Abstract
Over a century's worth of research suggests that, for a variety of verbal and perceptual memory phenomena, retrieval modifies memory in two ways. First, retrieved information is strengthened and thus more easily retrieved on subsequent recall tests. Secondly, when information is stored or organized in memory in such a way that multiple representations share a common retrieval cue, and are thus subject to interference, related information that is not retrieved becomes less recallable. Such retrieval-induced forgetting is a highly robust and well-understood phenomenon. Over the past decade, several experiments have demonstrated that retrieval-induced forgetting also persists for motor responses–both simple and complex, and that this impairment is observed for representations stored in both declarative and procedural memory. While several review papers have focused on retrieval-induced forgetting of verbal and perceptual information, to date no literature review has focused exclusively on retrieval-induced forgetting of motor responses or actions. In addition, while retrieval-induced forgetting has been documented extensively within the domain of declarative memory, only a few papers have been published documenting retrieval-induced forgetting in procedural memory. Therefore, this review paper will explore these findings and their primary implications, most notably that when multiple motor programs are initiated via a shared retrieval cue, inhibitory suppression is recruited to decrease accessibility of related, but inappropriate actions in service of potentiating retrieval of appropriate actions. More broadly, the same processes that govern selective retrieval in cognition are implicated in the selective retrieval of motor actions.
Introduction
A recent study estimates that the average internet user has 100 online accounts that require passwords (Pilar et al., 2012; Pearman et al., 2017). Unsurprisingly, 33% of Americans report frequent forgetting of passwords, unless they are written down (Brown et al., 2004). Cipresso et al. (2012) recommend using their PsychoPass method as a means of creating complex, secure, but easy-to-remember passwords. To use this technique, all one has to do is memorize a single letter on the keyboard and a series of subsequent motor actions. For example, A-press one key to the left-press one key up-press one key to the right. This method creates an infinite number of passwords that are all random and nonsensical, and thus difficult for hackers to penetrate. With the advent of the internet and related technologies however, individuals are often faced with the difficult task of memorizing and executing a series of similar motor sequences, such as when entering a password or PIN into an ATM. If these motor sequences are stored and organized in memory in such a way that they share a common retrieval cue, these motor sequences could be subject to interference. For example, if multiple motor sequences share the same effector with which the movement is initiated, the motor programs may compete for retrieval access. Successful execution of movement may thus require resolution of interference between motor programs (Tempel and Frings, 2013, 2014a,b, 2015, 2016, 2017). Avid ping pong players for instance, may struggle to play tennis because, while the shots are the same for both ping-pong and tennis, the former requires extensive movements of the wrist, while the latter is played without using the wrist. If both motor programs are activated simultaneously, the more familiar, dominant, and better practiced wrist sequences used for ping pong could interfere with the lesser practiced non-wrist movement sequences used for tennis, thus blocking them and preventing them from being executed properly (Tempel and Frings, 2013).
Retrieval-induced forgetting
One possible mechanism that may resolve interference between motor programs is retrieval inhibition (Bjork and Bjork, 1988; Bjork, 1989). Retrieval inhibition has been implicated as the primary mechanism for resolving interference in several different types of verbal memory phenomena, including retrieval-induced forgetting (Anderson et al., 1994). Retrieval-induced forgetting (RIF) is primarily investigated via a retrieval practice paradigm that consists of three phases (see Figure 1). During the initial learning phase, participants study a series of word pairs comprised of a category label and an exemplar derived from that category. For example, FRUIT: LEMON, FISH: HERRING, METALS: SILVER. Participants typically learn a total of 48 category-exemplar pairs–eight categories, each containing six exemplars. Subsequently, during the retrieval practice phase, participants are given a category label followed by a two-letter stem cue (e.g., FRUIT: LE, FISH: TR) and must generate the correct exemplar. Participants typically perform retrieval practice three times, on half of the exemplars from half of the studied categories. After a short distractor task, participants are tested on all of the category-exemplar pairs that they studied (FRUIT: L, FRUIT: B, FISH: T, FISH: H, METALS: S).
Figure 1

Retrieval practice paradigm and typical results. Retrieval-induced forgetting is investigated using the retrieval practice paradigm. During the initial study phase, participants study a series of category-exemplar pairs. Subsequently, during the retrieval practice phase, participants retrieve half of the exemplars from half of the studied categories. After a brief distractor task, participants are tested on all of the exemplars that they studied. Category-exemplar pairs that were subject to retrieval practice are better recalled than baseline items from other categories that were not featured during retrieval practice. Unpracticed category-exemplar pairs from practiced categories however, show impaired recall relative to baseline items, thus demonstrating retrieval-induced forgetting. Figure adapted from Anderson et al. (1994).
The pattern of results typically observed in this paradigm highlight the advantages and disadvantages conferred by retrieval practice. The advantage is that, unsurprisingly, items that were subject to retrieval practice (henceforth termed Rp+ items; e.g., FRUIT: LEMON; FISH: TROUT) are recalled better than items from other categories that were not practiced (henceforth termed Nrp or baseline items; e.g., METALS: SILVER). The costs of retrieval practice are demonstrated when recall of unpracticed exemplars from practiced categories (e.g., FRUIT: BANANA, FISH: HERRING; henceforth termed Rp– items) is compared to recall for Nrp or baseline items. While theories of spreading activation (Collins and Loftus, 1975; Anderson, 1983), would predict that studying some items sharing a category cue would facilitate recall of related items from that category, access to related items is subsequently impaired. That is, repeated retrieval practice of FRUIT: LEMON impedes later recollection of FRUIT: BANANNA on the final test. This phenomenon of impaired recall of related exemplars following retrieval practice is known as retrieval-induced forgetting. Retrieval-induced forgetting is an extremely robust and thoroughly replicated phenomenon. It has been observed with a variety of materials, including pictures (Maxcey and Woodman, 2014; Maxcey et al., 2021), text passages (Carroll et al., 2007), factual propositions (Anderson and Bell, 2001), phonological categories (Bajo et al., 2006), visuospatial materials (Ciranni and Shimamura, 1999), and mathematical equations (Campbell and Thompson, 2012). Retrieval-induced forgetting has also been observed outside of the laboratory in a number of applied, real-world contexts including eyewitness memory (Shaw et al., 1995; MacLeod, 2002; Saunders and MacLeod, 2002) and education (Carroll et al., 2007; Little et al., 2011—for a review of RIF in applied contexts, see Storm et al., 2015).
There are several theories that attempt to explain why retrieval-induced forgetting occurs (see Figure 2). The inhibitory account of retrieval-induced forgetting contends that during the initial retrieval practice phase, when one attempts to recall LEMON in response to the cue FRUIT: LE, the word FRUIT activates all of the members of that category, creating interference. In order to resolve such interference, individuals must suppress or select against the related exemplars, deactivating them and making it easier to retrieve the correct exemplar. The consequence of such suppression is that those items are less recallable on the final test (Anderson et al., 1994; Anderson and Spellman, 1995; Anderson, 2003). Various noninhibitory explanations have also been proposed to account for retrieval-induced forgetting. For instance, the associative interference account contends that when Rp+ items are subject to retrieval practice, the association between the category cue (e.g., FRUIT) and the exemplar (e.g., LEMON) becomes strengthened and highly accessible. This strengthened association causes LEMON to intrude perseveratively during the final test, thus blocking participants' access to related items from the same category, such as STRAWBERRY (for literature reviews of the interference account of RIF, see Verde, 2012; Raaijmakers and Jakab, 2013). Thus, the primary difference between the inhibitory account and the associative interference account is that while the former argues that retrieval-induced forgetting is caused by inhibition, recruited to suppress competing Rp– exemplars and resolve interference during retrieval practice, so as to facilitate selective retrieval of target Rp+ items, the latter assumes that retrieval practice strengthens the association between the category cue and target Rp+ exemplars, and this strengthened association prevents participants from recalling other nontarget items from the same category on the final test. There are numerous findings that uniquely support the inhibitory account, however (see Figure 3). For example, retrieval-induced forgetting is cue independent, strength independent, competition dependent, and retrieval specific (for literature reviews of the inhibitory account of RIF, see Anderson, 2003; Storm and Levy, 2012; Marsh and Anderson, 2022; for a meta-analytic review of RIF, see Murayama et al., 2014). Each of these properties of retrieval-induced forgetting will be discussed in turn.
Figure 2

The inhibitory account of retrieval-induced forgetting. The inhibitory account of retrieval-induced forgetting posits that during retrieval practice, when attempting to recall the target item LEMON in response to the cue FRUIT: LE, LEMON is activated, but so are competing nontarget items from the same category, such as APPLE, STRAWBERRY, and BANANNA. Both the target and nontarget items compete for retrieval access. To overcome such interference and facilitate retrieval of the target item, inhibitory suppression is recruited to select against and decrease the accessibility of nontarget competing items.
