 Gwendolyn Wirobski
Gwendolyn Wirobski Friederike Range
Friederike Range Laura Bonnet3
Laura Bonnet3 Zsófia Virányi
Zsófia Virányi- 1Domestication Lab, Konrad Lorenz Institute of Ethology, University of Veterinary Medicine Vienna, Vienna, Austria
- 2Comparative Cognition Group, Université de Neuchâtel, Faculty of Sciences, Neuchâtel, Switzerland
- 3Messerli Research Institute, University of Veterinary Medicine Vienna, Medical University of Vienna, University of Vienna, Vienna, Austria
Human–dog interactions are often ascribed to selection for increased cooperativeness during domestication, implying voluntary participation and mutual benefit. Alternatively, building on a possible selection for higher deference and submissive inclinations toward more dominant individuals (deferential behavior or super-dominance hypotheses), human–dog interactions might rather rely on following the human lead. Here, we compare three components of tractability, i.e., the willingness to comply with human directions, of young, hand-raised wolves and dogs using a set of behavioral tests to investigate their 1) trainability (coming when called with and without a distraction, following commands), 2) responsiveness to persuasion (fetch and retrieve a ball), and 3) readiness to accept coercion (brushing, muzzling, lying on the back while being petted). Based on the hypothesis that dogs show higher compliance with human directions than wolves, we predicted that dogs would respond to being called and follow learned commands faster than wolves. Dogs should also be more responsive to persuasion and accept coercion more readily than wolves. In contrast to our predictions, we found no difference between latencies of coming when called or when asked to sit, but wolves took longer to lie down on command than dogs. In the fetch and retrieve task, dogs retrieved the ball more often than wolves at 9 weeks of age, but not at 6 and 8 weeks, and it was more difficult to get the ball back from the wolves than from the dogs, at least when the partner was a stranger. Surprisingly, there were no significant differences between wolves and dogs in the brushing and the muzzling tests. In the final test, dogs accepted coercive handling when lying on their backs and remained lying down as long as without restraint, whereas the opposite was the case for wolves. In conclusion, the findings support the deferential behavior hypothesis whereby dogs more readily accept the leading role of the human partners, regardless of relationship strength and interaction style (persuasive vs. coercive), but wolves comply only if there is mutual trust and the interaction does not involve physical restrictions.
1 Introduction
Humans like to see interactions with domesticated animals, especially companion animals and pets, as cooperative, which implies a voluntary, mutually beneficial, purposeful, and coordinated interaction between individuals of the same or different species whereby individuals gain something together that they could not have achieved alone (Duguid and Melis, 2020). But how much do these interactions really qualify as cooperative? May they not rather be described as compliance, where the non-human animal conforms to a request, command, or set of rules, often without resistance or objection? In the context of behavior, it typically involves an individual or animal following instructions or expectations set by another, either voluntarily or because of external influence, such as training, persuasion, or coercion. Regarding dogs, compliance has also been referred to as tractability (Ujfalussy et al., 2020) (see Box 1 for definitions).
Box 1. Key terms and their definitions as used in this manuscript.
Tractability: The quality of being easily handled, controlled, and directed by humans as well as accepting limitations (“willingness to comply”) as an outcome of either coercion, persuasion, or training.
Trainability: The ability to learn an action over the course of repeated interactions and building up routines that are not easily influenced by distractions (“how much a certain behavior can be put on cue”).
Persuasion: The process of influencing someone’s behavior without duress but rather using enticement and/or positive rewards.
Coercion: The use of physical force, threat or social pressure to initiate or maintain a certain behavior.
According to Frank’s information‐processing hypothesis (Frank, 1980, 2011), dogs are predisposed to attend rapidly to human cues and translate them into action. This suggests that the domestication process has shaped dogs to efficiently process and act on human signals, resulting in higher trainability compared to similarly socialized wolves (Frank, 1980, 2011). While wolves and dogs share the basic capacity to learn from humans, dogs’ increased orientation toward human leadership yielded shorter latencies to follow commands (Vasconcellos et al., 2016). This difference could be attributed to wolves being less interested in following human directions than dogs in training interactions. Alternatively, it may be grounded in wolves’ greater vigilance and environmental awareness, which reflect adaptive strategies rather than reduced interest in humans. Indeed, orienting toward humans in such contexts may involve a trade-off with significant ecological implications for a non-domesticated species.
Alternatively, compliance could also be established through persuasion by the human partner. Dogs’ heightened human-directed social motivation, or hypersociability (vonHoldt et al., 2017; Boada and Wirobski, 2024; Bentosela et al., 2016), could have made them especially receptive to being influenced through enticement and/or positive rewards. By contrast, wolves may require stronger incentives or a deeper bond with the handler before deferring to human guidance. However, previous studies showed that persuasive strategies may work equally well in wolves and dogs if rewards of the right kind and of high enough incentive are picked (Klinghammer and Goodmann, 1987; Range et al., 2012). Nevertheless, the relationship with the human partner may be a crucial factor determining how willingly animals will follow human directions in a certain situation. Animals may be more willing to comply with people they know and trust than with strangers, especially in somewhat uncomfortable situations or when putting themselves in a vulnerable position. In well-known, routine situations such as simple obedience tasks, the relationship with the partner might not matter as much (Klinghammer and Goodmann, 1987; Kerepesi et al., 2015).
More recently, Frank’s thoughts have been developed further as the deferential behavior (Virányi and Range, 2014; Range et al., 2019) and the super-dominance hypotheses (Wynne, 2021). Using coercion, the human partner may leverage the power asymmetry in the relationship to “force” the animal’s compliance through punishment, physical restraint, or withholding of resources such as food. Unlike wolves, dogs might be more inclined to accept this form of control because of selection for deference during their domestication process (Frank, 2011; Range et al., 2019) and a higher propensity to accept more dominant individuals (Range et al., 2015; Wynne, 2021). Dominancy, in this case, may be understood not as a personality trait or outcome of repeated competition, but rather suggest that dogs have evolved a greater tolerance for being guided or controlled by socially more assertive or directive partners (e.g., humans), compared to wolves, in line with the idea of deference to leadership (Range et al., 2019). This makes dogs safer, more controllable, and more malleable social partners than wolves.
Experimental evidence for the deferential behavior hypothesis comes from studies using standardized paradigms, such as the cooperative string-pulling task, where hand-raised wolves and dogs were tested with human partners on their ability to solve two apparatuses in a coordinated fashion (Range et al., 2019). While human–dog and human–wolf dyads were similarly successful, the main difference was that wolves took the lead toward the second apparatus significantly more often and contested the human partner more often (rope stealing) than dogs (Range et al., 2019). Hence, wolves—given adequate socialization with humans—are as successful in cooperation with humans as domestic dogs, but their interaction styles differ. While dogs appear to preferentially comply with human leadership, wolves act more independently and sometimes challenge their human partners (Klinghammer and Goodmann, 1987). Furthermore, a recent study compared hand-raised wolves’ and dogs’ behavioral and physiological reactions to being invited to a petting session by differently bonded human partners. Results showed that while dogs spent more time with human partners than wolves, they also displayed more stress-related behaviors than wolves and had increased cortisol levels, suggesting that they followed the “invitation” despite being uncomfortable in the situation (Wirobski et al., 2021). While together these results open the possibility that dogs, at least sometimes, interact with people out of compliance (Kaminski, 2009) rather than their own motivation to do so. To date, whether dogs and wolves differ in their trainability, receptiveness toward persuasion, or acceptance of coercion, particularly at a very young age, has received very little attention.
A previous study by Ujfalussy et al. (2020) used a test battery including retrieving an object, responding to being called, sitting on request, allowing themselves to be brushed, and walking while wearing a muzzle, to gauge overall tractability in hand-raised wolf and dog pups. The authors assumed that all these tasks required the animals to be tractable, in that they had to “comply with and follow human actions, and either tolerate some physical restriction or give access to a target object.” However, how the animals were “asked” to comply differed between tasks. In at least one task, the experimenter initially tried to engage the animals gently, hence measuring their receptiveness to persuasion (fetch and retrieve task). They reported that dogs outperformed wolves when gentle encouragement was used (fetch and retrieve) and that wolves were slower to obey trained commands (calling, sitting). In restraint tasks (brushing), wolves initially showed more resistance, though this difference vanished by 4 months, when changes in housing limited comparability.
Applying the same tasks, we set out—not to replicate the previous study by Ujfalussy et al.—but to distinguish between trainability, persuasion, and mild coercion, and complement it by including an additional “lying on the back” test to directly assess willingness to accept human leadership in a vulnerable position. We tested young wolves and dogs, hand-raised and kept under similar conditions throughout the whole study period, to identify traits that may be deeply rooted in the domestication process and more likely to reflect evolved predispositions. Specifically, the animals participated in six different tests: 1) calling in the absence of food with a familiar or unfamiliar person and calling in the presence of inaccessible food, 2) fetching and retrieving a paper ball with a familiar person or with a stranger, 3) following commands (sit and lie down), 4) being brushed, 5) walking on a leash while wearing a muzzle, and 6) lying on the back while being petted by a familiar person. Importantly, in the lying on the back test, the method of getting the animals to lie on their backs differed between the experimenters, allowing for testing for an effect of a “persuasive” style (verbal encouragement) vs. a more “coercive” style (mild physical force) on the animals’ behavioral reactions.
We hypothesized that dogs and wolves would differ in how they react to tasks using coercion or persuasion by a human partner, but that they would not differ in their trainability (success in responding to learned commands) (see Table 1 for a detailed overview of all the tests and predictions). Briefly, in the tasks testing acceptance of coercion (brushing, muzzling), we predicted that dogs would be more willing to be handled (higher success, shorter latencies, longer durations) than wolves, while displaying fewer agonistic (biting into the brush or getting rid of the muzzle attempts). We further predicted that experimenters would manage to get dogs to lie down on their backs faster than wolves, that dogs would stay lying longer, struggle less, and show fewer biting attempts than wolves, mediated by the style of the experimenter, with dogs accepting a more coercive method than wolves, but fewer differences when the experimenter uses the persuasive method.
