 Molly Kent1
Molly Kent1 Brendan Crockett2Aditya Narayanan2Isabel Dilandro2Ava Wagner2Andrés Mauco2Gabriella Handford1Paean Luby2
Brendan Crockett2Aditya Narayanan2Isabel Dilandro2Ava Wagner2Andrés Mauco2Gabriella Handford1Paean Luby2 Richard Hunter3Troy Richter3Jonathan Richardson2Olivia Harding2
Richard Hunter3Troy Richter3Jonathan Richardson2Olivia Harding2 Joanna Jacob2
Joanna Jacob2 Sarah Catherine Hartvigsen2
Sarah Catherine Hartvigsen2 Kelly Lambert2*
Kelly Lambert2*- 1Department of Biology, Virginia Military Institute, Lexington, VA, United States
- 2Departments of Biology and Psychology, University of Richmond, Richmond, VA, United States
- 3Department of Psychology, University of Massachusetts Boston, Boston, MA, United States
Although chronic stress is typically associated with disease and negative outcomes, wild animals often survive—and even thrive—in environments marked by recurring threats. In this study, we investigated neurobiological differences in vigilance-associated and stress-coping systems, as well as resilience, in wild and laboratory-bred rats (Rattus norvegicus) to explore potential survival-related adaptations in wild populations. Consistent with previous findings, wild rats exposed to chronic environmental stressors exhibited heightened hypothalamic-pituitary-adrenal axis activity, evidenced by enlarged adrenal glands and elevated fecal metabolites of glucocorticoids. Brains were processed using thionin staining for neural and glial cell counts and immunohistochemistry for immunoreactive targets. Wild rats exhibited neuronal and glial cell modifications in vigilance-related cortical regions, including the auditory and piriform cortices. Additional adaptations included increased glial cell density in the lateral habenula—a region implicated in coping appraisal—and enhanced microvascular density in the hippocampus. These findings underscore the value of incorporating wild animals exposed to naturalistic stressors into the study of adaptive and maladaptive stress responses. Integrating wild and laboratory models may yield translational insights for developing therapeutic strategies targeting stress- and anxiety-related disorders.
1 Introduction
Laboratory-housed rats live in highly controlled environments with limited engagement opportunities, leading to predictable, sedentary behavior (Makowska and Weary, 2016). In contrast, wild rats face continuous high-risk challenges, requiring heightened vigilance to detect and respond to environmental threats such as predators, toxins, and other hazards (Koizumi et al., 2018; Lambert, 2023; Clinchy et al., 2013). By comparing selectively-bred laboratory rats to their wild Rattus norvegicus counterparts, we can gain a deeper understanding of how animals maintain vigilance in response to the unpredictable threats encountered in natural habitats.
In the mid-20th century, neuroscience pioneer Curt Richter compared adrenal gland weights of wild rats to laboratory R. norvegicus, reporting that the wild rats’ adrenal glands were approximately 300% larger than those of laboratory populations (Richter, 1949). This finding was recently replicated when our lab reported that the adrenal glands of urban wild-trapped R. norvegicus were 320% larger than weight and sex-matched laboratory-bred rats, accompanied by a 600% increase in fecal corticosterone (CORT) metabolites (FCMs; Jacob et al., 2022). These observations suggest that heightened hypothalamic-pituitary-adrenal (HPA) axis activity in wild rats is a neurobiological correlate of enhanced vigilance, defined as sustained attention accompanied by tonic alertness (Oken et al., 2006). An understanding of the neurobiological correlates of prolonged threat assessment in wild habitats is informative, considering that survival in the wild depends on appropriate levels of vigilance (Karaer et al., 2023).
In addition to reported adrenal gland differences between wild and domestic rats, brain structures involved in survival-related behavior also appear to differ between wild and laboratory rats. For example, larger amygdala volumes have been reported in wild rats and rabbits compared to domestic species (Koizumi et al., 2018; Brusini et al., 2018); further, in animals exposed to chronic stress, neural atrophy has been observed in brain areas such as the hippocampus (McEwen et al., 2016). Despite the documented association between chronic stress and brain atrophy in specific brain structures, wild rats in our previous research exhibited relatively larger brain volumes and increased neuronal density in the cerebellum, a brain area that is important for movement adaptation (Jacob et al., 2022). Together, these findings point to the existence of significant neurobiological adaptations to heightened vigilance in wild rats (Carrilho et al., 2024; Koizumi et al., 2018). Interestingly, a recent study reported smaller cerebella and comparable numbers of Purkinje neurons in wild R. norvegicus compared to laboratory strains; however, the wild rats were F3 generations from wild-trapped rats, different than wild-trapped rats (Waner-Mariquito et al., 2025). This discrepancy suggests that, whereas domestication may have an impact on rat cerebella volume, exposure to wild habitats throughout the rats’ development is also an influential factor (Jacob et al., 2022).
Neuroendocrine adaptations to chronically stressful environments may be driven by key molecular mechanisms such as mineralocorticoid receptor (MR) activity and expression. Whereas glucocorticoid receptors (GRs) are located throughout the brain, MRs are localized in the limbic areas, especially the hippocampus and amygdala (ter Heegde et al., 2015). Within the hippocampus, the highest MR concentration is in the CA2 region, an area associated with plasticity functions, especially related to memory, social recognition, and coping appraisal (McCann et al, 2021; Joëls and de Kloet, 2017). MRs are colocalized with GRs in the hippocampus but have a higher affinity (i.e., 10 times higher) to glucocorticoids than observed in GRs (de Kloet, 2022). Additionally, higher levels of glucocorticoids have been associated with heightened behavioral vigilance (Voellmy et al., 2014), as well as neural vigilance (i.e., neural reactivity; Henckens et al., 2016), in response to biologically relevant threatening stimuli.
Further, MR expression affects the transcription of genes related to neuroplasticity, including growth factors and cell-adhesion molecules (ter Heegde et al., 2015). Thus, past research suggests that MR engagement is involved in both the initial appraisal of stressful stimuli and subsequent memory systems of the experience, likely influencing future responses to similar stressful contexts (McCann et al., 2021). In clinical investigations, lower MR activation and the putative effect on stress appraisal and coping are associated with psychiatric conditions such as depression, anxiety disorders, and schizophrenia (ter Heegde et al., 2015). In support of these observations, antidepressants such as imipramine have been shown to increase hippocampal MR mRNA expression in the rat hippocampus (Brady et al., 1991, Seckl and Fink, 1992).
Also of interest in understanding stress adaptation in chronically stressful wild habitats is the steroid hormone dehydroepiandrosterone (DHEA). DHEA and its sulfate (DHEA-S) are produced in the adrenal glands of humans and the gonads of rats and mice (Maninger et al., 2009). Additionally, DHEA is produced in the brain and has been found to modulate glutamate [i.e., N-methyl-D-aspartate (NMDA)] and Gamma-Aminobutyric Acid (GABA) neurochemical systems (Maninger et al., 2009). Generally, DHEA is known as a marker of acute stress with additional anti-glucocorticoid properties (Dutheil et al., 2021). Further, DHEA levels are inversely related to the intensity of depressive episodes and may have clinical relevance for other psychiatric and neurologic conditions such as anxiety disorders and dementia (Peixoto et al., 2020; Sripada et al., 2013; Rasmusson et al., 2004; Souza-Teodoro et al., 2024), as well as providing neuroregenerative effects following brain injuries (Rahmani et al., 2013). Consequently, both MR and DHEA functions have been associated with emotional resilience in the face of prolonged stress exposure.
Enriched laboratory environments, consisting of larger physical spaces, social housing, and the introduction of novel stimuli, result in neural and behavioral adaptations that appear consistent with rodents housed in natural habitats (Lambert et al., 2015; Lambert, 2023). For example, rats housed in complex environments have thicker cortices, increased hippocampal neuroplasticity markers, and more complex neuronal structures than laboratory control animals (Mohammed et al., 2002; Han et al., 2022). Additionally, in models of cerebral stroke, animals housed in enriched environments exhibit increased angiogenesis and faster recovery rates (Gresita et al., 2022; Shen et al., 2020). Thus, microvasculature changes related to varying experiences or environments can lead to impactful outcomes such as altered cerebrovascular architecture and vascular pruning (Bogorad et al., 2019).
