 Zhi-Wei Kang
Zhi-Wei Kang Fang-Hua Liu
Fang-Hua Liu Rui-Ping Pang1
Rui-Ping Pang1 Hong-Gang Tian
Hong-Gang Tian Tong-Xian Liu
Tong-Xian Liu- 1State Key Laboratory of Crop Stress Biology for the Arid Areas, Key Laboratory of Northwest Loess Plateau Crop Pest Management of Ministry of Agriculture, Northwest A&F University, Yangling, China
- 2Department of Entomology, University of Georgia, Athens, GA, United States
- 3State Key Laboratory of Integrated Management of Pest and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
The integrated pest management (IPM) strategy was developed and used in combination with pesticides and beneficial biological control agents. To further develop IPM efficiency, it is important to evaluate the side effects of pesticides on biological control agents. Aphidius gifuensis is one of the most important aphid natural enemies and has been successfully used to control Myzys persicae and other aphid species. Imidacloprid (IMD) is a popular pesticide used worldwide and is highly toxic to non-target arthropods. Here, we investigated the short-term sublethal toxicity of IMD in Aphidius gifuensis and its impact on the biological performance and gene expression of this parasitoid. We found that sublethal IMD doses had a significant negative effect on the life history traits of female A. gifuensis, including shortening the lifespan and lowering parasitic capacity. Moreover, exposure to sublethal IMD also adversely affected the response of A. gifuensis to aphid-infested plant volatiles. Based on the transcriptome analysis, we found that the exposure to sublethal IMD doses significantly affected expression of genes involved in the central nervous system, energy metabolism, olfactory, and detoxification system of A. gifuensis. RT-qPCR also revealed that short term expose to sublethal IMD doses significantly induced the gene expression of genes related to the central nervous system (nAChRa7, nAChRa9, TbH, OAR1, NFR, TYR, and DAR1), olfactory system (OR28 and IR8a1), and detoxification system (CYP49p3, CYP6a2, and POD), while it suppressed the expression of genes involved in the central nervous system (nAChRa4 and nAChRb1), olfactory system (Orco1, IR8a2, and GR1), and detoxification system (GST2). Furthermore, exposure to sublethal doses of IMD also significantly increased the activities of CarEs and POD, whereas we observed no influence on the activities of CAT, GST, and SOD. Our results indicate that sublethal IMD doses might adversely affect the biological performance of A. gifuensis by altering gene expression related to the function of olfactory, nervous, energy metabolism, and detoxification systems. Thus, how the use of pesticides directly affect insect population should be considered when used in conjunction with natural pest parasitoids in IPM strategies.
Introduction
Over the past decade, numerous pesticides have been developed and introduced into agriculture, forestry, horticulture, grain storage, and public/personal health. Around the world, more than 2 million tons of pesticides are used annually (De et al., 2014). About 24.3, 18.2, and 9.7 kg/ha of pesticides were used in 12 villages in six counties in Guangdong, Jiangxi, and Hebei provinces, in China (Zhang et al., 2015)w. The global pesticide cost is estimated to be $81.1 billion by 2021. However, the intensive use of pesticides has posed selective pressure on targeted pest species to develop pesticide resistance or pest resurgence (Desneux et al., 2007; Tabashnik et al., 2009). Over 500 species are resistant to at least one type of pesticide (De et al., 2014). For example, the diamondback moth (Plutella xylostella) has developed a resistance to over 91 pesticides, all within 3 years (2015–2017), Dysdercus koenigii has developed a very high resistance to acetamiprid (from 33 to 433-fold) and imidacloprid (from 21 to 173-fold) in Punjab, Pakistan (Zhang et al., 2016; Saeed et al., 2018). P. xylostella is also resistant to Bacillus thuringiensis and its derivatives. This higher resistance of pests lead to the development of novel pesticides and an increase in the quantity and frequency of pesticide application, which not only facilitates the resistance in the target pests but also results in environment contamination. In Thailand, the average pesticide residues found in surface water was 1.3757 ± 0.5014 mg/L (dicrotophos in summer) and 0.3629 ± 0.4338 mg/L (ethion in winter), and the average ethion residues in soil was 42.2893 ± 39.0711 mg/kg (summer), and 90 ± 24.16443 mg/kg (winter) (Harnpicharnchai et al., 2013). The persistent nature of pesticides has entered into various food chains and has bioaccumulated in higher trophic levels inlcuding bees, birds, and mammals (Bayen et al., 2005; Desneux et al., 2007; Kapoor et al., 2011; Dicks, 2013). Thus, to some extent, the adverse effects of pesticides have outweighed the benefits associated with their use.
To minimize chemical pesticides use, various candidate biological control agents have been evaluated, such as the application of trap crop systems, and entomopathogenic fungi, bacteria, predators, and parasitoids (Shah and Pell, 2003; Shelton and Badenes-Perez, 2006; Yang et al., 2009; Walker et al., 2017). For example, blue fluorescent light is widely used in rice paddy fields to control the rice stem borer, Chilo suppressalis Walker, and Tryporyza incertulas Walker moths (Ishikura, 1950). Alfalfa and mungbean are used as a trap crop in cotton fields for managing lygus bugs, Lygus Hesperus, and the mirid Apolygus lucorum, respectively (Godfrey and Leigh, 1994; Lu et al., 2009). Two parasitic wasps Trichogrammatoidea bactrae fumata Nagaraja and Trichogrammatoidea cojuangcoi Nagaraja are successfully applied to control the cocoa pod borer, Conopomorpha cramerella Snellen, in the field (Lim and Chong, 1987; Alias et al., 2005). However, these biological control agents such as trap crop systems and commercial inundated releases of parasitoids and predators may not be capable of reducing pest densities to levels that avoid economic losses in a timely manner (Yang et al., 2011). Thus, the proper amalgamation of various control techniques into a unified system may provide a powerful tool to keep pest population levels low and to avoid economic damage. However, amalgamation of various pest control techniques also poses a major challenge: how do we take advantage of each biological technique?
The pesticides that are used in pest management programs must be effective in controlling pests and have a low impact on non-target organisms, such as natural enemies (Desneux et al., 2007; Lu et al., 2009). To determine the residual period of control for an insecticide, is essential to plan insect management strategies, which will influence the spraying frequency and the release time of natural enemies, and in turn, affect the pest control cost. Thus, the residual and toxic effects of pesticides are the most serious bottlenecks in the successful use of pesticides and natural enemies.
Aphids are key insect pests that are responsible for major agricultural losses, particularly because they are vectors of various plant viruses (Van Emden and Harrington, 2017). In the Australian grain industry alone, aphid-related plant injuries, either through direct feeding or virus transfer, represent a potential economic cost of $200–480 million/year (Murray et al., 2013; Valenzuela and Hoffmann, 2015). Current management strategies for broadacre aphids rely primarily on pesticides, either through seed dressings or foliar applications (Dedryver et al., 2010; Chollet et al., 2014). However, due to the strong adaptation and fecundity of aphids, they have developed strong resistance to various pesticides. For example, the green peach aphid (Myzus persicae) is resistant to more than 70 different types of synthetic insecticides (Silva et al., 2012).
Imidacloprid (IMD) is one of the most extensively used pesticides in the world (Li et al., 2018). It is sprayed directly onto plants or used as a seed or soil treatment on a number of agricultural products to control a variety of insect pests including plant- and leafhoppers, aphids, termites, whiteflies, and thrips (Li et al., 2018). However, IMD is highly persistent and toxic to non-target animals, including bees (Dicks, 2013). When a bumblebee (Bombus terrestris) colony was treated with IMD at a sublethal concentration, it significantly reduced the growth rate and production of queens and workers (Laycock et al., 2012; Whitehorn et al., 2012). In addition, there was a significant decrease in the fecundity of Orius insidiosus, Orius tristicolor, Hippodamia convergens, and Chrysoperla carnea, which are natural enemies of aphids, when treated with sublethal concentrations of IMD (Mizell and Sconyers, 1992; Sclar et al., 1998; Studebaker and Kring, 2003; Rogers et al., 2007; Funderburk et al., 2013).
Aphidius gifuensis Ashmead (Hymenoptera: Braconidae) is one of the most widely distributed and dominant endoparasitoid of pest aphids, including M. persicae and Sitobion avenae (Fabricius), and are successfully applied in greenhouses for controlling vegetable aphids and in fields for tobacco aphid (M. persicae, also known as green peach aphid) management in China (Yang et al., 2009, 2011; Ali et al., 2016; Kang et al., 2017a; Yang F. et al., 2017). Furthermore, Yang et al. (2009) reported that after augmentative releases of A. gifuensis, the frequency and quantity of pesticide application could be sustained at a low level for 8 years. However, A. gifuensis is sensitive to various agrochemicals (Ohta and Takeda, 2015). For example, after 14 days of exposure to residual permethrin and IMD, also showed high toxicities to A. gifuensis (Kobori and Amano, 2004). In this work, we not only evaluated the toxicity of IMD in A. gifuensis, but also investigated the biological performance of A. gifuensis exposed to sublethal doses of IMD. We hypothesized that sublethal doses of IMD would disrupt parasitoids performance through regulating some genes on the molecular level. Transcriptome technology was applied to explore which of the parasitoid genes could be modulated by IMD and how A. gifuensis adjusts its detoxification system to respond to the exposure of IMD.
Materials and Methods
Insect Species
Aphidius gifuensis used in this work were maintained on M. persicae, which was reared on chili pepper (Capsicum annuum L., var. “Lingxiudajiao F1”) at 25 ± 1°C with a 16 h light: 8 h dark photoperiod and a relative humidity of 60 ± 5% in an air-conditioned insectary.
Performance of A. gifuensis Exposure to IMD
We used three different dilution magnifications to evaluate the toxicity of IMD on A. gifuensis, and distilled water was used as a control. Five plastic vials (length: 8 cm; diameter: 4 cm) were treated with 1 ml IMD or H2O. The IMD was swirled inside the vials for 30 s and allowed to air-dry in a hood to simulate the pesticide residues. At the time of exposure, twenty 2-day old A. gifuensis female adults were introduced into a vial. Twenty-four hours later, the mortality of A. gifuensis was counted and living parasitoids (at least 15) were individually collected to test the effects of IMD on the parasitism, longevity and sex ratio of offspring as described by Kang et al. (2017a) with little modification. Chili pepper plant with 200 second- or third- instar M. persicae were placed into a plastic cage (diameter: 13 cm; height: 30 cm) with screen mesh caps. Then, five females of A. gifuensis from different treatments (control (CK) or IMD), were introduced into each rearing cage for 8 h. After the parasitism, the aphids and Chili pepper complex was maintained in an incubator. Ten days later, the number of mummified aphids and the sex ratio of all wasps emerging from these mummified aphids were recorded. Five biological replicates were conducted in this work.
To analyze the effect of sublethal doses of IMD on the orientation behavior and gene expression of A. gifuensis, the LC20 of IMD was used and 24 h later, surviving parasitoids were collected and separated into two groups: one group with thirty living parasitoids was flash-frozen in liquid nitrogen and stored at −80°C for the gene expression analysis; the remaining parasitoids were placed into a PCR tube for orientation behavior. Y-tube olfactometers were used to assess the oriented responses of A. gifuensis toward healthy and aphid-infested plants. Y-tube was conducted as described by Kang et al. (2018a,b). In total, 100 living parasitoids were tested for the orientation behavior.