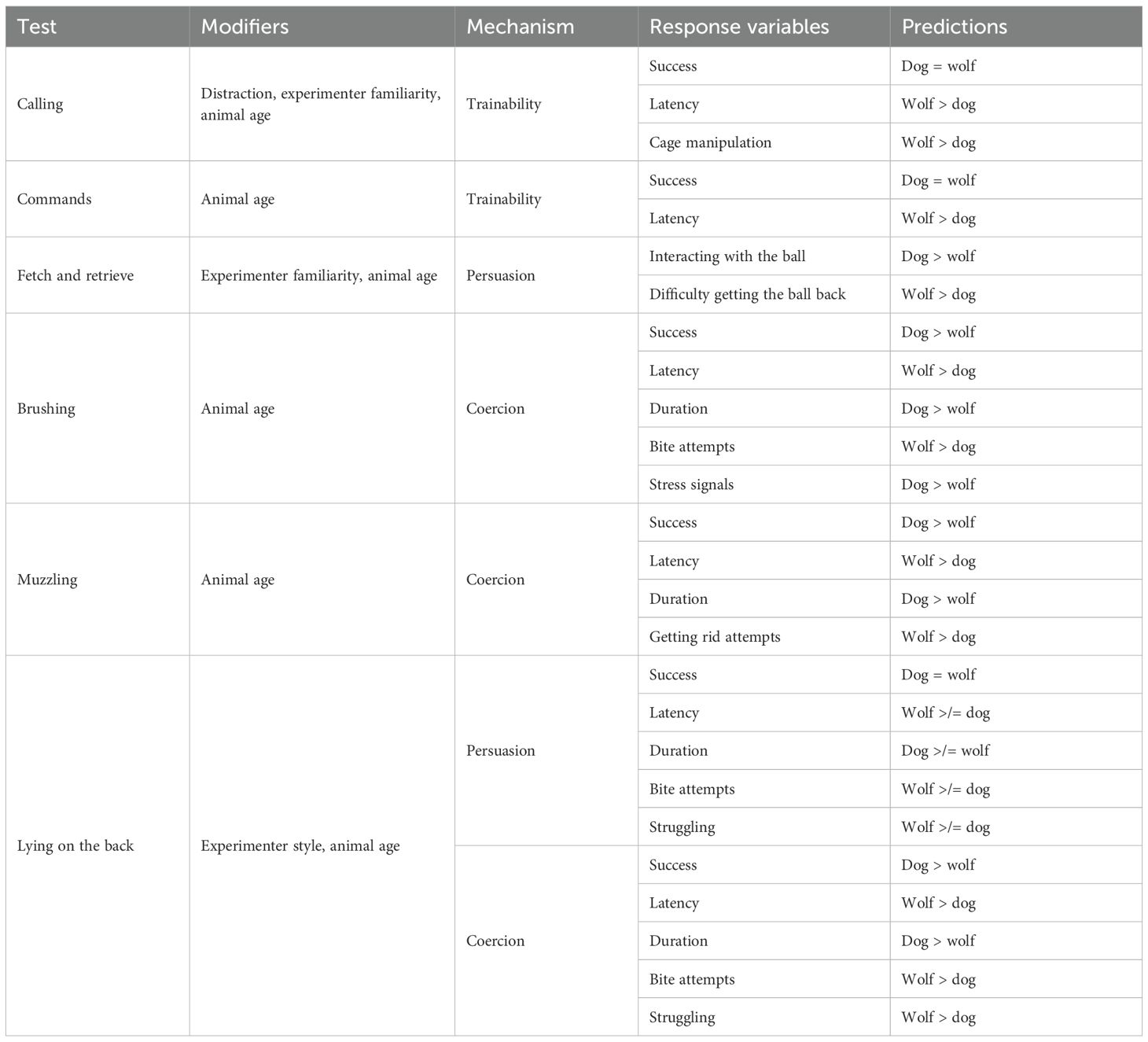
Table 1. Overview of the tests, modifiers, mechanisms, response variables, and predictions.
2 Materials and methods
2.1 Ethical statement
In Austria, no special permission was needed for the use of animals in socio-cognitive studies at the time the studies were conducted (Tierversuchsgesetz 2012 – TVG 2012). The relevant committee that allows research to run without special permission regarding animals is the Tierversuchskommission am Bundesministerium für Wissenschaft und Forschung (Austria). This research complies with the current Austrian laws on animal protection. All methods were carried out in accordance with relevant guidelines and regulations.
2.2 Subjects
A total of 18 hand-raised wolf and 37 dog puppies raised and housed under similar conditions at the Core Facility Wolf Science Center (CF-WSC), Ernstbrunn, Austria, participated in the tests. Due to changes in availability, the number of subjects involved in each test differed (see Supplementary Tables S1, S2 for sample sizes per test). The dog puppies were all mongrels and came from various animal shelters in Hungary or were born at the CF-WSC. All wolves originated from the US (North-American wolves) and came from zoos or private farms located in the USA, Canada, Switzerland, or Austria. All animals participating in this study were hand-raised in peer groups by a group of professional animal trainers, researchers, and students starting when they were approximately 10 days old. All animals were bottle-fed and spent 24 h a day, 7 days a week with their hand-raisers and their conspecific peers until 4–5 months old, when they were integrated into packs of adult animals living in large outdoor enclosures (for details, please see Range and Virányi, 2014).
2.3 Experimental procedures
Wolf pups were tested between 2008 and 2012 and dog pups between 2009 and 2014. All tests were conducted indoors and recorded (see Supplementary Video 1). The animals’ hand-raisers were considered familiar experimenters, whereas researchers who had never met the animals before the test were considered strangers. If a test was conducted twice or multiple tests were run on the same day, the same person acted as a stranger in all these tests. The calling tests started at the age of 3 weeks, with no prior training before the data collection, and so did the commands task. There was no prior training for the fetch and retrieve test. In the muzzle test, the animals were already used to being walked on a leash (but not while wearing the muzzle), whereas brushing and lying on the back were novel situations.
2.3.1 Task 1: calling (without distraction)
The handler (a hand-raiser) entered the experimental room holding the pup in her arms. She positioned herself on one side of the room, 2–3 m from the experimenter, facing her. Then, the handler put the pup down on the floor in front of her, restraining the pup with her hands. The experimenter crouched down, clapped her hands twice, and started to call the pup in a high-pitched voice. At this point, the handler let the pup go and did not interact with it anymore. The experimenter went on calling the pup until it came close enough to be petted and picked up or until 60 s had elapsed. The test was composed of two successive trials; in the second trial, the experimenter and the handler changed their places and repeated the same procedure in the opposite direction. The task was repeated five times at the ages of 3, 4, 5, 6, and 8 weeks. This differed from the methodology in the work of Ujfalussy et al. (2020), who tested the animals until the age of 24 weeks. However, some of their animals were rehomed at 8–10 weeks old, and others at 24 weeks old, which could have affected the results. In the present study, we were most interested in the earlier differences between wolves and dogs, as they may better inform us about inherent differences related to domestication. While the experimenter was familiar at the ages of 3, 4, and 5 weeks, she was a stranger at 6 and 8 weeks of age. At 3 weeks of age, the distance between the handler and experimenter was 2 m; afterward, it was 3 me for all the tests.
2.3.1.1 Calling (with distraction)
A handler placed the pup in front of a closed metal wire cage containing a bowl of food (the food was, thus, inaccessible for the pup) (Figure 1). As soon as the pup started to sniff the food and/or tried to get access to it by manipulating the cage, a second experimenter familiar with the animal started to call the pup in a high-pitched voice. She continued calling until the pup approached her, and she could pet it or until 60 s had elapsed. The task was conducted once at the age of 9 weeks.
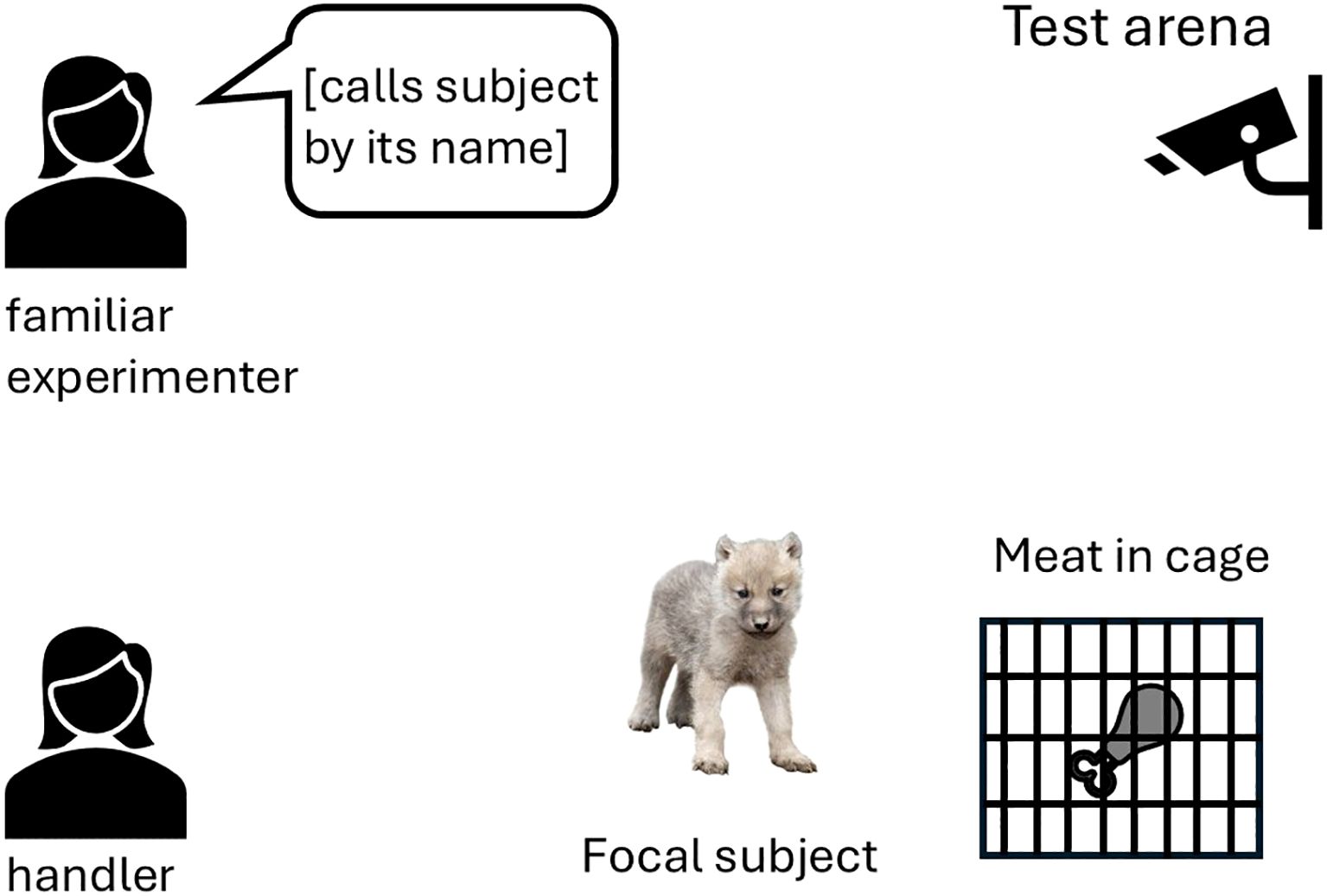
Figure 1. Setup of the calling test with distraction (inaccessible food in a metal wire cage).
