnun, N., and Golland, Y. (2012). Finding behavioral and network indicators of brain vulnerability. Front. Hum. Neurosci. 6:10. doi: 10.3389/fnhum.2012.00010
Lewis, J. D., Evans, A. C., Pruett, J. R., Botteron, K. N., Mckinstry, R. C., Zwaigenbaum, L., et al. (2017). The emergence of network inefficiencies in infants with autism spectrum disorder. Biol. Psychiatry 82, 176–185. doi: 10.1016/j.biopsych.2017.03.006
Lickliter, R. (2011). The integrated development of sensory organization. Clin. Perinatol. 38, 591–603. doi: 10.1016/j.clp.2011.08.007
Lydon, S., Healy, O., Reed, P., Mulhern, T., Hughes, B. M., and Goodwin, M. S. (2016). A systematic review of physological reactivity to stimuli in autism. Dev. Neurorehabil. 19, 335–355. doi: 10.3109/17518423.2014.971975
Mandy, W., and Lai, M.-C. (2016). Annual research review: the role of the environment in the developmental psychopathology of autism spectrum condition. J. Child Psychol. Psychiatry 57, 271–292. doi: 10.1111/jcpp.12501
Manohar, H., Prvallika, M., Kandasamy, P., Chandrasekaran, V., and Rajikumar, R. P. (2018). Role of exclusive breastfeeding in conferring protection in children at-risk for autism spectrum disorder: results from a sibling case-control study. J. Neurosci. Rural Practice 9, 132–136. doi: 10.4103/jnrp.jnrp_331_17
Mansfield, A. K., and Cordova, J. V. (2007). “A behavioral perspective on adult attachment style, intimacy, and relationship health,” in Understanding Behavior Disorders, eds D. Woods, and J. Kanter, (Oakland CA: Context Press), 389–416.
Marchesi, C., Ossola, P., Amerio, A., Daniel, B. D., Tonna, M., and De Panfilis, C. (2016). Clinical management of perinatal anxiety disorders: A systematic review. J. Affect. Disord. 190, 543–550. doi: 10.1016/j.jad.2015.11.004
Marin, O. (2016). Developmental timing and critical windows for the treatment of psychiatric disorders. Nat. Med. 22:1229. doi: 10.1038/nm.4225
Martiniuk, A. L. C., Vujovich-Dunn, C., Park, M., Yu, W., and Lucas, B. R. (2017). Plagiocephaly and developmental delay: a systematic review. J. Dev. Behav. Pediatr. 38, 67–78. doi: 10.1097/DBP.0000000000000376
Masi, A., Quintana, D. S., Glozier, N., Lloyd, A. R., Hickie, A. B., and Guastella, A. J. (2014). Cytokine aberrations in autism spectrum disorder: a systematic review and meta-analysis. Mol. Psychiatry 20, 440–446. doi: 10.1038/mp.2014.59
Matas, E., Bock, J., and Braun, K. (2016). The impact of parent-infant interaction on epigenetic plasticity mediating synaptic adaptations in the infant brain. Psychopathology 49, 201–210. doi: 10.1159/000448055
McCallum, S. M., Rowe, H. J., Gurrin, L. C., Quinlivan, J. A., Rosenthal, D. A., and Fisher, J. R. W. (2011). Unsettled infant behaviour and health service use: a cross-sectional community survey in Melbourne. Aust. J. Paediatr. Child Health 47, 818–823. doi: 10.1111/j.1440-1754.2011.02032.x
McCleery, J. P., Elliott, N. A., Sampanis, D. S., and Stefanidou, C. A. (2013). Motor development and motor resonance difficulties in autism: relevance to early intervention for language and communication skills. Front. Integr. Neurosci. 7:30. doi: 10.3389/fnint.2013.00030
