 Keith McAllister†
Keith McAllister† Caleb J. Axelrod
Caleb J. Axelrod Frédéric Laberge
Frédéric Laberge- Department of Integrative Biology, University of Guelph, Guelph, ON, Canada
Studying the factors that determine cognitive performance in animals is challenging under natural conditions but necessary to ensure that the laboratory test results are relevant to wild populations. We took advantage of nest fidelity in parental male pumpkinseed sunfish (Lepomis gibbosus) to conduct a novel object recognition (NOR) assay in the field. We assessed consistent individual differences in behaviour across the object familiarisation and test phases of the NOR assay and collected anatomical measurements, including brain size, after assay completion. We hypothesised that brain size would influence pumpkinseed cognitive ability and predict NOR performance after accounting for individual behavioural differences. Parental males showed repeatable reactions to nest disturbance and to the presence of objects at the nest periphery between assay phases, as well as correlated object investigation behaviours. We found evidence of novel object recognition memory at the population level, although it varied widely among individuals. Individual differences in object interactivity did not influence NOR performance, but relative brain mass (corrected for body length) did. Parental male pumpkinseed with relatively larger brains performed more poorly than males with relatively smaller brains. We interpret this negative relationship between brain size and NOR performance in the context of severe energy limitations faced by parental males during reproduction. Specifically, males maintaining energetically costly larger brains are likely operating near their upper energetic limit, with little or no spare resources available for investment in demanding learning processes. If this is the case, our findings emphasise that relationships between brain size and cognitive test performance may depend on energy availability and expenditure rates.
Introduction
Cognition is commonly defined as the ability of individuals to process, integrate, and use information from their environment, and it has often been suggested as a key driver of variation in animal performance and fitness (Shettleworth, 2010). Animal cognition is tested using a variety of learning, memory, or problem-solving paradigms. The assumption is often made that performance on these tests represents a direct measure of cognitive ability (Benson-Amram et al., 2016; Buechel et al., 2018; Kotrschal et al., 2013; MacLean et al., 2014). However, a variety of factors beyond cognitive ability can influence performance in tests designed to evaluate animal cognition (Boogert et al., 2018). These include consistent individual differences in behaviour within populations (Carere and Locurto, 2011; Sih and Del Giudice, 2012), external environmental factors (physical, social, developmental), and internal factors (reproductive status, health, stress) (Cauchoix et al., 2020; Kotrschal and Taborsky, 2010; Lambert and Guillette, 2021; Lupien et al., 2009). This potentially complex web of influences on test performance is difficult to untangle. Accurate estimates of cognitive ability and its variation within and across species require accounting for these additional influences on performance in cognitive tests.
There is long-standing interest in uncovering relationships between brain size and cognitive abilities among vertebrates (see Healy, 2021; Jerison, 1973; Striedter, 2005) and invertebrates (Greenspan and van Swinderen, 2004). Increased brain size is thought to occur only when sufficient cognitive challenges are present because the high energetic costs of growth and maintenance of nervous tissue should deter unnecessary brain enlargement (Niven and Laughlin, 2008). Changes in the efficiency of neural activity patterns without changes in brain size may also affect cognition (Chittka and Niven, 2009; Dubois et al., 2018), especially if brain size is constrained, but such differences are often understudied because they are harder to measure than brain size. Evidence linking variation in whole brain size (relative to body size or absolute size) to specific cognitive abilities has been obtained in comparative studies (e.g., Benson-Amram et al., 2016; MacLean et al., 2014; Reader et al., 2011). Comparing cognitive abilities among species is challenging due to difficulties in establishing equivalent testing contexts (Macphail, 1982) and isolating causal from correlated factors that drive variation among species (Peiman and Robinson, 2017). It is therefore also advantageous to evaluate variation in cognitive performance and brain size within species (see Logan et al., 2018). In support of this view, artificial selection for brain size affects some complex forms of cognitive performance in the guppy (Poecilia reticulata), such as numerical discrimination and executive functions that support goal-directed behaviours (e.g., Buechel et al., 2018; Kotrschal et al., 2013; Triki et al., 2023). The ability to perform different cognitive tasks also correlates positively with brain size in Long–Evans rats (Anderson, 1993).
Individual behavioural differences unrelated to brain size or cognitive ability may also influence performance in cognitive tests. The closely related concepts of animal personality and behavioural syndromes refer to stable behavioural differences expressed across time and contexts among individuals of the same species (Carere and Locurto, 2011; Sih and Bell, 2008). Personality focuses on repeatable behavioural differences, whereas syndromes emphasise correlations between different behavioural traits within individuals. Individual differences, such as those along the familiar shy–bold continuum (i.e., shy: less active, less exploratory versus bold: more active, more exploratory; Wilson et al., 1994), can correlate with variation in learning task performance due to differences in exploration. For example, a positive correlation between exploratory tendency and learning performance, independent of stress, emotional reactivity, or physical attributes, has been observed in outbred CD-1 mice (Matzel et al., 2006). The potential link between personality and cognition led Sih and Del Giudice (2012) to propose the concept of cognitive syndrome. A cognitive syndrome implies correlated suites of individual cognitive characteristics that result in different cognitive styles among individuals, influencing the ways animals acquire, process, store, or act on information, independent of overall cognitive ability. Many examples have shown a relationship between cognitive styles and learning performance (Daniel and Bhat, 2020; Gibelli et al., 2019; Jones et al., 2020; Lucon-Xiccato and Bisazza, 2017; Mazza et al., 2018; Schuster et al., 2017). Interestingly, Burns and Rodd (2008) showed that cognitive styles (i.e., speed vs. accuracy of decisions) in guppies are associated with brain morphology—hasty decisions were associated with a smaller telencephalon—but not with performance in a spatial memory task. In the same species, artificial selection for brain size influenced personality and performance on a numerical discrimination task in a sex-specific manner (Kotrschal et al., 2013; Kotrschal et al., 2014). These last studies highlight the complexity and potential variability of relationships between personality, brain size, and cognition.
Studying cognition in natural conditions can be challenging (Morand-Ferron et al., 2016; Salena et al., 2021), so most cognitive testing is conducted under controlled laboratory conditions. However, captive housing and domestication can confound cognitive results by profoundly altering phenotypes (Milla et al., 2021). For example, transferring wild fish into captivity can reduce brain size within 2–6 weeks through phenotypic plasticity (Joyce and Brown, 2020; Park et al., 2012; Turschwell and White, 2016). Captivity can also induce foraging deficits on natural prey in fish (Olla et al., 1998; Vehanen et al., 2009). Fish personality differences that are relatively stable in natural conditions also seem to disappear under laboratory housing (Wilson et al., 1993). Collectively, physical and social environments, particularly impoverished laboratory conditions, likely affect cognitive testing and may limit the relevance of laboratory results for inferring cognition in natural populations. Studies examining multiple potential determinants of cognitive performance in natural conditions are challenging and therefore rare, but they are necessary to improve our understanding of animal cognition.
Here, we performed cognitive testing under natural conditions using a pumpkinseed sunfish (Lepomis gibbosus) ecotype system, which exhibits variation in relative brain size between littoral and pelagic lake habitats without differences in gross brain morphology (Axelrod et al., 2018). This species also shows individual variation in behaviour along the shy–bold continuum in natural populations (Wilson et al., 1993). Novel object recognition (NOR) testing (Ennaceur and Delacour, 1988; Hamilton, 2018; Sivakumaran et al., 2018) on nest-guarding “parental” male pumpkinseed during the reproductive season allowed us to evaluate consistent behavioural differences across test phases as well as recognition memory performance. Anatomical measurements, including brain size, were obtained by collecting individuals after testing was completed. Although we are unaware of evidence directly linking brain size to NOR performance, we expected that larger brains would enhance NOR by supporting increased cognitive processing, thereby facilitating performance on demanding tasks. NOR is likely cognitively demanding because detecting novelty requires substantial memory storage capacity to discriminate novel from familiar features in the environment (e.g., Brady et al., 2008). Moreover, relevant work in guppies suggests that larger brains may enhance performance in more cognitively demanding tasks, such as reversal learning and numerical discrimination, but not in simple colour or spatial discriminations (Buechel et al., 2018; Burns and Rodd, 2008; Kotrschal et al., 2013). The key questions we addressed were whether pumpkinseed exhibit NOR in a natural setting and whether NOR performance is associated with brain size and/or personality traits. We hypothesised that variation in whole brain size (amplified by the mean difference between littoral and pelagic ecotypes) would influence cognitive ability, predicting that larger brains would improve NOR performance after accounting for individual differences in behaviour.
Methods
Study system
Behavioural data were collected from nest-guarding (hereafter “parental”) male pumpkinseed at littoral and pelagic sites in July 2019 (n = 62) and 2020 (n = 33) during daylight hours in Ashby Lake, Addington Highlands, Ontario, Canada (45.0944° N, 77.3496° W), a small (surface area, 2.59 km2; maximum depth, 36.6 m), post-glacial and oligotrophic inland lake. Two pumpkinseed ecotypes coexist in Ashby Lake by inhabiting either the inshore littoral or offshore pelagic lake habitat during the summer growing season (Robinson et al., 1993, Robinson et al., 2000; Gillespie and Fox, 2003; Jastrebski and Robinson, 2004; Berchtold et al., 2015). This population is typical of at least 12 polyphenic populations in the region (Weese et al., 2012). During the summer spawning and growing season, littoral individuals are common in bays with soft sediments, macrophyte vegetation, and coarse woody debris, and feed on large but cryptic benthic macroinvertebrates as well as floating insect prey. Higher densities of pelagic individuals occupy numerous hard rock shoals surrounded by deep, open-water habitat and feed on locally abundant zooplankton prey (Daphnia spp.) (Jarvis et al., 2020). Ecotypes differ along a continuum of variation in body and head anatomical traits related to feeding ecology (Robinson et al., 1993, Robinson et al., 2000; Gillespie and Fox, 2003; Jastrebski and Robinson, 2004; Robinson and Wilson, 1996; Weese et al., 2012; Jarvis et al., 2020). Our previous work showed that the mean brain size, adjusted for body size, is 8.3% larger in adult littoral compared to pelagic individuals, with no differences in the size of five brain regions (Axelrod et al., 2018). Ecotypes can interbreed (Jarvis et al., 2017) and are genetically similar (Weese et al., 2012; Colborne et al., 2016), thus reducing large genetic effects that might confound ecotype comparisons.
Male reproduction provides an ideal opportunity to reliably observe and sample mature male pumpkinseed in the wild because of their intense nest-guarding behaviour. Parental males construct solitary nests in either habitat (Jastrebski and Robinson, 2004; Colborne et al., 2016), typically in water less than 2 m deep from late May to early August (Danylchuk and Fox, 1996). Cuckoldry is possible but has not been observed in Ashby Lake (Gross, 1980; Rios-Cardenas and Webster, 2005). Parental male reproduction is costly. Parental males can lose between 6% and 15% of their wet body weight while nesting (Colgan and Gross, 1977; Gross, 1980, Gross, 1982; Coleman and Fischer, 1991; Rios-Cardenas and Webster, 2005) and face increased mortality postreproduction (Gross, 1980; Gillooly and Baylis, 1999). Successful parental male reproduction ranges from 11 to 21 days (mean, 15 days; Danylchuk and Fox, 1996) depending on water temperature (Cargnelli and Neff, 2006), and has four phases (Colgan and Gross, 1977): territory establishment and nest construction; active courting of and spawning with reproductive females who subsequently leave the nest after spawning; a larval brooding phase; and a vacating phase as larvae disperse and the male terminates reproduction. Successful egg development requires continuous fanning of oxygen-rich water beginning immediately following fertilisation and persists for a minimum of 3 days for each spawned clutch (Gross, 1980; Coleman and Fischer, 1991), longer for multiple clutches (Cooke et al., 2008). Throughout, parental males also vigorously and aggressively defend against predatory intruders (Keenleyside, 1972; Colgan and Gross, 1977; Gross, 1980; Cooke et al., 2008). Parental males may feed opportunistically, but rarely at the expense of parental brood care or nest defence (Thorp et al., 1989). Reproductive success increases with male body condition (Danylchuk and Fox, 1996; Cargnelli and Neff, 2006). Variation in energy reserves and metabolism over winter, largely due to size differences, permits larger males to nest earlier than smaller males, who must feed longer prior to nesting because of their greater winter energy deficit (Danylchuk and Fox, 1996; Cargnelli and Neff, 2006). Larger males in very good condition may spawn a second time (Danylchuk and Fox, 1996; Rios-Cardenas and Webster, 2005). Throughout July, we located and uniquely identified natural nest sites by swimming and boat surveys in both habitats. The lower number of natural nests at pelagic shoals was supplemented by installing shallow plastic basins (diameter, 36 cm; height, 12 cm) filled with 3 cm of the preferred nesting substrate of coarse gravel (n = 37). Our study is necessarily limited to nesting males, so the results are correspondingly limited in their generalisability to the whole population, although we argue they still provide an effective study model for the use of cognitive tests in a natural setting.
