 Johanna Gjøen
Johanna Gjøen Per Jensen
Per Jensen- AVIAN Behavioral Physiology and Genomics Group, IFM Biology, Linköping University, Linköping, Sweden
Domestication involves selection for tameness, which can lead to correlated changes in behavior, morphology, and brain physiology. One part of the brain that has become relatively larger in domesticated chickens, as well as in Red Junglefowl (Gallus gallus) selected for increased tameness, is the cerebellum, which is involved with an extensive variety of behaviors. This study explored the impact of selection for high (HF) or low (LF) fear of humans in Red junglefowl (RJF) on cerebellum-linked traits such as motor coordination, balance, and social navigation. Using an obstacle course, a rotarod test, and a social recognition test we assessed the performance of HF and LF birds from the 14th generation of selection lines. While HF birds completed the obstacle course more quickly (p < 0.001), there were no significant differences in locomotory coordination. LF birds exhibited more reorientation pauses, potentially linked to exploratory behavior and reduced fearfulness. In the rotarod test, the latency time did not reach statistical significance (p = 0.1). The social recognition test revealed a clear social preference in the HF line, with preference for a non-aggressive rooster, but no significant preferences in the LF line. Our findings suggest that the selection for tameness, previously associated with increased cerebellar size, influences the approach to transverse an obstacle course in order to regain social contact, and social navigation as measured by choice of social partner. Furthermore, a possible but non-significant link to motor control, as measured by the ability to stay on a rotarod, was observed. This suggests that the tameness-induced changes in cerebellum may affect social behavior such as social navigation, rather than affecting motor control.
Introduction
Animal domestication refers to the process by which animals adjust genetically to living within a human-controlled environment. Domestication causes a range of phenotypic changes in animals, but the reduction in fear responses towards humans stands out as the most essential, as this allows a life in human presence, hence is needed in order to achieve domestication in the first place (Lindberg et al., 2007; Agnvall et al., 2018; Fitak et al., 2020; Dou et al., 2023). Studies have revealed that domesticated animals often show a range of phenotypic changes, such as decrease in pigmentation, altered estrous, altered body mass, altered tails, floppy ears, reduced brain size, and increased docility, often referred to as the domestication syndrome (Collarini et al., 2022).
Fear responses can be reduced across generations through active and passive selection (Galef, 1970; Nagayama et al., 2018). Tameness can be categorized into two types: active and passive. Active tameness is when animals are motivated to explore humans and interact with them, while passive tameness is a reluctance to avoid humans (Nagayama et al., 2018). Tameness exists to some degree in all domesticated animal species (Albert et al., 2009).
Classic experiments, such as Belyaev’s long-term fox study, demonstrated that selecting animals for tameness alone produces a suite of traits typical to domestication syndrome, including altered morphology, physiology, and behavior. Follow-up studies in mink (Neovison vison) and rats (Rattus Norwegicus) confirmed that selection for reduced fear or aggression reliably induces domestication-linked changes, supporting tameness as a pivotal driver in domesticated species evolution (Belyaev, 1979; Naumenko et al., 1989; Trut, 1998, 1999; Malmkvist and Hansen, 2002; Trapezov et al., 2012).
Applying the same strategy to Red Junglefowl (Gallus gallus), we bred lines for low and high fear of humans. Over the generations, birds selected for low fear rapidly developed traits matching those of domestic chickens: increased body and egg size, more dominant behavior, higher feed efficiency, and smaller overall brain size. Crucially, these low-fear birds also showed relatively enlarged cerebellums, linking tameness selection not just to behavior and morphology, but to cerebellar adaptations that may shape social and motor traits in domestication contexts (Agnvall et al., 2012, 2014, 2015; Agnvall and Jensen, 2016; Henriksen et al., 2016; Agnvall et al., 2017, 2018; Katajamaa and Jensen, 2020; Katajamaa et al., 2021).
One of the defining aspects of the domestication syndrome is a reduction in brain size relative to body mass (Kruska, 2007; Rehkamper et al., 2008; Wright et al., 2020). However, not all parts of the brain follow this pattern, and in fact, cerebellum has been found to be relatively larger in domesticated chickens (Henriksen et al., 2016) and our previous studies found that the same was true in Red Junglefowl selected for reduced fear of humans (Katajamaa and Jensen, 2020; Katajamaa et al., 2021; Gjøen et al., 2023). The cerebellum is a complex structure traditionally associated with motor control but now recognized for being important in cognitive and social processes as well. Studies on humans suggest that the cerebellum contributes to functions such as social cognition and emotional regulation (Van Overwalle et al., 2020).
Studies on cerebellar structure in birds suggest its design is closely linked to ecological needs and behavioral adaptations. For instance, large-brained birds like crows, parrots, and woodpeckers exhibit larger trigeminal and visual regions of the cerebellum, aligning with their visually guided, goal-directed behaviors. In contrast, nocturnal predators like owls show expanded vestibular and somatosensory regions to support their hunting strategies (Sultan, 2005). The cerebellum also has an important role in sorting sensory inputs and coordinating movement (Sultan and Glickstein, 2008). Bird species known to use tools have a more folded cerebellar cortex, indicating that cognitive abilities may be linked to increased surface area rather than overall size (Iwaniuk et al., 2006). Cerebellum plays a role in social behavior, both in humans and other animals (Parkins, 1997; Molinari et al., 2009; Leggio and Molinari, 2015; Heleven et al., 2019; Hosaka et al., 2024). For example, in mice, the cerebellum has been implicated in social reward mechanisms, with studies showing its direct influence on regions involved in social reward processing (Carta et al., 2019) and it has been linked to fear memory (Sacchetti et al., 2002; Frontera et al., 2020; Katajamaa et al., 2021; Stingo-Hirmas et al., 2022) as well as avoidance behavior (Dahhaoui et al., 1990). Hence, the cerebellum is important in shaping behavior and cognition, and the fact that its relative size is preserved or even increased during chicken domestication and during selection for increased tameness suggests that some aspects of its functions are essential for a life among humans.
Here, using two lines selected for low (LF) and high (HF) fear of humans known to differ in relative cerebellum size, we investigate the effects of this selection on social and motor behaviors that have previously been linked to variations in cerebellar functions. Recent research on mice suggests that the cerebellum supports social behaviors through distinct mechanisms separate from those governing motor control (Van Der Heijden, 2024). However, the current study does not include direct anatomical measurements; instead, our approach examines cerebellum-linked behaviors as indicators of underlying neural adaptations related to domestication and selection for reduced fear of humans. The aim of the present study was to assess locomotory control, motor balance and social cognition in the two selection lines.
Materials and methods
Ethical note
The experiments were approved by the Linköping Animal Ethics Committee under license no. 10492-2023, with all procedures conducted accordingly.
Animals and housing
A total of 40 Red Junglefowl (20 LF; 60% males and 40% females, 20 HF; 50% males and 50% females) were used in this study. All chicks originated from established selection lines. Birds were housed in stable, mixed-sex groups throughout rearing and testing. Chicks were hatched at the same facility and raised under identical environmental and nutritional conditions, including standardized pen dimensions, feeding schedules, and lighting regimes. Prior to the present study, the birds participated in a play behavior experiment from day 6 to day 53 post-hatch, during which they were tested in enriched play arenas twice weekly in groups of three to four individuals. Housing conditions during rearing included sex-mixed groups of 7–12 individuals in solid-floor cages (0.7 × 0.68 × 0.57 m) with sawdust bedding, heat lamps (removed at four weeks), perches, and ad libitum access to food and water (Oscarsson et al., 2025). All individuals were included based on randomly assigned ID lists, with no selection based on temperament or prior behavior, except from pedigree (HF or LF). Handling and testing protocols ensured minimal self-selection bias.
These lines had been selected over 14 generations for either high or low fear of humans. The selection was determined by the individual fear score obtained using a standardized fear-of-human test conducted at 12 weeks old. For details of the test and the selection program, see (Agnvall et al., 2012). Briefly, each bird was tested individually in a controlled arena (3 m x 90 x 180) enclosed with soft plastic mesh to prevent escape. The arena was built and kept in a lab room with externally controlled lightning. Two experimenters conducted the test: one (Person A) managed the birds and lightning outside the arena, while the other (Person B) conducted the test inside. Person B used a timer with ten second intervals to score fear responses on a five-point ethogram, ranging from calm (1) to freezing or escape attempts (5). The test began in darkness, with Person A handing the bird to Person B, who places it in the center of the arena before signaling readiness with a brief flashlight cue. Once the room light turned on, the three-minute test started, with behavior recorded every 10 seconds. At one and two minutes, Person B stepped closer to the bird to assess its response to increasing proximity. The test concluded after three minutes, and the bird was picked up and taken back to its home area.