RNA Sequencing
Total RNA was extracted from whole bodies of five female A. gifuensis using RNAiso Plus (Takara Bio, Tokyo, Japan), following the manufacturer's instructions. The high quality RNA was used for the further cDNA synthesis and Illumina library generation, which was completed at the Novogene Bioinformatics Technology Co., Ltd. (Beijing, China).
De novo Assembly and Gene Annotation
Transcriptome de novo assembly was conducted using a short read assembling program—Trinity with min_kmer_cov set to 2 by default and all other parameters set to default (Grabherr et al., 2011). In order to get comprehensive information about the genes, we aligned the unigenes larger than 150 bp to nr, Nt, KEGG, Swiss-Prot, and COG databases, with e-value < 10−5. With nr annotation, we used the Blast2GO program to get GO annotation of unigenes (Conesa et al., 2005). The WEGO software was used next to perform GO functional classification for all unigenes (Ye et al., 2006). The unigene expression levels were calculated by fragments per kb per million reads (FPKM) method, using the formula, FPKM (A) = 103 (106 C)/NL (A: Unigene A; C: number of fragments that uniquely aligned to Unigene A; N: the total number of fragments that uniquely aligned to all Unigenes; L: the base number in the CDS of Unigene A). The FPKM method eliminates the influence of different gene lengths and sequencing levels on the calculation of gene expression; therefore, the calculated gene expression can be directly used for comparing differences in gene expression across samples.
Expression Analysis
Heat map analysis was performed by the R package of pheatmap (http://www.r-project.org/; R Foundation for Statistical Computing, Wien, Austria). Heatmap plots present the binary log of fold-change of IMD/CK for each gene with a three-color scale (navy, white and firebrick).
RT-qPCR was performed to validate the expression of several genes in A. gifuensis. Total RNA was extracted from five whole bodies of 2-day old female A. gifuensis, and cDNA was then synthesized from 1 μg total RNA using a PrimeScript® RT reagent Kit with gDNA Eraser (perfect Real Time) (Takara, Tokyo, Japan) according to the manufacturer's protocol. Specific gene primers were designed by Primer Premier 6 (PREMIER Biosoft International, Palo Alto, CA, USA), which are presented in Table S1. In total, three biological replicates, with three technical replicates were conducted, and the qPCR was performed as previously described (Kang et al., 2017b). However, in this study, we used a 2−ΔCt method to evaluate the expression of selected genes (Eakteiman et al., 2018).
Enzyme Activity Assay
The activities of CarE, SOD, CAT, POD, and GST were measured using commercially available assay kits (Nanjing Jiancheng Bioengineering Institute, Jiangsu, China) as described previously (Kang et al., 2017a).
Data Analyses
The comparison of performance parameters was subjected to a one-way analysis of variance (ANOVA) followed by the separation of means by the Fisher's protected least significant difference (LSD) test at P = 0.05. The gene expression profiles were determined by a student's t-test at P < 0.05. The orientation behavior of A. gifuensis under the different treatments was separated by the Chi-square test (P < 0.05). A generalized linear mixed-effects model (GLMM) with a binomial family with the cbind function was then performed to analyze the response to the treatment: yes or no. Except for GLMM, SPSS 22.0 (SPSS Inc., Chicago, IL, USA) was used for the data analyses. GLMMs were performed in the R programming environment (version 3.5.1).
Results
Exposure of Sublethal Does of IMD Impaired the Performance of A. gifuensis
The influence of IMD on the mortality, parasitism, longevity and female proportion in offspring of A. gifuensis are shown in Table 1. Exposure to IMD significantly increased the mortality of female and male adults, and decreased the longevities of surviving female and male adults, as well as the parasitism of surviving female adults (Mortality: Female: F = 152.071, P < 0.001; Male: F = 62.448, P < 0.001; Longevity: Female: F = 27.952, P < 0.001; Male: F = 26.069, P < 0.001; Parasitism: F = 19.991, P < 0.001). However, exposure to IMD did not influence the female proportion of offspring produced by surviving female adults, compared to healthy female adults (F = 0.725, P < 0.504). Furthermore, IMD significantly reduced the sensitivity of A. gifuensis to the volatiles from aphid infested plants (Healthy wasps: χ2 = 20.045, P < 0.001; IMD treated wasps: χ2 = 0.636, P = 0.425, Figure 1). The GLMM analysis also revealed that IMD changed the response of A. gifuensis to these volatiles (P = 0.009).
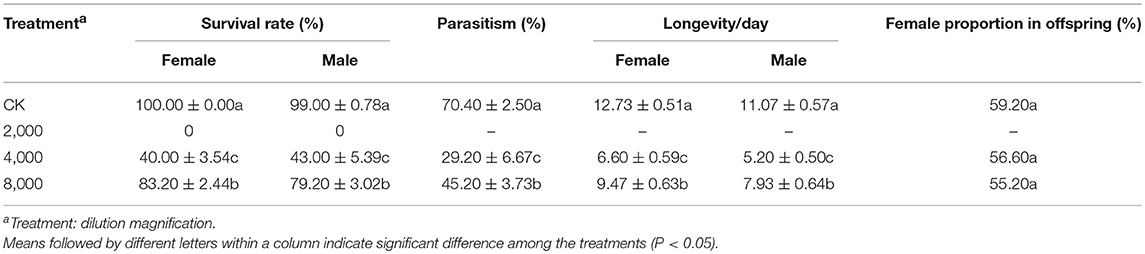
Table 1. The side effects of IMD on the parasitism, longevity, and female proportion in offspring of A. gifuensis.
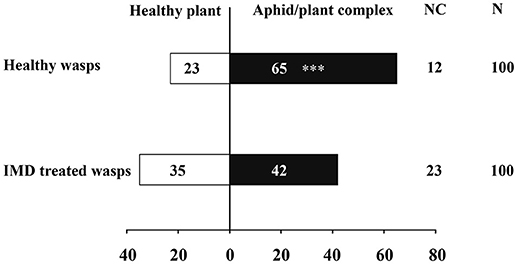
Figure 1. Effects of IMD exposure on the orientation behavior of aphid endoparasitoid, Aphidius gifuensis on aphid infested and control plants. Asterisk over the bars designate a significant difference based on GLMM and Chi-square test: *P < 0.05, **P < 0.01, ***P < 0.001. N = 100.
An Overview of the Transcriptome
The transcriptome assembly was performed using the Trinity program, with an optimal K-mer length set to 25. A total of 48,033,980 and 53,409,010 raw reads were obtained from CK and IMD treatment groups, respectively. After removing adaptor sequences, low quality sequences and N-containing sequences, 46,760,944, and 51,668,492 clean reads were generated form the CK and IMD raw data, respectively. The assemblies produced 81,727 transcripts with a maximum sequence length of 19,224 bp and a N50 transcript length of 1,284 bp (Table 2). Furthermore, the GC content of the CK and IMD treatment groups were 31.47 and 30.76%, respectively. The quality of RNA samples and the expression file of genes were supplied as Table S2 and Datasheet 1.
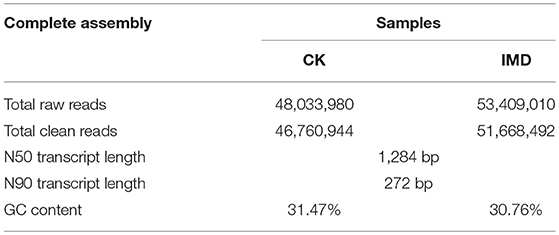
Table 2. Assembly summary of the A. gifuensis transcriptome.
Functional Gene Annotation and Classification
GO enrichment indicated that genes involved in the cellular process, metabolic process, single-organism process, biological regulation and the regulation of the biological process in the category of the biological process, cell, cell part, membrane and organelle in the category of cellular component, and binding and catalytic activity in the category of molecular function were dominant (Figure 2). The neuroactive ligand-receptor interaction, cAMP signaling pathway and MAPK signaling pathway were the major enrichment pathways in the IMD treatment group (Figure 3).
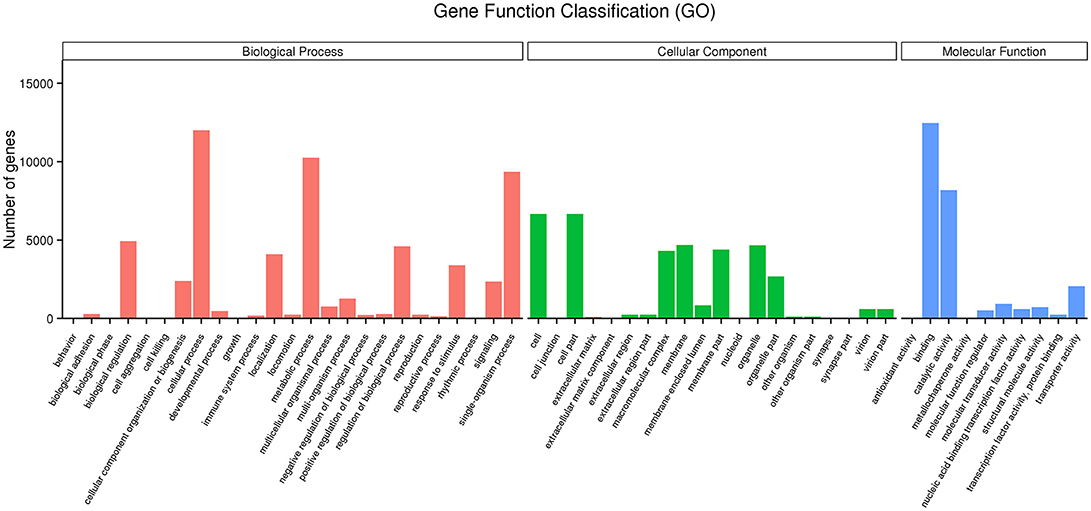
Figure 2. Functional annotation of Aphidius gifuensis transcripts based on gene ontology (GO) categorization.
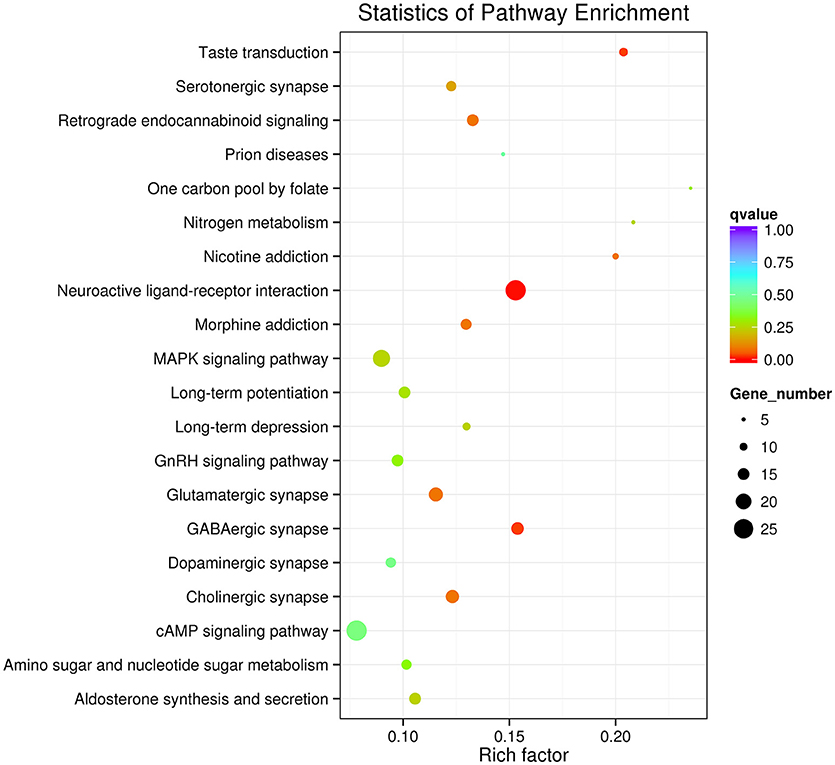
Figure 3. Top 20 enriched Kyoto Encyclopedia of Genes and Genomics (KEGG) pathways of Aphidius gifuensis after the exposure of sublethal does of IMD.