Although enriched laboratory environments mimic some aspects of natural habitats, they lack the persistent exposure to unpredictable stressors that wild rats face, which may drive more robust neurobiological adaptations. Even natural-enriched laboratory environments that contain elements of species-relevant environmental stimuli fall short of replicating the continuous presence of threats that characterize authentic natural habitats (Bardi et al., 2016). Additionally, in contrast to the constant provision of food, water, shelter, and temperature regulation in the laboratory, these variables are scarce and unpredictable for wild rats. To survive such threats and challenges, wild rats must accurately assess risk and employ appropriate coping and decision-making strategies to respond in a manner that maximizes the probability of survival. Accordingly, heightened sensitivity in sensory and motor cortical areas likely facilitates responsiveness to such real-time threats in wild habitats, leading to enhanced survival (Krubitzer et al., 2011). Also of putative relevance, prior research with varying experiential training suggests that the lateral habenula (LHb), an epithalamic brain structure, is involved in coping appraisal as it facilitates freeze responses in threatening contexts (Hones and Mizumori, 2022). Serving as an interface between internal emotional processes and external context, the LHb facilitates rapid and efficient responses to threatening stimuli (Baker et al., 2022). Thus, in addition to limbic structures such as the hippocampus and amygdala (Koizumi et al., 2018; Zanette et al., 2019), several brain areas are involved in decision-making and stress responses—responses that shape vigilance systems and, ultimately, survival, in wild animals.
Due to their apparent tolerance of high corticosterone levels, wild rats provide a potentially valuable model for the investigation of stress adaptation mechanisms during long-term vigilant states. As previously stated, although high HPA activity is generally related to negative brain and health outcomes (Bremner, 2006; McEwen et al., 2016; Bremner and Wittbrodt, 2020), previous research in our lab indicates that wild rats have healthy brain weights and cerebellar neuronal density, with no evidence of compromised tissue thus far (Jacob et al., 2022). Given these findings, the current study further investigated the neural substrates of vigilance and resilience in wild rats. Specifically, male and female wild and laboratory-matched rats were evaluated to determine the impact of their respective habitats on the following targeted mechanisms of neural vigilance and resilience: (1) hippocampal MR and GR immunoreactivity, (2) limbic and cortical microvasculature density, and (3) neuronal/glial ratios in cortical areas important for responsivity to stressful stimuli. Additionally, markers of HPA activity were assessed, including CORT and DHEA fecal metabolites, as well as relative adrenal weights. Due to their role in both stress and immune functions, spleen weights were also collected. Based on prior findings, it was hypothesized that the wild rats would have higher levels of MR-immunoreactive (MR-ir), region-specific modifications of GR-immunoreactive (GR-ir), increased microvasculature coverage (MVC), and evidence of altered neuronal and glial cell density in targeted cortical areas associated with responses to external threats (e.g., motor, somatosensory, pyriform, anterior cingulate, and auditory cortical areas), as well as modifications in the bed nucleus of the stria terminalis (BNST), LHb, and basolateral amygdala (BLA). Spleens and adrenal glands were hypothesized to be larger in wild rats due to increased antigen and threat exposure. An enhanced understanding of these mechanisms in wild rats offers critical insights into stress adaptation that laboratory models alone cannot provide.
2 Method
2.1 Animals
Over a three-month period during the summer of 2021, male (n=4) and female (n=7) wild rats (R. norvegicus) were live-trapped in urban Richmond, VA, USA, using Tomahawk live traps, Model 603.5 (Tomahawk Live Traps, Hazelhurst, WI). Once the wild animals were anesthetized, ages were estimated using a combination of body weight and a visual inspection of the developmental stage of reproductive organs in males (e.g., descended testicles in males indicated sexual maturity). In addition to tracking body weight, the viscera of the trapped female rats were examined and further dissected to confirm that the nipples, uterus, and ovaries showed no evidence of pregnancy, parturition, or lactation. One female and three males were estimated to be juveniles or pre-adults based on these criteria. As described in the results section, body weight ratios and covariates were used to statistically control for varying body weights.
Laboratory control Long Evans rats (Male = 4, Female = 7; Inotiv, Lafayette, IN) were weight- and sex-matched to the wild rats. Rats were acclimated to standard laboratory housing conditions and were housed in ventilated rat cages measuring 43cm (L) x 34cm (W) x 20cm (H) (Innovive, San Diego, CA)– for a minimum of five days prior to tissue collection. Males were pair-housed upon arrival; additionally, four females were pair-housed, whereas three females were housed together in a larger cage [60cm (L) x 30cm (W) x 30cm (H)] to accommodate the larger group of animals following their arrival to the lab. All cages were lined with pine bedding and ad libitum access to a standard rat chow diet and water was provided for the laboratory rats. The vivarium space was maintained on a 12-hour light cycle from 7:00 AM to 7:00 PM, with temperature maintained at 22°C ± 1°C and humidity between 45% and 55%. This study was carried out in accordance with the recommendations of, and approved by, the University of Richmond’s Institutional Animal Care and Use Committee.
For hormone and gross anatomy assessments, both male and female animals were included in the statistical analyses. For neural/cellular quantifications, including immunohistochemical experiments, only female animals were included in statistical analyses due to a tissue processing error in the males.
2.2 Trapping and tissue collection
The traps were pre-baited for 48 hours and set at approximately 8:00 PM following the initial exposure period. All traps were retrieved the following morning at approximately 8:00 AM. Following capture, rats were transported in their secured live trap to the outdoor field laboratory at the University of Richmond. After an initial sedation with isoflurane, both laboratory and wild rats were given an intraperitoneal (ip) injection of 2,2,2-tribromoethanol (400 mg/kg, previously marketed as Avertin™). Subsequent responses were measured through a toe- or tail-pinch; once the animals were determined to be nonresponsive, they were euthanized via rapid guillotine.
Brains were immediately extracted and hemisected along the cerebral fissure. The right hemisphere was snap-frozen on dry ice for another ongoing investigation focused on transposable element (TE) assay assessment. The left hemisphere was postfixed in 4% paraformaldehyde for 24 hours, then transferred sequentially into 10%, 20%, and 30% sucrose solutions for cyroprotection.
Spleens and adrenal glands were excised and individually placed into pre-weighed, labeled, 5mL polypropylene screw-cap tubes stored on wet ice. Organ weights were then collected on fresh tissue by weighing the vial containing the dissected organ and subtracting the previously recorded initial vial weight. This dissection and weighing procedure minimized exposure to any pathogens contained within the organs of the wild rats.
2.3 Hormone assessments
Glucocorticoids (primarily CORT in rodents, measured here via FCMs), as well as fecal DHEA metabolites (FDMs) were extracted from fecal boluses and evaluated using an ELISA kit (Enzo Life Sciences, Farmingdale, NY). For wild rats, fecal samples were collected from the rectum or distal colon during tissue collection and immediately stored at -80°C in microcentrifuge tubes until analysis. Following arrival to the vivarium, fresh fecal samples were collected from the laboratory animals prior to being placed in new home cages. To minimize circadian effects, the fecal collection period coincided with a similar time of day (appx. 10:00 AM). The rationale for collecting fecal samples immediately upon arrival to the laboratory was to capture the circulating CORT and DHEA levels during transport stress, as an attempt to mimic the trap and relocation stress in the wild rats (Jacob et al., 2022). Previous work in our laboratory indicates that CORT and DHEA levels lag approximately 12 hours behind circulating blood concentrations. This differs from the more immediate peak circulating concentrations in the blood following exposure to stressful stimuli (Bardi et al., 2010).
To prepare samples for analysis, the fecal samples were removed from cryo-storage and allowed to thaw to room temperature on the benchtop in their closed vials. Vials containing the fecal samples were then weighed and placed in an enclosed incubator set to 65°C ± 5°C situated inside a biological safety cabinet, with the snap tops set slightly ajar to allow for moisture to escape and dried overnight. The following day, the dried fecal samples were weighed again and assessed for adequate moisture loss determined by an approximate 50% reduction in weight and a dry, hard consistency.
To extract the CORT and DHEA metabolites, a slurry was made using the ratio of 0.09 g fecal material:1.0 mL methanol, adjusted to each sample’s final dried weight. A metal straight-tip dissection probe was used to break up the dried fecal samples to fully incorporate the fecal material into the methanol mixture. To prevent cross-contamination, the probe was sanitized between samples using fresh methanol or 100% ethanol and wiped dry. Following homogenization, the samples were allowed to sit at room temperature for up to 30 minutes while the solids separated and sank to the bottom of the solution.
Samples were then analyzed according to the Enzo Life Sciences ELISA kit protocol (Farmingdale, NY). Briefly, 20 µl of the supernatant (i.e., the clear liquid above the settled solids) was drawn from each sample, and after ensuring no solid material was contained in the aliquot, it was placed in a clean, labeled microcentrifuge tube. Assay buffer was added at the recommended 1:20 dilution and mixed well before being plated in duplicate. The final dilution in the wells equaled 1:40 after following the manufacturer’s instructions. Concentrations were measured against a standard curve and prepared per the manufacturer’s protocol. Plates were read on a BioTek ELx800 Microplate Reader with BioTek Gen5 detection software (BioTek Instruments, Winooski, VT) at 405 nM. Final concentrations (pg/mL) were calculated by converting net Optical Density (OD) values (sample OD value – blank OD value) via extrapolation values calculated based on the standard curve. A coefficient of variation (CV) limit (±10%) was set for OD values between duplicate wells for each sample. If samples fell above the standard curve, they were diluted and repeated to ensure alignment with the kit parameters. According to the manufacturer, both the CORT and DHEA kits exhibit high specificity for the target hormones, with cross-reactivity to structurally related steroids of less than 1%.