All birds were hatched at Linköping University, Sweden. The chicks were vaccinated against Marek’s disease and marked with a wing clip on day one after hatch. After hatch, they were kept in animal cages (0.7 x 0.68 x 0.57 m) until eight weeks of age, when they were transferred to the university’s animal research facility. Here, they were all kept in the same pen (3 x 3 x 3 m) with ad libitum access to food (commercial hen feed) and water. At arrival to the research facility, sexual dimorphism is unmistakable, so sex was written down along with ID by test personal. After 12 weeks of age, they were transferred to their permanent home pen. This was an aviary pen (3 x 3 x 3 m) with three levels, nests, perches and feed (commercial hen feed) and water ad lib, and from 14 weeks of age access to an outdoor area (same size as the indoor pen). The housing facility operated on a 12-hour light/dark cycle (lights on at 07:00h and off at 19:00h), ensuring consistent exposure to naturalistic day and night conditions throughout the study.
Test one: obstacle course
This test was designed to assess motor control, landscape navigation and social motivation. Birds (aged nine weeks) were required to navigate an obstacle course measuring 3.6 m in length, 120 cm in width, and 180 cm in height to regain social contact with its flock mates that were kept in an adjacent pen.
Procedure
The test start point was 4.45 m away from the home pen. Each bird was gently captured, its ID recorded and then placed at the starting point. The obstacle course included 6 elements for the birds to cross (Figure 1).
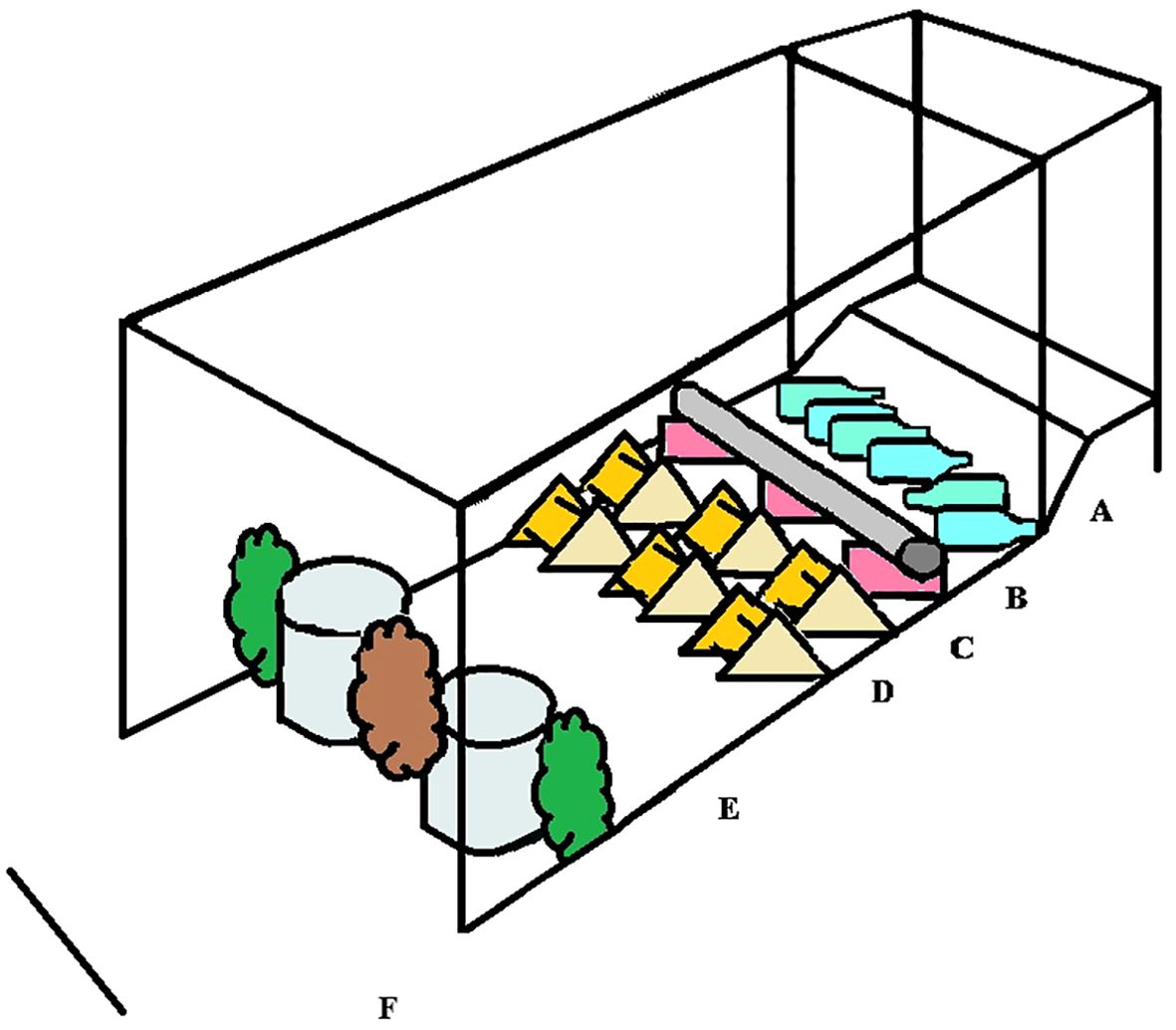
Figure 1. A sketch of the obstacle course used to test locomotory coordination. The birds started on the elevated area to the left of “A”. (A) 60 cm long slope with 25˚incline, (B) pool of 15 empty 1.5 L empty soda bottles, (C) a tube, (D) two rows of triangular perches, (E) smaller open area ending with two buckets and three artificial branches blocking the path, and (F) a final open area close to the goal.
The test ended when both legs of the bird crossed the finish line, located 40 cm from the pen with social companions. The bird could see and hear the cage mates throughout the test, as the starting point was elevated 32 cm above ground level. The test person was hiding behind a covered wall during the test. Each bird was tested three times, on three different days, during a week, but with one day in between each test day.
All tests were video-taped, and the following behavioral variables were recorded for each bird on each test occasion: frequency of Motor error (Bird trips over or collides with objects in the arena), duration of Reorientation pause (The birds stops and turns its front body towards section the bird just crossed to “check” with stretched neck), and latency. All coding was done using Solomon coder (Solomon coder beta 19.18.02).
Test two: rotarod test
The rotarod test (carried out when the birds were 11 weeks old) assessed motor coordination and balance. The apparatus consisted of a rod (50.5 cm wide, circumference 120 cm, diameter 38 cm) elevated 44 cm above the ground. Soft material and blankets cushioned potential falls from the rod. A roof, located 29 cm above the rod, prevented escape as well as flying attempts (Figure 2).

Figure 2. The rotarod apparatus used to assess balance in Red junglefowl during this study.
Procedure
Each bird (aged 11 weeks) was captured in their home pen and carefully carried to the adjacent test room. Its ID was recorded and then the bird was placed at the center of the rod. The rotation began after the bird voluntarily faced the wall, indicating readiness to stay on the rod. The participant in the experiment was responsible for manually regulating the speed of the rotarod from behind a concealed partition. By operating the apparatus through a handle, which was connected to a disk marked with various levels, and utilizing a timer relayed via earphones, the participant was able to adjust the handle to facilitate movements of either one, two, or, in the final moments, three steps per second. Rotation speed was then gradually increased during five minutes, or until the bird fell off the rod:
● 4 cm/s for the first two minutes,
● 6 cm/s for the next two minutes, and
● 12 cm/s for the final minute.
Latency to fall and frequency wing-balancing behavior were recorded from video footage. Tests ended after five minutes if the bird did not fall. Data were recorded using Solomon coder (Solomon coder beta 19.18.02).
Test three: social recognition test
To evaluate aspects of social cognition, birds were subjected to a Social Recognition (SR) test at 13 weeks of age. Birds were first exposed to two different video recordings, one after the other: one featuring an aggressive rooster and the other a rooster in a neutral state. After that, the birds were given a choice to see which of the two individuals they would choose to affiliate with.