Genes Involved in the Central Nervous and Olfactory Systems Are Differentially Expressed in Response to Sublethal Doses of IMD
The first gene groups we examined focused on the central nervous and olfactory systems, which are the target of the IMD and influence target insect behavior. For the target of the IMD, we identified 15 acetylcholine receptors: 11 neuronal acetylcholine receptors and four muscarinic acetylcholine receptors (Table 3). Among these target genes, nAChRα4 and nAChRβ1 were significantly down-regulated in response to the IMD. Only nAChRα7 and nAChRα9 exhibited higher transcript abundances in the IMD treated A. gifuensis when compared to CK samples. Furthermore, no significant differences were detected in the rest of the nAChRs genes.
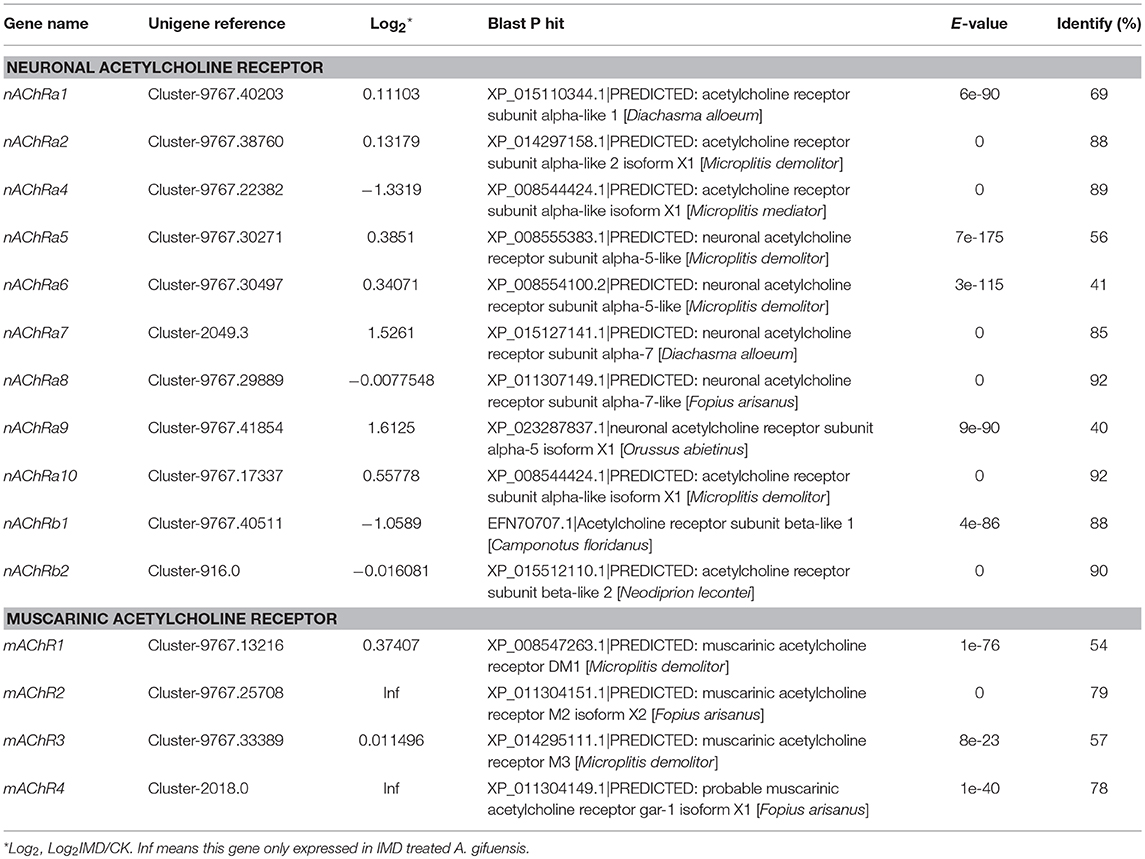
Table 3. The identified acetylcholine receptors in A. gifuensis.
Apart from the potential target genes, we also analyzed the impact of IMD exposure on olfactory systems to explain the impaired orientation behavior we observed in A. gifuensis treated with IMD. We found that a very low proportion of olfactory related genes exhibited significant differences between the treatments and control samples (Figure 4). The decrease in the mean FPKM values for the odorant co-receptor (Cluster-8038.0), odorant receptors (Cluster-1578.0, Cluster-4221.1, and Cluster-3108.1), chemosensory protein (Cluster-8527.0), gustatory receptors (Cluster-7667.0 and Cluster-4878.0), and ionotropic receptors (Cluster-9767.24401 and Cluster-1662.0) was particularly striking. On the contrary, exposure to IMD significantly up-regulated the expression of the odorant-binding protein (Cluster-1704.0), odorant receptor (Cluster-3211.0), gustatory receptor (Cluster-9767.3034), and the ionotropic receptor (Cluster-6117.0), which were effected the most by IMD treatment in their gene group.
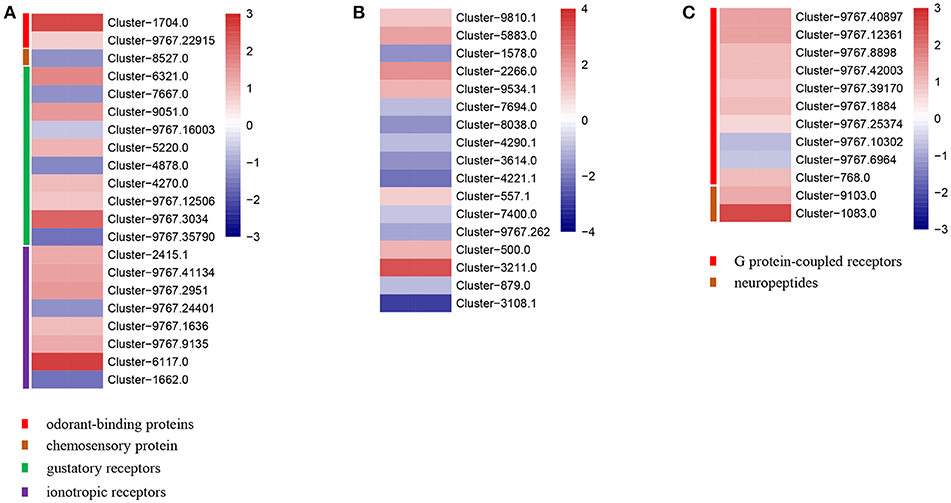
Figure 4. Heatmaps of expressions for genes with neuron and odorant related functions. (A) Genes identified as odorant-binding proteins, chemosesory proteins, gustatory receptors, and ionotropic receptors. (B) Genes identified as odorant receptors. (C) Genes involved in neuron functions.
Furthermore, exposure to IMD also influenced the expression of genes involved in the central neurons. The dopamine receptor 1 (Cluster-9767.1884), tryptophan 5-hydroxylase (Cluster-1083.0), neuropeptide FF receptor (Cluster-9767.40897) were significantly higher in IMD treatments compared to CK treatments.
Effects of Sublethal Doses of IMD on Detoxification Progress, Antioxidant System, and Biomolecule Damage Genes in A. gifuensis
We found that defense genes, such as cytochrome P450 (CYP4c1: Cluster-5030.0 and Cluster-9767.42126; CYP6a2: Cluster-9767.4090; CYP9p3: Cluster-9767.18925), cyt b5 (Cluster-6200.1), peroxidase (POD: Cluster-9767.17490), carboxylesterase (CarE, Cluster-9767.29708), glutathione S-transferaes (GSTs, Cluster-9767.30914), and heat shock proteins (HSPs, Cluster-9767.16364, and Cluster-9767.39176), were highly expressed in the IMD treated A. gifuensis (Figure 5), while three P450s (Cluster-9767.38298, Cluster-9767.30384, and Cluster-9767.36002), POD (Cluster-9767.24511), and HSP (Cluster-9767.32708) exhibited lower transcript abundances in the IMD treated A. gifuensis than in the CK group (Figure 5).
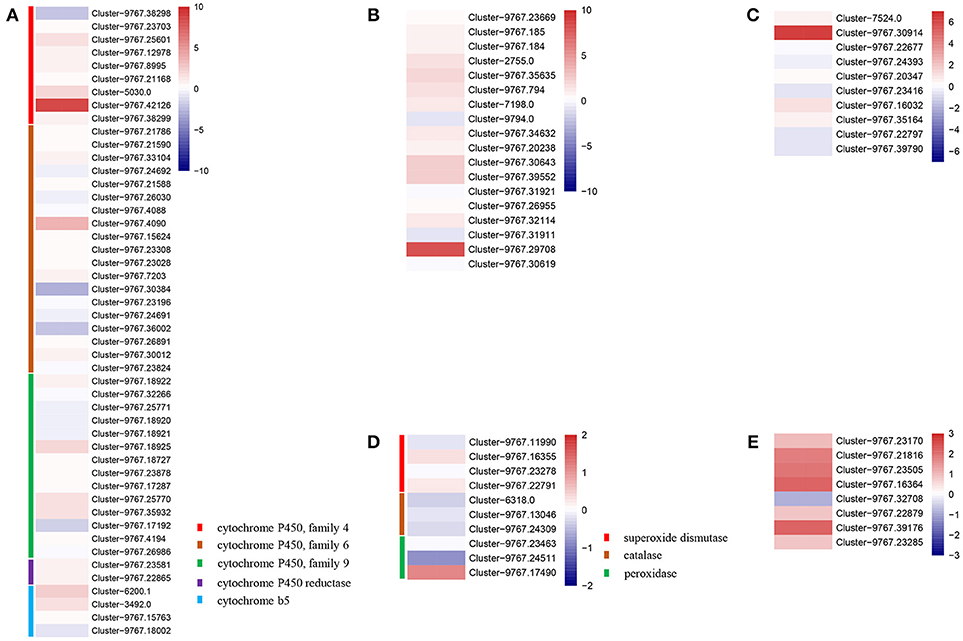
Figure 5. Heatmaps of expressions for genes with detoxification and stress response. (A) Genes involved in cytochrome P450. (B) Genes identified as carboxylesterase. (C) Genes identified as glutathione S-transferees. (D) Genes involved in antioxidant enzymes. (E) Gene identified as heat-shock proteins.
