 Theodore Spanos1
Theodore Spanos1 Amira Ghodbane1
Amira Ghodbane1 Aya Rezazga1
Aya Rezazga1 Márta Ludman2
Márta Ludman2 Károly Fátyol2
Károly Fátyol2 Toufic Elbeaino3
Toufic Elbeaino3 Ioannis Livieratos1*
Ioannis Livieratos1*- 1Department of Sustainable Agriculture, International Center for Advanced Mediterranean Agronomic Studies (CIHEAM), Mediterranean Agronomic Institute of Chania (MAICh), Chania, Greece
- 2Institute of Genetics and Biotechnology, Hungarian University of Agriculture and Life Sciences, Gödöllő, Hungary
- 3Department of Integrated Pest Management of Fruit Trees and Vegetable Crops, International Center for Advanced Mediterranean Agronomic Studies (CIHEAM), Mediterranean Agronomic Institute of Bari (IAMB), Valenzano, Italy
The phenomenon whereby plants infected with a mild isolate of a virus are cross-protected from subsequent “challenge” inoculation with a severe isolate of the same virus resembles mammalian vaccination and was first described almost 100 years ago. In this study, the ability of wild type (wt), ago2- and DCL2/4-deficient Nicotiana benthamiana plants “vaccinated” with the mild (Sp13) PepMV isolate to exert a cross-protective response against “challenge”-inoculation with a severe (PCH, aggressive) isolate was investigated. Initially in wt and ago2-deficient plants, two interval-times (5 and 10 days) between inoculations were used and virus accumulation was analyzed in newly-formed (systemic) leaves. In follow up experiments, wt and DCL2/4-deficient plants were inoculated using a single interval-time (5 days) and both inoculated (local) and systemic leaves were analyzed. In both experiments, leaves were collected 3 days post “challenge”-inoculation and immunoblot, northern blot and RT-qPCR assays demonstrated the complete blockage of the PCH aggressive isolate, which remained at undetectable levels. Using a similar experimental set-up with wt and ago2-deficient N. benthamiana plants and the genetically distinct Potato virus X as the “challenge”-inoculum, complete blockage of superinfection was not observed but instead a significant reduction of PVX RNA levels systemically. Collectively, our results cannot entirely exclude the complete redundancy of two essential components of RNA silencing, but clearly suggest the activation of an efficient and durable superinfection protection mechanism, which is distinct to it.
1 Introduction
Cross-protection (1) is a phenomenon in which infection of plants with a mild isolate of a virus protects them from subsequent infection with a severe isolate of the same virus. Cross-protection can be considered equivalent to mammalian vaccination, and the terms “pre-immunization” and “vaccination” have been used in some studies wherein mild, attenuated, or non-symptomatic plant virus isolates or mutants have acted as “vaccines” (2, 3). Cross-protection works best when the “vaccine” is genetically very close to the “challenge” isolate, but its exact mechanism remains elusive (2, 4, 5). In the late 1990s, cross-protection was attributed to RNA silencing; specifically, infection with a mild isolate activates the plant surveillance system at the molecular level, and the highly homologous “challenge” isolate is specifically recognized and degraded (6, 7). Typically, double-stranded RNAs (dsRNAs) of the primary virus (i.e., structural regions of the genome and replication intermediates) are recognized and cleaved by an RNase III-type enzyme (DCL) to produce small interfering RNAs (siRNAs) 21–24 nucleotides (nt) in length. siRNAs are then incorporated into Argonaute (AGO) protein-containing RNA-induced silencing complexes that guide the recognition and degradation of the severe genetically homologous “challenge” virus (reviewed in Hull, 2002). As siRNAs also represent a mobile systemic signal for the generation of systemic defense responses (8), this model fits the systemic character of cross-protection.
In some cases, cross-protection has worked in plant mutants with two or three DCL genes knocked out, implying a mechanism distinct from RNA silencing (9, 10). Superinfection exclusion (SIE) has been recently proposed for plants (11, 12). This model postulates that certain activities of the mild virus engage essential capacity in the host (i.e., replication complex formation sites, inhibition of uncoating) to which the “challenge” virus isolate has been restricted or has no access. In the cases of Citrus tristeza virus [p33, CTV (13)], Turnip crinkle virus [p28, TCV (14)], and Turnip mosaic virus [p3 and NIa-Pro; TuMV (15)], specific viral proteins represent the elicitors and targets of SIE, resulting in restricted replication of the “challenge” isolate.
The first successful application of cross-protection targeted severe CTV strains that caused significant economic damage (16). Other successful examples involved Zucchini mosaic virus (17), Tomato mosaic virus (18), and Papaya ringspot virus (19). Additionally, several mild isolates of the mechanically-transmitted cosmopolitan Pepino mosaic virus (PepMV) were identified and used successfully for cross-protection in many countries since the early 2000s (20, 21). However, no tomato varieties resistant to PepMV are commercially available; therefore, phytosanitary controls and early destruction of infected plants are the only alternative control strategies in the field (21). An attenuated cross-protective PepMV variant was generated by alignment-guided mutagenesis of the viral coat protein (CP) (22).
PepMV has a 6.4 kb single-stranded (ss) positive-sense RNA genome with a 5′-methylguanosine cap and a 3′-polyA tail (23). The PepMV genome contains five open reading frames, flanked by 5′- and 3′- untranslated regions (UTRs) of approximately 86 and 64 nt, respectively. The RNA-dependent RNA polymerase (RdRp) is encoded by the genomic RNA, whereas three proteins (triple gene block proteins 1–3; TGBp1–3) and the CP are encoded by 3′-co-terminal sgRNAs. TGBp1 is the main silencing suppressor of the virus, and together with TGBp2 and 3, is required for virus movement within the plant. CP, apart from coating viral RNA, is also a mild silencing suppressor (24) and is required for cell-to-cell movement of the virus. An RNA pseudoknot in the PepMV 3′-UTR includes part of the polyA tail and is required for virus replication (25). From the host perspective, the hierarchical roles of several AGOs in anti-PepMV defense have been reported, with the predominant contribution of AGO2 (26, 27). Several tomato proteins interact with PepMV-encoded proteins (28, 29), and the calcium-permeable channel 4.1 and glutathione S-transferase SlGSTU38 have recently been fully characterized as susceptibility factors for PepMV infections (30, 31).
This study investigated the cross-protection of ago2- and DCL2/4-deficient Nicotiana benthamiana plants against superinfection of an aggressive PepMV isolate following “vaccination” with a mild isolate. Protein and RNA analyses of leaves collected at 3 days post inoculation (dpi) showed that the aggressive isolate was completely blocked locally and systemically, suggesting that a mechanism other than RNA silencing is responsible for the observed phenomenon.
2 Materials and methods
2.1 Virus inoculum, plant material, polyclonal antiserum production, and experimental set up
Tomato leaves infected with the Spanish PepMV-Sp13 isolate [EU mild pathotype (23)] were provided by Dr. M. Aranda (CISC, Murcia, Spain). Tomato leaves infected with PepMV-PCH 06/104 [CH2 aggressive pathotype (32)] were provided by Dr. I. Hanssen (Scientia Terrae Research Institute, Belgium). Tomato leaves infected with Potato virus X (PVX) and Bamboo mosaic virus (BaMV) were provided by Dr. R. Olsthoorn (Leiden Institute of Chemistry, The Netherlands).
Plant DCL and AGO2 are critical components of the plant RNA silencing pathway. CRISPR/Cas9-generated ago2 mutant (33) and DCL2/4-suppressed (34) N. benthamiana seeds were donated by Dr. K. Fatyol (Agricultural Biotechnology Institute, Gödöllő, Hungary) and Dr. K. Kalantidis (University of Crete), respectively.
In all experiments, the third real leaf of three plants of each N. benthamiana genotype was mechanically inoculated using 1 µg of a purified potexvirus virion preparation in phosphate buffered saline (PBS). PepMV-Sp13 (mild), -PCH 06/104 (aggressive), BaMV, and PVX virions were purified from approximately 50 g of systemically infected N. benthamiana leaves using the protocol of Abou Haidar and co-workers (35). To prepare circa 500mg specific anti-mild PepMV IgG, approximately 1.5 mg of PepMV-Sp13 virions were purified and intradermally injected into a rabbit (days 1, 7, 14; Davids Biotechnologie GmbH, Germany). The crude serum was collected in a final bleed (day 28) and PepMV-Sp13-CP IgGs was purified by passage through a protein A column and eluted in 0.1 M sodium acetate; 0.02% sodium azide, and stored at -20°C.
For single virus infections, three plants from each of the N. benthamiana genotypes (wild-type [wt], DCL2/4-deficient, NahG, ago2) were mechanically inoculated with each of the four potexviruses (PepMV-Sp13, -PCH 06/104, PVX, and BaMV). In single and double infections in the cross-protection experiments, three N. benthamiana plants from each genotype (wt, ago2-mutant, and DCL2/4-deficient) were “vaccinated” with PepMV-Sp13 or PBS (mock) 5 or 10 days prior to subsequent challenge inoculation. Systemic and local leaves were collected 3 days after “challenge” inoculation for protein and RNA analyses.
2.2 RNA extraction and northern blot analysis
Half of each leaf sample was ground in liquid nitrogen. Total RNA was extracted using TRIzol reagent (Invitrogen) in accordance with the manufacturer’s instructions, and the concentration and RNA quality were assessed using a NanoPhotometer Pearl (Implen). RNA samples (1 μg) were separated by electrophoresis on 1% formaldehyde agarose gels and blotted onto Hybond N membranes (GE Healthcare). Following a previously described method (24), northern blots were hybridized with a digoxin (DIG)-labeled riboprobe corresponding to the negative sense of each viral (PepMV-Sp13, PepMV-PCH 06/104, PVX, and BaMV) CP gene. Chemiluminescent detection was performed using the DIG Detection Kit (Roche).