Figure 3

Properties of retrieval-induced forgetting. The inhibitory account of RIF is the best understood explanation for RIF and is supported by a wealth of evidence. RIF caused by inhibition exhibits four key properties—cue independence, strength independence, retrieval specificity, and competition dependence. The table above includes a definition for each property, an example from the RIF literature as it pertains to verbal memory, and an example of the same property replicated in motor memory.
Properties of retrieval-induced forgetting that are consistent with the inhibitory account
Cue independence
Anderson and Spellman (1995) were the first to demonstrate that retrieval-induced forgetting persists on the final test not just when participants are asked to generate an exemplar using the initially studied cue, or to recall BANANA when given the cue FRUIT: B, for example. Reliable retrieval-induced forgetting is also observed when participants are asked to recall the item with a totally novel or independent cue (e.g., MONKEY: B). Why is this a problem for the associative interference account of retrieval-induced forgetting? The interference account maintains that what gets strengthened during retrieval practice is the association between the exemplar and the initially studied category cue. Therefore, if an independent probe or novel category cue is used to retrieve the exemplar, there should be no memory impairment. In contrast, the inhibitory account maintains that during retrieval-practice of Rp+ items, competing Rp– items are suppressed. Said differently, according to inhibitory account, the exemplars themselves are suppressed to resolve interference, while the associative interference account contends that the association between the initially studied category cue and the practiced exemplars are strengthened, thus blocking or preventing recall of related, unpracticed items on the final test. The fact that retrieval-induced forgetting persists with a novel cue is thus consistent with the inhibitory account. The cue independence property of retrieval-induced forgetting is highly robust and generalizable (Anderson et al., 2000a,b; Anderson and Bell, 2001; Saunders and MacLeod, 2002; Johnson and Anderson, 2004; Veling and van Knippenberg, 2004; MacLeod and Saunders, 2005; Bajo et al., 2006; Aslan et al., 2007; Levy et al., 2007; Hulbert and Shivde, 2012). When Murayama et al. (2014) examined all of the published studies that have utilized an independent cue on the final test in their meta-analysis, a significant retrieval-induced forgetting effect was found, though it was smaller than the retrieval-induced forgetting effect observed when the studied cues were used on final test.
Despite the relatively high incidence of cue independence in studies of retrieval-induced forgetting, some failures to replicate have occurred. In addition, some have raised the concern that “independent” cues may not be truly independent. For example, according to the covert-cuing hypothesis, when participants are exposed to a novel or independent category cue during the final test (e.g., MONKEY), instead of using that category cue alone to retrieve the appropriate exemplar (e.g., BANANA), participants covertly generate the initially studied cue (e.g., FRUIT) and use it to facilitate retrieval of the exemplar. Because participants are therefore still making use of the association between the exemplar and the studied cue, this explanation is consistent with the interference account (Williams and Zacks, 2001; Perfect et al., 2004; Camp et al., 2007, 2009). That is, according to the covert-cuing hypothesis, if participants covertly retrieve FRUIT when presented with MONKEY: BANANA, all of the interfering exemplars associated with the FRUIT category could become activated and prevent successful recall of BANANA. Thus, even independent cues could be subject to associative interference. In support of the covert-cuing hypothesis, Camp et al. (2005) discovered that when participants did not know that the study and test phases were related, no retrieval-induced forgetting to independent cues was observed. However, when participants were explicitly informed that the study and test phases were related, significant cue-independent forgetting was observed. However, other researchers have found significant cue-independent forgetting in situations where the use of previously studied cues was very unlikely to occur (Shivde and Anderson, 2001; Johnson and Anderson, 2004), thus challenging the generalizability of Camp et al.'s (2005) results.
Other evidence that independent cues may not be truly independent comes from experiments demonstrating cue-enhancement effects. Camp et al. (2009) demonstrated that manipulating the accessibility of studied cues can affect recall even when participants are given an independent cue at test. In the procedure adopted by Camp and colleagues, participants learned 24 category-exemplar pairs (e.g., ROPE: SAILING, SUNFLOWER: YELLOW) and were subsequently tested on all 24 items using an independent cue (e.g., SPORT, COLOR). Prior to the learning phase however, a subset of the cues were presented to participants without the associated exemplar. Each cue was presented twice. During the first presentation of the cue, participants' task was to study each cue for 2 s and rate its pleasantness on a scale from 1 to 5. On the second presentation, each cue was again presented for 2 s, and participants were instructed to rate each item's relative frequency of use in the Dutch language on a scale from 1 to 5. Interestingly, the results revealed that prior exposure to the studied cue facilitated recall of the associated item, even though an independent cue was used. That is, recall of YELLOW in response to the cue COLOR was better when participants received additional exposure to the studied cue SUNFLOWER. If the recall of YELLOW given the cue COLOR is dependent on how accessible SUNFLOWER is, COLOR cannot be considered an independent cue. Therefore, even though Camp and colleagues' paradigm did not specifically investigate retrieval-induced forgetting, their results call into question the assumption that independent cues are truly independent. However, Huddleston and Anderson (2012) demonstrated that the cue-enhancement effects observed by Camp et al. only occur when there is a strong semantic relationship between the initially studied cue and the independent cue. For example, during the learning phase Camp et al. presented word pairs such as BEAK: DUCK and ZOO: TIGER. The independent probes they choose for these items were BIRD and ANIMAL respectively. Given the high degree of semantic similarity between the studied cues and their independent cues, it seems likely that participants would generate both cues during retrieval, thus facilitating recall of the target item. However, when Huddleston and Anderson used studied cues and independent probes that were totally unrelated (e.g., presenting GATE: DAISY during the study phase and later cuing it with the novel probe FLOWER; GATE and FLOWER have no semantic relation), no cue-enhancement effect was observed.
Thus, such cue-enhancement effects may simply be an artifact of the type of study materials selected. Furthermore, in some cases covert-cuing has abolished cue-independent forgetting instead of increasing it. For example, when Weller et al. (2013) explicitly instructed participants to adopt a covert-cuing strategy and use previously studied cues to help them recall the associated exemplar on a final test utilizing independent cues, no cue-independent retrieval-induced forgetting effect was observed. Why does covert-cuing eliminate retrieval-induced forgetting? Covert-cuing can often mask the effects of inhibition, due to the fact that Rp– items are made more accessible when augmented by the presence of both the initially studied cue and the independent probe. Recall of baseline or Nrp items also increases with access to studied cues, but Rp– items incur more of a retrieval benefit given the fact that the studied cues are highly accessible as a result of prior retrieval practice (Marsh and Anderson, 2022).
Strength independence and retrieval specificity
According to the interference account of retrieval-induced forgetting, the magnitude of the impairment effect observed for Rp– items should be directly related to the degree to which Rp+ items are strengthened during retrieval-practice. Said differently, retrieval-induced forgetting attributed solely to interference dynamics during the final test should be strength dependent. If retrieval practice strengthens the association between the category cue and practiced exemplars, and this strengthened association blocks successful recollection of weaker Rp– items, the more accessible the Rp+ items are, the more difficult it should be to access the Rp– items. The most straightforward way to test this prediction is investigate whether the observed facilitation effect for Rp+ items relative to baseline items positively correlates with the degree of retrieval-induced forgetting observed. Several researchers have done this (Hanslmayr et al., 2010; Staudigl et al., 2010; Hulbert and Shivde, 2012). Most strikingly however, when Murayama et al. (2014) examined this correlation in their meta-analysis, no relationship was observed between the facilitation of Rp+ items and the recall of Rp– items, provided the final test appropriately indexed forgetting caused by inhibition. Related findings of note are that providing feedback during retrieval-practice (Erdman and Chan, 2013; Murayama et al., 2014; Tempel and Frings, 2018) or increasing the number of retrieval practice trials fails to increase the amount of retrieval-induced forgetting observed (Macrae and MacLeod, 1999), provided the test used controls for output interference and thus appropriately measures forgetting due to inhibitory mechanisms. Therefore, the degree to which Rp+ items are strengthened during retrieval practice is totally unrelated to the size of the impairment observed for Rp– items. Retrieval-induced forgetting is therefore considered to be strength-independent.
Several experiments have demonstrated that extra study in place of retrieval practice fails to elicit retrieval-induced forgetting, despite the fact that significant strengthening of Rp+ items occurs in both cases (Anderson et al., 2000a,b; Anderson and Bell, 2001; Shivde and Anderson, 2001; Bäuml, 2002; Buchli et al., 2016; Rupprecht and Bäuml, 2016, 2017). For example, if after the initial learning phase participants are again presented with the intact category-exemplar pair FRUIT: ORANGE, and are prompted to restudy it numerous times, no retrieval-induced forgetting occurs. Retrieval-induced forgetting is only present in the standard retrieval practice condition, wherein participants are given the category-plus-letter stem cue FRUIT: OR and are prompted to recall the exemplar ORANGE. When Murayama et al. (2014) examined all of the published experiments that included restudy in place of retrieval-practice in their meta-analysis, while restudy produced significant strengthening of Rp+ items, no significant decrease in recall of Rp– items was observed.