Behavioural observations
Parental male behaviour was assessed from remote video recordings made by placing a small low-intrusion camera (Wasp®, Ariss, Ontario, Canada) approximately 1 m away from the nest perimeter to simultaneously view the nest and its resident parental male. We used a spontaneous NOR task to estimate the cognitive performance of parental males (Blaser and Heyser, 2015; Lucon-Xiccato and Dadda, 2014; Sivakumaran et al., 2018). This task normally relies on the innate predisposition of subjects to explore new rather than familiar objects. This exploration bias is considered to reflect the sensitivity of memory for familiar objects and, consequently, the ability to discriminate between old and new stimuli.
The NOR task involves two phases: the initial object familiarisation phase and a test phase with the introduction of a novel object. In the object familiarisation phase, a snorkel diver placed the camera and two identical buoyant objects, A1 and A2, just inside the nest perimeter on opposite sides of the nest relative to the camera view (Figure 1A). Parental male behaviour was then recorded for 30 min. After this time, the diver returned to remove objects A1 and A2 and replaced them with a new version of the original objects (A3) and one novel object, B1, placed at the locations of the original objects (Figure 1B). Another 30 min of male behaviour was recorded with the new objects in place. The parental male fled the nest during both installations of the novel objects, but typically quickly returned in less than a minute after the diver departed, most likely to resume guarding the spawn. Object types A and B were D-shaped pool noodle half sections (orange, 51 mm long × 25 mm maximum width) and ping pong balls (white, 40 mm diametre), respectively (Figure 1). They floated, anchored to stones with transparent fishing line. We minimised risks of intrinsic bias or curiosity effects by using a cross-over design, assigning half of the individuals to one of the two objects during the familiarisation phase (Koivisto et al., 2025) and counterbalancing the side of the novel object to control for side bias.

Figure 1. Novel object recognition assay. (A) Familiarisation phase, in which two identical objects, A1 and A2, are placed just inside the nest perimeter. The nest-guarding male pumpkinseed is visible on the right side of the nest in this panel. (B) Test phase, in which a new specimen of the old object (A3) and a novel object B1 are placed within the nest perimeter.
Upon completion of the test, the parental male was captured using a long-handled net, euthanised, and preserved in 10% neutral buffered formalin for brain size measurement. Capture time following the NOR assay varied because male responses to net intrusion differed, with some males captured immediately upon return and others requiring a longer period, typically less than 10 min. Stress following capture was minimised by immediately passing the netted male to assistants on a closely moored boat for anaesthesia verified by tail pinching (< 3 min in low-dose clove oil solution), followed by euthanasia in a 0.1 mg/ml solution of clove oil for a minimum of 5 min. Methods of capture, handling, and euthanasia of animals were approved by the University of Guelph Animal Care Committee (Protocol No. 4180) under the guidelines of the Canadian Council on Animal Care. Scientific sampling was licensed through the Ontario Ministry of Natural Resources (Permit No. 1092282 and No. 1095398).
Video recordings were scored for male behaviour using JWatcher software (version 1.0, 2000). Behaviour of males collected in 2019 was scored by KM, while behaviour of 2020 males was scored by MD (i.e., observer and year are confounded). Only behaviours focused on the buoyant objects were considered (i.e., interactions with anchoring rocks or lines were not included). First, counts of interactions with objects were made separately for contact with the mouth, contact with the tail, and circling of the object. Approaching an object within half a body length was also considered an interaction and counted. Separate counts were obtained for the two objects. Infrequent interactions with artificial objects involving contacts and circling resulted in little variation among individuals. To increase variation, we calculated two object interaction statistics from these count data. The first summed all noncontact interactions with objects (NCIO = sum of counts of approaches plus circles around objects), and the second summed all contact interactions with objects (CIO = sum of counts of contacts with mouth and tail). Second, elapsed time was determined for four behaviours during each phase of the assay: (1) latency to return (LTR) to the nest after diver-induced disruption, (2) latency until first interaction (LTI) with an object after return to the nest, (3) total time spent investigating objects (TTI), defined as the time between the first and last object interaction during a phase of the assay, and (4) total time spent on each of the right and left sides of the nest, where side occupancy was determined by head position. LTR may reflect how an individual reacts to disturbance, while LTI may reflect curiosity or fear regarding the objects installed at the nest periphery by the diver. TTI likely reflects willingness to explore and gain information about objects. High counts of interactions with objects (noncontact or contact) may reflect a willingness to investigate objects, while nest side occupancy may reflect a less overt object or nesting preference. Most fish ceased interactions with objects before the end of the 30-min period of each assay phase, which we interpret as indicating that the assay duration was sufficient to assess individual behaviour.
Repeatability of scoring between observers was evaluated using the intraclass correlation coefficient (ICC), estimated from a subsample of 15 nesting male trials from 2019 independently scored by each observer (Bell et al., 2009). We used a restricted maximum likelihood (REML) model to estimate the ICC as the per cent variance in behaviour due to a random fish effect, after accounting for fixed effects of habitat, test phase, and observer. Mean repeatability for fish (averaged over separate analyses of left and right side counts; Supplementary Table S1) was high for time spent on each side (ICCmean = 0.93), object circling (ICCmean = 0.92), moderate for latency to return (ICC = 0.74), total time interacting (ICC = 0.64), mouth contacts (ICCmean = 0.67), and object approaches (ICCmean = 0.60), and lowest for tail contacts with object (ICCmean = 0.29), indicating greater variation in observer error.
Novel object recognition performance
We used two different variables to assess NOR performance: the sum count of noncontact interactions with objects (NCIO) and the time spent on each side of the nest. Contact interactions with objects (CIO) were not used as a measure of NOR performance because only 28% of individuals expressed contact behaviours. For each variable, discrimination ratios (DR) were calculated for both the object familiarisation and test phases of NOR assays using the formula: DR = (Tnew − Told) / Ttotal, where Tnew represents time or counts measured on the side of the novel object, Told represents time or counts on the side of the original object, and Ttotal represents the summed time or counts on both sides. The side of the novel object in the test phase determined the “new” side in the object familiarisation phase for the purpose of DR calculations. The change in discrimination ratios between the two phases of the assays (ΔDR = DRtest − DRfamiliarisation) was used to quantify NOR performance while controlling for individual object or side preferences. Our main interest was in the general ability to remember and discriminate objects rather than in preference or avoidance, so absolute values of ΔDR (|ΔDR|) are analysed. As a result, our measure of novel object recognition memory includes individuals expressing either novel object preference or avoidance.
Brain size measurements
Brain mass was used to estimate brain size. Brains of males preserved for less than 7 months were removed by dissection as per Axelrod et al. (2018), trimmed to remove excess fat and cranial nerves, blotted to remove excess formaldehyde, and weighed using an Accu-124D scale (Fisher Scientific, Waltham, MA, USA) to a precision of 0.0001 g. Standard length (± 1 mm) was also measured and used to control for and explore the effects of body size on performance. Body condition was also estimated using the Scaled Mass Index (Peig and Green, 2010).
Year, habitat, and standard length all had significant effects on raw brain mass (Supplementary Table S2). Our focus on relationships between brain size relative to body size, personality, and measures of NOR performance, therefore, required adjusting brain mass for year and body size effects. We estimated “relative” brain mass as the residual values of log10-transformed brain mass regressed against log10-transformed standard length and a year factor (YrSz-adj. BM). Standard length was chosen over body mass for size correction because it is a more stable estimate of body size. We also considered “absolute” brain mass in a few models below, estimated as brain mass adjusted only for the effect of year (Yr-adj. BM). Brain mass was not adjusted for habitat because brain mass differences between ecotypes were either included as a habitat parameter in models or removed through the model selection process.
Statistics
Ecotype differences in behaviour and brain mass
We evaluated the influences of habitat on brain mass and five behavioural response variables (mean LTR, mean LTI, NCIO1, CIO1, TTI1) while statistically controlling for standard length using univariate ANCOVA models. Standard length, brain mass, and the five behaviours were normalised with a log10-transformation after adding one to each value (excluding standard length). An interaction between habitat and standard length was not included in the final models, as it was consistently nonsignificant in preliminary analyses. We used these univariate analyses to help interpret the multivariate variation in personality described below.
Individual differences in behaviour
Personality can influence cognitive performance in fishes (e.g., Daniel and Bhat, 2020), so we developed individual personality scores as parameters in some models. Consistency of behaviour differences among individuals over time (repeatability) was assessed with the intraclass correlation coefficient of LTR, LTI, NCIO, and CIO between phase one and phase two of the NOR assay. Consistent differences expressed across contexts, possibly indicative of individual personality, were first assessed through pairwise Spearman’s rank correlations among different behaviours. Here, individual values of LTR and LTI for each phase of the NOR assay were averaged to obtain mean LTR and mean LTI values. Only TTI1, CIO1, and NCIO1 rank values from phase one were used to avoid the effects of object learning on scores in phase two. Higher correlation coefficients indicate stronger evidence of consistent individual differences in behaviour and help interpret multivariate personality scores estimated below. Variation in shy–bold personality has been assessed in juvenile pumpkinseed under natural and artificial conditions (Wilson et al., 1993), but not under natural conditions using parental males. We applied an exploratory common factor analysis (CFA) approach to identify latent “personality” variables that contribute to multivariate behavioural correlations. Factor analysis was chosen over principal components because CFA accounts for behavioural measurement error (indicated by variation in behavioural ICC scores above). Accounting for error avoids inflated estimates of component variance and of loadings, especially when only a few behaviours are considered (Snook and Gorsuch, 1989). Additionally, CFA permits statistical tests of the relevant number of latent components (Schmitt, 2011). Common factors were estimated using maximum likelihood of log10-transformed values (plus one) of five behaviours (mean LTR and LTI, TTI1, CIO1, and NCIO1). A Factorparsimax rotation was employed because exploratory analyses are often subject to larger cross-loadings among behaviours and among common factors (Schmitt, 2011). Six individuals with missing values of LTI in either assay phase were removed from the CFA. Predictors of individual common factor scores one and two (CF1 and CF2) were then assessed using general linear models including parameters for nesting year (2019, 2020), habitat (littoral, pelagic), and log10-transformations of standard length and relative brain mass (YrSz-adjusted BM). Including explicit effects of year, habitat, and body size on variation in CF1 and CF2 here allowed us to evaluate these effects independently of the relative brain mass effect.
Predictors of NOR performance
First, we assessed whether parental males as a group expressed novel object recognition using separate one-sample t-tests on |ΔDR| values obtained with nest side occupancy (time spent |ΔDR|) and noncontact interactions with objects (NCIO |ΔDR|). This determined whether mean absolute recognition values (i.e., considering preference and avoidance behaviour together) departed significantly from a null hypothesis of zero. This test was performed for littoral and pelagic fish combined (the whole parental population), and again for each ecotype separately in case habitat influenced object recognition ability. Histograms of |ΔDR| and standard ΔDR were also plotted to assess variation in NOR performance among individuals.
Next, we assessed predictors of each NOR performance variable using general linear models that include fixed factors for habitat, year, individual differences in behaviour (CF1 and CF2 scores), brain mass, and two-way interactions between each CF and brain mass. Two versions of brain mass were modelled separately: relative brain mass (i.e., YrSz-adj. BM) and absolute brain mass (i.e., only Yr-adj. BM). Models that included relative brain mass also included log10-transformed standard length to partition body size effects and relative brain mass effects on NOR performance separately, because body size may influence behaviour for ecological reasons independent of a brain contribution. Models including absolute brain mass did not include log10 standard length to avoid collinearity.