McKenna, J. J., Ball, H. L., and Gettler, L. T. (2007). Mother-infant cosleeping, breastfeeding and sudden infant death syndrome: what biological anthropology has discovered about normal infant sleep and pediatric sleep medicine. Yearb. Physical. Anthropol. 50, 133–161. doi: 10.1002/ajpa.20736
Middlemiss, W., Granger, D. A., Goldberg, W. A., and Nathans, L. (2012). Asynchrony of mother-infant hypothalamic-pituitary-adrenal axis activity following extinction of infant crying responses induced during the transition to sleep. Early Hum. Dev. 88, 227–232. doi: 10.1016/j.earlhumdev.2011.08.010
Milidou, I., Lindhard, M. S., Sondergaard, C., Olsen, J., and Henriksen, T. B. (2015). Developmental coordination disorder in children with a history of infantile colic. J. Pediatr. 167, 725–730. doi: 10.1016/j.jpeds.2015.06.005
Mindell, J., and Lee, C. (2015). Sleep, mood, and development in infants. Infant. Behav. Dev. 41, 102–107. doi: 10.1016/j.infbeh.2015.08.004
Monteiro, F., Fonseca, A., Pereira, M., Alves, S., and Canavarro, M. C. (2018). What protects at-risk postpartum women from developing depressive and anxiety symptoms? The role of acceptance-focused processes and self-compassion. J. Affect. Disord. 246, 522–529. doi: 10.1016/j.jad.2018.12.124
Moore, E. R., Berman, N., Anderson, G. C., and Medley, N. (2016). Early skin-to-skin contact for mothers and their healthy newborn infants. Cochrane Database Syst. Rev. 5:CD003519. doi: 10.1002/14651858.CD14003519.pub14651854
NHMRC (2017). Report on the Evidence: Promoting Social and Emotional Development and Wellbeing of Infants in Pregnancy and the First Year of Life. Canberra: NHMRC.
Nugent, J. K. (2013). The competent newborn and the neonatal behavioral assessment scale: T. Berry Brazelton’s legacy. J. Child Adolesc. Psychiatr. Nurs. 26, 173–179. doi: 10.1111/jcap.12043
O’Shea, J. E., Foster, J. P., O’Donnell, C. P. F., Breathnach, D., Jacobs, S. E., Todd, D. A., et al. (2017). Frenotomy for tongue-tie in newborn infants (Review). Cochrane Database Syst. Rev. 3:CD011065.
Ozonoff, S., Young, G. S., Carter, A., Messinger, D., Yirmiya, N., Zwaigenbaum, L., et al. (2011). Recurrence risk for autism spectrum disorders: a baby siblings research consortium study. Pediatrics 128, e488–e495. doi: 10.1542/peds.2010-2825
Paquet, A., Olliac, B., Bouvard, M., Golse, B., and Vaivre-Douret, L. (2016). The semiology of motor disorders in autism spectrum disorders as highlighted from a standardized neuropsychomotor assessment. Front. Psychol. 7:1292. doi: 10.3389/fpsyg.2016.01292
Paquet, A., Olliac, B., Golse, B., and Vaivre-Douret, L. (2019). Nature of motor impairments in autism spectrum disorder: a comparison with developmental coordination disorder. J. Clin. Exp. Neuropsychol. 41, 1–14. doi: 10.1080/13803395.2018.1483486
Partty, A., and Kalliomaki, M. (2017). Infant colic is still a mysterious disorder of the microbiota-gut-brain axis. Acta Paediatr. 106, 528–529. doi: 10.1111/apa.13754
Partty, A., Kalliomaki, M., Salminen, S., and Isolauri, E. (2017). Infantile colic is associated with low-grade systematic inflammation. J. Pediatr. Gastroenterol. Nutrit. 64, 691–695. doi: 10.1097/mpg.0000000000001340