2.3 Protein extraction and immunoblot analysis
Total proteins were extracted in PBS from collected leaves in microfuge tubes using liquid nitrogen. Following a short centrifugation to precipitate the debris, the supernatants were mixed (1:1 v/v) with protein loading buffer and subjected to SDS-PAGE. For immunoblot analysis, proteins were transferred onto a AmershamTM HybondTM P 0.45 PVDF blotting membrane (GE Healthcare) and hybridized overnight against the PepMV-Sp13 (generated in this study) or PVX-CP IgGs (Loewe; Germany; Cat. No. 07037). Following incubation with alkaline phosphatase-conjugated goat anti-rabbit IgGs diluted (1:7500) in PBS, virus CPs were visualized using NBT-BCIP substrate (Promega).
2.4 Real-time qPCR
Approximately 500 ng of total RNA was treated with DNase I (Thermo Scientific) to remove any genomic DNA contamination before complementary DNA (cDNA) synthesis was performed with an oligo (dT18) primer using PrimeScript reverse transcriptase (Takara) in accordance with the manufacturer’s instructions.
qPCR was performed using a SaCycler-96 Real-Time PCR system (Sacace Biotechnologies, Italy). The 10 μL PCR mix consisted of 1× KAPA SYBR Fast qPCR Master Mix (Kapa Biosystems), 200 nM gene-specific primers, and 5 ng cDNA template. An initial denaturation step at 95°C for 3 min was followed by 40 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 20 s, and extension at 72°C for 20 s. A melting curve analysis protocol was executed in the temperature range of 60 to 95°C. Oligonucleotide primers for qPCR (Table 1) were designed using PRIMER3 software (Whitehead Institute for Biomedical Research, https://www-genome.wi.mit.edu/cgi-bin/primer/primer3.cgi/).
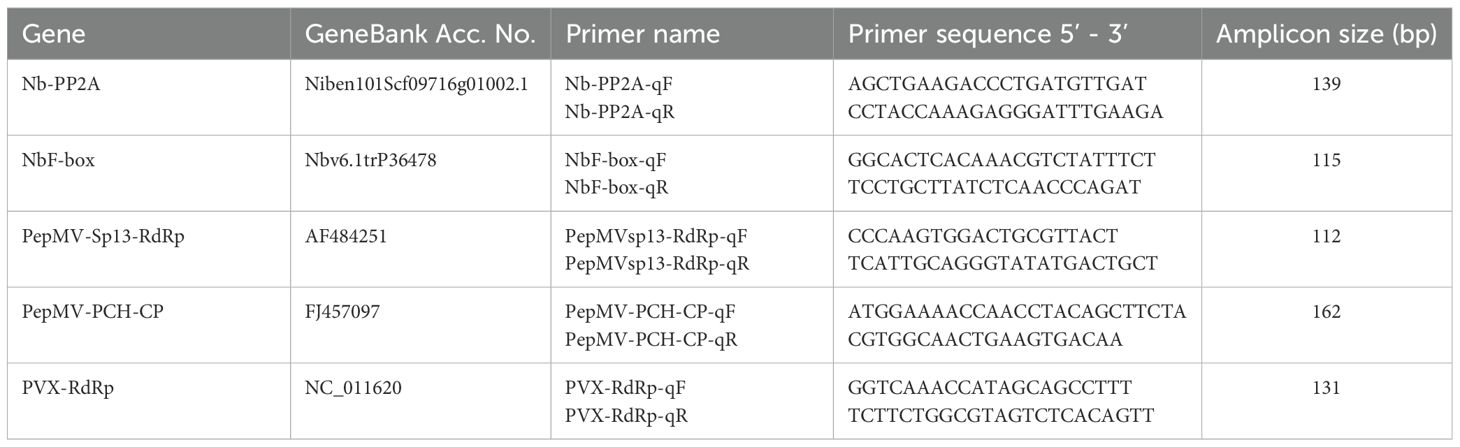
Table 1. Details for the genes and primers used in the study.
Total RNA extraction, cDNA synthesis, and qPCR were performed using materials from individual N. benthamiana plants, each representing a single biological replicate, to generate three distinct technical values. Data were analyzed using the 2−ΔΔCt method (36) and presented as relative levels of gene expression. The geometric means of the reference genes PP2a and F-box were used to normalize the qPCR data. Statistical analysis was performed using the SPSS software (version 19; IBM). Data were presented as means ± SEM (Standard Error of the Mean). One-way analysis of variance (ANOVA) was employed to test the effect of plant genotype on virus accumulation and two-way ANOVA was conducted to assess the combined treatment of cross protection and plant genotype on virus accumulation. Differences were considered to be significant when the p-value was <0.05. Means were ranked by using Tukey’s HSD post hoc test at the 5% level of significance.
3 Results
3.1 Differentiation of PepMV isolates
For the needs of our study, PepMV-Sp13 virions were purified to serve both as inoculum for subsequent “vaccinations” and the production of a polyclonal antiserum. Purified PepMV-PCH 06/104 and PVX virions served as inoculum for “challenge” mechanical inoculations. Their SDS-PAGE analysis showed that despite their nearly identical estimated molecular weights (25.13 and 25 kDa), the CP of the aggressive isolate PCH consistently run distinctly higher than its mild homolog. This property can be used to distinguish the two (Supplementary Figure 1A). Western blot analysis of single virus-infected leaf extracts showed no apparent cross-reactivity of the IgGs produced with plant proteins, PVX-infected plant extracts, or PVX virions (Supplementary Figure 1B). Commercially available PVX CP polyclonal antiserum reacted positively only with PVX-purified virions and infected plant extracts (Supplementary Figure 1B).
The specificity of four DIG-labeled RNA probes synthesized from the PepMV-Sp13, -PCH 06/104, PVX, and BaMV CP genes was examined by northern blot hybridization (Figure 1) against total RNAs extracted from systemically infected leaves of four N. benthamiana genotypes (wt, NahG, DCL2/4, and ago2). Each probe was hybridized with the genomic and subgenomic RNAs of the homologous potexvirus isolate with no apparent cross-reactions with the RNA extracts from mock-inoculated plants or plants infected with any of the other three potexviruses (Figure 1). In the case of the two PepMV isolates, this observation may be explained considering the degree of the nucleotide identity (77.9%) between their CP genes with the highest variability localized at the 5′-terminus. Finally, two pairs of oligonucleotide primers (Table 1), which exhibited specificity in their ability to amplify each PepMV isolate (Supplementary Figure 1C), were tested using RT-qPCR. Overall, several diagnostic tools and methods were generated to distinguish between the two PepMV isolates in subsequent mixed infections.
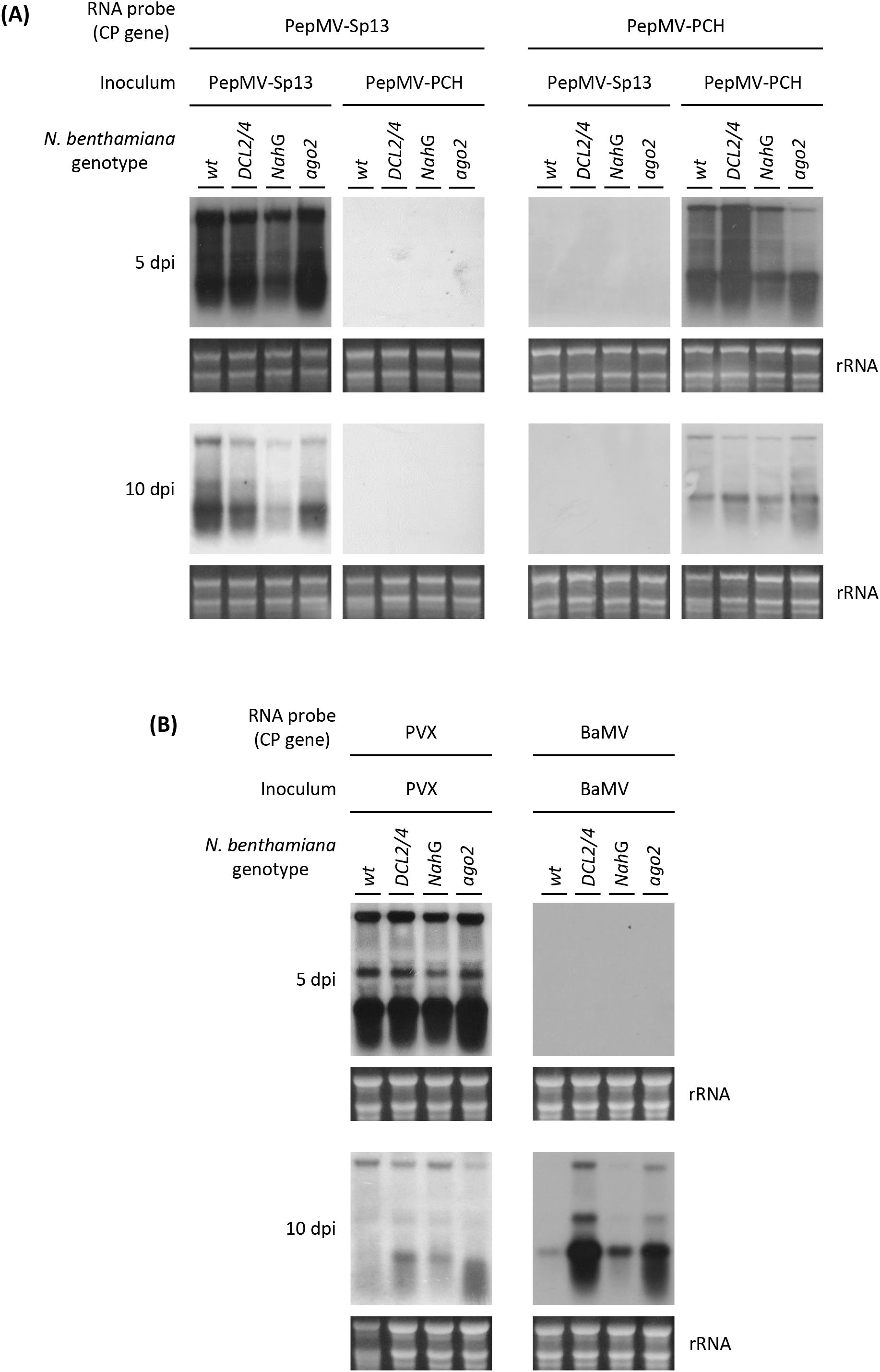
Figure 1. Northern blot hybridization analyses of total RNA extracts from four Nicotiana benthamiana genotypes (wt, DCL2/4, NahG, and ago2) infected with each of the following potexviruses: (A) Pepino mosaic virus mild (PepMV-Sp13), aggressive isolate (PepMV-PCH), (B) Potato virus X (PVX), and Bamboo mosaic virus (BaMV). Three plants from each genotype were mechanically inoculated with mock (0.1 M Tris-borate buffer) or purified virions (20 μgr). Newly formed leaves were collected at 5 or 10 dpi, and total RNA was extracted, pooled, and Northern blot analyzed using DIG-labeled negative (-)-stranded RNA probes from the full-length CP gene of each virus. For PepMV, 1 μg of total RNA was used; for PVX and BaMV, 5 μg of total RNA was used.
