 Vladimir Dinets1,2*
Vladimir Dinets1,2*- 1Psychology Department, University of Tennessee, Knoxville, TN, United States
- 2Department of Mathematics and Computer Science, Rutgers University, Newark, NJ, United States
Introduction: Urban birds have to adapt to their complex and challenging environment. In the present study, a particularly remarkable behavioral adaptation by Cooper’s hawk (Accipiter cooperi) was observed.
Methods: Observations were conducted visually in urban habitat in New Jersey, where Cooper’s hawks are mostly winter residents.
Results: An immature Cooper’s hawk was observed hunting birds near a road intersection using queues of cars waiting for green light as cover. The queues grew sufficiently long only when pedestrian crossing regime was activated at the streetlight. The hawk apparently learned to prepare for attack when sound signals indicated the activation of pedestrian crossing regime.
Discussion: The observed behavior required having a mental map of the area and understanding the connection between the sound signals and the change in traffic pattern – a remarkable intellectual feat for a young bird that likely had just moved into the city. Such level of understanding and use of human traffic patterns by a wild animal has never been reported before.
Introduction
Urban birds often develop remarkable behavioral adaptations to their complex and challenging environment. Moving vehicles are an important feature of that environment, and birds learn not just to avoid the danger they represent, but also to exploit them in various ways. Crows (Corvus spp.) drop walnuts, small vertebrates, shelled mollusks and other prey/food items onto busy roads to have them killed and/or crushed by cars (Maple, 1974; Grobecker and Pietsch, 1978; Nihei, 1995; Nihei and Higuchi, 2001; Marzluff and Angell, 2007; Kim Anderson, pers. comm.), with various populations having invented/learned this technique at different times and some not at all (Cristol et al., 1997; Nihei and Higuchi, 2001). Carrion-eating birds routinely monitor or patrol busy roads to immediately snatch roadkill (Santos et al., 2011; Planillo et al., 2015; Harju et al., 2018). Passerines glean dead insects from cars (Jokimäki and Kaisanlahti-Jokimäki, 2023) and sometimes even nest in moving cars, trains and boats (Turner, 2015). There are observations of small birds using moving cars as mobile shelters from pursuing raptors (Daan Drukker, pers. comm.; Hadija Mustafa, pers. comm.).
Accipiter hawks are stealthy hunters that routinely and masterfully use various objects as cover to get closer to their prey before the final charge (Roth and Lima, 2003) and combine long waits in ambush with lightning-fast chases (Thiollay, 1994). To hunt this way, they must be able to plan ahead, to know and understand the prey’s behavior, particularly its movement patterns, and to be highly observant – in short, they need remarkable cognitive abilities. Their intelligence and flexibility make them highly successful hunters in both natural and human-modified environments: moving into cities can actually improve their hunting success (Malan and Robinson, 1999; Estes and Mannan, 2003; Rutz, 2006), so many Accipiter species are now nesting and/or wintering in cities around the world (Thiollay, 1994). Urban hawks develop novel ways of hunting not observed in their “rural” conspecifics. Rutz (2006), who studied Eurasian goshawks (A. gentlis) in Hamburg (Germany), listed three such techniques: “(i) Peregrine-like perched hunts from exposed vantage points like roof-tops and TV aerials; (ii) ‘backyard jumping’ and low quartering flights in residential areas where buildings and other objects are used for cover during surprise attacks; and (iii) foraging after sunset under artificial light conditions”. Other researchers reported urban Eurasian sparrowhawks (A. nisus) chasing prey into narrow streets with no escape routes (Dietrich, 1982) or into large glass windows (Theo Squires, pers. comm.); the latter tactic has also been reported for sharp-shinned hawks (A. striatus) (Klem, 1981). In the city of Kharkiv (Ukraine), Eurasian sparrowhawks have long been known to use moving cars and streetcars as cover to sneak up on their prey undetected (Krivitsky and Reutsky, 2003), and a Eurasian goshawk has recently been observed doing so as well (Dmitry Strelkov, pers. comm.).
