 Nicolò Mangraviti
Nicolò Mangraviti Serena Castelli
Serena Castelli- 1Department of Neuroscience and Physiology, NYU Grossman School of Medicine, NYU Langone Health, New York, NY, United States
- 2Department for the Promotion of Human Science and Quality of Life, San Raffaele Open University, Rome, Italy
Cancer cells are primarily characterized by uncontrolled proliferation, which is closely linked to an increased energy demand. According to this, cancer cells preferentially utilize the glycolytic pathway for energy production. Metabolic regulation in cancer occurs at multiple levels, including post-translational modifications, which are rapid and energy-efficient, as they do not require enzyme degradation. In this context, non-coding RNA-mediated control plays a crucial role, as these small RNAs can directly modulate the mRNAs of key metabolic enzymes or induce epigenetic modulations, ultimately inducing alterations in cancer metabolism. In this mini-review, we summarize how non-coding RNAs (ncRNAs), including microRNAs, long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs), modulate core metabolic pathways, including glycolysis, mitochondrial respiration, lipid metabolism, and amino acid biosynthesis, in cancer. We also highlight recent findings that uncover novel regulatory mechanisms and propose that ncRNAs serve as dynamic integrators of metabolic and oncogenic signals. Understanding these interactions may open new avenues for biomarker identification and ncRNAs-targeted therapies development.
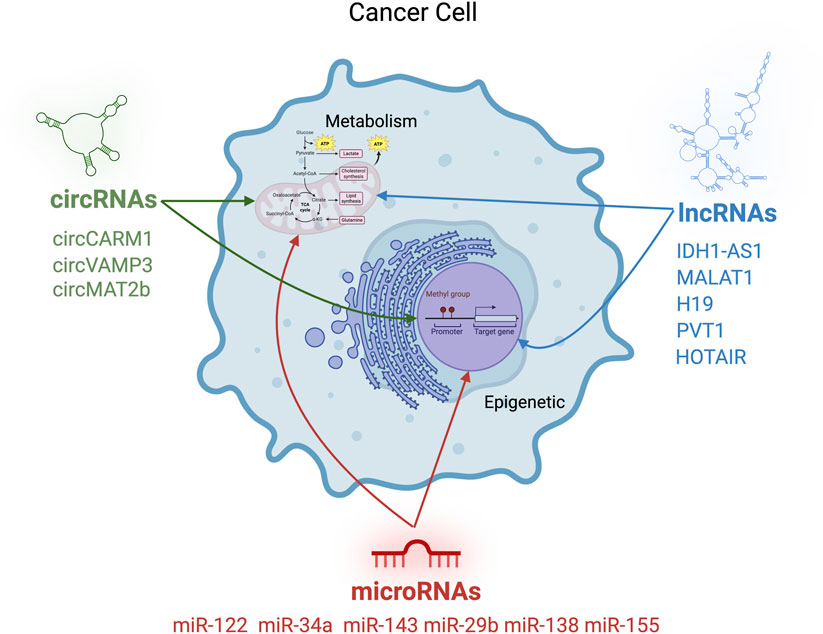
1 Introduction
Cancer metabolism is characterized by dramatic shifts in nutrient utilization, including enhanced aerobic glycolysis (the Warburg effect), increased lipid biosynthesis, and reprogramming of amino acid and mitochondrial metabolism (Mathew et al., 2024). While transcriptional regulators such as MYC and HIF-1α have been well studied in this context (Dong et al., 2020; Taneja et al., 2024), accumulating evidence reveals that non-coding RNAs (ncRNAs) are also central to this process. These molecules do not code for proteins but exert regulatory functions through RNA-RNA, RNA-DNA, or RNA-protein interactions. Although approximately 75% of the human genome is transcribed into RNA, only about 3% of these transcripts encode proteins. The majority of non-protein-coding transcripts correspond to ncRNAs (Yan and Bu, 2021; Zong et al., 2023).
NcRNAs can be broadly classified into three main categories. The first group includes housekeeping ncRNAs, such as tRNAs, rRNAs, snRNAs, and snoRNAs, which play essential structural and functional roles in cell biology. These RNAs are constitutively expressed across all cell types. The second group consists of small ncRNAs, including microRNAs (miRNAs) which will be the focus of this review small interfering RNAs (siRNAs), PIWI-interacting RNAs (piRNAs), and small Cajal body-specific RNAs (scaRNAs). These molecules are widely involved in gene expression regulation at multiple levels, acting both on chromatin and on specific mRNA targets. The third group comprises long ncRNAs (lncRNAs), which include long intergenic non-coding RNAs (lincRNAs) and circular RNAs (circRNAs), discussed below, as well as enhancer RNAs (eRNAs) and natural antisense transcripts (NATs) (Eldakhakhny et al., 2024; Fu, 2014). ncRNAs regulate gene expression through a variety of mechanisms, which are applied across multiple cellular pathways. These include pathways involved in differentiation (Mishra et al., 2023), proliferation, apoptosis (Tüncel et al., 2022; Wu et al., 2024), inflammation (Zhang et al., 2024), and, notably, metabolism. miRNAs have emerged as key regulators of metabolic processes, with growing evidence supporting the functional specificity of distinct miRNA families in targeting defined metabolic pathways. Several miRNA clusters are found to be overexpressed in various tumour types, contributing to oncogenesis by downregulating the expression of specific tumour suppressor proteins. This subset of miRNAs modulates mitochondrial metabolism, acting as oncogenic drivers through the suppression of oxidative phosphorylation (Thirunavukkarasu et al., 2024). Conversely, certain miRNAs function as tumor suppressors by downregulating the expression of key factors involved in glycolytic metabolism, such as glucose transporters (GLUTs). These tumor-suppressive miRNAs are frequently downregulated in cancer, thereby facilitating the metabolic reprogramming that supports tumor growth and progression (Safi et al., 2023). LncRNAs exert their regulatory functions through interactions with metabolic enzymes, modulating protein levels via mechanisms that include post-translational modifications, epigenetic alterations, lncRNA-mRNA binding that interferes with translation and miRNAs sponging. Through these diverse molecular mechanisms, lncRNAs can function either as oncogenic drivers or as tumor suppressors, depending on the context (Safi et al., 2023; Tan et al., 2021; Alluri et al., 2025). In addition, circRNAs also play a role in regulating metabolic reprogramming in cancer. As with other non-coding RNAs, their pro- or anti-tumorigenic functions are highly context-dependent and vary across different cancer types. A key feature of circRNAs is their ability to mediate long-distance cellular communication, partly due to their presence in extracellular vesicles such as exosomes (Liu et al., 2022). In this context, circRNAs have been primarily investigated for their involvement in the metastatic spread to distant peripheral sites from the primary tumor, as well as for their potential role as biomarkers for both diagnostic and prognostic purposes. Given the vast diversity of ncRNAs and, more importantly, their context-dependent roles across different tumor types, this review aims to summarize and integrate current knowledge on the involvement of ncRNAs, specifically, miRNAs, lncRNAs, and circRNAs, in the regulation of cancer metabolism.