Procedure
Birds were first transported from their home pen in two groups of five to a showroom. This contained two enclosures with non-see-through walls (70 * 160 cm), and one side had a video screen at floor level. Here, the chickens were shown two different videos, each five minutes long. The first depicted a rooster being aggressive (during a sparring display towards another invisible rooster on the other side of a mesh), and the other a rooster being calm and neutral (standing relaxed and moving a few steps in a relaxed position). After seeing both videos in groups of five, birds were transported individually to a Y-maze set up in a neighboring lab room, where they entered through a narrow entrance which was closed silently behind them. The maze consisted of a central corridor (70 cm long, 120 cm high) that branched into two lanes (140 x 60 cm). In the distant end of one lane a video showed a loop of the aggressive rooster, but this time being calm and neutral, and the other showed a looped video of the neutral rooster (from a different video than the one they had previously seen). The screens used for displaying the rooster were 52 x 33 cm. The first choice of lane was recorded for each bird. A lane was considered “chosen” when the bird’s entire body, excluding the tail, crossed into it. The test person left the room and closed the door after the birds was placed in the maze. After the test, the bird was carried back to their home pen. Video data were analyzed using Solomon Coder.
Statistics
Generalized linear models (GzLM) were employed in JMP (JMP Pro 18) to analyze effects of line, sex, and their interaction, as well as effect of trial (first, second or third) for data obtained in the obstacle course. Significance was set at P-value <0.05, and P <0.1 was interpreted as a tendency. Data from the rotarod were tested with Kaplan-Meier survival analysis in R-studio (RStudio Team,4.3.0, 2020). Data from the Social Recognition test were analyzed using binomial test in R-studio (RStudio Team,4.3.0, 2020). R studio was also used to analyze whether presentation order influenced the choices in the Y-maze, using Chi-square test of independence.
Results
HF birds had a significantly shorter latency to finish the obstacle course and pass the goal line (Figure 3, χ² = 13.90, P < 0.001). Furthermore, the trial number had a significant effect as well (Figure 3, χ² = 15.31, P < 0.001). The effect of sex was not statistically significant, but there was a tendency suggesting that females had lower values than males (χ² = 3.34, p = 0.0675). The number of motor errors did not differ between the selection lines (Figure 4, χ² = 1.42, P = 0.234). However, there were fewer motor errors with successive trial numbers (Figure 4, χ² = 4.12, P = 0.042) and females performed fewer motor errors than males (Figure 4, χ² = 6.59, P = 0.010).
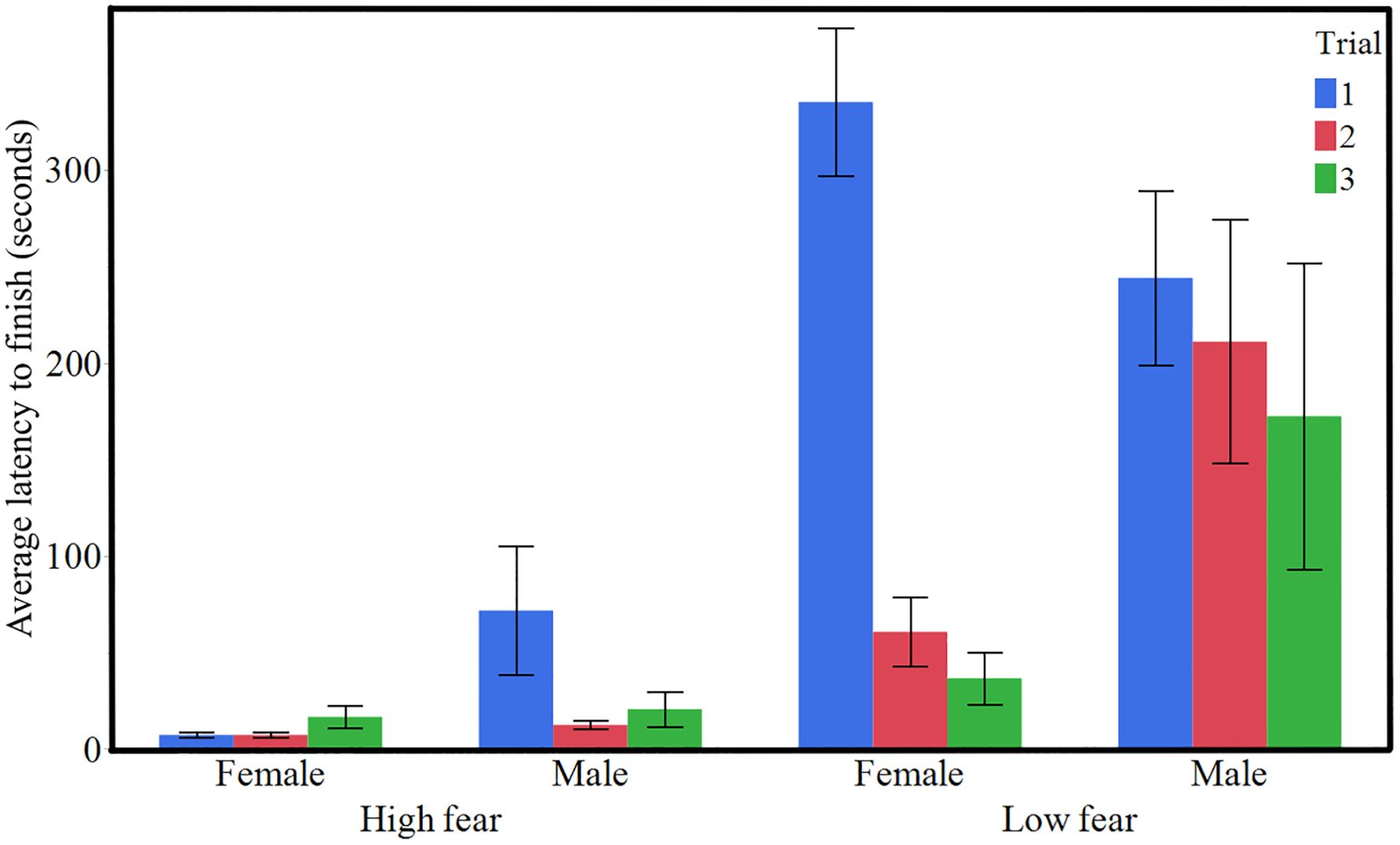
Figure 3. Mean latency (± SE) to complete the obstacle course across three trials for HF (high fear) and LF (low fear) lines of Red Junglefowl (n = 20 HF, n = 18 LF).
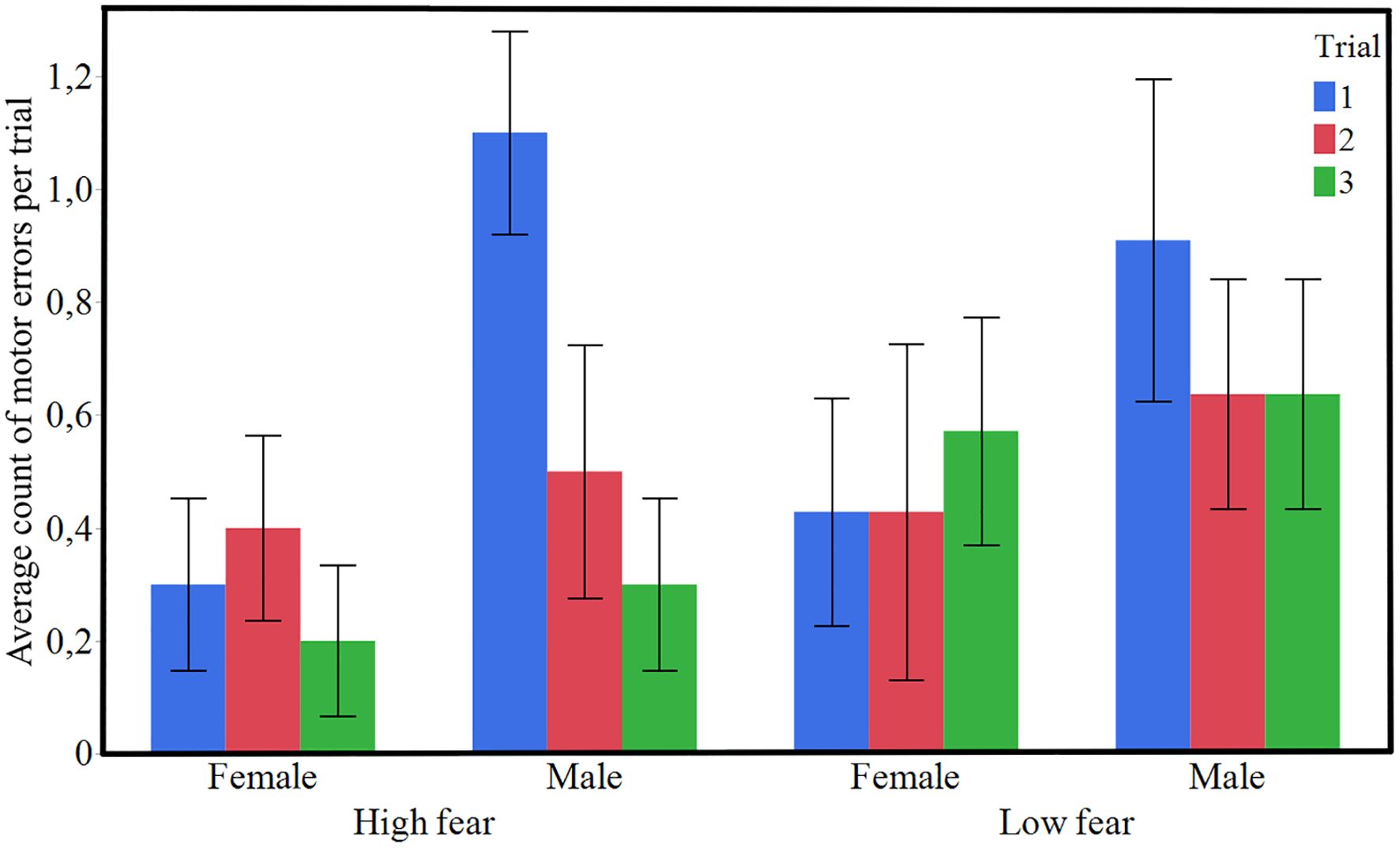
Figure 4. Average numbers ((± SE) of motor errors per trial across the three trials of the obstacle course test for HF (High fear) and LF (Low fear) lines of Red junglefowl (n = 20 HF, n = 18 LF).