Patel, S., Day, T. N., Neil, J., and Mazefsky, C. A. (2018). Association between anger rumination and autism symptom severity, depression symptoms, aggression, and general dysregulation in adolescents with autism spectrum disorder. Autism 21, 181–189. doi: 10.1177/1362361316633566
Paterson, S., Wolff, J. J., Winder-Patel, B., Zwaigenbaum, L., Estes, A., Pandey, J., et al. (2019). The importance of temperament for understanding early manifestations of Autism Spectrum Disorder in high-risk infants. J. Autism Dev. Disord. 49, 2849–2863. doi: 10.1007/s10803-019-04003-2
Penn, A. H., Carver, L. J., Herbert, C. A., Lai, T. S., McIntire, M. J., Howard, J. T., et al. (2016). Breast milk protects against gastrointestinal symptoms in infants at high risk for autism during early development. J. Pediatr. Gastroenterol. Nutr. 2, 317–327. doi: 10.1097/MPG.0000000000000907
Pennestri, M.-H., Laganiere, C., Bouvette-Turcot, A.-A., Pokhvisneva, I., Steiner, M., Jeaney, M. J., et al. (2018). Uninterrupted infant sleep, development, and maternal mood. Pediatrics 142, e20174330. doi: 10.1542/peds.2017-4330
Petinou, K., and Minaidou, D. (2017). Neurobiological bases of autism spectrum disorders and implications for early intervention: a brief overview. Folia Phoniatr. Logop. 69, 38–42. doi: 10.1159/000479181
Peverill, S., Smither, I. M., duku, E., Szatmari, P., Vallancourt, T., Volden, J., et al. (2019). Developmental trajectories of feeding problems in children with autism spectrum disorder. J. Pediatr. Psychol. 44, 988–998. doi: 10.1093/jpepsy/jsz1033
Phagava, H., Muratori, F., Einspieler, C., Maestro, S., Apicella, F., Guzzetta, A., et al. (2008). General movements in infants with autism spectrum disorders. Georgian Med. News 156, 100–105.
Pierce, K., Courchesne, E., and Bacon, E. (2016). To screen or not to screen universally for autism is not the question: why the task force got it wrong. J. Pedatr. 176, 182–194. doi: 10.1016/j.jpeds.2016.06.004
Plsek, P., and Greenhalgh, T. (2001). The challenge of complexity in health care. Br. Med. J. 323, 625–627.
Pote, I., Wang, S., Sethna, V., Blasi, A., Daley, E., Kuklisova-Murgasova, M., et al. (2019). Familial risk of autism alters subcortical and cerebellar brain anatomy in infants and predicts the emergence of repetitive behaviors in early childhood. Autism Res. 12, 614–627. doi: 10.1002/aur.2083
Price, A., Quach, J., Wake, M., Bittman, M., and Hiscock, H. (2015). Cross-sectional sleep thresholds for optimal health and well-being in Australian 4-9-year-olds. Sleep Med. 22, 83–90. doi: 10.1016/j.sleep.2015.1008.1013
Price, A. M., Wake, M., Ukoumunne, O. C., and Hiscock, H. (2012). Outcomes at six years of age for children with infant sleep problems: longitudinal community-based study. Sleep Med. 13, 991–998. doi: 10.1016/j.sleep.2012.04.014
Renfrew, M., Pokhrel, S., Quigley, M., McCormick, F., Fox-Rushby, J., Dodds, R., et al. (2012). Preventing Disease and Saving Resources: the Potential Contribution of Increasing Breastfeeding Rates in the UK. London: Unicef UK.