2 MicroRNAs: small regulators with broad impact
MiRNAs are a class of ncRNAs that regulate gene expression post-transcriptionally, acting after mRNA synthesis and before its translation into protein. MiRNAs exert their function by recognizing and binding, with high specificity, to complementary sequences typically located in the 3′ untranslated region (3′ UTR) of target mRNAs. This interaction inhibits translation by promoting deadenylation and decapping, ultimately leading to mRNA degradation (O’Brien et al., 2018). Additionally, miRNAs have been shown to bind other regions of the mRNA, including the 5′ UTR and coding sequences, which can also suppress translation (Xu et al., 2014). Interestingly, miRNA binding to promoter regions has also been associated with upregulation of gene expression, indicating that miRNAs may exert both repressive and activating regulatory functions depending on the binding context (Dharap et al., 2013).
A growing body of evidence links the expression of specific miRNA clusters to metabolic reprogramming in cancer cells. For instance, miR-122, often downregulated in hepatocellular carcinoma, suppresses glycolysis and lipid metabolism by targeting pyruvate kinase and FASN (Chun, 2022). Similarly, miR-34a and miR-143 regulate key enzymes in glucose metabolism and the TCA cycle (Zhao et al., 2016; Zhang and Schulze, 2016). These miRNAs often function as tumor suppressors and are dysregulated in multiple cancer types. Beyond their direct involvement in the post-transcriptional regulation of metabolism-related mRNAs, microRNAs also contribute to epigenetic regulation mechanisms, a key aspect explored in this review.
In addition to being regulated by epigenetic mechanisms, miRNAs themselves can exert epigenetic effects, primarily by modulating the expression of key epigenetic enzymes such as DNMTs, TETs, HDACs, and EZH2 (Sato et al., 2011). Due to this specific regulatory activity, such miRNAs are referred to as epi-miRNAs; several of them are included in Table 1. Among them, miR-29b, part of the miR-29 family targets both DNMTs and TET enzymes. The downregulation of miR-29b leads to increased expression of DNMT3A, which in turn promotes the silencing of the tumor suppressor gene PTEN, a negative regulator of glycolysis and a positive regulator of energy expenditure (Yao et al., 2019). MiR-138 has been shown to downregulate the histone demethylase Lysine demethylase 5B (KDM5b), down-regulating key genes of lipid metabolism (including FASN and ACLY), thereby suppressing the proliferation and migration of breast cancer cells (Denis et al., 2016). KDM5b, indeed, is overexpressed in multiple cancer types, and elevated expression levels of this enzyme are associated with decreased survival (He et al., 2022). KDM5b has emerged as a promoter of cancer-associated lipid metabolic reprogramming, and its downregulation has been proposed as a potential therapeutic strategy in various tumors, including breast cancer, liver cancer, and pancreatic ductal adenocarcinoma (Jose et al., 2020; Zhang et al., 2019). MiR-137 is another microRNA implicated in epigenetic regulation associated with metabolic reprogramming. Among its targets is lysine (K)-specific demethylase 1A (LSD1), an epigenetic modifier known to exert widespread effects on genomic methylation (Yao et al., 2019). LSD1 has been reported to enhance glucose uptake, promoting glycolysis over mitochondrial metabolism and thereby contributing to the Warburg effect. Among the underlying mechanisms, LSD1 stabilizes HIF-1α, facilitating the expression of glycolytic genes such as hexokinase. In parallel, LSD1 promotes the degradation of key regulators of mitochondrial biogenesis, including PGC-1α. Additionally, LSD1 upregulates c-MYC, a well-known oncogenic factor that further supports glycolysis and tumor cell proliferation (Cheng et al., 2025). Epi-miRNAs have also been associated with the ability of cancer cells to adapt to stress conditions, such as hypoxia, which is characteristic of the inner regions of solid tumors. Notably, miR-155 has been shown to affect H3K36me2 expression under hypoxic conditions by repressing the histone lysine demethylase KDM2a, thereby limiting its activity. The miR-155–KDM2a interaction is essential for preventing excessive reactive oxygen species (ROS) production and for maintaining proper regulation of mitochondrial gene expression (Nakagawa et al., 2024). Epi-miRNAs have also been shown to play a key role in regulating the metabolism of immune cells, which are typically present in the tumor microenvironment. In these cells, metabolic pathways are closely linked to polarization and differentiation into either a pro-inflammatory, glycolytic phenotype, or an anti-inflammatory, oxidative phenotype. In this context, a representative example is epi-miR-143, which targets DNA methyltransferase 3A (DNMT3A). This miRNA can modulate the expression of pro-glycolytic genes, such as hexokinase and GLUT1, thereby influencing the metabolic programming of immune cells (Alshahrani et al., 2022). Based on this, the interest toward epi-miRNAs is growing since targeting a single miRNA can influence a broad network of downstream targets. In the case of epi-miRNAs, this effect is further amplified, as epigenetic regulators can induce widespread changes across the entire genome.