The behavior termed “Reorientation pause” was significantly more common in the LF birds (Figure 5, χ² = 9.64, P = 0.0019). It was also significantly affected by sex, being more common in males (Figure 5, χ² = 7.49, P = 0.0062) and was less frequent with successive trials (Figure 5, χ² = 4.97, P = 0.0258). There was a significant interaction effect between line and sex (Figure 5, χ² = 10.86, P = 0.0010).
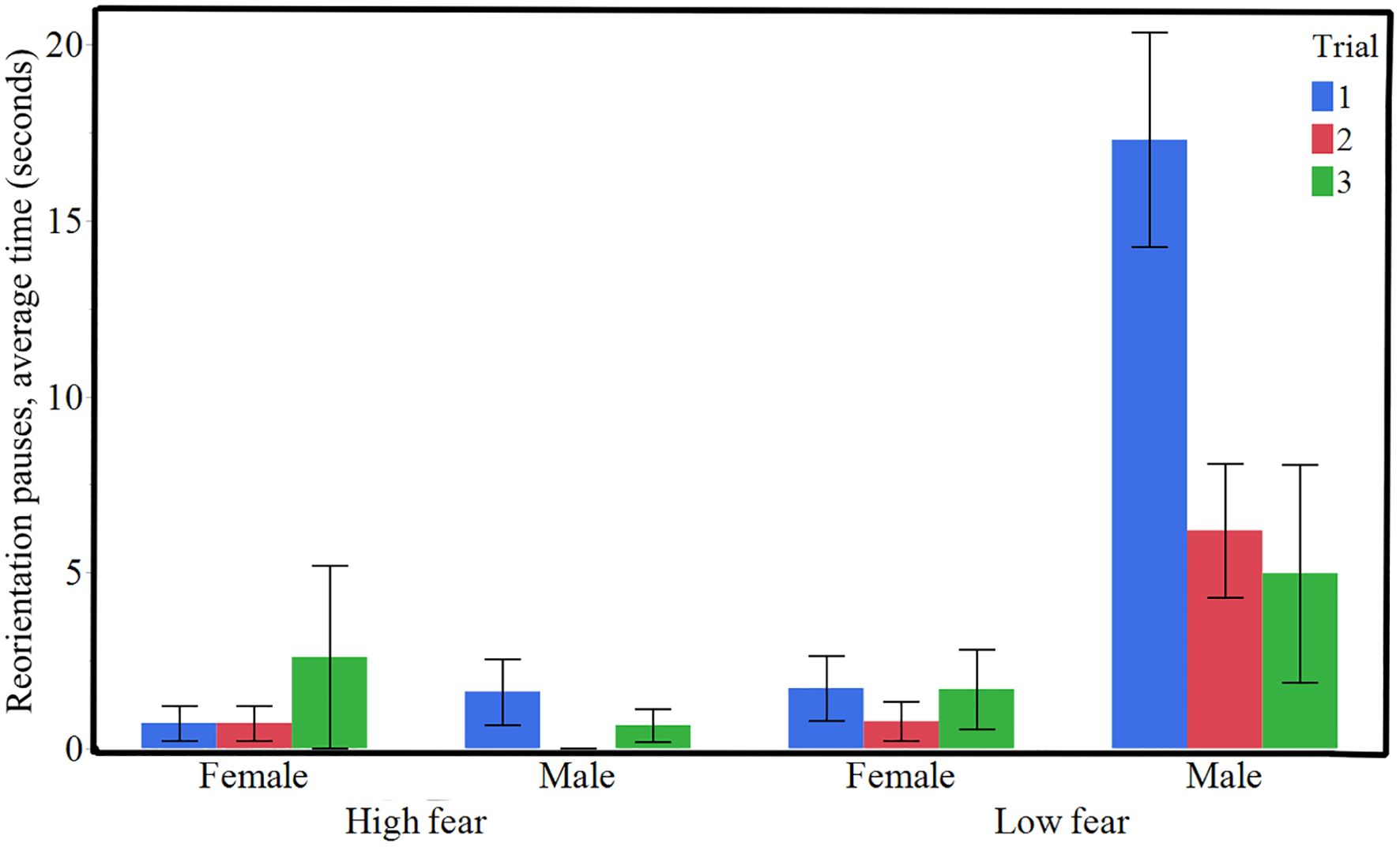
Figure 5. Mean duration (± SE) of reorientation pauses in the obstacle course test in HF (High fear) and LF (Low fear) lines of Red junglefowl (n = 20 HF, n = 18 LF).
There was a weak tendency that LF birds stayed on the rotarod longer than HF (χ² = 2.7, P = 0.1), as shown by the survival analysis in Figure 6. The average latency for HF birds to fall off was 229.33 ± 22.49 seconds, while LF birds had average latency of 267.753 ± 17.88 seconds. There were no effects of sex and no interactions between sex and selection line on this result.