Cooper’s hawk (A. cooperi) began to utilize cities as breeding and (more commonly) wintering habitat in the 1970s and quickly became the most successful colonizer of cities among Accipiter hawks (see review by Rosenfield et al., 2018). This medium-sized, hypercarnivorous raptor is an exception to the general trend of small, generalist raptors being more successful in cities (Cooper et al., 2022; Headland et al., 2023). However, the only novel hunting technique adapted specifically for urban environments reported for this species to date is hunting around artificial lights (Boal, 1997).
In 2021–2022, I repeatedly observed Cooper’s hawk(s) using cars backed up at a particular intersection as cover for approaching the prey undetected, and using the sound signals for visually impaired pedestrians (hereafter “sound signals”) as a clue that the red light phase of the streetlight will be longer and more cars will line up. This appears to be the most advanced case of raptors adapting to use traffic patterns reported to date.
Methods
Study site
The study was conducted in the town of West Orange, New Jersey, USA, at the intersection of two streets hereafter named Main Street and Side Street to protect residents’ privacy. Main Street was wider and busier (Figure 1). Side Street usually had little traffic, but during morning rush hours, small queues of cars formed before the intersection when the streetlight was in red phase. During most red light phases the queue was up to 4 cars long and did not stretch past house #4. However, if a pedestrian pressed a button at the streetlight on Main Street in order to cross it, the red light phase became much longer, increasing from 30 to 90 sec, and the queue of cars could stretch up to house #10 (Figure 1). When the button was pressed, sound signals were activated at the streetlight and continued for 45 sec. At the time of observation there were 2–6 (av. 3) pedestrian crossings per hour, meaning that 1 out of 10–30 red phases was longer than others, and that the sound signals were on for ~3.75% of the time.
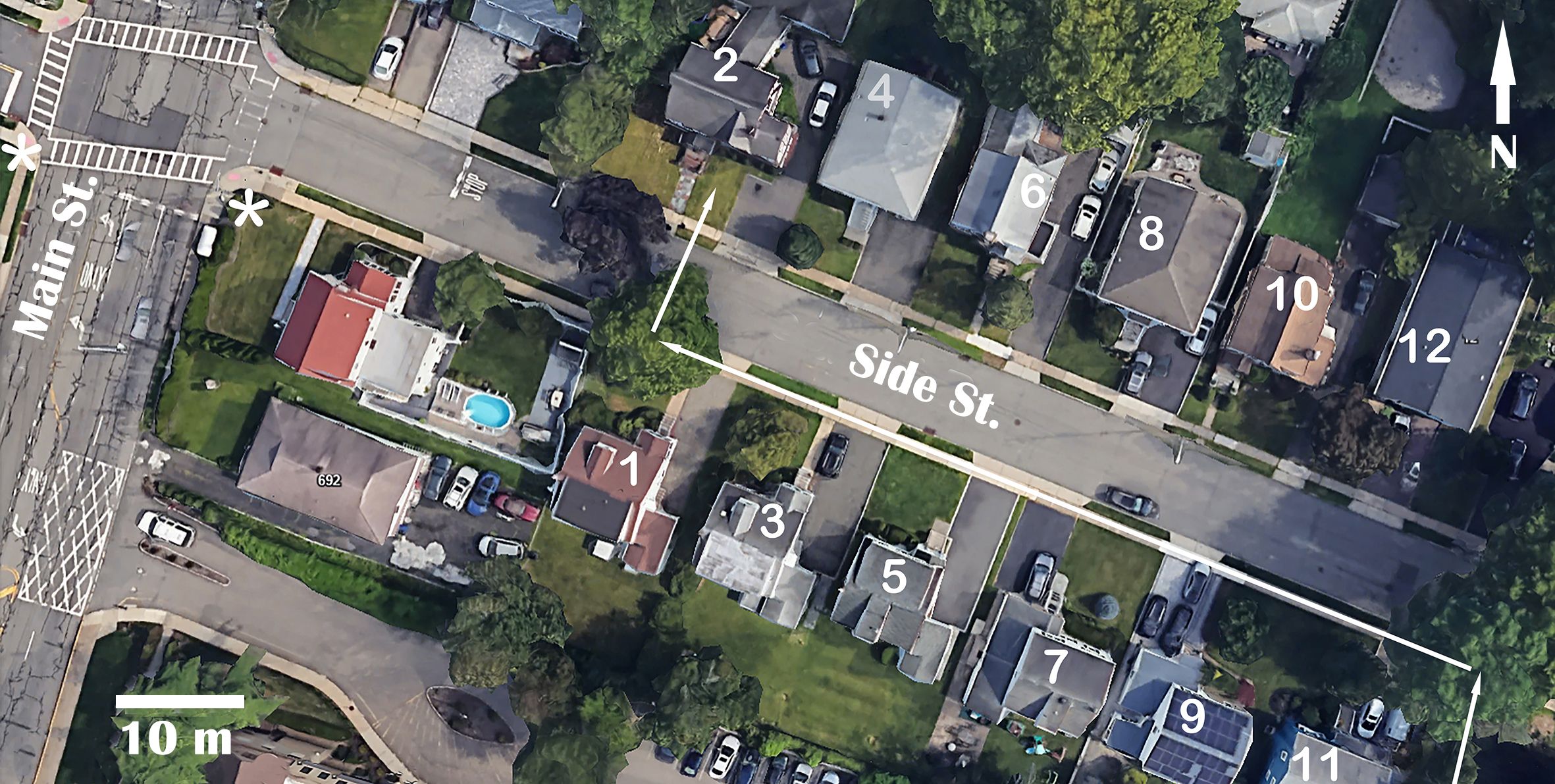