The number of parameters (up to eight) risks overparameterising NOR performance models. Thus, we first selected models using Bayesian (BIC) and Akaike information criteria (AICc). Four models were compared for each of two NOR performance response variables. In addition to the full model (e.g., eight predictors: year, habitat, SL, YrSz-adj. brain mass, CF1, CF2, plus two interactions between YrSz-adj. brain mass and CF1, and CF2, respectively), we evaluated three reduced models: a main effects model (year, habitat, SL, YrSz-adj. brain mass, CF1, CF2); a brain and behaviour interaction model (YrSz-adj. brain mass, CF1, CF2, and two interactions between YrSz-adj. brain mass and CF1, and CF2); and a brain and behaviour main effects model (YrSz-adj. brain mass, CF1, CF2). We resolved disagreements between BIC and AICc selection in favour of BIC because its model selection is more consistent with changing sample size (Brewer et al., 2016). All statistical analyses were performed using JMP Pro statistical analysis software version 18 or GraphPad Prism version 8.4.3.
Results
Assay participation
NOR performance was assessed on 89 individuals, representing 94% of nests sampled. Four individuals were not sampled because they did not return to the nest after the diver disturbance, and two individuals were rejected due to the improper positioning of the camera to view behaviour. Basic information summarising variation in the behaviours measured in the two phases of the NOR assay is presented in Supplementary Table S3. An additional six individuals were rejected from the NOR performance analyses that follow: four because of faulty brain dissection, and two littoral individuals because of very high leverage on model fit.
Ecotype differences
Consistent with prior findings of differences in brain size between ecotypes of nonnesting pumpkinseed, the brains of nest-guarding males in the littoral habitat were on average 5.2% heavier than their counterparts in the pelagic habitat (tHab = 2.36, p = 0.021, adj. mean ± SE littoral = 0.129 ± 0.0016 g, pelagic = 0.123 ± 0.0016 g). As expected, standard length had a strong positive effect on brain mass (tSL = 13.7, p = 0.0001), and an unexpected effect between years was also evident (tYr = − 8.38, p = 0.0001; Supplementary Figure S1). The relative brain mass of individuals collected in 2020 was, on average, 23.5% heavier than those from 2019, but the reason for this difference is unclear. To account for this difference between years, we included a year effect in our models. There is also consistent evidence of a year effect on body size and body condition (all p ≤ 0.008; Supplementary Table S2), extending year effects beyond the effect on relative brain mass. On average, parental males in 2019 were larger than those in 2020, but the mean condition of parental males in 2019 was 5% lower than that of those in 2020.
There was also strong evidence of habitat effects on nest-guarding male behaviour (except for mean LTI, p = 0.50) that generally reflected greater object interactivity at pelagic compared to littoral nests (Supplementary Table S4). Pelagic males returned more quickly to their nest (low mean LTR; p = 0.016), had greater numbers of noncontact and contact interactions with artificial objects there (higher NCIO1, p = 0.0016, and CIO1, p = 0.0003 in phase 1) and showed longer total time intervals investigating objects (longer TTI1 in phase 1; p = 0.032) after accounting for effects of standard length and year (Supplementary Table S4). A significant influence of year only occurred for NCIO1, with counts higher in 2020 compared to 2019.
NOR performance: population patterns
We found evidence of novel object recognition memory in the parental male population, justifying further exploration of the factors that influence performance below. The mean time spent |ΔDR| for all individuals combined was 0.17 and differed significantly from zero (one-sample t-test: t88 = 7.3, p < 0.0001). The mean NCIO |ΔDR| was 0.64, which also differed significantly from zero (one-sample t test: t71 = 10.7, p < 0.0001). Evidence for object recognition varied widely among individuals, with many fish expressing either positive or negative ΔDR values, respectively indicating preference or avoidance of the novel object, and some expressing ΔDR values of zero, showing no evidence of object discrimination (Figure 2). There was no evidence that NOR performance was influenced by habitat because the 95% CI of mean recognition responses for both time spent and NCIO overlapped between littoral and pelagic individuals. The mean [95% CI] recognition response in time spent |ΔDR| for littoral individuals was 0.21 [0.13–0.29] (one-sample t-test: t49 = 5.5, p < 0.0001), and in pelagic individuals it was 0.11 [0.09–0.13] (one-sample t-test: t38 = 9.5, p < 0.0001). The mean [95% CI] recognition response in NCIO |ΔDR| for littoral individuals was 0.67 [0.50–0.84] (one-sample t-test: t35 = 8.1, p < 0.0001) and in pelagic individuals it was 0.61 [0.43–0.78] (one-sample t-test: t35 = 7.0, p < 0.0001).
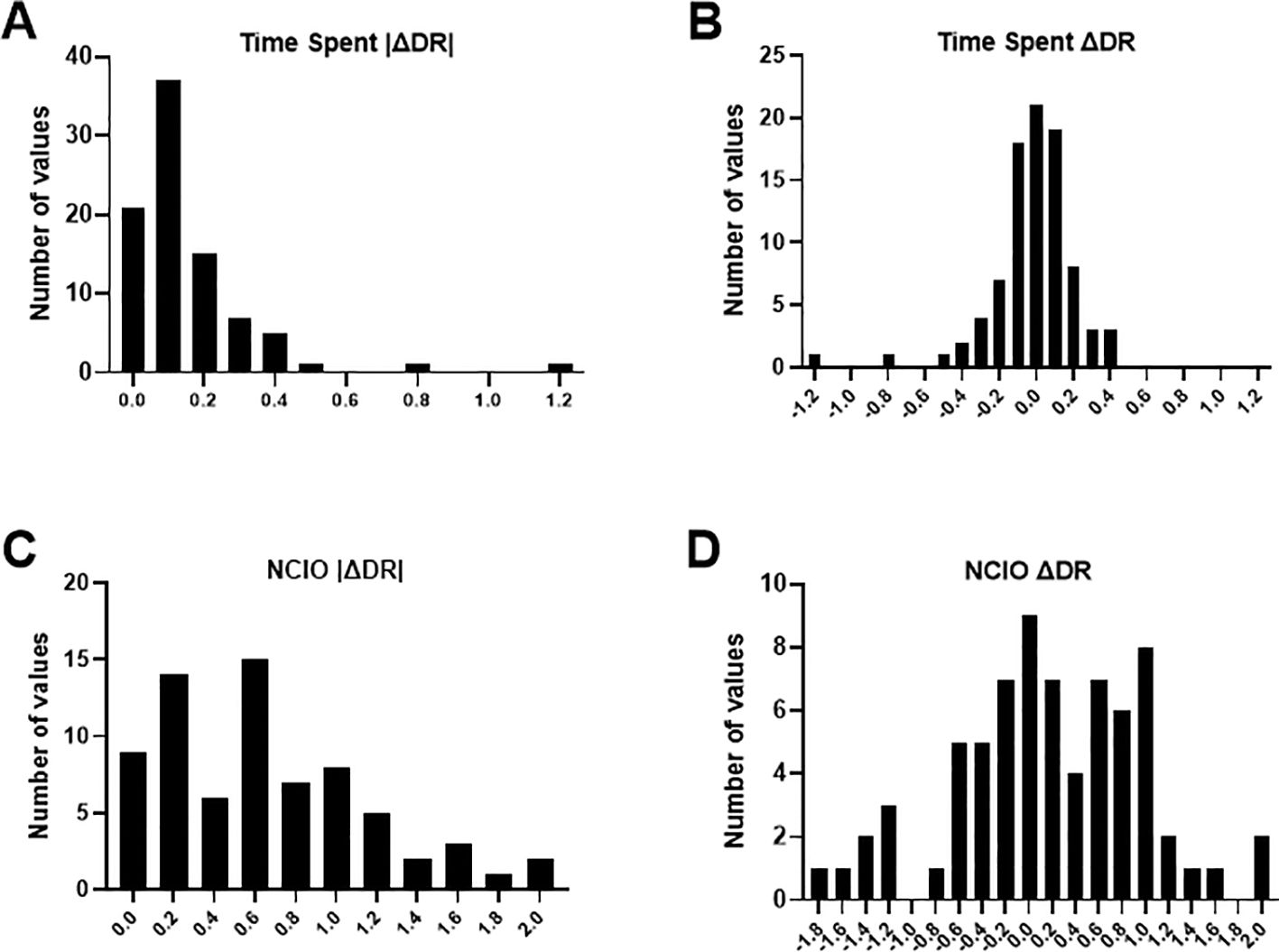
Figure 2. Frequency distribution of the change in discrimination ratios between the two phases of the novel object recognition assays (ΔDR) among pumpkinseed individuals. (A) Absolute values of ΔDR based on nest-side occupancy (time spent). (B) Standard values of ΔDR based on nest-side occupancy. (C) Absolute values of ΔDR based on counts of approaches toward and circling around objects (NCIO, noncontact interactions with objects). (D) Standard values of ΔDR based on noncontact interactions with objects.
Individual differences in behaviour
There was evidence of consistent individual differences in behaviour among parental males. All behaviours except LTI demonstrated significant intraclass correlations between the two phases of the NOR assay, consistent with behaviour repeatability (Table 1). There was also some evidence of consistent individual behavioural differences across behaviours indicative of personality in pumpkinseed. Mean LTI and TTI were negatively correlated (Table 1), and TTI was not constrained by the 30-min duration of assay phases because most individuals ceased interacting with the objects before the end of the assay. The common factor analysis further suggested personality in the parental male pumpkinseed. Two common factors (CF1 and CF2) were sufficient to explain a significant proportion of behavioural covariation (CF1 = 38.1% and CF2 = 20.0%; whole model χ2 = 189.9, df = 10, p = 0.0001) (Figure 3). Larger CF1 values identify individuals that tended to quickly begin investigating objects after return to the nest (low LTI; rotated factor loading = − 0.445), spend a longer time investigating objects (high TTI; loading = 0.986), and have more noncontact and contact interactions (high NCIO and CIO; respective factor loadings = 0.727, 0.451) (Supplementary Figure S2A). Mean LTR hardly contributed to CF1 (loading = 0.042) and is not significantly correlated with any other variable (Table 1). In contrast, larger CF2 values identify individuals that more quickly return to the nest site after disturbance (mean LTR loading = − 0.319) and made more noncontact and contact interactions with the artificial objects (high NCIO and CIO; respective factor loadings = 0.565, 0.692) (Supplementary Figure S2B).