2.3.2 Task 2: commands
The experimenter, a familiar person, holding a piece of food in her hand, tried to make the animal sit two times using either a verbal command, a hand signal, or both, to guide the action of the animal. If the animal sat down, the experimenter also asked it to lie down two times, again using either a verbal command or hand signal to direct the animal. Each successful command was rewarded with a piece of food and verbal praise from the experimenter. The procedure was carried out repeatedly with each animal without a leash at the ages of 7–24 weeks.
2.3.3 Task 3: fetch and retrieve
An experimenter moved a paper ball in front of the pup, inviting it to play with it. As soon as the pup’s attention was focused on the ball, she threw it away. The ball was thrown approximately 1 m away from the puppy. If the pup followed the ball and took it in its mouth, the experimenter asked it to bring it back by calling the pup for up to 15 s. If the pup returned to the experimenter, she tried to retrieve the ball; if not, the experimenter went to the puppy to retrieve the ball. In order to get the ball back from the pup, the experimenter first reached her hand toward the pup and was calling it in a friendly voice, then, if this did not work, she followed the pup and picked the ball up if the pup dropped it, or if needed, she gently held the pup’s body and inserted her finger into its mouth in order to retrieve the ball. The experimenter was also allowed to use another ball to distract the pup if she failed with the methods cited before or if the pup showed aggressive behavior. This procedure was repeated three times on each of two occasions at 6 and 8 weeks of age with an unfamiliar experimenter and again at the age of 9 weeks with a familiar experimenter.
2.3.4 Task 4: brushing
The familiar experimenter tried to brush the fur of the animal continuously for a duration of 30 s with a dog grooming brush while keeping the animal on a loose leash. A brushing attempt was considered successful if the animal accepted the brushing for a minimum of 3 s without struggling or following the brush in an attempt to grab it. This test was carried out twice with the animals at the ages of 12–13 and 16–17 weeks.
2.3.5 Task 5: muzzling
The familiar experimenter put a standard muzzle on the animal’s snout and started walking with the animal on the leash for up to 60 s, verbally encouraging the animal to come along. No treats were used in the tests analyzed for this study. The outcome was considered successful if the animal moved without attempting to get rid of the muzzle (scratching, head shaking, rubbing against the experimenter) for at least 3 s. The animals were already used to leash walking before the muzzle was introduced for the first time during the test sessions. This test was carried out twice with the animals aged 16–17 and 24 weeks.
2.3.6 Task 6: lying on the back
The familiar experimenter tried to get the animal to lie on its back for up to 2 min. The experimenter could attempt to achieve this by using either verbal encouragement (touching/petting the animals but without restraining or directing their movement or preventing them from getting up) or physical restraint (using mild force to get the animal on its side or back by holding its legs and gently pushing down). The attempt was considered successful when the animals’ back or side touched the floor while being petted by the experimenter. Each experimenter tested a wolf and a dog at least once. This test was carried out twice with the animals aged 12–13 and 16–17 weeks.
2.4 Behavioral coding
BORIS (Friard and Gamba, 2016) and Loopy software (http://loopb.io, loopbio gmbh, Vienna, Austria) were used for behavioral coding of the test videos (see Supplementary Table S3 for a detailed ethogram). Interobserver reliability (IOR) coding was done by a second rater on 20% of the videos for each test. IOR outcomes ranged from good to excellent. Detailed results were as follows: calling test: “calling success” 100% agreement, “calling latency” R = 0.99 (Pearson correlation coefficient); commands: “latency to sit” ICC = 0.82 (interclass correlation coefficient), “latency to lie down” ICC = 0.99; fetch and retrieve test: “follow ball” 95% agreement, “contact with the ball” 82% agreement, “ball in mouth” 86% agreement, “retrieve ball” 94% agreement, “difficulty taking the ball from the animal” 100% agreement; brushing test: “duration of acceptance” ICC = 0.85, “bite attempts” R = 0.88, “stress signals” R = 0.88; muzzling: “latency to accept” R = 0.99, “duration of acceptance” R = 0.97, “scratch off attempt” R = 0.93; lying on the back test: “latency to lie down” R = 0.96, “duration of lying down” R = 0.99, “bite attempts” R = 0.93.
2.5 Statistical data analyses
2.5.1 Calling without distraction (models 1a and 1b)
We investigated whether the individual established contact with the caller (no/yes; thereafter contact success; model 1a) and the latency until this contact was established (contact latency; model 1b) by fitting two generalized linear mixed models (GLMM; Baayen, 2008). The contact success model was fitted with binomial error structure and logit link function (McCullagh, 1989), and the contact latency model was a Cox proportional hazards (a.k.a. survival model). In the contact latency model, we set the latency to the duration of the experiment when no contact was established and defined the “hazard” as the incidence of contact establishment. All models included the same fixed effect structure, namely, the three-way interaction between species (dog or wolf), subject age (thereafter “age”), and familiarity with the experimenter (familiar/stranger; thereafter “familiarity”) and all two-way interactions and main effects this encompassed, and the trial number. Their random effect structures were also identical regarding the random intercepts included, which were those of the individual animal, the litter it originated from, and the identity of the experimenter.
2.5.1.1 Calling with distraction (models 2a to 2b)
Here, we investigated two response variables and hence fitted two models. These were cage manipulation (no/yes; binomial error structure and logit link function; model 2a) and latency until the animal established contact with the familiar caller (survival model; model 2b). All models comprised species, age, and their interaction as fixed effects. We included the interaction between species and age and the random intercepts for the identity of the experimenters and the litter (no random slope was theoretically identifiable, and each individual animal contributed only one data point).
2.5.2 Commands (models 3a to 3c)
To examine the behavior of the animals after a “sit” or a “lie” command, we fitted three models (all but one subject succeeded in following the sit command, so no model was fitted for sitting success). The behavior we considered after the sit command was the latency until the animal sat (model 3a; Gaussian error distribution and identity link). The behaviors we considered after the lie command were whether it was successful (no/yes; model 3b; binomial error structure and logit link function), and the latency until the animal lay down (survival model, 3c). All models comprised species and the interaction with age as the key test predictor with fixed effects and included trial number as a further control predictor. All models also comprised the same random intercept effects of the ID of the animal, litter, and experimenter ID (see Supplementary Table S1 for further details).
2.5.3 Fetch and retrieve (models 4a to 4e)
In the fetch and retrieve test, we coded five different behaviors and hence fitted five models. Four of these (following the ball, 4a; ball contact, 4b; ball in the pup’s mouth, 4c; and retrieving the ball, 4d) we fitted with binomial error structure and logit link function. The difficulty of taking the ball from the pup by the experimenter was rated on an ordinal scale, and hence, we fitted a cumulative logit link model (model 4e), as required for such data. The fixed effects were the same in all models and included species, age, familiarity with the experimenter, and trial number. Since we were not only interested in species effects but also how these depended on age and familiarity with the experimenter, we also included the interactions between these latter two predictors and species. We included random intercept effects for the identity of the litter, the animal, and the experimenter and all theoretically identifiable random slopes (see Supplementary Table S1 for further details).
2.5.4 Brushing (models 5a to 5e)
To investigate the behavior of wolf and dog pups in response to being brushed by a familiar experimenter, we fitted five models. These were the experimenter’s success of brushing the animal (no/yes; binomial error structure and logit link function; model 5a); latency to success, i.e., the animals’ acceptance of being brushed (survival model; 5b); duration of accepting to be brushed as a proportion of the total duration of the brushing attempt (model with beta error structure and logit link function; 5c); and finally, the number of biting attempts (zero-inflated negative binomial model; 5d) and avoidance or stress-related behaviors (count model with Poisson error distribution; 5e) during the brushing attempts. All models comprised species as the test predictor, age as a fixed effect, and random effects of animal ID, experimenter ID, and dyad ID. We included only tests by experimenters who tested at least one dog and one wolf.
2.5.5 Muzzling (models 6a to 6d)
For the muzzling test, we looked at the experimenter’s success of putting the muzzle on the animal (no/yes; binomial error structure and logit link function; model 6a); the latency to success, i.e., the animals’ acceptance of the muzzle and walking calmly without attempting to get rid of it (survival model; 6b); the duration of accepting as a proportion of the total duration of the test (model with beta error structure and logit link function; 6c); and finally, the number of attempts of getting rid of the muzzle (scratching muzzle with front limb; count model with Poisson error distribution; 6d). All models comprised species as the test predictor, age as a fixed effect, and random effects of animal ID, experimenter ID, and dyad ID. We included only tests by experimenters who tested at least one dog and one wolf.