Renz-Poster, H., and De Bock, F. (2018). Deformational plagiocephaly: the case for an evolutionary mismatch. Evol., Med. Public Health 2018, 180–185. doi: 10.1093/emph/eoy019
Rhoads, J. M., Collins, J., Fatheree, N. Y., Hashmi, S. S., Taylor, C. M., Luo, M., et al. (2018). Infant colic represents gut inflammation and dysbiosis. J. Pedatr. 203, 55–61. doi: 10.1016/j.jpeds.2018.07.042
Rimer, R., and Hiscock, H. (2014). National survey of Australian paediatricians’ approach to infant crying. J. Paediatr. Child Health 50, 202–207. doi: 10.1111/jpc.12452
Robertson, C. E., and Baron-Cohen, S. (2017). Sensory perception in autism. Nat. Rev. Neurosci. 18, 671–684. doi: 10.1038/nrn.2017.1112
Rogers, S. J., Vismara, L., Wagner, A. L., McCormick, C. E. B., Young, G. S., and Ozonoff, S. (2014). Autism treatment in the first year of life: a pilot study of Infant Start, a parent-implemented intervention for symptomatic infants. J Autism Dev. Disord. 44, 2981–2995. doi: 10.1007/s10803-014-2202-y
Ruiz, F. (2010). A review of acceptance and commitment therapy (ACT) empiral evidence: correlational, experimental psychopathology, component and outcome studies. Int. J. Psychol. Psychol. Ther. 10, 125–162.
Santos, I., Matijasevich, A., Capilheira, M., Anselmi, L., and Barros, F. (2015). Excessive crying at 3 months of age and behavioural problems at 4 years age: a prospective cohort study. J. Epidemiol. Commun. Health 69, 654–659. doi: 10.1136/jech-2014-204568
Satterstrom, A., Kosmicki, J., Wangk, J., Breen, M. S., De Rubeis, S., An, J.-Y., et al. (2018). Novel genes for autism implicate both excitatory and inhibitory cell lineages in risk. bioRxiv. [preprint].
Schmid, G., and Wolke, D. (2014). Preschool regulatory problems and attention-deficit/hyperactivity and cognitive deficits at school age in children born at risk: different phenotypes of dysregulation? Early Hum. Dev. 90, 399–405. doi: 10.1016/j.earlhumdev.2014.05.001
Schmied, V., Beake, S., Sheehan, A., McCourt, C., and Dykes, F. (2011). Women’s perceptions and experiences of breastfeeding support: a metasynthesis. Birth 38, 49–60. doi: 10.1111/j.1523-536X.2010.00446.x
Schore, A. N. (2001). Effects of a secure attachment relationship on right brain development, affect reglation, and infant mental health. Infant Men. Health J. 22, 7–66. doi: 10.1002/1097-0355(200101/04)22:1<7::aid-imhj2>3.0.co;2-n
Schultz, S. T., Klonoff-Cohen, H. S., and Wingard, D. L. (2006). Breastfeeding infant formula supplementation, and austistic disorder: the results of a parent survey. Int. Breastfeed. J. 1:16.
Shafer, R. L., Newell, K. M., Lewis, M. H., and Bodfish, J. W. (2017). A cohesive framework for motor stereotypy in typical and atypical deelopment: the role of sensorimotor integration. Front. Integr. Neurosci. 11:19. doi: 10.3389/fnint.2017
Sheinkopf, S. J., Levine, T. P., McCormick, C. E. B., Puggioni, G., Conradt, E., Lagasse, L. L., et al. (2019). Developmental trajectories of autonomic functioning in autism from birth to early childhood. Biol. Psychol. 142, 13–18. doi: 10.1016/j.biopsycho.2019.01.003
Shen, M. D., and Piven, J. (2017). Brain and behavior development in autism from birth through infancy. Dialogues Clin. Neurosci. 19, 325–333.
Shonkoff, J. P., and Phillips, D. A. (eds) (2000). From Neurons to Neighbourhoods: the Science of Early Childhood Development. Washington: National Academy Press.