2.4 Immunohistochemistry and cytoarchitectural analyses
Due to tissue integrity challenges during the perfusion process, histological experiments were only conducted on female wild and female laboratory rats. The left cerebrum of each animal was mounted by immersing the posterior portion of the hemisphere into optimal cutting temperature (OCT) compound and then positioning it in the cryostat (HM525 NX Thermo Fischer; Waltham MA) set to -25°C. Once thoroughly frozen, 40 µm-thick free-floating sections were collected from selected regions, including the anterior hippocampus, BLA, LHb, auditory cortex, cingulate cortex, motor cortex, piriform cortex, and somatosensory cortex and subsequently placed in Phosphate Buffered Saline (PBS) + 0.1% sodium azide. For each region of interest, and for each histological procedure, duplicate sections were separated by every 6th consecutive section, resulting in a minimum distance of 200µm between sections to ensure each cell was only counted once. All target regions were assessed for neuron and glial cell densities via thionin-stained cells.
For thionin-stained tissue (utilized to visualize cell bodies), the sections were mounted onto 25x75x1 mm gelatin-coated slides (Globe Scientific, Mahwah, NJ) and dried at room temperature overnight before being exposed to the thionin histological protocol (0.1% dilution) and cover-slipped using Permount mounting medium (Fisher Scientific). Cellular counts were gathered from the targeted regions (i.e., dorsal hippocampus, motor cortex, LHh, piriform cortex, auditory cortex, anterior cingulate cortex, and BLA). Brain tissue was analyzed using the Zeiss Axioscope M.2 light microscope with the software Neurolucida 360 (MBF Bioscience, Williston, VT, USA). Neurons and glial cells (broadly defined) were counted using unique digital markers assigned to each cell within a defined 200x300 µm visual field set in the region of interest at 40x magnification. Cell counts were collected from a minimum of two tissue sections (and an average of four sections) per region for each animal and averaged for a final representative estimate.
For the immunohistochemistry processing, all free-floating sections were washed in PBS for 10 minutes to remove any residual paraformaldehyde and sodium azide. Tissue designated for GR, MR, and CD-31 (a marker of microvasculature used in addition to microvasculature-MR staining) was exposed to an antigen retrieval step consisting of placing sections in PBS containing 10mM sodium citrate for 30 minutes in a 75°C water bath. The sections were then allowed to cool to room temperature before proceeding with the staining protocol. Immunoreactivity was assessed using the following primary antibodies and concentrations: anti-mineralocorticoid receptor [rMR1-18 9C2 IgG1; 1:400 dilution gifted from Gomez-Sanchez Laboratory, University of Mississippi Medical Center (Gomez-Sanchez et al., 2006)], anti-glucocorticoid receptor (GR Ab-226; 1:750 dilution; Signalway Antibody, Greenville, MD, USA), and anti-CD-31 (Abcam TLD-3A12, Eugene OR, dilution of 1:100). To visualize immunoreactivity, sections were exposed to 3,3′-Diaminobenzidine (i.e., DAB substrate) for approximately 10 minutes. Stained sections were mounted onto gelatin-coated slides and cleared and cover slipped with Permount (Fisher Scientific). GR-ir and MR-ir cells were quantified using Neurolucida 360 software (MBF Bioscience, Williston, VT, USA) at 40x magnification in the following regions: cornu ammonus (CA) -1, -2, and -3 of the hippocampus, auditory cortex, BLA, and anterior cingulate cortex. A minimum of two tissue sections per region and an average of four sections per hemisphere were evaluated and cell counts were averaged for a final representative estimate. A standard-sized field of vision (i.e., 200 x 300 um) was used for all quantifications at 40x. For all targeted proteins, negative controls—prepared by omitting the primary antibody during the staining procedure—were included to verify the specificity of the observed immunoreactivity.
For MVC analyses, the percent area of stained microvasculature within the anterior cingulate cortex and the dentate gyrus in MR-ir or CD-31-immunoreactive (CD-31-ir) tissue was determined using an image thresholding tool to highlight targeted immunoreactive tissue (BIOQUANT Image Analysis Co., Nashville, TN, USA) at 40x magnification. The dependent variable for MVC analysis was the percent of visual field (200x300 µm) containing MR-ir or CD-31-ir tissue, which was determined with Ilastik interactive machine learning image classification system (Ilastik.com).
2.5 Statistical analyses
Statistical analyses were conducted using SPSS Statistics, version 29 (IBM). Endocrine data were analyzed via a two-way ANCOVA (2 × 2; sex × environment), with body weight included as a covariate. Where appropriate, Tukey’s post hoc tests were applied for pairwise comparisons. Spleen and adrenal gland data, expressed as gland-to-body weight ratios to account for body weight differences, were analyzed using two-way ANOVAs. Immunohistochemistry results were assessed using two-tailed independent samples t-tests to compare wild and laboratory female rats. Data met the assumptions of normality unless otherwise stated. Statistical significance was set at α = 0.05. Due to the multiple comparisons conducted for the neural assessments conducted in the females, a Bonferroni correction was applied to control the family-wise error rate for t-tests, with each brain region (e.g., Lhb) treated as a separate family. Adjusted alpha levels were calculated as αadjusted = α/m, and individual p-values were evaluated against these thresholds. All figures were generated using GraphPad Prism, version 9.0 (GraphPad Software; www.graphpad.com). Graphs display the mean ± standard error of the mean (SEM), with individual data points shown when appropriate. To reduce redundancy, means for data presented in figures are not repeated in the text. However, for non-significant trends not shown graphically, group means are included in the narrative to facilitate interpretation. Similar to the graphs, all reported means are expressed as mean ± SEM.
3 Results
3.1 Anatomical measurements
A 2 × 2 ANOVA revealed a significant interaction between sex and habitat for spleen-to-body weight ratios (F1,18=6.053, p = 0.024, ηp² = 0.252). Post hoc comparisons indicated no significant difference between male groups; however, wild female rats had significantly heavier spleens relative to body weight compared to laboratory females (Figure 1A). Additionally, a 2 × 2 ANOVA revealed a main effect of habitat on adrenal-to-body weight ratios (F1,18=55.103, p < 0.001, ηp² = 0.754), with wild rats showing significantly greater relative adrenal weights than laboratory rats (Figure 1B).
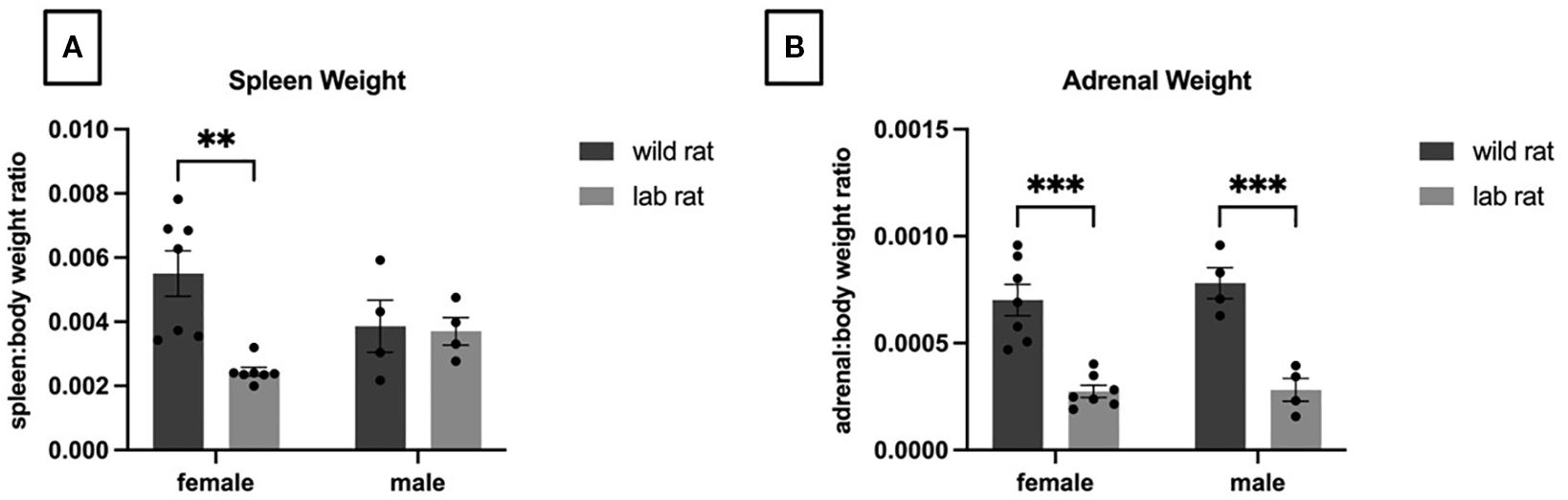
Figure 1. Organ Weights. A 2x2 ANOVA indicated a significant interaction (p=.02) of sex and habitat on spleen weights; specifically, wild females had heavier spleens whereas no effect of habitat was observed in males (A). Focusing on adrenal weights (B), a 2x2 ANOVA indicated a significant effect of habitat with heavier adrenals observed in the wild rats (p=.001). See text for additional statistical information.