Sublethal Doses of IMD Altered Expression of Genes Involved in Metabolic Signaling
To investigate the impact of IMD on energy metabolism, we analyzed the expression of genes involved in fatty acid, sugar, and amino acid metabolism (Figure 6). We found that almost all the genes involved in fatty acid metabolism were expressed at a higher level in the IMD treated A. gifuensis (Figure 6A), while only Cluster-9767.37118 and Cluster-6642.1 were expressed at a lower level of the IMD treatment. Consistent with fatty acid metabolism, the majority of genes that regulate sugar and amino acid metabolism, also exhibited higher mean FPKM values in the IMD treated A. gifuensis, whereas the expression of Cluster-9767.30238, Cluster-9767.39562, Cluster-8931.0, and Cluster-9767.35416 in sugar metabolism and Cluster-2788.0, Cluster-6624.0, Cluster-6867.0, Cluster-9767.27231, Cluster-9767.29443, Cluster-9767.39986, and Cluster-9767.5013 in amino acid metabolism, were down-regulated in response to the IMD treatment (Figures 6B,C).
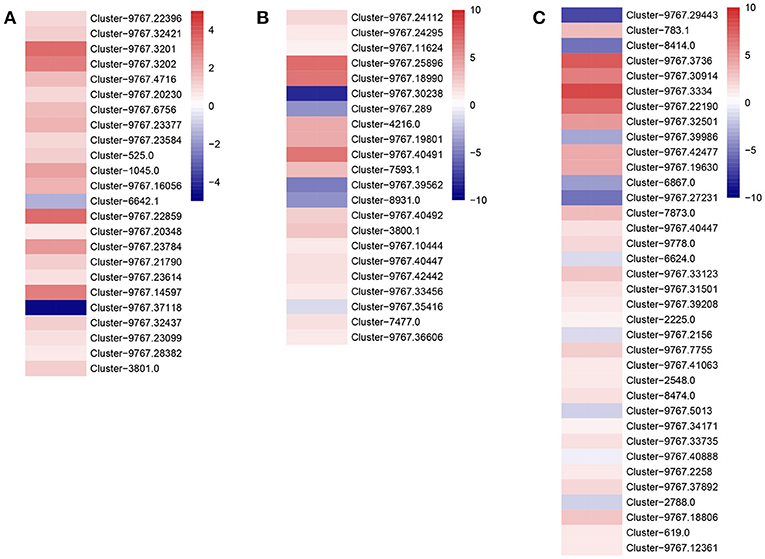
Figure 6. Heatmaps of expressions for genes with energy metabolic signaling. (A) Genes with functions in fatty acid metabolism. (B) Genes with functions in sugar metabolism. (C) Genes with functions in amino acid metabolism.
Validation of Transcriptome Data by qPCR
To confirm the transcriptome data, we conducted the RT-qPCR of several genes identified in the transcriptome that were IMD-sensitive. Exposure to IMD significantly increased the expression of CYP6a2, CYP9P3, POD, OR28, IR8a1, nAChRa7, nAChRa9, TbH, OAR1, NFR, TYR, and DAR1, whereas the expression of GST2, nAChRa4, nAChRb1, ORco, IR8a2, and GR1 decreased (Figure 7). Furthermore, exposure to IMD did not influence the expression of GST5, SOD1, and SOD2 (Figure S1).
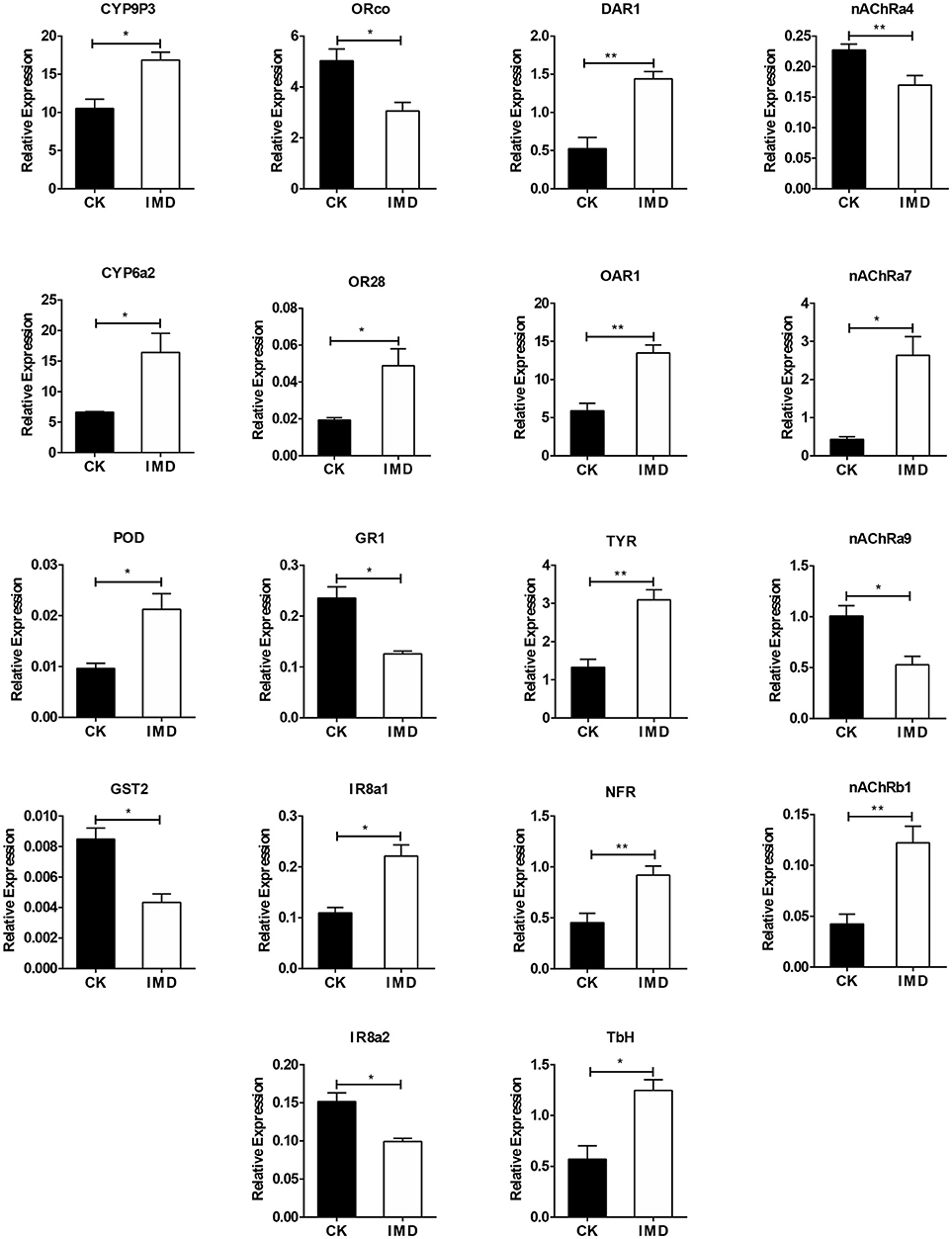
Figure 7. Validation of expression of selected genes using qPCR. DAR1, dopamine receptor 1; SNF, short neuropeptide F; NFR, neuropeptide FF receptor 2; TYR, putative tyramine receptor 2; TbH, tyramine beta-hydroxylase. Asterisk over the bars designate a significant difference based on student's t-test: *P < 0.05, **P < 0.01, ***P < 0.001, and the error bars is ± SE bars. N = 3.
Activities of CarEs, POD, and GSTs in A. gifuensis After IMD Exposure
Exposure to IMD significantly induced the activities of POD and CarEs, while it had no significant influence on SOD, CAT and GST activity (POD: t = −11.648, df = 4, P < 0.001; CarE: t = −10.552, df = 4, P = 0.003; SOD: t = 0.843, df = 4, P = 0.4467; CAT: t = 0.6523, df = 4, P = 0.2298; GST: t = 1.886, df = 4, P = 0.1323; Figure 8 and Figure S2).
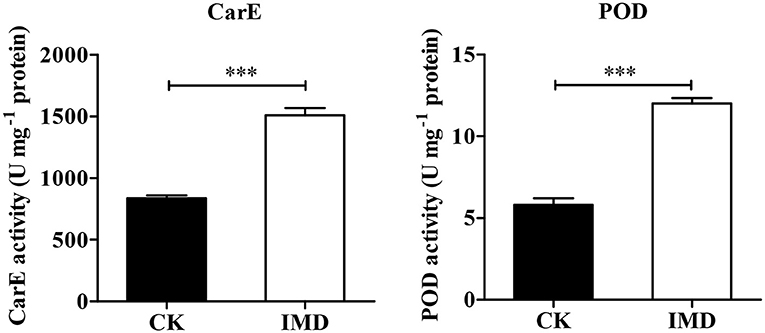
Figure 8. Activities of CarE and POD. Asterisk over the bars designate a significant difference based on student's t-test: *P < 0.05, **P < 0.01, ***P < 0.001, and the error bars is ± SE bars. N = 3.
Discussion
IPM program improvements requires an understanding of how pesticides influence natural enemies of the pests that are being targeted. Therefore, the effects of sublethal doses of pesticides are important for improving IPM programs. In this work, we found that oral ingestion of sublethal doses of IMD, adversely affected parasitoid performance, including the survival rate, parasitic capacity, and longevity of female adults, which was consistent with the performance of the Aphidius colemani, Microplitis mediator, O. insidiosus, C. flavipes, and Trichogramma species exposed to pesticides (D'Avila et al., 2018; Fontes et al., 2018). In the M. mediator, exposure to flonicamid, pymetrozine, spinosad, and thiacloprid reduced its parasitization activity, percentage of parasitism and female longevity. In addition, IMD impaired the longevity and parasitic capacity of Trichogramma species including Trichogramma achaeae, T. chilonis, T. platneri, and T. pretiosum (Khan and Ruberson, 2017; Fontes et al., 2018). The exposure to pesticides also adversely affected the biocontrol efficiency of pest predators (Moscardini et al., 2013; Nawaz et al., 2017). For example, IMD significantly repressed egg hatching, nymph survival and adult fecundity of the predatory bug, Orius albidipennis (Sabahi et al., 2010; Moscardini et al., 2013). Similarly, sublethal doses of diazinon, fenitrothion, and chlorpyrifos exhibited adverse effects on the biological performance of Andrallus spinidens, which is a predator of rice lepidopterous larvae (Gholamzadeh-Chitgar et al., 2015). Similar to chemical pesticides, other biological agents like entomopathogenic fungi and bacteria also adversely affect the biological performance of parasitoids and predators (Potrich et al., 2017). In addition, a high occurrence of wing deformities was observed when mummies of A. gifuensis were exposed to IMD (44.44%), acetamiprid (67.25%), and thiamethoxam (33.33%) (Sun et al., 2014). All of these results indicated that pesticide exposure adversely influenced the performance of natural enemies, which also means that the effectiveness of natural enemies can be reduced by the application of pesticide.