Other research has demonstrated that significant retrieval-induced forgetting can occur without strengthening any items during the retrieval-practice phase. In perhaps the most striking demonstration of strength independence, after presenting a series of category-exemplar pairs to participants, Storm et al. (2006) observed significant retrieval-induced forgetting regardless of whether retrieval-practice was possible or impossible. During the possible retrieval practice trials, participants were shown category-plus-two-letter stem cues like FRUIT: LE for which they could successfully generate the corresponding exemplar LEMON. Impossible retrieval practice trials, in contrast, included cues such as FRUIT: LU (there is no corresponding fruit that begins with the letters LU). When Murayama et al. (2014) compared experiments that utilized the impossible retrieval practice manipulation to standard retrieval-induced forgetting studies, the effect size of the observed retrieval-induced forgetting effect in the impossible paradigm was statistically significant and similar to that observed in the standard paradigm. Thus, what appears to be a critical prerequisite for retrieval-induced forgetting is not the strengthening of practiced exemplars during retrieval practice, but rather the retrieval attempt itself. That is, retrieval-induced forgetting is strength independent and retrieval specific, in the sense that impairment of related, unpracticed Rp– items is only induced if Rp+ items are actively retrieved from long-term memory during retrieval-practice.
While these results are incompatible with strength-dependent associative interference accounts of retrieval-induced forgetting, they are highly consistent with the inhibition account. The inhibitory account of retrieval-induced forgetting posits that when one attempts to selectively retrieve target Rp+ items during retrieval practice, the target item is activated in response to the category cue, but so are all of the other related, nontarget Rp– items associated with the same category cue. Coactivation of target Rp+ and nontarget Rp– items thus creates retrieval competition. Inhibitory suppression is therefore recruited as an adaptive mechanism to reduce the accessibility of competing nontarget items, and in so doing facilitate selective retrieval of target or Rp+ items. The consequence of the inhibitory suppression of competing items during retrieval practice is that, by virtue of their reduced accessibility, those items are more difficult to recall on the final test. If inhibition acts to reduce the accessibility of interfering or competing nontarget items to enhance retrieval of target items, it is the retrieval attempt in the face of competition/interference that matters rather than the strength of the target item. In contrast, the associative interference account of retrieval-induced forgetting assumes that impairment of nontarget Rp– items is entirely a consequence of strengthening the association between the target Rp+ items and their respective category cues. If retrieval of target items is impossible and therefore unsuccessful, no impairment of nontarget Rp– items should occur. Similarly, strengthening the cue-target association by any means should result in retrieval-induced forgetting. Therefore, in sum, the collective observations that retrieval-induced forgetting persists when successful retrieval of Rp+ items is impossible (Storm et al., 2006) and fails to emerge during restudy despite significant strengthening of Rp+ items (Anderson et al., 2000a,b; Bäuml, 2002; Buchli et al., 2016; Rupprecht and Bäuml, 2016, 2017), provide compelling support in favor of the inhibitory account, and invalidate many of the key assertions of the associative interference account.
In addition to the extensive behavioral evidence discussed above, neuroscientists have established converging evidence in support of the strength independence property of retrieval-induced forgetting. For example, a multitude of studies employing a variety of different neuroimaging techniques have demonstrated that the anterior cingulate cortex and the lateral prefrontal cortex are engaged during retrieval practice, but not during restudy, and that activity in these regions correlates with retrieval-induced forgetting (Kuhl et al., 2007; Wimber et al., 2008, 2015).
Further evidence implicating the lateral prefrontal cortex as a structure that is related to retrieval-induced forgetting comes from studies investigating stress. After the initial learning phase, in which participants memorized a series of category-exemplar pairs, Koessler et al. (2009) instructed participants to either complete the Trier social stress test, in which they were required to give a speech and perform arithmetic problems in front of an audience or complete a control task in which participants wrote a speech and solved arithmetic problems by themselves. Subsequently, both groups performed retrieval practice on a subset of the items they learned and were later tested on all the category-exemplar pairs. The final test revealed significant retrieval-induced forgetting in the control condition, but no retrieval induced forgetting when participants completed the stressful task. Why? Stress increases salivary cortisol, which disrupts proper functioning of the lateral prefrontal cortex. Critically, while stress abolished retrieval-induced forgetting, participants showed a normal facilitation effect for Rp+ items during retrieval practice. Thus, retrieval-induced forgetting can be eliminated with no impact on the facilitation of practiced items. Similarly, when Penolazzi et al. (2014) applied cathodal transcranial current stimulation to the dorsolateral prefrontal cortex (to inhibit or disrupt brain activity in this region) during retrieval practice, no retrieval-induced forgetting effect occurred. Again, while retrieval-induced forgetting was eliminated in participants who received cathodal transcranial current stimulation, they showed levels of facilitation for Rp+ items that were comparable to those of control participants (Marsh and Anderson, 2022). Thus, there are a multitude of behavioral and neuroscientific studies that convincingly demonstrate that the magnitude of the facilitation effect for Rp+ items is entirely unrelated to the magnitude of the forgetting effect for Rp– items.
Competition dependence
As stated above, another critical prerequisite that must be met for retrieval-induced forgetting to occur is retrieval competition. If there are no interfering or competing nontarget Rp– items present during selective retrieval of target Rp+ items, there should be no memory impairment observed for nontarget Rp– items on the final test.
Relatedly, a multitude of studies have demonstrated that the magnitude of the retrieval-induced forgetting effect is directly related to the degree to which nontarget items compete for retrieval access. Said differently, the resultant retrieval-induced forgetting effect is larger when competing nontarget items are stronger or more accessible during retrieval practice than when nontarget items are weaker or less accessible during retrieval practice. Thus, retrieval-induced forgetting caused by inhibition is competition dependent. For example, Anderson et al. (1994) found that more retrieval-induced forgetting occurred for items high in taxonomic frequency (e.g., FRUIT: ORANGE) than for items lower in taxonomic frequency (e.g., FRUIT: GUAVA). Items that are more readily encountered come to mind more easily and are thus more likely to intrude and disrupt successful retrieval of target items. Murayama et al. (2014) examined the combined effects of 12 studies manipulating taxonomic frequency. Indeed, the retrieval-induced forgetting effect observed for high-frequency items was larger than that observed for low-frequency items. Furthermore, when the final test used appropriately indexed inhibitory processes, significant retrieval-induced forgetting was only observed for high-frequency items, not low-frequency items.
Similarly, Shivde and Anderson (2001) observed significant retrieval-induced forgetting when participants practiced retrieving the less dominant or less frequently encountered meaning of a homonym (ARM: MISSILE) and thus had to suppress the more dominant or frequently encountered semantic association (ARM: SHOULDER). However, no retrieval-induced forgetting effect was found in the opposite condition, when participants practiced the dominant association. The lack of impairment observed in the latter case was due to the fact that limited prior exposure to the less dominant meaning reduced the potential for retrieval competition.
In a particularly clever demonstration of competition dependence, Levy et al. (2007) examined the consequences of practicing words in a foreign language during second language acquisition. For novice Spanish speakers, repeated retrieval practice trials requiring the generation of Spanish words subsequently impaired recall of the corresponding English word. When prompted to generate English words during retrieval-practice however, novices failed to exhibit a retrieval-induced forgetting effect for Spanish words. Because English words were highly accessible competitors, more inhibitory suppression was required to reduce their accessibility and facilitate recall of the Spanish words. In contrast, because novices were only just beginning to acquire Spanish words, these weaker, less dominant items were much less likely to elicit significant retrieval competition and hence less retrieval-induced forgetting. Interestingly, bilingual speakers failed to show a retrieval-induced forgetting effect for English words when prompted to generate the Spanish equivalents during retrieval practice, presumably because their fluency in both languages made the English items less competitive or intrusive during retrieval-practice (but see Runnqvist and Costa, 2012).
Further support for the property of competition dependence comes from studies of retrieval-induced forgetting that have used directed forgetting instructions in an attempt to reduce the degree to which nontarget items interfered during retrieval practice. For instance, when participants were prompted to forget a list of category-exemplar pairs after study and prior to retrieval practice, thereby eliminating the potential for proactive interference from nontarget items during target retrieval, no retrieval-induced forgetting occurred (Storm et al., 2007; Bäuml and Samenieh, 2010). In contrast, when participants were prompted to remember the list, creating the usual degree of interference between target and nontarget items, a retrieval-induced forgetting effect emerged.