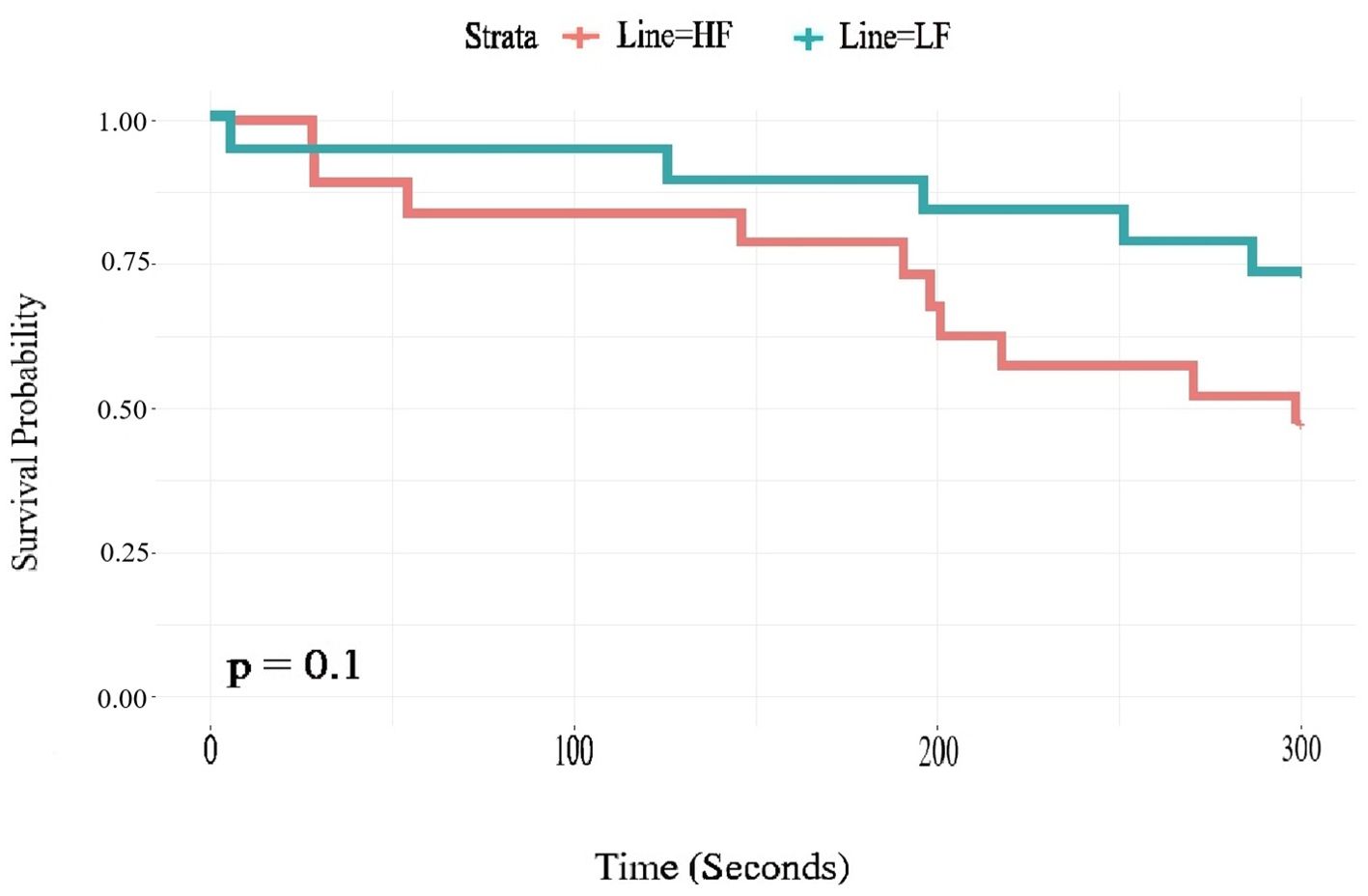
Figure 6. Survival analysis of the time taken to fall off the rod in the rotarod test in HF (High fear) and LF (Low fear) lines of Red junglefowl (n = 20 HF, n = 18 LF). The graph shows the proportion of tested birds that stayed on the rod (Y-axis; Survival probability) for different durations of the test (X-axis).
In the social recognition test, our null-hypothesis was that the birds would choose arms in the Y-maze randomly. Hence, we expected a proportion of choices equal to 0.5 in both selection lines. As seen in Figure 7, HF birds deviated significantly from this proportion as determined by a binomial test (P = 0.032), choosing the arm with the previously friendly rooster more. LF birds did not show any significant deviation from the expected proportion (P = 0.17), although the males chose the previously friendly rooster in twice as many cases as the previously aggressive one. To address the potential confound of presentation order, we conducted a chi-square analysis examining whether video order influenced choice behavior. The analysis revealed no significant effect of presentation order on video choice overall (χ² = 0.69, p = 0.407) or within either selection line (HF: χ² = 1.50, p = 0.221; LF: χ² = 0.00, p = 1.000). While there was a numerical trend toward higher friendly-video selection when it was presented first (72.2%) versus second (52.9%), this difference was not statistically significant and is unlikely to have confounded the main line comparison results.
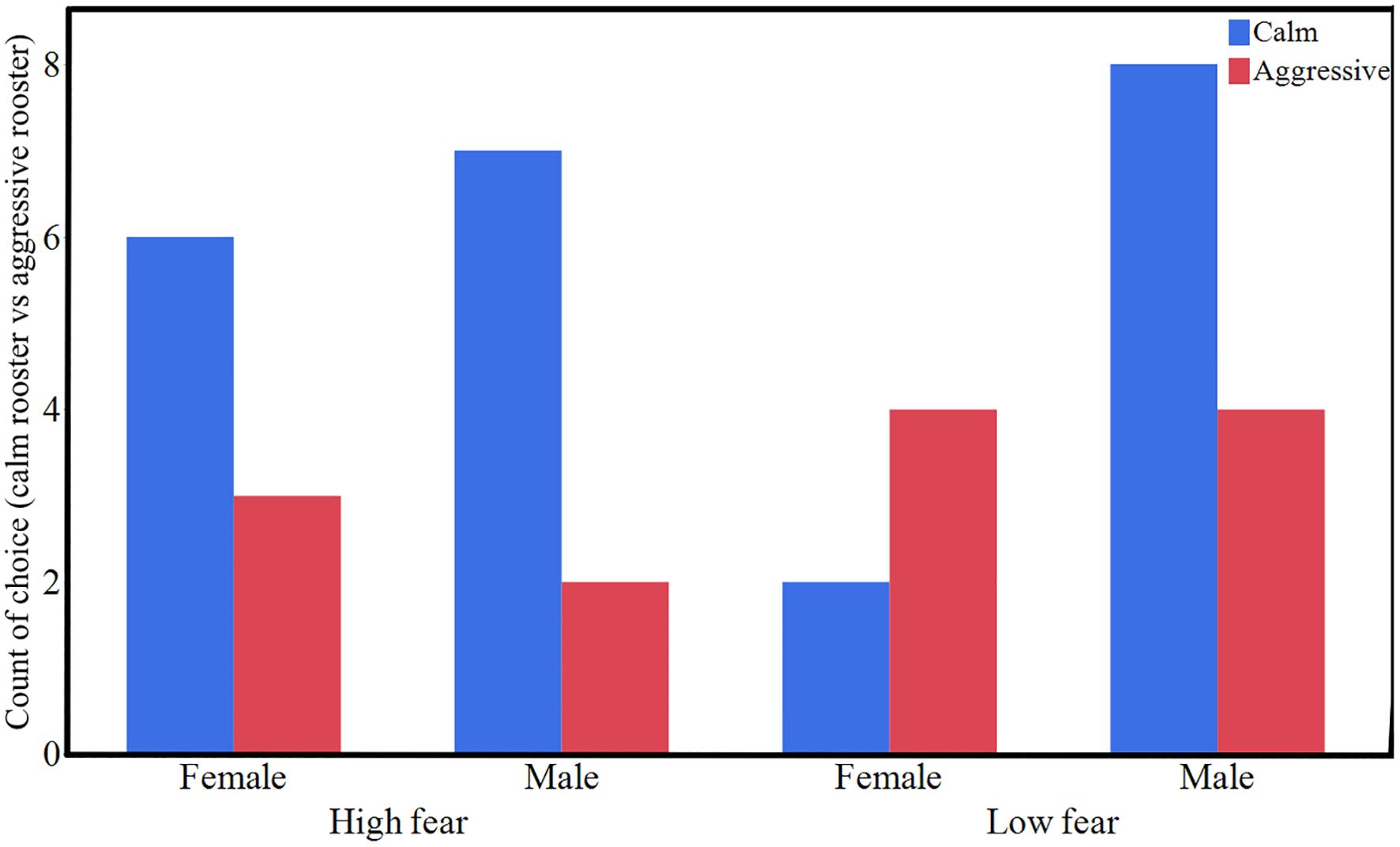
Figure 7. Number of birds from the two selection lines choosing to approach a video of a rooster previously observed to be in a calm non-aggressive state (blue) versus the number of birds choosing the rooster observed to be aggressive (red) (n = 20 HF, n = 18 LF).
Discussion
Previous research on earlier generations of these selection lines, has identified variations in relative cerebellar size and neuronal density between Red Junglefowl selected for low fear (LF) and high fear (HF) of humans (Katajamaa and Jensen, 2020; Katajamaa et al., 2021; Gjøen et al., 2023). This is in line with findings of general chicken domestication and its effects on cerebellar anatomy (Racicot et al., 2021; Stingo-Hirmas et al., 2022). However, the implications of these differences and their role in elucidating behavioral adaptations associated with domestication remain ambiguous. The present study aims to examine how long-term selection for high (HF) or low (LF) fear of humans in Red Junglefowl influenced cerebellum-associated traits, specifically motor coordination, balance, and social recognition, using an obstacle course, a rotarod test, and a social recognition (SR) test. HF birds completed the obstacle course significantly faster than LF birds, though coordination did not differ between the lines. In the rotarod test, balance and endurance were similar, with a weak but non-significant tendency for LF birds to perform better. The SR test revealed no significant preference for a previously neutral versus aggressive roster in LF birds, but HF birds showed a significant preference for the neutral rooster. These findings suggest that bidirectional selection for fear of humans may have affected social behavior in Red Junglefowl but not motor ability to the same extent even though relative size of cerebellum has previously been shown to be larger in the LF birds (Gjøen et al., 2023).