ιn cross-protection studies, the time-interval between “vaccination” and “challenge” inoculation is important (4, 5, 21). Single infections using purified virions of PepMV-Sp13, -PCH 06/104, and PVX on four genetically distinct N. benthamiana plant genotypes, unlike BaMV, showed that they reached high RNA levels at 5 dpi but significantly decreased at 10 dpi (Figure 1). During this short period, no alterations on the induced symptomatology were observed (Supplementary Figure 2), but the significant drop of the viral RNA levels resembles the “recovery” phenomenon first described in tobamoviruses (37) and later connected with RNA silencing and cross-protection (6). As our experimental plant growth conditions typically induced flowering in N. benthamiana plants no later than 15 dpi, two time-intervals (5 and 10 days) were adapted in the cross-protection experiments. As BaMV infection was detectable after 10 dpi (Figure 1), it was decided that no “challenge” inoculations would be carried out with this virus.
3.2 PepMV cross-protection in ago2 plants blocks superinfection with the aggressive isolate systemically
The potential involvement of RNA silencing in cross-protection was initially investigated in N. benthamiana ago2-mutant plants because of the dominant role of this protein in anti-PepMV defense (26, 27). In all superinfected plants, both PepMV-Sp13 CP and RNAs were detected systemically, unlike the “challenge” PepMV aggressive isolate (Figures 2, 3). Protein sequencing analysis revealed that the systemic leaf samples contained only the PepMV mild CP and not the aggressive one (data not shown). Overall, protein and RNA analyses failed to detect any PepMV-PCH 06/104 accumulation in newly formed leaves of all “vaccinated” plants. A second set of experiments (time interval: 10 days) generated identical results to those described above (data not shown) and collectively indicated the existence of a protective mechanism that efficiently targets the “challenge” virus isolate in newly formed leaves for several days.
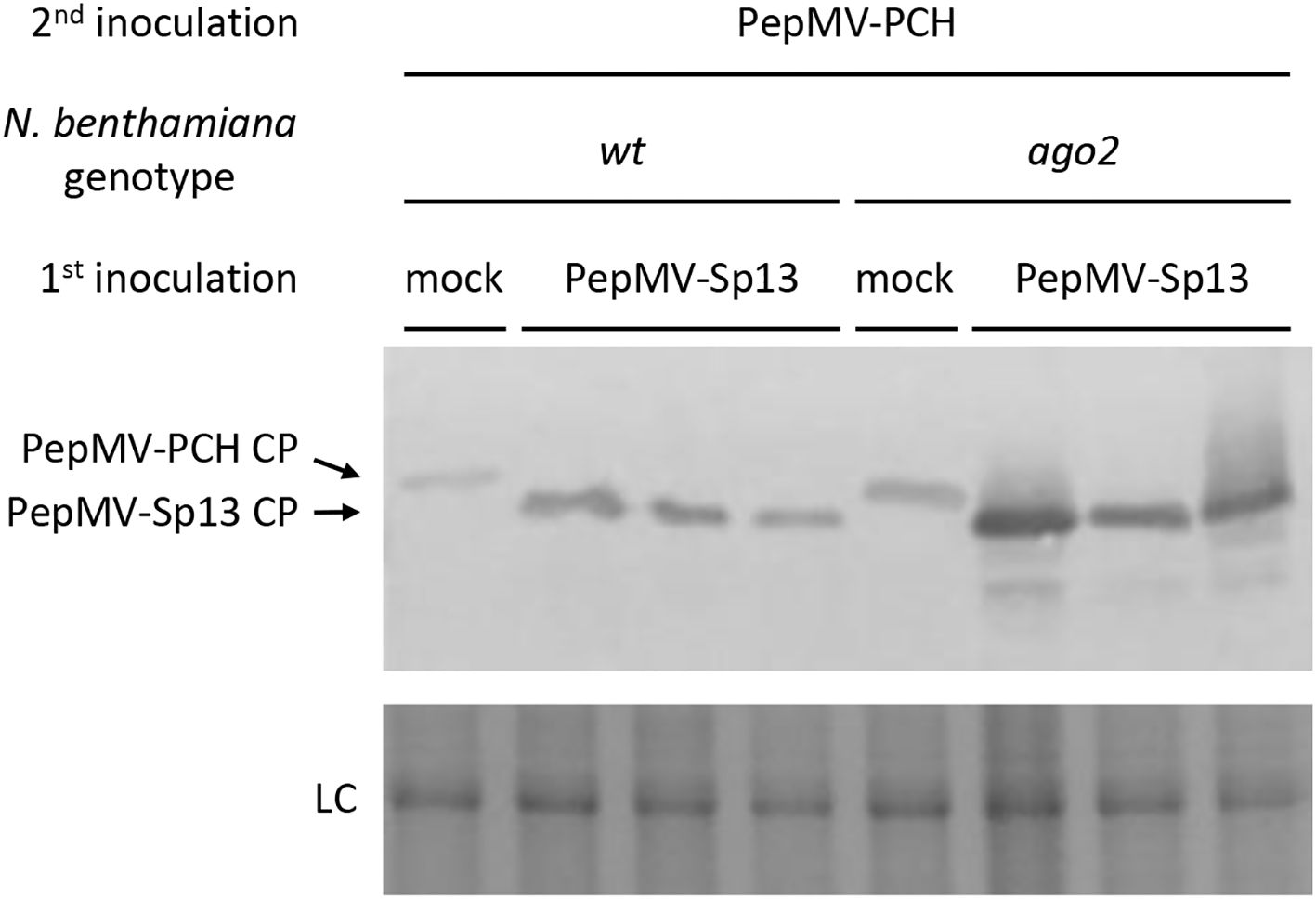
Figure 2. Western blot analysis of total leaf plant proteins using the α-PepMV-Sp13 polyclonal antiserum (dilution: 1:3000; this study). Nicotiana benthamiana wt and ago2-mutant plants were either mock- or PepMV mild (Sp13) isolate mechanically inoculated and then “challenge”-inoculated with the aggressive (PCH) isolate after 5 days. Newly formed leaves were collected 3 days post “challenge” inoculation for analysis. Coomassie Brilliant Blue R-250 staining of the rubisco large subunit was used as a protein loading control (LC).
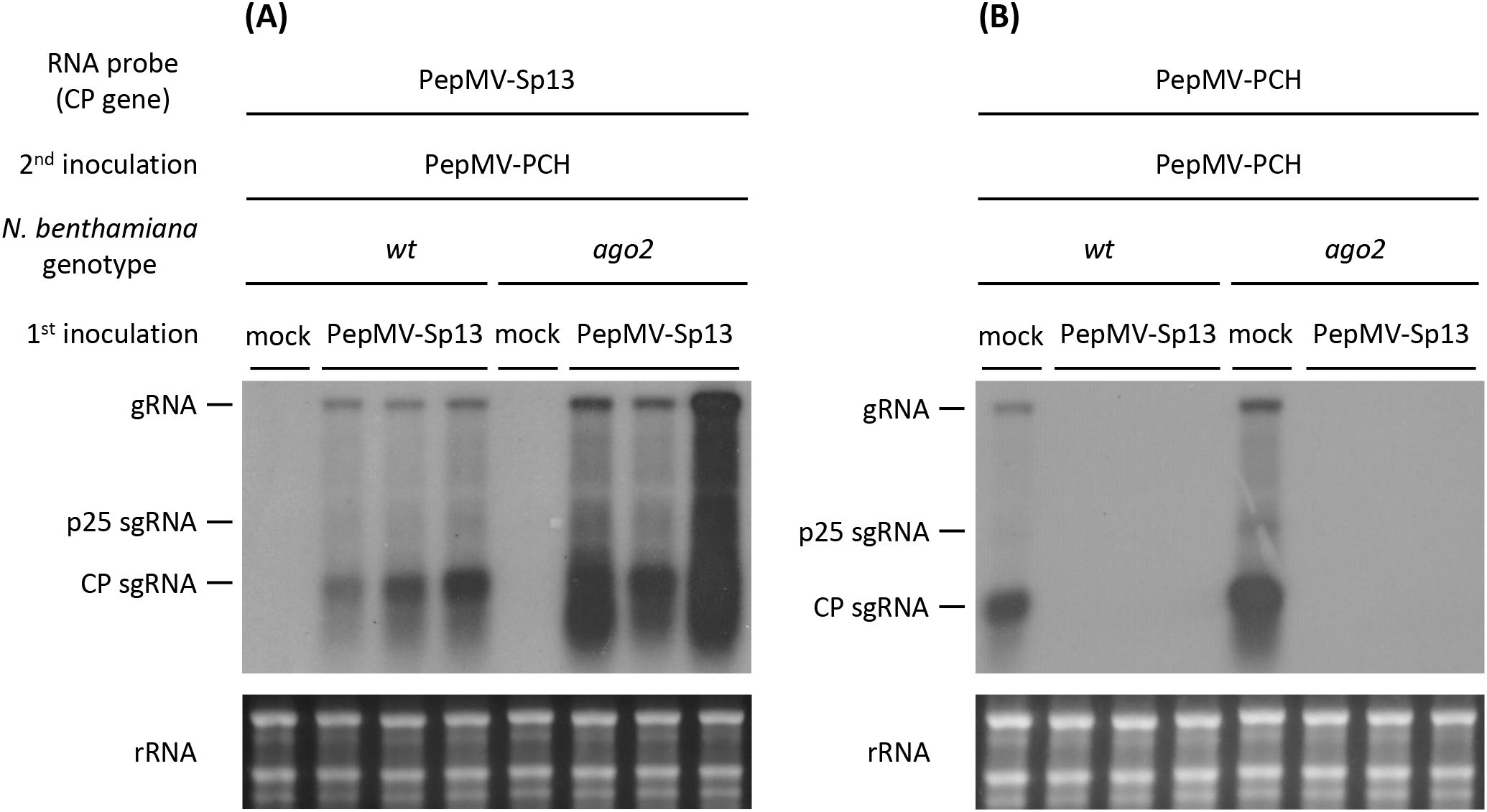
Figure 3. Northern blot analysis of total RNAs extracted from newly formed leaves of Nicotiana benthamiana (wt and ago2-mutant) plants that were either mock- or PepMV-Sp13 (mild) isolate “vaccinated” prior to “challenge” inoculation with the PepMV-PCH (aggressive) isolate (5 days later). Total RNAs were hybridized against negative-stranded DIG-labeled RNA probes from (A) the mild or (B) aggressive PepMV isolate CP genes.