Finally, various neuroimaging studies have also provided support for the competition dependence property of retrieval-induced forgetting. As noted above, with respect to the neural substrates of retrieval-induced forgetting, the anterior cingulate cortex and the lateral prefrontal cortex are activated during retrieval practice but not restudy, and this activity correlates with retrieval-induced forgetting. In addition to selective retrieval, these structures are also activated when response conflict is detected and resolved via cognitive control. Thus, activity in these regions should provide a neurological index of interference during retrieval practice and the resolution of such interference via inhibition. In support of this hypothesis, when Kuhl et al. (2007) placed participants in a fMRI scanner and conducted a typical retrieval-induced forgetting experiment utilizing the retrieval practice paradigm, the researchers found that activation in these regions gradually decreased over retrieval practice trials, reflecting a decline in demand for cognitive control and conflict detection as competing items were suppressed. Furthermore, the greater the decline in demands placed on the lateral prefrontal cortex, the greater the resultant retrieval-induced forgetting effect. That is, individuals who exhibited higher amounts of retrieval-induced forgetting showed significant declines in lateral prefrontal cortex activation between the first retrieval practice trial, when they would likely incur the most interference from competitors and the third retrieval practice trial, when suppression of competitors via inhibition would be expected to reduce the amount of interference incurred. In contrast, those who exhibited less retrieval-induced forgetting showed a smaller decline in lateral prefrontal cortex activity between the first and third retrieval-practice trials (Marsh and Anderson, 2022).
Hellerstedt and Johansson (2014) provided additional neuroscientific evidence in support of competition dependence when they collected event-related potentials from participants while they completed a retrieval-induced forgetting task comprised of taxonomically strong and weak exemplars. Consistent with the behavioral results obtained by Anderson et al. (1994), when competing Rp– items were strong or high in taxonomic frequency, more retrieval-induced forgetting was observed than if they were weak or low in taxonomic frequency. In addition, the researchers were able to identify competition-specific event-related potential modulates over anterior regions, and these neural substrates of retrieval competition predicted the degree of retrieval-induced forgetting observed.
Retrieval-induced forgetting of simple motor responses
As the preceding review suggests, there is ample evidence in support of the assertion that retrieval-induced forgetting results from an inhibitory suppression mechanism recruited to resolve interference between competing exemplars during selective retrieval. A reasonable assumption is that a similar retrieval-induced forgetting effect could be observed for motor responses, provided (a) such motor responses were organized in memory in such a way that they shared a common retrieval cue, and (b) presentation of said cue created response competition between associated motor programs.
Tempel and Frings (2013) adapted the retrieval practice paradigm to examine whether inhibition is recruited to resolve interference between motor programs when they share a common category label or retrieval cue (see Figure 4). During the study phase, participants learned a series of two-finger motor sequences, each requiring either the left or right hand. It was assumed that the hand was the effector and would thus serve as the category cue given the well-established finding that people often use hands as discriminators in categorization tasks. For example, regardless of whether their dominate hand is left or right, individuals tend to associate positive valence with their dominate hand. Furthermore, hands have been used to categorize stimuli in binary classification tasks (Lakens et al., 2011). Lastly, it has been demonstrated that people classify small numbers more quickly with their left hand and large numbers more quickly with their right hand (Dehaene et al., 1993). After learning a series of two-finger motor sequences requiring either the left or right hand, half of the motor sequences on one hand were practiced. When all of the motor sequences were tested following a distractor task, clear retrieval-induced forgetting was present. That is, unpracticed motor sequences on the practiced hand (Rp– items) were recalled worse than unpracticed items on the unpracticed hand (Nrp items). Tempel and Frings (2014a) replicated this finding in a subsequent experiment. Participants first learned 12 movement sequences using a joystick. Each movement was randomly assigned to either the left or right hand. During retrieval practice, participants practiced half of the sequences corresponding to either the left or right hand. The final test results revealed significant retrieval-induced forgetting of the remaining unpracticed sequences from the practiced hand. In addition, these results again demonstrated that hands were used as category cues and thus organized associated memories in such a way that they later gave rise to interference. Reppa et al. (2013) observed a similar pattern of results. That is, when participants learned a sequence of touch-screen responses associated with 10 novel objects, significant retrieval-induced forgetting resulted when the sequences were organized by hand only, button only, or both hand and button.
Figure 4
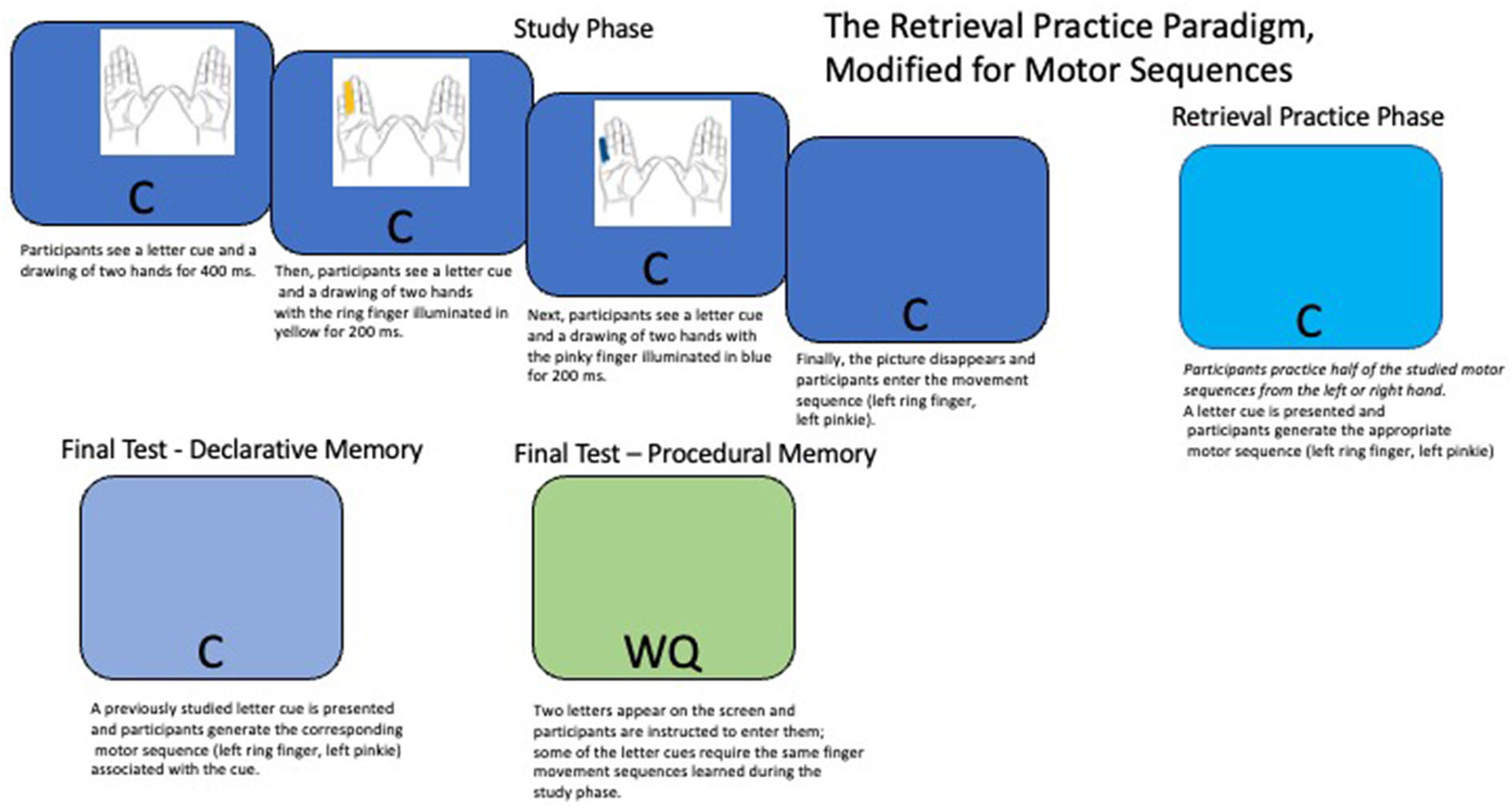
The retrieval practice paradigm, modified for motor sequences. During the study phase, participants learn a series of two finger motor sequences that are initiated by the left or right hand (e.g., left ring finger, left pinkie). On a given trial, participants first see a letter cue (e.g., C) and a drawing of two hands for 400 milliseconds. Then, the first finger in the sequence is illuminated in yellow for 200 milliseconds, and the second finger in the sequence is illuminated in blue for an additional 200 milliseconds. Finally, the picture disappears and participants see the letter cue by itself, at which point they are instructed to execute the appropriate motor sequence. During retrieval practice, again a letter cue is presented by itself, and participants are instructed to execute the motor sequence associated with that letter cue. During the final test phase, participants again generate the appropriate motor sequence when presented with the studied cue (declarative memory test), before subsequently being presented with a novel two letter cue (e.g., WG) and being instructed to type in the sequence of letters on a keyboard. Some of the two letter cues require the same finger movement sequences as those studied previously (procedural memory test). Figure adapted from Tempel and Frings (2014a).