3.2 Endocrine assays
A 2 × 2 ANCOVA revealed a significant main effect of habitat on fecal CORT metabolites (FCMs), with wild rats exhibiting significantly higher FCM levels than laboratory rats (F1,17=34.09, p < 0.001, ηp² = 0.670; Figure 2A). No significant differences in FCM were observed between males and females. A separate 2 × 2 ANCOVA indicated a significant main effect of sex on fecal DHEA metabolite (FDM) levels, with females showing significantly higher levels than males (F1,17=11.14, p = 0.004, ηp² = 0.396; Figure 2B). Although not statistically significant, a trend toward higher FDM levels in wild rats compared to laboratory rats was observed (F1,17=3.54, p = 0.077, ηp² = 0.172; also depicted in Figure 2B). A third 2 × 2 ANCOVA revealed significant main effects of both habitat and sex on the DHEA: CORT metabolite ratio. Laboratory rats exhibited a significantly higher DHEA: CORT ratio compared to wild rats (F1,17=8.023, p = 0.011, ηp² = 0.321), and females exhibited a higher ratio than males (F1,17=4.788, p = 0.043, ηp² = 0.220; Figure 2C).
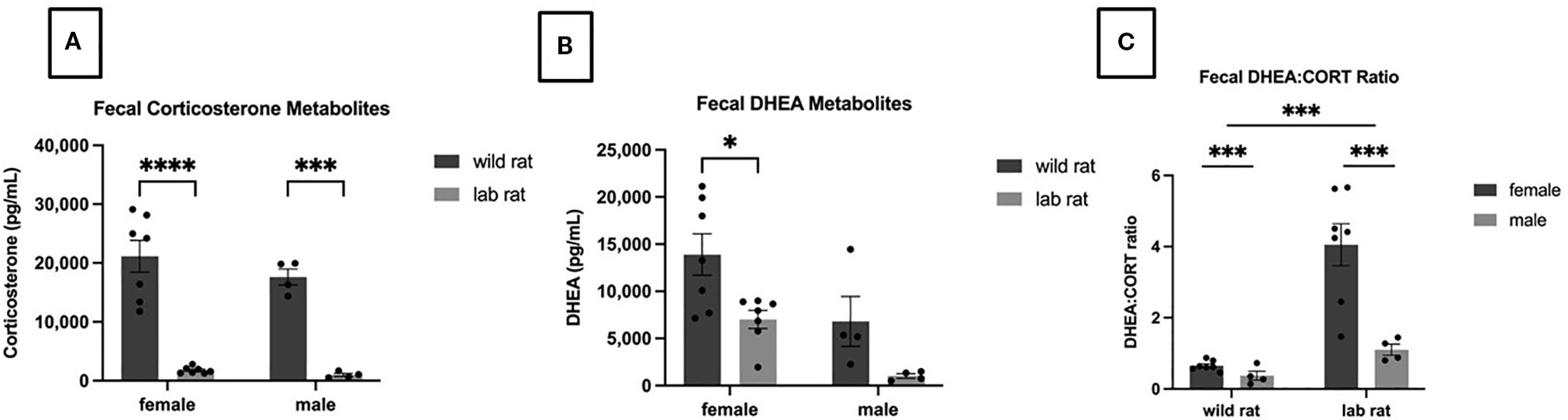
Figure 2. Endocrinological metabolites. A 2x2 ANCOVA revealed a significant effect of habitat on fecal corticosterone metabolites (A); specifically, wild rats had higher FCM levels than lab rats (p=.001). A main effect of sex was observed in the Fecal DHEA metabolites (B) with the females exhibiting higher levels than males and a nonsignificant trend suggested that wild rats had higher FDA levels than lab rats. Focusing on the DHEA/CORT Ratio (C), significant main effects were observed for sex (females had higher ratios; p=.04) and habitat (lab rats had higher ratios; p=.01).
To confirm that transport was a stressor for the laboratory rats—as indicated by elevated FCM levels—a repeated-measures analysis was conducted. FCM levels were significantly higher immediately following transport compared to after five days of acclimation (F1,9=17.27, p = 0.002, ηp² = 0.657). Samples taken from lab rats directly after transport (to control for the relocation factor in the comparison with the wild rats, as described above) exhibited significantly higher FCM levels compared to samples taken after acclimation to the lab for five days F1,9=5.17.27, p=0.002, ηp2=0.657 (arrival = 2278.08±360.46; acclimation = 694.16±107.075; means provided since these data are not included in a figure).
3.3.1 Thionine detection of neuronal and glial cells
Thionine-stained tissue was examined in wild and laboratory females (n = 7 per group). For the lateral habenula, after Bonferroni correction (α = 0.0167 for three comparisons within this brain region family), an independent samples t-test revealed a significant effect of habitat on the number of neurons in the assessed LHb visual field (t7 = 3.352, p = 0.012, d = 2.248), with wild females exhibiting significantly more neurons than laboratory females (Figure 3A). The ratio of neurons to glial cells in the LHb also differed significantly between groups (t7 = 3.409, p = 0.011, d = 2.287), again with wild females showing a higher ratio (Figure 3B). No significant differences were observed in the number of glial cells in the LHb.
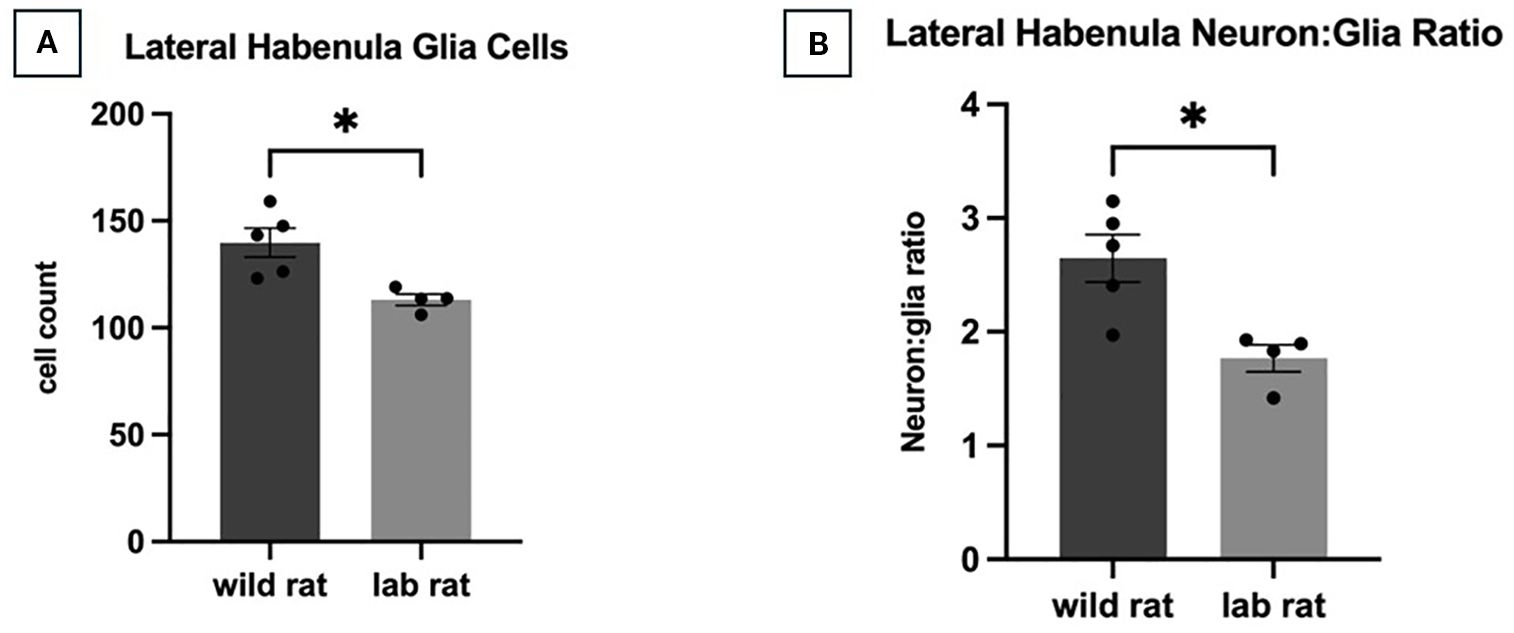
Figure 3. Lateral Habenula Cellular Profile. Compared to laboratory-bred rats, an independent samples t-test revealed that wild rats had more neurons in the sampled visual field of the lateral habenula (A; p=.012) and a higher ratio of neurons to glia (B; p=.011).