RT-qPCR quantification of RNAs from both PepMV isolates in the “vaccinated” and mock-inoculated (control) groups of plants was also performed. As shown in Figure 4, the RNAs of the PepMV-PCH 06/104 isolate remained undetectable in both wt and ago2 plants pre-inoculated with Sp13 but reached high levels in the control group (mock-pre-inoculated). The highest RNA levels of PepMV-PCH 06/104 were observed in the N. benthamiana ago2 plants (time interval: 5 days; Figure 4A), a result consistent with the predominant role of AGO2 in anti-PepMV defense (26, 27). RT-qPCR confirmed the results of western and northern blots, which revealed the presence of the PepMV-Sp13 isolate RNAs in both wt and ago2 plants (data not shown) and showed that “vaccination” with the mild PepMV isolate induces an efficient and active protective mechanism for several days, completely restricting the aggressive PepMV isolate from newly formed leaves. Previous PepMV studies showed that the anti-PepMV activities of AGO1A, AGO5, and AGO10 become apparent only when AGO2 is mutated (27) and as a result, the involvement of RNA silencing cannot be entirely excluded from the observed phenomenon.
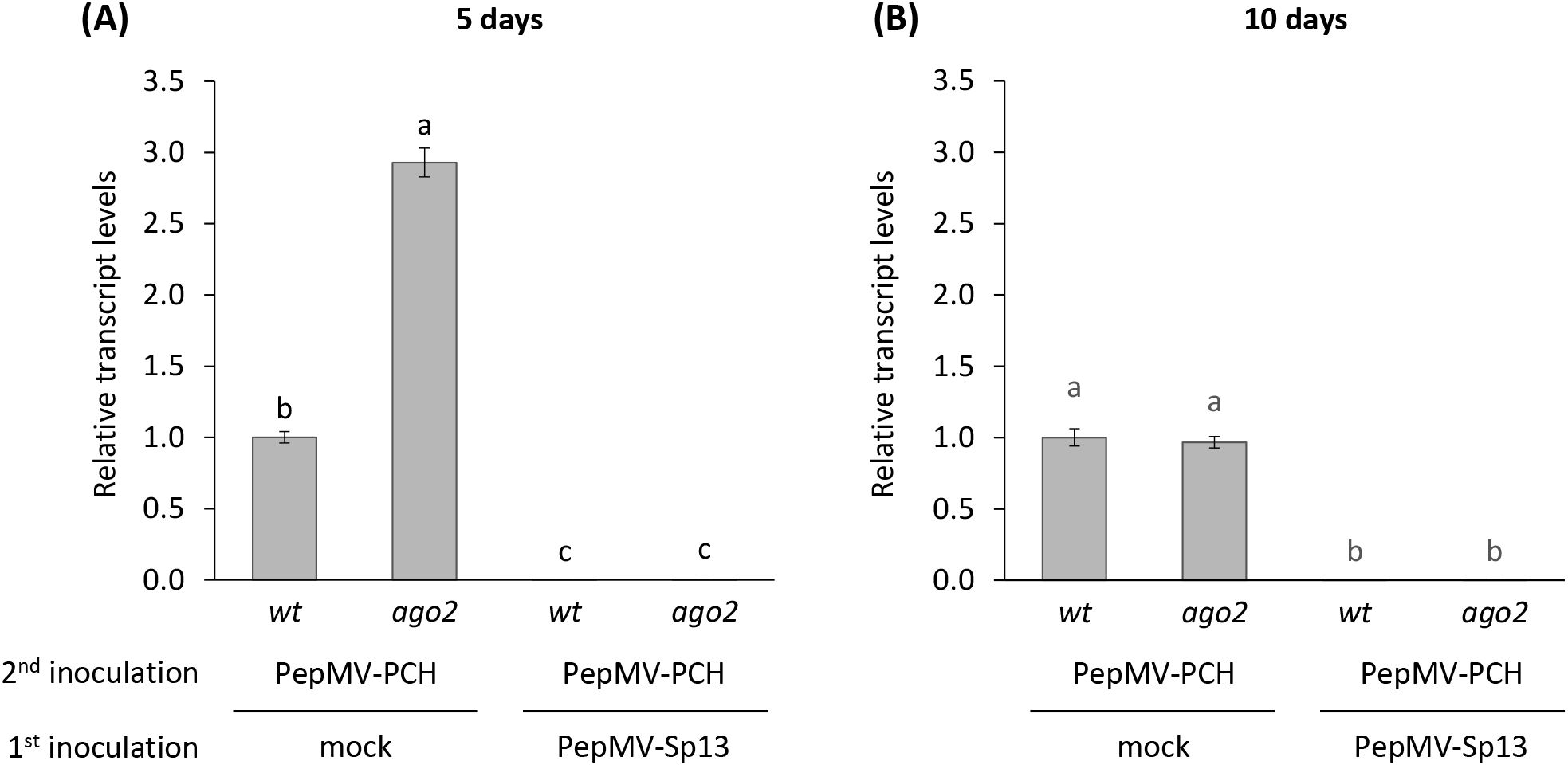
Figure 4. Relative RNA transcript levels of the PepMV-PCH (aggressive) isolate in cross-protection experiments using two Nicotiana benthamiana genotypes (wt and ago2). Plants were either mock- or PepMV-Sp13-”vaccinated” and then “challenge”-inoculated with the aggressive PepMV isolate after 5 or 10 days (A, B), respectively. Newly formed leaves were collected 3 days after “challenge” inoculation and analysed. qPCR data were normalized to the geometric mean of the reference genes PP2a and F-box. Graphical data are presented as mean ± standard error of the mean (s.e.m.), with n = 3 biological replicates. Letters indicate statistical significance based on a two-way ANOVA with Tukey’s HSD post hoc analysis (p < 0.05); samples sharing letters are not significantly different.
3.3 PVX superinfection is not efficiently arrested in ago2 plants
To better comprehend the nature of the cross-protection mechanism induced by PepMV-Sp13 pre-inoculation, we examined its efficiency and specificity against an alternative “challenge” virus, PVX, the prototype potexvirus and a candidate virus in mixed infections on tomato plants in the field. As above, PepMV mild isolate RNAs from systemic leaves of the “vaccinated” plants were verified using the homologous CP RNA probe, but PVX RNAs were detectable only after long exposures with greatly decreased levels when compared with extracts from mock-pre-inoculated plants (control, Figure 5). The presence of PVX CPs in western blots indicated sporadic PVX infections in some PepMV-Sp13 pre-inoculated wt and ago2 N. benthamiana plants (Figure 6) but RT-qPCR analysis provided additional support that PVX RNA levels were significantly reduced when compared to mock-pre-inoculated plants (Figure 7). These results indicate that also for the PepMV-PVX combination, a cross-protective mechanism is initiated, which nevertheless does not completely block superinfection possibly because of the genetically distant nature of the “challenge” virus.
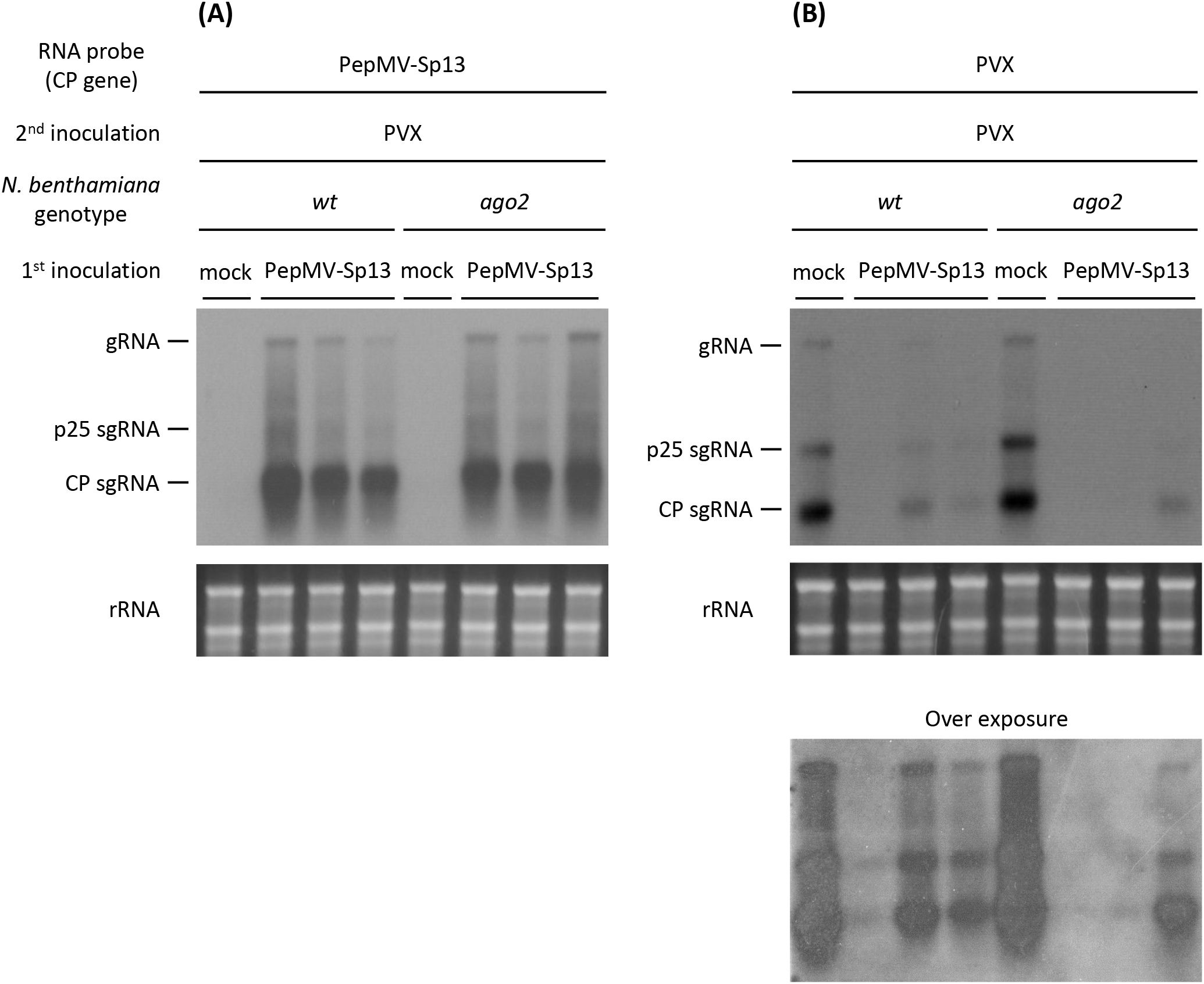
Figure 5. Northern blot analysis of total RNAs extracted from newly formed leaves of Nicotiana benthamiana (wt and ago2-mutant) plants that were either mock- or PepMV-Sp13 (mild) isolate “vaccinated” and then “challenge” PVX-inoculated after 5 days. Newly formed leaves were collected 3 days after “challenge” inoculation. RNAs were hybridized against negative-stranded DIG-labeled RNA probes from (A) the PepMV mild isolate or (B) PVX CP genes.