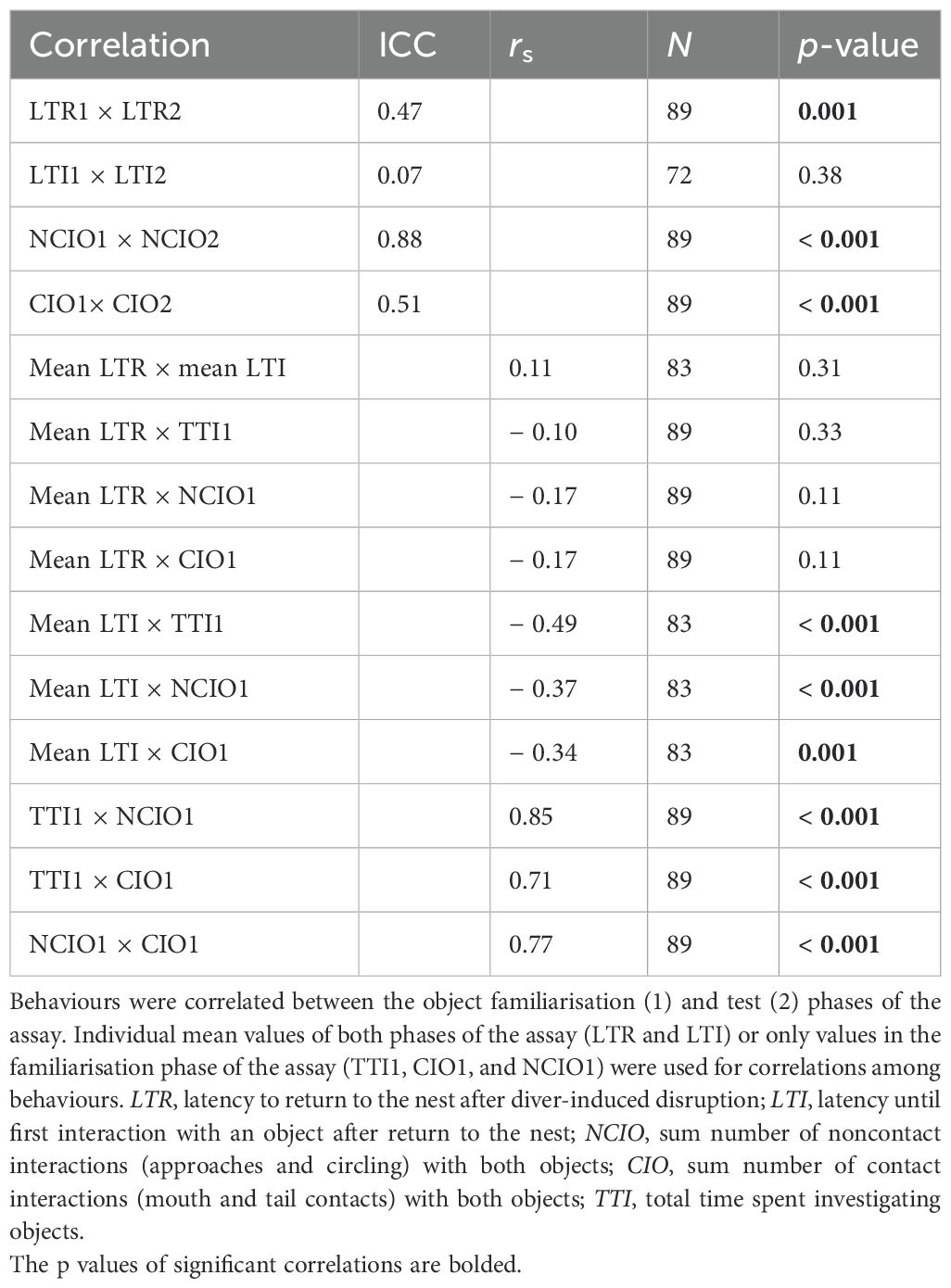
Table 1. Intraclass correlation coefficient (ICC) evaluates repeatability (between phases), and Spearman’s rank correlation (rs) evaluates consistency (among behaviours) in pumpkinseed nest-guarding behaviours expressed during the novel object recognition assay.
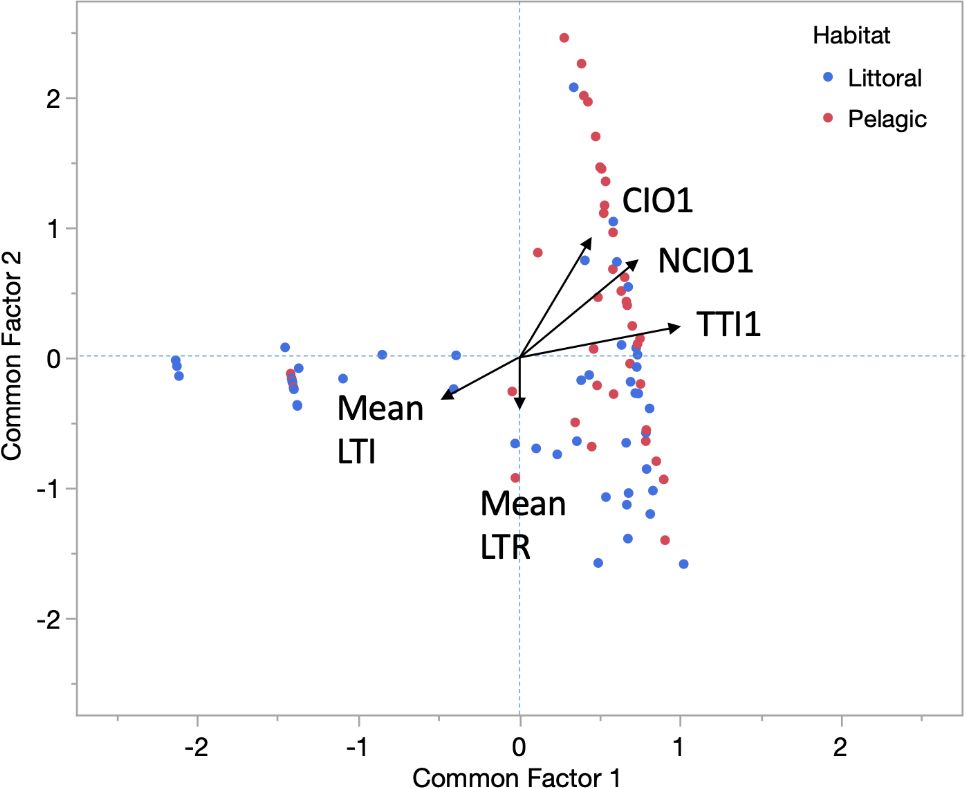
Figure 3. Factor analysis of behavioural patterns expressed by nest-guarding male pumpkinseed combining fish from both habitats (littoral blue and pelagic red). The analysis identified two common factors accounting for a total of 58.1% of the observed variation in five male behaviours. Nesting male behaviour scores are plotted in the space defined by two common factors, CF1 (x-axis, 38.1% of variation in behaviour) and CF2 (y-axis, 20.0% of variation in behaviour), for means of log10-transformed values of LTR and LTI averaged over the two phases of the novel object recognition assay, and TTI1, CIO1, and NCIO1 from phase one of the assay. Black vectors represent correlation loadings of each behaviour with each common factor. Abbreviations: LTR, latency to return to the nest after diver-induced disruption; LTI, latency until first interaction with an object after return to the nest; TTI, total time spent investigating objects; NCIO, total number of noncontact approaches and circling of objects; CIO, total number of mouth and tail contacts with objects.
Variation in both CF1 and CF2 was positively influenced by standard length, although the statistical evidence was marginal for CF2 (Table 2). Thus, larger individuals tended to investigate objects with greater effort (higher CF1) and were more nest vigilant (higher CF2). Furthermore, CF2 (but not CF1) was also influenced by habitat. Individuals with higher values of CF2 tended to be from the pelagic habitat, consistent with lower values of mean LTR and higher values of NCIO and CIO observed at pelagic nest sites (Supplementary Table S4). An effect of year on CF1 was also consistent with a year effect on NCIO1 that loaded strongly on CF1 (Supplementary Table S4).
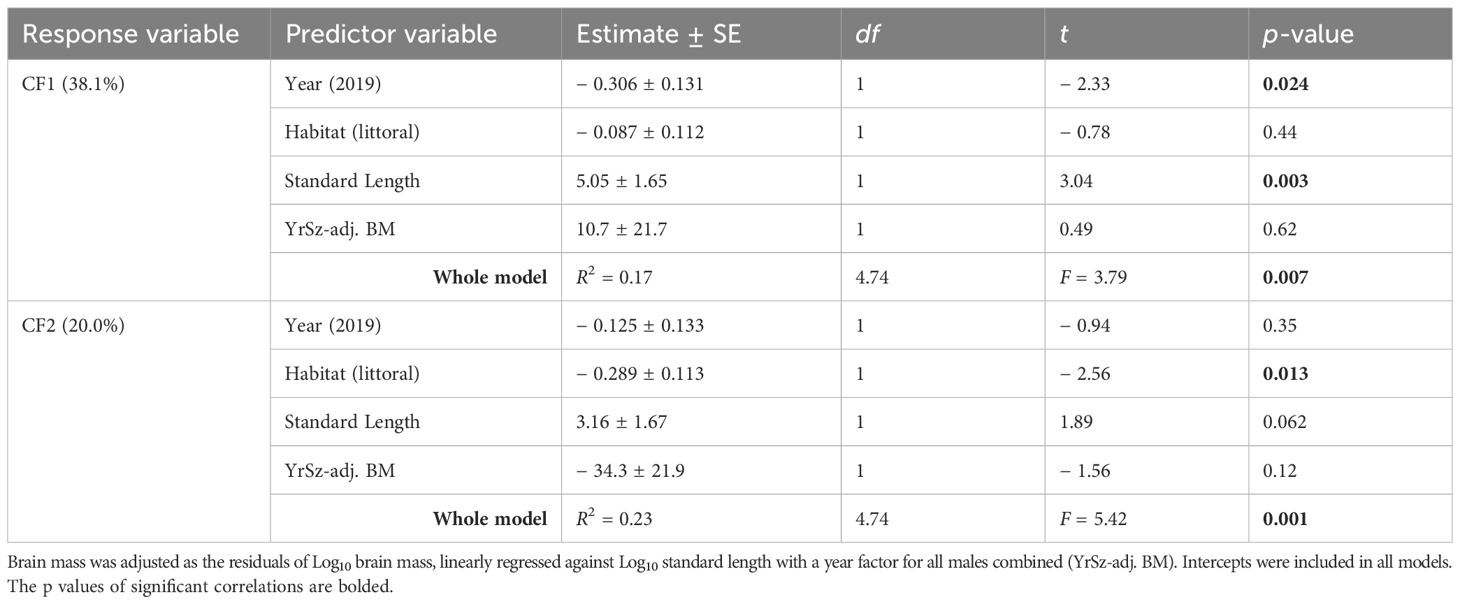
Table 2. Summary of general linear models evaluating the influence of habitat, year, and morphology on variation in common factors CF1 and CF2.
Predictors of NOR performance
The model selection process identified the simplest brain and behaviour main effects models as the most parsimonious, and we used this model to evaluate the effects of relative brain mass on NOR performance (Supplementary Table S5), but these simple models had low explanatory power (R2 of 4% and 9%). Relative brain mass had a negative effect on both time spent and NCIO |ΔDR|, although this was significant only for NCIO (Table 3; Figure 4). Neither CF1 nor CF2 was significantly related to either time spent or NCIO |ΔDR|. A more complex main effects model was ranked 0.8 AICc units above the simplest relative brain mass and behaviour main model for time spent |ΔDR | (Supplementary Table S5), and this model yielded qualitatively similar results to those reported above. Additional modelling indicated that absolute brain mass was not related to either time spent or NCIO |ΔDR|. The explanatory power of these additional models, including absolute brain mass, was also very low (R2 = 0.01 and 0.04, respectively; Supplementary Tables S6, S7).
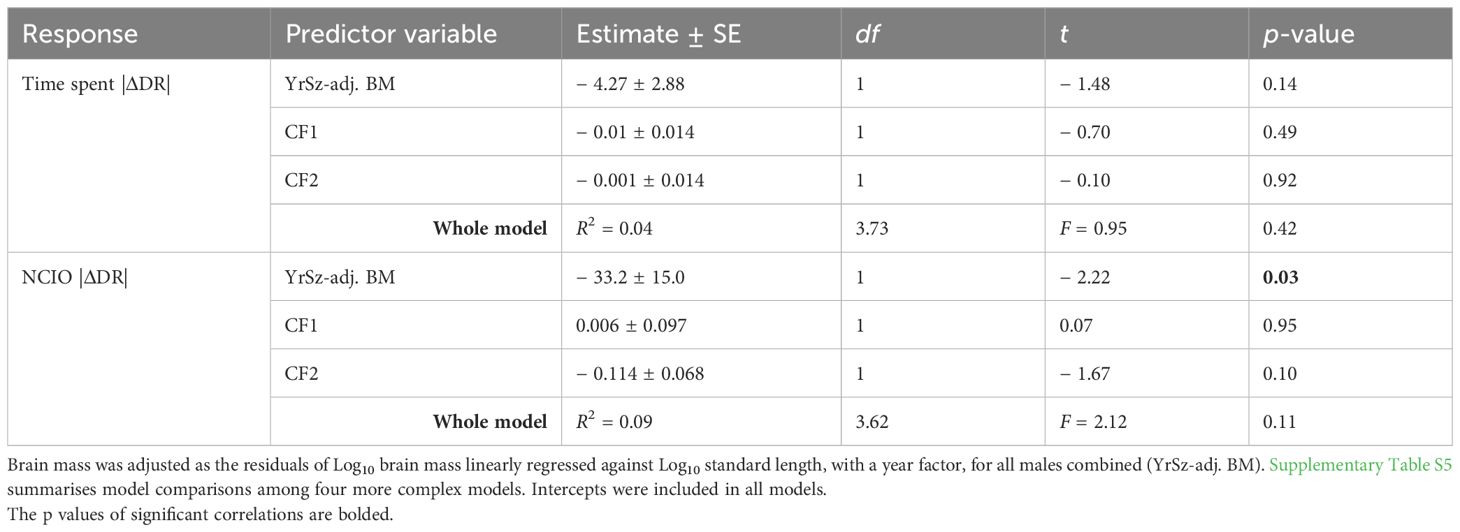
Table 3. Summary of reduced general linear models evaluating the influence of relative brain mass, after accounting for common behaviour factors (CF1 and CF2), on response measures of pumpkinseed novel object recognition performance (absolute values of delta DR time spent and noncontact interactions with objects [NCIO]).