2.5.6 Lying on the back (models 7a to 7d)
To test the behavior of animals in the lying on the back test, we examined the following: the experimenter’s success of getting the animal to lie on its back (no/yes; binomial error structure and logit link function; model 7a), the latency thereof (survival model; 7b), and the duration of lying on the back as the proportion of the total duration of the test (model with beta error structure and logit link function; 7c). Finally, we investigated the number of biting attempts during the test (count model with Poisson error distribution followed by a negative binomial model to address zero inflation; 7d). Models on success, latency, and proportions comprised the interaction between species and method (i.e., whether the experimenter used physical restraint or verbal encouragement) as the test predictor. Age was included as a control predictor. All models included random effects of animal and experimenter ID, as well as dyad ID. We included only tests by experimenters who tested at least one dog and one wolf.
2.6 General considerations
We fitted all models in R (version 4.2.1; R Core Team, 2022) using either the function glmmTMB (models with beta error distribution) of the package glmmTMB (version 1.0.0; Brooks et al., 2017), coxme (proportional hazards models) of the package coxme (version 2.2-14; Therneau, 2019), clmm (model with an ordinal response) of the package ordinal (version 2019.12-10; Christensen, 2019), or glmer of the package lme4 (version 1.1-21; Bates et al., 2015). To avoid overconfident models and keep the type I error rate at the nominal level of 0.05, we included random effects into all models (Schielzeth and Forstmeier, 2009; Barr et al., 2013). Initially, we included all theoretically identifiable random slopes and parameters for the correlations among random intercepts and slopes, but due to convergence or identifiability problems, and for computational feasibility, we had to exclude most of the correlation parameters (see Supplementary Tables S1, S2 for the random slopes and correlation parameters included in the final full models). Full models were tested against their null model, which lacked the test predictor but retained control and random effects (Forstmeier and Schielzeth, 2011) using a likelihood ratio test (Dobson, 2002). Outputs of all full-null model comparisons can be found in Supplementary Table S4. If the full-null model comparison revealed no significance, no further tests were performed, but following a significant full-null model comparison, the significance of individual fixed effects was tested by dropping them from the model one at a time (Barr et al., 2013). In case the highest order interaction in each full model did not reveal significance, we removed it to infer about the lower order terms it encompassed. We proceeded correspondingly with non-significant two-way interactions. To further investigate the effect in case of a significant interaction term, we used estimated marginal means (package emmeans version 1.10.3). To ease model convergence and achieve easier interpretable model estimates, we z-transformed age and trial number (if present in a model) to a mean of zero and a standard deviation of one prior to fitting the models. In case of factors (i.e., categorical predictor variables) being present as random slopes in a model, we manually dummy-coded and then centered them before including them in the random slopes part. For all models fitted using glmer, we estimated using the optimizer “bobyqa.” For models fitted using the function glmer or glmmTMB, we determined confidence intervals of model estimates and fitted values by means of a parametric bootstrap (N = 1,000 bootstraps; functions bootMer and simulate of the packages lme4 and glmmTMB, respectively; we are not aware of an option to get confidence intervals for models fitted using the function clmm or coxme). For all models, we established model stability by dropping levels of random effects one at a time, fitting the full model to each of the derived subsets, and then comparing the estimates derived with those obtained from the model based on all data. Full results for all models can be found in Table 2. Results of stability checks can be found for all full models with a significant full-null model comparison in the Supplementary Information (Supplementary Tables S5–S14).
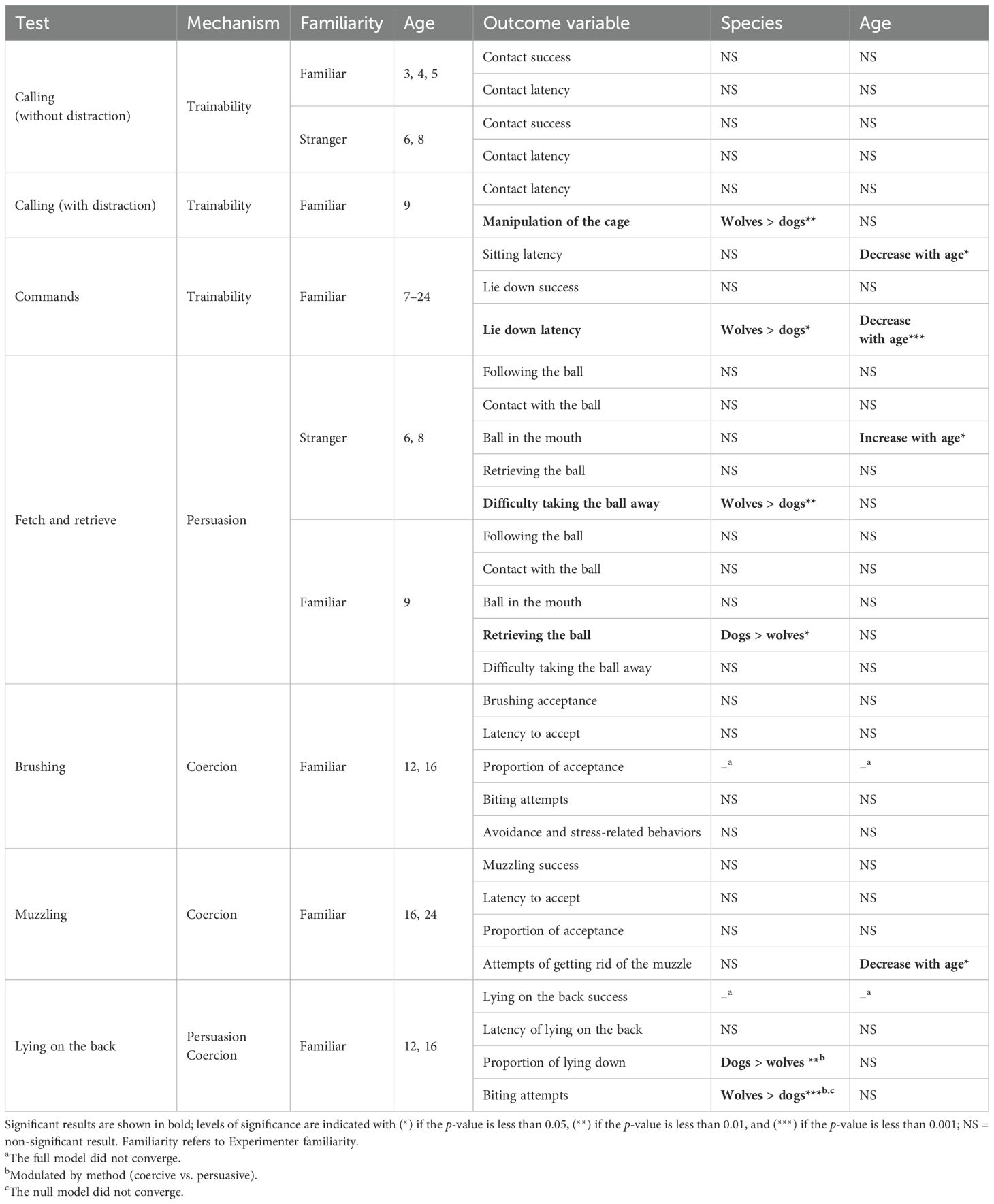
Table 2. Summary of all results for species and age of the subjects (in weeks) in each test.
3 Results
3.1 Calling without distraction (models 1a and 1b)
When investigating whether wolves and dogs differed in their propensity to come when being called either by a familiar person or a stranger, the full-null model comparison did not reveal any significant differences in success (model 1a, Supplementary Table S4) or in latency (model 1b; Supplementary Table S4) to come to the experimenter, suggesting that wolves and dogs were similarly obedient across ages and independent of the person who called them.
3.2 Calling with distraction (models 2a and 2b)
To further investigate whether wolves and dogs differ in responsiveness to calling with a distraction present, the full-null model comparisons revealed significance for cage manipulation (model 2a), but not for latency to contact (model 2b; Supplementary Table S4), again suggesting that responsiveness to being called did not differ between the two species. After removal of the non-significant interaction between species and age from the cage manipulation model 2a (for full results, see Supplementary Tables S5, S6), we found a clearly significant difference in that wolves were more likely to manipulate the cage than dogs (Figure 2; Table 2).