Figure 1. The study area. The numbers are house numbers mentioned in the text. The route used by the hawk to attack a flock of birds feeding in front of house #2 is shown with white arrows. The hawk appeared in the tree in front of house #11 as soon as sound signals at the streetlight at the intersection (marked with white asterisks) indicated that red light will be longer than usual, and attacked when the queue of cars reached house #8, making it possible for the hawk to move to the tree in front of house #1 without being visible to potential prey.
Residents of house #2 regularly ate their evening meals in front of the house, leaving breadcrumbs and other small leftovers that attracted a small flock of house sparrows (Passer domesticus), mourning doves (Zenaida macroura), and sometimes European starlings (Sturnus vulgaris) next morning.
Cooper’s hawks are uncommon but regular winter residents in West Orange, but to my knowledge they have never been recorded nesting there, and are very rarely observed in summer, with just three summer records on eBird in the last ten years (eBird, 2024).
Observations
In November 2021, while driving through the intersection every morning, I noticed that an immature Cooper’s hawk (presumably the same individual) was using cars that queued on Side Street waiting for green light as a cover to approach the flock of birds feeding in front of house #2. I then conducted a series of observations from a car parked in front of house #9 (Figure 1).
The observations were conducted between 07:30 and 09:00 Eastern Standard Time, for 30–60 minutes a day, for a total of 12 hours, on 18 days between 5 December 2021 and 3 March 2022.
Systematic observations were conducted only on weekdays (there were no car queues on weekends) and only on mornings without precipitation on that and previous days (if there was rain or snow the previous day, the residents of house #2 did not eat outside, so there was no bird flock feeding on the leftovers in the morning). On some days the observations were not conducted because the flock was not present for no obvious reason. It is possible that on some or all of those days, the hawk attack had happened earlier, scaring away the birds.