Shui, A. M., Katz, T., Malow, B. A., and Mazurek, M. O. (2018). Predicting sleep problems in children with autism spectrum disorders. Res. Dev. Disabi. 83, 270–279. doi: 10.1016/j.ridd.2018.10.002
Sidor, A., Fischer, C., and Cierpka, M. (2017). The link between infant regulatory problems, temperament traits, maternal depressive symptoms and children’s psychopathological symptoms at age three: a longitudinal study in a German at-risk sample. Child Adolesc. Psychiatr. Men. Health 11, 10. doi: 10.1186/s13034-017-0148-5
Simard, V., Chevalier, V., and Bedard, M.-M. (2017). Sleep and attachment in early childhood: a series of meta-analyses. Attach. Hum. Dev. 19, 298–321. doi: 10.1080/14616734.2017.1293703
Simpson, E. A., Miller, G. M., Ferrari, P. F., Suomi, S. J., and Paukner, A. (2016). Neonatal imitation and early social experience predict gaze following abilities in infant monkeys. Sci. Rep. 6:20233. doi: 10.1038/srep20233
Siniscalco, D. (2015). Cmmentary: the impact of neuroimmune iterations in autism spectrum disorder. Front. Psychiatry 6:121. doi: 10.3389/fpsyt.2015.00145
Sloman, I., Bellinger, D. C., and Krentzel, C. P. (1990). Infantile colic and transient developmental lag in the first year of life. Child Psychiatry Hum. Dev. 21, 25–36. doi: 10.1007/bf00709925
St James-Roberts, I., Alvarez, M., Csipke, E., Abramsky, T., Goodwin, J., and Sorgenfrei, E. (2006). Infant crying and sleeping in London, copenhagen and when parents adopt a “proximal” form of care. Pediatrics 117, e1146–e1155. doi: 10.1542/peds.2005-2387
Steel, N., Abdelhamid, A., and Stokes, T. (2014). A review of clinical practice guidelines found that they were often based on evidence of uncertain relevance to primary care patients. J. Clin. Epidemiol. 67, 1251–1257. doi: 10.1016/j.jclinepi.2014.05.020
Stein, A., Pearson, R. M., Goodman, S. H., Rapa, E., Rahman, A., McCallum, M., et al. (2014). Effects of perinatal mental disorders on the fetus and child. Lancet 384, 1800–1819. doi: 10.1016/S0140-6736(14)61277-0
Steiner, A. M. (2013). Pivotal response treatment for infants at-risk for Autism spectrum disorders: a pilot study. J. Autism Dev. Disord. 43, 91–102. doi: 10.1007/s10803-012-1542-8
Stephens, R. L., Watson, L. R., Crais, E. R., and Reznick, J. S. (2018). Infant quantitative risk for autism spectrum disorder predicts executive function in early childhood. Autism Res. 11, 1532–1541. doi: 10.1002/aur.2024
Stevenson, R. A., Segers, M., Ncube, B. L., Black, K. R., Bebko, J. M., Ferber, S., et al. (2018). The cascading influence of multisensory processing on speech perception in autism. Autism 22, 609–624. doi: 10.1177/1362361317704413
Sujan, A. C., Oberg, A. S., Quinn, P. D., and D’Onofrio, B. M. (2019). Annual Research review: maternal antidepressant use during pregnancy and offspring neurodevelopmental problems - a critical review and recommendations for future research. J. Child Psychol. Psychiatry 2019, 356–376. doi: 10.1111/jcpp.13004
Sung, V., D’Amico, F., Cabana, M. D., Chau, K., Koren, G., Savino, F., et al. (2017). Lactobacillus reuteri to treat infant colic: a meta-analysis. Pediatrics 141, e20171811. doi: 10.1542/peds.2017-1811
Suren, P., Roth, C., Bresnahan, M., Haugen, M., Hornig, M., Hirtz, D., et al. (2013). Association between maternal use of folic acid supplements and risk of Autism Spectrum Disorders in children. JAMA 309, 570–577. doi: 10.1001/jama.2012.155925
Svensson, K. E., Velandia, M., Matthiesen, A.-S. T., Welles-Nystrom, B. L., and Widstrom, A.-M. E. (2013). Effects of mother-infant skin-to-skin contact on severe latch-on problems in older infants: a randomized trial. Int. Breastfeed. J. 8:1. doi: 10.1186/1746-4358-8-1
Swain, J. E., Konrath, S., Dayton, C. J., and Finegood, E. D. (2013). Toward a neuroscience of interactive parent-infant dyad empathy. Behav. Brain Sci. 36, 438–439. doi: 10.1017/S0140525X12002063
Synergies Economic Consulting (2011). Economic Costs of Autism Spectrum Disorder in Australia. Brisbane, QLD: Synergies Economic Consulting.