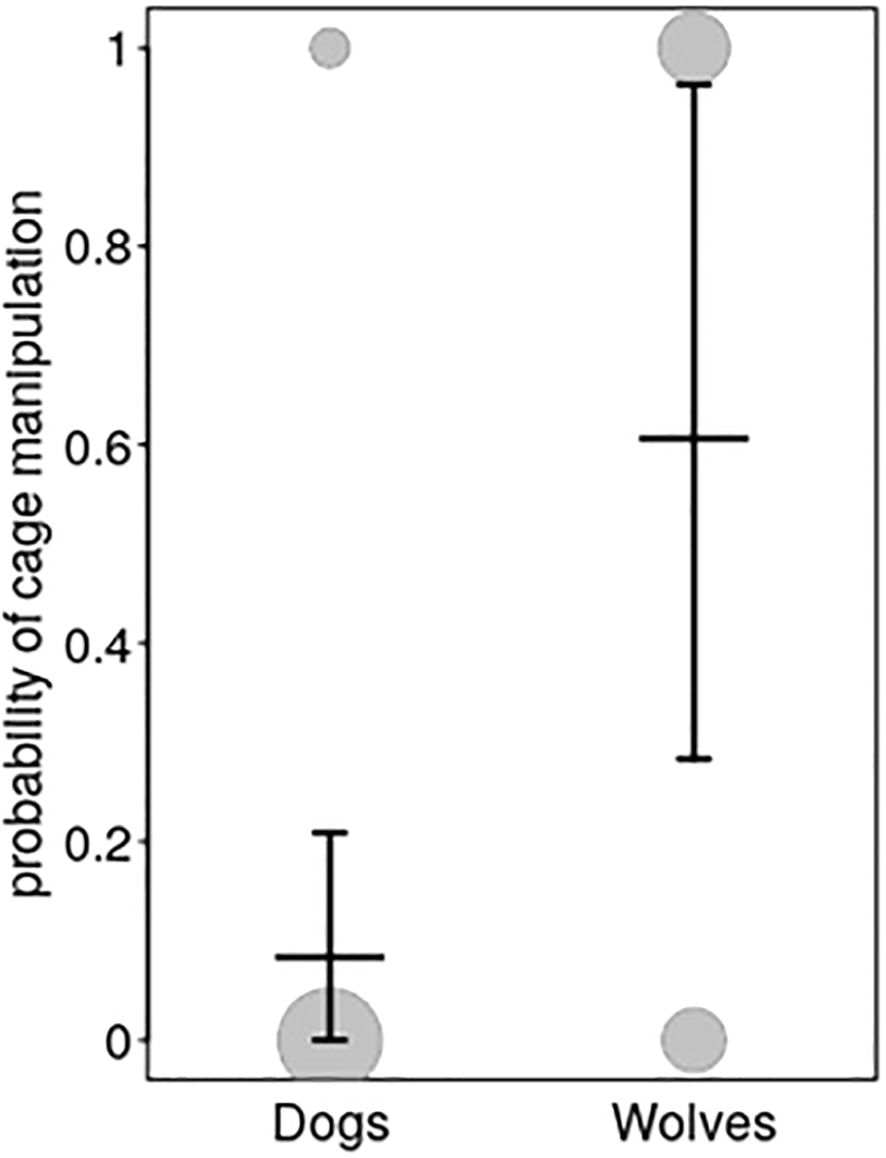
Figure 2. Probability of cage manipulation (model 2a) of dogs and wolves during the calling test in the presence of food. Dots show the observed response, whereby the area of the dots depicts the number of experiments in which dogs and wolves did and did not manipulate the cage (N = 3 to 21). Horizontal lines with error bars depict the fitted model and its confidence limits, with age being centered to a mean of zero.
3.3 Commands (models 3a to 3c)
All but one subject (wolf, 17 weeks) successfully followed the sit command during the trials. There was no difference between wolves and dogs in the latency to sit (model 3a; Figure 3; Supplementary Table S4). Furthermore, wolves and dogs did not differ in their success to lie down after the lie command (model 3b; Supplementary Table S4). However, the reduced model fitted to investigate lying down latency (model 3c) revealed significant main effects of species and age (Supplementary Tables S7, S8). Specifically, wolves took longer than dogs to lie down following the command (Figure 3; Table 2), and both followed the command faster with increasing age (Table 2).
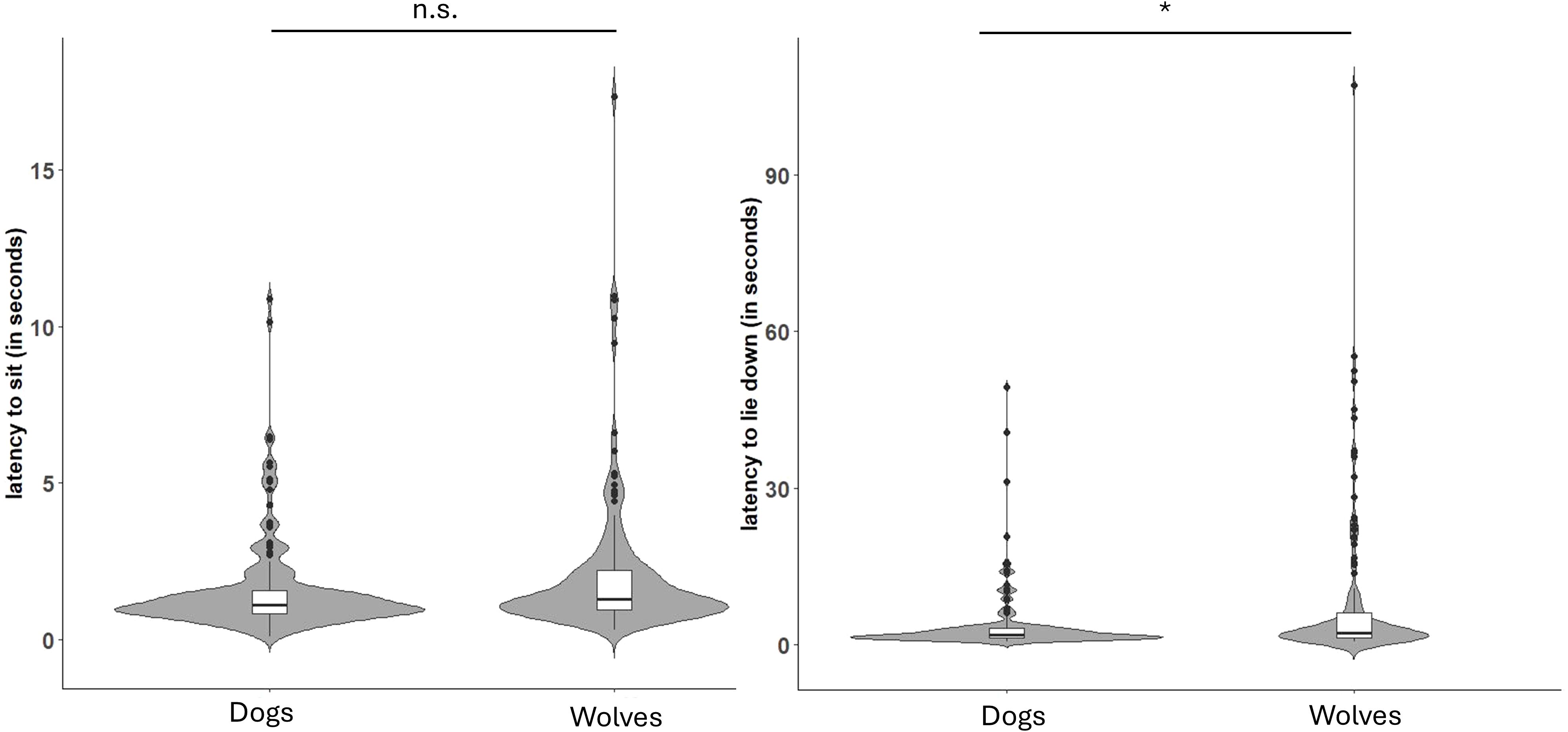
Figure 3. Latencies of dogs and wolves to sit (model 3a, on the left) and lie down (model 3c, on the right) on command. Boxes with thick horizontal lines depict quartiles and medians of the response. The asterisk (*) indicates a p-value of less than 0.05, and n.s. indicates a non-significant result.
3.4 Fetch and retrieve (models 4a to 4e)
In the fetch and retrieve test, we investigated whether wolves and dogs would differ in their motivation to follow a thrown ball, pick it up, return it to the experimenter when called, and give it up when the experimenter tried to take it. Of the full-null model comparisons, only two (retrieving, model 4d, and difficulty taking the ball away, model 4e) revealed a significant difference (Supplementary Table S4). After removing all non-significant interactions from these models, we found a significant species–familiarity interaction in the retrieving model 4d (Supplementary Tables S9, S10) and a significant species difference in the difficulty of taking the ball model 4e (Supplementary Tables S11, S12). More specifically, the probability of retrieving was higher for dogs than wolves and was not affected by whether the animal was tested with a familiar person or a stranger, whereas for wolves, the probability of retrieving was higher when tested with a stranger as compared to a familiar person (Figure 4A; Table 2). Regarding the difficulty of getting the ball back from the animal, this was higher in wolves than in dogs (Figure 4B; Table 2).