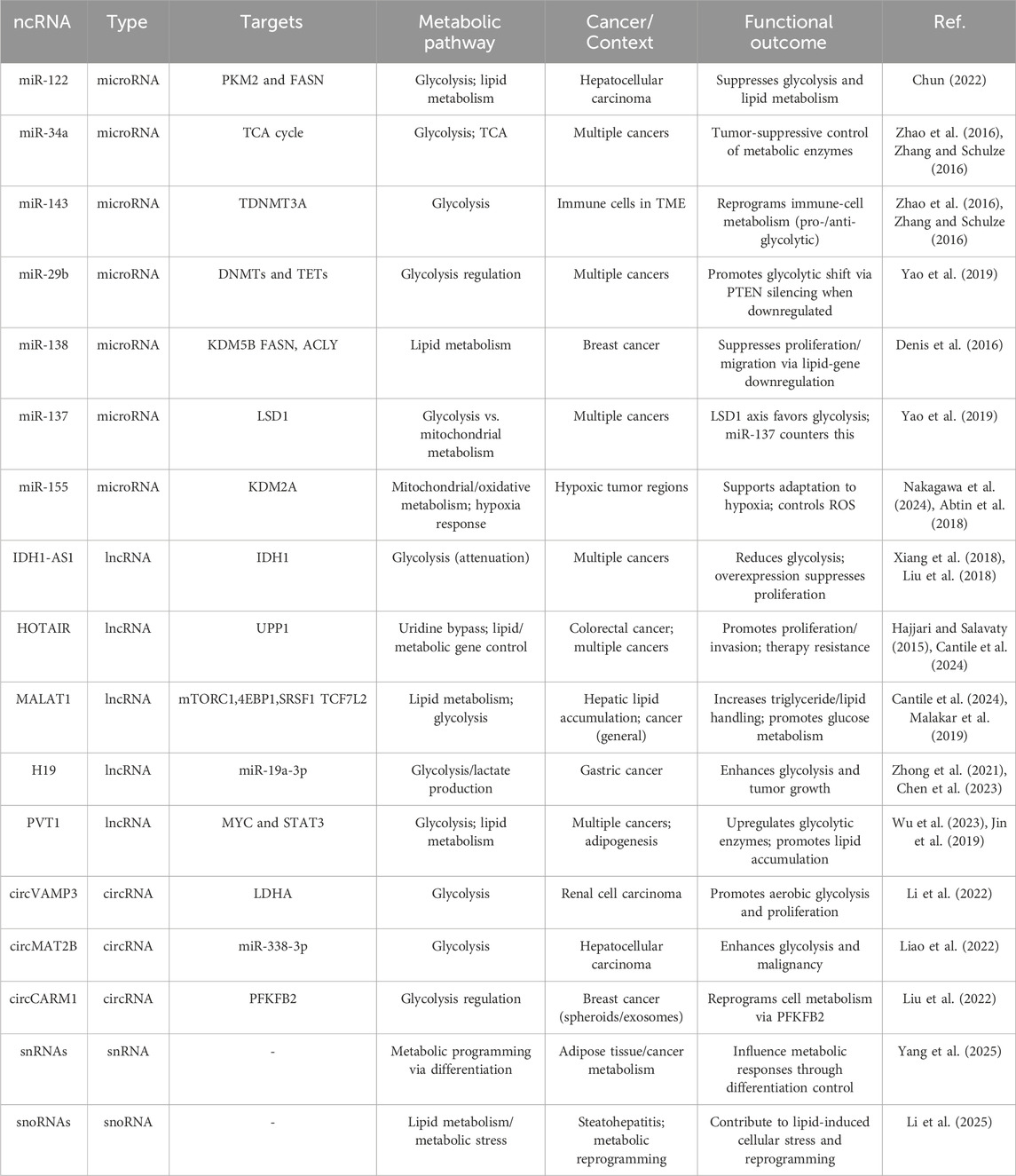
Table 1. Non-coding RNAs involved in metabolic regulation in cancer.
2.1 Long non-coding RNAs: The mastermind of metabolic adaptation
Emerging evidence indicates that lncRNAs play a central role in orchestrating and reinforcing cancer cells’ metabolic reprogramming. LncRNAs are transcripts longer than 500 nucleotides that lack coding potential but exert broad regulatory effects through diverse mechanisms (Palma et al., 2025). Functionally, they can act as guides, recruiting chromatin-modifying complexes to specific loci; scaffolds, assembling multiprotein complexes; decoys, sequestering transcription factors or miRNAs; and signals, reflecting the activity of distinct transcriptional programs (Mangraviti and De Windt, 2022). These categories often overlap, and many lncRNAs operate through multiple mechanisms simultaneously. In cancer, lncRNAs are emerging as pivotal integrators of epigenetic remodeling and metabolic reprogramming, enabling tumor cells to dynamically adapt to nutrient availability and therapeutic pressure.
Among the best characterized oncogenic lncRNAs is IDH1-AS1, which functions as a metabolic regulator by enhancing IDH1 homodimerization (Wu et al., 2020). This activity increases α-ketoglutarate levels and reduces ROS, resulting in downregulation of HIF-1α and attenuation of aerobic glycolysis. However, in cancer cells under normoxic conditions, the oncogene c-MYC transcriptionally represses IDH1-AS1, thereby maintaining HIF-1α activity and sustaining glycolytic metabolism. Experimentally, IDH1-AS1 overexpression suppresses tumor cell proliferation, whereas its knockdown promotes proliferation and tumor growth in xenograft models, highlighting the therapeutic potential of restoring IDH1-AS1 expression (Xiang et al., 2018; Liu et al., 2018).