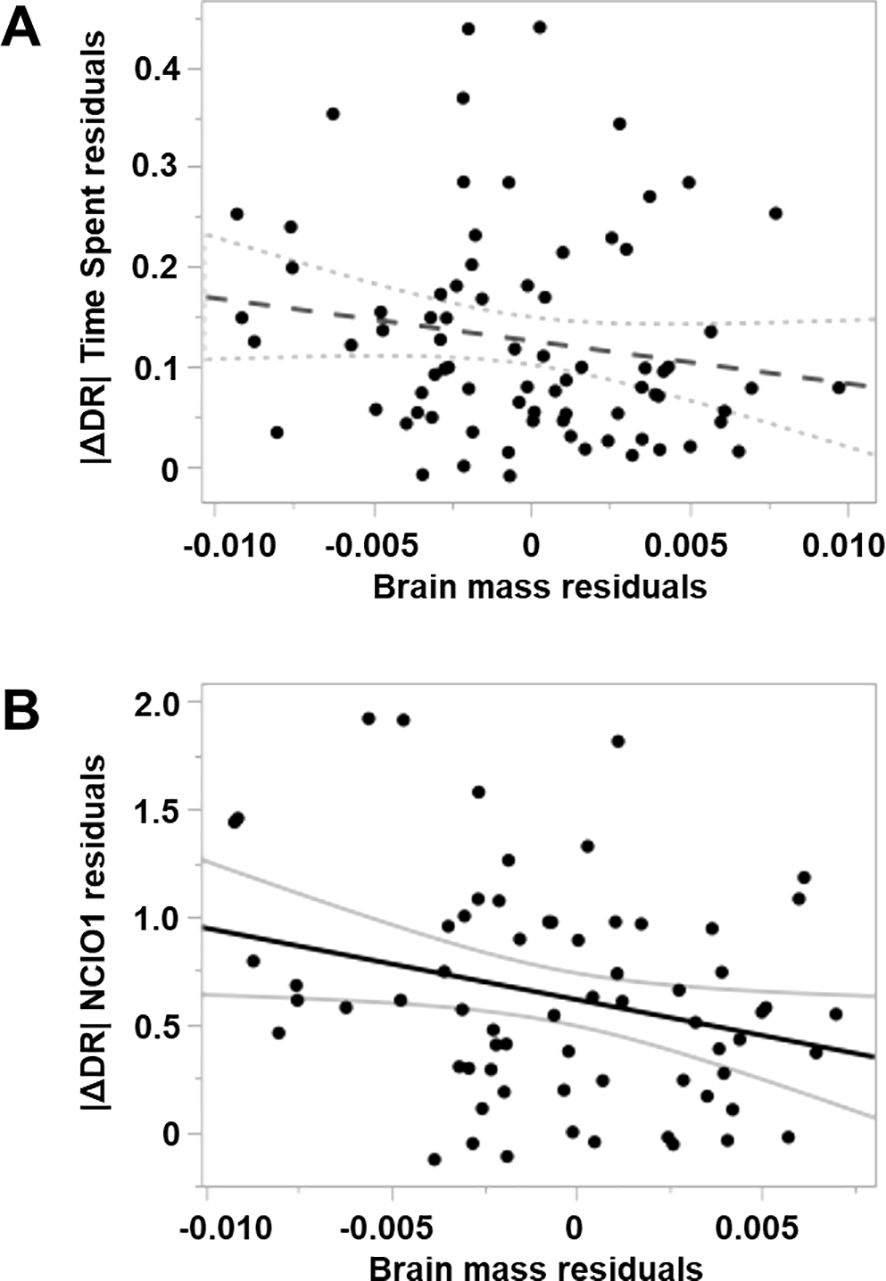
Figure 4. Partial regression relationships between residual novel object recognition performance (time spent or NCIO1) after accounting for two behavioural variables (CF1 and CF2), plotted against residual brain mass (adjusted for body size and year effects) (complete model summaries in Table 3). (A) Novel object recognition performance, estimated as the absolute values of ΔDR for nest side occupancy (time spent), is not significantly related to relative brain mass (p = 0.14, dashed best-fit line and 95% CI indicate the nonsignificant trend). (B) Novel object recognition performance, estimated as the absolute values of ΔDR for noncontact interactions with objects (NCIO1), is negatively related to relative brain mass (p = 0.03; best-fit line and 95% CI).
Discussion
We set out to evaluate the links between brain size, personality, and NOR performance in parental male pumpkinseed tested under natural conditions. Individuals with larger brains did not show better performance as we predicted. Instead, we found some evidence for the opposite relationship, in which individuals with relatively larger brains performed poorly in comparison to those with smaller brains. We also found no effect of behaviours related to object exploration tendency explaining variation in NOR performance, despite repeatability and consistency of these behaviours among parental male pumpkinseed.
NOR performance
NOR was previously studied in different fish species under controlled laboratory conditions (Hamilton, 2018), but others have argued that overreliance on captive-bred fish and lab-based studies may hinder our ability to gain generalisable insights about fish cognition (Salena et al., 2021). Here, we took advantage of nest fidelity during the reproductive period of parental male pumpkinseed to adapt the NOR assay for testing wild fish under field conditions. We believe that this is the first application of NOR to fish living in their natural environment, emphasising that NOR is a versatile assay that can be used as a tool to study fish cognition under a variety of circumstances when fish display a tendency to investigate objects. Evidence that parental males expressed NOR memory at the population level, despite a strong focus on nest defence and lack of feeding, suggests that the NOR assay is relevant to the cognitive ecology of this wild fish.
The chosen objects displayed at the nest periphery were detected because they produced overt responses by most individuals. However, responses to the novel object during the test phase of the NOR assay were highly variable. The expected preference for the novel object, based on the behaviour of rodents, is not always seen in fish (Hamilton, 2018). Adult guppies and zebrafish preferred novel objects in NOR assays (Braida et al., 2014; DePasquale et al., 2021; Lucon-Xiccato and Dadda, 2014; Lucon-Xiccato and Dadda, 2016; Magyary, 2019; Oliveira et al., 2015), while another study in zebrafish, newborn guppies, and a cichlid species showed the opposite pattern of preference for the familiar object (May et al., 2016; Miletto Petrazzini et al., 2012; Wallace and Hofmann, 2021), which can also be conceived as avoidance of the novel object. High variability expressed by parental male pumpkinseed individuals suggests that NOR could be expressed differently within the population, and justified our use of absolute values of ΔDRs to assess performance, effectively accounting for individual variation in preference or avoidance of the novel object. Moreover, NCIO produced larger |ΔDR| values than nest side preference, which may have provided greater performance variation among test subjects to effectively assess predictors of NOR performance, considering that we only found a significant predictor of performance with NCIO. Contacts with objects happened too infrequently to be a good measure of NOR performance, but this feature may be species-specific, because mouth contacts with floating objects have been used to assess NOR performance in zebrafish (Magyary, 2019). The most frequent behaviour represented by NCIO was by far approaches toward the objects (within half a body length but without contact with the object), indicating that counting approaches toward objects, regardless of whether contact with the object occurs, could be the best measure to use in fish NOR assays. This aligns with the use of measures of spatial proximity to the objects in most previous studies of fish NOR (Braida et al., 2014; DePasquale et al., 2021; Lucon-Xiccato and Dadda, 2014, Lucon-Xiccato and Dadda, 2016; May et al., 2016; Miletto Petrazzini et al., 2012; Oliveira et al., 2015; Wallace and Hofmann, 2021).
The only factor associated with parental male NOR performance in our analyses was relative brain mass when performance was assessed using NCIO |ΔDR|. The negative relationship between relative brain mass and NCIO |ΔDR| suggests that learning during nesting tends to be constrained in males with larger brains. This result was consistent across model selection exercises, indirectly providing evidence that this is a true characteristic of our system, despite the low explanatory power of these models. This low explanatory power is common in field-based studies with high data variability and underlines the need to account for other factors when assessing performance in cognitive tests of this kind.
The negative relationship between relative brain mass and NOR performance is initially counterintuitive and indeed runs contrary to previous tests of links between cognitive performance and brain size (Benson-Amram et al., 2016; Buechel et al., 2018; Kotrschal et al., 2013; MacLean et al., 2014). We propose that this result may reflect an energy constraint imposed by large brain size. Parental male pumpkinseed face strong energetic constraints during the breeding cycle because they stop active feeding. Parental males lose between 6% and 9% of their body mass during spawning through constant nest defence and brood care, and risk increased mortality after reproduction (Gillooly and Baylis, 1999; Gross, 1980; Rios-Cardenas and Webster, 2005). Investing in a larger brain should come at a high energetic cost because nervous tissue is especially costly to maintain (Niven and Laughlin, 2008; Padamsey and Rochefort, 2023; Tait et al., 2024). Thus, large-brained pumpkinseed may bear energetic costs that push them closer to the limit of their energy budget than small-brained pumpkinseed. There is also evidence that learning processes can be energetically costly, as demonstrated by increased energy uptake in brain tissue during learning (McNay et al., 2000; Plaçais et al., 2017), increased susceptibility to stress following learning (Mery and Kawecki, 2005; Jauman et al., 2013), and the inhibition of memory to favour survival during food shortage (Plaçais and Preat, 2013). Although the evidence for these energetic costs favours learning that is dependent on protein synthesis and leads to long-term memory, which is not directly relevant to the approximately one-hour time frame of pumpkinseed NOR memory observed here, other work also established high energetic costs for shorter working memory processes (McNay et al., 2000). With this in mind, we hypothesise that the negative relationship between relative brain mass and NOR performance reflects an energy expenditure limit reached by males with large relative brain size during breeding, inhibiting their ability to learn. Conversely, males with smaller relative brain mass, and thus lower energetic maintenance costs, had sufficient energy reserves to invest in learning processes that allowed at least some of them to express NOR memory. NOR learning should involve an energetic cost for this interpretation to be valid. Interestingly, male-specific inhibition of spatial learning during nest guarding has been observed in another fish species, the Cocos Frillgoby (Carbia and Brown, 2020), suggesting that learning processes in general may be subject to energetic constraints during periods of no or diminished foraging in fishes. Although individual male pumpkinseed vary in vigilance and clutch defence during nest guarding (Rios-Cardenas and Webster, 2005), we exclude a role for increased vigilance in explaining the relationship between relative brain mass and NOR performance because relative brain mass was not associated with CF2, our best proxy for male vigilance near the nest (discussed below), and CF2 did not influence NOR performance. If correct, our finding implies that energetic constraints on learning may depend on both the availability of external resources and the status of internal reserves, which can vary based on individual characteristics influencing the rate of energy consumption. Our approach highlights the need to carefully evaluate multiple potential factors that contribute to NOR performance in the field.