Recently, Schmidt and Tempel (2022) extended this pattern of results to include eye movements. The researchers utilized a gaze direction task, teaching participants to fix their eyes on a grid of squares presented on a computer screen. They learned a series of eye movements requiring them to look to the left or the right, and subsequently practiced a subset of the movements corresponding to one direction. When participants were tested on all of the eye-movement sequences, significant retrieval-induced forgetting occurred for unpracticed sequences in the practiced direction. Thus, a variety of motor responses, in multiple modalities, are subject to retrieval-induced forgetting.
Interestingly, Tempel and Frings (2017) demonstrated that retrieval-induced forgetting of motor sequences is modality specific. Participants learned a series of motor sequences corresponding to the index finger on either the left or right hand. In the subsequent retrieval practice phase, participants either practiced half of the motor sequences associated with one hand or completed a visual search task related to the same sequences. That is, participants in the motor retrieval practice condition executed the appropriate sequence of movements in response to a letter cue. Participants in the visual retrieval practice condition simply clicked on the correct squares corresponding to a visual depiction of the task. Retrieval-induced forgetting of motor sequences only persisted in the motor retrieval practice condition. Consistent with transfer-appropriate processing effects in verbal memory (Morris et al., 1977), when the final test was manipulated to either match or not match the task required during retrieval practice, retrieval-induced forgetting only occurred when the two tasks were matching. How did the authors explain these results? In order for the interference that underlies retrieval-induced forgetting to occur, the type of representation activated during retrieval practice must also be activated at test. Said differently, when representations of one modality are activated, retrieval only elicits interference between representations of that same modality. Thus, the after-effects of such retrieval competition, namely, the suppression of competing exemplars, are only evident when the final test requires participants to activate representations within the same modality. Practice of motor actions elicits interference and subsequent suppression of related motor representations; practice of visual representations elicits interference and subsequent suppression of related visual representations.
Tempel and Frings (2014b) extended their results to include not just cued recall, a measure of declarative memory, but an implicit letter dyads task, a measure of procedural memory (see Figure 4). Participants again studied several two-finger motor sequences on either the left or right hand (e.g., LEFT HAND: INDEX FINGER, PINKIE; RIGHT HAND: RING FINGER, MIDDLE FINGER). Participants would first see a letter cue (e.g., C) followed by a picture of two hands with the appropriate fingers colored in yellow and blue respectively. Participants subsequently practiced half of the motor sequences from one hand. Replicating previous findings, on the final test of all the motor sequences, when cued to elicit the motor sequence given the studied cue, recall of the unpracticed items from the practiced hand was below the rate of recall for unpracticed sequences from the unpracticed hand. Participants were also asked to enter two letter sequences (e.g., WQ), a subset of which required the same finger movements as the previously learned motor sequences. Thus, while the first task required participants to retrieve previously studied cue-movement sequence associations and was therefore a test of explicit memory, the second task presented a novel cue that was not previously studied, thus providing an implicit measure of memory for the motor sequences. Execution of these movements was significantly impaired as compared to those that were not previously learned. That is, when the researchers compared reaction times for Rp- motor sequences to reaction times for Nrp or baseline motor sequences, Rp- reaction times were significantly longer. Thus, inhibition was recruited to resolve interference in both declarative and procedural memory. The fact that significant retrieval-induced forgetting was observed in the letter sequences task also suggests that suppression of motor sequences is cue independent, since memory for the motor sequences was tested using cues other than those presented during the initial study phase. Tempel and Frings (2015) also demonstrated significant retrieval-induced forgetting of motor sequences on explicit and implicit memory tests when the motor sequences were cued not by effector but rather by the direction of the movement. Participants learned a series of mouse click sequences, half of which prompted them to move forward and half of which prompted them to move backward. During retrieval practice, participants repeated half of the sequences in one direction (either backward or forward). When participants were explicitly cued to generate the remaining unpracticed movements in the same direction, significant retrieval-induced forgetting occurred. The same pattern occurred on an implicit test prompting participants to execute the same movements in response to novel letter cues. Thus, there is converging evidence to suggest that retrieval dynamics operate similarly in procedural and declarative memory, and that retrieval-induced forgetting of motor responses is cue independent.
In addition to providing compelling evidence that retrieval-induced forgetting of motor sequences is cue independent, Tempel and Frings (2016) demonstrated that retrieval-induced forgetting of motor sequences is both strength independent and retrieval specific. Participants again learned a series of two-finger motor sequences carried out by either the left or right hand. Subsequently, participants either (a) practiced executing half of the motor sequences of one hand in response to a letter cue or (b) restudied those same motor sequences. While significant retrieval-induced forgetting was observed in the retrieval practice condition, none was observed in the extra study condition. Thus, strengthening of motor sequences in the absence of retrieval competition was not sufficient to induce forgetting.
Retrieval-induced forgetting of motor sequences is also competition dependent. Tempel et al. (2016) investigated whether retrieval-induced forgetting of motor sequences would persist if retrieval practice took place prior to the study phase. In such a case, we would expect no need for the resolution of interference between motor programs, as there are no previously learned motor sequences to intrude perseveratively and create response competition. Indeed, no retrieval-induced forgetting was observed when retrieval practice preceded the study phase, demonstrating that interference during retrieval is a necessary component of retrieval-induced forgetting of motor sequences. Tempel et al. (2016) conducted a second experiment in which they implemented a directed forgetting instruction similar to the one utilized by Storm et al. (2007). As predicted, when participants were prompted to forget the recently learned motor sequences, this forget instruction prevented the requisite interference or response competition from occurring. In the absence of such interference, there was no need for inhibitory suppression, thus no retrieval-induced forgetting occurred. Collectively, all of these results provide compelling support for the inhibitory account. That is, they strongly suggest that inhibition is recruited during selective retrieval of motor responses to ameliorate interference created by competing motor programs.
Retrieval-induced forgetting of complex motor responses
All of the preceding examples reference instances where participants learned and subsequently forgot a subset of fairly simplistic motor responses. Is memory for more complex motor actions susceptible to retrieval-induced forgetting? Koutstaal et al. (1999) were the first to demonstrate retrieval-induced forgetting for complex actions, in samples of both older and younger adults. During the study phase, participants were prompted to perform a series of everyday actions (e.g., retrieving a tissue from the Kleenex box, tracing a boomerang). During retrieval practice, participants were shown a series of photographs depicting other people performing a subset of the actions, and they were asked to think back to when they performed the actions themselves during the initial study phase. On a final test in which participants were asked to perform all of the actions they learned, unpracticed actions from practiced object cues were impaired relative to unpracticed actions from unpracticed object cues.
Sharman (2011) investigated whether retrieval-induced forgetting would occur for complex actions that were either bizarre or familiar. During the learning phase, participants executed the actions using a specific object (Bizarre: balancing a pencil on a cup; Familiar: sharpening a pencil). They subsequently practiced executing a subset of those actions in response to a picture of the object cue. Retrieval-induced forgetting persisted for both bizarre and familiar actions. Sharman also demonstrated that retrieval-induced forgetting of complex actions was retrieval specific. Some participants executed a subset of the actions during retrieval practice, while others observed someone else doing them. Significant retrieval-induced forgetting only persisted when participants performed the actions themselves, due to the fact that passive observation does not include an active retrieval component.
Tempel et al. (2015) demonstrated retrieval-induced forgetting for complex actions in the form of dance moves. During the study phase, participants learned 4 dances, each comprised of 8 steps. Participants watched a video of an instructor performing each sequence of dance moves. While watching, participants were instructed to execute the movements along with the instructor and memorize the name corresponding to each dance. In the subsequent practice phase, participants performed two of the four dances they initially learned. They watched another video in which the instructor demonstrated the first two moves, before prompting participants to execute the remaining six dance moves themselves. Following retrieval practice and a distractor task, participants were tested on all four dances (without a demonstration first). Significant retrieval-induced forgetting of the dance moves occurred, but only when participants participated in retrieval practice (not extra study), thus indicating that retrieval-induced forgetting of complex actions is strength independent and retrieval specific.