In cortical regions, a t-test indicated a significant difference in the number of neurons in the auditory cortex (t9 = 4.602, p = 0.001, d = 2.885), which was revealed following a Bonferroni correction (0.05/4, adjusted α = 0.0125) for four comparisons in this brain region. Wild females had significantly more neurons in this region compared to laboratory females (Figure 4). Although wild females had a 16% increase in glial cells in the auditory cortex relative to laboratory females (wild: M=154.15 ± 7.29; lab: M=131.97 ± 1.67), this difference was not statistically significant. Similarly, no significant difference was found in the neuron-to-glia ratio in the auditory cortex. In the piriform cortex, a Bonferroni-corrected t-test (0.05/3, adjusted α = 0.0167) revealed a significant effect of habitat on glial cell counts (t8 = 3.965, p = 0.004, d = 2.559), with wild rats displaying more glial cells than laboratory rats (Figure 5). No significant differences were found in the number of neurons or the neuron-to-glia ratio in the piriform cortex. In the somatosensory cortex, no significant differences were observed in the number of glial cells (Bonferroni correction: 0.05/3, α = 0.0167), though wild females exhibited a 16.2% increase compared to laboratory females (wild: M=78.68 ± 4.85; lab: M= 67.71 ± 1.76). There were also no significant differences in neuron counts or the neuron-to-glial cell ratio in this region. In the motor cortex, habitat had no significant effect on the number of neurons, glial cells, or the neuron-to-glial cell ratio.
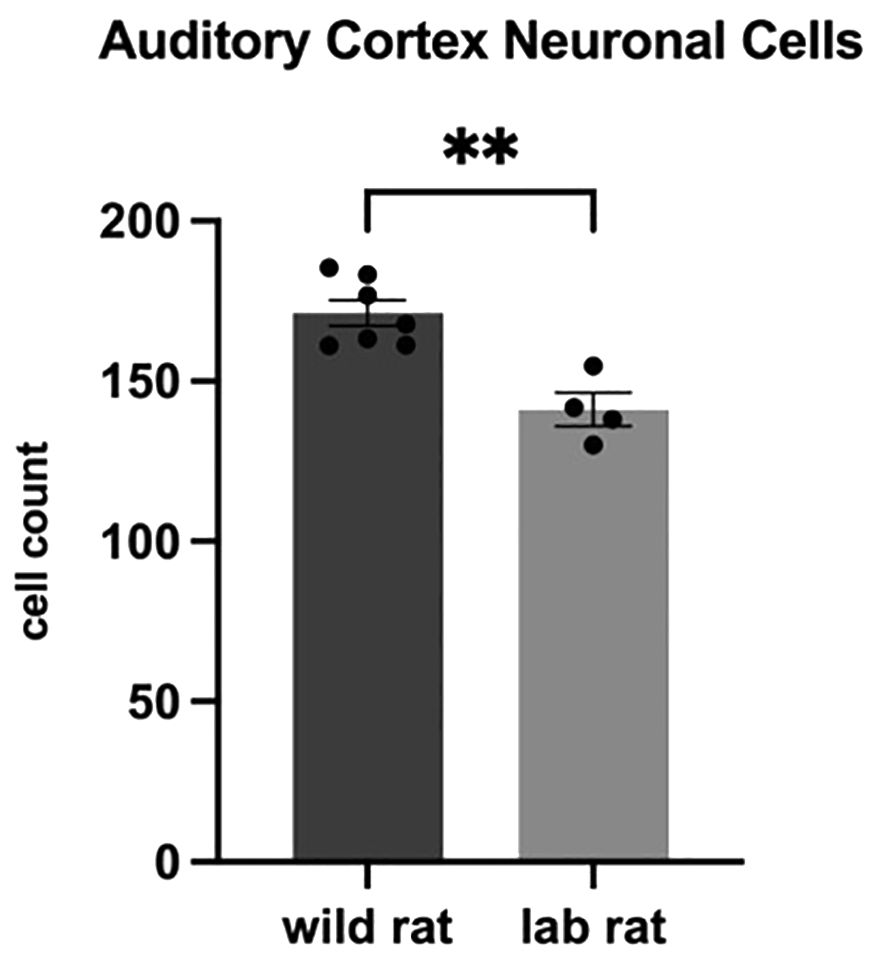
Figure 4. Auditory Cortex Neuronal Density. An independent samples t-test indicated that wild rats had higher numbers of neurons in the targeted visual field of the auditory cortex compared to lab rats (p=.001).

Figure 5. Piriform Cortex Glia Cells. An independent samples t-test indicated that wild rats had higher numbers of glial cells in the targeted visual field of the piriform cortex than lab rats (p=.004).
In the dentate gyrus, no significant differences were found between groups in neuron counts, glial cell counts, or neuron-to-glia ratio. However, wild females had a 26.4% higher glial cell count than laboratory females (wild: M=82.16 ± 6.99; lab: M=64.99 ± 4.29). Similarly, no significant group differences were observed in the bed nucleus of the stria terminalis (BNST) for any of the assessed metrics.
3.3.2 Corticosteroid and mineralocorticoid receptor immunoreactive cells
GR-ir and MR-ir cells were assessed in several brain regions of female rats, including the hippocampal CA1, CA2, and CA3 areas; the motor, auditory, and anterior cingulate cortices; and the basolateral amygdala (BLA).
A t-test with Bonferroni correction using brain area as family (0.05/3, adjusted α = 0.0167) revealed no statistically significant effect of habitat on the number of GR-ir cells in CA2. However, wild rats exhibited a 36% increase in GR-ir cells compared to laboratory rats (wild: M=57.8 ± 4.97; lab: M=42.5 ± 1.94). In CA3, no significant difference was detected after Bonferroni correction (0.05/3, adjusted α = 0.0167; wild: M=49.64 ± 4.04; lab: M=40.0 ± 1.74). Similarly, in CA1, no statistically significant difference was found (wild: M=58.67 ± 6.47; lab: M=44.73 ± 2.90). No significant differences in GR-ir cells were observed in the remaining targeted areas (i.e., motor cortex, auditory cortex, BLA, and anterior cingulate cortex).
Focusing on MR-ir cells, a t-test revealed no significant difference in CA2 after Bonferroni correction (0.05/3, adjusted α = 0.0167). Nevertheless, wild rats displayed a 26% increase in MR-ir cell counts compared to laboratory rats (wild: M=58.59 ± 3.36; lab: M=46.29 ± 6.48). No significant differences in MR-ir cells were observed in CA1 or CA3. Additionally, the ratio of MR-ir:GR-ir cells did not differ significantly between groups in CA1, CA2, or CA3 hippocampal areas.
3.3.3 Microvasculature immunoreactivity
Immunoreactivities to MR and CD-31 were used to assess microvasculature coverage (MVC) in the anterior cingulate cortex and dentate gyrus. A significant effect of habitat on MR-ir cells was observed in the dentate gyrus following Bonferroni correction (0.05/6, adjusted α = 0.00833), with wild females exhibiting significantly higher MR-ir cell counts compared to laboratory females (t9 = 3.403, p = 0.008, d = 2.06; Figure 6). In contrast, no significant difference in MR-ir cells was found between groups in the anterior cingulate cortex. Additionally, CD-31 immunoreactivity in the dentate gyrus did not differ significantly between wild and laboratory females.

Figure 6. Microvasculature. An independent t-test indicated that the wild rats had higher MR immunoreactive tissue in the targeted visual field of the dentate gyrus than the lab rats, indicating enhanced microvasculature (p=.008).
4 Discussion
The goal of this study was to explore neurobiological variables associated with vigilance and emotional resilience in wild R. norvegicus to provide insights into survival-related responses that laboratory models alone cannot fully capture. To differentiate these two concepts, we viewed vigilance as an index of sensory, cortical, and habenular neural adaptations, while resilience was assessed via stress hormone profiles and receptor distributions. The findings replicate and extend earlier reports from our laboratory indicating heightened stress reactivity in wild rats compared to laboratory-bred rodents (Jacob et al., 2022). Specifically, our previous findings indicated that wild rats exhibit a marked increase in the fecal metabolites of CORT and DHEA, alongside heavier adrenal and spleen weights. In the present study, wild rats likewise exhibited significantly higher FCMs than laboratory rats, as well as heavier adrenal and spleen weights. Due to the modest sample sizes (a challenge often associated with field studies utilizing trapped animals), findings that did not meet adjusted significance thresholds should be interpreted with caution, as it was our intent to minimize both Type I and Type II errors. Accordingly, effect size estimates and consistent patterns across multiple variables point to potential biologically relevant trends deserving further investigation (as described below). These current findings, combined with previous research outcomes, suggest that wild rats exhibit adaptive stress responses that support sustained vigilance, likely contributing to their survival in high-risk environments.