Results
In 12 hours of observation, 6 attempted attacks were observed. From the side of Side Street where I parked the car, I could see the hawk approaching the part of the street where the prey flock was located, but not the attack itself, so I do not know how many attacks were successful. Once the hawk was seen flying away back across Side Street with a house sparrow in its talons, and once it was observed eating a mourning dove on the ground near the site of attack. After each attack, the bird flock scattered and never returned on the same morning; more than one attack attempt per day has never been observed.
It took me the first two observations to determine the entire course of attacks; it was the same during the remaining four attacks. The hawk would first fly into the tree in front of house #11 (Figure 1) and perch on a low branch near the trunk for up to one minute while the queue of cars was getting longer. The cars obstructed the hawk’s view of the prey and vice versa. As soon as the queue reached house #8, the hawk would take off, fly ~65 m (less than 1 m above ground) along the pedestrian sidewalk on the southern side of Side Street, make a 90° right turn under the tree in front of house #1, and cross Side Street between cars toward the prey flock feeding in front of house #2 (Figure 1).
While observing the fourth attack, I noticed that the hawk appeared in the tree in front of house #11 when the sound signals at the streetlight were on. I realized that the sound accurately predicted that the car queue would be longer than usual. In two subsequently observed attacks, the hawk also appeared in the tree when the sound signals were on, but before the long car queue formed. That was not a random occurrence (arithmetic probability P= 0.03753 ≈ 0.000053, as the signal was on for only ~3.75% of the time). In all three cases, the signal was no longer active by the time the hawk attacked. It is unknown if the first three observed attacks also started when the signals were on.
The hawk was never seen on weekends, even though I drove or walked through the intersection many times. In the winter of 2022–2023 I twice accidentally observed an adult Cooper’s hawk using the same method at the same location. It is very likely that it was the same individual as the immature hawk the year before. These two observations were not included in the analysis because total observation time was not recorded and the identity of the bird was not certain. In the summer of 2023, the sound signals at the streetlight stopped working for unknown reason, and the residents of house #2 stopped leaving leftovers. No hawks were ever observed at the intersection after that.
Discussion
This is a case study, and as such it would be difficult or impossible to replicate. However, it does not particularly stand out among existing observations of complex hunting behavior of raptors: various species can be taught to selectively hunt certain prey species other than their own preferred ones (Bildstein, 2006); others hunt in cooperating and coordinating packs (Bednarz, 1988), use tools to procure food and nesting material (Stoyanova et al., 2010; Santiago, 2025), and use burning sticks to start wildfires, facilitating prey detection (Bonta et al., 2017).
The observations presented here could be explained as a result of simple Pavlovian training, but that would be an obvious oversimplification. To hunt as observed, the hawk had to develop a precise mental map of the street (as the target flock was invisible to the hawk until the last stage of attack), notice the connection between the sound signals and the length of the car queue, and figure out that only longer queues provided cover for the entire approach. The ability to plot the most efficient approach to a food item obscured by obstacles has been documented in corvids (i.e., Bugnyar et al., 2004) and some mammals (i.e., Dumas et al., 2006), but not in birds of prey. However, tests have shown that birds of prey can be as good at inventing and learning as parrots and corvids (Smith and Colbert-White, 2023).
The behavior described here is an impressive feat of intelligence, going a long way to explain the species’ ability to successfully colonize such unusual and dangerous environment as urban landscape. This intelligence was likely pre-existing rather than evolved in the novel environment. Red foxes (Vulpes vulpes) have been found to become bolder but not more intelligent in cities (Morton et al., 2023), even though they’ve been living there longer than Cooper’s hawks. Urban chimango caracaras (Milvago chimango) show faster learning and lower neophobia than rural ones, but their learning process is similar (Biondi et al., 2024). Moreover, most Cooper’s hawks wintering in cities in our area are visitors from non-urban areas (Boyle, 2011), so it is likely that the immature hawk I observed was a very recent immigrant to urban habitat. One can only imagine the level of knowledge and understanding of the environment possessed by hawks living in more natural landscapes.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the study was purely observational.