In addition to biological performance, we also investigated the side effects of IMD on the orientation behaviors of A. gifuensis after IMD treatment. We found that exposure of IMD significantly reduced the sensitivity of A. gifuensis to the volatiles produced by aphid infested plants. Consistent with this results, consuming IMD or aldicarb contaminated floral nectar, also reduced the response of Microplitis croceipes to the odors from its host Helicoverpa zea infested cotton (Stapel et al., 2000). In Anagrus nilaparvatae, survivors of IMD exposure had no response to volatiles from Nilaparvata lugens-infested rice (Liu et al., 2010). In addition, exposure to pyrethroids impaired the host-searching and oviposition behavior of the aphid parasitoids Aphidius ervi and Aphidius colemani, and Trissolcus basalis, which is an egg parasitic wasp of the southern green stinkbug, Nezara viridula (Ahmad and Hodgson, 1998; Salerno et al., 2002; Desneux et al., 2004a). Furthermore, Wang D. et al. (2017) found that exposure to beta-cypermethrin significantly decreased pheromone perception in male Trichogramma chilonis. All of these results indicate that IMD exposure impairs or reduces the sensitivity of the A. gifuensis olfactory system, thereby disrupting host searching and parasitizing.
To explore the potential mechanism of the negative effects of IMD on A. gifuensis, transcriptome technology was used to comprehensively analyze the gene expression of A. gifuensis in response to sublethal doses of IMD exposure. Our transcriptomic analysis pointed to a profound regulation of genes principally related to the olfactory and neuronal systems. The most down-regulated genes were the odorant co-receptor (Cluster-8038.0), which is the most important odorant receptor in the detection of odorants; gustatory receptor 1 (Cluster-7667.0), a sugar receptor that is associated with host aphid discrimination; and the neuropeptides capa receptor (Cluster-9767.10302), a G protein-coupled receptor for the capa peptides and an important signaling molecule in the regulation of a wide range of physiological processes (Kang et al., 2017b; Schoofs et al., 2017). These results are generally consistent with recent studies of the interaction of neonicotinoid with OBPs and CSPs. For example, CSP3 and OBP21 were downregulated in honey bees exposed to thiamethoxam (Shi et al., 2017). Furthermore, a sublethal dose of IMD inhibited the binding affinity of OBP2 and CSP1 to a floral volatile β-ionone in Apis cerana and GOBP2 to a tea volatile E-2-hexenal in Agrotis ipsilon (Li et al., 2015, 2017a,b). Interestingly, in addition to these down-regulated genes, a considerable number of genes were up-regulated, such as the odorant-binding protein (Cluster-1704.0), gustatory receptor (Cluster-9767.3034), ionotropic receptor (Cluster-6117.0), and odorant receptor (Cluster-3211.0). Similarly, a single brief exposure to pesticides dramatically increased CSP expression in Bombyx mori (abamectin) and Bemisia tabaci (thiamethoxam) (Xuan et al., 2015; Liu G. et al., 2016). All of these results indicate that the impairment of olfactory systems from sublethal doses of some pesticides could be involved the disordered orientation behavior.
As a neurotoxin and agonist of nAChRs, IMD had high binding affinity for nAChRs, thereby disrupting neurotransmission (Ffrench-Constant et al., 2016). IMD was thought to impede information receiving and processing in N. vitripennis, which led to the disruption of sexual communication and foraging behavior (Cook et al., 2016; Tappert et al., 2017). Furthermore, in Solenopsis invicta, when treated with 0.25 μg/ml IMD, there was a significant reduction in food consumption, digging and foraging behavior, while the neurotoxins flubendiamide and indoxacarb increased the walking time of Copidosoma truncatellum (Wang L. et al., 2015; Ramos et al., 2018). In the current work, we found that nAChRa4 and nAChRb1 were down-regulated, which was consistent with the expression of nAChRs in Rhopalosiphum padi (Wang K. et al., 2017). Similar to the findings of Desneux et al. (2004b) in A. ervi, we also found that some of the IMD exposed females bended their abdomen forward as they were attacking aphid, while no aphids were present. All of these results revealed that sublethal doses of IMD not only impaired the olfactory system of A. gifuensis, but also disrupted the neurotransmission that influences their behavior. Furthermore, these results also indicate that the development of specific and environmentally safe pesticides, that present little or no harm to natural enemies of pest insects, are needed.
In addition to the effect IMD had on the olfactory and neuron systems, we also investigated the impact of IMD on the detoxification systems in A. gifuensis. Cytochrome P450 monooxygenases (P450s), carboxyl esterases (CarEs), and glutathione S-transferees (GSTs) are three major multiline enzyme families that are responsible for xenobiotic metabolism in most insect species (Li et al., 2007; Hsu et al., 2012; Chaimanee et al., 2016; Gong and Diao, 2017; Magesh et al., 2017; Traverso et al., 2017).
P450s are a group of important stress response-related genes that play significant roles in several physiological processes, including hormone metabolism, the adaptation to natural and synthetic toxins, and insecticide detoxification. As we know, overexpression of the gene coding of the P450 clades (CYP4, CYP6, and CYP9), contributes considerably to insecticide-resistance (Li et al., 2007; Bass et al., 2014). For example, in B. tabaci and M. persicae, over-expression of the cytochrome P450 genes CYP6CM1 and CYP6CY3, contribute to neonicotinoid insecticide resistance, as these enzymes can catalyze a more rapid conversion of imidacloprid to its less active form, 5-hydroxy-imidacloprid (Karunker et al., 2008; Puinean et al., 2010). Furthermore, CYP6AY1 and CYP6ER1 were highly overexpressed in the IMD resistant strain of N. lugens compared to the susceptible strain (Yang Y. X. et al., 2017). In A. mellifera, coumaphos and IMD treatment significantly decreased the expression of CYP306A1, CYP4G11, and CYP6AS14, whereas pyrethroid bifenthrin induced the expression of CYP9Q1 and CYP9Q2 but repressed the expression of CYP9Q3 (Mao et al., 2011; Chaimanee et al., 2016). In-vitro, CYP9Q1, CYP9Q2, and CYP9Q3 detoxify coumaphos independently and tau-fluvalinate with the cooperation of CarEs (Mao et al., 2011). Additionaly, CYP9Q1 and CYP9Q3 also contributed to the metabolism of quercetin (Mao et al., 2011). In this work, the most up-regulated genes were CYP4c1 and CYP6a2 (Cluster-9767.42126), which are associated with the IMD resistance in N. lugens; and cyt b5 (Cluster-6200.1), which is the electron transfer partners of P450 proteins and which modify the catalytic activity of P450 proteins (Paine et al., 2005; Ding et al., 2013).
CarEs are involved in the metabolic detoxification of dietary and environmental xenobiotics in insects (Xie et al., 2017; Wu et al., 2018). A higher expression or activity of CarEs have been reported in the insecticide resistance strains of many insect species such as M. persicae, R. padi, Aphis gossypii, Pediculus humanus capitis, P. xylostella, and Bactrocera dorsalis (Hsu et al., 2012; Gong et al., 2013, 2014; Kwon et al., 2014; Wang L. et al., 2015; Wang L. L. et al., 2015; Xie et al., 2017). In A. mellifera, the induction of CarE activity by IMD, acetamiprid, pymetrozine, and pyridalyl was observed, while malathion and permethrin significantly inhibited CarE activity (Yu et al., 1984; Suh and Shim, 1988; Badawy et al., 2014; Li Z. et al., 2017). Furthermore, in P. xylostella, CarEs activity was positively correlated with resistance to spinosyn, beta-cypermethrin, chlorpyrifos, and abamectin (Gong et al., 2013). Moreover, RNA interference-mediated gene silencing (RNAi) tests revealed that the knock-down of CarE genes led to a decreased tolerance to some pesticides (Wang L. L. et al., 2015). In B. dorsalis, the knock-down of CarE4 and CarE6 significantly decreased the resistance to malathion, and the detoxification of malathion was observed when CarE4 and CarE6 were heterologously expressed (Wang L. L. et al., 2015). Furthermore, in Lygus lineolaris, IMD exposure significantly increased the expression of 13 esterase genes (Zhu and Luttrell, 2015). In this work, we found that the expression and enzyme activity of CarEs in IMD treated A. gifuensis were significantly higher than that in CK treatment, especially carboxylesterase (Cluster-9767.29708).
GSTs are part of another important detoxification enzyme family. GST activity in larvae, pupae, and nurse bees, but not in foragers, was induced by pyrethroid flumethrin (Nielsen et al., 2000). In the eastern honey bee Apis cerana cerana, the sigma-class AccGSTS1 was up-regulated by phoxim, cyhalothrin and acaricide and the theta-class GST gene GSTT1 and omega-class GST gene GSTO2 was induced by cyhalothrin, phoxim, pyridaben, and paraquat, indicating that they might be involved in the stress response to pesticides (Yan et al., 2013; Zhang et al., 2013; Liu S. et al., 2016). Furthermore, formetanate increased the activity of GST, whereas IMD and dimethoate had no influence on GST activity in A. mellifera (Li Z. et al., 2017; Staron et al., 2017). However, in this work, only one GST (Cluster-9767.30914) was found to be highly expressed in the IMD treated A. gifuensis compared to the CK treatment, while the rest of the GSTs did not show any response to sublethal doses of IMD treatment. Similarly, in Lygus lineolaris, only four of the 19 GSTs were significantly down-regulated after IMD exposure, while the rest of these genes did not show any detectable difference in expression (Zhu and Luttrell, 2015). All of these results indicate that GST might not be responsible for IMD resistance in A. gifuensis.
In addition to these three major detoxification pathways, other interrelated pathways might also contribute to the response of xenobiotics, such as superoxide dismutase (SOD), catalase, POD, and HSPs (Chaimanee et al., 2016). In honeybee queens, exposure to IMD and coumaphos significantly depressed the expression of SOD and thioredoxin peroxidase (Chaimanee et al., 2016). Conversely, the expression of catalase, SOD and thioredoxin peroxidase was significantly increased in worker bees (Chaimanee et al., 2016). In this work, POD (Cluster-9767.17490) and HSPs (Cluster-9767.16364 and Cluster-9767.39176) were up-regulated in IMD treated A. gifuensis, which is consistent with the HSP expression profiles in beta-cypermethrin treated R. padi (Li Y. T. et al., 2017). Further, exposure to IMD significantly decreased the expression of GSTs. Together, these results indicate that detoxification and stress response systems are critical for protecting A. gifuensis from IMD damage.
To support or drive detoxification processes, the increased energy production through the up-regulation of enzymes involved in ATP synthesis, sugar metabolism, fatty acid metabolism, glycolysis, and the tricarboxylic acid (TCA) cycle were investigated. Our transcriptome data revealed that IMD treatment altered the expression of genes in energy-producing metabolic pathways such as fatty acid metabolism and sugar metabolism. Consistent with this finding, exposure to neonicotinoid also led to increased energy usage in honey bees (du Rand et al., 2017). Furthermore, exposure to a sublethal dose of beta-cypermethrin, led to an increase in respiratory quotient and respiratory rates in Harmonia axyridis, which is often coupled with the status of energy metabolism (Xiao et al., 2017). All of these results suggest that insects increase their energetic cost when undergoing detoxification after pesticide exposure. The increase in their energetic cost might result in the decrease of longevity and parasitism.