In the current study, increased glia cell density and microvascularization (MVC) were observed in the dentate gyrus with no effects observed in the anterior cingulate cortex (it is important to point out that the MVC effect was not observed in the CD-31 data, perhaps a function of the nonperfused but postfixed tissue). While not statistically significant, the observed increase in GR-ir cells in CA2 suggests that the distribution and responsivity of these receptors deserve further investigation. These observations in the hippocampus point to potential neurobiological adaptations related to threat, resilience, and memory processing in this brain area—responses critical for survival in a wild habitat. Additionally, wild rats had a higher neuron-to-glial cell ratio and neuronal density in the LHb, increased neuron density in the auditory cortex, and increased glia in the piriform cortex. Altogether, these results point to altered sensory processing and stress adaptation in wild rats; however, considering that the brain data were collected in female rats, caution should be taken when generalizing to males.
The neurobiological findings in the current study reflect potential mechanisms underlying vigilance and threat responsiveness, traits critical for survival in natural environments. For example, in the current dataset, elevated glucocorticoid levels may lead to increased arousal and sensitivity to environmental threats (Sapolsky et al., 2000; Sapolsky, 2021). The variability in stress hormone responses observed in wild rats may also reflect more fine-tuned regulation of stress hormones during specific environmental challenges (Boyle et al., 2021). However, a limitation of this study is the lack of longitudinal hormone measurements, which makes it unclear whether stress markers decrease in non-threatening contexts, such as relatively safer nesting areas. Interestingly, when wild wood mice were captured in a previous study and relocated to a laboratory habitat, their CORT levels decreased in a dramatic fashion over the course of four weeks, suggesting considerable flexibility in the wild mouse HPA-axis responsivity (Carrilho et al., 2024).
Contrary to our hypothesis, we observed lower DHEA/CORT ratios in wild rats—a finding typically seen in less resilient laboratory rodents, as well as less resilient humans (Morgan et al., 2009; Bardi et al., 2012; Lambert et al, 2020). Although lower DHEA: CORT ratios in wild rats may initially appear inconsistent with prior resilience models derived from laboratory rodents, several biological factors may contribute to this discrepancy. First, differences in reproductive status among wild females—despite the absence of overt pregnancy signs—could affect DHEA production, given gonadal influences on this hormone. Additionally, fecal hormone sampling reflects integrated hormonal activity over time and may not capture transient DHEA peaks or regulatory feedback cycles. It is also plausible that wild rats, through evolutionary adaptations, may have developed altered receptor sensitivity, enhanced local synthesis, or alternative coping mechanisms—variables that require further exploration. These findings underscore the importance of considering ecological and physiological context when interpreting hormonal indicators of resilience.
Extending from neurobiological elements of the HPA system, the LHb, a key region for processing aversive and rewarding stimuli (Mondoloni et al., 2022), exhibited increased glial immunoreactivity in wild rats. This region integrates multimodal sensory inputs and contributes to decision-making processes crucial for responding to environmental threats (Baker et al., 2022). Although not significant, GR- and MR-ir modifications in the CA2 region of the hippocampus may also influence spatial memory and navigation, functions critical for identifying and avoiding potential threats in the wild (Bhasin and Nair, 2022). This area is in need of further investigation, but altered CA2 activity could help explain the well-documented phenomenon of bait shyness in wild rats (Rzóska, 1953). Finally, neuronal and glial modifications were observed in several cortical areas relevant to real-time responses to threats in wild habitats. Specifically, altered cellular profiles in the piriform and auditory cortical areas are noteworthy given their contributions to olfactory and auditory detection of threatening stimuli. Together, these sensory systems provide valuable information about the presence of predators or other threats in an animal’s environment (Bao et al., 2016).
Evidence of increased cerebral MVC in wild rats, particularly in the dentate gyrus, was observed in the current study. Increased MVC may enhance nutrient transport and blood flow essential for real-time responsiveness to environmental challenges (Graff et al., 2021). Additionally, sympathetic-induced peripheral vasoconstriction and increased blood flow facilitate successful escape from predators (Charkoudian and Rabbitts, 2009), a response influenced by MR-mediated changes in blood pressure and vascular remodeling (DuPont and Jaffe, 2017). Although the microvasculature in the anterior cingulate cortex of the wild rats was not statistically significantly different from that of their laboratory counterparts, the values of the wild rats were also higher than those of their laboratory counterparts. Providing support for the role of MVC profiles in vigilant responses, modified cerebral blood flow has been associated with resilient responses to natural predator stimuli in laboratory rats (Kondashevskaya et al., 2022). Consequently, future research should explore the role of microvasculature in facilitating rapid escape behaviors and threat responsiveness. Additionally, although it wasn’t possible to perfuse the animals in the current study due to limitations of working in a field site, the use of perfused wild-trapped brain tissue would be informative for future investigations.
An additional area of interest for future exploration is the potential role of transposable elements (TEs), such as Long Interspersed Nuclear Elements (LINE1), in genomic plasticity and immune function in wild rats adapting to dynamic environments (Zhang et al., 2020; DeRosa et al., 2022; Lapp and Hunter, 2019). Preliminary data from our laboratories, obtained from a separate cohort, suggest possible differences in LINE1 expression between wild and laboratory rats, warranting further targeted investigation.
As previously mentioned, wild animals face persistent stressors such as predation, seasonal changes, and anthropogenic habitat destruction, making the wild rat a valuable model for investigating adaptive responses essential for survival in unpredictable environments (Sopinka et al., 2015). The distinct stress responses observed between wild and laboratory animals underscore the importance of investigating wild models in addition to traditional laboratory models to capture the full spectrum of neurobiological adaptations to chronic stress. Whereas laboratory animals often exhibit altered neural structure (e.g., hippocampal atrophy) and suppressed immune function when exposed to chronic stress (Kim et al., 2015; Selye, 1973), wild animals appear to have evolved unique coping mechanisms that mitigate the adverse health effects associated with high corticosterone levels.
While the current findings are specific to urban wild R norvegicus populations, they offer valuable insights into coping mechanisms shaped by natural ecological conditions. Additional investigations across diverse species and habitats are needed to determine the generalizability of these neurobiological adaptations. Although wild habitats differ markedly from standard laboratory settings—where animals experience limited sensory input and minimal cognitive demands—it is important to integrate species-relevant and ecologically meaningful features into laboratory environments when possible (Lambert, 2023). Ultimately, validating laboratory-based findings with data from wild populations across varied naturalistic contexts is essential for ensuring the translational relevance of preclinical models.
Findings from the current study emphasize the importance of neuroethological approaches—examining wild animals in natural habitats—as a means to uncover adaptive stress responses with potential relevance for human health. While the underlying mechanisms remain to be determined, such resilience may point to biological processes that, if better understood, could eventually inform novel strategies for managing chronic stress in humans. Neuroendocrine and vascular adaptations, in particular, represent promising avenues for future investigations, including modulating glucocorticoid receptor (GR) activity or enhancing MVC support in stress-sensitive brain areas. Additionally, information about corticotropin globulin protein (CBG; also known as transcortin) that binds to circulating corticosterone and prevents its entry into the central nervous system (CNS) will provide further clarification about the wild rats’ management of elevated peripheral CORT levels (Meyer et al., 2016). Thus, while translational applications are speculative at this stage, the wild rat model underscores the importance of investigating animals in naturalistic contexts to reveal resilience and vigilance pathways that may not be detectable in laboratory settings.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by University of Richmond Institutional Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
MK: Conceptualization, Supervision, Data curation, Writing – review & editing, Writing – original draft, Investigation, Methodology, Formal Analysis. BC: Writing – review & editing, Investigation, Methodology, Data curation. AN: Data curation, Methodology, Investigation, Writing – review & editing. ID: Investigation, Data curation, Writing – review & editing, Methodology. AW: Writing – review & editing, Data curation. AM: Investigation, Writing – review & editing, Data curation. GH: Data curation, Writing – review & editing. PL: Data curation, Methodology, Software, Formal Analysis, Investigation, Writing – review & editing. RH: Writing – review & editing, Formal Analysis, Data curation, Investigation, Conceptualization. TR: Data curation, Investigation, Conceptualization, Writing – review & editing. JR: Writing – review & editing, Methodology, Data curation, Investigation. OH: Formal Analysis, Data curation, Investigation, Writing – review & editing. JJ: Project administration, Formal Analysis, Data curation, Investigation, Supervision, Writing – review & editing, Conceptualization, Methodology. SH: Writing – review & editing, Investigation, Data curation, Methodology. KL: Data curation, Methodology, Conceptualization, Writing – review & editing, Investigation, Supervision, Formal Analysis, Writing – original draft, Visualization, Resources, Funding acquisition, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was supported by the University of Richmond School of Arts and Sciences and the Department of Psychology, as well as the Department of Biology at the Virginia Military Institute.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Generative AI was used as an editing tool in places once the manuscript was written.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Baker P. M., Mathis V., Lecourtier L., Simmons S. C., Nugent F. S., Hill S., et al. (2022). Lateral Habenula beyond avoidance: roles in stress, memory, and decision-making with implications for psychiatric disorders. Front. Syst. Neurosci. 16, 826475. doi: 10.3389/fnsys.2022.826475
Bao X., Raguet L. L. G., Cole S. M., Howard J. D., and Gottfried J. A. (2016). The role of piriform associative connections in odor categorization. eLife 5. doi: 10.7554/elife.13732
Bardi M., Hampton J. E., and Lambert K. G. (2010). Fecal dehydroepiandrosterone (DHEA) immunoreactivity as a noninvasive index of circulating DHEA activity in young male laboratory rats. Comp. Med. 60, 455–460.