HOTAIR exemplifies the guide function. It is significantly overexpressed in numerous cancers, including colorectal, breast, brain, lung, pancreatic, and gastric cancers (Hajjari and Salavaty, 2015). By predominantly recruiting PRC2, HOTAIR regulates a wide range of targets in trans, modulating both gene expression and alternative splicing, thereby influencing adipocyte development and human fat distribution (Kuo et al., 2022a). In addition, HOTAIR promotes H3K27me3 deposition and transcriptional silencing of genes enriched in nervous system developmental pathways and lipid metabolism (Kuo et al., 2022b). Its epigenetic activity has been implicated in resistance to chemotherapy, linking metabolic regulation to therapy adaptation. In colorectal cancer cells, HOTAIR drives proliferation and invasion by upregulating the metabolic enzyme UPP1 via recruitment of the histone methyltransferase EZH2, which deposits H3K27me3 on the UPP1 promoter. This HOTAIR–EZH2–UPP1 axis enables cancer cells to exploit uridine as an alternative energy source, a process known as the uridine bypass, thereby sustaining tumor growth under glucose limitation (Chen et al., 2024). Moreover, by coupling metabolic regulation with autophagy control, HOTAIR reduces the efficacy of radiotherapy and chemotherapy (Cantile et al., 2024).
Another well-characterized oncogenic lncRNA is MALAT1, whose upregulation contributes to hepatic lipid accumulation in response to oleic acid. MALAT1 regulates a network of lipid metabolism–related genes (Khalifa et al., 2025), particularly those involved in triglyceride synthesis and lipid handling, rather than canonical regulators such as SREBP-1 or PPARγ. These findings suggest that MALAT1 modulates both transcriptional and post-transcriptional layers of lipid metabolism. In cancer, MALAT1 promotes glucose metabolism through the mTORC1–4EBP1 signaling axis, where it increases SRSF1 expression to enhance translation of the transcription factor TCF7L2 (Malakar et al., 2019).
The imprinted lncRNA H19 illustrates the interplay between metabolism, chromatin state, and environmental cues. H19 is overexpressed in several cancers, including gastric cancer, where it correlates with poor prognosis. In normal physiology, H19 regulates processes such as neurogenesis and neural stem cell differentiation (Zhong et al., 2021), but it can also promote glucose consumption and lactate production by modulating metabolic enzymes. In gastric cancer, H19 enhances glycolysis and tumor growth by sponging miR-19a-3p, thereby upregulating the glycolytic enzyme PGK1. Silencing H19 reduces glycolysis, proliferation, and tumor size in vivo, confirming its oncogenic metabolic role (Chen et al., 2023).
Another important lncRNA reinforcing this epigenetic–metabolic axis is PVT1. This lncRNA is a critical regulator of differentiation and development (Wu et al., 2023). In the context of adipogenesis, PVT1 is upregulated during 3T3-L1 differentiation and in adipose tissue, where it promotes lipid accumulation by activating PPARγ, C/EBPα, and aP2, likely through interaction with STAT3, while enhancing fatty acid synthesis and inhibiting oxidation (Archambault et al., 2021). In cancer, PVT1 is frequently co-amplified with MYC. PVT1 stabilizes MYC protein, promotes expression of glycolytic enzymes, and thereby enhances tumor energy metabolism (Jin et al., 2019).
3 The new player on the board in metabolic regulation: CirRNA, SnoRNAs, tiRNAs
Although miRNAs and lncRNAs are now broadly recognized as major regulators of metabolic function and cancer progression, recent years have seen growing attention to additional classes of ncRNAs with similar functions. Among the most relevant are circRNAs, a large class of single-stranded RNA molecules that form covalently closed continuous loops with their 5′ and 3′ ends joined together (Awas et al., 2018). This unique structure lacks the typical termini of linear RNAs and often does not include a poly(A) tail. Mechanistically, circRNAs are generated through back-splicing, a process in which a downstream 5′ splice site joins to an upstream 3′ splice site of the same or different exon(s) within the pre-mRNA, resulting in a circular transcript. Important examples include circVAMP3, which promotes aerobic glycolysis and proliferation by regulating LDHA in renal cell carcinoma (Li et al., 2022), and circMAT2B, which similarly promotes glycolysis and malignancy in hepatocellular carcinoma through the miR-338-3p/PKM2 axis (Ferrera et al., 2025). Other examples of circRNAs found to be closely related to metabolic reprogramming are circMAT2B, circSPECC1, circC3P1 and circRPN2 (Liao et al., 2022).
Despite the fact that miRNAs, circRNAs, and lncRNAs are still considered the three major classes of ncRNAs in cancer metabolism, additional and less clearly defined RNA species are emerging as relevant regulators. Examples include small nuclear RNAs (snRNAs), which are non-coding RNAs of approximately 100–300 nucleotides localized in the nucleus and functioning as essential components of the spliceosome (Webster and Ghalei, 2023), and small nucleolar RNAs (snoRNAs), small RNAs of 60–300 nucleotides mainly located in the nucleolus that guide chemical modifications of other RNAs, particularly ribosomal RNAs (rRNAs), transfer RNAs (tRNAs), and snRNAs (Bratkovič et al., 2020). In particular, snoRNAs can impact various metabolic pathways, including the p53 pathway, which acts as a key regulator of tumor oxidative metabolism. They also influence the PI3K/AKT pathway and the MAPK signaling cascade, which is itself associated with the regulation of oxidative stress and autophagy (Li et al., 2025; Desideri et al., 2025). Compelling evidence suggests that snoRNAs contribute to lipid-induced cellular stress and metabolic reprogramming, including pathological processes such as steatohepatitis, whereas snRNAs have been implicated in the control of proper adipose tissue differentiation, thereby strongly influencing metabolic responses in cancer (Yang et al., 2025).