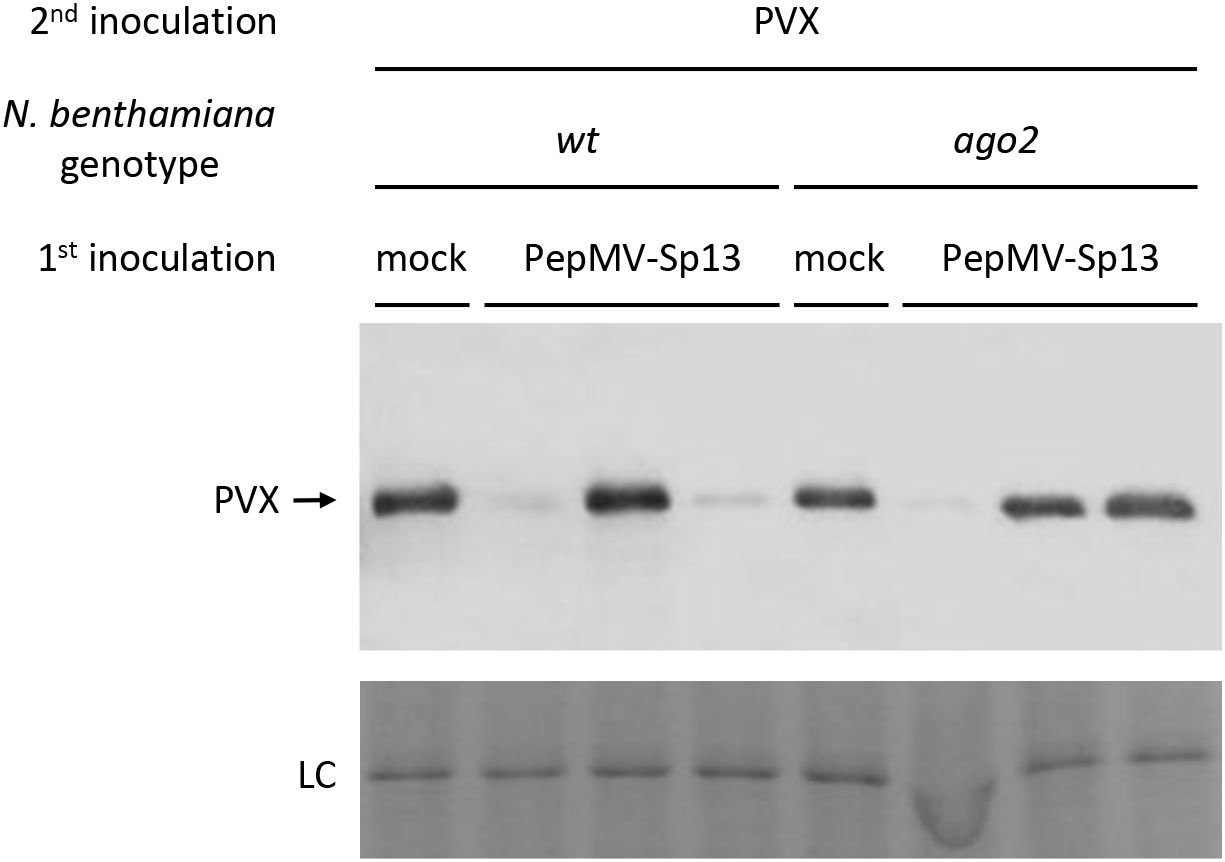
Figure 6. Western blot analysis of total leaf plant proteins collected from cross-protection experiments in N. benthamiana (wt and ago2-mutant) plants using PepMV mild and PVX virions. The plants were either mock- or PepMV-Sp13 “vaccinated” and then “challenge” inoculated using PVX virions after 5 days. Newly formed leaves were collected 3 days after “challenge” inoculation, and protein extracts were separated in 12% SDS-PAGE, membrane blotted, and hybridized against the PVX polyclonal antiserum (dilution 1:2000; Loewe Biochemica). Coomassie Brilliant Blue R-250 staining of the rubisco large subunit was used as a protein loading control (LC).
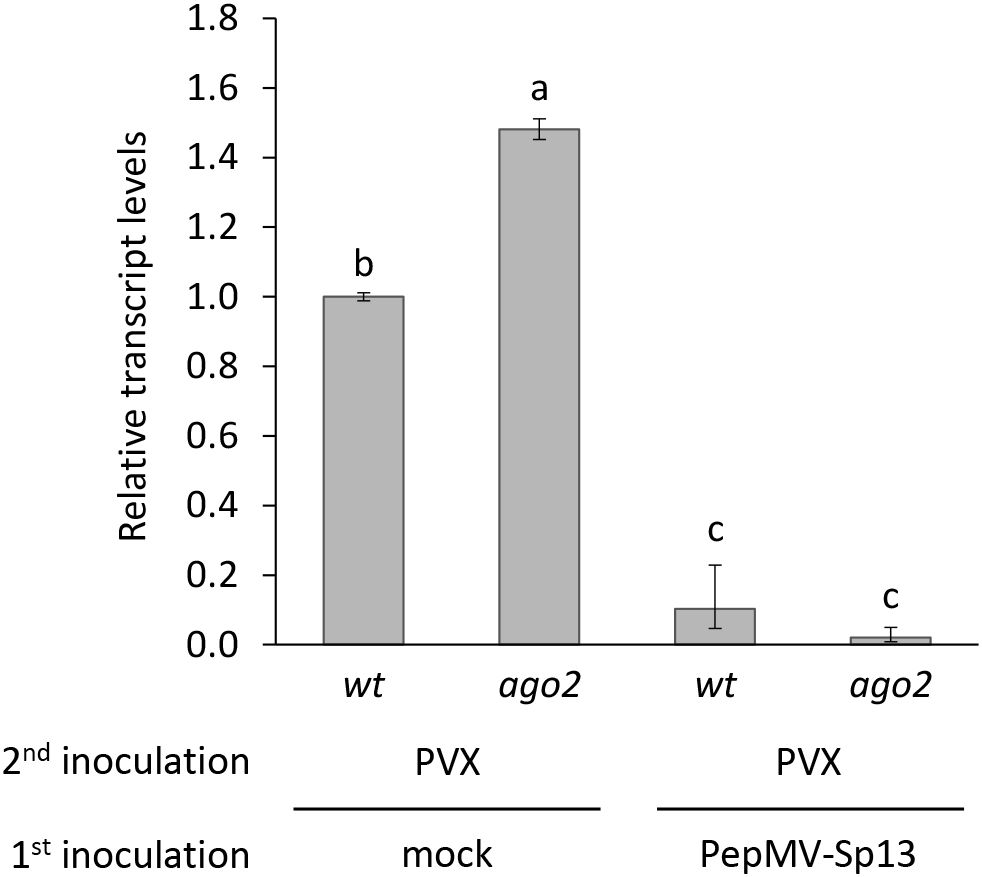
Figure 7. Relative RNA transcript levels of PVX in cross-protection experiments in two Nicotiana benthamiana genotypes (wt and ago2). Plants were either mock- or PepMV-Sp13-”vaccinated” and after 5 days “challenge”-inoculated with PVX virions. Newly-formed leaves were collected 3 days after “challenge” inoculation and analysed. qPCR data were normalized to the geometric mean of the reference genes PP2a and F-box. Graphical data are presented as mean ± standard error of the mean (s.e.m.), with n = 3 biological replicates. Letters indicate statistical significance based on a two-way ANOVA with Tukey’s HSD post hoc analysis (p < 0.05); samples sharing letters are not significantly different.
3.4 Pre-inoculated DCL2/4-suppressed N. benthamiana plants with the PepMV mild isolate completely restrict severe isolate superinfection in inoculated and newly formed leaves
Dicers are essential for RNA silencing in plants and are responsible for the recognition and cleavage of dsRNA-producing siRNAs as part of the antiviral defense system. We used N. benthamiana DCL2/4-deficient plants to examine whether pre-inoculation with the PepMV mild isolate protects plants from superinfection with the aggressive isolate. Similar to the ago2 mutants, cross-protection was activated, exhibiting “immunity” throughout the experimentation. Both western and hybridization analyses showed no detectable levels of CP and RNA of the PepMV aggressive isolate in the systemic and also local leaves (Figures 8, 9). These results were confirmed by RT-qPCR analyses (Figure 10), where the “challenge” virus isolate remained (unlike the “vaccine”; data not shown) undetectable at 8 days post “vaccination”. Overall, the generated data are more in line with a model that does not require triggering of a defense response and rather suggest the “exclusion” of the aggressive isolate from sites or resources. Interpreting these results cannot entirely exclude the complete redundancy of AGO2 and DCL2/4, but additional reasoning for the non-involvement of RNA silencing as a prime mechanism for the observed “immunity” has been generated.