Bardi M., Rhone A. P., Franssen C. L., Hampton J. E., Shea E. A., Hyer M. M., et al (2012). Behavioral training and predisposed coping strategies interact to influence resilience in male Long-Evans rats: Implications for depression. Stress 15(3), 306–317. doi: https://doi.org/10.3109/10253890.2011.623739
Bardi M., Kaufman C., Franssen C., Hyer M. M., Rzucidlo A., Brown M., et al. (2016). Paper or plastic? Exploring the effects of natural enrichment on behavioural and neuroendocrine responses in long-evans rats. J. Neuroendocrinol 28. doi: 10.1111/jne.12383
Bhasin G. and Nair I. R. (2022). Dynamic hippocampal CA2 responses to contextual spatial novelty. Front. Syst. Neurosci. 16, 923911. doi: 10.3389/fnsys.2022.923911
Bogorad M. I., DeStefano J. G., Linville R. M., Wong A. D., and Searson P. C. (2019). Cerebrovascular plasticity: Processes that lead to changes in the architecture of brain microvessels. J. Cereb. Blood Flow Metab. 39, 1413–1432. doi: 10.1177/0271678X19855875
Boyle S. A., de la Sancha N. U., Pérez P., and Kabelik D. (2021). Small mammal glucocorticoid concentrations vary with forest fragment size, trap type, and mammal taxa in the Interior Atlantic Forest. Sci. Rep. 11, 2111. doi: 10.1038/s41598-021-81073-2
Brady L. S., Whitfield H. J. Jr., Fox R. J., Gold P. W., and Herkenham M. (1991). Long-term antidepressant administration alters corticotropin-releasing hormone, tyrosine hydroxylase, and mineralocorticoid receptor gene expression in rat brain: Therapeutic implications. Journal of Clinical Investigation 87(3), 831–837. https://doi.org/10.1172/JCI115086
Bremner J. D. (2006). Stress and brain atrophy. CNS Neurological Disord. Drug Targets 5, 503–512. doi: 10.2174/187152706778559309
Bremner J. D. and Wittbrodt M. T. (2020). Stress, the brain, and trauma spectrum disorders. Int Rev Neurobiol 152, 1–22. doi: 10.1016/bs.irn.2020.01.004.
Brusini I., Carneiro M., Wang C., Rubin C.-J., Ring H., Afonso S., et al. (2018). Changes in brain architecture are consistent with altered fear processing in domestic rabbits. Proc. Natl. Acad. Sci. United States America 115, 7380–7385.
Carrilho M., Monarca R. I., Aparício G., Mathias M., da L., Tapisso J. T., et al. (2024). Physiological and behavioural adjustment of a wild rodent to laboratory conditions. Physiol. Behav. 273, 114385. doi: 10.1016/j.physbeh.2023.114385
Charkoudian N. and Rabbitts J. A. (2009). Sympathetic neural mechanisms in human cardiovascular health and disease. Mayo Clinic Proc. Mayo Clinic 84, 822–830. doi: 10.4065/84.9.822
Clinchy M., Sheriff M. J., and Zanette L. Y. (2013). Predator-induced stress and the ecology of fear. Funct. Ecol. 27, 56–65. doi: 10.1111/1365-2435.12007
de Kloet E. R. (2022). Brain mineralocorticoid and glucocorticoid receptor balance in neuroendocrine regulation and stress-related psychiatric etiopathologies. Curr. Opin. Endocrine Metab. Res. 24, 100352. doi: 10.1016/j.coemr.2022.100352
DeRosa H., Richter T., Wilkinson C., and Hunter R. G. (2022). Bridging the gap between environmental adversity and neuropsychiatric disorders: The role of transposable elements. Front. Genet. 13, 813510. doi: 10.3389/fgene.2022.813510
DuPont J. J. and Jaffe I. Z. (2017). 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: The role of the mineralocorticoid receptor in the vasculature. J. Endocrinol. 234, T67–T82. doi: 10.1530/JOE-17-0009
Dutheil F., de Saint Vincent S., Pereira B., Schmidt J., Moustafa F., Charkhabi M., et al. (2021). DHEA as a biomarker of stress: A systematic review and meta-analysis. Front. Psychiatry/Frontiers Res. Foundation 12, 688367. doi: 10.3389/fpsyt.2021.688367
Gomez-Sanchez C. E., de Rodriguez A. F., Romero D. G., Estess J., Warden M. P., Gomez-Sanchez M. T., et al. (2006). Development of a panel of monoclonal antibodies against the mineralocorticoid receptor. Endocrinology 147, 1343–1348. doi: 10.1210/en.2005-0860
Graff B. J., Payne S. J., and El-Bouri W. K. (2021). The ageing brain: Investigating the role of age in changes to the human cerebral microvasculature with an in silico model. Front. Aging Neurosci. 13, 632521. doi: 10.3389/fnagi.2021.632521
Gresita A., Mihai R., Hermann D. M., Amandei F. S., Capitanescu B., and Popa-Wagner A. (2022). Effect of environmental enrichment and isolation on behavioral and histological indices following focal ischemia in old rats. GeroScience 44, 211–228. doi: 10.1007/s11357-021-00432-z
Han Y., Yuan M., Guo Y.-S., Shen X.-Y., Gao Z.-K., and Bi X. (2022). The role of enriched environment in neural development and repair. Front. Cell. Neurosci. 16, 890666. doi: 10.3389/fncel.2022.890666
Henckens M. J., Klumpers F., Everaerd D., Kooijman S. C., van Wingen G. A., and Fernández G. (2016). Interindividual differences in stress sensitivity: basal and stress-induced cortisol levels differentially predict neural vigilance processing under stress. Soc. Cogn. Affect. Neurosci. 11, 663–673. doi: 10.1093/scan/nsv149
Hones V. I. and Mizumori S. J. Y. (2022). Response flexibility: The role of the lateral habenula. Front. Behav. Neurosci. 16, 852235. doi: 10.3389/fnbeh.2022.852235
Jacob J., Watanabe S., Richardson J., Gonzales N., Ploppert E., Lahvis G., et al. (2022). Divergent neural and endocrine responses in wild-caught and laboratory-bred rattus norvegicus. Behav. Brain Res. 432, 113978. doi: 10.1016/j.bbr.2022.113978
Joëls M. and de Kloet E. R. (2017). 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: The brain mineralocorticoid receptor: a saga in three episodes. J. Endocrinol. 234, T49–T66. doi: 10.1530/JOE-16-0660
Peixoto C., José Grande A., Gomes Carrilho C., Nardi A. E., Cardoso A., and Barciela Veras A. (2020). Dehydroepiandrosterone for depressive symptoms: A systematic review and meta-analysis of randomized controlled trials. J Neurosci Res 98(12), 2510–2528. doi: 10.1002/jnr.24721.