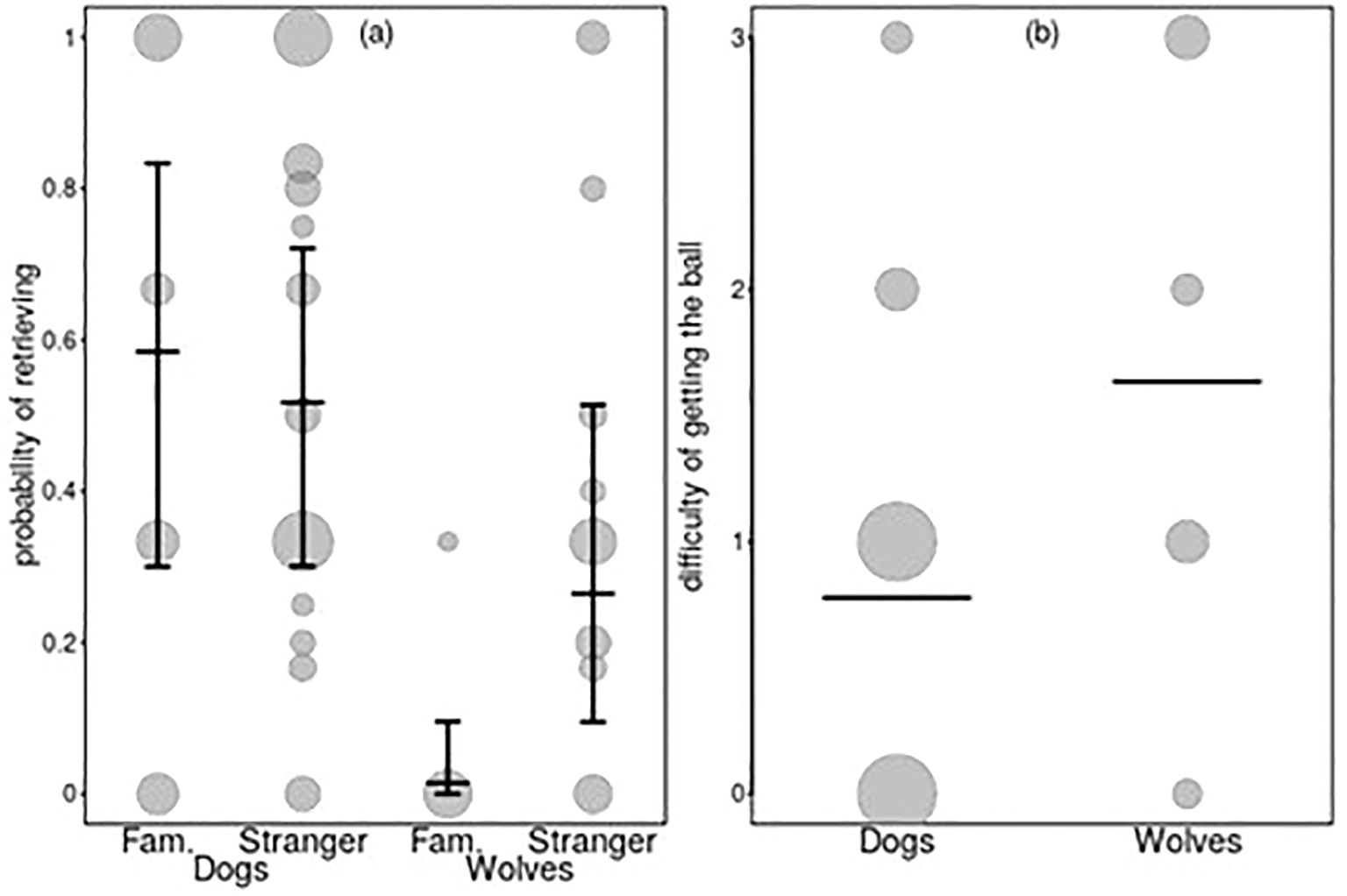
Figure 4. (a, b) Probability of retrieving (a, model 4d) and difficulty obtaining the ball from the animal (b, model 4e). Dots in (a) show the probability of retrieving per animal and familiarity, whereby the area of the dots depicts the total number of experiments summarized per dot (N = 3 to 30). Dots in (b) show the actual response, whereby the area of the dots depicts the relative frequency by which they occurred (N = 7 to 51). Horizontal line segments depict the fitted model, and error bars and its confidence limits for all the other terms in the respective model are centered to a mean of zero.
3.5 Brushing (models 5a to 5f)
When investigating whether wolves and dogs differed in their propensity and latency to accept brushing by the experimenter, neither full-null model comparison revealed a difference between them (Supplementary Table S4). The full model for the proportion of time accepting the brushing did not converge. No differences between wolves and dogs were found regarding biting attempts or avoidance and stress-related behaviors.
3.6 Muzzling (models 6a to 6e)
None of the full-null model comparisons revealed a difference between wolves and dogs in the muzzling test (Supplementary Table S4).
3.7 Lying on the back (models 7a to 7d)
The full model fitted to test for a difference between the experimenters’ success in getting wolves and dogs to lie on their backs (model 7a) did not converge, most likely because only 5 out of 44 tests were unsuccessful, and all of these were trials with wolves. The full-null model comparisons for models 7c (proportion of time the animal spent lying on the back) revealed significance, but not for model 7b (latency to lie on the back) (Supplementary Table S4). For model 7e (biting attempts), the null model did not converge. For the proportion of lying on the back (model 7c), the interaction between species and method (coercive or persuasive) was significant (Supplementary Table S13). In detail, although dogs remained lying on their backs longer on average when the experimenter had used physical restraint (coercive method) vs. persuasion, this difference was not significant (estimate = −0.84, SE = 0.68, adj. p = 0.60), whereas the opposite was the case for the wolves. Wolves remained on their backs longer when the experimenter had used persuasion instead of coercion (estimate = 1.24, SE = 0.42, adj. p = 0.031), and they remained down for longer than dogs in the persuasive condition (estimate = −2.16, SE = 0.73, adj. p = 0.028) (Figure 5; Table 2). Finally, wolves attempted to bite the experimenter’s hand more often than dogs, particularly when they used the persuasive method (Supplementary Table S14), but zero inflation was an issue, and the null model did not converge, so this finding must be interpreted cautiously.
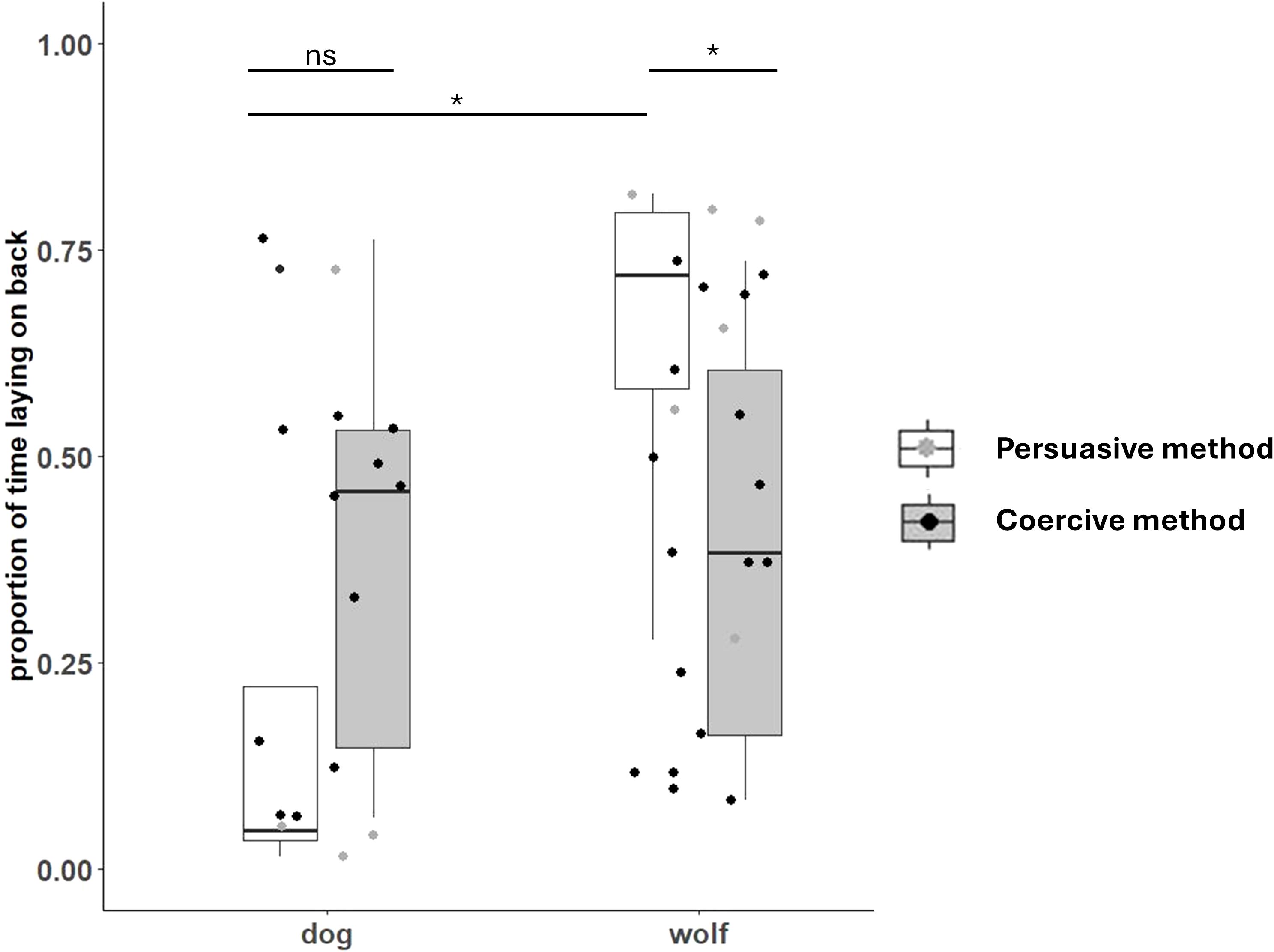
Figure 5. Proportion of test time dogs and wolves spent lying on their backs (model 7c) when the experimenter used either persuasion (verbal encouragement, white boxes) or coercion (physical restraint, gray boxes). Dots show the observed response. Boxes with thick horizontal lines depict quartiles and medians of the response.
4 Discussion
In the current study, we investigated the trainability, responsiveness to persuasion, and acceptance of coercion of young, similarly raised wolves and dogs, as possible mechanisms underlying the animals’ tractability. Wolves’ and dogs’ developmental trajectories are very similar, with olfaction being functional by 2 weeks of age, audition by 4 weeks, and vision by 6 weeks on average, but wolves explore their environment earlier than dogs (Lord, 2013). In the current study, we show that, already at 3 weeks, hand-raised wolf and dog pups show directed movement toward a human caller, suggesting that they are capable of meaningful engagement with social cues at this age. While early developmental stages present certain constraints, these are precisely the stages during which domestication-related differences are likely to emerge and are not yet influenced by experience.
4.1 Trainability
In line with our predictions, dogs and wolves showed similar success rates when asked to sit, lie down, or when being called—even in the presence of a distractor (inaccessible food; Figure 1). Both wolves and dogs responded faster to the commands with increasing age, demonstrating a similar learning effect over time. Distraction did not influence the results: Wolves, in line with previous research showing that wolf pups are more explorative (Marshall-Pescini et al., 2017) and more persistent than dog pups in object manipulation (Rao et al., 2018), manipulated the cage with the food significantly more than the dogs, which, however, did not affect their success or latency to reach the caller. In Ujfalussy and colleagues’ study, latencies to come after calling differed between wolves and dogs only at ages 16 and 24 weeks, when the animals were called away from conspecifics, which meant they had to be willing to move away from a significant social distraction. Wolves may prioritize interactions with their conspecifics over humans, whereas dogs may be more inclined to follow human invitations to interact, similar to their higher success rates when cooperating with humans than conspecific partners (Marshall-Pescini et al., 2018). Furthermore, although Ujfalussy et al. (2020) did not test their subjects using the lie down command, they found that dogs were quicker to sit down than wolves at ages 7 and 16 weeks but not at 9, 12, and 24 weeks. In contrast, we found a significant difference in the latency to follow the lie down command, with wolves taking longer than dogs. Lying down puts the animal into a compromised position, which they may perceive as a loss of control, as it becomes more difficult to get away from the situation. It is likely that dogs feel more comfortable putting themselves into such a situation than wolves because of their domestication process. Accepting the loss of control and going along with human directions appears to be a central feature of this process, as outlined by the deferential behavior hypothesis, which has shaped dogs into such useful partners for humans.