4 Discussion
The interplay between ncRNAs, epigenetic remodeling, and metabolic reprogramming defines a multilayered regulatory framework that grants cancer cells remarkable plasticity. MiRNAs, lncRNAs, and circRNAs converge on chromatin modifiers and metabolic enzymes to sustain glycolysis, repress mitochondrial respiration, and diversify lipid and amino acid metabolism. These molecules act within feedback loops integrating nutrient availability, metabolite levels, and chromatin state, allowing tumors to adapt to environmental and therapeutic pressures (Zong et al., 2023; Shin et al., 2025; Wang et al., 2022). For example, both miR-199a and miR-122 reduce the expression of pyruvate kinase M2 (PKM2) and are therefore downregulated under hypoxic conditions, which are common in cancer. This downregulation promotes glycolysis and supports the Warburg effect (Liao et al., 2022). In addition, hypoxia leads to the downregulation of miR-125a, which in turn results in the upregulation of hexokinase 2 (HK2), a key rate-limiting enzyme in the glycolytic pathway (Jin et al., 2017).
Despite progress, several challenges remain. ncRNAs often display context-dependent roles, similarly, circRNAs and lncRNAs that drive glycolysis in one setting may promote oxidative metabolism in another. Such dualities highlight the need for systematic studies across tumor types, ideally employing single-cell and spatial transcriptomics to capture heterogeneity. Such heterogeneity is associated with the fact that ncRNAs can exert their effects at multiple levels, and the response may depend on the basal expression level of specific genes, including those involved in the expression of metabolic enzymes. Many transcription factors altered in cancer are involved in metabolic regulation and are themselves regulated by ncRNAs, including the previously mentioned MYC. Therefore, the role of a specific ncRNA also depends on the basal expression of these factors, that could be different even the same tumor mass (Alfar et al., 2021). Another unresolved issue is causality within ncRNA–epigenetic–metabolic circuits: it is often unclear whether an ncRNA initiates remodeling or is induced by metabolic stress. Addressing this will require integrated approaches combining genetic perturbation, metabolomics, and chromatin profiling.
From a translational perspective, targeting ncRNAs remains promising but challenging. Antisense oligonucleotides, siRNAs, and CRISPR-based tools show potential, but specificity and delivery remain hurdles. Small molecules that disrupt lncRNA–protein interactions or circRNA biogenesis are emerging, while the stability of circRNAs and tissue specificity of many lncRNAs render them attractive biomarkers (Ruzi et al., 2025). In this regard, high-throughput screening of chemical libraries against RNA targets has led to the identification of promising lead compounds with high binding affinities. In this context, several small molecules have been identified capable of interfering with miRNA biogenesis (Ruzi et al., 2025; Syed et al., 2024). For example, oleuropein has been shown to reduce proliferation and induce apoptosis in breast cancer by downregulating miR-21 and miR-155, both oncomiRs which can favor glycolysis (Abtin et al., 2018; Hashemi et al., 2023). Among the different strategies that have been developed to suppress oncomiRs, there are anti-miRs (AMOs), miRNA sponges, miRNA masking, and small molecule inhibitors (SMIRs). AMOs are chemically modified synthetic antisense oligonucleotides of simple design and ease of synthesis. The main limitation is that their inhibitory effects are often transient due to dilution during cell division. MiRNA sponges are constructs containing multiple tandem binding sites complementary to a specific miRNA, effectively sequestering it and preventing interaction with its natural targets. MiRNA masks are single-stranded antisense oligonucleotides that bind to specific miRNA recognition elements on target mRNAs. Finally, SMIRs act at various stages of miRNA biogenesis (Arghiani and Shah, 2021).
Looking forward, integrating multi-omics with artificial intelligence and systems biology will be essential to map ncRNA–epigenetic–metabolic networks. Ultimately, resolving their mechanistic complexity may transform ncRNA-driven plasticity from an obstacle into a therapeutic vulnerability, advancing biomarker discovery and precision oncology.
Author contributions
NM: Writing – original draft. SC: Writing – review and editing, Conceptualization, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abtin, M., Alivand, M. R., Khaniani, M. S., Bastami, M., Zaeifizadeh, M., and Derakhshan, S. M. (2018). Simultaneous downregulation of miR-21 and miR-155 through oleuropein for breast cancer prevention and therapy. J. Cell. Biochem. 119 (9), 7151–7165. doi:10.1002/jcb.26754