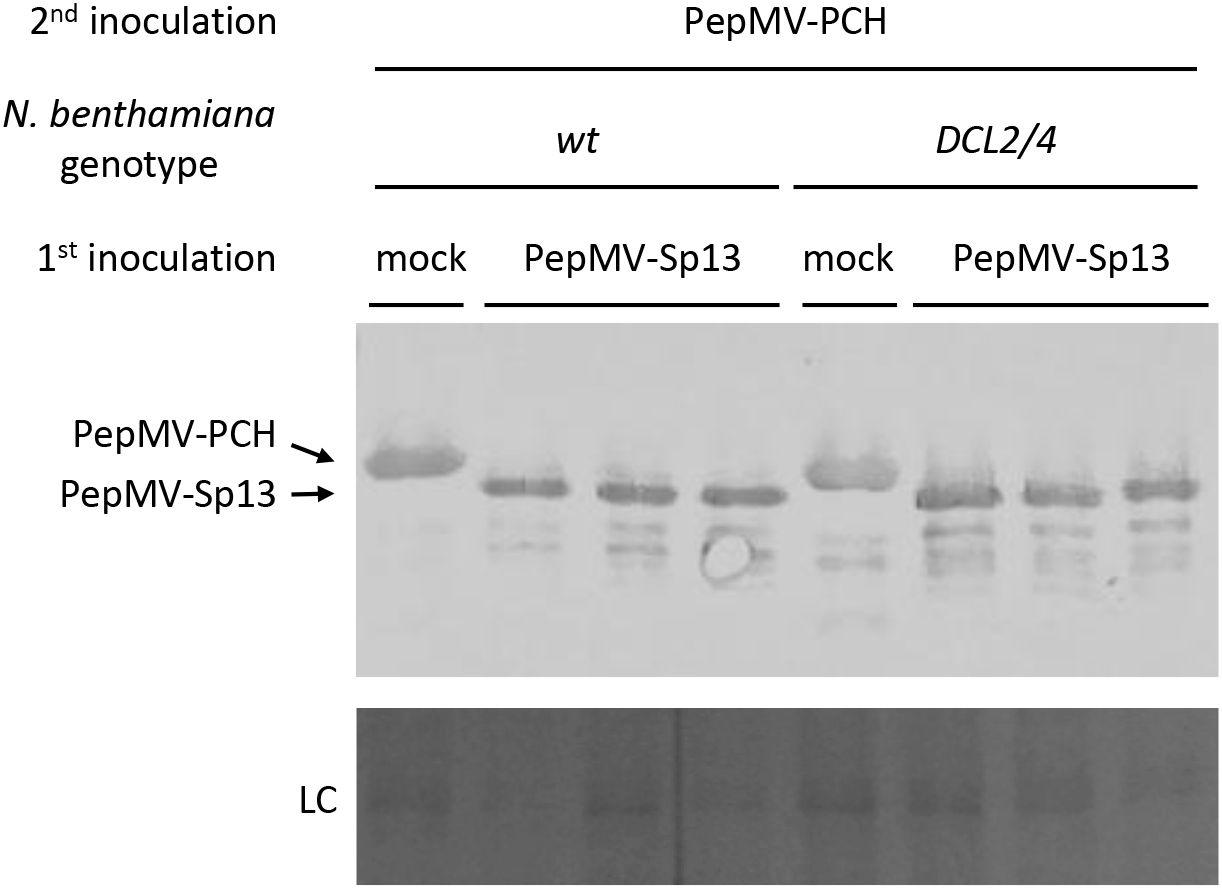
Figure 8. Western blot analysis of total leaf plant proteins collected from cross-protection experiments using PepMV mild and severe isolates to inoculate N. benthamiana (wt and DCL2/4-suppressed) plants. The plants were either mock- or PepMV-Sp13 “vaccinated” and 5 days later “challenge”-inoculated with the aggressive PepMV-PCH isolate. Inoculated leaves were collected 3 days after “challenge” inoculation, and protein extracts were separated in 12% SDS-PAGE, membrane blotted, and hybridized against the α-PepMV-Sp13 polyclonal antiserum (dilution 1:3000; this study). Coomassie Brilliant Blue R-250 staining of the rubisco large subunit was used as a protein loading control (LC).
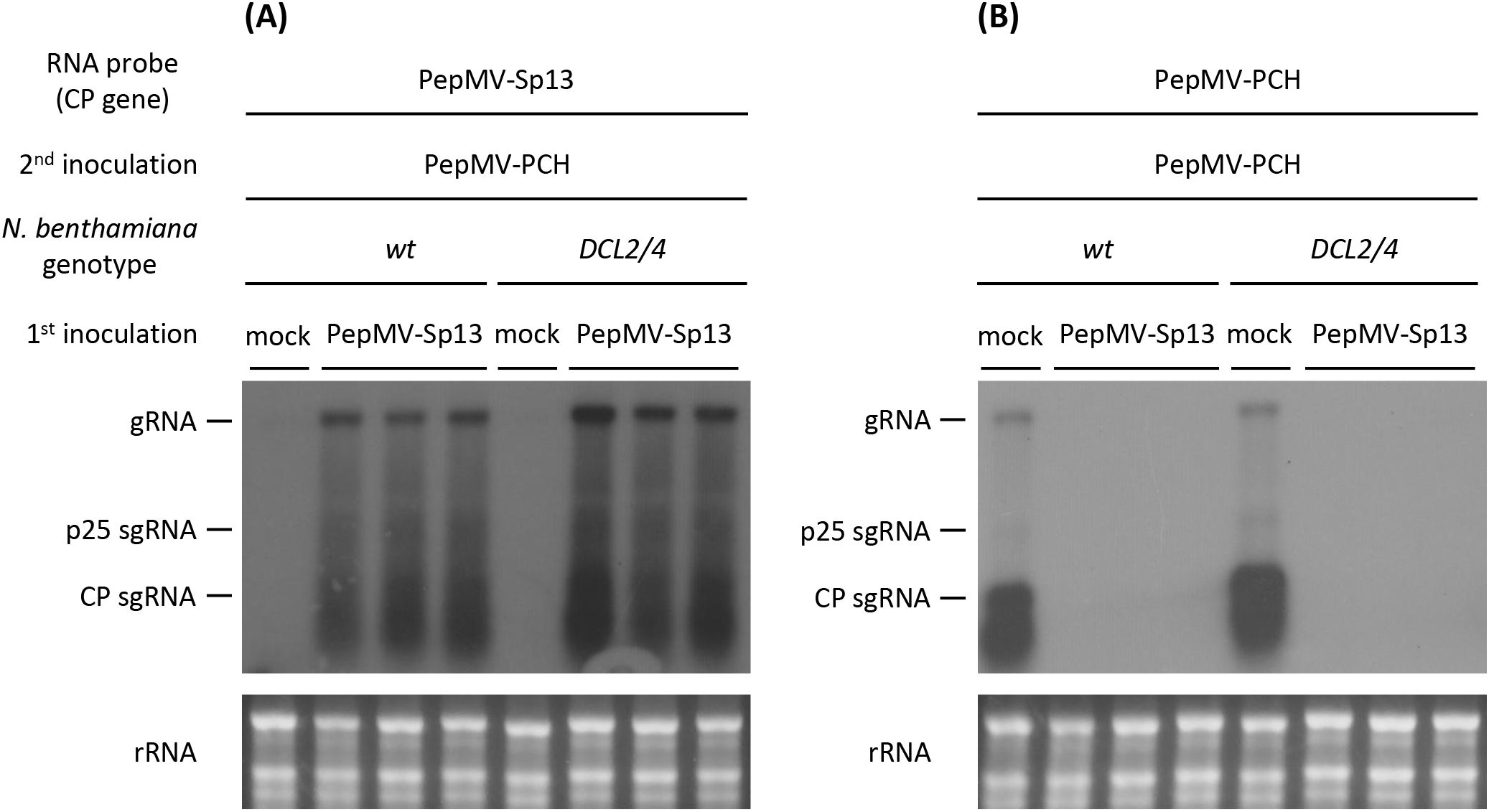
Figure 9. Northern blot analysis of total RNAs extracted from newly formed leaves of N. benthamiana (wt and DCL2/4 mutant) plants used in cross-protection experiments. The plants were either mock- or PepMV-Sp13 “vaccinated” and then “challenge” inoculated with PepMV-PCH virions after 5 days. Newly formed leaves were collected 3 days after “challenge” inoculation. RNAs were hybridized against negative-stranded DIG-labeled RNA probes from (A) PepMV-Sp13 or (B) -PCH isolate CP genes.