Author contributions
VD: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bednarz J. C. (1988). Cooperative hunting Harris’ hawks (Parabuteo unicinctus). Science 239, 1525–1527. Available at: http://links.jstor.org/sici?sici=0036-8075%2819880325%293%3A239%3A4847%3C1525%3ACHIHH%28%3E2.0.CO%3B2-L.
Bildstein K. L. (2006). Raptors: the curious nature of diurnal birds of prey (Ithaca, NY: Cornell University Press).
Biondi L. M., Medina A., Bonetti E. A., Paterlini C. A., Bó M. S. (2024). Cognitive flexibility in a generalist raptor: a comparative analysis along an urbanization gradient. Behav. Ecol. 35, arae025. doi: 10.1093/beheco/arae025
Boal C. W. (1997). The urban environment as an ecological trap for Cooper’s Hawks (Phoenix, AZ: University of Arizona).
Bonta M., Gosford R., Eussen D., Ferguson N., Loveless E., Witwer M. (2017). Intentional fire-spreading by “Firehawk” raptors in Northern Australia. J. Ethnobiol. 37, 700–718. doi: 10.2993/0278-0771-37.4.700
Boyle W. J. H. (2011). The birds of New Jersey: status and distribution (Princeton, NJ: Princeton University Press).
Bugnyar T., Stöwe M., Heinrich B. (2004). Ravens, Corvus corax, follow gaze direction of humans around obstacles. Proc. R. Soc London. B: Biol. Sci. 271, 1331–1336. doi: 10.1098/rspb.2004.2738
Cooper D. S., Shultz A. J., Şekercioğlu Ç.H., Osborn F. M., Blumstein D. T. (2022). Community science data suggest the most common raptors (Accipitridae) in urban centres are smaller, habitat-generalist species. Ibis 164, 771–784. doi: 10.1111/ibi.13047
Cristol D. A., Switzer P. V., Johnson K. L., Walke L. S. (1997). Crows do not use automobiles as nutcrackers: putting an anecdote to the test. Auk 114, 296–298. doi: 10.2307/4089172
Dietrich J. (1982). Zur Ökologie des Habichts – Accipiter gentilis – im Stadtverband Saarbrücken (Saarbrücken, Germany: Univercity of the Saarland).
Dumas C., St-Louis B., Routhier L. (2006). Decision making and interference in the domestic cat (Felis catus). J. Compar. Psychol. 120, 367–377. doi: 10.1037/0735-7036.120.4.367
eBird (2024). eBird: An online database of bird distribution and abundance (Itaca, NY: Cornell Lab of Ornithology). Available online at: http://www.ebird.org (Accessed November 25, 2024).
Estes W. A., Mannan R. W. (2003). Feeding behavior of Cooper’s Hawks at urban and rural nests in southeastern Arizona. Condor 105, 107–116. doi: 10.1093/condor/105.1.107
Grobecker D. B., Pietsch T. W. (1978). Crows use automobiles as nutcrackers. Auk 95, 760–761. doi: 10.2307/4089172
Harju S. M., Olson C. V., Hess J. E., Bedrosian B. (2018). Common raven movement and space use: influence of anthropogenic subsidies within greater sage-grouse nesting habitat. Ecosphere 9, e02348. doi: 10.1002/ecs2.2348
Headland T., Colombelli-Négrel D., Callaghan C. T., Sumasgutner S. C., Kleindorfer S., Sumasgutner P. (2023). Smaller Australian raptors have greater urban tolerance. Sci. Rep. 13, 11559. doi: 10.1038/s41598-023-38493-z
Jokimäki J., Kaisanlahti-Jokimäki M. L. (2023). Urban birds using insects on front panels of cars. Birds 4, 15–27. doi: 10.3390/birds4010002
Krivitsky I., Reutsky A. (2003). Птицы в нашем городе [Birds in our city] (Kharkiv, Ukraine: Nauka).