Alfardus, H., de los Angeles Estevez-Cebrero, M., Rowlinson, J., Aboalmaaly, A., Lourdusamy, A., Abdelrazig, S., et al. (2021). Intratumour heterogeneity in microRNAs expression regulates glioblastoma metabolism. Sci. Rep. 11 (1), 15908. doi:10.1038/s41598-021-95289-9
Alluri, A., Saxena, P., Mishra, A., and Gutti, R. K. (2025). Association of long non-coding RNA in lipid metabolism: implications in leukemia. Int. J. Biochem. and Cell Biol. 184, 106785. doi:10.1016/j.biocel.2025.106785
Alshahrani, S. H., Ibrahim, Y. S., Jalil, A. T., Altoum, A. A., Achmad, H., Zabibah, R. S., et al. (2022). Metabolic reprogramming by miRNAs in the tumor microenvironment: focused on immunometabolism. Front. Oncol. 12, 1042196. doi:10.3389/fonc.2022.1042196
Archambault, A. S., Tinto, F., Dumais, É., Rakotoarivelo, V., Kostrzewa, M., Plante, P. L., et al. (2021). Biosynthesis of the novel endogenous 15-Lipoxygenase metabolites N-13-Hydroxy-octodecadienoyl-ethanolamine and 13-Hydroxy-octodecadienoyl-glycerol by human neutrophils and eosinophils. Cells. 10 (9), 2322. doi:10.3390/cells10092322
Arghiani, N., and Shah, K. (2021). Modulating microRNAs in cancer: next-generation therapies. Cancer Biol. Med. 19 (3), 289–304. doi:10.20892/j.issn.2095-3941.2021.0294
Awasthi, R., Singh, A. K., Mishra, G., Maurya, A., Chellappan, D. K., Gupta, G., et al. (2018). An overview of circular RNAs. Adv. Exp. Med. Biol. 1087, 3–14. doi:10.1007/978-981-13-1426-1_1
Bratkovič, T., Božič, J., and Rogelj, B. (2020). Functional diversity of small nucleolar RNAs. Nucleic Acids Res. 48 (4), 1627–1651. doi:10.1093/nar/gkz1140
Cantile, M., Belli, V., Scognamiglio, G., Martorana, A., De Pietro, G., Tracey, M., et al. (2024). The role of HOTAIR in the modulation of resistance to anticancer therapy. Front. Mol. Biosci. 11, 1414651. doi:10.3389/fmolb.2024.1414651
Chen, S., Wang, H., Xu, P., Dang, S., and Tang, Y. (2023). H19 encourages aerobic glycolysis and cell growth in gastric cancer cells through the axis of microRNA-19a-3p and phosphoglycerate kinase 1. Sci. Rep. 13, 17181. doi:10.1038/s41598-023-43744-0
Chen, X., Wang, S., Jiang, X., Zhang, M., and Ding, Y. (2024). Long non-coding RNA HOTAIR: from pan-cancer analysis to colorectal cancer-related uridine metabolism. Aging (Albany NY) 16 (9), 7752–7773. doi:10.18632/aging.205781
Cheng, B., Xu, X., Fan, B., Ma, T., and Yu, B. (2025). Epigenetic roles of lysine-specific demethylase 1 (LSD1) in cancer metabolism. Drug Discov. Today 30 (7), 104397. doi:10.1016/j.drudis.2025.104397
Chun, K. H. (2022). Molecular targets and signaling pathways of microRNA-122 in hepatocellular carcinoma. Pharmaceutics 14 (7), 1380. doi:10.3390/pharmaceutics14071380
Denis, H., Van Grembergen, O., Delatte, B., Dedeurwaerder, S., Putmans, P., Calonne, E., et al. (2016). MicroRNAs regulate KDM5 histone demethylases in breast cancer cells. Mol. Biosyst. 12 (2), 404–413. doi:10.1039/c5mb00513b
Desideri, E., Castelli, S., and Ciriolo, M. R. (2025). MAPK signaling in the interplay between oxidative stress and autophagy. Antioxidants (Basel) 14 (6), 662. doi:10.3390/antiox14060662
Dharap, A., Pokrzywa, C., Murali, S., Pandi, G., and Vemuganti, R. (2013). MicroRNA miR-324-3p induces promoter-mediated expression of RelA gene. PLOS ONE 8 (11), e79467. doi:10.1371/journal.pone.0079467
Dong, Y., Tu, R., Liu, H., and Qing, G. (2020). Regulation of cancer cell metabolism: oncogenic MYC in the driver’s seat. Signal Transduct. Target. Ther. 5 (1), 124. doi:10.1038/s41392-020-00235-2
Eldakhakhny, B., Sutaih, A. M., Siddiqui, M. A., Aqeeli, Y. M., Awan, A. Z., Alsayegh, M. Y., et al. (2024). Exploring the role of noncoding RNAs in cancer diagnosis, prognosis, and precision medicine. Non-coding RNA Res. 9 (4), 1315–1323. doi:10.1016/j.ncrna.2024.06.015
Ferrera, F., Altosole, T., Tardito, S., Astone, G., Bernardi, C., Parodi, A., et al. (2025). Immunoregulatory properties of cell free DNA. Cell Mol. Life Sci. 82 (1), 320. doi:10.1007/s00018-025-05862-y
Fu, X. D. (2014). Non-coding RNA: a new frontier in regulatory biology. Natl. Sci. Rev. 1 (2), 190–204. doi:10.1093/nsr/nwu008
Hajjari, M., and Salavaty, A. (2015). HOTAIR: an oncogenic long non-coding RNA in different cancers. Cancer Biol. and Med. 12 (1), 1–9. doi:10.7497/j.issn.2095-3941.2015.0006
Hashemi, M., Mirdamadi, M. S. A., Talebi, Y., Khaniabad, N., Banaei, G., Daneii, P., et al. (2023). Pre-clinical and clinical importance of miR-21 in human cancers: tumorigenesis, therapy response, delivery approaches and targeting agents. Pharmacol. Res. 187, 106568. doi:10.1016/j.phrs.2022.106568
He, R., Xhabija, B., Gopi, L. K., Kurup, J. T., Xu, Z., Liu, Z., et al. (2022). H3K4 demethylase KDM5B regulates cancer cell identity and epigenetic plasticity. Oncogene 41 (21), 2958–2972. doi:10.1038/s41388-022-02311-z
Jin, F., Wang, Y., Zhu, Y., Li, S., Liu, Y., Chen, C., et al. (2017). The miR-125a/HK2 axis regulates cancer cell energy metabolism reprogramming in hepatocellular carcinoma. Sci. Rep. 7 (1), 3089. doi:10.1038/s41598-017-03407-3