4.2 Responsiveness to persuasion
In the fetch and retrieve test, in contrast with our predictions, dogs and wolves did not differ from each other with the stranger at ages 6 and 8 weeks. However, in the 9-week tests with a familiar experimenter, dogs retrieved the ball significantly more often than wolves (10 out of 33 dogs retrieved the ball at least once, whereas only 1 out of 18 wolves did so, Supplementary Figure S1). While the full model suggests that this effect is better explained by experimenter familiarity than age (as the interaction between species and age was not significant, Supplementary Table S11), age and experimenter familiarity were confounded due to the study design. Thus, wolves may either have lost interest in retrieving the ball with increasing age or they were less interested in retrieving the ball for a familiar person instead of a stranger. Interestingly, a recent study reported that 8-week-old wolf puppies of one litter (but not the others) spontaneously retrieved a ball for an unfamiliar person without prior training (Hansen-Wheat and Temrin, 2020). Such findings indicate that behavioral variation in response to human cues is already present in ancestral populations and could have served as a selection target during the early stages of domestication. Lastly, in the tests with the stranger at 6 and 8 weeks, it was significantly more difficult to get the ball back from the wolves than from the dogs. This aligns with the findings of Ujfalussy et al. (2020) of higher rates of aggressive behaviors in that task in wolves than in dogs and our prediction that it would be harder to persuade the wolves to give up a resource than the dogs, especially if no previous relationship had been built with their respective human partners.
4.3 Acceptance of coercion
In contrast to our predictions, we found no differences between wolves and dogs in the brushing and muzzling tests. In the previous study by Ujfalussy et al. (2020), wolves made more biting attempts during brushing than dogs, but they were required to lie down for the brushing; hence, these different results could be attributed to a perceived higher vulnerability compared to the present study, where animals experienced less restraint. In contrast, in the muzzle test, the animals were similarly accepting of wearing it in both studies. In the present study, wolves and dogs differed in the lying on the back test, which can be considered the most restrictive test in this battery. Specifically, dogs remained lying on their backs for slightly (although not significantly) longer when the experimenters employed a “coercive” style using physical restraint than when using a “persuasive” style of verbal encouragement and petting. Wolves, in contrast, stayed on their backs for longer when the experimenter used no physical restraint and did not attempt to prevent them from getting up again. Remarkably, wolves spent more time lying on their backs than dogs when the experimenter used persuasion but also tried to bite (in a playful rather than aggressive manner) more often than dogs. However, due to a lack of null model convergence, this latter result should be considered with caution. Accordingly, wolves may need more convincing than dogs that a situation is “safe” for them to go along with the human partner’s wishes. However, if the partner can establish a trusting relationship and does not violate this trust (i.e., by using physical restraint to prevent the animal from getting out of the situation), wolves can be at least as compliant as dogs. This is supported by previous research on human-directed attachment. Although both hand-raised wolves and dogs can form strong relationships with familiar humans (Hansen-Wheat et al., 2022, but see comment by Gácsi et al., 2023), wolves tend to be more selective and less trusting than dogs when interacting with familiar, unbonded people (Lazzaroni et al., 2020; Wirobski et al., 2021; Burkhard et al., 2023).
The lack of a significant difference in dogs’ response to coercive vs. persuasive handling is a bit surprising. Given that experimenters received minimal instructions before the test to create a natural situation, it is possible that they were more likely to use the coercive method first with the dogs and act more persuasively only with those dogs that were more difficult to handle. This could have resulted in them being less likely to stay lying down with the persuasive method. Importantly, neither wolves nor dogs appeared to have experienced the situation as stressful as evidenced by a very low occurrence of stress-related behaviors during the test situation (we recorded only two occurrences of stress-related behavior, yawning, scratching, or shaking, in two wolves and two dogs, respectively, which precluded any further statistical analyses). This low frequency indirectly supports the interpretation that species differences are not easily attributable to stress responses. In addition, lying on the back may have different functional meanings across contexts and species. In dogs, it has been described as facilitating social play (Norman et al., 2015), whereas in wolves, it may be tied more strongly to self-handicapping, which is related to dominance relationships outside of play (Essler et al., 2016; Cafazzo et al., 2018). In any case, elements of the test situations described in this study indeed show some overlap with typical canine conspecific play interactions (e.g., grooming, nibbling and mouthing behavior, rolling over). This similarity in form makes it difficult to clearly separate play from low-level physical coercion, and we cannot be sure how the animals perceived the situation. In this framework, the use of the term coercion is grounded in the asymmetry of the situation: the experimenter initiated and directed the interaction, whereas the animal was physically controlled by her. Nevertheless, none of the animals responded in a scared or fearful manner. The fact that some individuals (wolves more than dogs) engaged in play-biting supports the notion that they experienced the situation, at least initially, as a playful interaction. A carefully controlled follow-up experiment could use this setup, with each animal assigned to both handling conditions and standardized experimenter behavior, to further investigate those alternative explanations.
To sum up, there were no differences between wolves’ and dogs’ trainability, and we found no firm evidence that dogs respond stronger to persuasion than wolves. However, in situations that required giving up control and putting oneself in a more vulnerable position, such as lying down, dogs responded faster than wolves. While dogs were as accepting of the “coercive” as the “persuasive” style, in line with selection for deferential behavior, wolves responded better to the “persuasive” style, likely depending more on previous trust-building interactions with the human partner. This also aligns with previous studies where wolves were less willing to accept humans when they were physically restraining them (Zimen, 1971), forbidding something (Gácsi et al., 2013; Range et al., 2019; Ujfalussy et al., 2020), or taking the leading role (Range et al., 2019). The presence of such subtle differences in the acceptance of physical handling at a young age supports the notion that domestication selected for an increased willingness to follow human guidance, even before extensive training or social experience has occurred.
Finally, our results may also suggest that the hierarchical elements of the relationship with humans have different meanings for wolves or dogs. Dogs accept the human leadership role more readily, submitting to their handling, relinquishing control, and striving to meet expectations, including coming close and accepting manipulation, even if it causes them some discomfort (Wirobski et al., 2021). Conversely, wolves are only willing to endure potentially unpleasant situations if the relationship with the human is strong and built on previous trust, and the requests are respectful (i.e., expressed without coercion). If these elements are absent, wolves find no reason to comply and might respond with avoidance or even aggression. Such a difference in compliance consistently explains the behavior of dogs with conspecific and human interaction partners, i.e., accepting the leading role of higher-ranking individuals to avoid potentially dangerous conflicts. This interpretation also accounts for the fact that human-socialized wolves are cooperative with bonded humans in various situations (training, cooperative string-pulling, daily handling, and even medical procedures), but that their readiness to cooperate depends on both the relationship with (i.e., reflecting trust) and the interaction style of the human partner (i.e., persuasion vs. coercion).
Which specific aspects of the domestication process selected for these differences between wolves and dogs remains an open question. However, it is very likely that humans selected animals that accepted the inhibitory and leading role of humans more readily, as suggested by the “deferential behavior” and “super-dominance” hypotheses (Virányi and Range, 2014; Range et al., 2019; Wynne, 2021). Accordingly, after an initial selection against fear during the first phase of the domestication process, dogs were selected for increased deference to higher-ranking individuals to minimize conflicts over resources, ensure safe cohabitation, and promote cooperation in a way that humans lead and dogs follow.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements, because in Austria, no special permission was needed for the use of animals in socio-cognitive studies at the time the studies were conducted (Tierversuchsgesetz 2012 – TVG 2012). The relevant committee that allows research to run without special permission regarding animals is the Tierversuchskommission am Bundesministerium für Wissenschaft und Forschung (Austria). This research complies with the current Austrian laws on animal protection. All methods were carried out in accordance with relevant guidelines and regulations.
Author contributions
GW: Writing – review & editing, Formal Analysis, Project administration, Data curation, Writing – original draft, Visualization. FR: Supervision, Conceptualization, Methodology, Writing – review & editing, Resources, Writing – original draft, Investigation, Funding acquisition. LB: Data curation, Writing – original draft. ZV: Conceptualization, Methodology, Writing – review & editing, Supervision, Investigation, Funding acquisition.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded in part by the Austrian Science Fund (FWF) (10.55776/P34675) to FR and in part by the Vienna Science and Technology Fund (WWTF) through project CS11–026 to ZV. For open access purposes, the author has applied a CC BY public copyright license to any Author Accepted Manuscript version arising from this submission. Open Access funding provided by the University of Veterinary Medicine Vienna.
Acknowledgments
The Wolf Science Center was established by ZV, FR, and Kurt Kotrschal and later carried on by Marianne Heberlein and Aleksander Orlic as the Core Facility Wolf Science Center of the Vetmeduni Vienna. We thank all the animal trainers, students, helpers, and administrative staff who made this research possible. We thank Roger Mundry for statistical support and Kata Rille for video coding. We further thank the Game Park Ernstbrunn for hosting the CF-WSC.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fetho.2025.1616436/full#supplementary-material
References
Baayen R. H. (2008). A practical introduction to statistics using R. Analyzing linguistic data 2001. Cambridge: Cambridge University Press.