Thomas, K. A., Burr, R. L., Spieker, S., Lee, J., and Chen, J. (2014). Mother-infant circadian rhythm: development of individual patterns and dyadic synchrony. Early Hum. Dev. 90, 885–890. doi: 10.1016/j.earlhumdev.2014.09.005
Thompson, R. E., Kruske, S., Barclay, L., Linden, K., Gao, Y., and Kildea, S. V. (2016). Potential predictors of nipple trauma from an in-home breastfeeding programme: a cross-sectional study. Women Birth 29, 336–344. doi: 10.1016/j.wombi.2016.01.002
Tietz, A., Zietlow, A. L., and Reck, C. (2014). Maternal bonding in mothers with postpartum anxiety disorder: the crucial role of subclinical depressive symptoms and maternal avoidance behaviour. Arch. Womens Men. Health 17, 433–442. doi: 10.1007/s00737-014-0423-x
Tordjman, S., Somogyi, E., Coulon, N., Kermarrec, S., Cohen, D., Guillaume, B., et al. (2014). Gene x environment interactions in autism spectrum disorders: role of epigenetic mechanisms. Front. Psychiatry 5:53. doi: 10.3389/psyt.2014.00053
Torres, E. B., Brincker, M., Isenhower, R. W., Yanovich, P., Stigler, K. A., Numberger, J. I., et al. (2013). Autism: the micro-movement perspective. Front. Integr. Neurosci. 7:32. doi: 10.3389/fnint.2013.00032
Trevarthen, C., and Delafield-Butt, J. T. (2013). Autism as a developmental disorder in intentional movement and affective engagement. Front. Integr. Neurosci. 7:49. doi: 10.3389/fnint.2013.00049
Trutzer, I. M., Garcia-Cabezas, M. A., and Zikopoulos, B. (2019). Postnatal development and maturation of layer 1 in the lateral prefrontal cortex and its disruption in autism. Acta Neuropathol. Commun. 7:40. doi: 10.1186/s40478-40019-40684-40478
Tseng, P. T., Chen, Y. W., Stubbs, B., Carvalho, A. F., Whiteley, P., Tang, C. H., et al. (2017). Maternal breastfeeding and autism spectrum disorder in children: a systematic review and meta-analysis. Nutr. Neurosci. 22, 354–362. doi: 10.1080/1028415X.1022017
Tsivos, Z. L., Calam, R., Sanders, M. R., and Wittkowski, A. (2015). Interventions for postnatal depression assessing the mother-infant relationship and child develomental outcomes: a systematic review. Int. J. Women’s Health 23, 429–447. doi: 10.2147/IJWH.S75311
Tye, C., Runicles, A. K., Whitehouse, A. J. O., and Alvares, G. A. (2019). Characterizing the interplay between autism spectrum disorder and comorbid medical conditions: an integrative review. Front. Psychiatry 9:751.