With the wide use of pesticides in agriculture and horticulture, understanding how pesticides impair, and influence biological efficiency of natural enemy insect species and how natural enemies adjust their detoxification mechanisms to metabolize pesticides is very important. In this work, we found that exposure to sublethal doses of IMD significantly affected the biological performance of A. gifuensis, potentially through changes in the expression of genes involved in the nervous, olfactory, detoxification systems and energy metabolism. Our results indicated that pesticides may block some physiological or biochemical processes that lead to the disruption of the survival, growth, development, reproduction, and behavior of the natural enemies of insect pests. Based on these results, we not only elucidated the sublethal effects of pesticides on the natural enemies, but also contributed to a better understanding of how residual pesticides influence the biological performance of natural enemies and how natural enemies respond to environmental xenobiotics. Our results provide an insight on how to improve experimental approaches, to investigate IPM.
Author Contributions
Z-WK and T-XL designed the study. Z-WK, R-PP, and F-HL performed research. Z-WK, F-HL, and H-GT analyzed data. Z-WK wrote the manuscript. H-GT and T-XL edited the manuscript. Z-WK revised the manuscript.
Funding
This work was supported by the National Key Basic Research Program of China (973 Program) (No. 2013CB127600), China Agriculture Research System (CARS-23-D06), and the China Scholarship Council (award to Z-WK for two year's study abroad at the University of Georgia: 201706300121).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful for Dr. Shun-Hua Han (University of Georgia) for GLMM analysis and the assistance of all staff and students in the Key Laboratory of Applied Entomology, Northwest A&F University at Yangling, Shaanxi, China.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2018.01729/full#supplementary-material
Figure S1. qPCR results of SNF, GST5, SOD1 and SOD2. ns over the bars mean no significant difference, and the error bars is ± SE bars. N = 3.
Figure S2. Activities of SOD, CAT, and GST. ns over the bars mean no significant difference, and the error bars is ± SE bars. N = 3.
Table S1. Primers used for target genes and reference genes in qPCR.
Table S2. RNA quality of transcriptomic samples.
Datasheet 1. Gene expression differences between IMD and CK.
References
Ahmad, M., and Hodgson, C. (1998). The searching efficiency of Aphidius colemani Viereck after visiting insecticide treated plants at different time intervals. Entomon. 23, 185–190.
Ali, A., Desneux, N., Lu, Y. H., Liu, B., and Wu, K. M. (2016). Characterization of the natural enemy community attacking cotton aphid in the Bt cotton ecosystem in Northern China. Sci. Rep. 6:24273. doi: 10.1038/srep24273
Alias, A., Schilthuizen, M., and Sulaiman, H. (2005). “Biodiversity of egg parasitoids of the cocoa pod borer, Conopomorpha cramerella (Snellen) in cocoa smallholders' fields in Sabah, Malaysia,” in Proceedings of 4th Malaysian International Cocoa Conference, eds G. K. Denamany, L. Ling, A. N. Maisin, A. Che, S. B. Nuraziawati, and M. Yazik (Kuala Lumpur: Malaysian Cocoa Board), 486–489.
Badawy, M. E. I., Nasr, H. M., and Rabea, E. I. (2014). Toxicity and biochemical changes in the honey bee Apis mellifera exposed to four insecticides under laboratory conditions. Apidologie 46, 177–193. doi: 10.1007/s13592-014-0315-0
Bass, C., Puinean, A. M., Zimmer, C. T., Denholm, I., Field, L. M., Foster, S. P., et al. (2014). The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 51, 41–51. doi: 10.1016/j.ibmb.2014.05.003
Bayen, S., Giusti, P., Lee, H. K., Barlow, P. J., and Obard, J. P. (2005). Bioaccumulation of DDT pesticide in cultured Asian seabass following dietary exposure. J. Toxicol. Environ. Health A 68, 51–65. doi: 10.1080/15287390590524037
Chaimanee, V., Evans, J. D., Chen, Y., Jackson, C., and Pettis, J. S. (2016). Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 89, 1–8. doi: 10.1016/j.jinsphys.2016.03.004
Chollet, J. F., Couderchet, M., and Bonnemain, J. L. (2014). Crop protection: new strategies for sustainable development. Environ. Sci. Pollut. Res. 21, 4793–4796. doi: 10.1007/s11356-014-2567-7
Conesa, A., Götz, S., García-Gómez, J. M., Terol, J., Talón, M., and Robles, M. (2005). Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676. doi: 10.1093/bioinformatics/bti610
Cook, N., Green, J., Shuker, D. M., and Whitehorn, P. R. (2016). Exposure to the neonicotinoid imidacloprid disrupts sex allocation cue use during superparasitism in the parasitoid wasp Nasonia vitripennis. Ecol. Entomol. 41, 693–697. doi: 10.1111/een.12344
D'Avila, V. A., Barbosa, W. F., Guedes, R. N. C., and Cutler, G. C. (2018). Effects of spinosad, imidacloprid, and lambda-cyhalothrin on survival, parasitism, and reproduction of the aphid parasitoid Aphidius colemani. J. Econ. Entomol. 111, 1096–1103. doi: 10.1093/jee/toy055
De, A., Bose, R., Kumar, A., and Mozumdar, S. (2014). Targeted Delivery of Pesticides Using Biodegradable Polymeric Nanoparticles. New Delhi: Springer, 59–81. doi: 10.1007/978-81-322-1689-6
Dedryver, C. A., Le Ralec, A., and Fabre, F. (2010). The conflicting relationships between aphids and men: a review of aphid damage and control strategies. CR Biol. 333, 539–553. doi: 10.1016/j.crvi.2010.03.009
Desneux, N., Decourtye, A., and Delpuech, J. M. (2007). The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 52, 81–106. doi: 10.1146/annurev.ento.52.110405.091440
Desneux, N., Pham-Delegue, M. H., and Kaiser, L. (2004a). Effects of sub-lethal and lethal doses of lambda-cyhalothrin on oviposition experience and host-searching behaviour of a parasitic wasp, Aphidius ervi. Pest Manag. Sci. 60, 381–389. doi: 10.1002/ps.822
Desneux, N., Rafalimanana, H., and Kaiser, L. (2004b). Dose-response relationship in lethal and behavioural effects of different insecticides on the parasitic wasp Aphidius ervi. Chemosphere 54, 619–627. doi: 10.1016/j.chemosphere.2003.09.007
Ding, Z. P., Wen, Y. C., Yang, B. J., Zhang, Y. X., Liu, S. H., Liu, Z. W., et al. (2013). Biochemical mechanisms of imidacloprid resistance in Nilaparvata lugens: over-expression of cytochrome P450 CYP6AY1. Insect Biochem. Mol. Biol. 43, 1021–1027. doi: 10.1016/j.ibmb.2013.08.005
du Rand, E. E., Human, H., Smit, S., Beukes, M., Apostolides, Z., Nicolson, S. W., et al. (2017). Proteomic and metabolomic analysis reveals rapid and extensive nicotine detoxification ability in honey bee larvae. Insect Biochem. Mol. Biol. 82, 41–51. doi: 10.1016/j.ibmb.2017.01.011
Eakteiman, G., Moses-Koch, R., Moshitzky, P., Mestre-Rincon, N., Vassão, D. G., Luck, K., et al. (2018). Targeting detoxification genes by phloem-mediated RNAi: A new approach for controlling phloem-feeding insect pests. Insect Biochem. Mol. Biol. 100, 10–21. doi: 10.1016/j.ibmb.2018.05.008
Ffrench-Constant, R. H., Williamson, M. S., Davies, T. G., and Bass, C. (2016). Ion channels as insecticide targets. J. Neurogenet. 30, 163–177. doi: 10.1080/01677063.2016.1229781
Fontes, J., Roja, I. S., Tavares, J., and Oliveira, L. (2018). Lethal and sublethal effects of various pesticides on Trichogramma achaeae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 111, 1219–1226. doi: 10.1093/jee/toy064
Funderburk, J., Srivastava, M., Funderburk, C., and McManus, S. (2013). Evaluation of imidacloprid and cyantraniliprole for suitability in conservation biological control program for Orius insidiosus (Hemiptera: Anthocoridae) in field pepper. Fla. Entomol. 96, 229–231. doi: 10.1653/024.096.0131
Gholamzadeh-Chitgar, M., Hajizadeh, J., Ghadamyari, M., Karimi-Malati, A., and Hoda, H. (2015). Effects of sublethal concentration of diazinon, fenitrothion and chlorpyrifos on demographic and some biochemical parameters of predatory bug, Andrallus spinidens Fabricius (Hemiptera: Pentatomidae) in laboratory conditions. Int. J. Pest Manage. 61, 204–211. doi: 10.1080/09670874.2015.1035772
Godfrey, L., and Leigh, T. (1994). Alfalfa harvest strategy effect on Lygus bug (Hemiptera: Miridae) and insect predator population density: implications for use as trap crop in cotton. Environ. Entomol. 23, 1106–1118. doi: 10.1093/ee/23.5.1106
Gong, Y., and Diao, Q. (2017). Current knowledge of detoxification mechanisms of xenobiotic in honey bees. Ecotoxicology 26, 1–12. doi: 10.1007/s10646-016-1742-7
Gong, Y. H., Yu, X. R., Shang, Q. L., Shi, X. Y., and Gao, X. W. (2014). Oral delivery mediated RNA interference of a carboxylesterase gene results in reduced resistance to organophosphorus insecticides in the cotton Aphid, Aphis gossypii Glover. PLoS ONE 9:e102823. doi: 10.1371/journal.pone.0102823
Gong, Y. J., Wang, Z. H., Shi, B. C., Kang, Z. J., Zhu, L., Jin, G. H., et al. (2013). Correlation between pesticide resistance and enzyme activity in the diamondback moth, Plutella xylostella. J. Insect Sci. 13:135. doi: 10.1673/031.013.13501
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Harnpicharnchai, K., Chaiear, N., and Charerntanyarak, L. (2013). Residues of organophosphate pesticides used in vegetable cultivation in ambient air, surface water and soil in Bueng Niam chlo, Khon Kaen, Thailand. Southeast Asian. J. Trop. Med. Public Health 44, 1088–1097.