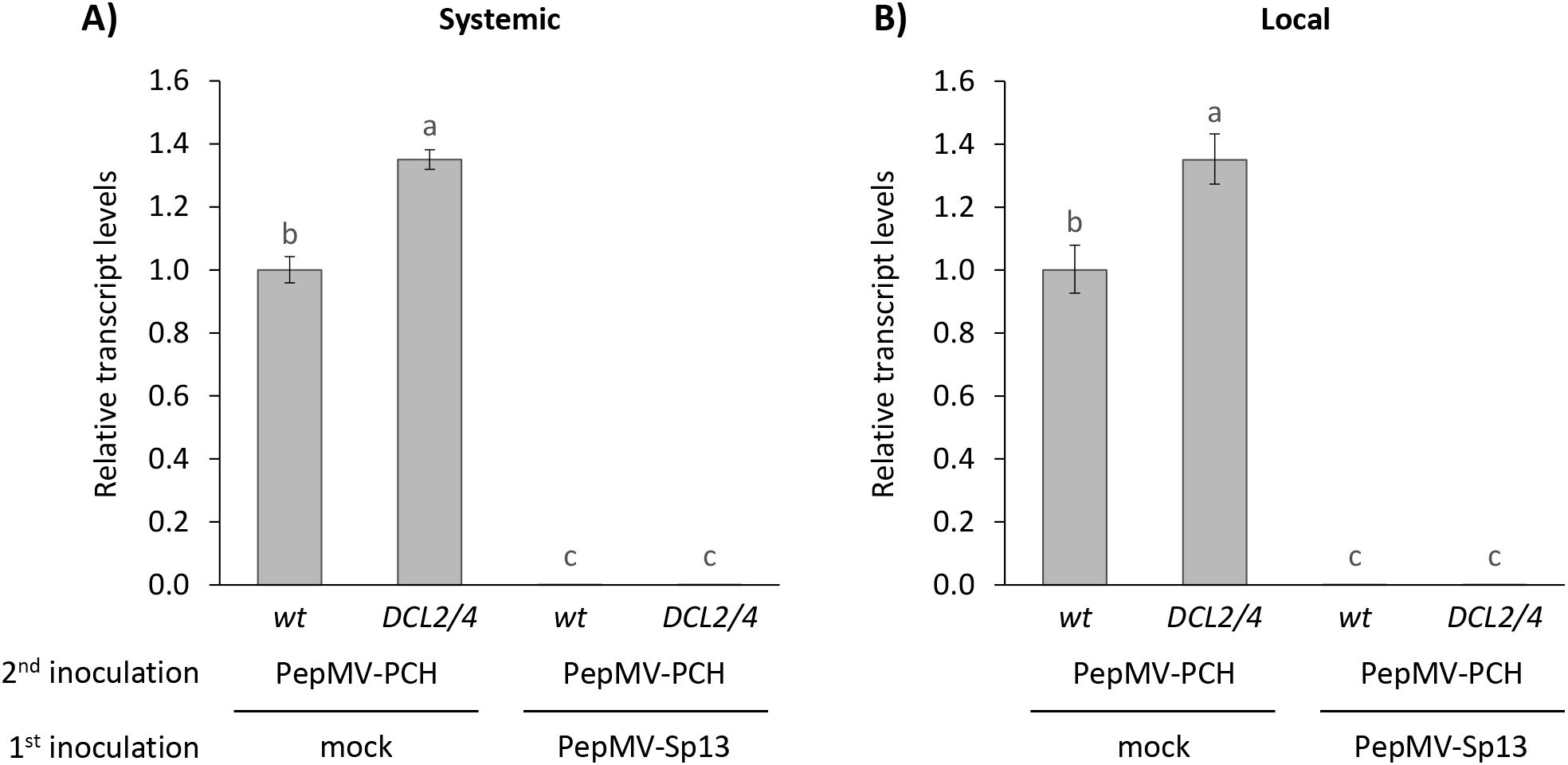
Figure 10. Relative RNA transcript levels of the “challenge” PepMV-PCH aggressive isolate in (A) newly formed (systemic) or (B) inoculated (local) leaves of Nicotiana benthamiana plants (wt or DCL2/4 genotypes) in cross-protection experiments. Plants were either mock- or PepMV-Sp13 “vaccinated” and 5 days later “challenge”-inoculated with the aggressive isolate. Leaves were collected 3 days after “challenge” inoculation for RT-qPCR analysis. qPCR data were normalized to the geometric mean of the reference genes PP2a and F-box. Graphical data are presented as mean ± standard error of the mean (s.e.m.), with n = 3 biological replicates. Letters indicate statistical significance based on a two-way ANOVA with Tukey’s HSD post hoc analysis (p < 0.05); samples sharing letters are not significantly different.
4 Discussion
Plant viruses pose a serious threat to food crops and cause significant annual economic losses. The identification of sustainable control measures for plant viruses is a key challenge in modern agriculture. The consequences of plant viral infections are frequently exacerbated by the impact of climate change on hosts and vectors, and by the mobility of symptomless infected plant propagative material worldwide. The production of virus-resistant cultivars by conventional breeding is a long and costly procedure; however, promising technologically advanced genetic approaches are not well accepted by consumers. Cross-protection, which shares similarities to mammalian “vaccination”, has successfully been employed in some cases but remains underexploited partly because of the lack of a convincing explanation for its mechanism (4, 5).
In our preliminary experiments, PepMV-Sp13, -PCH 06/104, or PVX infections (but not BaMV, which shares distinct biological properties) in newly formed leaves were associated with high viral RNA loads at 5 dpi, which were lower at 10 dpi. Although these alterations did not correspond with differential symptomatology (e.g., “dark green islands”), the lowered viral loads were reported in the 1990s as “recovery” and were also later associated with RNA-mediated cross-protection (6, 38). This observation helped to determine two specific post-infection points (5 and 10 dpi) to be followed in subsequent cross-protection experiments and indicated the activation of RNA silencing following primary inoculations. The results of the hybridization experiments were somewhat surprising, as both PepMV CP RNA probes were hybridized exclusively to their own isolates possibly due to the relatively low degree of nucleotide identity between the CP genes. Overall, several useful methods and tools were initially developed to discriminate between the two PepMV isolates.
Both PepMV-PCH and PVX accumulated at higher levels in mock-”vaccinated” N. benthamiana plants, in which elements of the RNA silencing mechanism had been compromised, when compared with wt plants, demonstrating the importance of both factors (AGO2 and DCL2/4) in RNA silencing and anti-viral defense upon infection. In the actual cross-protection experiments, similar to those conducted by Alcaide et al. (39) on tomatoes (cv. Moneymaker), “vaccination” with the PepMV mild isolate conferred protection against the aggressive isolate. Specifically in our case, superinfection was completely blocked in wt and in ago2 and DCL2/4-deficient N. benthamiana plants. As leaves were collected and analyzed at 8 and 13 days after “vaccination,” the arrest of the challenge isolate infection seems absolute in inoculated (wt, DCL2/4) and systemic (wt, ago2, DCL2/4) leaves. This result indicates the early activation and maintenance of an efficient systemic protection mechanism over approximately 2 weeks.
In general (4, 5), cross-protection works better against two genetically related viruses. To examine this, we also used PVX as a challenge virus, considering that numerous studies have reported that a pre-inoculated virus may also protect from superinfection by a genetically distinct “challenge” virus species. For example, pre-inoculation with Arabis mosaic virus-S protects Chenopodium quinoa from Grapevine fanleaf virus-F13 superinfection (40), and pre-inoculation with Soilborne mosaic virus protects sugar beet against subsequent Beet necrotic yellow vein virus infection (41). In the present study, a PepMV-PVX cross-protective mechanism was also induced in wt and ago2 plants that significantly reduced PVX RNA levels but did not completely block infection.
Previous PepMV studies have shown that the anti-PepMV activities of AGO1A, AGO5, and AGO10 become apparent only when AGO2 is mutated (27) and thus, the high multilevel redundancy of the AGO2 and DCL2/4 components cannot be excluded in our experimental set up. Ma et al. [2015 (42),] reported that AGO2 and AGO4 mutant plants can recover from Tobacco rattle virus-GFP inoculations and proposed an alternative theory involving translational repression of viral transcripts. In DCL1, DCL2, and DCL3 mutants, various Dicer functions appear redundant, as assessed by measuring Cucumber mosaic virus and TuMV titers and siRNA levels (9). Our results including the observed cross protection of the genetically distant PepMV-PVX seem to be more in line with a model that does not require the triggering of a sequence-specific defense response and rather suggest the “exclusion” of the “challenge” virus from essential sites or resources for infection. According to the SIE theory, related viral strains compete with each other to reach vital host-cell sites and resources and, once established, cannot be displaced by the competing strain. Previous studies on CTV, TCV, and TuMV (13–15) found that virus-encoded proteins (p33, p28, p3, and NIa-Pro) are involved in the role(s) of elicitors and targets of SIE as the primary mechanism for the observed protection. Relevant hypotheses need to be addressed for PepMV in the near future.
In conclusion, “vaccination” with the PepMV mild isolate completely blocks the aggressive isolate and significantly reduces PVX superinfection in RNA silencing-compromised N. benthamiana mutant plants, suggesting a prime protection mechanism other than RNA silencing.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
TS: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing – review & editing, Software. AG: Data curation, Formal Analysis, Investigation, Methodology, Software, Validation, Writing – review & editing. AR: Data curation, Formal Analysis, Investigation, Methodology, Software, Writing – review & editing. ML: Writing – review & editing, Resources. KF: Resources, Writing – review & editing. TE: Resources, Writing – review & editing, Investigation, Methodology, Software. IL: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work in K. Fátyol’s laboratory was supported by a grant from the National Research Development and Innovation Office, Hungary (K142626).