Jin, K., Wang, S., Zhang, Y., Xia, M., Mo, Y., Li, X., et al. (2019). Long non-coding RNA PVT1 interacts with MYC and its downstream molecules to synergistically promote tumorigenesis. Cell Mol. Life Sci. 76 (21), 4275–4289. doi:10.1007/s00018-019-03222-1
Jose, A., Shenoy, G. G., Sunil Rodrigues, G., Kumar, N. A. N., Munisamy, M., Thomas, L., et al. (2020). Histone demethylase KDM5B as a therapeutic target for cancer therapy. Cancers (Basel) 12 (8), 2121. doi:10.3390/cancers12082121
Khalifa, O., Ayoub, S., and Arredouani, A. (2025). Exploring the putative involvement of MALAT1 in mediating the beneficial effect of Exendin-4 on oleic acid-induced lipid accumulation in HepG2 cells. Biomedicines 13 (2), 370. doi:10.3390/biomedicines13020370
Kuo, F. C., Neville, M. J., Sabaratnam, R., Wesolowska-Andersen, A., Phillips, D., Wittemans, L. B. L., et al. (2022a). HOTAIR interacts with PRC2 complex regulating the regional preadipocyte transcriptome and human fat distribution. Cell Rep. 40 (4), 111136. doi:10.1016/j.celrep.2022.111136
Kuo, F. C., Huang, Y. C., Yen, M. R., Lee, C. H., Hsu, K. F., Yang, H. Y., et al. (2022b). Aberrant overexpression of HOTAIR inhibits abdominal adipogenesis through remodelling of genome-wide DNA methylation and transcription. Mol. Metab. 60, 101473. doi:10.1016/j.molmet.2022.101473
Li, J., Zhang, Q., Guan, Y., Liao, D., Jiang, D., Xiong, H., et al. (2022). Circular RNA circVAMP3 promotes aerobic glycolysis and proliferation by regulating LDHA in renal cell carcinoma. Cell Death Dis. 13 (5), 443. doi:10.1038/s41419-022-04863-0
Li, Y., Chen, X., Xiao, S., Wang, H., Li, B., Zhang, M., et al. (2025). Unlocking the life code: a review of SnoRNA functional diversity and disease relevance. Cell Commun. Signal 23, 266. doi:10.1186/s12964-025-02274-0
Liao, W., Du, J., Wang, Z., Feng, Q., Liao, M., Liu, H., et al. (2022). The role and mechanism of noncoding RNAs in regulation of metabolic reprogramming in hepatocellular carcinoma. Int. J. Cancer 151 (3), 337–347. doi:10.1002/ijc.34040
Liu, Z., Dai, J., and Shen, H. (2018). Systematic analysis reveals long noncoding RNAs regulating neighboring transcription factors in human cancers. Biochim. Biophys. Acta Mol. Basis Dis. 1864 (9 Pt B), 2785–2792. doi:10.1016/j.bbadis.2018.05.006
Liu, Y., Ma, L., Hua, F., Min, Z., Zhan, Y., Zhang, W., et al. (2022). Exosomal circCARM1 from spheroids reprograms cell metabolism by regulating PFKFB2 in breast cancer. Oncogene 41 (14), 2012–2025. doi:10.1038/s41388-021-02061-4
Malakar, P., Stein, I., Saragovi, A., Winkler, R., Stern-Ginossar, N., Berger, M., et al. (2019). Long noncoding RNA MALAT1 regulates cancer glucose metabolism by enhancing mTOR-Mediated translation of TCF7L2. Cancer Res. 79 (10), 2480–2493. doi:10.1158/0008-5472.can-18-1432
Mangraviti, N., and De Windt, L. J. (2022). Long non-coding RNAs in cardiac hypertrophy. Front. Mol. Med. 2, 836418. doi:10.3389/fmmed.2022.836418
Mathew, M., Nguyen, N. T., Bhutia, Y. D., Sivaprakasam, S., and Ganapathy, V. (2024). Metabolic signature of warburg effect in cancer: an effective and obligatory interplay between nutrient transporters and catabolic/anabolic pathways to promote tumor growth. Cancers (Basel) 16 (3), 504. doi:10.3390/cancers16030504
Mishra, A., Kumar, R., Mishra, S. N., Vijayaraghavalu, S., Tiwari, N. K., Shukla, G. C., et al. (2023). Differential expression of non-coding RNAs in stem cell development and therapeutics of bone disorders. Cells 12 (8), 1159. doi:10.3390/cells12081159
Nakagawa, R., Llorian, M., Varsani-Brown, S., Chakravarty, P., Camarillo, J. M., Barry, D., et al. (2024). Epi-microRNA mediated metabolic reprogramming counteracts hypoxia to preserve affinity maturation. Nat. Commun. 15 (1), 10516. doi:10.1038/s41467-024-54937-0
O’Brien, J., Hayder, H., Zayed, Y., and Peng, C. (2018). Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. (Lausanne) 9, 402. doi:10.3389/fendo.2018.00402
Palma, A., Buonaiuto, G., Ballarino, M., and Laneve, P. (2025). Genome biology of long non-coding RNAs in humans: a virtual karyotype. Comput. Struct. Biotechnol. J. 27, 575–584. doi:10.1016/j.csbj.2025.01.026
Ruzi, Z., Han, D., and Aierken, K. (2025). Advanced strategies for screening and identifying RNA-targeted small molecules: bridging therapeutic potential and innovation. Results Chem. 15, 102305. doi:10.1016/j.rechem.2025.102305
Safi, A., Saberiyan, M., Sanaei, M. J., Adelian, S., Davarani Asl, F., Zeinaly, M., et al. (2023). The role of noncoding RNAs in metabolic reprogramming of cancer cells. Cell. and Mol. Biol. Lett. 28 (1), 37. doi:10.1186/s11658-023-00447-8
Sato, F., Tsuchiya, S., Meltzer, S. J., and Shimizu, K. (2011). MicroRNAs and epigenetics. FEBS J. 278 (10), 1598–1609. doi:10.1111/j.1742-4658.2011.08089.x
Shin, C. H., Kim, K., Ho, C. W., Lee, J. W., Jo, M. J., Min, K. W., et al. (2025). Long noncoding RNAs regulating enzymatic reactions in cancer. Exp. Mol. Med. 57 (8), 1641–1650. doi:10.1038/s12276-025-01464-7