Inhibition as a domain general mechanism in cognitive and motor tasks
As noted above, there is ample evidence to suggest that retrieval-induced forgetting extends beyond the domain of verbal memory to include motor responses across multiple modalities and that this effect persists for representations stored in both declarative and procedural memory. Furthermore, inhibition is clearly implicated as the primary mechanism by which interference between competing motor programs is resolved in service of facilitating successful retrieval. More broadly, inhibition can thus be best understood as a domain general mechanism that operates similarly in cognitive and motor tasks. It should be noted however, that this assertion is not new. Thirteen years ago, Anderson and Weaver (2009) argued that inhibition of motor actions and inhibition that occurs during selective retrieval operate via similar mechanisms necessitated by situations that require response override. According to the authors, response override refers to “one's ability to stop strong or habitual responses” (Anderson and Weaver, 2009, p. 153). In cognitive tasks such as the retrieval practice paradigm, response override becomes necessary in order to “retrieve a target memory trace in the face of many competing memories” (Anderson and Weaver, 2009, p. 153). In motor tasks, “a stimulus may initiate multiple-compatible physical actions, only one of which should guide behavior,” thus contextually-inappropriate actions must be suppressed in order to facilitate execution of the desired behavior (Anderson and Weaver, 2009, p. 153). In support of this assertion, Schilling et al. (2014) found that successful response override of motor actions via the stop signal task reliably predicted retrieval-induced forgetting of verbal material when the final test used appropriately indexed inhibitory processes. That is, participants completed the stop signal task as well as a standard retrieval-induced forgetting task with category-exemplar pairs. In the stop signal task, participants are shown a series of shapes (i.e., squares, circles) and each shape corresponds to a particular key on the keyboard. On 75% of trials, participants are prompted to press the appropriate key when the corresponding shape appears on the screen. On 25% of the trials however, participants are prompted to withold their response. Schilling and colleagues found that participants who showed superior performance during the stop signal task, as indexed by faster reaction times, also showed greater retrieval-induced forgetting. Other cognitive paradigms presumed to require response override also generalize to the inhibition of motor responses. For example, Schmidt et al. (2022) utilized the think-no-think paradigm to (Anderson and Green, 2001) assess the consequences of intentionally suppressing a subset of motor sequences on their later recall. In the initial learning phase, participants studied a series of motor sequences and learned to associate each sequence with a letter cue. During the subsequent think-no-think phase, on think trials participants were trained to think of the associated motor sequence when presented with the studied cue. On no-think trials, participants were expressly instructed to suppress the associated motor sequence and any thoughts related to its execution when presented with the associated cue. When participants' memory was tested for all of the motor sequences, clear suppression-induced forgetting was present. That is, relative to baseline motor sequences that were not featured during the think-no think phase, sequences subject to suppression were more poorly recalled and executed more slowly. Thus, similar to the pattern of results observed in the retrieval practice paradigm, inhibitory suppression of motor responses in the think-no think paradigm reduced the accessibility of the motor representations stored in declarative memory and hindered their execution in procedural memory.
General discussion
Movement is a ubiquitous part of the human experience. The average adult will walk 75,000 miles–or the equivalent of 216,262,500 steps in their lifetime (Leach, 2022). Even sedentary individuals who spend most of their time seated in front of a computer will click their mouse 428 times per day, and complete 6,000 keystrokes on their keyboard within a 24-hour period (Buchannon, 2022). Unsurprisingly, movement sequences that are repeatedly generated or practiced will become more easily recallable, and the time required to execute these sequences will decrease (Tempel and Frings, 2019). However, such benefits of retrieval practice are not without costs, provided similar motor sequences interfere with one another and are thus subject to interference or retrieval competition. In such cases, unpracticed sequences from practiced categories are vulnerable to forgetting. However, such retrieval-induced forgetting is arguably adaptive and essential for proper functioning of the memory system, as the inhibitory control processes governing such forgetting allow for competing, nontarget motor responses to be suppressed so as to facilitate efficient recall and execution of target responses (Bjork and Bjork, 1988; Bjork, 1989; Anderson, 2003; Anderson and Weaver, 2009).
Consistent with this assertion, a multitude of experiments reviewed above suggest that motor sequences with features in common such as the hand initiating the movement, the direction of the movement, or the object pertaining to the movement, are vulnerable to interference dynamics in both declarative and procedural memory provided the movement sequences are organized and stored via this shared cue. Furthermore, because such forgetting has reliably been shown to be cue independent, strength independent, retrieval specific, and competition dependent, the primary mechanism responsible for this impairment is likely inhibition, and not associative interference. Finally, converging evidence from the literature on inhibitory control in cognitive and motor tasks points to the critical role of response override in facilitating successful selective retrieval in declarative memory and suppression of inappropriate actions in procedural memory.
Aside from adding to the literature on retrieval dynamics in declarative and procedural memory (Tempel and Frings, 2013, 2014a,b, 2015, 2016, 2017, 2019; Schmidt et al., 2022), what are the real-world implications of these collective findings? While the preponderance of studies investigating retrieval-induced forgetting of verbal, perceptual, and motor information have been conducted with contrived laboratory tasks, and thus future research should be aimed at investigating these processes in more naturalistic settings, the insights reviewed in this paper may be applicable to athletes, musicians, and anyone who is highly motivated to learn and properly execute sophisticated motor actions. While trained body movements that are subject to repeated practice are likely to be strengthened, as indexed by superior recall and more efficient execution (Tempel and Frings, 2019), associated body movements that are not practiced are vulnerable to forgetting. For example, a pianist may begin a piece of music by pressing a series of keys on the piano together in sequence to create a chord. Given the fact that there are many songs that start with the same chord, this sequence of keystrokes may become associated with numerous pieces of music. If the pianist practices one song many times, in preparation for a recital for example, such repeated practice may cause forgetting of other songs initiated by the same sequence of notes (Tempel and Frings, 2013). This impairment for related movements will only occur if training or practice procedures include an active retrieval component; observing others executing the proper movements or restudying the movement sequence is sufficient to strengthen the movement's representation in memory without triggering interference and the subsequent suppression of related movements (Sharman, 2011; Tempel et al., 2015, 2016). However, practicing without an active retrieval component may be ill-advised or impractical in many cases. In instances where active retrieval of a subset of motor sequences is necessary, adding a training component in which athletes or musicians review related, unpracticed motor sequences could reduce or abolish retrieval-induced forgetting entirely. There is ample evidence to suggest that relearning related information after retrieval practice eliminates retrieval-induced forgetting (Storm et al., 2008; Storm and Soares, 2022). While the effect of relearning on retrieval-induced forgetting has only been investigated with verbal materials, given the fact that motor responses are subject to similar dynamics in memory there are good reasons to expect similar results for motor sequences. Therefore, if there is a concern that related motor actions may become vulnerable to retrieval-induced forgetting after other movements are practiced, simple reexposure to these movements may prevent any forgetting. However, the different consequences of each type of practice should be considered when developing training protocols (Tempel and Frings, 2015; Tempel et al., 2015).
Statements
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Anderson J. R. (1983). A spreading activation theory of memory. J. Verbal Learn. Verbal Behav.22, 261–295. 10.1016/S0022-5371(83)90201-3
2
Anderson M. (2003). Rethinking interference theory: executive control and the mechanisms of forgetting. J. Mem. Lang.49, 415–445. 10.1016/j.jml.2003.08.006
3
Anderson M. C. Bell T. (2001). Forgetting our facts: the role of inhibitory processes in the loss of propositional knowledge. J. Exp. Psychol. Gen. 130, 544–570.
4
Anderson M. C. Bjork E. L. Bjork R. A. (2000a). Retrieval-induced forgetting: evidence for a recall-specific mechanism. Psychon. Bull. Rev.7, 522–530. 10.3758/BF03214366
5
Anderson M. C. Bjork R. A. Bjork E. L. (1994). Remembering can cause forgetting: retrieval dynamics in long-term memory. J. Exp. Psychol. Learn. Mem. Cogn. 20, 1063. 10.1037/0278-7393.20.5.1063
6
Anderson M. C. Green C. (2001). Suppressing unwanted memories by executive control. Nature410, 366–369. 10.1038/35066572
7
Anderson M. C. Green C. McCulloch K. C. (2000b). Similarity and inhibition in long-term memory: evidence for a two-factor theory. J. Exp. Psychol. Learn. Mem. Cogn. 26, 1141–1159. 10.1037/0278-7393.26.5.1141
8
Anderson M. C. Spellman B. A. (1995). On the status of inhibitory mechanisms in cognition: memory retrieval as a model case. Psychol. Rev. 102, 68–100.
9
Anderson M. C. Weaver C. (2009). Inhibitory control over action and memory, in Encyclopedia of Neuroscience, ed SquireL. (Amsterdam: Elsevier), 153–163. 10.1016/B978-008045046-9.00421-6
10
Aslan A. Bäuml K. H. Pastötter B. (2007). No inhibitory deficit in older adults' episodic memory. Psychol. Sci.18, 72–78. 10.1111/j.1467-9280.2007.01851.x
11
Bajo M. T. Gómez-Ariza C. J. Fernandez A. Marful A. (2006). Retrieval-induced forgetting in perceptually driven memory tests. J. Exp. Psychol. Learn. Mem. Cogn. 32, 1185–1194. 10.1037/0278-7393.32.5.1185
12
Bäuml K. H. (2002). Semantic generation can cause episodic forgetting. Psychol. Sci.13, 356–360. 10.1111/1467-9280.00464
13
Bäuml K. H. T. Samenieh A. (2010). The two faces of memory retrieval. Psychol. Sci. 21, 793–795.