Acknowledgments
The authors thank LabSupplies Scientific (Athens, Greece) for providing the Real-Time PCR machine (SACycler-96 Real PCR System; Sacace Biotechnologies, Italy) and Prof. J. Jones (Sainsbury Laboratory, UK) and K. Kalantidis (Univ. Crete, Greece) for providing NahG and DCL2/4-5i-suppressed transgenic N. benthamiana seeds, respectively. Special thanks to Ms Hadil Rbib and Rosa L. for valuable technical assistance and discussions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fviro.2025.1584535/full#supplementary-material
References
1. McKinney HH. Mosaic diseases in the Canary Islands, West Africa and Gibraltar. J Agric. (1929) 39:557–78. doi: 10.5555/19301100507
2. Hull R. Matthews’ Plant Virology. 4th. London and San Diego: Academic Press (2002). p. 1029. doi: 10.1016/B978-0-12-361160-4.X5050-6
3. Pechinger K, Chooi KM, MacDiarmid RM, Harper SJ, and Ziebell H. A new era for mild strain cross-protection. Viruses. (2019) 11:670. doi: 10.3390/v11070670
4. Gal-On A and Shiboleth YM. Cross-protection. In: Loebenstein G and Carr JP, editors. Natural Resistance Mechanisms of Plants to Viruses. Springer, Dordrecht (2006). p. 261–88. doi: 10.1007/1-4020-3780-5_12
5. Ziebell H and Carr JP. Cross-protection a century of mystery. Adv Virus Res. (2010) 76:211–64. doi: 10.1016/S0065-3527(10)76006-1
6. Ratcliff FG, MacFarlane SA, and Baulcombe DC. Gene silencing without DNA: RNA-mediated cross-protection between viruses. Plant Cell. (1999) 11:1207–16. doi: 10.1105/tpc.11.7.1207
8. Dunoyer P, Melnyk C, Molnar A, and Slotkin RK. Plant mobile small RNAs. Cold Spring Harb Perspect Biol. (2013) 5:a017897. doi: 10.1101/cshperspect.a017897
9. Ziebell H and Carr JP. Effects of dicer-like endoribonucleases 2 and 4 on infection of Arabidopsis thaliana by cucumber mosaic virus and a mutant virus lacking the 2b counter-defence protein gene. J Gen Virol. (2009) 90:2288–92. doi: 10.1099/vir.0.012070-0
10. Zhang XF, Guo J, Zhang X, Meulia T, Paul P, Madden LV, et al. Random plant viral variants attain temporal advantages during systemic infections and in turn resist other variants of the same virus. Sci Rep. (2015) 5:15346. doi: 10.1038/srep15346
11. Folimonova SY. Developing an understanding of cross-protection by citrus tristeza virus. Front Microbiol. (2013) 4:76. doi: 10.3389/fmicb.2013.00076
12. Zhang XF, Zhang S, Guo Q, Sun R, Wei T, and Qu F. A new mechanistic model for viral cross protection and superinfection exclusion. Front Plant Sci. (2018) 9:40. doi: 10.3389/fpls.2018.00040
13. Folimonova SY. Superinfection exclusion is an active virus-controlled function that requires a specific viral protein. J Virol. (2012) 86:5554–61. doi: 10.1128/JVI.00310-12
14. Zhang XF, Sun R, Guo Q, Zhang S, Meulia T, Halfmann R, et al. A self-perpetuating repressive state of a viral replication protein blocks superinfection by the same virus. PloS Pathog. (2017) 13:e1006253. doi: 10.1371/journal.ppat.1006253
15. Nunna H, Qu F, and Tatineni S. P3 and NIa-Pro of Turnip mosaic virus are independent elicitors of superinfection exclusion. Viruses. (2023) 15:1459. doi: 10.3390/v15071459
16. Grant TJ and Costa AS. A mild strain of the tristeza virus of citrus. Phytopathology. (1951) 41:114–22.
17. Yarden G, Hemo R, Livne H, Maoz E, Lev E, Lecoq H, et al. Cross-protection of Cucurbitaceae from Zucchini yellow mosaic Potyvirus. Acta Hortic. (2000) 510:349–59. doi: 10.17660/ActaHortic.2000.510.56
18. Rast ATB. MII-16, an artificial symptomless mutant of tobacco mosaic virus for seedling inoculation of tomato crops. Neth J Plant Pathol. (1972) 78:110–2. doi: 10.1007/BF01980475
19. Yeh SD, Gonsalves D, Wang HL, Namba R, and Chiu RJ. Control of papaya ringspot virus by cross protection. Plant Dis. (1988) 72:375–80. doi: 10.1094/PD-72-0375
20. Agüero J, Gómez-Aix C, Sempere RN, García-Villalba J, García-Núñez J, Hernando Y, et al. Stable and broad spectrum cross-protection against Pepino mosaic virus attained by mixed infection. Front Plant Sci. (2018) 9:1810. doi: 10.3389/fpls.2018.01810
21. Hernando Y and Aranda MA. Cross-protection against Pepino mosaic virus, more than a decade of efficient disease control. Ann Appl Biol. (2024) 184:174–82. doi: 10.1111/aab.12884
22. Chewachong GM, Miller SA, Blakeslee JJ, Francis DM, Morris TJ, and Qu F. Generation of an attenuated, cross-protective Pepino mosaic virus variant through alignment-guided mutagenesis of the viral capsid protein. Phytopathology. (2015) 105:126–34. doi: 10.1094/PHYTO-01-14-0018-R
23. Aguilar JM, Hernández-Gallardo MD, Cenis JL, Lacasa A, and Aranda MA. Complete sequence of the Pepino mosaic virus RNA genome. Arch Virol. (2002) 147:2009–15. doi: 10.1007/s00705-002-0848-9
24. Mathioudakis MM, Rodríguez-Moreno L, Sempere RN, Aranda MA, and Livieratos I. Multifaceted capsid proteins: multiple interactions suggest multiple roles for Pepino mosaic virus capsid protein. Mol Plant-Microbe Interact. (2014) 27:1356–69. doi: 10.1094/MPMI-07-14-0195-R
25. Olsthoorn RCL, Owen CA, and Livieratos IC. Role of an RNA pseudoknot involving the polyA tail in replication of Pepino mosaic potexvirus and related plant viruses. Sci Rep. (2022) 12:11532. doi: 10.1038/s41598-022-15598-5
26. Alcaide C, Donaire L, and Aranda MA. Transcriptome analyses unveiled differential regulation of AGO and DCL genes by Pepino mosaic virus strains. Mol Plant Path. (2022) 23:1592–607. doi: 10.1111/mpp.13249
27. Ludman M, Szalai G, Janda T, and Fátyol K. Hierarchical contribution of Argonaute proteins to antiviral protection. J Exp Bot. (2023) 74:6760–72. doi: 10.1093/jxb/erad327
28. Mathioudakis MM, Veiga RSL, Canto T, Medina V, Mossialos D, Makris AM, et al. Pepino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol Plant Path. (2013) 14:589–601. doi: 10.1111/mpp.12034
29. Mathioudakis MM, Khechmar S, Owen CA, Medina V, Ben Mansour K, Tomaszewska W, et al. A thioredoxin domain-containing protein interacts with Pepino mosaic virus triple gene block protein 1. Int J Mol Sci. (2018) 19:3747. doi: 10.1094/MPMI-07-14-0195-R
30. Ruiz-Ramón F, Rodríguez-Sepúlveda P, Bretó P, Donaire L, Hernando Y, and Aranda MA. The tomato calcium-permeable channel 4.1 (SlOSCA4. 1) is a susceptibility factor for Pepino mosaic virus. Plant Biotechnol J. (2023) 21:2140–54. doi: 10.1111/pbi.14119
31. Méndez-López E, Donaire L, Gosálvez B, Díaz-Vivancos P, Sánchez-Pina MA, Tilsner J, et al. Tomato SlGSTU38 interacts with the PepMV coat protein and promotes viral infection. New Phytol. (2023) 238:332–48. doi: 10.1111/nph.18728
32. Hanssen IM, Peter van Esse H, Ballester AR, Hogewoning SW, Parra NO, Paeleman A, et al. Differential tomato transcriptomic responses induced by pepino mosaic virus isolates with differential aggressiveness. Plant Physiol. (2011) 156:301–18. doi: 10.1104/pp.111.173906
33. Ludman M, Burgyán J, and Fátyol K. Crispr/Cas9 mediated inactivation of Argonaute 2 reveals its differential involvement in antiviral responses. Sci Rep. (2017) 7:1010. doi: 10.1038/s41598-017-01050-6
34. Katsarou K, Mitta E, Bardani E, Oulas A, Dadami E, and Kalantidis K. DCL-suppressed Nicotiana benthamiana plants: valuable tools in research and biotechnology. Mol Plant Pathol. (2019) 20:432–46. doi: 10.1111/mpp.12761
35. Abou Haidar MG, Xu H, and Hefferon KL. Potexvirus isolation and RNA extraction. Methods Mol Biol. (1998) 81:131–43. doi: 10.1385/0-89603-385-6:131
36. Taylor SC, Nadeau K, Abbasi M, Lachance C, Nguyen M, and Fenrich J. The ultimate qPCR experiment: producing publication quality, reproducible data the first time. Trends Biotechnol. (2019) 37:761–74. doi: 10.1016/j.tibtech.2018.12.002
37. Atkinson PH and Matthews REF. On the origin of dark green tissue in tobacco leaves infected with tobacco mosaic virus. Virology. (1970) 40:344–56. doi: 10.1016/0042-6822(70)90411-3
38. Lindbo JA and Dougherty WG. Plant pathology and RNAi: a brief history. Ann Rev Phytopathol. (2005) 43:191–204. doi: 10.1146/annurev.phyto.43.040204.140228
39. Alcaide C and Aranda MA. Determinants of persistent patterns of Pepino mosaic virus mixed infections. Front Microbiol. (2021) 12:694492. doi: 10.3389/fmicb.2021.694492
40. Huss B, Walter B, and Fuchs M. Cross-protection between Arabis mosaic virus and Grapevine fan leaf virus isolates in Chenopodium quinoa. Ann Appl Biol. (1989) 114:45–60. doi: 10.1111/j.1744-7348.1989.tb06786.x
41. Mahmood T and Rush CM. Evidence of cross-protection between Beet soilborne mosaic virus and Beet necrotic yellow vein virus in sugar beet. Plant Dis. (1999) 83:521–26. doi: 10.1094/PDIS.1999.83.6.521
Keywords: cross-protection, RNA silencing, superinfection exclusion, potexviruses, pepino mosaic virus, plant virus control
Citation: Spanos T, Ghodbane A, Rezazga A, Ludman M, Fátyol K, Elbeaino T and Livieratos I (2025) Pre-inoculation of ago2 and DCL2/4-deficient Nicotiana benthamiana plants with the Pepino mosaic virus EU mild isolate confers complete protection against superinfection with the aggressive isolate. Front. Virol. 5:1584535. doi: 10.3389/fviro.2025.1584535
Received: 27 February 2025; Accepted: 27 May 2025;
Published: 19 June 2025.
Edited by:
Leonardo Velasco, Andalusian Institute for Research and Training in Agriculture, Fisheries, Food and Ecological Production (IFAPA), SpainReviewed by:
Adrian Alejandro Valli, Spanish National Research Council (CSIC), SpainDirk Janssen, IFAPA Centro La Mojonera, Spain
Copyright © 2025 Spanos, Ghodbane, Rezazga, Ludman, Fátyol, Elbeaino and Livieratos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ioannis Livieratos, bGl2aWVyYXRvc0BtYWljaC5ncg==