Karaer M. C., Čebulj-Kadunc N., and Snoj T. (2023). Stress in wildlife: comparison of the stress response among domestic, captive, and free-ranging animals. Front. Veterinary Sci. 10, 1167016. doi: 10.3389/fvets.2023.1167016
Kim E. J., Pellman B., and Kim J. J. (2015). Stress effects on the hippocampus: a critical review. Learn. Memory 22, 411–416. doi: 10.1101/lm.037291.114
Koizumi R., Kiyokawa Y., Mikami K., Ishii A., Tanaka K. D., Tanikawa T., et al. (2018). Structural differences in the brain between wild and laboratory rats (Rattus norvegicus): Potential contribution to wariness. J. Veterinary Med. Science/the Japanese Soc. Veterinary Sci. 80, 1054–1060. doi: 10.1292/jvms.18-0052
Kondashevskaya M. V., Downey H. F., Tseilikman V. E., Alexandrin V. V., Artem’yeva K. A., Aleksankina V. V., et al. (2022). Cerebral blood flow in predator stress-resilient and -susceptible rats and mechanisms of resilience. Int. J. Mol. Sci. 23, 14729. doi: 10.3390/ijms232314729
Krubitzer L., Campi K. L., and Cooke D. F. (2011). All rodents are not the same: a modern synthesis of cortical organization 2011). Brain Behav. Evolution. 78, 51–93. doi: 10.1159/000327320
Lambert K. (2023). Wild brains: The value of neuroethological approaches in preclinical behavioral neuroscience animal models. Neurosci. Biobehav. Rev. 146, 105044. doi: 10.1016/j.neubiorev.2023.105044
Lambert K., Hunter R. G., Bartlett A. A., Lapp H. E., and Kent M. (2020). In search of optimal resilience ratios: Differential influences of neurobehavioral factors contributing to stress-resilience spectra. Front. Neuroendocrinol. 56, 100802. doi: 10.1016/j.yfrne.2019.100802
Lambert K. G., Nelson R. J., Jovanovic T., and Cerdá M. (2015). Brains in the city: Neurobiological effects of urbanization. Neurosci. Biobehav. Rev. 58, 107–122. doi: 10.1016/j.neubiorev.2015.04.007
Lapp H. E. and Hunter R. G. (2019). Early life exposures, neurodevelopmental disorders, and transposable elements. Neurobiol. Stress 11, 100174. doi: 10.1016/j.ynstr.2019.100174
Makowska I. J. and Weary D. M. (2016). The importance of burrowing, climbing and standing upright for laboratory rats. R. Soc. Open Sci. 3, 160136. doi: 10.1098/rsos.160136
Maninger N., Wolkowitz O. M., Reus V. I., Epel E. S., and Mellon S. H. (2009). Neurobiological and neuropsychiatric effects of dehydroepiandrosterone (DHEA) and DHEA sulfate (DHEAS). Front. Neuroendocrinol. 30, 65–91. doi: 10.1016/j.yfrne.2008.11.002
McCann K. E., Lustberg D. J., Shaughnessy E. K., Carstens K. E., Farris S., Alexander G. M., et al. (2021). Novel role for mineralocorticoid receptors in control of a neuronal phenotype. Mol. Psychiatry 26, 350–364. doi: 10.1038/s41380-019-0598-7
McEwen B. S., Nasca C., and Gray J. D. (2016). Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 41, 3–23. doi: 10.1038/npp.2015.171
Meyer E. J., Nenke M. A., Rankin W., Lewis J. G., and Torpy D. J. (2016). Corticosteroid-binding globulin: A review of basic and clinical advances. Horm. Metab. Res. 48, 359–371. doi: 10.1055/s-0042-108071
Mohammed A. H., Zhu S. W., Darmopil S., Hjerling-Leffler J., Ernfors P., Winblad B., et al. (2002). Environmental enrichment and the brain. Prog. Brain Res. 138, 109–133. doi: 10.1016/S0079-6123(02)38074-9
Mondoloni S., Mameli M., and Congiu M. (2022). Reward and aversion encoding in the lateral habenula for innate and learned behaviours. Trans. Psychiatry 12, 3. doi: 10.1038/s41398-022-01857-6
Morgan C. A. 3rd, Rasmusson A., Pietrzak R. H., Coric V., and Southwick S. M. (2009). Relationships among plasma dehydroepiandrosterone and dehydroepiandrosterone sulfate, cortisol, symptoms of dissociation, and objective performance in humans exposed to underwater navigation stress. Biol. Psychiatry 66, 334–340. doi: 10.1016/j.biopsych.2009.04.004
Oken B. S., Salinsky M. C., and Elsas S. M. (2006). Vigilance, alertness, or sustained attention: physiological basis and measurement. Clin. Neurophysiol. 117, 1885–1901. doi: 10.1016/j.clinph.2006.01.017
Rahmani A., Shoae-Hassani A., Keyhanvar P., Kheradmand D., and Darbandi-Azar A. (2013). Dehydroepiandrosterone stimulates nerve growth factor and brain derived neurotrophic factor in cortical neurons. Adv. Pharm. Pharmacol. Sci. 2013, 506191. doi: 10.1155/2013/506191
Rasmusson A. M., Vasek J., Lipschitz D. S., Vojvoda D., Mustone M. E., Shi Q., et al. (2004). An increased capacity for adrenal DHEA release is associated with decreased avoidance and negative mood symptoms in women with PTSD. Neuropsychopharmacology 29, 1546–1557. doi: 10.1038/sj.npp.1300432
Richter C. P. (1949). Domestication of the Norway rat and its implications for the problem of stress. Res. Publications - Assoc. Res. Nervous Ment. Dis. 29, 19–47.
Rzóska J. (1953). Bait shyness, a study in rat behaviour. Br. J. Anim. Behav. 1, 128–135. doi: 10.1016/S0950-5601(53)80011-0
Sapolsky R. M. (2021). Glucocorticoids, the evolution of the stress-response, and the primate predicament. Neurobiol. Stress 14, 100320. doi: 10.1016/j.ynstr.2021.100320
Sapolsky R. M., Romero L. M., and Munck A. U. (2000). How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocrine Rev. 21, 55–89. doi: 10.1210/edrv.21.1.0389
Seckl J. R. and Fink G. (1992). Antidepressants increase glucocorticoid and mineralocorticoid receptor mRNA expression in rat hippocampus in vivo. Neuroendocrinology 55, 621–626. doi: 10.1159/000126180
Shen X., Luo L., Wang F., Yu K., Xie H., Tian S., et al. (2020). An enriched environment enhances angiogenesis surrounding the cingulum in ischaemic stroke rats. Neural Plasticity 2020, 8840319. doi: 10.1155/2020/8840319
Sopinka N. M., Patterson L. D., Redfern J. C., Pleizier N. K., Belanger C. B., Midwood J. D., et al. (2015). Manipulating glucocorticoids in wild animals: basic and applied perspectives. Conserv. Physiol. 3, cov031. doi: 10.1093/conphys/cov031
Souza-Teodoro L. H., Davies N. M., Warren H. R., Andrade L. H. S. G., and Carvalho L. A. (2024). DHEA and response to antidepressant treatment: A Mendelian Randomization analysis. J. Psychiatr. Res. 173, 151–156. doi: 10.1016/j.jpsychires.2024.02.049
Sripada R. K., Marx C. E., King A. P., Rajaram N., Garfinkel S. N., Abelson J. L., et al. (2013). DHEA enhances emotion regulation neurocircuits and modulates memory for emotional stimuli. Neuropsychopharmacology 38, 1798–1807. doi: 10.1038/npp.2013.79
ter Heegde F., De Rijk R. H., and Vinkers C. H. (2015). The brain mineralocorticoid receptor and stress resilience. Psychoneuroendocrinology 52, 92–110. doi: 10.1016/j.psyneuen.2014.10.022
Voellmy I. K., Goncalves I. B., Barrette M. F., Monfort S. L., and Manser M. B. (2014). Mean fecal glucocorticoid metabolites are associated with vigilance, whereas immediate cortisol levels better reflect acute anti-predator responses in meerkats. Hormones Behav. 66, 759–765. doi: 10.1016/j.yhbeh.2014.08.008
Waner-Mariquito R., Williams L. A., Stark R. A., Stryjek R., Modlińska K., Pisula W., et al. (2025). Strain-specific differences in cerebellar anatomy between laboratory and wild rats. Brain Behavior Evol. 17, 1–12. doi: 10.1159/000546604
Zanette L. Y., Hobbs E. C., Witterick L. E., MacDougall-Shackleton S. A., and Clinchy M. (2019). Predator-induced fear causes PTSD-like changes in the brains and behaviour of wild animals. Sci. Rep. 9, 11474. doi: 10.1038/s41598-019-47684-6
Keywords: resilience, vigilance, stress response, coping, wild rodents
Citation: Kent M, Crockett B, Narayanan A, Dilandro I, Wagner A, Mauco A, Handford G, Luby P, Hunter R, Richter T, Richardson J, Harding O, Jacob J, Hartvigsen SC and Lambert K (2025) Comparative neurobiology of vigilance and stress adaptation systems in wild and laboratory rodents (Rattus norvegicus). Front. Ethol. 4:1644812. doi: 10.3389/fetho.2025.1644812
Received: 10 June 2025; Accepted: 31 August 2025;
Published: 23 September 2025.
Edited by:
Enrique Font, University of Valencia, SpainReviewed by:
Raffaele d’Isa, San Raffaele Scientific Institute (IRCCS), ItalyBoróka Bárdos, University of Kaposvar, Hungary
Copyright © 2025 Kent, Crockett, Narayanan, Dilandro, Wagner, Mauco, Handford, Luby, Hunter, Richter, Richardson, Harding, Jacob, Hartvigsen and Lambert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kelly Lambert, a2xhbWJlcnRAcmljaG1vbmQuZWR1