Observational studies, especially in nature, can suffer confounds when unknown factors influence the outcome in ways that are correlated with the effects of a known factor. Well-designed experimental manipulations can avoid this, but usually at the expense of ecological reality, which was an important goal here. The littoral ecotype has, on average, a larger relative brain size than the pelagic ecotype (Axelrod et al., 2018), and evaluating the effects of relative brain size on cognitive performance was our primary goal. However, by combining littoral and pelagic individuals to maximise variation in relative brain size in our NOR performance analyses, we risk a confound between brain size and habitat effects, should the latter also influence behaviour in the NOR test. For example, if the littoral habitat exposes individuals to greater ecological “novelty” than the pelagic habitat, then littoral fish might have a higher reactive threshold (i.e., lower interactivity) to the NOR test novelty compared with pelagic fish. If true, this would undermine our interpretation of the effects of relative brain size on NOR performance. Evidence of behavioural differences between habitats presented in Supplementary Table S4 makes such a confound plausible. We reject this habitat confound risk for two reasons. First, an individual’s NOR score removes individual differences in overall interactivity by calculating the difference in interactive behaviours taken in the familiarisation and test phases of the assay. Potential group differences in interactivity, such as those due to habitat, are removed in the same way. Second, we can evaluate the impact of a generic habitat effect on NOR performance by including it in the final NOR performance models shown in Table 3. Such post hoc analyses including a habitat parameter did not change the negative effect of relative brain size on NOR performance (Supplementary Table S8). Habitat is a significant predictor of NOR performance for time spent, but not for NCIO. Littoral fish had slightly higher average NOR scores for time spent than those in the pelagic habitat. Accounting for this habitat effect strengthened the trend for a negative effect of relative brain size on NOR performance measured using time spent (p = 0.07; Supplementary Table S8). The observational nature of our study precludes rejecting all possible confounds, but the protections just outlined suggest that confounds involving an influence of habitat on NOR performance are unlikely to undermine our basic observation of a negative effect of relative brain size on NOR performance.
Of note, there was a strong effect of year on parental male relative brain mass, body size, and body condition. The pattern of larger parental males in 2019 than in 2020, but better condition in 2020 than in 2019, is consistent with carryover effects on male spawning of a more severe winter in 2018–2019 compared to 2019–2020. Historical climate records of the nearest weather station (Bancroft, ON, 40 km distant) exhibit a lower mean winter monthly temperature and an additional month of ground snow cover in winter 2018–2019 than in 2019–2020. Smaller individuals face a higher probability of an energetic deficit in spring as winter length increases, because they have greater energy consumption than larger males during a period when food intake is severely reduced. This size-related energy deficit drives variation in the timing of spawning in parental males of different sizes over a single spawning season (Danylchuk and Fox, 1996) and is widely thought to generate increased risk of winter mortality in smaller compared to larger fish (reviewed in Suski and Ridgway, 2009). It seems reasonable, then, that a severe energy deficit arising from the 2018–2019 winter restricted spawning parentals to larger males in 2019, while the milder following winter permitted more smaller males to spawn and likely accounts for the higher condition of parental males in 2020. A severe energy deficit accruing over the 2018–2019 winter may also have triggered a plastic response to shrink relative brain size to minimise maintenance costs over the longer period of severely reduced food consumption. While our evidence of a plastic relative brain size response to winter severity is circumstantial, the consistent difference in relative brain size between ecotypes over years suggests that both ecotypes responded to various winter carryover effects in the same way. Importantly, post hoc analyses adding the effect of body condition (Scaled Mass Index) to the NOR performance models shown in Supplementary Table S9 did not change these results, and thus the negative relationship between relative brain mass and NOR performance appears impervious to year-to-year and ecotype differences in brain size.
Nest-guarding parental male personality
The exploratory common factor analysis revealed two groups of correlated behaviours that potentially reflect personality differences among parental male pumpkinseed sunfish. Although debate about animal personality continues (Roche et al., 2016), consistent individual differences in behaviour form the foundation of the concept (Carere and Locurto, 2011; Sih and Bell, 2008). We interpret the first common factor as a proactive exploration personality because individuals with higher CF1 values exhibited greater engagement with novel objects after returning to their nest (e.g., low latency to interact, longer total time interacting, higher noncontact and contact interactions with objects). LTR is less likely to represent object exploration behaviour compared with LTI and TTI, and more likely reflects a reaction to disturbance by the snorkel diver at the nest. We provisionally interpret variation in CF2 as differences in nesting vigilance personality. Males with higher values exhibited stronger responses to disturbance, such as returning more rapidly to their nest after the diver visit (low LTR) and showing greater reactivity toward novel objects (higher NCIO and CIO), independent of total time interacting. Personality traits, including exploration and boldness, are often associated with growth and body size (Biro and Stamps, 2008; Ferderer et al., 2022; Kelleher et al., 2017; Mayer et al., 2016). Larger males tended to be more proactively vigilant than smaller males in this study. This is consistent with the idea that when facing a trade-off between present and future reproductive investment (Sargent and Gross, 1986), parental males may optimise reproductive success by increasing current investment as they age (Coleman et al., 1985). Pelagic males also expressed greater vigilance, likely due to increased risk of nest invasion and fry predation (Popiel et al., 1996), because conspecific density is much higher at pelagic than littoral sites (Jarvis et al., 2020). Thus, intrinsic and extrinsic factors likely regulate personality differences here.
This is a rare demonstration of personality in the context of male reproduction in the wild, because both personalities identified on CF1 and CF2 map to several aspects of personality axes identified in fishes (e.g., shyness–boldness, exploration–avoidance, activity, aggressiveness, sociability; Conrad et al., 2011). However, two key uncertainties exist about these individual personality differences: the extent to which individual differences remain consistent across time and across contexts (Roche et al., 2016) other than nest-guarding; and how individual differences influence parental male reproductive success. Our standardised novel object recognition context provides only a narrow perspective on personality variation in these pumpkinseed, because we assess male behaviour over a short interval and only in the context of nest-guarding. Stable individual differences in behaviour over time have been reported in sunfish. Wilson and Godin (2009) reported that individual differences in shy–bold behaviour of wild juvenile bluegill sunfish (Lepomis macrochirus), assessed with sequential standardised laboratory tests, were maintained for up to 3 months in individuals released and recaptured from the field. However, evidence that individual differences in behaviour are stable across different natural contexts is weaker. Wilson et al. (1993) reported that differences in shyness–boldness assessed in juvenile pumpkinseed in the field were not consistent with standardised behavioural assessments under laboratory conditions. Furthermore, repeatable individual differences in behaviour observed in foraging or predation risk contexts in the field were inconsistent between those contexts (Coleman and Wilson, 1998). More tests are needed to confirm the concordance of differences measured during NOR assays with other methods used to assess personality (Toms et al., 2010).
The consequences of individual differences in parental male pumpkinseed behaviour here are also uncertain. While we found no evidence that personality influenced NOR performance in parental males, other studies suggest considerable scope for performance and fitness effects of personality in centrarchid sunfishes that are consistent with other species (e.g., Conrad et al., 2011; Mittelbach et al., 2014). In wild juvenile pumpkinseed, consistent individual differences in behaviour assessed in the field predicted individual differences in predation risk, diet, and parasite load (Wilson et al., 1993). In juvenile bluegill sunfish, boldness is also related to a movement syndrome involving increasingly intermittent locomotion (Wilson and Godin, 2009, Wilson and Godin, 2010), greater risk of capture by angling (Wilson et al., 2011), and greater aerobic capacity (Binder et al., 2016). However, evidence of an influence of personality on parental care and reproductive success is sparse. Male personality could influence competition among males for limited nest sites or influence female choice of parental males by signalling superior parental care (Conrad et al., 2011). For example, Mittelbach et al. (2014) reported that largemouth bass (Micropterus salmoides) assessed as bolder using laboratory assays have higher reproductive success in the field, although why is not clear. One hypothesis is that this may reflect correlated behaviour in male parental care that enhances offspring survival. Vargas et al. (2018) using zebrafish Danio rerio and Scherer et al. (2025) using the biparental cichlid Pelvicachromis pulcher separately reported that male “proactivity” and boldness, respectively, predict greater male parental care and subsequent offspring survival. Thus, the vigilance personality identified in parental male pumpkinseed here may influence nestling survival from conspecific nest raiding. Testing the function of personality on male reproductive success requires additional study.
Conclusions
We evaluated sunfish cognitive performance in a natural setting using a NOR test on parental pumpkinseed males during nest guarding and found that these sunfish demonstrate the ability to recognise novel objects, indicating that cognition can be measured in the wild using this approach. Contrary to our expectations and observations from laboratory experiments, we found that a larger brain size may hinder learning performance. We hypothesise that this reflects an energy expenditure limit reached by males with relatively larger brains during breeding. Differences between our results in a natural setting and previously reported associations between brain morphology and performance in laboratory experiments suggest that nest-guarding pumpkinseed may experience strong constraints on learning during the extended reproductive period. The pronounced effect of energetic context on cognition and its potential determinants warrant further investigation to improve our understanding of the relationships among animal personality, brain size, and cognitive performance.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.5683/SP3/TT6ZXP, Borealis, V2.
Ethics statement
The animal study was approved by University of Guelph Animal Care Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
KM: Conceptualization, Writing – review & editing, Investigation, Formal Analysis. MD: Formal Analysis, Conceptualization, Writing – review & editing, Investigation. CA: Investigation, Conceptualization, Writing – review & editing. BR: Funding acquisition, Conceptualization, Writing – original draft, Formal Analysis, Supervision. FL: Formal Analysis, Writing – original draft, Funding acquisition, Supervision.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by Discovery Grants from the National Science and Engineering Research Council of Canada to FL and BR.