Unwin, L. M., Bruz, I., Maybery, M. T., Reynolds, V., Ciccone, N., Dissanayake, C., et al. (2017). Acoustic properties of cries in 12-month old infants at high-risk of autism spectrum disorder. J. Autism Dev. Disord. 47, 2108–2119. doi: 10.1007/s10803-10017-13119-z
Valicenti-McDermott, M., Lawson, K., Hottinger, K., Seijo, R., Schechtman, M., Shulman, L., et al. (2019). Sleep problems in children with autism and other developmental disabilities: a brief report. J. Child Neurol. 34, 387–393. doi: 10.1177/0883073819836541
Verhoeff, M. E., Blanken, L. M. E., Kocevska, D., Mileva-Seitz, V. R., Jaddoe, V. W. V., White, T., et al. (2018). The bidirectional association between sleep problems and autism spectrum disorder: a population-based cohort study. Mol. Autism 9:8. doi: 10.1186/s13229-13018-10194-13228
Vik, T., Grote, V., Escribano, J., Socha, J., Verduci, E., Fritsch, M., et al. (2009). Infantile colic, prolonged crying and maternal postnatal depression. Acta Paediatr 8, 1344–1348. doi: 10.1111/j.1651-2227.2009.01317.x
Vivanti, G., Kasari, C., Green, J., Mandell, D., Maye, M., and Hudry, K. (2018). Implementing and evaluting early intervention for children with autism: where are the gaps and what should we do? Autism Res. 11, 16–23. doi: 10.1002/aur.1900
Von Kries, R., Kalies, H., and Papousek, M. (2006). Excessive crying beyond 3 months may herald other features of multiple regulatory problems. Arch. Pediatr. Adolesc. Med. 160, 508–511.
Wake, M., Morton-Allen, E., Poulakis, Z., Hiscock, H., and Gallagher, S. (2006). Prevalence, stability, and outcomes of cry-fuss and sleep problems in the first 2 years of life: prospective community-based study. Pediatrics 117, 836–842. doi: 10.1542/peds.2005-0775
Wan, M. W., Green, J., and Scott, J. (2018). A systematic review of parent-infant interaction in infants at risk of autism. Autism. 23, 811–820. doi: 10.1177/1362361318777484
Warner, B. B. (2018). The contribution of the gut microbiome to neurodevelopment and neuropsychiatric disorders. Pediatr. Res. 85, 216–224. doi: 10.1038/s41390-41018-10191-41399
Wass, S. V., De Barbaro, K., Clackson, K., and Leong, V. (2018). New meanings of thin-skinned: the contrasting attentional profiles of typical 12-month-olds who show high, and low, stress reactivity. Dev. Psychol. 54, 816–828. doi: 10.1037/dev0000428
Werchan, D. M., Baumgartner, H. A., Lewkowicz, D. J., and Amso, D. (2018). The origins of cortical multisensory dynamics: evidence from human infants. Dev. Cogn. Neurosci. 34, 75–81. doi: 10.1016/j.dcn.2018.07.002
Werner, E. A., Miller, M., Osborne, L., Kuzava, S., and Monk, C. (2015). Preventing postpartum depression: review and recommendations. Arch. Womens Men. Health 18, 41–60. doi: 10.1007/s00737-014-0475-y
Wess, J. M., Isaiah, A., Watkins, P. V., and Kanold, P. O. (2017). Subplate neurons are the first cortical neurons to respond to sensory stimuli. PNAS 114, 12602–12607. doi: 10.1073/pnas.1710793114
Whitehouse, A. J. O. (2017). Rethinking the clinical pathway for autism spectrum disorder and challenging the status quo. Int. J. Speech Lang. Pathol. 19, 208–217. doi: 10.1080/17549507.2016.1276963
Whitehouse, A. J. O., Varcin, K. J., Alvares, G. A., Barbaro, J., Bent, C., Bourtrus, M., et al. (2019). A randomised-controlled trial of a pre-emptive intervention for infants showing early behavioural risk signs of autism spectrum disorder. Lancet Child Adoles. Health. 3, 605–615. doi: 10.1016/S2352-4642(19)30184-1
Whittingham, K., and Douglas, P. S. (2014). Optimising parent-infant sleep from birth to 6 months: a new paradigm. Infant Men. Health J. 35, 614–623. doi: 10.1002/imhj.21455
Whittingham, K., and Douglas, P. S. (2016). “”Possums”: building contextual behavioural science into an innovative evidence-based approach to parenting support in early life,” in Psychotherapy in Parenthood and Beyond, ed. B. Kirkaldy, (Turin: Edizioni Minerva Medica), 43–56.