Although previous work has demonstrated selection−related changes in cerebellum size and neuron composition in these same Red Junglefowl lines (Henriksen et al., 2016; Agnvall et al., 2017; Katajamaa and Jensen, 2020; Katajamaa et al., 2021; Gjøen et al., 2023), our study did not include direct anatomical or histological measurements. Consequently, the present findings should be interpreted as functional indicators of cerebellar involvement in motor and social behaviors rather than direct evidence of underlying structural variation. Thus, while our study does not directly measure cerebellar volume or other variables related to brain function within the tested individuals, it uses standardized behavioral tests known to reflect cerebellar involvement to infer the impact of cerebellar size and related adaptations in domestication (Shiotsuki et al., 2010; Mohseni et al., 2020; Chao et al., 2021; Ninomiya et al., 2023). This strategy underscores the importance of behavior as the ultimate readout for brain function, supporting a more complete understanding of how cerebellar changes contribute to the domestication phenotype, beyond size alone.
Tameness is regarded a key driver for domestication, enabling animals to tolerate human handling and management, and may explain why only certain species were successfully domesticated (Price, 2002). Selection experiments worldwide have demonstrated that tameness has genetic components and can be altered through selection. One of the most well-known selection-experiments was initiated in the 1950’s by Dmitry Belyaev, using silver foxes, leading to traits such as piebald pigmentation, shorter tails, floppy ears, and increased social motivation towards humans (Belyaev, 1979). Similar changes have been observed in selection experiments in mink, rats, and rabbits (Trut, 1999; Albert et al., 2009; Trapezov et al., 2012; Singh et al., 2017) (Brusini et al., 2018). In guinea pigs, domesticated individuals show more sociopositive and maternal behaviors, reduced aggression, and lower stress responses compared to wild conspecifics (Kaiser et al., 2015). In Red Junglefowl, selection for tameness has been linked to changes in courtship behavior, increased crowing and food calls in males, higher activity levels, and decreased social proximity (Agnvall and Jensen, 2016; Katajamaa et al., 2018; Gjøen and Jensen, 2021, 2024). Additionally, selecting for tameness in RJF has resulted in domesticated-related traits such as increased size at hatch and adulthood, larger eggs, and enhanced feed efficiency (Agnvall et al., 2018). Importantly, RJF selected for low fear of humans have a smaller brain relative to body size, but larger cerebellum relative to total brain size, similar to domesticated chickens (Henriksen et al., 2016; Gjøen et al., 2023). This indicates that cerebellar controlled traits are linked to tameness and domestication, and the experiments reported here are the first attempt to assess behavioral effects that may possibly be linked to this.
The obstacle course test was performed since cerebellum is known to be involved in motor control as well as social behavior (Adamaszek et al., 2017; Olivito et al., 2023). It thus measures aspects of both these traits. In the obstacle course, HF birds preferred flying over walking, while LF birds paused more frequently to assess their surroundings. These pauses may reflect increased exploratory behavior, perhaps suggesting that LF birds were assessing potential threats, while HF showed higher social motivation. This supports previous research linking tameness to increased exploratory tendencies (Katajamaa et al., 2018; Gjøen and Jensen, 2021; Oscarsson and Jensen, 2024). A notable limitation of this study is the possibility that motor learning acquired during the obstacle course test may have affected subsequent performance on the rotarod apparatus. Research indicates that cerebellar-mediated motor learning can exhibit transfer effects between tasks, especially when both tasks require balance and coordination. It is plausible that the process of motor learning influenced the two selection lines in distinct manners, resulting in one group exhibiting better performance on the rotarod test compared to what might have been expected in the absence of potential training derived from the obstacle course. Given the two-week interval between the assessments, it is conceivable that the transfer effects associated with motor learning were not entirely mitigated, thereby complicating the interpretation of the observed differences in rotarod performance (Celnik, 2015).
The rotarod test is originally a standard test to measure neuromotor performance in rodents used for medical research (Stroobants et al., 2013). It has been used for over 50 years to test neuromotor changes linked to cerebellar dysfunction. Due to the previously demonstrated differences in relative cerebellum size between the selection lines, we hypothesized that motor skills might differ accordingly. Whereas balance and endurance were comparable, the fact that LF birds showed a weak but non-significant tendency for better performance, might suggest that the larger cerebellum in this selection line might be linked to improved motor control. However, it should be noted that standard rotarod tests often fail to detect subtle motor alterations in mice (Stroobants et al., 2013).
The SR test was performed to reveal potential differences in social memory, given the cerebellum’s role in cognitive and social functions, for example that the cerebellum is involved in social reward processing (Manto et al., 2024). In the SR test, HF birds showed a significant preference for the non-aggressive rooster, whereas LF birds exhibited no significant preference. However, LF was the only group that had more visits to the aggressive simulation, although not significant, which may warrant further investigation with a larger sample size. These results indicate a non-random choice of social affiliation that differ between the selection lines. Previous studies found that adult HF and LF females differ in social coping strategies with LF and White Leghorns preferentially affiliated with familiar birds during stress, while HF and unselected RJF females had a weaker such tendency under similar conditions (Gjøen and Jensen, 2024). The HF bird’s preference for the calm rooster suggests that selection for higher fearfulness may have preserved conflict-avoidance behaviors, possibly due to an increased risk sensitivity. Social grouping provides various benefits, such as protection through shared resources and anti-predator behaviors. Even low-ranking individuals gain advantages with these shared resources and protection through anti-predator behaviors, and this may have driven the HF birds to better predict social outcomes and strategically avoid conflict.
Evolutionary changes in cerebellum size is linked to social behavior, also observed in humans, where the cerebellum expanded rapidly compared to apes (Barton and Venditti, 2014). This suggests its crucial role in cognition and social behavior, highlighting its potential significance in domesticated chickens. Previous findings support the mosaic brain evolution hypothesis (Agnvall et al., 2017; Katajamaa and Jensen, 2020; Gjøen et al., 2023), which proposes that domestication can lead to selective growth in brain regions crucial for adapting to human-altered environments (Mehlhorn and Caspers, 2021).Domesticated chickens and LF birds have altered social strategies, including increased social play and different social exploration patterns, which have previously been linked to cerebellar function (Lewis and Barton, 2004; Gabrielle et al., 2022; Oscarsson and Jensen, 2023; Gjøen and Jensen, 2024). Since the cerebellum is involved in social orientation and processing social stimuli (Hosaka et al., 2024), its enlargement may reflect adaptations to changing social structures during domestication. Animal models show that cerebellar circuit disruptions impair social behaviors (Carta et al., 2019). The cerebellum is also linked to emotional regulation, so selection for tameness may additionally have allowed for better regulation of fear and more adaptive responses to human interaction.
While our study focused on behaviors linked to cerebellar function, it is likely that other brain regions involved in fear processing and social cognition also contribute to the observed line differences, particularly in the social recognition test. The amygdala, for instance, plays a key role in mediating fear responses, processing social cues, and forming emotionally salient memories (Ledoux, 2013; Phelps and Ledoux, 2005; Pessoa, 2010). Domestication and selection for tameness have been associated with reduced amygdala size and fear reactivity in several species, potentially including chickens (Brusini et al., 2018; Katajamaa and Jensen, 2020). The stronger preference of HF birds for the calm rooster, compared to LF birds, may therefore reflect differences in amygdala-mediated threat perception. Moreover, increasing evidence indicates functional interplay between the cerebellum and amygdala in shaping emotional and social behavior (Jung et al., 2022). Thus, while our findings highlight cerebellar involvement in motor and social tasks, the line differences likely arise from coordinated changes across multiple brain regions influencing tameness, fear, and social cognition during domestication.
A limitation of this study is the order of testing. Although birds were already classified as high fear HF or LF based on pedigree, the individual “fear-of-human” test, which is used to determine tameness scores for later breeding, was conducted after two behavioral assays. Prior experiences from these tests (e.g., habituation or learning) may have influenced fear responses, potentially altering tameness scores. Moreover, such effects may differ between HF and LF lines, introducing variability that could influence later performance in the social navigation test and complicate interpretation of group difference.