Acknowledgments
The authors wish to acknowledge Adam DeLeeuw for his assistance during fieldwork, as well as Bob and Christine Gautier and the Ashby Lake Protective Association for facilitating the fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fetho.2025.1683770/full#supplementary-material
References
Anderson B. (1993). Evidence from the rat for a general factor that underlies cognitive performance and that relates to brain size: intelligence? Neurosci. Lett. 153, 98–102. doi: 10.1016/0304-3940(93)90086-Z
Axelrod C. J., Laberge F., and Robinson B. W. (2018). Intraspecific brain size variation between coexisting sunfish ecotypes. Proc. B 285, 20181971. doi: 10.1098/rspb.2018.1971
Bell A. M., Hankison S. J., and Laskowsk K. L. (2009). The repeatability of behaviour: a meta-analysis. Anim. Behav. 77, 771–783. doi: 10.1016/j.anbehav.2008.12.022
Benson-Amram S., Dantzer B., Stricker G., Swanson E. M., and Holekamp K. E. (2016). Brain size predicts problem-solving ability in mammalian carnivores. Proc. Natl. Acad. Sci. U.S.A. 113, 2532–2537. doi: 10.1073/pnas.1505913113
Berchtold A. E., Colborne S. F., Longstaffe F. J., and Neff B. D. (2015). Ecomorphological patterns linking morphology and diet across three populations of pumpkinseed sunfish (Lepomis gibbosus). Can. J. Zool. 93, 289–297. doi: 10.1139/cjz-2014-0236
Binder T. R., Wilson A. D. M., Wilson S. M., Suski C. D., Godin J.-G., and Cooke S. J. (2016). Is there a pace-of-life syndrome linking boldness and metabolic capacity for locomotion in bluegill sunfish? Anim. Behav. 121, 175–183. doi: 10.1016/j.anbehav.2016.09.006
Biro P. A. and Stamps J. A. (2008). Are animal personality traits linked to life-history productivity? Trends Ecol. Evol. 23, 361–368. doi: 10.1016/j.tree.2008.04.003
Blaser R. and Heyser C. (2015). Spontaneous object recognition: A promising approach to the comparative study of memory. Front. Behav. Neurosci. 9. doi: 10.3389/fnbeh.2015.00183
Boogert N. J., Madden J. R., Morand-Ferron J., and Thornton A. (2018). Measuring and understanding individual differences in cognition. Philo. Trans. R. Soc B 373, 20170280. doi: 10.1098/rstb.2017.0280
Brady T. F., Konkle T., Alvarez G. A., and Oliva A. (2008). Visual long-term memory has a massive storage capacity for object details. Proc. Natl. Acad. Sci. U.S.A. 105, 14325–14329. doi: 10.1073/pnas.0803390105
Braida D., Ponzoni L., Martucci R., and Sala M. (2014). A new model to study visual attention in zebrafish. Prog. Neuropsychopharmacol. Biol. Psychiatry 55, 80–86. doi: 10.1016/j.pnpbp.2014.03.010
Brewer M. J., Butler A., and Cooksley S. L. (2016). The relative performance of AIC, AICc and BIC in the presence of unobserved heterogeneity. Methods Ecol. Evol. 7, 679–692. doi: 10.1111/2041-210X.12541
Buechel S. D., Boussard A., Kotrschal A., van der Bijl W., and Kolm N. (2018). Brain size affects performance in a reversal-learning test. Proc. B 285, 20172031. doi: 10.1098/rspb.2017.2031
Burns J. G. and Rodd F. H. (2008). Hastiness, brain size and predation regime affect the performance of wild guppies in a spatial memory task. Anim. Behav. 76, 911–922. doi: 10.1016/j.anbehav.2008.02.017
Carbia P. S. and Brown C. (2020). Seasonal variation of sexually dimorphic spatial learning implicates mating system in the intertidal Cocos Frillgoby (Bathygobius cocosensis). Anim. Cog. 23, 621–628. doi: 10.1007/s10071-020-01366-3
Carere C. and Locurto C. (2011). Interaction between animal personality and animal cognition. Curr. Zool. 57, 491–498. doi: 10.1093/czoolo/57.4.491
Cargnelli L. M. and Neff B. D. (2006). Condition-dependent nesting in bluegill sunfish Lepomis macrochirus. J. Anim. Ecol. 75, 627–633. doi: 10.1111/j.1365-2656.2006.01083.x
Cauchoix M., Chaine A. S., and Barragan-Jason G. (2020). Cognition in context: plasticity in cognitive performance in response to ongoing environmental variables. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00106
Chittka L. and Niven J. (2009). Are bigger brains better? Curr. Biol. 19, R995–R1008. doi: 10.1016/j.cub.2009.08.023
Colborne S. F., Garner S. R., Longstaffe F. J., and Neff B. D. (2016). Assortative mating but no evidence of genetic divergence in a species characterized by a trophic polymorphism. J. Evol. Biol. 29, 633–644. doi: 10.1111/jeb.12812
Coleman R. M. and Fischer R. U. (1991). Brood size, male fanning effort and the energetics of a nonshareable parental investment in bluegill sunfish, Lepomis macrochirus (Teleostei: Centrarchidae). Ethology 87, 177–188. doi: 10.1111/j.1439-0310.1991.tb00245.x
Coleman R. M., Gross M. R., and Sargent R. C. (1985). Parental investment decision rules: a test in bluegill sunfish. Behav. Ecol. Sociobiol. 18, 59–66. doi: 10.1007/BF00299238
Coleman K. and Wilson D. S. (1998). Shyness and boldness in pumpkinseed sunfish: individual differences are context-specific. Anim. Behav. 56, 927–936. doi: 10.1006/anbe.1998.0852
Colgan P. W. and Gross M. R. (1977). Dynamics of aggression in male pumpkinseed sunfish (Lepomis gibbosus) over the reproductive phase. Z. Tierpsychol. 43, 139–151. doi: 10.1111/j.1439-0310.1977.tb00064.x
Conrad J. L., Weinersmith K. L., Brodin T., Saltz J. B., and Sih A. (2011). Behavioural syndromes in fishes: a review with implications for ecology and fisheries management. J. Fish Biol. 78, 395–435. doi: 10.1111/j.1095-8649.2010.02874.x
Cooke S. J., Weatherhead D. H., Wahl D. H., and Philipp D. P. (2008). Parental care in response to natural variation in nest predation pressure in six sunfish (Centrarchidae: Teleostei) species. Ecol. Freshw. Fish 17, 628–638. doi: 10.1111/j.1600-0633.2008.00314.x
Daniel D. K. and Bhat A. (2020). Bolder and brighter? Exploring correlations between personality and cognitive abilities among individuals within a population of wild zebrafish, Danio rerio. Front. Behav. Neurosci. 14. doi: 10.3389/fnbeh.2020.00138
Danylchuk A. J. and Fox M. G. (1996). Size- and age-related variation in the seasonal timing of nesting activity, nest characteristics, and female choice of parental male pumpkinseed sunfish (Lepomis gibbosus). Can. J. Zool. 74, 1834–1840. doi: 10.1139/z96-206
DePasquale C., Kemerer N., White N., Yost M., Wolfkill J., Sturgill J., et al. (2021). The Influence of an enriched environment in enhancing recognition memory in zebrafish (Danio rerio). Front. Vet. Sci. 8. doi: 10.3389/fvets.2021.749746
Dubois J., Galdi P., Paul L. K., and Adolphs R. (2018). A distributed brain network predicts general intelligence from resting-state human neuroimaging data. Philo. Trans. R. Soc B 373, 20170284. doi: 10.1098/rstb.2017.0284
Ennaceur A. and Delacour J. (1988). A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 31, 47–59. doi: 10.1016/0166-4328(88)90157-X
Ferderer A., Davis A. R., and Wong M. Y. L. (2022). Temperature and body size influence personality and behavioural syndromes in an invasive crayfish. Anim. Behav. 190, 187–198. doi: 10.1016/j.anbehav.2022.06.009
Gibelli J., Aubin-Horth N., and Dubois F. (2019). Individual differences in anxiety are related to differences in learning performance and cognitive style. Anim. Behav. 157, 121–128. doi: 10.1016/j.anbehav.2019.09.008
Gillespie G. J. and Fox M. G. (2003). Morphological and life-history differentiation between littoral and pelagic forms of pumpkinseed. J. Fish Biol. 62, 1099–1115. doi: 10.1046/j.1095-8649.2003.00100.x
Gillooly J. F. and Baylis J. R. (1999). Reproductive success and the energetic cost of parental care in male smallmouth bass. J. Fish Biol. 54, 573–584. doi: 10.1111/j.1095-8649.1999.tb00636.x
Greenspan R. J. and van Swinderen B. (2004). Cognitive consonance: complex brain functions in the fruit fly and its relatives. Trends Neurosci. 27, 707–711. doi: 10.1016/j.tins.2004.10.002
Gross M. R. (1980). Sexual selection and the evolution of reproductive strategies in sunfishes (Lepomis: Centrarchidae) (Salt Lake City, Utah, USA: The University of Utah ProQuest Dissertations & Theses). 1980. 8017132.
Gross M. R. (1982). Sneakers, satellites and parentals: polymorphic mating strategies in North American sunfishes. Z. Tierpsychol. 60, 1–26. doi: 10.1111/j.1439-0310.1982.tb01073.x
Hamilton T. J. (2018). “Object novelty and object location recognition memory in fish - Recent advances,” in Handbook of object novelty recognition. Eds. Ennaceur A. and de Souza Silva M. A. (Academic Press, London), 151–161. doi: 10.1016/B978-0-12-812012-5.00009-4
Jarvis W. M. C., Comeau S. M., Colborne S. F., and Robinson B. W. (2017). Flexible mate choice may contribute to ecotype assortative mating in pumpkinseed sunfish (Lepomis gibbosus). J. Evol. Biol. 30, 1810–1820. doi: 10.1111/jeb.13127
Jarvis W. M. C., Peiman K. S., Cooke S. J., and Robinson B. W. (2020). Low connectivity between sympatric populations of sunfish ecotypes suggests ecological opportunity contributes to diversification. Evol. Ecol. 34, 391–410. doi: 10.1007/s10682-020-10042-4
Jastrebski C. J. and Robinson B. W. (2004). Natural selection and the evolution of replicated trophic polymorphisms in pumpkinseed sunfish (Lepomis gibbosus). Evol. Ecol. Res. 6, 285–305.
Jauman S., Scudelari R., and Naug D. (2013). Energetic cost of learning and memory can cause cognitive impairment in honeybees. Biol. Lett. 9, 20130149. doi: 10.1098/rsbl.2013.0149
Jones N. A. R., Webster M., Newport C., Templeton C. N., Schuster S., and Rendell L. (2020). Cognitive styles: speed–accuracy trade-offs underlie individual differences in archerfish. Anim. Behav. 160, 1–14. doi: 10.1016/j.anbehav.2019.11.019
Joyce B. J. and Brown G. E. (2020). Short-term captivity drives hypothalamic plasticity and asymmetry in wild-caught northern red bellied dace (Chrosomus eos). J. Fish Biol. 97, 577–582. doi: 10.1111/jfb.14408
Keenleyside M. H. A. (1972). Intraspecific intrusions into nests of spawning longear sunfish (Pisces: Centrarchidae). Copeia 1972, 272–278. doi: 10.2307/1442488
Kelleher S. R., Silla A. J., Dingemanse N. J., and Byrne P. G. (2017). Body size predicts between-individual differences in exploration behaviour in the southern corroboree frog. Anim. Behav. 129, 161–170. doi: 10.1016/j.anbehav.2017.05.013
Koivisto H., Maguire C., and Tanila H. (2025). Novel object recognition task for mice: Is it a test for memory, object neophobia or innate preference? Behav. Brain Res. 491, 115649. doi: 10.1016/j.bbr.2025.115649
Kotrschal A., Lievens E. J. P., Dahlbom J., Bundsen A., Semenova S., Sundvik M., et al. (2014). Artificial selection on relative brain size reveals a positive genetic correlation between brain size and proactive personality in the guppy. Evolution 68, 1339–1349. doi: 10.1111/evo.12341
Kotrschal A., Rogell B., Bundsen A., Svensson B., Zajitschek S., Brännström I., et al. (2013). Artificial selection on relative brain size in the guppy reveals costs and benefits of evolving a larger brain. Curr. Biol. 23, 168–171. doi: 10.1016/j.cub.2012.11.058
Kotrschal A. and Taborsky B. (2010). Environmental change enhances cognitive abilities in fish. PloS Biol. 8, e1000351. doi: 10.1371/journal.pbio.1000351
Lambert C. T. and Guillette L. M. (2021). The impact of environmental and social factors on learning abilities: a meta-analysis. Biol. Rev. 96, 2871–2889. doi: 10.1111/brv.12783
Logan C. J., Avin S., Boogert N., Buskell A., Cross F. R., Currie A., et al. (2018). Beyond brain size: Uncovering the neural correlates of behavioral and cognitive specialization. Comp. Cogn. Behav. Rev. 13, 55–89. doi: 10.3819/CCBR.2018.130008
Lucon-Xiccato T. and Bisazza A. (2017). Individual differences in cognition among teleost fishes. Behav. Processes 141, 184–195. doi: 10.1016/j.beproc.2017.01.015
Lucon-Xiccato T. and Dadda M. (2014). Assessing memory in zebrafish using the one-trial test. Behav. Processes 106, 1–4. doi: 10.1016/j.beproc.2014.03.010
Lucon-Xiccato T. and Dadda M. (2016). Guppies show behavioural but not cognitive sex differences in a novel object recognition test. PloS One 11, e0156589. doi: 10.1371/journal.pone.0156589
Lupien S. J., McEwen B. S., Gunnar M. R., and Heim C. (2009). Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 10, 434–445. doi: 10.1038/nrn2639
MacLean E. L., Hare B., Nunn C. L., Addessi E., Amici F., Anderson R. C., et al. (2014). The evolution of self-control. Proc. Natl. Acad. Sci. U.S.A. 111, E2140–E2148. doi: 10.1073/pnas.1323533111
Magyary I. (2019). Floating novel object recognition in adult zebrafish: a pilot study. Cogn. Process. 20, 359–362. doi: 10.1007/s10339-019-00910-5
Matzel L. D., Townsend D. A., Grossman H., Han Y. R., Hale G., Zappulla M., et al. (2006). Exploration in outbred mice covaries with general learning abilities irrespective of stress reactivity, emotionality, and physical attributes. Neurobiol. Learn. Mem. 86, 228–240. doi: 10.1016/j.nlm.2006.03.004
May Z., Morrill A., Holcombe A., Johnston T., Gallup J., Fouad K., et al. (2016). Object recognition memory in zebrafish. Behav. Brain Res. 296, 199–210. doi: 10.1016/j.bbr.2015.09.016
Mayer M., Shine R., and Brown G. P. (2016). Bigger babies are bolder: effects of body size on personality of hatchling snakes. Behaviour 153, 313–323. doi: 10.1163/1568539X-00003343
Mazza V., Eccard J. A., Zaccaroni M., Jacob J., and Dammhahn M. (2018). The fast and the flexible: cognitive style drives individual variation in cognition in a small mammal. Anim. Behav. 137, 119–132. doi: 10.1016/j.anbehav.2018.01.011
McNay E. C., Fries T. M., and Gold P. E. (2000). Decreases in rat extracellular hippocampal glucose concentration associated with cognitive demand during a spatial task. Proc. Natl. Acad. Sci. U.S.A. 97, 2881–2885. doi: 10.1073/pnas.050583697
Mery F. and Kawecki T. J. (2005). A cost of long-term memory in Drosophila. Science 308, 1148. doi: 10.1126/science.1111331
Miletto Petrazzini M. E., Agrillo C., Piffer L., Dadda M., and Bisazza A. (2012). Development and application of a new method to investigate cognition in newborn guppies. Behav. Brain Res. 233, 443–449. doi: 10.1016/j.bbr.2012.05.044
Milla S., Pasquet A., El Mohajer L., and Fontaine P. (2021). How domestication alters fish phenotypes. Rev. Aquacult. 13, 388–405. doi: 10.1111/raq.12480
Mittelbach G. G., Ballew N. G., and Kjelvik M. K. (2014). Fish behavioral types and their ecological consequences. Can. J. Fish. Aquat. Sci. 71, 927–944. doi: 10.1139/cjfas-2013-0558
Morand-Ferron J., Cole E. F., and Quinn J. L. (2016). Studying the evolutionary ecology of cognition in the wild: a review of practical and conceptual challenges. Biol. Rev. 91, 367–389. doi: 10.1111/brv.12174
Niven J. E. and Laughlin S. B. (2008). Energy limitation as a selective pressure on the evolution of sensory systems. J. Exp. Biol. 211, 1792–1804. doi: 10.1242/jeb.017574
Oliveira J., Silveira M., Chacon D., and Luchiari A. (2015). The zebrafish world of colors and shapes: preference and discrimination. Zebrafish 12, 166–173. doi: 10.1089/zeb.2014.1019
Olla B. L., Davis M. W., and Ryer C. H. (1998). Understanding how the hatchery environment represses or promotes the development of behavioral survival skills. Bull. Mar. Sci. 62, 531–550.