14
Bjork E. L. Bjork R. A. (1988). On the adaptive aspects of retrieval failure in autobiographical memory, in Practical Aspects of Memory: Current Research and Issues, Vol. 1. Memory in Everyday Life, eds GrunebergM. M.MorrisP. E.SykesR. N. (Hoboken, NJ: John Wiley and Sons), 283–288.
15
Bjork R. A. (1989). Retrieval inhibition as an adaptive mechanism in human memory, in Varieties of Memory and Consciousness: Essays in Honour of Endel Tulving, eds RoedigerH. L.CraikF. I. M. (Hillsdale, NJ: Erlbaum), 309–330.
16
Brown A. S. Bracken E. Zoccoli S. Douglas K. (2004). Generating and remembering passwords. Appl. Cogn. Psychol.18, 641–651. 10.1002/acp.1014
17
Buchannon J. (2022). Click Here to Improve Your Motor Skills. Science: Brain and Behavior. Available online at: https://www.science.org/content/article/click-here-improve-your-motor-skills#:~:text=But%20with%20daily%20practice%E2%80%94the,this%20amazing%20feat%20of%20dexterity (accessed October 26, 2022).
18
Buchli D. R. Storm B. C. Bjork R. A. (2016). Explaining retrieval-induced forgetting: a change in mental context between the study and restudy practice phases is not sufficient to cause forgetting. Q. J. Exp. Psychol.69, 1197–1209. 10.1080/17470218.2015.1076866
19
Camp G. Pecher D. Schmidt H. G. (2005). Retrieval-induced forgetting in implicit memory tests: the role of test awareness. Psychon. Bull. Rev.12, 490–494. 10.3758/BF03193793
20
Camp G. Pecher D. Schmidt H. G. (2007). No retrieval-induced forgetting using item-specific independent cues: evidence against a general inhibitory account. J. Exp. Psychol. Learn. Mem. Cogn. 33, 950–958. 10.1037/0278-7393.33.5.950
21
Camp G. Pecher D. Schmidt H. G. Zeelenberg R. (2009). Are independent probes truly independent?J. Exp. Psychol. Learn. Mem. Cogn. 35, 934–942. 10.1037/a0015536
22
Campbell J. I. D. Thompson V. A. (2012). Retrieval-induced forgetting of arithmetic facts. J. Exp. Psychol. Learn. Mem. Cogn. 38, 118–129. 10.1037/a0025056
23
Carroll M. Campbell-Ratcliffe J. Murnane H. Perfect T. (2007). Retrieval-induced forgetting in educational contexts: monitoring, expertise, text integration, and test format. Eur. J. Cogn. Psychol.19, 580–606. 10.1080/09541440701326071
24
Cipresso P. Serino S. Villani D. Repetto C. Sellitti L. Albani G. et al . (2012). Is your phone so smart to affect your state? An exploratory study based on psychophysiological measures. Neurocomputing84, 23–30. 10.1016/j.neucom.2011.12.027
25
Ciranni M. A. Shimamura A. P. (1999). Retrieval-induced forgetting in episodic memory. J. Exp. Psychol. Learn. Mem. Cogn. 25, 1403. 10.1037/0278-7393.25.6.1403
26
Collins A. M. Loftus E. F. (1975). A spreading-activation theory of semantic processing. Psychol. Rev.82, 407–428. 10.1037/0033-295X.82.6.407
27
Dehaene S. Bossini S. Giraux P. (1993). The mental representation of parity and number magnitude. J. Exp. Psychol. Gen.122, 371–396.
28
Erdman M. R. Chan J. C. (2013). Providing corrective feedback during retrieval practice does not increase retrieval-induced forgetting. J. Cogn. Psychol.25, 692–703. 10.1080/20445911.2013.790389
29
Hanslmayr S. Staudigl T. Aslan A. Bäuml K. H. (2010). Theta oscillations predict the detrimental effects of memory retrieval. Cogn. Affect. Behav. Neurosci.10, 329–338. 10.3758/CABN.10.3.329
30
Hellerstedt R. Johansson M. (2014). Electrophysiological correlates of competitor activation predict retrieval-induced forgetting. Cereb. Cortex24, 1619–1629. 10.1093/cercor/bht019
31
Huddleston E. Anderson M. C. (2012). Reassessing critiques of the independent probe method for studying inhibition. J. Exp. Psychol. Learn. Mem. Cogn. 38, 1408–1418. 10.1037/a0027092
32
Hulbert J. C. Shivde G. Anderson M. C. (2012). Evidence against associative blocking as a cause of cue-independent retrieval-induced forgetting. Exp. Psychol. 59, 11–21.
33
Johnson S. K. Anderson M. C. (2004). The role of inhibitory control in forgetting semantic knowledge. Psychol. Sci.15, 448–453. 10.1111/j.0956-7976.2004.00700.x
34
Koessler S. Engler H. Riether C. Kissler J. (2009). No retrieval-induced forgetting under stress. Psychol. Sci.20, 1356–1363. 10.1111/j.1467-9280.2009.02450.x
35
Koutstaal W. Schacter D. L. Johnson M. K. Galluccio L. (1999). Facilitation and impairment of event memory produced by photograph review. Mem. Cognit.27, 478–493. 10.3758/BF03211542
36
Kuhl B. A. Dudukovic N. M. Kahn I. Wagner A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. Nat. Neurosci.10, 908–914. 10.1038/nn1918
37
Lakens D. Schneider I. K. Jostmann N. B. Schubert T. W. (2011). Telling things apart: The distance between response keys influences categorization times. Psychol. Sci. 22, 887–890.
38
Leach N. (2022). How Many Steps Does the Average Person Take in a Lifetime? Alliance Work Partners. Availalble online at: https://www.awpnow.com/main/2019/09/27/fun-fact-friday-steps-over-a-lifetime (accessed October 26, 2022).
39
Levy B. J. McVeigh N. D. Marful A. Anderson M. C. (2007). Inhibiting your native language: the role of retrieval-induced forgetting during second-language acquisition. Psychol. Sci.18, 29–34. 10.1111/j.1467-9280.2007.01844.x
40
Little J. L. Storm B. C. Bjork E. L. (2011). The costs and benefits of testing text materials. Memory. 19, 346–359.
41
MacLeod M. (2002). Retrieval-induced forgetting in eyewitness memory: forgetting as a consequence of remembering. Appl. Cogn. Psychol.16, 135–149. 10.1002/acp.782
42
MacLeod M. D. Saunders J. (2005). The role of inhibitory control in the production of misinformation effects. J. Exp. Psychol. Learn. Mem. Cogn. 31, 964. 10.1037/0278-7393.31.5.964
43
Macrae C. N. MacLeod M. D. (1999). On recollections lost: When practice makes imperfect. J. Pers. Soc. Psychol. 77, 463–473.
44
Marsh L. C. Anderson M. C. (2022). Inhibition as a cause of forgetting, in Oxford Handbook of Human Memory, Volume 1: Foundations, eds KahanaM. J.WagnerA. D. (Oxford: Oxford University Press), 1–42.
45
Maxcey A. M. De Leon V. Janakiefski L. Megla E. Stallkamp S. Torres S. B et al . (2021). Induced forgetting of pictures across shifts in context. J. Exp. Psychol. Hum. Percept. Perform. 47, 1091–1102. 10.1037/xhp0000928
46
Maxcey A. M. Woodman G. F. (2014). Forgetting induced by recognition of visual images. Vis. Cogn.22, 789–808. 10.1080/13506285.2014.917134
47
Morris C. D. Bransford J. D. Franks J. J. (1977). Levels of processing versus transfer appropriate processing. J. Verb. Learn. Verb. Behav. 16, 519–533.
48
Murayama K. Miyatsu T. Buchli D. Storm B. C. (2014). Forgetting as a consequence of retrieval: a meta-analytic review of retrieval-induced forgetting. Psychol. Bull.140, 1383–1409. 10.1037/a0037505
49
Pearman S. Thomas J. Naeini P. E. Habib H. Bauer L. Christin N. et al . (2017). Let's go in for a closer look: Observing passwords in their natural habitat, in Proceedings of the 2017 ACM SIGSAC Conference on Computer and Communications Security (ACM), 295–310.