In conclusion, our findings suggest that the selection for tameness, where studies on previous generations show increased relative cerebellar size, influences the ability to transverse an obstacle course in order to regain social contact, and social navigation as measured by choice of social partner. Furthermore, a possible but non-significant link to motor control, as measured by the ability to stay on a rotarod, was observed. HF birds showed significant preference for the non-aggressive rooster, whereas LF birds did not exhibit the same selective behavior. This suggests that the tameness-induced changes in cerebellum may affect both social and motor related traits.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Linköping Animal Ethics Committee under license no. 14916-2018. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
JG: Methodology, Data curation, Investigation, Writing – review & editing, Visualization, Writing – original draft, Conceptualization, Formal Analysis. PJ: Methodology, Supervision, Conceptualization, Writing – review & editing, Writing – original draft, Funding acquisition, Project administration, Resources.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Swedish Research Council (grant no 2019-04869).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adamaszek M., D’agata F., Ferrucci R., Habas C., Keulen S., Kirkby K., et al. (2017). Consensus paper: cerebellum and emotion. Cerebellum 16, 552–576. doi: 10.1007/s12311-016-0815-8
Agnvall B., Ali A., Olby S., and Jensen P. (2014). Red Junglefowl (Gallus gallus) selected for low fear of humans are larger, more dominant and produce larger offspring. Animal 8, 1498–1505. doi: 10.1017/S1751731114001426
Agnvall B., Bélteky J., and Jensen P. (2017). Brain size is reduced by selection for tameness in Red Junglefowl– correlated effects in vital organs. Sci. Rep. 7, 3306. doi: 10.1038/s41598-017-03236-4
Agnvall B., Bélteky J., Katajamaa R., and Jensen P. (2018). Is evolution of domestication driven by tameness? A selective review with focus on chickens. Appl. Anim. Behav. Sci. 205, 227–233. doi: 10.1016/j.applanim.2017.09.006
Agnvall B. and Jensen P. (2016). Effects of divergent selection for fear of humans on behavior in Red Junglefowl. PloS One 11, e0166075. doi: 10.1371/journal.pone.0166075
Agnvall B., Jöngren M., Strandberg E., and Jensen P. (2012). Heritability and genetic correlations of fear-related behavior in red junglefowl–possible implications for early domestication. PloS One 7, e35162. doi: 10.1371/journal.pone.0035162
Agnvall B., Katajamaa R., Altimiras J., and Jensen P. (2015). Is domestication driven by reduced fear of humans? Boldness, metabolism and serotonin levels in divergently selected red junglefowl (Gallus gallus). Biol. Lett. 11, 20150509. doi: 10.1098/rsbl.2015.0509
Albert F. W., Carlborg O. R., Plyusnina I., Besnier F., Hedwig D., Lautenschläger S., et al. (2009). Genetic architecture of tameness in a rat model of animal domestication. Genetics 182, 541–554. doi: 10.1534/genetics.109.102186
Barton R. A. and Venditti C. (2014). Rapid evolution of the cerebellum in humans and other great apes. Curr. Biol. 24, 2440–2444. doi: 10.1016/j.cub.2014.08.056
Belyaev D. K. (1979). Destabilizing selection as a factor in domestication. J. Heredity 70, 301–308. doi: 10.1093/oxfordjournals.jhered.a109263
Brusini I., Carneiro M., Wang C., Rubin C.-J., Ring H., Afonso S., et al. (2018). Changes in brain architecture are consistent with altered fear processing in domestic rabbits. Proc. Natl. Acad. Sci. 115, 7380–7385. doi: 10.1073/pnas.1801024115
Carta I., Chen C. H., Schott A. L., Dorizan S., and Khodakhah K. (2019). Cerebellar modulation of the reward circuitry and social behavior. Science 363, eaav0581. doi: 10.1126/science.aav0581
Celnik P. (2015). Understanding and modulating motor learning with cerebellar stimulation. Cerebellum 14, 171–174. doi: 10.1007/s12311-014-0607-y
Chao O. Y., Zhang H., Pathak S. S., Huston J. P., and Yang Y. M. (2021). Functional convergence of motor and social processes in lobule IV/V of the mouse cerebellum. Cerebellum 20, 836–852. doi: 10.1007/s12311-021-01246-7
Collarini E., Gioia M., Cordoni G., and Norscia I. (2022). Does the domestication syndrome apply to the domestic pig? Not completely. Animals 12, 2458. doi: 10.3390/ani12182458
Dahhaoui M., Caston J., Auvray N., and Reber A. (1990). Role of the cerebellum in an avoidance conditioning task in the rat. Physiol. Behav. 47, 1175–1180. doi: 10.1016/0031-9384(90)90369-F
Dou M., Li M., Zheng Z., Chen Q., Wu Y., Wang J., et al. (2023). A missense mutation in RRM1 contributes to animal tameness. Sci. Adv. 9. doi: 10.1126/sciadv.adf4068
Fitak R. R., Mohandesan E., Corander J., Yadamsuren A., Chuluunbat B., Abdelhadi O., et al. (2020). Genomic signatures of domestication in Old World camels. Commun. Biol. 3. doi: 10.1038/s42003-020-1039-5
Frontera J., Aissa H., Sala R., Mailhes-Hamon C., Georgescu I., Léna C., et al. (2020). Bidirectional control of fear memories by cerebellar neurons projecting to the ventrolateral periaqueductal grey. Nat. Commun. 11, 5207. doi: 10.1038/s41467-020-18953-0
Gabrielle L., Rebecca O., Louise H., Johanna G., and Per J. (2022). Play ontogeny in young chickens is affected by domestication and early stress. Sci. Rep. 12, 13576. doi: 10.1038/s41598-022-17617-x
Galef B. G. Jr (1970). Aggression and timidity: responses to novelty in feral Norway rats. J. Comp. Physiol. Psychol. 70, 370. doi: 10.1037/h0028719
Gjøen J., Cunha F., and Jensen P. (2023). Selection for reduced fear of humans changes brain and cerebellum size in red junglefowl in line with effects of chicken domestication. Brain Sci. 13, 988. doi: 10.3390/brainsci13070988
Gjøen J. and Jensen P. (2021). Selection for reduced fear of humans changes intra-specific social behavior in red junglefowl—Implications for chicken domestication. Genes 13, 43.