Padamsey Z. and Rochefort N. L. (2023). Paying the brain’s energy bill. Curr. Opin. Neurobiol. 78, 102668. doi: 10.1016/j.conb.2022.102668
Park P. J., Chase I., and Bell M. A. (2012). Phenotypic plasticity of the threespine stickleback Gasterosteus aculeatus telencephalon in response to experience in captivity. Curr. Zool. 58, 189–210. doi: 10.1093/czoolo/58.1.189
Peig J. and Green A. J. (2010). The paradigm of body condition: a critical reappraisal of current methods based on mass and length. Funct. Ecol. 24, 1323–1332. doi: 10.1111/j.1365-2435.2010.01751.x
Peiman K. S. and Robinson B. W. (2017). Comparative analyses of phenotypic trait covariation within and among populations. Am. Nat. 190, 451–468. doi: 10.1086/693482
Plaçais P.-Y., de Tredern É., Scheunemann L., Trannoy S., Goguel V., Han K.-A., et al. (2017). Upregulated energy metabolism in the Drosophila mushroom body is the trigger for long-term memory. Nat. Commun. 8, 15510. doi: 10.1038/ncomms15510
Plaçais P.-Y. and Preat T. (2013). To favor survival under food shortage, the brain disables costly memory. Science 339, 440–442. doi: 10.1126/science.1226018
Popiel S. A., Pérez-Fuentetaja A., McQueen D. J., and Collins N. C. (1996). Determinants of nesting success in the pumpkinseed (Lepomis gibbosus): a comparison of two populations under different risks from predation. Copeia 1996, 649–658. doi: 10.2307/1447529
Reader S. M., Hager Y., and Laland K. N. (2011). The evolution of primate general and cultural intelligence. Philo. Trans. R. Soc B 366, 1017–1027. doi: 10.1098/rstb.2010.0342
Rios-Cardenas O. and Webster M. S. (2005). Paternity and paternal effort in the pumpkinseed sunfish. Behav. Ecol. 16, 914–921. doi: 10.1093/beheco/ari076
Robinson B. W. and Wilson D. S. (1996). Genetic variation and phenotypic plasticity in a trophically polymorphic population of pumpkinseed sunfish (Lepomis gibbosus). Evol. Ecol. 10, 631–652. doi: 10.1007/BF01237711
Robinson B. W., Wilson D. S., and Margosian A. S. (2000). A pluralistic analysis of character release in pumpkinseed sunfish (Lepomis gibbosus). Ecology 81, 2799–2812. doi: 10.1890/0012-9658(2000)081[2799:APAOCR]2.0.CO;2
Robinson B. W., Wilson D. S., Margosian A. S., and Lotito P. T. (1993). Ecological and morphological differentiation of pumpkinseed sunfish in lakes without bluegill sunfish. Evol. Ecol. 7, 451–464. doi: 10.1007/BF01237641
Roche D. G., Careau V., and Binning S. A. (2016). Demystifying animal ‘personality’ (or not): why individual variation matters to experimental biologists. J. Exp. Biol. 219, 3832–3843. doi: 10.1242/jeb.146712
Salena M. G., Turko A. J., Singh A., Pathak A., Hughes E., Brown C., et al. (2021). Understanding fish cognition: a review and appraisal of current practices. Anim. Cogn. 24, 395–406. doi: 10.1007/s10071-021-01488-2
Sargent R. C. and Gross M. R. (1986). “Williams’ principle: an explanation of parental care in teleost fishes,” in The behaviour of teleost fishes. Ed. Pitcher T. J. (Springer, Boston, MA), 275–293. doi: 10.1007/978-1-4684-8261-4_11
Scherer U., Niemelä P. T., and Schuett W. (2025). Male activity under risk predicts paternal care and reproductive success in a bi-parental cichlid. Behav. Ecol. 36, araf021. doi: 10.1093/beheco/araf021
Schmitt T. A. (2011). Current methodological considerations in exploratory and confirmatory factor analysis. J. Psychoeduc. Assess. 29, 304–321. doi: 10.1177/0734282911406653
Schuster A. C., Zimmermann U., Hauer C., and Foerster K. (2017). A behavioural syndrome, but less evidence for a relationship with cognitive traits in a spatial orientation context. Front. Zool. 14, 19. doi: 10.1186/s12983-017-0204-2
Sih A. and Bell A. M. (2008). Insights for behavioral ecology from behavioral syndromes. Adv. Stud. Behav. 38, 227–281. doi: 10.1016/S0065-3454(08)00005-3
Sih A. and Del Giudice M. (2012). Linking behavioural syndromes and cognition: a behavioural ecology perspective. Philo. Trans. R. Soc B 367, 2762–2772. doi: 10.1098/rstb.2012.0216
Sivakumaran M. H., Mackenzie A. K., Callan I. R., Ainge J. A., and O’Connor A. R. (2018). The discrimination ratio derived from novel object recognition tasks as a measure of recognition memory sensitivity, not bias. Sci. Rep. 8, 11579. doi: 10.1038/s41598-018-30030-7
Snook S. C. and Gorsuch R. L. (1989). Component analysis versus common factor analysis: A Monte Carlo study. Psych. Bull. 106, 148–154. doi: 10.1037/0033-2909.106.1.148
Suski C. D. and Ridgway M. S. (2009). “Winter biology of centrarchid fishes,” in Centrarchid fishes: diversity, biology, and conservation. Eds. Cooke S. J. and Philipp D. P. (Wiley Blackwell, Hoboken, NJ), 264–292. doi: 10.1002/9781444316032.ch9
Tait C., Chicco A. J., and Naug D. (2024). Brain energy metabolism as an underlying basis of slow and fast cognitive phenotypes in honeybees. J. Exp. Biol. 227, jeb247835. doi: 10.1242/jeb.249359
Thorp J. H., Goldsmith L. D., Polgreen J. A., and Mayer L. M. (1989). Foraging Patterns of Nesting and Nonnesting Sunfish (Centrarchidae: Lepomis auritus and L. gibbosus). Can. J. Fish. Aquat. Sci. 46, 1342–1346. doi: 10.1139/f89-172
Toms C. N., Echevarria D. J., and Jouandot D. J. (2010). A methodological review of personality-related studies in fish: Focus on the shy-bold axis of behavior. Int. J. Comp. Psychol. 23, 1–25. doi: 10.46867/ijcp.2010.23.01.08
Triki Z., Fong S., Amcoff M., Vàsquez-Nilsson S., and Kolm N. (2023). Experimental expansion of relative telencephalon size improves the main executive function abilities in guppy. PNAS Nexus 2, 1–11. doi: 10.1093/pnasnexus/pgad129
Turschwell M. P. and White C. R. (2016). The effects of laboratory housing and spatial enrichment on brain size and metabolic rate in the eastern mosquitofish, Gambusia holbrooki. Biol. Open 5, 205–210. doi: 10.1242/bio.015024
Vargas R., Mackenzie S., and Rey S. (2018). ‘Love at first sight’: The effect of personality and colouration patterns in the reproductive success of zebrafish (Danio rerio). PLOS One 13, e0203320. doi: 10.1371/journal.pone.0203320
Vehanen T., Huusko A., and Hokki R. (2009). Competition between hatchery-raised and wild brown trout Salmo trutta in enclosures – do hatchery releases have negative effects on wild populations? Ecol. Freshw. Fish 18, 261–268. doi: 10.1111/j.1600-0633.2008.00343.x
Wallace K. J. and Hofmann H. A. (2021). Equal performance but distinct behaviors: sex differences in a novel object recognition task and spatial maze in a highly social cichlid fish. Anim. Cogn. 24, 1057–1073. doi: 10.1007/s10071-021-01498-0
Weese D. J., Ferguson M. M., and Robinson B. W. (2012). Contemporary and historical evolutionary processes interact to shape patterns of within-lake phenotypic divergences in polyphenic pumpkinseed sunfish, Lepomis gibbosus. Ecol. Evol. 2, 574–592. doi: 10.1002/ece3.72
Wilson A. D. M., Binder T. R., McGrath K. P., Cooke S. J., and Godin J.-G. (2011). Capture technique and fish personality: angling targets timid bluegill sunfish, Lepomis macrochirus. Can. J. Fish. Aquat. Sci. 68, 749–757. doi: 10.1139/f2011-019
Wilson D. S., Clark A. B., Coleman K., and Dearstyne T. (1994). Shyness and boldness in humans and other animals. Trends Ecol. Evol. 9, 442–446. doi: 10.1016/0169-5347(94)90134-1
Wilson D. S., Coleman K., Clark A. B., and Biederman L. (1993). Shy-bold continuum in pumpkinseed sunfish (Lepomis gibbosus): An ecological study of a psychological trait. J. Comp. Psychol. 107, 250–260. doi: 10.1037/0735-7036.107.3.250
Wilson A. D. M. and Godin J.-G. (2009). Boldness and behavioral syndromes in the bluegill sunfish, Lepomis macrochirus. Behav. Ecol. 20, 231–237. doi: 10.1093/beheco/arp018
Keywords: animal personality, brain size, cognition, fish, learning and memory, novel object recognition
Citation: McAllister K, Dolan M, Axelrod CJ, Robinson BW and Laberge F (2025) Assessing cognitive performance in nature: brain size and personality correlates of novel object recognition in nest-guarding male pumpkinseed sunfish. Front. Ethol. 4:1683770. doi: 10.3389/fetho.2025.1683770
Received: 11 August 2025; Accepted: 27 October 2025;
Published: 10 November 2025.
Edited by:
Zonghang Zhang, Shantou University, ChinaReviewed by:
Brendan J. Joyce, Concordia University, CanadaAnnabell Klinke, Macquarie University, Australia
Copyright © 2025 McAllister, Dolan, Axelrod, Robinson and Laberge. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédéric Laberge, ZmxhYmVyZ2VAdW9ndWVscGguY2E=
†Present addresses: Keith McAllister, Stantec Consulting Ltd., Waterloo, ON, Canada
Maria Dolan, Department of Integrative Biology, Wilfrid Laurier University, Waterloo, ON, Canada
Caleb J. Axelrod, Department of Biology, Swarthmore College, Swarthmore, PA, United States