Malan G., Robinson E. (1999). The diet of the Black Sparrowhawk Accipiter melanoleucus (Aves: Accipitridae) in South Africa: hunting columbids in man-altered environments. Durban Museum Novitates 24, 43–47.
Marzluff J. M., Angell T. (2007). In the company of crows and ravens (New Haven, CT: Yale University Press).
Morton F. B., Gartner M., Norrie E. M., Haddou Y., Soulsbury C. D., Adaway K. A. (2023). Urban foxes are bolder but not more innovative than their rural conspecifics. Anim. Behav. 203, 101–113. doi: 10.1016/j.anbehav.2023.07.003
Nihei Y. (1995). Variations of behaviour of carrion crows Corvus corone using automobiles as nutcrackers. Japan. J. Ornithol. 44, 21–35. doi: 10.3838/jjo.44.21
Nihei Y., Higuchi H. (2001). When and where did crows learn to use automobiles as nutcrackers. Tohoku Psychologica Folia 60, 93–97. Available at: http://hdl.handle.net/10097/54761.
Planillo A., Kramer-SChadt S., Malo J. E. (2015). Transport infrastructure shapes foraging habitat in a raptor community. PLoS One 10, e0118604. doi: 10.1371/journal.pone.0118604
Rosenfield R. N., Mannan R. W., Millsap B. A. (2018). “Cooper’s hawks: the bold backyard hunters,” in Urban raptors: ecology and conservation of birds of prey in cities. Eds. Boal C. W., Dykstra C. R. (Island Press, Washington, D.C), 93–109.
Roth T. C., Lima S. L. (2003). Hunting behavior and diet of Cooper’s hawks: an urban view of the small-bird-in-winter paradigm. Condor 105, 474–483. Available at: https://www.jstor.org/stable/1370672.
Rutz C. (2006). Home range size, habitat use, activity patterns and hunting behaviour of urban-breeding Northern Goshawks Accipiter gentilis. Ardea 94, 185–202.
Santiago J. M. (2025). Why and how Egyptian vultures use tools for feeding. Ethol. Ecol. Evol. 37, 128–138. doi: 10.1080/03949370.2024.2437351
Santos S. M., Carvalho F., Mira A. (2011). How long do the dead survive on the road? Carcass persistence probability and implications for road-kill monitoring surveys. PloS One 6, e25383. doi: 10.1371/journal.pone.0025383
Smith C. R., Colbert-White E. (2023). Raptors’ Natural history influences their response to the string-pull task. Int. J. Compar. Psychol. 36. Available at: https://escholarship.org/uc/item/2rv3c28t ( Accessed April 2, 2025).
Stoyanova Y., Stefanov N., Schmutz J. K. (2010). Twig used as a tool by the Egyptian vulture (Neophron percnopterus). J. @ Raptor Res. 44, 154–156. doi: 10.3356/JRR-09-20.1
Thiollay J. M. (1994). “Family accipitridae (Hawks),” in Handbook of the birds of the world. Vol. 2. New World vultures to Guineafowl. Eds. del Hoyo J., Elliott A., Sargatal J. (Lynx Edicions, Barcelona, Spain), 52–205.
Keywords: Accipiter cooperi, complex behavior, Cooper’s hawk, mental map, predation, raptor, traffic
Citation: Dinets V (2025) Street smarts: a remarkable adaptation in a city-wintering raptor. Front. Ethol. 4:1539103. doi: 10.3389/fetho.2025.1539103
Received: 03 December 2024; Accepted: 07 April 2025;
Published: 23 May 2025.
Edited by:
Klaudia Modlinska, Polish Academy of Sciences, PolandReviewed by:
Gonzalo Ángel Ramírez-Cruz, National Autonomous University of Mexico, MexicoBrandon Mak, Technical University of Munich, Germany
Francisco Guil, Ministry for the Ecological Transition and the Demographic Challenge (MITERD), Spain
Copyright © 2025 Dinets. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vladimir Dinets, ZGluZXRzQGdtYWlsLmNvbQ==