Hsu, J. C., Chien, T. Y., Hu, C. C., Chen, M. J., Wu, W. J., Feng, H. T., et al. (2012). Discovery of genes related to insecticide resistance in Bactrocera dorsalis by functional genomic analysis of a de novo assembled transcriptome. PLoS ONE 7:e40950. doi: 10.1371/journal.pone.0040950
Kang, Z. W., Liu, F. H., Liu, X., Yu, W. B., Tan, X. L., Zhang, S. Z., et al. (2017a). The potential coordination of the heat-shock proteins and antioxidant enzyme genes of Aphidius gifuensis in response to thermal stress. Front. Physiol. 8:976. doi: 10.3389/fphys.2017.00976
Kang, Z. W., Liu, F. H., Tan, X. L., Zhang, Z. F., Zhu, J. Y., Tian, H. G., et al. (2018a). Infection of powdery mildew reduces the fitness of grain aphids (Sitobion avenae) through restricted nutrition and induced defense response in wheat. Front. Plant Sci. 9:778. doi: 10.3389/fpls.2018.00778
Kang, Z. W., Liu, F. H., Zhang, Z. F., Tian, H. G., and Liu, T. X. (2018b). Volatile β-ocimene can regulate developmental performance of peach aphid Myzus persicae through activation of defense responses in Chinese cabbage Brassica pekinensis. Front. Plant Sci. 9:708. doi: 10.3389/fpls.2018.00708
Kang, Z. W., Tian, H. G., Liu, F. H., Liu, X., Jing, X. F., and Liu, T. X. (2017b). Identification and expression analysis of chemosensory receptor genes in an aphid endoparasitoid Aphidius gifuensis. Sci. Rep. 7:3939. doi: 10.1038/s41598-017-03988-z
Kapoor, U., Srivastava, M. K., and Srivastava, L. P. (2011). Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats. Food Chem. Toxicol. 49, 3086–3089. doi: 10.1016/j.fct.2011.09.009
Karunker, I., Benting, J., Lueke, B., Ponge, T., Nauen, R., Roditakis, E., et al. (2008). Over-expression of cytochrome P450 CYP6CM1 is associated with high resistance to imidacloprid in the B and Q biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Mol. Biol. 38, 634–644. doi: 10.1016/j.ibmb.2008.03.008
Khan, M. A., and Ruberson, J. R. (2017). Lethal effects of selected novel pesticides on immature stages of Trichogramma pretiosum (Hymenoptera: Trichogrammatidae). Pest Manag. Sci. 73, 2465–2472. doi: 10.1002/ps.4639
Kobori, Y., and Amano, H. (2004). Effects of agrochemicals on life-history parameters of Aphidius gifuensis Ashmead (Hymenoptera: Braconidae). Appl. Entomol. Zool. 39, 255–261. doi: 10.1303/aez.2004.255
Kwon, D. H., Kim, J. H., Kim, Y. H., Yoon, K. S., Clark, J. M., and Lee, S. H. (2014). Identification and characterization of an esterase involved in malathion resistance in the head louse Pediculus humanus capitis. Pestic. Biochem. Physiol. 112, 13–18. doi: 10.1016/j.pestbp.2014.05.006
Laycock, I., Lenthall, K. M., Barratt, A. T., and Cresswell, J. E. (2012). Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology 21, 1937–1945. doi: 10.1007/s10646-012-0927-y
Li, H., Tan, J., Song, X., Wu, F., Tang, M., Hua, Q., et al. (2017a). Sublethal doses of neonicotinoid imidacloprid can interact with honey bee chemosensory protein 1 (CSP1) and inhibit its function. Biochem. Biophys. Res. Commun. 486, 391–397. doi: 10.1016/j.bbrc.2017.03.051
Li, H., Wu, F., Zhao, L., Tan, J., Jiang, H., and Hu, F. (2015). Neonicotinoid insecticide interact with honeybee odorant-binding protein: implication for olfactory dysfunction. Int. J. Biol. Macromol. 81, 624–630. doi: 10.1016/j.ijbiomac.2015.08.055
Li, H., Zhao, L., Fu, X., Song, X., Wu, F., Tang, M., et al. (2017b). Physicochemical evidence on sublethal neonicotinoid imidacloprid interacting with an odorant-binding protein from the tea geometrid moth, Ectropis obliqua. J. Agric. Food Chem. 65, 3276–3284. doi: 10.1021/acs.jafc.7b00597
Li, X., Schuler, M. A., and Berenbaum, M. R. (2007). Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 52, 231–253. doi: 10.1146/annurev.ento.51.110104.151104
Li, Y. F., An, J. J., Dang, Z. H., Pan, W. L., and Gao, Z. L. (2018). Systemic control efficacy of neonicotinoids seeds dressing on English grain aphid (Hemiptera: Aphididae). J. Asia-Pac. Entomol. 21, 430–435. doi: 10.1016/j.aspen.2018.01.003
Li, Y. T., Zhao, Q., Duan, X. L., Song, C., and Chen, M. H. (2017). Transcription of four Rhopalosiphum padi (L.) heat shock protein genes and their responses to heat stress and insecticide exposure. Comp. Biochem. Physiol. A 205, 48–57. doi: 10.1016/j.cbpa.2016.12.021
Li, Z., Li, M., He, J., Zhao, X., Chaimanee, V., Huang, W. F., et al. (2017). Differential physiological effects of neonicotinoid insecticides on honey bees: a comparison between Apis mellifera and Apis cerana. Pestic. Biochem. Physiol. 140, 1–8. doi: 10.1016/j.pestbp.2017.06.010
Lim, G. T., and Chong, T. C. (1987). “Biological control of cocoa pod borer by periodic release of Trichogrammatoidea bactrae fumata Nagaraja in Sabah, Malaysia,” in Management of the Cocoa Pod Borer, eds P. A. C. Ooi, L. G. Chan, K. K. Chong, T. C. Hai, M. J. Mamat, H. C. Tuck, and L. G. Soon (Kuala Lumpur: The Malaysian Plant Protection Society), 71–80.
Liu, F., Bao, S. W., Song, Y., Lu, H. Y., and Xu, J. X. (2010). Effects of imidacloprid on the orientation behavior and parasitizing capacity of Anagrus nilaparvatae, an egg parasitoid of Nilaparvata lugens. BioControl 55, 473–483. doi: 10.1007/s10526-010-9276-z
Liu, G., Ma, H., Xie, H., Xuan, N., Guo, X., Fan, Z., et al. (2016). Biotype characterization, developmental profiling, insecticide response and binding property of Bemisia tabaci chemosensory proteins: role of CSP in insect defense. PLoS ONE 11:e0154706. doi: 10.1371/journal.pone.0154706
Liu, S., Liu, F., Jia, H., Yan, Y., Wang, H., Guo, X., et al. (2016). A glutathione S-transferase gene associated with antioxidant properties isolated from Apis cerana cerana. Naturwissenschaften 103:43. doi: 10.1007/s00114-016-1362-3
Lu, Y. H., Wu, K. M., Wyckhuys, K. A. G., and Guo, Y. Y. (2009). Potential of mungbean, Vigna radiatus as a trap crop for managing Apolygus lucorum (Hemiptera: Miridae) on Bt cotton. Crop Prot. 28, 77–81. doi: 10.1016/j.cropro.2008.08.018
Magesh, V., Zhu, Z., Tang, T., Chen, S., Li, L., Wang, L., et al. (2017). Toxicity of Neonicotinoids to honey bees and detoxification mechanism in honey bees. IOSR J. Environ. Sci. Toxicol. Food Technol. 11, 102–110. doi: 10.9790/2402-110401102110
Mao, W., Schuler, M. A., and Berenbaum, M. R. (2011). CYP9Q-mediated detoxification of acaricides in the honey bee (Apis mellifera). Proc. Natl. Acad. Sci. U.S.A. 108, 12657–12662. doi: 10.1073/pnas.1109535108
Mizell, R. F., and Sconyers, M. C. (1992). Toxicity of imidacloprid to selected arthropod predators in the laboratory. Fla. Entomol. 75, 277–280. doi: 10.2307/3495632
Moscardini, V. F., Gontijo Pda, C., Carvalho, G. A., Oliveira, R. L., Maia, J. B., and Silva, F. F. (2013). Toxicity and sublethal effects of seven insecticides to eggs of the flower bug Orius insidiosus (Say) (Hemiptera: Anthocoridae). Chemosphere 92, 490–496. doi: 10.1016/j.chemosphere.2013.01.111
Murray, D. A., Clarke, M. B., and Ronning, D. A. (2013). Estimating invertebrate pest losses in six major Australian grain crops. Aust. J. Entomol. 52, 227–241. doi: 10.1111/aen.12017
Nawaz, M., Cai, W., Jing, Z., Zhou, X., Mabubu, J. I., and Hua, H. (2017). Toxicity and sublethal effects of chlorantraniliprole on the development and fecundity of a non-specific predator, the multicolored Asian lady beetle, Harmonia axyridis (Pallas). Chemosphere 178, 496–503. doi: 10.1016/j.chemosphere.2017.03.082
Nielsen, S. A., Brodsgaard, C. J., and Hansen, H. (2000). Effects on detoxification enzymes in different life stages of honey bees (Apis mellifera L., Hymenoptera: Apidae) treated with a synthetic pyrethroid (flumethrin). Altern. Lab. Anim. 28, 437–443.
Ohta, I., and Takeda, M. (2015). Acute toxicities of 42 pesticides used for green peppers to an aphid parasitoid, Aphidius gifuensis (Hymenoptera: Braconidae), in adult and mummy stages. Appl. Entomol. Zool. 50, 207–212. doi: 10.1007/s13355-015-0323-1
Paine, M. J., Scrutton, N. S., Munro, A. W., Gutierrez, A., Roberts, G. C., and Wolf, C. R. (2005). “Electron transfer partners of cytochrome P450,” in Cytochrome P450: Structure, Mechanism, and Biochemistry, ed P. Ortiz De Montellano (New York, NY: Kluwer Academic/Plenum Publishers), 115–138.
Potrich, M., Alves, L. F. A., Lozano, E. R., Bonini, A. K., and Neves, P. M. O. J. (2017). Potential side effects of the entomopathogenic Fungus Metarhizium anisopliae on the egg parasitoid Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) under controlled conditions. J. Econ. Entomol. 110, 2318–2324. doi: 10.1093/jee/tox257
Puinean, A. M., Foster, S. P., Oliphant, L., Denholm, I., Field, L. M., Millar, N. S., et al. (2010). Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 6:e1000999. doi: 10.1371/journal.pgen.1000999
Ramos, R. S., de Araujo, V. C. R., Pereira, R. R., Martins, J. C., Queiroz, O. S., Silva, R. S., et al. (2018). Investigation of the lethal and behavioral effects of commercial insecticides on the parasitoid wasp Copidosoma truncatellum. Chemosphere 191, 770–778. doi: 10.1016/j.chemosphere.2017.10.113
Rogers, M. A., Krischik, V. A., and Martin, L. A. (2007). Effect of soil application of imidacloprid on survival of adult green lacewing, Chrysoperla carnea (Neuroptera: Chrysopidae), used for biological control in greenhouse. Biol. Control 42, 172–177. doi: 10.1016/j.biocontrol.2007.05.006
Sabahi, Q., Talebi, K., Kavousi, A., and Sheikhi Garjan, A. (2010). Effects of imidacloprid, dichlorvos, pymetrozine and abamectin, on life table parameters of the predatory bug, Orius albidipennis (Hemiptera: Anthocoridae). J. Entomol. Soc. Iran. 30, 1–11.