Barr D. J., Levy R., Scheepers C., and Tily H. J. (2013). Random effects structure for confirmatory hypothesis testing: Keep it maximal. J. Memory Lang. 68, 255–278. doi: 10.1016/j.jml.2012.11.001
Bates D., Mächler M., Bolker B. M., and Walker S. C. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67. doi: 10.18637/jss.v067.i01
Bentosela M., Wynne C. D. L., D’Orazio M., Elgier A., and Udell M. A. R. (2016). Sociability and gazing toward humans in dogs and wolves: Simple behaviors with broad implications. J. Exp. Anal. Behav. 105, 68–75. doi: 10.1002/jeab.191
Boada M. and Wirobski G. (2024). Human-directed sociability in the domestic dog: a Tinbergian approach. Neurosci. Biobehav. Rev. 168, 105947. doi: 10.1016/j.neubiorev.2024.105947
Brooks M. E., Kristensen K., Van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Burkhard M. E., Range F., Ward S. J., and Robinson L. M. (2023). Bonded by nature: Humans form equally strong and reciprocated bonds with similar raised dogs and wolves. Front. Psychol. 13. doi: 10.3389/fpsyg.2022.1044940
Cafazzo S., Marshall-Pescini S., Essler J. L., Virányi Z., Kotrschal K., and Range F. (2018). In wolves, play behaviour reflects the partners’ affiliative and dominance relationship. Anim. Behav. 141, 137–150. doi: 10.1016/j.anbehav.2018.04.017
Christensen R. H. B. (2019). ordinal—Regression models for ordinal data. R Package version 10, 54. Available online at: https://github.com/runehaubo/ordinal (Accesed June 8, 2025).
Duguid S. and Melis A. P. (2020). How animals collaborate: underlying proximate mechanisms. Wiley Interdiscip. Rev.: Cogn. Sci. 11, e1529. doi: 10.1002/wcs.1529
Essler J. L., Cafazzo S., Marshall-Pescini S., Virányi Z., Kotrschal K., and Range F. (2016). Play behavior in wolves: Using the ‘50: 50’rule to test for egalitarian play styles. PloS One 11, e0154150. doi: 10.1371/journal.pone.0154150
Forstmeier W. and Schielzeth H. (2011). Cryptic multiple hypotheses testing in linear models: Overestimated effect sizes and the winner’s curse. Behav. Ecol. Sociobiol. 65, 47–55. doi: 10.1007/s00265-010-1038-5
Frank H. (1980). Evolution of canine information processing under conditions of natural and artificial selection. Z. für Tierpsychologie 53, 389–399. doi: 10.1111/j.1439-0310.1980.tb01059.x
Frank H. (2011). Wolves, dogs, rearing and reinforcement: complex interactions underlying species differences in training and problem-solving performance. Behav. Genet. 41, 830–839. doi: 10.1007/s10519-011-9454-5
Friard O. and Gamba M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Gácsi M., Miklósi Á., and Topál J. (2023). Comment on “Human‐directed attachment behaviour in wolves suggests standing ancestral variation for human‐dog attachment bonds”. Ecol. Evol. 13 (9), e10514. doi: 10.1002/ece3.10514
Gácsi M., Vas J., Topál J., and Miklósi Á. (2013). Wolves do not join the dance: Sophisticated aggression control by adjusting to human social signals in dogs. Appl. Anim. Behav. Sci. 145, 109–122. doi: 10.1016/j.applanim.2013.02.007
Hansen-Wheat C., Larsson L., Berner P., and Temrin H. (2022). Human-directed attachment behavior in wolves suggests standing ancestral variation for human–dog attachment bonds. Ecol. Evol. 12, e9299. doi: 10.1002/ece3.9299
Hansen-Wheat C. and Temrin H. (2020). Intrinsic ball retrieving in wolf puppies suggests standing ancestral variation for human-directed play behavior. IScience 23. doi: 10.1016/j.isci.2019.100811
Kaminski J. (2009). Dogs (Canis familiaris) are adapted to receive human communication. In Neurobiology of “Umwelt” How Living Beings Perceive the World (Berlin, Heidelberg: Springer Berlin Heidelberg).
Kerepesi A., Dóka A., and Miklósi Á. (2015). Dogs and their human companions: The effect of familiarity on dog–human interactions. Behav. Process. 110, 27–36. doi: 10.1016/j.beproc.2014.02.005
Klinghammer E. and Goodmann P. A. (1987). “Socialization and management of wolves in captivitiy,” in Man and Wolf: Advances, Issues, and Problems in Captive Wolf Research (Netherlands: Springer Dordrecht). 978-90-6193-614-5.
Lazzaroni M., Range F., Backes J., Portele K., Scheck K., and Marshall-Pescini S. (2020). The effect of domestication and experience on the social interaction of dogs and wolves with a human companion. Front. Psychol. 11, 508171. doi: 10.3389/fpsyg.2020.00785
Lord K. (2013). A comparison of the sensory development of wolves (Canis lupus lupus) and dogs (Canis lupus familiaris). Ethology 119, 110–120. doi: 10.1111/eth.12044
Marshall-Pescini S., Basin C., and Range F. (2018). A task-experienced partner does not help dogs be as successful as wolves in a cooperative string-pulling task. Sci. Rep. 8, 16049. doi: 10.1038/s41598-018-33771-7
Marshall-Pescini S., Virányi Z., Kubinyi E., and Range F. (2017). Motivational factors underlying problem solving: comparing wolf and dog puppies’ Explorative and neophobic behaviors at 5, 6, and 8 weeks of age. Front. Psychol. 8. doi: 10.3389/fpsyg.2017.00180
McCullagh P. (1989). Generalized Linear Models: Bd. 2nd ed Vol. 37 (Routledge: eBook Collection (EBSCOhost). Available online at: https://search.ebscohost.com/login.aspx?direct=true&db=nlebk&AN=1999050&site=ehost-live (Accessed July 23, 2025).
Norman K., Pellis S., Barrett L., and Henzi S. P. (2015). Down but not out: Supine postures as facilitators of play in domestic dogs. Behav. Process. 110, 88–95. doi: 10.1016/j.beproc.2014.09.001
Range F., Leitner K., and Virányi Z. (2012). The influence of the relationship and motivation on inequity aversion in dogs. Soc. Justice Res. 25, 170–194. doi: 10.1007/s11211-012-0155-x
Range F., Marshall-Pescini S., Kratz C., and Virányi Z. (2019). Wolves lead and dogs follow, but they both cooperate with humans. Sci. Rep. 9, 2–11. doi: 10.1038/s41598-019-40468-y
Range F., Ritter C., and Virányi Z. (2015). Testing the myth: tolerant dogs and aggressive wolves. Proc. R. Soc. B: Biol. Sci. 282, 20150220. doi: 10.1098/rspb.2015.0220
Range F. and Virányi Z. (2014). Wolves are better imitators of conspecifics than dogs. PloS One 9, e86559. doi: 10.1371/journal.pone.0086559
Rao A., Bernasconi L., Lazzaroni M., Marshall-Pescini S., and Range F. (2018). Differences in persistence between dogs and wolves in an unsolvable task in the absence of humans. PeerJ 6, e5944. doi: 10.7717/peerj.5944
R Core Team (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/.
Schielzeth H. and Forstmeier W. (2009). Conclusions beyond support: Overconfident estimates in mixed models. Behav. Ecol. 20, 416–420. doi: 10.1093/beheco/arn145
Ujfalussy D. J., Virányi Z., Gácsi M., Faragó T., Pogány Á., Bereczky B. M., et al. (2020). Comparing the tractability of young hand-raised wolves (Canis lupus) and dogs (Canis familiaris). Sci. Rep. 10, 14678. doi: 10.1038/s41598-020-71687-3
Vasconcellos S., Virányi Z., Range F., and Ades C. (2016). Training reduces stress in human-socialised wolves to the same degree as in dogs. PloS One 11.9 (2016), e0162389. doi: 10.1371/journal.pone.0162389
Virányi Z. and Range F. (2014). “On the way to a better understanding of dog domestication: aggression and cooperativeness in dogs and wolves,” in The Social Dog: Behavior and Cognition (Amsterdam, Netherlands: Elsevier). doi: 10.1016/B978-0-12-407818-5.00002-4
vonHoldt B. M., Shuldiner E., Koch I. J., Kartzinel R. Y., Hogan A., Brubaker L., et al. (2017). Structural variants in genes associated with human Williams-Beuren syndrome underlie stereotypical hypersociability in domestic dogs. Sci. Adv. 3, e1700398. doi: 10.1126/sciadv.1700398
Wirobski G., Range F., Schaebs F. S., Palme R., Deschner T., and Marshall-Pescini S. (2021). Life experience rather than domestication accounts for dogs’ increased oxytocin release during social contact with humans. Sci. Rep. 11, 1–12. doi: 10.1038/s41598-021-93922-1
Wynne C. D. (2021). The indispensable dog. Front. Psychol. 12, 656529. doi: 10.3389/fpsyg.2021.656529
Keywords: tractability, trainability, persuasion, coercion, dog–wolf comparison, domestication, deferential behavior hypothesis
Citation: Wirobski G, Range F, Bonnet L and Virányi Z (2025) Trainability, persuasion, or coercion: a study of human–dog and human–wolf interactions in early life. Front. Ethol. 4:1616436. doi: 10.3389/fetho.2025.1616436
Received: 22 April 2025; Accepted: 25 July 2025;
Published: 20 August 2025.
Edited by:
Enrique Font, University of Valencia, SpainReviewed by:
Anne M. Burrows, Duquesne University, United StatesLugosi Anna, Eötvös Loránd University, Hungary
Copyright © 2025 Wirobski, Range, Bonnet and Virányi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gwendolyn Wirobski, R3dlbmRvbHluLndpcm9ic2tpQHVuaW5lLmNo; Friederike Range, RnJpZWRlcmlrZS5SYW5nZUB2ZXRtZWR1bmkuYWMuYXQ=
†These authors have contributed equally to this work and share first authorship