Syed, R. U., Banu, H., Alshammrani, A., Alshammari, M. D., G, S. K., Kadimpati, K. K., et al. (2024). MicroRNA-21 (miR-21) in breast cancer: from apoptosis dysregulation to therapeutic opportunities. Pathology - Res. Pract. 262, 155572. doi:10.1016/j.prp.2024.155572
Tan, Y. T., Lin, J. F., Li, T., Li, J. J., Xu, R. H., and Ju, H. Q. (2021). LncRNA-mediated posttranslational modifications and reprogramming of energy metabolism in cancer. Cancer Commun. 41 (2), 109–120. doi:10.1002/cac2.12108
Taneja, N., Chauhan, A., Kulshreshtha, R., and Singh, S. (2024). HIF-1 mediated metabolic reprogramming in cancer: mechanisms and therapeutic implications. Life Sci. 352, 122890. doi:10.1016/j.lfs.2024.122890
Thirunavukkarasu, S., Banerjee, S., Tantray, I., and Ojha, R. (2024). Non-coding RNA and reprogrammed mitochondrial metabolism in genitourinary cancer. Front. Genet. 15, 1364389. doi:10.3389/fgene.2024.1364389
Tüncel, Ö., Kara, M., Yaylak, B., Erdoğan, İ., and Akgül, B. (2022). Noncoding RNAs in apoptosis: identification and function. Turk J. Biol. 46 (1), 1–40. doi:10.3906/biy-2109-35
Wang, Y., Li, Q., Wang, S., Wang, B. J., Jin, Y., Hu, H., et al. (2022). The role of noncoding RNAs in cancer lipid metabolism. Front. Oncol. 12, 1026257. doi:10.3389/fonc.2022.1026257
Webster, S. F., and Ghalei, H. (2023). Maturation of small nucleolar RNAs: from production to function. RNA Biol. 20 (1), 715–736. doi:10.1080/15476286.2023.2254540
Wu, S., Ding, L., Xu, H., Gao, J., Shao, Y., Zhang, S., et al. (2020). The long non-coding RNA IDH1-AS1 promotes prostate cancer progression by enhancing IDH1 enzyme activity. Onco Targets Ther. 13, 7897–7906. doi:10.2147/OTT.S251915
Wu, F., Zhu, Y., Zhou, C., Gui, W., Li, H., and Lin, X. (2023). Regulation mechanism and pathogenic role of lncRNA plasmacytoma variant translocation 1 (PVT1) in human diseases. Genes and Dis. 10 (3), 901–914. doi:10.1016/j.gendis.2022.05.037
Wu, H., Liu, Y., and Liu, C. (2024). The interregulatory circuit between non-coding RNA and apoptotic signaling in diabetic cardiomyopathy. Non-coding RNA Res. 9 (4), 1080–1097. doi:10.1016/j.ncrna.2024.06.011
Xiang, S., Gu, H., Jin, L., Thorne, R. F., Zhang, X. D., and Wu, M. (2018). LncRNA IDH1-AS1 links the functions of c-Myc and HIF1α via IDH1 to regulate the warburg effect. Proc. Natl. Acad. Sci. U. S. A. 115 (7), E1465–E1474. doi:10.1073/pnas.1711257115
Xu, W., San Lucas, A., Wang, Z., and Liu, Y. (2014). Identifying microRNA targets in different gene regions. BMC Bioinforma. 15 (7), S4. doi:10.1186/1471-2105-15-s7-s4
Yan, H., and Bu, P. (2021). Non-coding RNA in cancer. Essays Biochem. 65 (4), 625–639. doi:10.1042/ebc20200032
Yang, Y., Zhang, G., Yi, T., Yang, S., Wu, S., Zhang, Y., et al. (2025). snRNA-seq reveals subcutaneous white adipose tissue remodeling upon return to thermoneutrality after cold stimulation. Front. Cell Dev. Biol. 13, 1578180. doi:10.3389/fcell.2025.1578180
Yao, Q., Chen, Y., and Zhou, X. (2019). The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 51, 11–17. doi:10.1016/j.cbpa.2019.01.024
Zhang, X., and Schulze, P. C. (2016). MicroRNAs in heart failure: non-coding regulators of metabolic function. Biochimica Biophysica Acta (BBA) - Mol. Basis Dis. 1862 (12), 2276–2287. doi:10.1016/j.bbadis.2016.08.009
Zhang, Z. G., Zhang, H. S., Sun, H. L., Liu, H. Y., Liu, M. Y., and Zhou, Z. (2019). KDM5B promotes breast cancer cell proliferation and migration via AMPK-Mediated lipid metabolism reprogramming. Exp. Cell Res. 379 (2), 182–190. doi:10.1016/j.yexcr.2019.04.006
Zhang, Y., Liu, H., Niu, M., Wang, Y., Xu, R., Guo, Y., et al. (2024). Roles of long noncoding RNAs in human inflammatory diseases. Cell Death Discov. 10 (1), 235. doi:10.1038/s41420-024-02002-6
Zhao, L., Mao, Y., Zhao, Y., Cao, Y., and Chen, X. (2016). Role of multifaceted regulators in cancer glucose metabolism and their clinical significance. Oncotarget 7 (21), 31572–31585. doi:10.18632/oncotarget.7765
Zhong, L., Liu, P., Fan, J., and Luo, Y. (2021). Long non-coding RNA H19: physiological functions and involvements in central nervous system disorders. Neurochem. Int. 148, 105072. doi:10.1016/j.neuint.2021.105072
Keywords: ncRNA, metabolic reprogramming, circRNAs, microRNA, lncRNA
Citation: Mangraviti N and Castelli S (2025) Non-coding RNAs at the intersection of epigenetics and cancer metabolism. Front. Epigenet. Epigenom. 3:1699969. doi: 10.3389/freae.2025.1699969
Received: 05 September 2025; Accepted: 10 October 2025;
Published: 20 October 2025.
Edited by:
Andre J. van Wijnen, University of Vermont, United StatesReviewed by:
Marios Agelopoulos, Biomedical Research Foundation of the Academy of Athens (BRFAA), GreeceCopyright © 2025 Mangraviti and Castelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Serena Castelli, c2VyZW5hLmNhc3RlbGxpQHVuaXJvbWE1Lml0