Gjøen J. and Jensen P. (2024). Social preferences in chickens–effects of domestication and tameness. Front. Anim. Sci. 5, 1487688. doi: 10.3389/fanim.2024.1487688
Heleven E., Van Dun K., and Van Overwalle F. (2019). The posterior Cerebellum is involved in constructing Social Action Sequences: An fMRI Study. Sci. Rep. 9, 11110–11110. doi: 10.1038/s41598-019-46962-7
Henriksen R., Johnsson M., Andersson L., Jensen P., and Wright D. (2016). The domesticated brain: genetics of brain mass and brain structure in an avian species. Sci. Rep. 6, 1–9. doi: 10.1038/srep34031
Hosaka S., Hosokawa M., Hibi M., and Shimizu T. (2024). The zebrafish cerebellar neural circuits are involved in orienting behavior. eNeuro 11. doi: 10.1523/ENEURO.0141-24.2024
Iwaniuk A. N., Hurd P. L., and Wylie D. R. (2006). Comparative morphology of the avian cerebellum: I. Degree of foliation. Brain Behav. Evol. 68, 45–62. doi: 10.1159/000093530
Jung S. J., Vlasov K., D’ambra A. F., Parigi A., Baya M., Frez E. P., et al. (2022). Novel cerebello-amygdala connections provide missing link between cerebellum and limbic system. Front. Syst. Neurosci. 16. doi: 10.3389/fnsys.2022.879634
Kaiser S., Hennessy M. B., and Sachser N. (2015). Domestication affects the structure, development and stability of biobehavioral profiles. Front. Zool 12, S19. doi: 10.1186/1742-9994-12-S1-S19
Katajamaa R. and Jensen P. (2020). Selection for reduced fear in red junglefowl changes brain composition and affects fear memory. R. Soc. Open Sci. 7, 200628. doi: 10.1098/rsos.200628
Katajamaa R., Larsson L. H., Lundberg P., Sörensen I., and Jensen P. (2018). Activity, social and sexual behavior in Red Junglefowl selected for divergent levels of fear of humans. PloS One 13, e0204303. doi: 10.1371/journal.pone.0204303
Katajamaa R., Wright D., Henriksen R., and Jensen P. (2021). Cerebellum size is related to fear memory and domestication of chickens. Biol. Lett. 17, 20200790. doi: 10.1098/rsbl.2020.0790
Kruska D. (2007). The effects of domestication on brain size. In: KAAS J. H. (ed.) Evolution of Nervous Systems. 3, 143–153. Oxford: Academic Press. doi: 10.1016/B0-12-370878-8/00054-9
Leggio M. and Molinari M. (2015). Cerebellar sequencing: a trick for predicting the future. Cerebellum 14, 35–38. doi: 10.1007/s12311-014-0616-x
Lewis K. P. and Barton R. A. (2004). Playing for keeps: evolutionary relationships between social play and the cerebellum in nonhuman primates. Hum. Nat. 15, 5–21. doi: 10.1007/s12110-004-1001-0
Lindberg J., Björnerfeldt S., Bakken M., Vilà C., Jazin E., and Saetre P. (2007). Selection for tameness modulates the expression of heme related genes in silver foxes. Behav. Brain Functions 3. doi: 10.1186/1744-9081-3-18
Malmkvist J. and Hansen S. W. (2002). Generalization of fear in farm mink, Mustela vison, genetically selected for behavior towards humans. Anim. Behav. 64, 487–501. doi: 10.1006/anbe.2002.3058
Manto M., Adamaszek M., Apps R., Carlson E., Guarque-Chabrera J., Heleven E., et al. (2024). Consensus paper: cerebellum and reward. Cerebellum 23, 2169–2192. doi: 10.1007/s12311-024-01702-0
Mehlhorn J. and Caspers S. (2021). The effects of domestication on the brain and behavior of the chicken in the light of evolution. Brain Behav. Evolution. 95 (6), 287–301. doi: 10.1159/000516787
Mohseni F., Behnam S. G., and Rafaiee R. (2020). Review paper: A review of the historical evolutionary process of dry and water maze tests in rodents. Basic Clin. Neurosci. 11, 389–401. doi: 10.32598/bcn.11.4.1425.1
Molinari M., Restuccia D., and Leggio M. G. (2009). State estimation, response prediction, and cerebellar sensory processing for behavioral control. Cerebellum 8, 399–402. doi: 10.1007/s12311-009-0112-x
Nagayama H., Matsumoto Y., Tanave A., Nihei M., Goto T., and Koide T. (2018). Measuring active and passive tameness separately in mice. J. Vis. Exp. 138. doi: 10.3791/58048
Naumenko E. V., Popova N. K., Nikulina E. M., Dygalo N. N., Shishkina G. T., Borodin P. M., et al. (1989). Behavior, adrenocortical activity, and brain monoamines in Norway rats selected for reduced aggressiveness towards man. Pharmacol. Biochem. Behav. 33, 85–91. doi: 10.1016/0091-3057(89)90434-6
Ninomiya A., Amano I., and Koibuchi N. (2023). A versatile, behavioral method to investigate thyroid hormone effects on cerebellar function. J. Visualized Experiments 2023, e65940. doi: 10.3791/65940
Olivito G., Siciliano L., Clausi S., Lupo M., Baiocco R., Gragnani A., et al. (2023). The cerebellum gets social: evidence from an exploratory study of cerebellar, neurodevelopmental, and psychiatric disorders. Biomedicines 11, 309. doi: 10.3390/biomedicines11020309
Oscarsson R., Gjøen J., and Jensen P. (2025). Selection for tameness alters play-like behavior in red junglefowl in line with effects of domestication. Biol. Lett. 21, 20240607. doi: 10.1098/rsbl.2024.0607
Oscarsson R. and Jensen P. (2023). Potential domestication and tameness effects on prosocial behavior in chickens. PloS One 18, e0287213. doi: 10.1371/journal.pone.0287213
Oscarsson R. and Jensen P. (2024). Male chicks play more than females–sex differences in chicken play ontogeny. Front. Ethology 3, 1392378. doi: 10.3389/fetho.2024.1392378
Parkins E. (1997). Cerebellum and cerebrum in adaptive control and cognition: a review. Biol. cybernetics 77, 79–87. doi: 10.1007/s004220050369
Pessoa L. (2010). Emotion and cognition and the amygdala: from “what is it?“ to “what's to be done”. Neuropsychol. 48, 3416–3429. doi: 10.1016/j.neuropsychologia.2010.06.038
Phelps E. A. and Ledoux J. E. (2005). Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron 48, 175–187. doi: 10.1016/j.neuron.2005.09.025
Price E. O. (2002). Pre-adaptations for domestication animal domestication and behavior (allingford Oxon, UK, CABI Publishing).
Racicot K. J., Popic C., Cunha F., Wright D., Henriksen R., and Iwaniuk A. N. (2021). The cerebellar anatomy of red junglefowl and white leghorn chickens: insights into the effects of domestication on the cerebellum. R. Soc. Open Sci. 8, 211002. doi: 10.1098/rsos.211002
Rehkamper G., Frahm H. D., and Cnotka J. (2008). Mosaic evolution and adaptive brain component alteration under domestication seen on the background of evolutionary theory. Brain Behav. Evol. 71, 115–126. doi: 10.1159/000111458
Sacchetti B., Baldi E., Lorenzini C. A., and Bucherelli C. (2002). Cerebellar role in fear-conditioning consolidation. Proc. Natl. Acad. Sci. 99, 8406–8411. doi: 10.1073/pnas.112660399
Shiotsuki H., Yoshimi K., Shimo Y., Funayama M., Takamatsu Y., Ikeda K., et al. (2010). A Rotarod Test For evaluation of motor skill learning. J. Neurosci. Methods 189, 180–185. doi: 10.1016/j.jneumeth.2010.03.026
Singh N., Albert F. W., Plyusnina I., Trut L., Pääbo S., and Harvati K. (2017). Facial shape differences between rats selected for tame and aggressive behaviors. PloS One 12 (4), e0175043. doi: 10.1371/journal.pone.0175043
Stingo-Hirmas D., Cunha F., Cardoso R. F., Carra L. G., Rönnegård L., Wright D., et al. (2022). Proportional cerebellum size predicts fear habituation in chickens. Front. Physiol. 124. doi: 10.3389/fphys.2022.826178
Stroobants S., Gantois I., Pooters T., and D’hooge R. (2013). Increased gait variability in mice with small cerebellar cortex lesions and normal rotarod performance. Behav. Brain Res. 241, 32–37. doi: 10.1016/j.bbr.2012.11.034
Sultan F. (2005). Why some bird brains are larger than others. Curr. Biol. 15, R649–R650. doi: 10.1016/j.cub.2005.08.043
Sultan F. and Glickstein M. (2008). The cerebellum: Comparative and animal studies. Cerebellum 6, 168–176. doi: 10.1080/14734220701332486
Trapezov O. V., Trapezova L. I., and Sergeev E. G. (2012). Coat color mutations and defensive reaction towards man in farm-bred minks and sables (Wageningen: Wageningen Academic Publishers), 396–403.
Trut L. (1998). The evolutionary concept of destabilizing selection: status quo In commemoration of DK Belyaev. J. Anim. Breed. Genet. 115, 415–431. doi: 10.1111/j.1439-0388.1998.tb00364.x
Trut L. N. (1999). Early Canid Domestication: The Farm-Fox Experiment: Foxes bred for tamability in a 40-year experiment exhibit remarkable transformations that suggest an interplay between behavioral genetics and development. Am. Scientist 87, 160–169. doi: 10.1511/1999.20.160
Van Der Heijden M. E. (2024). Converging and diverging cerebellar pathways for motor and social behaviors in mice. Cerebellum 23, 1754–1767. doi: 10.1007/s12311-024-01706-w
Van Overwalle F., Van De Steen F., Van Dun K., and Heleven E. (2020). Connectivity between the cerebrum and cerebellum during social and non-social sequencing using dynamic causal modelling. NeuroImage 206, 116326. doi: 10.1016/j.neuroimage.2019.116326
Keywords: tameness, Red Junglefowl (RJF), cerebellum, motor behavior, social behavior, animal cognition, selection experiment, animal domestication
Citation: Gjøen J and Jensen P (2025) Cerebellar related social and motor behavior in Red Junglefowl selected for high or low fear of humans. Front. Ethol. 4:1699920. doi: 10.3389/fetho.2025.1699920
Received: 05 September 2025; Accepted: 29 October 2025;
Published: 17 November 2025.
Edited by:
Matteo Chincarini, University of Teramo, ItalyReviewed by:
Scott Nunes, University of San Francisco, United StatesUsama T. Mahmoud, Assiut University, Egypt
Copyright © 2025 Gjøen and Jensen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Per Jensen, cGVyLmplbnNlbkBsaXUuc2U=