Whittingham, K., Sanders, M. R., McKinlay, L., and Boyd, R. N. (2014). Interventions to reduce behavioral problems in children with cerebral palsy: an RCT. Pediatics 133, e1249. doi: 10.1542/peds.2013-3620
Whyatt, C., and Craig, C. (2013). Sensory-motor problems in Autism. Front. Integr. Neurosci. 7:51. doi: 10.3389/fnint.2013.00051
Willfors, C., Carlsson, T., Anderlid, B.-M., Nordgren, A., Kostrzewa, E., Berggren, S., et al. (2017). Medical history of discordant twins and environmental etiologies of autism. Transal. Psychiatry 7, e1014. doi: 10.1038/tp.2016.269
Williams, K. E., Berthelsen, D., Walker, S., and Nicholson, J. M. (2017). A developmental cascade model of behavioral sleep problems and emotional and attentional self-regulation across early childhood. Behav. Sleep Med. 15, 1–21. doi: 10.1080/15402002.2015.1065410
Williams, K. E., Nicholson, J. M., Walker, S., and Berthelsen, D. (2016). Early childhood profiles of sleep problems and self-regulation predict later school adjustment. Br. J. Educ. Psychol. 86, 331–350. doi: 10.1111/bjep.12109
Winsper, C., and Wolke, D. (2014). Infant and toddler crying, sleeping and feeding problems and trajectories of dysregulated behavior across childhood. J. Abnorm. Child Psychol. 42, 831–843. doi: 10.1007/s10802-013-9813-1
Wolke, D., Bilgin, A., and Samara, M. (2017). Systematic review and meta-analysis: fussing and crying durations and prevalence of colic in infants. J. Pedatr. 185, 55–61. doi: 10.1016/j.jpeds.2017.02.020
Woods, N. K., Woods, N. F., Blackburn, S. T., and Sanders, E. A. (2016). Interventions that enhance breastfeeding initiation, duration and exclusivity: a systematic review. MCN 41, 299–307. doi: 10.1097/NMC.0000000000000264
Yu, C., and Smith, L. B. (2017). Multiple sensory-motor pathways lead to coordinated visual attention. Cogn. Sci. 41, (Suppl. 1), 5–31. doi: 10.1111/cogs.12366
Zeevenhooven, J., Browne, P. D. L., Hoir, M. P., De Weerth, C., and Benninga, M. A. (2018). Infant colic: mechanisms and management. Nat. Rev. 15, 479–496. doi: 10.1038/s41575-018-0008-7
Zeifman, D. M., and St James-Roberts, I. (2017). Parenting the crying infant. Curr. Opin. Psychol. 15, 149–154. doi: 10.1016/j.copsyc.2017.02.009
Zikopoulos, B., and Barbas, H. (2010). Changes in prefrontal axons may disrupt the network in autism. J. Neurosci. 30, 14595–14609. doi: 10.1523/JNEUROSCI.2257-10.2010
Zikopoulos, B., and Barbas, H. (2013). Altered neural connectivity in excitatory and inhibitory cortical circuits in autism. Front. Hum. Neurosci. 7:609. doi: 10.3389/fnhum.2013.00609
Keywords: autism (ASD), pre-emptive, intervention, environmental factors, complex adaptive systems
Citation: Douglas PS (2019) Pre-emptive Intervention for Autism Spectrum Disorder: Theoretical Foundations and Clinical Translation. Front. Integr. Neurosci. 13:66. doi: 10.3389/fnint.2019.00066
Received: 07 July 2019; Accepted: 04 November 2019;
Published: 19 November 2019.
Edited by:
Elizabeth B. Torres, Rutgers, The State University of New Jersey, United StatesReviewed by:
Miguel Ángel García-Cabezas, Boston University, United StatesKuan Hong Wang, University of Rochester, United States
Copyright © 2019 Douglas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pamela S. Douglas, cC5kb3VnbGFzQHBvc3N1bXNvbmxpbmUuY29t