50
Penolazzi B. Stramaccia D. F. Braga M. Mondini S. Galfano G. (2014). Human memory retrieval and inhibitory control in the brain: beyond correlational evidence. J. Neurosci.34, 6606–6610. 10.1523/JNEUROSCI.0349-14.2014
51
Perfect T. J. Stark L.-J. Tree J. J. Moulin C. J. A. Ahmed L. Hutter R. et al . (2004). Transfer appropriate forgetting: the cue-dependent nature of retrieval-induced forgetting. J. Mem. Lang.51, 399–417. 10.1016/j.jml.2004.06.003
52
Pilar D. R. Jaeger A. Gomes C. F. A. Stein L. M. (2012). Passwords usage and human memory limitations: a survey across age and educational background. PLoS ONE7, e51067. 10.1371/journal.pone.0051067
53
Raaijmakers J. G. W. Jakab E. (2013). Rethinking inhibition theory: on the problematic status of the inhibition theory for forgetting. J. Mem. Lang.68, 98–122. 10.1016/j.jml.2012.10.002
54
Reppa I. Worth E. R. Greville W. J. Saunders J. (2013). The representation of response effector and response location in episodic memory for newly acquired actions: evidence from retrieval-induced forgetting. Acta Psychol.143, 210–217. 10.1016/j.actpsy.2013.03.007
55
Runnqvist E. Costa A. (2012). Is retrieval-induced forgetting behind the bilingual disadvantage in word production?Biling.: Lang. Cogn.15, 365–377. 10.1017/S1366728911000034
56
Rupprecht J. Bäuml K.-H. T. (2016). Retrieval-induced forgetting in item recognition: retrieval specificity revisited. J. Mem. Lang.86, 97–118. 10.1016/j.jml.2015.09.003
57
Rupprecht J. Bäuml K.-H. T. (2017). Retrieval-induced versus context-induced forgetting: can restudy preceded by context change simulate retrieval-induced forgetting?J. Mem. Lang.93, 259–275. 10.1016/j.jml.2016.10.006
58
Saunders J. MacLeod M. D. (2002). New evidence on the suggestibility of memory: the role of retrieval-induced forgetting in misinformation effects. J. Exp. Psychol. Appl.8, 127–142. 10.1037/1076-898X.8.2.127
59
Schilling C. J. Storm B. C. Anderson M. C. (2014). Examining the costs and benefits of inhibition in memory retrieval. Cognition133, 358–370. 10.1016/j.cognition.2014.07.003
60
Schmidt M. Anderson M. C. Tempel T. (2022). Suppression-induced forgetting of motor sequences. Cognition230, 105–292. 10.1016/j.cognition.2022.105292
61
Schmidt M. Tempel T. (2022). Retrieval-induced forgetting of eye movements. PsyArXiv [Preprint]. 10.31234/osf.io/kjqhd
62
Sharman S. J. (2011). Retrieval-induced forgetting of performed and observed bizarre and familiar actions. Exp. Psychol.58, 361–369. 10.1027/1618-3169/a000103
63
Shaw J. S. Bjork R. A. Handal A. (1995). Retrieval-induced forgetting in an eyewitness-memory paradigm. Psychon. Bull. Rev.2, 249–253. 10.3758/BF03210965
64
Shivde G. Anderson M. C. (2001). The role of inhibition in meaning selection: Insights from retrieva linduced forgetting. Understanding endogenous emotions View project, in On the Consequences of Meaning Selection: Perspectives on Resolving Lexical Ambiguity, ed GorfeinD. S. (American Psychological Association), 175–190.
65
Staudigl T. Hanslmayr S. Bäuml K. H. T. (2010). Theta oscillations reflect the dynamics of interference in episodic memory retrieval. J. Neurosci.30, 11356–11362. 10.1523/JNEUROSCI.0637-10.2010
66
Storm B. C. Angello G. Buchli D. R. Koppel R. H. Little J. L. Nestojko J. F. et al . (2015). A review of retrieval-induced forgetting in the contexts of learning, eyewitness memory, social cognition, autobiographical memory, and creative cognition. Psychol. Learn. Motiv.62, 141–194. 10.1016/bs.plm.2014.09.005
67
Storm B. C. Bjork E. L. Bjork R. A. (2007). When intended remembering leads to unintended forgetting. Q. J. Exp. Psychol.60, 909–915. 10.1080/17470210701288706
68
Storm B. C. Bjork E. L. Bjork R. A. (2008). Accelerated relearning after retrieval-induced forgetting: the benefit of being forgotten. J. Exp. Psychol. Learn. Mem. Cogn. 34, 230. 10.1037/0278-7393.34.1.230
69
Storm B. C. Bjork E. L. Bjork R. A. Nestojko J. F. (2006). Is retrieval success a necessary condition for retrieval-induced forgetting?Psychon. Bull. Rev.13, 1023–1027. 10.3758/BF03213919
70
Storm B. C. Levy B. J. (2012). A progress report on the inhibitory account of retrieval-induced forgetting. Mem. Cognit.40, 827–843. 10.3758/s13421-012-0211-7
71
Storm B. C. Soares J. S. (2022). Relearning can eliminate the effect of retrieval-induced forgetting. Psychol. Res.86, 1725–1736. 10.1007/s00426-021-01601-7
72
Tempel T. Aslan A. Frings C. (2016). Competition dependence of retrieval-induced forgetting in motor memory. Mem. Cognit.44, 671–680. 10.3758/s13421-015-0578-3
73
Tempel T. Frings C. (2013). Resolving interference between body movements: retrieval-induced forgetting of motor sequences. J. Exp. Psychol. Learn. Mem. Cogn. 39, 1152–1161. 10.1037/a0030336
74
Tempel T. Frings C. (2014a). Interference within hands: retrieval-induced forgetting of left and right hand movements. Acta Psychol.148, 1–5. 10.1016/j.actpsy.2014.01.003
75
Tempel T. Frings C. (2014b). Forgetting motor programmes: retrieval dynamics in procedural memory. Memory22, 1116–1125. 10.1080/09658211.2013.871293
76
Tempel T. Frings C. (2015). Categorization by movement direction: retrieval-induced forgetting of motor sequences grouped by motion features. Q. J. Exp. Psychol.68, 473–486. 10.1080/17470218.2014.945098
77
Tempel T. Frings C. (2016). How motor practice shapes memory: retrieval but not extra study can cause forgetting. Memory24, 903–915. 10.1080/09658211.2015.1059858
78
Tempel T. Frings C. (2017). Retrieval-induced forgetting is retrieval-modality specific: evidence from motor memory. Cognition162, 143–152. 10.1016/j.cognition.2017.02.005
79
Tempel T. Frings C. (2018). Feedback increases benefits but not costs of retrieval practice: retrieval-induced forgetting is strength independent. Psychon. Bull. Rev.25, 636–642. 10.3758/s13423-018-1450-9
80
Tempel T. Frings C. (2019). Testing enhances motor practice. Mem. Cognit.47, 1270–1283. 10.3758/s13421-019-00932-6
81
Tempel T. Loran I. Frings C. (2015). Dancing your moves away: how memory retrieval shapes complex motor action. J. Exp. Psychol. Appl.21, 300–312. 10.1037/xap0000052
82
Veling H. van Knippenberg A. (2004). Remembering can cause inhibition: retrieval-induced inhibition as cue independent process. J. Exp. Psychol. Learn. Mem. Cogn. 30, 315–318. 10.1037/0278-7393.30.2.315
83
Verde M. F. (2012). Retrieval-induced forgetting and inhibition, in Psychology of Learning and Motivation, Vol. 56 (Amsterdam: Elsevier), 47–80. 10.1016/B978-0-12-394393-4.00002-9
84
Weller P. D. Anderson M. C. Gómez-Ariza C. J. Bajo M. T. (2013). On the status of cue independence as a criterion for memory inhibition: evidence against the covert blocking hypothesis. J. Exp. Psychol. Learn. Mem. Cogn. 39, 1232–1245. 10.1037/a0030335
85
Williams C. C. Zacks R. T. (2001). Is retrieval-induced forgetting an inhibitory process?Am. J. Psychol.114, 329. 10.2307/1423685
86
Wimber M. Alink A. Charest I. Kriegeskorte N. Anderson M. C. (2015). Retrieval induces adaptive forgetting of competing memories via cortical pattern suppression. Nat. Neurosci.18, 582–589. 10.1038/nn.3973
87
Wimber M. Bäuml K. H. Bergström Z. Markopoulos G. Heinze H. J. Richardson-Klavehn A. et al . (2008). Neural markers of inhibition in human memory retrieval. J. Neurosci.28, 13419–13427. 10.1523/JNEUROSCI.1916-08.2008
Summary
Keywords
retrieval-induced forgetting, inhibition, movement, procedural memory, declarative memory
Citation
Buchli DR (2023) Retrieval-induced forgetting in declarative and procedural memory: Inhibitory suppression resolves interference between motor responses. Front. Cognit. 1:1081181. doi: 10.3389/fcogn.2022.1081181
Received
27 October 2022
Accepted
28 December 2022
Published
18 January 2023
Volume
1 - 2022
Edited by
Petra Jansen, University of Regensburg, Germany
Reviewed by
Jerwen Jou, The University of Texas Rio Grande Valley, United States; David Ian Anderson, San Francisco State University, United States
Updates
Copyright
© 2023 Buchli.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dorothy R. Buchli ✉ buchli_dr@mercer.edu
This article was submitted to Cognition and Movement, a section of the journal Frontiers in Cognition
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.