Saeed, R., Abbas, N., Razaq, M., Mahmood, Z., Naveed, M., and Rehman, H. M. U. (2018). Field evolved resistance to pyrethroids, neonicotinoids and biopesticides in Dysdercus koenigii (Hemiptera: Pyrrhocoridae) from Punjab, Pakistan. Chemosphere 213, 149–155. doi: 10.1016/j.chemosphere.2018.09.042
Salerno, G., Colazza, S., and Conti, E. (2002). Sub-lethal effects of deltamethrin on walking behaviour and response to host kairomone of the egg parasitoid Trissolcus basalis. Pest Manag. Sci. 58, 663–668. doi: 10.1002/ps.492
Schoofs, L., De Loof, A., and Van Hiel, M. B. (2017). Neuropeptides as regulators of behavior in insects. Annu. Rev. Entomol. 62, 35–52. doi: 10.1146/annurev-ento-031616-035500
Sclar, D. C, Gerace, D., and Cranshaw, W. S. (1998). Observations of population increases and injury by spider mites (Acari: Tetranychidae) on ornamental plants treated with imidacloprid. J. Econ. Entomol. 91, 250–255. doi: 10.1093/jee/91.1.250
Shah, P. A., and Pell, J. K. (2003). Entomopathogenic fungi as biological control agents. Appl. Microbiol. Biotechnol. 61, 413–423. doi: 10.1007/s00253-003-1240-8
Shelton, A. M., and Badenes-Perez, F. R. (2006). Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 51, 285–308. doi: 10.1146/annurev.ento.51.110104.150959
Shi, T. F., Wang, Y. F., Liu, F., Qi, L., and Yu, L. S. (2017). Sublethal effects of the neonicotinoid insecticide thiamethoxam on the transcriptome of the honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 110, 2283–2289. doi: 10.1093/jee/tox262
Silva, A. X., Jander, G., Samaniego, H., Ramsey, J. S., and Figueroa, C. C. (2012). Insecticide resistance mechanisms in the green peach aphid Myzus persicae (Hemiptera: Aphididae) I: a transcriptomic survey. PLoS ONE 7:e36366. doi: 10.1371/journal.pone.0036366
Stapel, J. O., Cortesero, A. M., and Lewis, W. J. (2000). Disruptive sublethal effects of insecticides on biological control: altered foraging ability and life span of a parasitoid after feeding on extrafloral nectar of cotton treated with systemic insecticides. Biol. Control 17, 243–249. doi: 10.1006/bcon.1999.0795
Staron, M., Sabo, R., Sobekova, A., Sabova, L., Legath, J., Lohajova, L., et al. (2017). Formetanate toxicity and changes in antioxidant enzyme system of Apis mellifera larvae. Environ. Sci. Pollut. Res. Int. 24, 14060–14070. doi: 10.1007/s11356-017-8966-9
Studebaker, G. E., and Kring, T. J. (2003). Effects of insecticides on Orius insidiosus (Hemiptera: Anthocoridae), measured by field, greenhouse and petri dish bioassays. Fla. Entomol. 86, 178–185. doi: 10.1653/0015-4040(2003)086[0178:EOIOOI]2.0.CO;2
Suh, Y.T., and Shim, J. H. (1988). Enzyme activities of a honeybee Apis-mellifera l. associated with the degradation of some insecticides. Agric. Chem. Biotechnol. 31, 241–248.
Sun, Z. J., Chen, D., Jia, F. Z., Zhang, C. H., Tang, S. H., Ren, G. W., et al. (2014). Effect of six conventional insecticides on Aphidius gifuensis Ashmead in tobacco fields. Plant Prot. 40, 185–189. doi: 10.3969/j.issn.0529-1542.2014.04.038
Tabashnik, B. E., Van Rensburg, J. B. J., and Carrière, Y. (2009). Field-evolved insect resistance to Bt crops: definition, theory, and data. J. Econ. Entomol. 102, 2011–2025. doi: 10.1603/029.102.0601
Tappert, L., Pokorny, T., Hofferberth, J., and Ruther, J. (2017). Sublethal doses of imidacloprid disrupt sexual communication and host finding in a parasitoid wasp. Sci. Rep. 7:42756. doi: 10.1038/srep42756
Traverso, L., Lavore, A., Sierra, I., Palacio, V., Martinez-Barnetche, J., Latorre-Estivalis, J. M., et al. (2017). Comparative and functional triatomine genomics reveals reductions and expansions in insecticide resistance-related gene families. PLoS Negl. Trop. Dis. 11:e0005313. doi: 10.1371/journal.pntd.0005313
Valenzuela, I., and Hoffmann, A. A. (2015). Effects of aphid feeding and associated virus injury on grain crops in A ustralia. Aust. Entomol. 54, 292–305. doi: 10.1111/aen.12122
Van Emden, H. F., and Harrington, R. (2017). Aphids as Crop Pests. Wallingford: CAB International. doi: 10.1079/9781780647098.0000
Walker, J. T., Suckling, D. M., and Wearing, C. H. (2017). Past, present, and future of integrated control of apple pests: the New Zealand experience. Annu. Rev. Entomol. 62, 231–248. doi: 10.1146/annurev-ento-031616-035626
Wang, D., Lü, L., and He, Y. (2017). Effects of insecticides on sex pheromone communication and mating behavior in Trichogramma chilonis. J. Pest Sci. 91, 65–78. doi: 10.1007/s10340-017-0864-x
Wang, K., Zhang, M., Huang, Y., Yang, Z., Su, S., and Chen, M. H. (2017). Characterisation of imidacloprid resistance in the bird cherry-oat aphid, Rhopalosiphum padi, a serious pest on wheat crops. Pest Manag. Sci. 74, 1457–1465. doi: 10.1002/ps.4834
Wang, L., Zeng, L., and Chen, J. (2015). Sublethal effect of imidacloprid on Solenopsis invicta (Hymenoptera: Formicidae) feeding, digging, and foraging behavior. Environ. Entomol. 44, 1544–1552. doi: 10.1093/ee/nvv127
Wang, L. L., Huang, Y., Lu, X. P., Jiang, X. Z., Smagghe, G., Feng, Z. J., et al. (2015). Overexpression of two alpha-esterase genes mediates metabolic resistance to malathion in the oriental fruit fly, Bactrocera dorsalis (Hendel). Insect Mol. Biol. 24, 467–479. doi: 10.1111/imb.12173
Whitehorn, P. R., O'connor, S., Wackers, F. L., and Goulson, D. (2012). Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 336, 351–352. doi: 10.1126/science.1215025
Wu, X. M., Xu, B. Y., Si, F. L., Li, J., Yan, Z. T., Yan, Z. W., et al. (2018). Identification of carboxylesterase genes associated with pyrethroid resistance in the malaria vector Anopheles sinensis (Diptera: Culicidae). Pest Manag. Sci. 74, 159–169. doi: 10.1002/ps.4672
Xiao, D., Tan, X., Wang, W., Zhang, F., Desneux, N., and Wang, S. (2017). Modification of flight and locomotion performances, respiratory metabolism, and transcriptome expression in the lady beetle Harmonia axyridis through sublethal pesticide exposure. Front. Physiol. 8:33. doi: 10.3389/fphys.2017.00033
Xie, M., Ren, N. N., You, Y. C., Chen, W. J., Song, Q. S., and You, M. S. (2017). Molecular characterisation of two alpha-esterase genes involving chlorpyrifos detoxification in the diamondback moth, Plutella xylostella. Pest Manag. Sci. 73, 1204–1212. doi: 10.1002/ps.4445
Xuan, N., Guo, X., Xie, H. Y., Lou, Q. N., Lu, X. B., Liu, G. X., et al. (2015). Increased expression of CSP and CYP genes in adult silkworm females exposed to avermectins. Insect Sci. 22, 203–219. doi: 10.1111/1744-7917.12116
Yan, H., Jia, H., Gao, H., Guo, X., and Xu, B. (2013). Identification, genomic organization, and oxidative stress response of a sigma class glutathione S-transferase gene (AccGSTS1) in the honey bee, Apis cerana cerana. Cell Stress Chaperon. 18, 415–426. doi: 10.1007/s12192-012-0394-7
Yang, F., Wu, Y. K., Xu, L., Wang, Q., Yao, Z. W., Zikic, V., et al. (2017). Species composition and richness of aphid parasitoid wasps in cotton fields in northern China. Sci. Rep. 7:9799. doi: 10.1038/s41598-017-10345-7
Yang, S., Wei, J. N., Yang, S. Y., and Kuang, R. P. (2011). Current status and future trends of augmentative release of Aphidius gifuensis for control of Myzus persicae in China's Yunnan province. J. Entomol. Res. Soc. 13, 87–99.
Yang, S., Yang, S. Y., Zhang, C. P., Wei, J. N., and Kuang, R. P. (2009). Population dynamics of Myzus persicae on tobacco in Yunnan Province, China, before and after augmentative releases of Aphidius gifuensis. Biocontrol Sci. Techn. 19, 219–228. doi: 10.1080/09583150802696525
Yang, Y. X., Yu, N., Zhang, J. H., Zhang, Y. X., and Liu, Z. W. (2017). Induction of P450 genes in Nilaparvata lugens and Sogatella furcifera by two neonicotinoid insecticides. Insect Sci. 25, 401–408. doi: 10.1111/1744-7917.12440
Ye, J., Fang, L., Zheng, H., Zhang, Y., Chen, J. L., Zhang, Z. J., et al. (2006). WEGO: a web tool for plotting GO annotations. Nucleic Acids Res. 34, W293–W297. doi: 10.1093/nar/gkl031
Yu, S. J., Robinson, F. A., and Nation, J. L. (1984). Detoxication capacity in the honey bee, Apis mellifera L. pestic. Biochem. Physiol. 22, 360–368. doi: 10.1016/0048-3575(84)90029-4
Zhang, C., Hu, R. F., Shi, G. M., Jin, Y. H., Robson, M. G., and Huang, X. S. (2015). Overuse or underuse? An observation of pesticide use in China. Sci. Total Environ. 538, 1–6. doi: 10.1016/j.scitotenv.2015.08.031
Zhang, S. Z., Zhang, X. L., Shen, J., Mao, K. K., You, H., and Li, J. H. (2016). Susceptibility of field populations of the diamondback moth, Plutella xylostella, to a selection of insecticides in Central China. Pestic. Biochem. Physiol. 132, 38–46. doi: 10.1016/j.pestbp.2016.01.007
Zhang, Y., Yan, H., Lu, W., Li, Y., Guo, X., and Xu, B. (2013). A novel Omega-class glutathione S-transferase gene in Apis cerana cerana: molecular characterisation of GSTO2 and its protective effects in oxidative stress. Cell Stress Chaperon. 18, 503–516. doi: 10.1007/s12192-013-0406-2
Keywords: Aphidius gifuensis, imidacloprid, biological performance, transcriptome, integrated pest management
Citation: Kang Z-W, Liu F-H, Pang R-P, Tian H-G and Liu T-X (2018) Effect of Sublethal Doses of Imidacloprid on the Biological Performance of Aphid Endoparasitoid Aphidius gifuensis (Hymenoptera: Aphidiidae) and Influence on Its Related Gene Expression. Front. Physiol. 9:1729. doi: 10.3389/fphys.2018.01729
Received: 06 July 2018; Accepted: 16 November 2018;
Published: 11 December 2018.
Edited by:
Bin Tang, Hangzhou Normal University, ChinaReviewed by:
Abid Ali, University of Agriculture, Faisalabad, PakistanThorben Müller, Bielefeld University, Germany
Rakesh Kumar Seth, University of Delhi, India
Copyright © 2018 Kang, Liu, Pang, Tian and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hong-Gang Tian, dGlhbmhnQG53c3VhZi5lZHUuY24=
Tong-Xian Liu, dHhsaXVAbndzdWFmLmVkdS5jbg==.
†These authors have contributed equally to this work